
Sistemi riproduttivi
Sistemi riproduttivi
I sistemi riproduttivi sono caratterizzati dai comportamenti riguardanti le relazioni tra partner sessuali e le cure parentali prestate da individui di entrambi i sessi. Essi comprendono il numero di partner di un individuo, il modo con cui sono acquisiti i partner, le modalità e la durata delle cure parentali. Due caratteristiche fondamentali dell'ambiente sono associate all'evoluzione delle varie forme di sistemi riproduttivi: la distribuzione delle risorse nello spazio e nel tempo. Studi recenti hanno evidenziato che anche le scelte dirette del partner possono svolgere un ruolo ed è stata introdotta la distinzione tra partner sociali e partner genetici. Nuovi progressi in campo teorico hanno permesso di associare in modo più diretto questi aspetti della selezione sessuale alle cure parentali offerte dai partner, mentre studi fìlogenetici stanno estendendo le nostre conoscenze al campo delle transizioni storiche tra i diversi tipi di sistemi riproduttivi.
I maschi degli elefanti marini del Nord (Miro unga angustirostris), che pesano circa 2300 kg, hanno qualcosa in comune con i maschi di acaro d'acqua (Unionicola Jormosa) lunghi appena 5 mm. In entrambe le specie essi combattono vigorosamente per conquistare partner sessuali e i vincitori possono ottenere l'accesso esclusivo a più di 50 femmine durante una singola stagione riproduttiva. Lo studio dei sistemi riproduttivi degli animali ha l'obiettivo di spiegare le ragioni che sono alla base dell'evoluzione di differenze di comportamento così marcate come quelle che si osservano tra i sessi. Forse queste specie condividono somiglianze in certe caratteristiche chiave del loro habitat e della loro storia naturale. Di fatto, in altre specie le differenze ambientali sono in grado di spiegare l'evoluzione di differenti sistemi riproduttivi. I sistemi riproduttivi possono essere definiti come descrizioni del comportamento inerente la formazione e l'organizzazione del legame di coppia e delle cure parentali da parte degli individui di ambo i sessi (Reynolds, 1996). Essi descrivono la variabilità che si osserva tra individui di entrambi i sessi nei seguenti aspetti del comportamento riproduttivo: modalità e durata delle cure parentali; modalità e durata dei legami di coppia; numero di partner acquisiti ('genetici' e 'sociali'); forme di corteggiamento, coercizione e competizione (inclusa quella tra spermatozoi di maschi diversi); difesa e offerta di risorse utili al partner; grado di scelta del partner (inclusa quella tra spermatozoi). Queste componenti comprendono quelle riportate in origine da S.T. Emlen e L.W. Oring (1977) nella loro classica formulazione dei sistemi di accoppiamento (mating systems), ma anche numerosi aspetti del comportamento riproduttivo emersi da nuovi studi, effettuati dall'inizio degli anni Ottanta. Oggi, infatti, viene data molta importanza alla distinzione tra partner sociali e partner genetici, agli studi sulla competizione tra spermatozoi, sulla coercizione e scelta del partner, con un'enfasi crescente sulla variabilità di comportamento tra individui di entrambi i sessi, rinunciando alla precedente tendenza a focalizzare l'attenzione soprattutto sul sesso maschile. Attualmente, nella letteratura scientifica la locuzione sistema riproduttivo viene preferita a sistema di accoppiamento, in quanto quest'ultima implica una restrizione al solo comportamento di coppia (che è il dominio della selezione sessuale), mentre il primo include anche il comportamento parentale. Quest'ultimo costituisce una caratteristica fondamentale di ogni descrizione e spiegazione degli aspetti sociali della riproduzione. Per comprendere la diversità dei sistemi riproduttivi degli animali è necessario considerare la selezione che agisce su ciascun sesso nel momento in cui un individuo fornisce cure parentali o compete per un partner.
Differenze tra i sessi nella competizione per riprodursi
Charles Darwin (1871) pose le fondamenta della moderna teoria dei sistemi riproduttivi enfatizzando le differenze tra i sessi nella competizione per la riproduzione. Nella maggioranza delle specie uno dei due sessi (in genere quello maschile) tenta di accoppiarsi con più individui dell'altro sesso. La variabilità esistente tra i maschi relativamente alla capacità di avere successo nella competizione per le femmine, potrebbe aver esercitato una fortissima influenza sull'evoluzione di 'armi', come i palchi dei cervi, o di tratti vistosi da esibire per attrarre il sesso opposto, come le piume degli uccelli del paradiso e dei fagiani. Darwin definì questa competizione e la relativa scelta del partner come selezione sessuale. La descrizione della selezione sessuale, in particolare della variabilità nel numero di partner ottenuti da ciascun sesso e del modo con cui sono conquistati, permette di delineare, almeno in parte, le caratteristiche del sistema riproduttivo di una popolazione. Il passo successivo consiste nel comprendere l'evoluzione delle cure parentali, che includono qualsiasi comportamento che aumenti la fitness della prole di un animale. Infatti il comportamento parentale è fortemente legato alla selezione sessuale e fornisce elementi importanti ai ricercatori che tentano di costruire una teoria completa dei sistemi riproduttivi. Se uno dei due sessi fornisce poca o nessuna cura alla propria prole, esso tenderà a rendere massimo il proprio successo riproduttivo accoppiandosi col maggior numero di partner possibile. Così, i maschi del fagiano di monte (Tetrao tetrix), degli elefanti marini e dei coleotteri lucanidi, che non offrono alcuna cura parentale, sono più grandi rispetto alle femmine probabilmente a causa di un adattamento evolutosi in risposta alla necessità di lottare per ottenere un partner. Al contrario, le femmine di queste specie ottengono scarsi benefici materiali dall'accoppiarsi con più maschi e sono libere di concentrare la propria scelta sugli esemplari migliori, forse basandosi sulle qualità genetiche o sull'assenza di malattie trasmissibili per via sessuale. La loro fitness viene enormemente accresciuta dall'impegno nelle cure parentali. In altre parole, le femmine investono la loro energia nella prole proveniente da uno o pochi accoppiamenti, invece di impegnarsi nella ricerca di accoppiamenti multipli (Andersson e lwasa, 1996).
Relazioni tra cure parentali e selezione sessuale
Mentre le cure parentali e la selezione sessuale sono chiaramente collegate, un problema fondamentale è capire come esse siano coevolute e se siamo in grado di distinguere in modo chiaro le relazioni di causa ed effetto che legano i due fenomeni. Le teorie sulla selezione sessuale tendono a considerare i diversi tipi di cure parentali come punti di partenza, mentre le teorie riguardanti le cure parentali talora considerano un punto di partenza la selezione sessuale. Tuttavia gli esperimenti hanno dimostrato che ciascuna delle due forme di comportamento può cambiare se l'altra viene opportunamente modificata. Per esempio, nel pesce ciclide Herotilapia multispinosa entrambi i genitori proteggono la prole, ma il maschio abbandona presto la femmina e la prole se gli si mette a disposizione un numero elevato di femmine non accoppiate, dimostrando che le cure paterne sono influenzate dalle opportunità offerte dalla selezione sessuale. Quindi, tali opportunità possono imporre condizioni alle cure parentali.
Al contrario, in un'altra specie di pesci, il ghiozzetto minuto (Pomatoschistus minutus), il maschio fornisce tutte le cure senza aiuto da parte della femmina. Tuttavia, riducendo artificialmente il periodo di sviluppo degli embrioni, si dà la possibilità al maschio di liberarsi in anticipo dagli impegni parentali e di tornare a competere per altre femmine (Kvarnemo, 1994). In questo caso sono le cure parentali a porre le condizioni per la selezione sessuale. Dunque sarebbe sbagliato assumere una singola direzione di causalità nel rapporto di scambio che si crea tra selezione sessuale e cure parentali.
