
Storie naturali
Storie naturali
La diversità della vita sulla Terra è anche una diversità di storie naturali. Le piante superiori e gli animali raggiungono l'età della maturità in un tempo che varia da poche ore a un secolo, differiscono in dimensioni, da quelle di microscopici rotiferi e nematodi a quelle di sequoie e balenottere azzurre, in fecondità, dalla 'manciata' di prole generata da esseri umani, albatros e scarabei ai miliardi di discendenti prodotti da ostriche e orchidee, infine in durata di vita da ore a secoli. Quali sono le cause evolutive che hanno prodotto questa diversità? La teoria delle storie naturali spiega che le condizioni dei caratteri delle storie naturali sono determinate dall'interazione tra tassi di mortalità estrinseca e conflitti intrinseci. Tuttavia ciò lascia aperta la questione del perché i conflitti assumano lo forma che osserviamo. La risposta a questa domanda è stata ricercata nell'origine e nelle conseguenze delle costrizioni filogenetiche che differiscono per le linee evolutive. Senza una visione che consideri gli elementi filogenetici, lo teoria delle storie naturali non è in grado di rendere pienamente conto della diversità dei cicli vitali osservata sulla Terra.
L'evoluzione delle storie naturali come risultato dell'interazione di più fattori
Perché raggiungere la maturità a una certa età e con certe dimensioni, non prima né dopo, né più grandi né più piccoli? Perché produrre una prole più o meno numerosa? Perché avere una vita più o meno lunga? È inevitabile l'invecchiamento? Queste domande permettono di determinare il campo dell'evoluzione delle storie naturali e questo saggio offre una traccia per poter dare una risposta. L'evoluzione delle storie naturali condivide con la selezione sessuale e la selezione per consanguinei i requisiti per trasformare l'evoluzione in una teoria che sia in grado di formulare con successo previsioni sui fenotipi. Questo campo di ricerca deve il suo successo al fatto di essersi focalizzato sull'impatto che pressioni selettive, legate all'età e alle dimensioni, hanno sull'evoluzione dei tassi di fecondità e di mortalità, anch'essi legati all'età e alle dimensioni, in relazione ai tradeoff (conflitti risolvibili con compromessi).
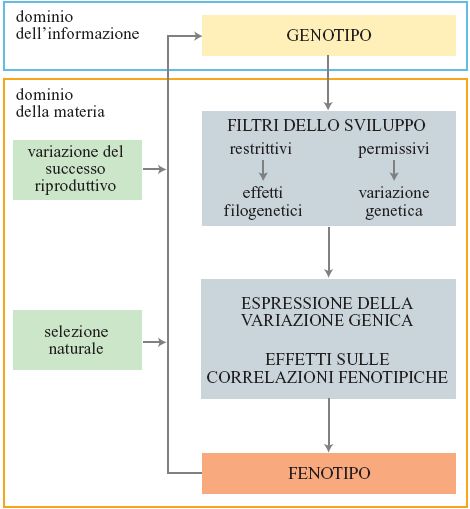
Gli effetti combinati di quattro fattori - demografia, genetica dei caratteri quantitativi, tradeoff e limiti filogenetici - sono alla base della diversità delle storie naturali. Le cause della diversità prendono due direzioni: la prima parte dalla variazione a livello genico e, attraverso l'espressione dei geni durante lo sviluppo, la morfologia e la fisiologia, arriva fino al fenotipo; la seconda, di ritorno, parte dalla variazione tra fenotipi in fecondità e sopravvivenza (selezione naturale) fino a modificazioni nelle frequenze geniche (fig. 1). Una rappresentazione grossolana del processo che include gran parte degli elementi essenziali vede l'evoluzione delle storie naturali come un'interazione tra tassi di mortalità estrinseci, tradeoff intrinseci e vincoli filo genetici che limitano la variazione dei caratteri.
La selezione naturale agisce quando gli individui di una popolazione differiscono per ereditari età nella possibilità di sopravvivenza e nella fecondità. Il primo fattore determinante, quello demografico, è importante in quanto sopravvivenza e fecondità variano con età e dimensioni. La demografia congiunge la variabilità, specifica per età e per dimensioni, nella sopravvivenza e nella fecondità alla variabilità nella fitness (idoneità evolutiva) ed esprime inoltre quanto sia forte la selezione naturale su differenti caratteri. Tuttavia, essa non dice nulla su come una popolazione risponderà a breve termine alla selezione; per questo abbiamo bisogno di sapere quanta variabilità genetica è presente e fino a che punto i caratteri sottoposti a selezione siano subordinati a tradeoff.
La maggior parte dei caratteri delle storie naturali è influenzata da molti geni che esercitano piccoli effetti. Il loro effetto combinato può essere stimato con i metodi della genetica quantitativa, che costituiscono lo strumento appropriato per analizzare i fattori genetici dei caratteri ecologicamente più importanti, il secondo fattore determinante. Sviluppatasi per l'allevamento di piante e animali, la genetica quantitativa può essere estesa a popolazioni naturali se si includono gli effetti della plasticità fenotipica, la quale ha due importanti ruoli nell'evoluzione. Per prima cosa, modificando le relazioni dei caratteri con la fitness, la plasticità fenotipica modifica le pressioni selettive sui caratteri stessi attraverso gli ambienti. In secondo luogo, cambiando l'espressione della variabilità genetica, essa modifica la risposta genetica alla selezione attraverso gli ambienti.
I tradeoff sono il terzo elemento determinante. Un tradeoff si verifica quando un aumento nella fitness, ottenuto con una modifica in un carattere, è associato a una diminuzione nella fitness di un altro carattere. La direzione che l'evoluzione della storia naturale prenderà in risposta a una nuova pressione selettiva dipende dalla forza relativa dei tradeoff presenti. Tra i tradeoff più spesso incontrati vi sono quello tra il numero di figli prodotti e il loro tasso di sopravvivenza da giovani e quello tra l'investimento riproduttivo e la sopravvivenza da adulti.