Resta da vedere quante specie mostrano questa flessibilità. Recentemente è stato dimostrato che nella rondine (Hirundo rustica) i maschi con code particolarmente lunghe forniscono meno cure ai loro nidiacei rispetto ai maschi con code più corte (De Lope e M0ller, 1993). Tuttavia, esperimenti in cui la lunghezza della coda del maschio era stata allungata artificialmente attaccando segmenti addizionali alle penne centrali, hanno mostrato che le femmine continuano a preferire i maschi con le code 'vere' più lunghe. Ciò suggerisce che i maschi considerati più attraenti possono aumentare il proprio successo riproduttivo riducendo le cure parentali al fine di trascorrere più tempo a corteggiare le compagne. Si tratta di un altro esempio del fatto che le cure parentali sono condizionate almeno in parte dalla selezione sessuale e che non si verifica soltanto il caso opposto. Sembra dunque logico ritenere che lo sviluppo delle risposte facoltative in entrambe le direzioni si possa estendere a tempi evolutivi lunghi, e così si può prevedere che le cure parentali e la selezione sessuale continuino a coevolvere con effetti reciproci di retroazione.
Il rapporto di scambio appena descritto tra cure parentali e selezione sessuale non implica che il sesso che si occupa della prole debba essere necessariamente sottoposto a una selezione sessuale meno intensa. l maschi di Betta splendens, una specie nota come pesce combattente, sono così aggressivi che gli acquario fili devono tenerli sempre separati da altre specie. Tuttavia i maschi forniscono tutte le cure parentali, costruiscono e custodiscono elaborati nidi assemblati con bolle d'aria aspirate dalla superficie e proteggono gli embrioni e i giovani dai predatori. Il punto è che i maschi possono occuparsi contemporaneamente di uova deposte da più di una femmina. In tal modo essi possono allevare i giovani con ritmi più intensi di quelli con cui le femmine possono produrre gruppi di uova. Inoltre, accoppiarsi con una femmina non limita la possibilità di un maschio di accoppiarsi con un'altra subito dopo. Siamo dunque in grado di spiegare perché in questa specie i maschi vanno incontro a una selezione sessuale più intensa rispetto alle femmine, nonostante forniscano tutte le cure parentali possibili. Spiegazioni analoghe possono essere utilizzate per molte altre specie di pesci, per esempio per gran parte delle popolazioni di spinarello (gasterosteidi).
Tassi potenziali di riproduzione
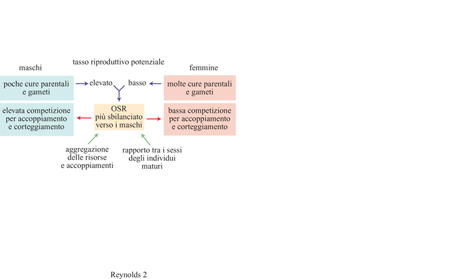
La relazione tra competizione per il partner e cure parentali è stata formalizzata in base al tasso potenziale di riproduzione di ciascun sesso, definito come la velocità massima con la quale gli individui possono produrre prole se hanno un accesso illimitato ai membri del sesso opposto (CluttonBrock e Parker, 1992). A parità di altri fattori, il sesso con il tasso potenziale di riproduzione più elevato ricaverà il maggior vantaggio dalla competizione per il partner (fig. 2).
Questa formulazione rende conto dei comportamenti osservati nelle specie precedentemente descritte, per esempio nei pesci combattenti, nei quali, sebbene i maschi provvedano a tutte le cure dopo la fecondazione, questa attività richiede meno tempo e meno energia per nidiata rispetto a quanto avviene in specie come gli uccelli, che nutrono o riscaldano la loro prole o i mammiferi che allattano i loro cuccioli. Di conseguenza, i maschi tendono a competere maggiormente rispetto alle femmine per ottenere accoppiamenti multipli.
Oltre a fornire una spiegazione per le differenze che si osservano nella competizione per il partner, i diversi tassi potenziali di riproduzione nei due sessi sono serviti anche per lo studio della scelta del partner, l'altro aspetto fondamentale della selezione sessuale (Johnstone et al., 1996). Il sesso che ha il tasso potenziale di riproduzione più basso in genere quello femminile - può permettersi di essere più selettivo nella scelta del compagno, in quanto il sesso opposto sarà raramente numericamente carente.
Consideriamo una specie come il combattente (Philomachus pugnax), un uccello limicolo in cui la femmina provvede da sola alle cure parentali. Se una femmina rifiuta i primi tre maschi che incontra in un' arena di esibizione, essa può sempre accoppiarsi con il quarto maschio e può ancora raggiungere il suo massimo riproduttivo stagionale consistente in una nidiata di quattro pulcini (forse dotati di migliore qualità genetica o forse provenienti da un maschio con una scarsa probabilità di trasmettere una malattia per via sessuale). Tuttavia, se un maschio rifiuta l'opportunità di accoppiarsi con una femmina, ciò comporterà la perdita di una nidiata per la sua fitness, in quanto il suo potenziale riproduttivo è limitato solo dal tempo di accoppiamento e dal tasso di produzione di spermatozoi e non da tre settimane di incubazione più un'altra settimana circa di cure parentali, come accade invece per la femmina.
l recenti modelli proposti dalla teoria dei giochi, basati su queste considerazioni e sulle differenze nei due sessi dei costi risultanti dalla perdita di opportunità di accoppiamento, hanno dimostrato che le femmine dovrebbero essere più propense a rifiutare i maschi qualora debbano dedicare più tempo alle cure parentali, anche se esistono differenze sostanziali tra i sessi nella variabilità della qualità del partner (Johnstone et al., 1996). In tal modo, a parità di altri fattori, i costi dovuti alla perdita di opportunità di accoppiamento, legati alle differenze nei potenziali riproduttivi di ciascun sesso, potrebbero determinare la direzione della selezione sessuale.
Rapporto funzionale tra i sessi
Un modo efficace per individuare le differenze tra i sessi nelle opportunità per l'accoppiamento è costituito dal rapporto funzionale tra i sessi od OSR (Operational Sex Ratio), definito per la prima volta da Emlen e Oring (1977) come il rapporto tra numero di femmine disponibili e numero di maschi, oggi calcolato perlopiù come numero di individui di un sesso pronti per l'accoppiamento diviso il numero totale di individui di entrambi i sessi disponibili ad accoppiarsi. Le disparità nei potenziali di riproduzione svolgono un ruolo primario in questo rapporto, poiché determinano la velocità con cui i membri di ciascun sesso saranno esclusi dal totale di partner disponibili dopo la fecondazione (v. figura 2). Inoltre, nelle specie in cui i maschi non forniscono cure parentali, il valore di OSR sarà maggiore per il sesso maschile. Al contrario, in specie come il falaropo beccolargo, un uccello limicolo in cui il maschio fornisce da solo le cure parentali per un totale di circa 40 giorni, comprendenti incubazione e allevamento della prole, mentre la femmina s'impegna per circa 12 giorni nella produzione delle uova, il valore di OSR sarà molto più grande per il sesso femminile. Valori di OSR prossimi a 0,5 sono tipici delle specie monogame in cui entrambi i sessi si dedicano alla prole e nessuno dei due abbandona il compagno.
Altre differenze tra i sessi che contribuiscono a determinare il valore di OSR sono (v. figura 2): il rapporto numerico tra i sessi e la mortalità; le risposte comportamentali al rischio di predazione; l'emigrazione e l'immigrazione; l'età in cui si raggiunge la maturità e la durata della vita riproduttiva; la distribuzione dei sessi nello spazio e nel tempo. Nelle specie in cui un sesso combatte per l'accesso all'altro sesso e mostra vistosi segnali di esibizione, come nel caso dei fagiani, il valore di OSR può essere notevolmente spostato rispetto a 0,5, poiché il sesso più competitivo sarà numericamente ridotto o vedrà ostacolate le sue attività di corteggiamento e combattimento a causa della predazione.