Il quarto e ultimo elemento determinante può essere inferito dalla biologia comparata. I paragoni tra i taxa di ordine superiore fanno sorgere quesiti di notevole impatto. Gli organismi che ritardano la maturità vivono più a lungo? I mammiferi con figliate più numerose hanno prole più piccola nel caso in cui entrambi i caratteri sono adattati al peso della madre? A tali interrogativi si può rispondere utilizzando metodi che tengano conto degli effetti della dimensione corporea e del taxon di appartenenza rispetto ai tipi di variazione osservati. Tali metodi permettono di chiarire quanto di certe caratteristiche sia da attribuire alla storia e alla filogenesi e quanto a processi microevolutivi che hanno operato all'interno della popolazione locale nel recente passato.
Esaminiamo ora l'applicazione di alcuni di questi metodi ai più importanti caratteri delle storie naturali.
L'età e le dimensioni alla maturità come caratteri cruciali
Il raggiungimento della maturità divide la vita in una fase preparatoria e una di realizzazione. L'età in cui viene raggiunta la maturità è un punto cruciale per la selezione naturale. La fitness spesso è più sensibile a mutamenti in questo carattere che in qualsiasi altro. Col raggiungimento della maturità, la pressione selettiva cambia drasticamente; infatti, prima della maturità l'invecchiamento non può evolvere, ma inizia soltanto dopo.
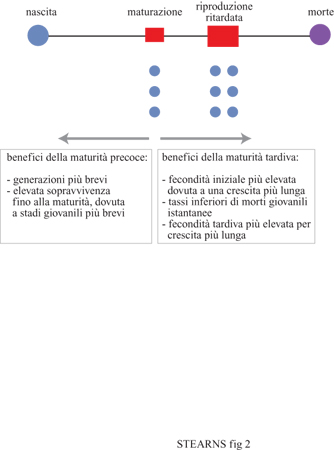
Età e dimensioni alla maturità si modificano evolutivamente quando la pressione demografica verso l'anticipazione della maturità è bilanciata dai benefici di una maturità ritardata (fig. 2). Secondo J. Kozlowski (1992) "Gli organismi piccoli in genere sono tali non perché essere piccoli migliori la fecondità o abbassi la mortalità. Essi sono piccoli perché per diventare grandi ci vuole tempo e con un'alta mortalità l'investimento nella crescita non sarebbe mai compensato da un'aumentata fecondità".
Il principale beneficio di una maturità precoce consiste nel fatto che uno sviluppo rapido aumenta la probabilità di sopravvivenza per riprodursi. Gli organismi che raggiungono prima la maturità hanno anche una fitness maggiore, in quanto la loro prole nasce prima e comincia a sua volta a riprodursi prima. l costi di una maturità precoce corrispondono ai benefici di una maturità ritardata; esaminiamo i due più importanti. Innanzi tutto, se ritardare la maturità permette una crescita ulteriore e se la fecondità aumenta con le dimensioni, la maturità ritardata porta a una maggiore fecondità. Per questa ragione la maturità è ritardata fino al punto in cui la fitness ottenuta attraverso un'aumentata fecondità è bilanciata dalla fitness perduta attraverso un tempo di generazione più lungo e una minore sopravvivenza alla maturità. In secondo luogo, se ritardare la maturità migliora la qualità della prole o delle cure parentali, allora migliora anche i tassi giornalieri di sopravvivenza della prole. Per questa ragione la maturità è ritardata fino al punto in cui la fitness ottenuta tramite l'aumento del tasso di sopravvivenza giovanile, dovuto alla maggiore età della madre al primo parto, bilancia sia la diminuzione di fitness causata da un tempo di generazione più lungo, sia quella porzione di sopravvivenza alla maturità attribuibile semplicemente al ritardo e non a una variazione nel tasso giornaliero di sopravvivenza giovanile.
Il fatto che età e dimensioni alla maturità varino tra specie molto affini, tra popolazioni della stessa specie e tra differenti individui della stessa popolazione suggerisce che questi caratteri sono in grado di rispondere rapidamente alla selezione naturale. L'età alla maturità risponde alla selezione artificiale nelle tarme della farina e nei moscerini della frutta; la variazione genetica per età e dimensioni alla maturità è stata misurata anche in molte altre specie. Tutto ciò suggerisce che entrambi questi caratteri possano essere modulati dalla selezione naturale secondo le condizioni locali nell'ambito della stessa popolazione. D'altra parte, il fatto che la dimensione corporea e la filo genesi influiscano fortemente su età e dimensioni alla maturità suggerisce che esistano importanti limiti imposti a tali caratteri dalla storia e dal disegno dell'evoluzione.
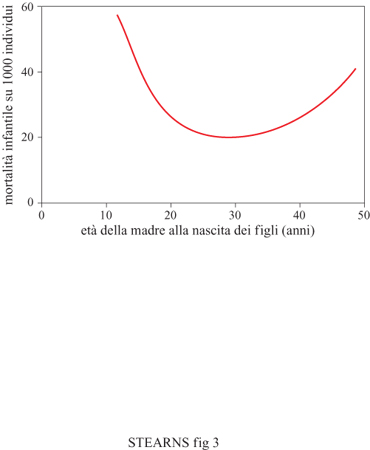
Il principio di ottimizzazione implica che il valore di un carattere sia determinato da un equilibrio bilanciato di costi e benefici per la fitness. Possiamo essere certi che un costo della maturità precoce, cioè il tasso di mortalità istantanea della prole, aumenta rapidamente quando le femmine raggiungono l'età matura prima. La capacità di femmine più giovani e più piccole di produrre prole in grado di sopravvivere deve diminuire con l'età; a un certo punto la femmina stessa sarà così piccola e poco sviluppata da non poter produrre prole (fig. 3).
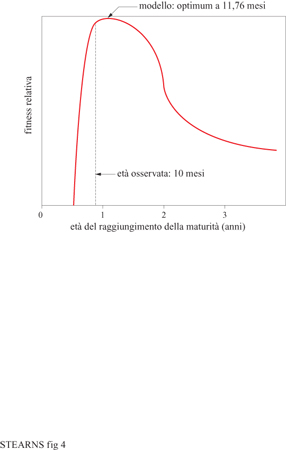
Il più importante beneficio di una maturità tardiva è che il tempo addizionale per lo sviluppo permette all'organismo di crescere di più in dimensioni e di guadagnare in fecondità. La fecondità aumenta con le dimensioni nei pesci, negli anfibi, nei rettili, in molti invertebrati e nella maggior parte delle piante superiori. Di conseguenza, una maturità precoce porta a tempi di generazione più brevi e a un periodo più breve di esposizione a mortalità prima della maturità; una maturità tardiva comporta invece aumentata fecondità e tassi inferiori di mortalità giovanile. L'optimum dovrebbe esistere a un'età intermedia. Il difficile compito dei ricercatori è stimare i costi e i relativi benefici, elaborare modelli delle loro interazioni, quindi predire l'optimum e confrontarlo con i dati (fig. 4). l modelli confrontati con dati provenienti da mosche, lucertole, salamandre e pesci hanno dato buoni risultati spiegando, con percentuali comprese tra l'80 e l'88%, la variazione di età alla maturità nelle popolazioni naturali (Bell, 1980; Roff, 1981; Stearns e Koella, 1986; Kozlowski, 1992). L'età alla maturità sembrerebbe in genere collocarsi in un optimum intermedio.
La diversità nell'investimento riproduttivo
Il secondo carattere principale del ciclo vitale è la programmazione specie-specifica dell'investimento riproduttivo. Quanta prole dovrà essere prodotta per ciascun tentativo di riproduzione? Molti organismi condividono il record minimo di un solo figlio. Il singolo figlio di una balenottera azzurra alla nascita è grande quanto un esemplare adulto di elefante africano e raggiunge la metà del suo peso da adulto nei primi sei mesi di vita. Tuttavia, le balene neonate non sono poi così grandi se paragonate alle loro madri. Tra i mammiferi, i pipistrelli hanno la prole più grande in proporzione alle dimensioni corporee e il record spetta alla specie Pipistrellus pipistrellus. Questa specie partorisce gemelli il cui peso totale alla nascita è pari al 50% del peso della madre dopo che ha partorito; e quest'ultima deve volare e alimentarsi durante la gestazione. Il kiwi, un uccello di dimensioni pari a un grosso pollo domestico, depone l'uovo più grande per le sue dimensioni, cinque volte più grande del più voluminoso uovo di gallina domestica selezionata per produrre uova di grosse dimensioni. Il suo investimento riproduttivo, pari al 20% del peso, appare tuttavia modesto rispetto a quello di Dermophis mexicanus, un anfibio tropicale che genera una prole che pesa fino al 65% del peso della madre.
La specie che presenta il più basso potenziale riproduttivo nel corso della vita non è un grosso mammifero né un uccello - elefanti, balene, specie umana e albatros hanno tutti un potenziale riproduttivo di 10÷20 figli - ma un silfide, che può avere solo una stagione riproduttiva in cui depone meno di 10, o forse 5, uova. All'altro lato della scala molti organismi producono prole piccola e numerosa. l minuscoli semi di orchidea ammontano da centinaia di milioni fino a miliardi. l grandi molluschi bivalvi e alcuni grandi pesci (storione comune, pesce gatto, merluzzo atlantico) liberano centinaia di milioni di minuscole uova per ogni deposizione. Il record mondiale di mortalità giovanile è detenuto probabilmente da una specie di orchidea, i cui semi, che pesano meno di un microgrammo, hanno una probabilità di sopravvivenza alla riproduzione pari a circa l caso su 1 miliardo.
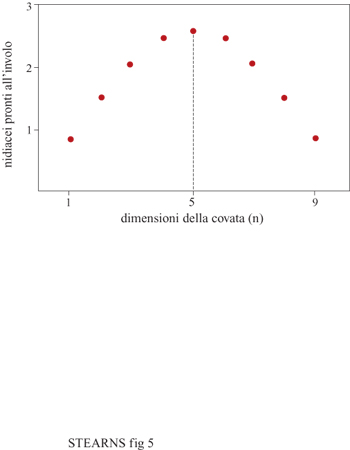
L'analisi dell'evoluzione della numerosità della prole iniziò con l'ornitologo inglese D. Lack il quale suggerì che le specie di uccelli a prole inetta producono una covata con un numero di uova tale da far involare più prole (covata di Lack). Le moderne teorie analizzano gli effetti che causano deviazioni dalla covata di Lack. Tradeoff con altri caratteri parentali e della prole, variazioni temporali nella dimensione ottimale della covata e conflitti genitori-prole sono fattori che portano a una riduzione della covata di Lack. Invece, sia l'opportunità di aborto selettivo, sia la potenzialità di ridurre flessibilmente le nidiate per modulare la dimensione della covata rispetto a livelli non predicibili di risorse sono fattori che portano a un aumento della covata di Lack (fig. 5).