Ruolo dei fattori ambientali
Aggregazione temporale delle risorse e dei partner sessuali
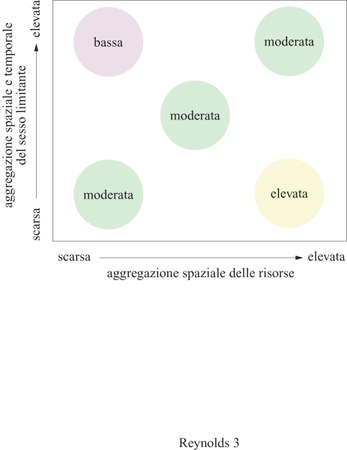
La predazione costituisce solo uno dei fattori ambientali che possono condizionare il sistema riproduttivo di una popolazione. In realtà uno degli obiettivi classici della ricerca in questo senso è stato quello di trovare una corrispondenza di un dato sistema riproduttivo con un determinato ambiente. Per esempio, tra gli anuri vi è la tendenza, nelle specie che hanno stagioni riproduttive prolungate, a subire una selezione sessuale più intensa rispetto alle specie in cui la stagione riproduttiva si conclude nel giro di pochi giorni. È stato proposto che le specie con la stagione riproduttiva prolungata siano spesso territoriali, mentre quelle con stagione riproduttiva breve (explosive breeders) mostrerebbero una competizione diretta basata sulla lotta tra maschi per l'accesso alle femmine. Dunque, la brevità delle stagioni riproduttive e il raggruppamento temporaneo di femmine (fig. 3) riducono la capacità dei singoli maschi di monopolizzare risorse e partner sessuali (Emlen e Oring, 1977).
Aggregazione spaziale delle risorse
A differenza degli effetti dovuti all'aggregazione temporale, il raggruppamento spaziale delle risorse riproduttive e degli individui appartenenti al sesso limitante (cioè con un potenziale di riproduzione inferiore) è in grado di promuovere la selezione sessuale (v. figura 3). Per esempio, tra i pinnipedi (foche e specie affini) il grado di poliginia è correlato a quello di aggregazione delle femmine durante la stagione riproduttiva. Questo fattore, a sua volta, dipende dall'habitat riproduttivo: nelle specie che si riproducono in isole remote, un maschio può monopolizzare molte femmine, mentre nelle specie che si riproducono su banchise di ghiaccio, le femmine sono più disperse e quindi più difficili da difendere. Queste differenze nella tendenza alla poliginia sono in relazione alla diversa dimensione corporea tra i sessi, con i maschi molto più grandi delle femmine nel caso di specie altamente poligine, nelle quali l'abilità nella lotta viene fortemente premiata. Per esempio, i maschi di elefante marino pesano 2300 kg, mentre le femmine solo 900 kg. La facilità con cui si può difendere una risorsa riproduttiva può spiegare anche l'esempio della specie di acaro d'acqua altamente poligina, Unionicola formosa. Questa specie si riproduce in America Settentrionale all'interno del guscio di molluschi di acqua dolce (Anodonta imbecillis) che rappresentano una risorsa discreta e difendibile in grado di sostenere molte femmine e di essere monopolizzata da singoli maschi.
Gli esempi dei pinnipedi e degli acari d'acqua dimostranv che la distribuzione delle risorse determina la dispersione delle femmine che, a sua volta, condiziona la distribuzione dei maschi. Tale ipotesi può essere saggiata modificando la dispersione delle risorse o rimuovendo alcuni individui di un sesso e valutando il conseguente effetto sull'altro sesso. Per esempio, in un esperimento sull'arvicola di Sundevall, Clethrionomys rufocanus, la distribuzione spaziale di ciascun sesso fu modificata spostando in diversi punti individui tenuti in gabbia (Ims, 1988). I maschi liberi di muoversi seguirono la dispersione delle femmine, aggregandosi ai gruppi costruiti artificialmente, invece le femmine libere di muoversi non modificarono la loro area familiare (home range) in seguito alle manipolazioni.
Ci si potrebbe aspettare che questa semplice relazione causale, che lega la distribuzione delle risorse alla dispersione delle femmine e questa alla distribuzione dei maschi, si sia evoluta soprattutto nelle specie in cui i maschi non forniscono alcun vantaggio alle femmine, come avviene in gran parte dei roditori, che tipicamente non offrono cure paterne. Tuttavia, una ricerca condotta in Finlandia su un altro roditore dello stesso genere, l'arvicola rossastra, Clethrionomys glareolus, ha mostrato preferenze attive delle femmine nei confronti di maschi socialmente dominanti anche quando la competizione tra maschi era stata sperimentalmente esclusa durante i test di scelta (Home e Ylonen, 1996). Studi effettuati su altre specie di roditori hanno dimostrato anche che la scelta femminile si baserebbe sulla familiarità e la parentela. Così, sebbene la dispersione delle risorse sia indubbiamente importante nel porre limiti al numero e alla qualità degli individui incontrati, e quindi alla probabilità di selezione sessuale, sembra esistere un ulteriore effetto dovuto alla scelta diretta dei maschi operata dalle femmine.
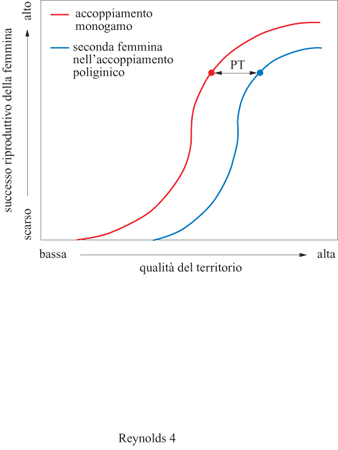
La teoria secondo cui la femmina sceglie tra i maschi che gestiscono le risorse disponibili, è stata formalizzata tramite il modello della soglia di poliginia (Orians, 1969). Quest'idea fu ispirata da alcune specie di uccelli che nidificano nelle zone paludose, come lo scricciolo di palude (Cistothorus palustris) e l'ittero alirosse (Agelaius phoeniceus). Immaginiamo una femmina di quest'ultima specie al suo arrivo in primavera in un'area palustre divisa in territori dai maschi. Alcuni maschi hanno già una femmina nei loro territori, altri devono ancora conquistare una compagna. Questa femmina appena arrivata dovrebbe sistemarsi con un maschio che ha già una compagna, condividendo le risorse territoriali con un'altra femmina, o dovrebbe piuttosto scegliere un maschio non accoppiato? Secondo il modello della soglia di poliginia, questa decisione dipenderà dalle differenze nella qualità dei territori, dal momento che la femmina in questione potrebbe riuscire ad allevare un numero maggiore di figli condividendo un territorio di alta qualità con un'altra femmina, piuttosto che sfruttando da sola un territorio di bassa qualità (fig. 4).
Nella sua semplicità, il modello della soglia di poliginia è interessante perché permette previsioni generali sul modo in cui la variazione in qualità tra i membri di un sesso e le risorse che essi gestiscono possono tradursi nello scopo principale dei sistemi riproduttivi, cioè l'ottenimento del maggior numero di partner possibile. In particolare esso prevede che una femmina che sceglie di accoppiarsi in una situazione di poliginia dovrebbe optare per un maschio in grado di garantirle un successo riproduttivo pari almeno a quello che avrebbe avuto scegliendo di restare sola con un altro maschio in un contesto riproduttivo di bassa qualità. Purtroppo nessuno degli esperimenti condotti dagli anni Settanta in poi, soprattutto sugli uccelli, è riuscito a suffragare tale modello. Esso sembrerebbe non essere valido per una serie di ragioni, tra cui il fatto che in alcune specie le femmine non hanno la possibilità di scegliere liberamente a causa di molestie da parte dei maschi, di aggressioni da parte di femmine già insediate o di una conoscenza imperfetta della qualità delle risorse al momento dell'insediamento. Così, sebbene il modello della soglia di poliginia sia un utile punto di partenza per comprendere i modelli di accoppiamento nelle situazioni in cui vengono offerte risorse, è necessario tener conto di altri elementi della biologia delle diverse specie per capire quali maschi riescono a conquistare numerose femmine.
Prevedibilità delle risorse disponibili e difesa del partner
Qualora la distribuzione nello spazio di risorse come il cibo per gli adulti e la prole sia imprevedibile, le risorse non saranno difendibili in maniera economica e ciò potrebbe determinare una riduzione degli accoppiamenti multipli col risultato di una selezione sessuale meno intensa. Queste sono le condizioni in cui si trovano specie come i falaropi (Phalaropus lobatus, P. fulicaria e Steganopus wilsoni) e il piviere tortolino (Charadrius morinellus), uccelli limicoli che nidificano nelle regioni artiche (Phalaropus), sulle cime delle montagne oltre il limite degli alberi (Charadrius) e nelle praterie dell'America Settentrionale (Steganopus). Ciascuna di queste specie può essere considerata opportunista, dato che per alimentarsi si basa su un'imprevedibile disponibilità di invertebrati, è soggetta ai capricci dello scioglimento tardivo delle nevi in primavera e, nel caso di Steganopus, a occasionali periodi di siccità. Inoltre, in queste specie le cure parentali sono fornite solo dal maschio, mentre le femmine mostrano un potenziale di riproduzione più elevato e sono quindi sottoposte a una selezione sessuale più marcata: infatti le aggressioni tra femmine sono più violente ed è questo il sesso che mostra il piumaggio più brillante. Tuttavia la selezione sessuale è limitata dall'imprevedibilità dell'abbondanza di cibo, resa ancor più forte dalla brevità della stagione riproduttiva. Perciò le femmine in queste quattro specie non sono in grado di difendere territori che assicurino il cibo o i siti di nidificazione richiesti dai maschi. Piuttosto, il loro sistema riproduttivo comprende comportamenti come la sorveglianza del partner, in cui le femmine scortano ciascun maschio frnché la deposizione è stata completata e respingono le rivali. Per questo motivo, le femmine che hanno più successo sono in grado di accoppiarsi con un massimo di due maschi per ogni stagione riproduttiva. Tale selezione sessuale ridotta è in contrasto con quanto avviene in altri uccelli limicoli che presentano cure parentali maschili, come il piro-piro macchiato, una specie nidificante in aree temperate, e la jacana spinosa (Jacana spinosa), tipica delle regioni tropicali. Questi uccelli nidificano lungo le rive di specchi di acqua dolce, che costituiscono una fonte di cibo stabile. La territorialità è dunque un'opzione e ciascuna femmina entra in competizione per attrarre il maggior numero di maschi che nidificano nell'area che essa difende. La combinazione di legami di coppia simultanei con numerosi maschi con stagioni riproduttive più lunghe determina una selezione sessuale più intensa, che in queste specie si riflette in un più accentuato dimorfismo nella taglia, ma non nel piumaggio.
Limitazione delle risorse e variabilità intraspecifica
Gli studi sulla variabilità intraspecifica dei sistemi riproduttivi hanno aiutato a chiarire il ruolo della limitazione delle risorse nel determinare le tipologie dei sistemi di accoppiamento e l'intensità della selezione sessuale.
Il ghiozzetto minuto è un piccolo pesce bentonico che si riproduce lungo le coste di gran parte d'Europa. I maschi scavano nidi sotto i gusci dei mitili o sotto le pietre e attirano una o più femmine per deporre le uova all'interno dei nidi. I maschi sorvegliano e ventilano le uova fino alla schiusa che avviene dopo 1 ÷ 3 settimane, a seconda della temperatura dell'acqua. E. Forsgren e collaboratori (1996) hanno confrontato il comportamento riproduttivo del ghiozzetto minuto in due popolazioni che differivano per disponibilità di gusci per i siti di nidificazione. Un esperimento condotto sul campo ha mostrato che, aggiungendo nidi artificiali in una popolazione del Mar Baltico in cui la densità naturale era solo di 0,2 nidi/m², tutti i 30 nuovi nidi aggiunti furono occupati dai maschi nel giro di 54 ore. Al contrario, in una popolazione della costa occidentale svedese in cui la densità naturale era di 9,6 nidi/m², solo 17 dei 30 siti artificiali furono occupati dopo 96 ore. Nell'area a bassa densità di nidi, i maschi erano significativamente più grandi delle femmine; questo dato concorda con l'osservazione di una più intensa competizione per i siti di nidificazione che può aver determinato la selezione degli individui di dimensioni corporee maggiori. Nell'area a elevata densità di nidi non fu riscontrata alcuna differenza nella dimensione corporea tra i sessi.
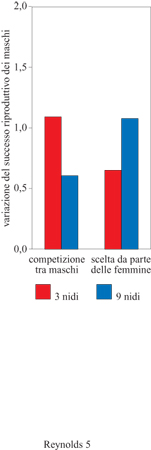
Questi risultati furono confermati da manipolazioni effettuate in laboratorio sul numero di siti di nidificazione. Quando a sei maschi e sei femmine di ghiozzo furono offerti alternativamente tre o nove siti per il nido, la variazione tra maschi nel successo riproduttivo derivante dalla competizione per il partner fu più marcata nel gruppo a cui erano stati offerti tre siti. Al contrario, la variazione nel successo dei maschi dovuta alla scelta da parte delle femmine tendeva a essere inferiore rispetto al gruppo a cui erano stati offerti nove siti (fig. 5).
Gli stessi risultati furono osservati nei pesci di entrambe le popolazioni, maladdove la disponi bilità di siti di nidificazione era già in natura più scarsa, l'effetto di questo fattore sulla scelta femminile risultò più blando. Dunque risorse riproduttive limitate possono condurre a una più intensa selezione sessuale dovuta alla competizione tra maschi, riducendo nel contempo l'importanza della scelta operata dalle femmine.
Un'elegante serie di esperimenti ha mostrato l'importanza della limitazione di cibo nei sistemi riproduttivi delle cavallette (tettigonidi). In una specie australiana i maschi forniscono alle femmine spermatofore nutrienti che le femmine mangiano mentre vengono fertilizzate fondendo le con le loro uova. D.T. Gwynne e L.W. Simmons (1990) hanno dimostrato sperimentalmente che quando il cibo era abbondante, i maschi erano capaci di produrre rapidamente queste spermatofore, di conseguenza per le femmine non era necessario competere per i maschi. Tuttavia, quando la disponibilità di cibo era limitata, pochi maschi erano capaci di offrire spermatofore, di conseguenza le femmine divenivano più competitive e si accoppiavano più spesso di quanto non facessero quando il cibo era abbondante. Così la direzione della selezione sessuale all'interno di una singola popolazione può modificarsi in base alla quantità di cibo disponibile.

Uno degli esempi più sorprendenti del ruolo svolto dalle risorse nel determinare le caratteristiche dei sistemi riproduttivi degli animali è fornito dalla passera scopaiola, Prunella modularis (fig. 6). Questi uccelli possono nidificare in coppie o in gruppi sociali che variano dalla poliandria alla poliginia a entrambi (poliginandria) (Davies 1992). N.E. Davies e collaboratori hanno dimostrato che gran parte di questa variabilità può essere attribuita alla distribuzione del cibo. Quando il cibo era limitato, l'home range delle femmine era ampliato, rendendo difficile per i maschi il compito di monopolizzare le femmine e provocando la formazione di legami di coppia poliandrici, costituiti da una femmina e da due maschi che l'aiutavano ad allevare la prole. Al contrario, quando veniva fornito cibo, le aree familiari delle femmine si riducevano e i maschi erano in grado di monopolizzare le femmine e di creare legamimonogami. La flessibilità del sistema riproduttivo della passera scopaiola mostra l'importanza delle limitazioni imposte dalla risorsa cibo e, nel contempo, fornisce un esempio paradigmatico del ruolo svolto dall'aggregazione temporale e spaziale delle risorse (v. figura 2), dal momento che Davies ha mostrato che i legami di coppia poligini tendono a verificarsi quando il cibo è abbondante e che due femmine che occupano l'area familiare di uno stesso maschio non si riproducono nello stesso periodo.
Competizione spermatica
Studi condotti in un'ampia varietà di gruppi tassonomici hanno dimostrato che le femmine spesso si accoppiano con più di un maschio durante il loro periodo fertile. Ciò porta alla competizione spermatica e alla possibilità di scelta degli spermatozoi da parte delle femmine, sia nelle specie a fecondazione interna che in quelle a fecondazione esterna (Birkhead e M0ller, 1992). In molte specie questa forma di selezione sessuale a livello gametico ha esercitato un'enorme influenza sull'evoluzione dei sistemi riproduttivi. Per esempio, a causa delle copulazioni extracoppia (vedi oltre), specie che un tempo si ritenevano monogame hanno rivelato maggiori probabilità di selezione sessuale di quanto si potesse immaginare. Gli adattamenti dovuti alla selezione sessuale comprendono un incremento del volume dell'eiaculato e della lunghezza degli spermatozoi, il comportamento di sorveglianza del partner e l'evoluzione di comportamenti alternativi nei maschi (v. anche il saggio di M. Gomendio, Competizione spermatica).