La dimensione della covata può anche essere limitata dalla filo genesi; in tal caso non è possibile molta variabilità. Tutti gli uccelli dell'ordine dei procellariformi depongono un solo uovo e alcune specie lo fanno solo in certi anni. Questi uccelli, che si nutrono soprattutto in mare, differiscono nella distanza dei tragitti compiuti per alimentarsi, nelle dimensioni corporee e nell'età alla maturità. La specie più grande dell'ordine, l'albatros urlatore, impiega fino a trentatré giorni per effettuare un singolo volo di foraggiamento, coprendo distanze fino a 15.000 km. In tal caso non vi è da stupirsi che la dimensione di covata consista di un solo uovo, dato che il singolo nidiaceo, dotato di caratteristiche fisiologiche che lo rendono pronto al digiuno prolungato, deve attendere fino a un mese per un pasto; questi uccelli possono vivere 50÷60 anni e deporre un totale di 20÷ 25 uova nel corso della vita. Tuttavia, non è chiaro perché una dimensione di covata di un solo uovo sia la soluzione migliore anche per le specie più piccole del gruppo, che si alimentano a distanze inferiori. Esse sembrerebbero essere limitate dalla filo genesi piuttosto che essersi adattate alle loro particolari condizioni ecologiche.
Il modello dello sforzo riproduttivo
Il punto centrale della teoria delle storie naturali consta di modelli che riguardano l'ottimizzazione dello sforzo riproduttivo rispetto all'intero corso della vita prendendo in considerazione i costi della riproduzione. Questo tipo di approccio è detto modello dello sforzo riproduttivo (Williams, 1966; Charlesworth, 1973; Schaffer, 1974; Leon, 1976; Michod, 1979; Kozlowski e Uchmanski, 1987). Le sue previsioni sono confermate da esperimenti effettuati sui gheppi in Olanda (Daan et al., 1990), sui guppy a Trinidad (Reznick et al., 1990) e sui moscerini della frutta in laboratorio (Stearns et al., 1996).
Lo studio sui gheppi ha fornito dati di insolitamente buona qualità che si sono conformati alla teoria fm nei dettagli. L'idea alla base dello studio fu suggerita da G. Williams; egli evidenziò che la fitness dei genitori consiste di due componenti: il valore riproduttivo della prole che essi stanno producendo in quel momento sommato al loro valore riproduttivo residuo (cioè a tutti i loro successivi contributi alla fitness).
Per un uccello il valore riproduttivo è pari al valore riproduttivo di un uovo moltiplicato per la sua dimensione di covata; il valore riproduttivo di un uovo equivale alla somma data dalla probabilità che esso divenga un uccello che sopravvive fino al momento di riprodursi una volta moltiplicata per la dimensione della sua prima covata più la probabilità che esso sopravviva fino a riprodursi due volte moltiplicata per la dimensione della sua seconda covata e così via per tutte le possibili covate. Il valore riproduttivo residuo di un genitore equivale alla probabilità di sopravvivere fino a riprodursi un'altra volta moltiplicata per la quantità attesa di prole più la probabilità di sopravvivere fino a riprodursi un'altra volta moltiplicata per la quantità attesa di prole e così via fino alla morte.
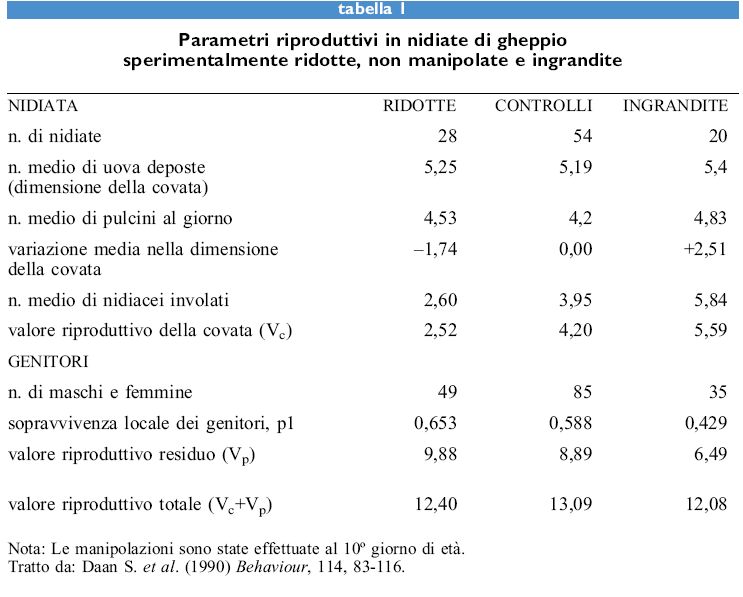
S. Daan e collaboratori hanno calcolato il valore riproduttivo della covata e il valore riproduttivo residuo dei genitori in covate sperimentalmente ridotte e aumentate nel loro numero di uova. Se i gheppi bilanciassero correttamente costi e benefici dell'investimento riproduttivo, il valore riproduttivo totale delle covate realmente deposte dovrebbe essere più grande sia di quello delle covate ridotte sia di quello delle covate incrementate. Questi uccelli sono stati studiati presso il Mare del Nord nei Paesi Bassi dove la sopravvivenza è buona e le covate sono numerose; tuttavia la probabilità di sopravvivere fino a uno, due o tre anni di età dipende dalla data in cui le uova sono deposte e così avviene per la probabilità di successo riproduttivo. Gli omitologi olandesi, quando tennero conto di questi fattori, trovarono che la dimensione di covata effettivamente deposta era quella che possedeva il più alto valore riproduttivo. Questo risultato è di notevole importanza, in quanto la selezione degli effetti da misurare era stata effettuata vari anni prima del calcolo del valore riproduttivo ed era stata misurata la maggior parte dei possibili costi della riproduzione, inclusi quelli sorti solo nella generazione successiva (tab. I).
Un'altra notevole conferma dei modelli di sforzo riproduttivo è stata ottenuta tramite esperimenti condotti in natura nel guppy, Poecilia reticulata, effettuati da D.N. Reznick e collaboratori. l guppy sono piccoli pesci vivipari che vivono a Trinidad in corsi d'acqua poco profondi (e anche negli acquari di casa di tutto il mondo). In alcuni luoghi il loro principale predatore è un pesce ciclide, Crenicichla alta, che può predare i guppy grandi e sessualmente maturi, causando un elevato grado di mortalità negli individui di tutte le dimensioni. In altri luoghi il loro principale predatore è il pesce ciprinodontide Rivulus hartii che preda per lo più guppy piccoli e giovani, causando un grado inferiore di mortalità in individui di tutte le dimensioni. Prima che gli esperimenti iniziassero, Reznick trovò che i guppy che vivono nei luoghi dove il principale predatore è C. alta raggiungono prima la maturità, compiono uno sforzo riproduttivo maggiore e hanno una prole più numerosa e più piccola rispetto ai guppy predati da R. hartii. Le differenze erano ereditabili e corrispondevano alle previsioni dei modelli di sforzo riproduttivo.
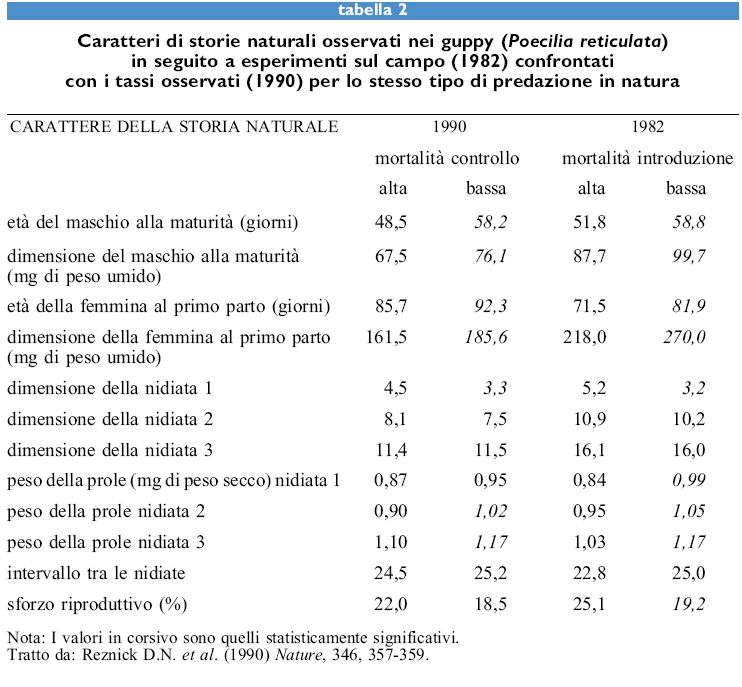
Per dimostrare che la fenomenologia osservata era causata dalla predazione, Reznick e collaboratori manipolarono le popolazioni naturali cambiando i predatori per aumentare i tassi di mortalità giovanile e diminuire quelli di mortalità adulta. Ciò modificò il tasso di mortalità in ciascuna classe di età. Dopo 11 anni, cioè dopo 30÷60 generazioni, fu osservata una significativa evoluzione della storia naturale nella direzione predetta (tab. 2).
Infatti, prima che la manipolazione iniziasse, nel 1976, quando la mortalità negli adulti era relativamente alta, sia i maschi che le femmine maturavano precocemente ed erano più piccoli; inoltre le femmine avevano prime nidiate più numerose, prole più piccola, un intervallo tra le nidiate più breve e sostenevano uno sforzo riproduttivo più elevato. Dopo 11 anni i guppy avevano evoluto gran parte delle differenze osservate in popolazioni che avevano subito pressioni selettive simili per un periodo di tempo molto più lungo. L'età e la dimensione alla maturità, la dimensione della prima nidiata, lo sforzo riproduttivo precoce e la dimensione della prole nelle prime due nidiate erano già cambiati e corrispondevano qualitativamente e quantitativamente alle differenze trovate nelle popolazioni non manipolate. La modificazione nella intensità dei tassi di mortalità, provocata da una manipolazione della pressione predatoria, aveva condotto a un rapido cambio evolutivo dei tratti di storia naturale nella direzione prevista.
La rapidità del cambiamento è sorprendente. Assumendo che le popolazioni non manipolate fossero all'equilibrio evolutivo, il raggiungimento dell'equilibrio in seguito a un'importante perturbazione ha impiegato appena 30÷60 generazioni, cioè Il anni, per gran parte dei caratteri misurati. l caratteri che erano cambiati di più erano quelli che si osservavano precocemente nel corso della vita, confermando un assunto basilare della teoria evolutiva dell'invecchiamento: la selezione sulle prestazioni riproduttive che avvengono più precocemente nel corso della vita è più forte di quella sulle prestazioni riproduttive successive.
La terza conferma dei modelli sullo sforzo riproduttivo è stata ottenuta nei moscerini della frutta con esperimenti sull'evoluzione condotti in laboratorio (Stearns et al., 1996). Il primo esperimento consisteva nel mantenere una mortalità costantemente elevata degli adulti; il secondo nel mantenere una mortalità costantemente bassa. Per assicurare che gli effetti degli esperimenti fossero dovuti a differenze nella mortalità e non nella densità, sia la densità degli adulti sia quella delle larve sono state mantenute uguali in tutti gli esperimenti. L'evoluzione dello sforzo riproduttivo e di età e dimensione alla maturità è stata rapida e ha confermato il modello dello sforzo riproduttivo: in condizioni di elevata mortalità i moscerini schiudevano prima, aumentavano la fecondità precocemente, la diminuivano in seguito e avevano una vita più breve. La velocità a cui cambiavano i caratteri corrispondeva all'intensità delle pressioni selettive previste dal modello dello sforzo riproduttivo. l tradeoff implicati - le relazioni tra età e dimensione alla schiusa e tra dimensione alla schiusa e fecondità - avevano anch' essi subito un'evoluzione. Dunque, quando una modificazione evolutiva comporta costi, si presentano ulteriori modificazioni per ridurli. Dato che l'unica differenza tra i due esperimenti era il livello della mortalità degli adulti, questo studio ha stabilito che differenze in tassi di mortalità estrinseci, interagendo con tradeoff intrinseci, costituiscono una condizione sufficiente per l'evoluzione di caratteri legati alle storie naturali.