Alcuni sistemi riproduttivi insoliti
Monogamia sociale
La monogamia sociale è favorita da diverse condizioni: l) selezione sessuale limitata da brevi stagioni riproduttive e sincronia riproduttiva del sesso con il più basso potenziale di riproduzione (v. figura 2); 2) selezione sessuale limitata da risorse fortemente disperse e dunque difficili da monopolizzare (v. figura 2); impegno di entrambi i genitori nell'allevare la prole; 4) probabilità di competizione spermatica che favorisce la sorveglianza della compagna da parte dei maschi, restringendo in tal modo le opportunità di accoppiamenti multipli.
Sebbene circa il 90% delle specie di uccelli siano socialmente monogame, cioè costituite da un maschio che forma un legame di coppia con una sola femmina, tale sistema riproduttivo rappresenta un'eccezione negli altri gruppi tassonomici, compresi i mammiferi in cui l'allattamento, prerogativa del sesso femminile, concede ai maschi di gran parte delle specie più opportunità per cercare altre compagne. Negli uccelli la monogamia viene spiegata in genere con la necessità per la prole di essere alimentata da entrambi i genitori ed effettivamente è stato dimostrato che legami di coppia non monogami negli uccelli si sono evoluti più spesso in quelle specie in cui i giovani sono in grado di nutrirsi da soli, come i tetraonidi e gran parte degli uccelli limicoli.
La monogamia in certi mammiferi carnivori come il licaone (Lycaon pictus) e il lupo (Canis lupus) potrebbe essere spiegata considerando che i maschi ottengono più vantaggi dal nutrire la prole e dal fornire altre forme di cure parentali piuttosto che dall'abbandono della femmina dopo l'accoppiamento. Specie come queste, che si nutrono di grandi prede pericolose e difficili da catturare, sono selezionate per una caccia di tipo cooperativo, che porta a complesse organizzazioni sociali caratterizzate da rigide gerarchie di dominanza e dalla tendenza della coppia dominante a monopolizzare l'accoppiamento all'interno del branco. La monogamia sociale è dunque una proprietà emergente del comportamento di ricerca del cibo, che ha le sue radici nel contesto ambientale. Gli scarabei sii fidi (Nicrophorus spp.) forniscono un altro caso eccezionale di monogamia sociale (nell'ambito degli insetti) che può essere spiegato in base ai benefici provenienti dalle cure di entrambi i genitori. Essi depongono le uova nelle carcasse di vertebrati, una risorsa effimera, presente in quantità limitata e difficile da trovare. Così entrambi i membri della coppia rimangono nelle vicinanze della carcassa difendendola da altri individui finché non si sviluppano le larve. Vi è una certa flessibilità in questo sistema, dato che maschi di alcune specie tentano di attirare altre femmine sulla carcassa.

La monogamia non esclude la selezione sessuale, in quanto sono ancora favoriti gli individui che distinguono i partner in base alla loro qualità. Così molti uccelli marini sono monomorfici e presentano colorazioni vivaci; ci si aspetterebbe una scelta nell'accoppiamento sulla base di tali tratti se questi costituissero segnali di qualità parentale o genetica. Di fatto le femmine possono richiedere ai maschi l'offerta di doni nuziali prima di concedersi per l'accoppiamento (fig. 7).
Allevamento in comune
Se le risorse riproduttive sono sufficientemente scarse, la competizione potrebbe essere così intensa che gli animali subordinati potrebbero trarre vantaggio dall'unione con una coppia dominante all'interno di un gruppo. Si tratta di una forma di allevamento cooperativo in cui i giovani animali possono aiutare a difendere il territorio e forse ad allevare la prole di una coppia dominante in cambio della possibilità di ereditare il territorio nel futuro. Le ghiandaie della Florida (Aphelocoma coerulescens) costituiscono un esempio di questo adattamento (Fitzpatrick e Woolfenden, 1988). Questi uccelli formano popolazioni stabili che nidificano in boscaglie aperte con prevalenza di querce, distribuendo tutti i siti di nidificazione disponibili all'interno di territori permanenti. A una coppia nidificante si possono unire fino a sei individui che non si riproducono, in genere la prole degli anni precedenti, che aiutano la coppia ad allevare un numero di giovani leggermente superiore a quanto sarebbero in grado di fare i genitori da soli e aumentano le probabilità di sopravvivenza degli adulti. In questo modo la coppia nidificante ottiene un vantaggio e gli aiutanti aumentano leggermente la loro fitness complessiva mentre aspettano che si liberi un territorio. L'allevamento in comune che si osserva nella passera scopaiola, di cui si è già discusso, rappresenta un caso differente, in quanto i membri del gruppo che cooperano (e che sono in conflitto) non sono imparentati. Tuttavia, come avviene nelle ghiandaie, gli individui vengono a trovarsi insieme a causa di particolari fattori ambientali, nella fattispecie la distribuzione e l'abbondanza di risorse nel territorio.
Arene di esibizione comune
Nelle arene di esibizione comune (lek) i maschi difendono piccoli territori simbolici e offrono alle femmine soltanto la possibilità di accoppiarsi. Questa mancanza di istinto parentale da parte dei maschi crea un netto squilibrio nei potenziali di riproduzione dei sessi e, di conseguenza, una forte deviazione verso il sesso maschile del valore di OSR; ciò potrebbe comportare un'elevata variabilità tra maschi nel successo riproduttivo e un'intensa selezione sessuale. Il lek è un sistema riproduttivo raro e con una distribuzione irregolare nei diversi gruppi tassonomici; questa strategia si è evo Iuta almeno 14 volte negli uccelli (per esempio nel gallo forcello, nei manachini, in alcuni colibrì, in una specie di pigliamosche dei tropici), nei mammiferi (per esempio nel kob dell'Uganda, Kobus kob thomasi, nello pteropo dalla testa a martello, Hypsignathus monstrosus), in alcune specie di rane e rospi (per esempio ilidi come Ololygon rubra o il rospo calamita BuJo calamita) e nei pesci ciclidi africani (per esempio Cyrtocara eucinostomus). Le specie che adottano questo sistema riproduttivo hanno ricevuto molta attenzione poiché mostrano una selezione sessuale molto intensa. In realtà vi è scarso consenso tra i ricercatori sul perché un tale sistema si sia evoluto; tra le molte ipotesi, alcune mettono in primo piano il ruolo della scelta femminile, altre il ruolo svolto dalla competizione tra maschi (H6glund e Alatalo, 1995). Per esempio, un'analisi comparativa tra le specie di tetraonidi ha suggerito che nei casi in cui le femmine controllano aree familiari vaste che si sovrappongono (in risposta a risorse di cibo o habitat di nidificazione dispersi), i maschi tendono a esibirsi nelle aree di sovrapposizione, in cui hanno una maggiore possibilità di incontrare le femmine. Le arene possono essere localizzate in hot spots (punti caldi), aree in cui vi è un intenso traffico di femmine che visitano la zona per altri scopi. In alcune specie potrebbero essere stati favoriti i maschi che si raggruppano nelle arene consentendo alle femmine di ridurre i costi energetici legati alla ricerca del compagno e facilitando i confronti.
In contrasto con questi modelli di evoluzione del sistema dellek, basati sui vantaggi per il sesso femminile, quelli che si riferiscono al 'punto di vista' maschile suggeriscono che i lek si sarebbero formati a causa della dominanza tra maschi. Così è stato formulato il modello hot shot (colpo caldo) in alternativa al modello hot spot, sostenendo che i maschi subordinati o meno attraenti tendono ad aggregarsi intorno agli hot shot dei dominanti per intercettare le femmine. Il modello black hole (buco nero) propone invece che le arene si formino perché i maschi traggono vantaggi dalla tendenza delle femmine a essere attratte dai gruppi, come avviene negli ungulati, tra i quali è raro che le femmine operino una scelta attiva dei maschi presenti all'interno del lek.