La durata della vita e l'invecchiamento
Il terzo importante carattere della storia naturale è la durata della vita riproduttiva, che varia grandemente a seconda delle specie.
La distribuzione della durata della vita negli animali varia da circa una settimana nei rotiferi a circa un secolo in alcune tartarughe, primati, bivalvi e pesci di grandi dimensioni. Per gran parte degli animali la durata massima della vita è inferiore ai 30 anni, ma animali longevi, con una durata della vita dell'ordine di un secolo, si sono evoluti indipendentemente tra i bivalvi, i pesci, i rettili e i mammiferi. Gli invertebrati che vivono più a lungo sono gli anemoni di mare, le aragoste e i bivalvi; alcuni bivalvi possono vivere fino a 220 anni. l meno longevi sono i rotiferi, gli insetti e certi crostacei; alcuni vivono meno di una settimana. Il record massimo di durata della vita in cattività tra i mammiferi che vivono negli zoo di tutto il mondo è detenuto dall'elefante africano, 57 anni, insieme al cavallo domestico; al secondo posto troviamo l'echidna, con i suoi 50 anni di vita. Alcuni esseri umani vivono fino a 120 anni. La lunghezza della vita nelle piante varia da settimane fino a molti secoli. Tutto ciò necessita di una spIegazione.
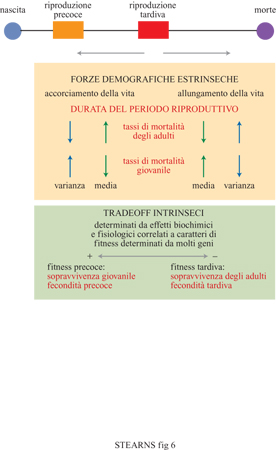
La teoria della storia naturale considera l'evoluzione della durata della vita riproduttiva come un equilibrio tra la selezione per aumentare il numero degli eventi riproduttivi nel corso della vita e gli effetti che aumentano le cause intrinseche di mortalità con l'avanzare dell'età. La prima allunga la vita, i secondi la accorciano. Le pressioni selettive età-specifiche possono regolare la lunghezza della vita verso un optimum intermedio attraverso l'interazione della selezione con i tradeoff intrinseci dell'organismo, indipendentemente dall'invecchiamento. Le pressioni selettive che allungano la vita diminuiscono il valore riproduttivo della prole e aumentano il valore riproduttivo degli adulti. Esse comprendono tassi inferiori di mortalità in età adulta, tassi più elevati di mortalità in età giovanile e un'aumentata variabilità nei tassi di mortalità giovanile tra un evento riproduttivo e il successivo (fig. 6).
L'analisi delle pressioni selettive che allungano la durata della vita riproduttiva inizia con la distinzione tra semelparità (un unico evento riproduttivo nella vita) e iteroparità (più di un evento riproduttivo nella vita). Tutti i modelli concordano nel fatto che, paragonando tra loro forme molto affini semelpare e iteropare con storie naturali simili per altri aspetti (età e dimensioni alla maturità, condizioni fisiologiche), le forme semelpare dovrebbero avere una fecondità maggiore; questo è ciò che sembra verificarsi effettivamente. Le distribuzioni delle prestazioni riproduttive delle piante semelpare sono da 1,5 a 10 volte più grandi di quelle delle loro parenti iteropare.
Dato che la semelparità dovrebbe evolvere più facilmente nelle specie a maturazione precoce piuttosto che in quelle a maturazione tardiva, le piante a rosetta semelpare a maturazione tardiva delle regioni tropicali alpine e subalpine e del deserto rivestono particolare interesse. T.P. Young (1990) eseguì un esemplare studio demografico a lungo termine su un certo tipo di gruppo, le lobelie del Monte Kenya, dove la specie semelpara Lobelia telekii coesiste con un'altra lobelia strettamente imparentata, la specie iteropara Lobelia keniensis. La specie iteropara L. keniensis fiorisce solo una volta ogni 7 o 14 anni. Per prevedere dove le piante sarebbero state semelpare e dove sarebbero state iteropare, Y oung applicò un modello sviluppato da E. Charnove W. Shaffer che indicava al di sotto di quali tassi di mortalità ci si sarebbe aspettati l'iteroparità o la semelparità. l dati demografici a lungo termine erano consistenti con le previsioni del modello.
Lo studio di Young sulla lobelia del Monte Kenya suggerisce che la durata della vita riproduttiva è adattata alle condizioni locali. Tuttavia gli effetti di non adattamento dell'invecchiamento si sovrappongono al modello adattativo determinato dalla distribuzione ottimale delle risorse per il mantenimento, la crescita e la riproduzione. Perché molti organismi diventano più suscettibili alla morte quando invecchiano? Perché non dovrebbero vivere per sempre se evitassero eventi accidentali e violenti?
Gli effetti dell'invecchiamento aumentano il tasso di mortalità e diminuiscono il tasso di fertilità in tarda età oltre i livelli previsti dalla distribuzione ottimale. La migliore misura dell'invecchiamento è quella che combina entrambi gli effetti: in una popolazione che invecchia il prodotto della sopravvivenza età-specifica per il grado di fecondità decresce con l'età. Una condizione necessaria per l'evoluzione dell'invecchiamento è che gli organismi realizzino una separazione tra linea somatica e linea germinale, dato che la forza della selezione declina con l'età della linea somatica e non con l'età della linea germinale. Comunque la divisione tra linea germinale e linea somatica non riguarda necessariamente lo stadio dello sviluppo; può anche essere raggiunta dal ritorno allo stadio di singola cellula nel ciclo vitale quando la singola cellula diventa in effetti la linea germinale.