Nel considerare l'evoluzione di questa strategia è importante ricordare che, sebbene vi possa essere alla base una singola causa, è forse più probabile che il sistema riproduttivo dei lek si sia evoluto per ragioni diverse in differenti gruppi animali. Per esempio, le femmine degli ungulati sembrano avere meno libertà di scelta tra i maschi presenti nellek rispetto a quanto si osserva negli uccelli, in quanto i maschi sono capaci di raggruppare fisicamente le femmine. Dunque le molestie da parte dei maschi, che si verificano negli ungulati e in certi uccelli come il galletto di roccia della Guyana (Rupicola rupi cola ), in cui le femmine talora vengono gettate a terra dai maschi, creano per ciascun sesso un diverso bilancio di costi e benefici derivanti dall'aggregazione, mentre in specie come il combattente, una volta che ha attratto una femmina verso la sua area di corteggiamento, il maschio china il capo in avanti e rimane immobile. Il contatto diretto con le femmine è molto ridotto, e ciò conferisce a queste ultime maggiori opportunità per la scelta del partner nell'arena. Si dovrebbe anche notare che le arene si osservano in habitat che variano da brughiere aride, nel caso del gallo della salvia (Centrocercus urophasianus), a fitte foreste, nel caso di molte specie di manachini, e che i rischi di predazione per gli individui possono essere praticamente nulli come nel caso del manachino barbabianca di Trinidad (Manacus manacus trinitatis), fino a diventare estremamente elevati nel caso del galletto di roccia della Guyana. È possibile riconoscere anche una differenza tra specie a fecondazione interna e specie a fecondazione esterna se in quest'ultimo caso le femmine mostrano maggiori possibilità di controllo. Come per l'allevamento in comune, il termine "lek" potrebbe perciò nascondere molteplici fenomeni pur identificando un singolo sistema riproduttivo.
Inversione dei ruoli nel corteggiamento
L'inversione dei ruoli nel corteggiamento (fenomeno noto anche come inversione dei ruoli sessuali) si riferisce al sesso verso cui si rivolge la selezione sessuale, non alle differenze tra i sessi nelle cure parentali. l maschi di spinarello (Gasterosteus aculeatus) offrono cure alla prole, ma questa specie mostra in genere ruoli di corteggiamento 'convenzionali' perché i maschi di solito sono il sesso più competitivo, essendo più attivi delle femmine nel corteggiamento e nella competizione per il partner.
Abbiamo incontrato esempi di specie con inversione dei ruoli nel corteggiamento nel paragrafo sulla prevedibilità delle risorse e la difesa del partner, per esempio nel caso dei falaropi, delle jacane, del piviere torto lino e dei piropiro macchiati. Queste specie hanno in comune il fatto che i maschi si dedicano da soli alle cure parentali. A causa degli elevati costi energetici e dei lunghi periodi di cura richiesti dalla prole rispetto a pesci come lo spinarello, i maschi possono prendersi cura solo di una nidiata alla volta e il rapporto funzionale tra i sessi è dunque spostato in modo tale che i maschi limitano il tasso di riproduzione delle femmine. Il risultato può essere un'intensa competizione tra femmine per i partner sessuali.
L'esempio degli uccelli limicoli dimostra che, come nel caso dei lek, sembra improbabile che l'inversione dei ruoli nel corteggiamento possa essere attribuita a un singolo fattore ambientale. Le popolazioni di jacana vanno incontro a notevoli perdite di covate a causa della predazione, ed è stato suggerito che ciò potrebbe spiegare la tendenza evolutiva dei maschi a impegnarsi nelle cure parentali, mentre le loro compagne recuperano riserve di energia e si preparano a una rapida sostituzione della covata. Tuttavia non ci sono prove che i limicoli nidificanti nelle zone temperate e artiche subiscano una predazione più marcata rispetto alle specie in cui entrambi i genitori si dedicano alla prole, e un recente studio comparativo condotto su 96 specie non ha trovato alcuna relazione generale tra cure parentali e predazione delle covate (Reynolds e Székely, 1997). È stato ipotizzato anche che, nel caso di uccelli limicoli che depongono un numero di uova molto grande rispetto alle dimensioni corporee o che depongono le uova rapidamente, potrebbero essere favoriti dalla selezione naturale i maschi che tentano di ridurre lo stress della propria partner quando depone le uova, assumendo il compito dell'incubazione. Tuttavia, i dati comparativi non confermano nemmeno questa ipotesi. Potrebbe essere che nelle jacane l'incubazione delle uova da parte dei maschi, con la conseguente inversione dei ruoli, si sia evo Iuta a causa della necessità per le femmine di deporre rapidamente covate di sostituzione in ambienti tropicali caratterizzati da elevate pressioni di predazione. Invece, nelle specie come i falaropi e il piviere tortolino, le cure parentali maschili avrebbero compensato la tendenza delle femmine ad abbandonare le covate nel momento in cui esse debbano fare i conti con imprevedibili variazioni della disponibilità degli invertebrati di cui si nutrono. Sfortunatamente le cure maschili e l'inversione dei ruoli negli uccelli sono tratti così filogeneticamente conservativi che risulta arduo effettuare confronti più rigorosi dei loro diversi percorsi evolutivi.
Nel regno animale gran parte dei casi di inversione dei ruoli nel corteggiamento si osservano in specie in cui i maschi che si dedicano alle cure parentali sono incapaci di dedicarsi a più di una nidiata in un breve lasso di tempo. Per esempio, le femmine degli emitteri belostomatidi attaccano le loro uova sulla schiena dei maschi. l maschi poi puliscono, arieggiano e proteggono le uova per tre settimane, cioè fino alla loro schiusa. l maschi non possono trasportare le covate di due femmine nello stesso tempo e, nella specie Belostoma flumineum, in cui le femmine possono produrre rapidamente una seconda covata, all'inizio della stagione lo spazio sulla schiena del maschio diventa limitante per le femmine. l ruoli nel corteggiamento non sono del tutto invertiti, ma i maschi sembrerebbero controllare la decisione finale dell'accoppiamento.
Una specie studiata in dettaglio in cui si osserva l'inversione dei ruoli nel corteggiamento è il pesce ago (Nerophis ophidion). Le femmine depongono le uova sulla superficie ventrale del maschio e il maschio trasporta gli embrioni mediamente per quattro settimane fino alla schiusa (Berglund et al., 1989). Le femmine possono produrre uova 1,8 volte più velocemente di quanto i maschi siano in grado di accudirle. Sulla base di questa differenza nel tasso potenziale di riproduzione ci si attenderebbe l'inversione dei ruoli nel corteggiamento. In accordo con questa previsione le femmine effettuano una danza di corteggiamento esibendo ai maschi vivaci colori nuziali blu.
Competizione per la riproduzione negli ermafroditi
Negli ermafroditi un individuo produce sia gameti maschili sia femminili, ma non necessariamente nello stesso momento. Per esempio gli ermafroditi sequenziali possono essere sia proteroginici, comportandosi prima come femmine, sia proterandrici, iniziando come maschi e divenendo femmine in seguito. L'inversione sessuale è favorita quando la relazione tra il successo riproduttivo e la dimensione corporea o l'età differiscono tra i sessi. Tali differenze sono previste dal modello generale del sistema riproduttivo che mette in relazione la forma e l'intensità della selezione sessuale con l'ecologia e l'impegno parentale.
Esempi di specie proteroginiche sono forniti da molti pesci noti della barriera corallina come i labridi, i pesci pappagallo (scaridi) e le cernie (epinefelidi). Inoltre, in alcune specie che mostrano il fenomeno dell'ermafroditismo, certi individui cominciano come maschi e rimangono tali per tutta la vita. Nella donzella fasciata (Thalassoma bifasciatum) i maschi nella fase terminale sviluppano un colore bluastro interrotto da una banda bianca ben delineata con bordi neri nella parte mediana del corpo. Questi maschi sono fortemente territoriali e monopolizzano i siti di deposizione collocati su affioramenti di corallo. Le femmine visitano questi maschi per la deposizione e gli embrioni pelagici in seguito vanno alla deriva allontanandosi dalla barriera. Il modello del vantaggio derivante dalla taglia corporea evidenzia come i maschi di piccole dimensioni non avrebbero possibilità di competere contro tali possessori di territori se adottassero le stesse tattiche comportamentali e se mostrassero colori sessuali marcati (Warner, 1988). Perciò i giovani pesci possono iniziare la vita come femmine, in grado di deporre le uova senza competere per i partner, oppure come maschi primari che adottano la colorazione e il comportamento delle femmine. Questi si uniscono ai gruppi di accoppiamento e partecipano a un'intensa competizione spermatica con altri individui opportunisti e con il dominante che detiene il territorio. Tipicamente le femmine più grandi cambiano sesso e, in seguito, aspettano il momento favorevole come maschi 'satelliti', ottenendo nessuno o pochi accoppiamenti, ma rimanendo pronti a inserirsi quando muore il maschio dominante in un gruppo sociale. A questo punto il successo riproduttivo come maschio dominante può essere molto più consistente di quello ottenibile come femmina. Così la direzione dell'inversione sessuale, il momento in cui avviene e quale individuo diventa maschio, possono essere spiegati dall'intensa selezione sessuale dovuta a potenziali riproduttivi differenti in un sistema in cui i maschi sono in grado di monopolizzare risorse critiche per la riproduzione.
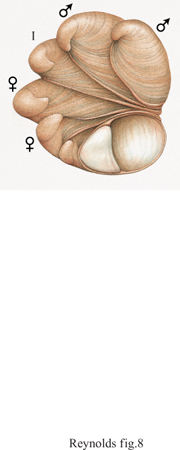
L'inversione sessuale nell'altra direzione, cioè il passaggio da maschio a femmina (proterandria), tende a verificarsi in specie in cui il successo riproduttivo del maschio non è in relazione alla sua dimensione o all'età. Gli individui tenderebbero a massimizzare il loro successo riproduttivo iniziando come maschi, poi trasformandosi in femmine e traendo così benefici per la propria fecondità legati alla taglia. Gli esempi più illuminanti di questa strategia comprendono alcune specie di pesci come i pesci pagliaccio (Amphiprion), in cui le risorse riproduttive sono ampiamente disperse e sono difese da ambedue i sessi, mentre la competizione diretta tra maschi assume una rilevanza minore. Ulteriori esempi sono costituiti dai gamberetti pandalidi, non territoriali, e dalle chiocciole a pantofola (Crepidula fornicata), che formano ammassi contenenti fino a 20 individui, di cui gran parte di quelli in basso sono femmine di grandi dimensioni, mentre i maschi, più piccoli, si trovano in alto (fig. 8). È stato dimostrato sperimentalmente che la rimozione di una femmina fa sì che nell'ammasso il maschio più vicino cresca rapidamente e diventi femmina. In questa inversione sessuale l'individuo trae vantaggio dalla maggiore fecondità legata alla taglia corporea e dal fatto di non essere più soggetto a competizione spermatica.
Negli ermafroditi simultanei come certi serranidi (Hypoplectrus) , piccoli pesci che vivono nelle barriere coralline, il tasso potenziale di riproduzione nella fase maschile è più elevato che nella fase femminile, dato che le uova sono più grandi degli spermatozoi e più costose da produrre. Nessuno dei due sessi si dedica alla cura degli embrioni. Ciò provoca un potenziale conflitto tra partner dovuto alla tendenza di entrambi a offrire solo spermatozoi. Questi pesci hanno evoluto una soluzione che soddisfa molto bene le previsioni della teoria della cooperazione. Gli individui di una coppia si rendono 'pan per focaccia' scambiandosi gameti in una sequenza ripetuta in cui prima si donano uova, poi spermatozoi. Di fatto questa soluzione basata sulla reciprocità si è evoluta indipendentemente in almeno due altri phyla: gli anellidi e alcune specie di molluschi. Nonostante l'apparente monogamia di questo sistema, ci si potrebbe ancora attendere l'azione della selezione sessuale a causa delle differenze nella qualità come partner e nella competitività esistenti tra gli individui. Inoltre, in diverse specie di pesci della barriera corallina studiate approfonditamente, si è riscontrato che l'individuo più grande della coppia continua a ottenere il successo riproduttivo più elevato, in parte perché svolge il ruolo del maschio con una frequenza sproporzionata e perché a volte riesce a evitare di comportarsi come femmina.
Conflitti tra i sessi nella riproduzione
I sistemi riproduttivi sono il risultato di conflitti. Questi conflitti si verificano tra individui dello stesso sesso e di sesso diverso anche nelle specie che formano legami di coppia prolungati. Ciò è stato dimostrato in modo evidente dagli studi di Davies (1992) sulle passere scopaiole, in cui alcuni individui ottengono un vantaggio riproduttivo a spese di altri in base al numero di uccelli presenti in un gruppo nidificante (v. figura 6).
Una femmina ottiene un ottimo successo riproduttivo quando si accoppia con due maschi, in quanto la sua prole viene nutrita da entrambi i compagni, ma non quando deve condividere le prestazioni di un singolo maschio con un'altra femmina. Al contrario, un individuo maschio ha un successo riproduttivo più elevato se si accoppia con due femmine, ma non nel caso in cui si trovi in habitat poveri in cui potrebbe essere necessario l'aiuto di un altro maschio per sfamare i nidiacei di una singola femmina. Il successo riproduttivo del maschio si riduce drasticamente quando la femmina è condivisa con un altro maschio a causa della perdita di paternità. La monogamia comporta un successo riproduttivo intermedio per entrambi i sessi. Tra i conflitti che derivano da un tale comportamento riproduttivo, ci può essere il caso di femmine che adescano un altro maschio mentre il loro primo compagno tenta di evitare che l'altro riesca a raggiungere il suo territorio e ad accoppiarsi con la femmina.
Mentre la teoria classica dei sistemi riproduttivi ha trascurato gran parte di questi conflitti tra sessi tendendo a centrare l'attenzione sul punto di vista del sesso dominante o prevalentemente poligamo (in genere quello maschile), le ricerche condotte dall'inizio degli anni Novanta hanno dimostrato che in molte specie le femmine svolgono un ruolo assai più attivo di quanto si pensasse in precedenza. Questo aspetto comprende la competizione tra femmine per i partner in specie che normalmente mostrano ruoli 'convenzionali' nel corteggiamento, come è stato osservato nel caso del gallo della salvia e dei pavoni, due specie che adottano il sistema riproduttivo del lek. Le femmine possono anche incitare i maschi a competere, come avviene negli elefanti marini, in cui le femmine emettono richiami più forti se montate da un maschio subordinato. Ricerche dettagliate sui tratti riproduttivi femminili in molti insetti e in altre specie hanno fatto ritenere che le femmine possano essere in grado di selezionare tra gli eiaculati di maschi differenti, fenomeno talora detto cryptic female choice (scelta femminile criptica).
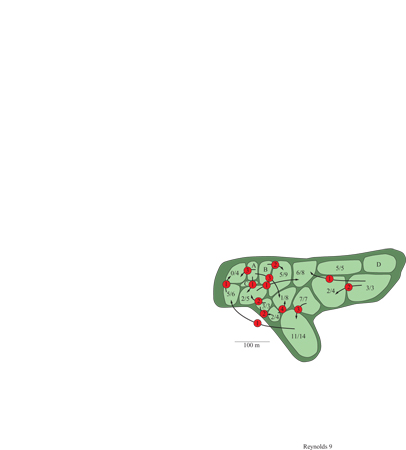
Gli accoppiamenti extracoppia nelle specie di uccelli apparentemente monogame hanno proposto la sfida più grande alla nostra comprensione dei sistemi riproduttivi. Questo fenomeno è venuto alla luce negli anni Novanta grazie alla possibilità di analizzare il DNA parentale e grazie ad altre tecnologie molecolari. Per esempio nella cinciallegra, Parus major, una specie socialmente monogama, in media il 17 ÷ 20% dei giovani sono generati da un maschio diverso dal partner putativo della femmina. La paternità extracoppia è presente anche nella rondine, con il 22% di giovani 'illegittimi', nella rondine degli alberi (Tachycineta bicolor), con percentuali del 50÷ 70%, e nel ministro (Passerina cyanea), con valori intorno al 35%. Altrettanto sorprendente è stata la scoperta che le femmine di ittero alirosse s'impegnano in così tante copulazioni al di fuori della coppia che il successo riproduttivo apparente del maschio (basato sul numero di femmine presenti nel suo territorio) ha poco a che vedere con il suo valore effettivo (fig. 9).