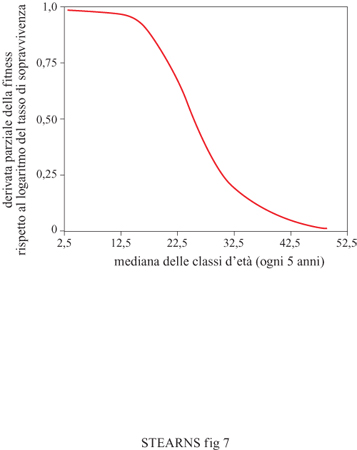
A. Weismann nel 19° secolo notò che l'invecchiamento è una proprietà della linea somatica, mentre R. Fisher e IB.S. Haldane sapevano già negli anni Trenta che la forza della selezione declina con l'età. W.D. Hamilton (1966) e B. Charlesworth (1973) hanno fornito una chiara dimostrazione matematica del modo in cui le pressioni selettive sul tasso di sopravvivenza diminuiscono con l'età e come lo fanno a differenti livelli in organismi con differenti storie naturali. Questa è la prima idea chiave nell'evoluzione dell'invecchiamento. Negli uomini che vivono in paesi industrializzati, la selezione per aumentare il tasso di sopravvivenza cade pressoché a zero quando si raggiungono i 45 anni e ciò non è incoraggiante per le persone di mezza età (fig. 7).
Ottimizzazione della storia naturale e variabilità genetica non adattativa
Un'altra idea chiave per la teoria delle storie naturali è stata suggerita da P.B. Medawar (1946), secondo il quale la genetica e la fisiologia interagirebbero con le forze declinanti della selezione nel produrre un degrado età-specifico di tutti gli aspetti funzionali dell'organismo, portando al decadimento delle funzioni riproduttive e della capacità di sopravvivenza quando l'organismo invecchia. Un meccanismo proposto da Medawar è quello delle mutazioni con effetti invecchiamento-specifici. Se mutazioni deleterie fossero mantenute nella popolazione da un equilibrio fra mutazione e selezione, se la forza della selezione diminuisse con l'invecchiamento e se certe mutazioni avessero effetti deleteri specifici per l'invecchiamento, queste mutazioni dovrebbero essere più frequenti in proporzione all'invecchiamento della classe di età su cui agiscono. Tali considerazioni sono oggi note come ipotesi dell'accumulo delle mutazioni.
G.C. Williams (1957) propose, in aggiunta, un altro meccanismo che sembra verificarsi frequentemente: l'esistenza di geni che hanno effetti contrastanti in classi di età iniziali e finali. Consideriamo un gene con effetti positivi che si manifestano precocemente nella vita, durante lo stadio giovanile o nel primo periodo riproduttivo, e con effetti deleteri che si manifestano tardi nella vita. Il suo effetto netto sulla fitness sarà positivo poiché le classi di età più giovani offrono un contributo alla fitness più ampio di quello che offrono le classi di età più vecchie. Tali mutazioni si diffonderanno e, se non sono stabilizzate nella popolazione, manterranno una correlazione genetica negativa tra i caratteri espressi precocemente e tardivamente nel corso della vita. Questa idea è oggi nota come ipotesi pleiotropica antagonista, antagonista in quanto gli effetti sono positivi in una classe di età e negativi in un'altra e pleiotropica perché ciascuno di questi geni influenza due o più caratteri.
Così la risposta evoluzionistica alla domanda "perché invecchiamo?" consta di tre parti. Innanzi tutto, deve esservi una separazione tra la linea germinale e quella somatica, dato che invecchia il soma, non la linea germinale (avere un singolo stadio cellulare durante il ciclo vitale è sufficiente a definire una linea germinale). Inoltre, la forza della selezione declina con l'età: oltre una certa età gli organismi diventano irrilevanti per l'evoluzione. Infine, date queste condizioni si possono osservare due tipi di effetti genetici: l'accumulo di mutazioni con effetti più forti sulle classi di età più avanzate rispetto a quelle più giovani e l'accumulo di geni di cui beneficiano le classi più giovani a spese di quelle più avanzate. La maturità è il punto della storia naturale in cui dovrebbero iniziare a verificarsi questi effetti, che prima non dovrebbero essere evidenti. Così l'invecchiamento dovrebbe seguire l'inizio della riproduzione, con un decadimento diffuso delle funzioni fisiologiche e biochimiche causato da diversi geni con piccoli effetti che producono l'invecchiamento che non rappresenta un adattamento quanto il sottoprodotto della selezione per prestazioni riproduttive più precoci nella vita. Non può essere escluso l'invecchiamento causato da geni con grandi effetti, ma non ci si aspetta che si tratti della situazione più comune. La morte cellulare geneticamente programmata si verifica; tuttavia non esiste una morte dell'organismo geneticamente programmata, esistono soltanto prestazioni riproduttive geneticamente programmate che possono avere la morte come effetto collaterale.