È ironico, o forse non casuale, osservare come questi uccelli abbiano ispirato una delle pietre miliari della teoria dei sistemi riproduttivi, il modello della soglia di poliginia (Orians, 1969), e siano stati i protagonisti principali dei tentativi di saggiarne la validità. Le femmine controllano gli accoppiamenti in questa e in molte altre specie di uccelli: dobbiamo dunque concentrarci sul loro comportamento di scelta del compagno per risolvere questo rompicapo. Sono state proposte molte soluzioni possibili (tab. I); una spiegazione è che alcuni maschi sarebbero particolarmente esperti come partner sociali a causa della loro abilità a procurarsi territori di alta qualità e a dedicarsi alla prole, ma questi caratteri potrebbero avere una debole base genetica. Le femmine potrebbero perciò scegliere tali maschi come partner sociali, ma cercarne altri per le loro qualità genetiche. Nel caso dell'ittero alirosse, tuttavia, i maschi si dedicano assai poco alla prole e, nonostante i ripetuti tentativi dei ricercatori di misurare la qualità dei loro territori, sono emerse poche correlazioni tra i fattori analizzati. Resta da capire quanto questa idea sia applicabile al caso di altri gruppi tassonomici.
Un'altra possibilità per spiegare le cause della ricerca di fecondazioni extracoppia è che la competizione tra femmine potrebbe limitare le possibilità di scelta per gli individui quando si stabiliscono in territori. Ciò significa che ad alcune femmine si offrirebbero maschi geneticamente inferiori. Le femmine potrebbero anche avere ottenuto informazioni imperfette sulla qualità dei loro compagni al momento di insediarsi, ma scoprirne più tardi i difetti. In entrambi i casi potrebbero essere favorite le femmine che cercano di rimediare a una cattiva situazione sociale e di migliorare la qualità genetica della loro prole ricorrendo a copulazioni extracoppia.
Transizioni filogenetiche nei sistemi riproduttivi
Recentemente sono stati effettuati tentativi per ricostruire le transizioni evolutive tra gli aspetti chiave dei sistemi riproduttivi degli animali. L'approccio 'storico' tenta di individuare le modificazioni di un carattere avvenute all'interno di ciascuna linea filetica (o albero filogenetico). Questo tipo di analisi fa da complemento alle osservazioni dirette e agli esperimenti riguardanti i singoli gruppi tassonomici, fornendo lo strumento principale per trarre conclusioni sulla coevoluzione e sulle sequenze evolutive degli eventi. Per esempio, B. Sillén-Tullberg e H. Temrin (1994) hanno utilizzato l'albero filogenetico degli uccelli per valutare l'ipotesi che legami di coppia di breve durata (il presupposto per la poligamia sociale) si siano evoluti con maggiore probabilità nei gruppi tassonomici in cui i giovani sono in grado di nutrirsi da soli. L'idea era che se i giovani erano capaci di nutrirsi da soli, senza che fosse necessario che i genitori portassero cibo fino al nido, come avviene in effetti nel caso dei tetraonidi, uno dei genitori sarebbe stato sufficientemente libero dagli impegni parentali e avrebbe potuto abbandonare il partner per creare nuove opportunità di accoppiamento. Quando Sillén-Tullberg e Temrin analizzarono la durata del legame di coppia e le modalità di alimentazione dei giovani nella filogenesi degli uccelli, trovarono una conferma per la loro ipotesi: giovani che si alimentavano da soli e legami di coppia brevi coesistevano nelle linee evolutive più spesso di quanto ci si potesse attendere se ciò fosse stato dovuto al caso. Dunque questa analisi lega le cure parentali e la selezione sessuale a un'altra caratteristica chiave della storia naturale degli uccelli.
Con un numero sufficiente di informazioni provenienti da gruppi tassonomici diversi è stato anche possibile ricostruire i percorsi evolutivi di alcuni cambiamenti avvenuti nei sistemi riproduttivi animali. In un altro studio sugli uccelli, T. Székely e ID. Reynolds (1995) hanno mostrato che negli uccelli limicoli, un gruppo comprendente circa 200 specie di piro-piro, pivieri e specie simili, la forma più antica di cure parentali consisteva nell'impegno di ambo i sessi a rimanere con la prole fino al momento dell'involo. Tuttavia nelle prime fasi della storia evolutiva della più importante famiglia di limicoli, gli scolopacidi (che include i piro-piro, i beccaccini, i falaropi, le jacane e specie simili), si osservava una transizione precoce verso cure fornite soprattutto dal maschio. Da allora vi è stata una tendenza ripetuta a un ritorno verso un maggior impegno da parte della femmina rispetto al maschio. L'aumento della quantità relativa di cure offerte dalla femmina corrispondeva a una riduzione della taglia corporea dei maschi rispetto alle femmine. Ciò è in linea con l'idea che, dopo il mutamento ancora non spiegato verso cure soprattutto maschili avvenuto milioni di anni fa, la selezione sessuale sui maschi abbia fatto in modo che gruppi differenti siano tornati ripetutamente a cure maschili ridotte. Ulteriori analisi delle transizioni evolutive nei sistemi riproduttivi in questi e altri gruppi tassonomici saranno possibili nel momento in cui sarà disponibile un maggior numero di informazioni filo genetiche.
Bibliografia citata
ANDERSSON, M., IWAsA, Y. (1996) Sexual selection. Trends Ecol. Evol., Il, 53-58.
BERGLUND, A., ROSENQVIST, G., SVENSSON, L (1989) Reproductive success of females limited by males in two pipefish species. Am. Nat., 133, 506-516.
BIRKHEAD, T.R., M0LLER, A.P. (1992) Sperm competition in birds, Londra, Academic Press.
CLUTTON-BROCK, T.H., PARKER, G.A. (1992) Potential reproductive rates and the operation of sexual selection. Quart. Rev. biol., 67, 437-455.
DARWIN, CH. (1871) The descent of man, and selection in relation to sex, Londra, Murray.
DAVIEs, N.B. (1992) Dunnock behaviour and sodal evolution, Oxford, Oxford University Press.
DE LOPE, F., M0LLER, A.P. (1993) Female reproductive effort depends on the degree of omamentation of their mates. EvoIution, 47, 1152-1160.
EMLEN, S.T., ORING, L.W. (1977) Ecology, sexual selection, and the evolution of animaI mating systems. Science, 197, 215-223.
FITZPATRICK, J.W., WOOLFENDEN, G.E. (1988) Components of lifetime reproductive success in the Florida scrub jay. In Reproductive Success, a c. di Clutton-Brock T.H., Chicago, University of Chicago Press.
FORSGREN, E., KVARNEMO, C., LINDSTROM, K. (1996) Mode of sexual selection determined by resource abundance in two sand goby populations. Evolution, 50, 646-654.
GWYNNE, D.T., SIMMONS, L.W. (1990) Experimental reversal of courtship roles in an insect. Nature, 346, 172-174.
HOGLUND, J., ALATALO, R.V. (1995) Leks, Princeton, Princeton University Press.
HORNE, T.J., YLONEN, H. (1996) Female bank voles (CIethrionomys glareolus) prefer dominant males; but what if there is no choice? Behav. Ecol. Sociobiol., 38, 401-405.
IMS, R.A. (1988) Spatial clumping of sexually receptive females induces space sharing among male voles. Nature, 335, 541-543.
JOHNSTONE, R.A., REYNOLDS, J.D., DEuTscH, J.C. (1996) MutuaI mate choice and sex differences in choosiness. Evolution, 50, 1382-1391.
KVARNEMO, C. (1994) Temperature differentially affects male and female reproductive rates in the sand goby: consequences for the operational sex ratio. Proc. R. Soc. Lond. Biol. Sci. 256, 151-156.
ORIANS, G.H. (1969) On the evolution of mating systems in birds and mammals. Am. Nat., 103, 589-603.
REYNOLDS, J.D. (1996) AnimaI breeding systems. Trends Ecol., Evol., Il, 68-72.
REYNOLDS, J.D., SZÉKELY, T. (1997) The evolution of parental care in shorebirds: life-histories, ecology and sexual selection. Behav. Ecol., 8, 126-134.
SILLÉN-TULLBERG, B., TEMRIN, H. (1994) A phylogenetic analysis of the relationship between polygamy, pair bond length, and the characteristics of young in birds. In PhyIogenetics and Ecology, a c. di Vane-Wright D., Eggleton P., Londra, Academic Press.
SZÉKELY, T., REYNOLDS, J.D. (1995) Evolutionary transitions in parental care in shorebirds. Proc. R. Soc. Lond. Biol. Sci. 262, 57-64.
WARNER, R.R. (1988) Sex change and the size-advantage mode!. Trends Ecol. Evol., 3, 133-136.