Ne consegue che la fitness è massima a un livello di investimento in attività di riparazione che è inferiore a quello che sarebbe richiesto per una sopravvivenza indefinita. Ecco perché cresciamo, invecchiamo e moriamo. Questa intuizione, sviluppata da T.B.L. Kirkwood (1987) e chiamata teoria del soma disponibile, stabilisce una connessione tra la teoria evoluzionistica dell'invecchiamento e la gerontologia. Tale teoria prevede che l'invecchiamento dovrebbe essere il prodotto dell'accumulo di danni somatici irreparabili e che le specie con differente longevità dovrebbero esibire corrispondenti differenze nei livelli di riparazione somatica. Tuttavia la riparazione è costosa: più del 2% del bilancio energetico di una cellula è speso nella riparazione del DNA e nella correzione dei moduli di lettura genici, nei processi che determinano l'accuratezza della sintesi proteica, nel ricambio delle proteine e nell'inattivazione dei radicali dell'ossigeno. Inoltre i geni che influenzano i processi di riparazione dovrebbero avere effetti antagonisti pleiotropici. Diminuzioni nella mortalità estrinseca dell'adulto aumenteranno il valore dell'investimento nella riparazione; a ciò dovrebbe essere accoppiata la diminuzione dell'investimento nella crescita e nella riproduzione, con conseguenti durate della vita superiori e sforzi riproduttivi inferiori. Al contrario, quando la mortalità estrinseca dell'adulto aumenta, l'investimento nella riparazione perde valore; dovremmo quindi osservare spostamenti verso una minore riparazione e un più alto investimento nella riproduzione.
Queste idee sono state verificate tramite esperimenti su moscerini della frutta eseguiti, tra gli altri, da M. Rose, L. Partridge, E. Hillesheim e B. Zwaan (Steams e Kaiser, 1996) con risultati in accordo con la teoria evolutiva: quando la forza della selezione su classi di età più vecchie viene aumentata, permettendo solo agli organismi più vecchi di riprodursi, l'invecchiamento è posticipato. In alcuni casi l'aumento della durata della vita è accompagnato da una ridotta fecondità negli stadi precoci della vita, in altri è accompagnato da modificazioni nei tassi di crescita larvale, in quelli di sopravvivenza e nell'abilità competitiva. Quando la selezione è applicata all'altro termine della vita, la dimensione alla maturità, i moscerini più piccoli che si evolvono vivono più a lungo e hanno una fecondità inferiore.
Le evidenze sperimentali sull'evoluzione dell'invecchiamento possono essere riassunte come segue: la durata della vita risponde rapidamente alla selezione di laboratorio, in modo conforme alla teoria evolutiva. Una vita più lunga deve comportare certi costi, sia una riduzione della prestazione negli stadi precoci della vita, sia una minore fecondità, minori dimensioni fisiche o una riduzione della capacità larvale e dell'abilità competitiva. Queste relazioni tra i caratteri espressi precocemente e tardivamente nella vita confermano il meccanismo suggerito da Williams (1957).
Le sfide del futuro
L'analisi dell'evoluzione delle storie naturali ha rivelato molti aspetti riguardanti le pressioni selettive che plasmano i cicli vitali e alcuni meccanismi genetici alla base della diversità. È stato compiuto il primo passo per spiegare perché il pianeta è occupato da organismi che mostrano una così grande varietà di dimensioni, durata della vita, età alla maturità e investimenti riproduttivi. Le parti meno chiare e più interessanti di questa analisi sono costituite dai meccanismi alla base dei tradeoff e dall'origine evolutiva dei limiti filo genetici all'adattamento. Lo sviluppo e l'applicazione delle metodologie necessarie per analizzare questi due effetti costituisce una sfida impegnativa.
Bibliografia citata
BELL, O. (1980) The costs of reproduction and their consequences. Am. Nat., 116, 45-76.
CHARLESWORTH, B. (1973) Selection in populations with overlapping generations. V. Natural selection and life histories. Am. Nat., 107, 303-311.
DAAN, S., DUKSTRA, C., TINBERGEN, J.M. (1990) Family planning in the kestrel (Falco tinnunculus): the ultimate control of covariation of laying date and clutch size. Behavior, 114, 83-116.
HAMILTON, W.D. (1966) The moulding of senescence by natural selection. J. Theor. Biol., 12, 12-45.
KIRKWOOD, T.B.L. (1987) Immortality of the germ line versus disposability of the soma. In Evolution of longevity in animals: a comparative approach, a c. di Woodhead A.D.H., Thompson K.H., New York, Plenum Press, pp. 209-218.
KOZLOWSKI, J. (1992) Optimal allocation of resources to growth and reproduction, implications for age and size at maturity. Trends in Ecol. Evol., 7, 15-19.
KOZLOWSKI, J., UCHMANSKI, J. (1987) Optimal individuaI growth and reproduction in perennial species with indeterminate growth. Evol. Ecol., 1, 214-230.
LEON, J.A. (1976) Life histories as adaptive strategies. J. Theor. Biol., 60, 301-335.
MEDAWAR, P.B. (1946) Old age and natural death. Modern Quarterly, l, 30-56.
MICHOD, R.E. (1979) Evolution of life histories in response to agespecific mortality factors. Am. Nat., 113, 531-550.
REZNICK, D.N., BRYGA, H., ENDLER, J.A. (1990) Experimentally induced life-history evolution in a natural population. Nature, 346, 357-359.
ROFF, DA (1981) On being the right size. Am. Nat., 118, 405-422.
SCHAFFER, W.M. (1974) Selection for optimal life histories: the effects of age structure. Ecology, 5, 291-303.
STEARNS, S.C., KAISER, M. (1996) Effect of fitness components of enhanced expression of elongation factor EF-la. III. Tradeoff analysis. Evolution, 50, 795-806.
STEARNS, S.C., KAISER, M., BLARER, A. (1996) A case study in experimental evolution: reproductive effort and induced responses in Drosophila melanogaster. Plant Species Biology, 11, 97-105.
STEARNS, S.C., KOELLA, J. (1986) The evolution of phenotypic plasticity in life-history traits: predictions for norms of reaction for age- and size-at-maturity. Evolution, 40, 893-913.
WILLIAMS, O.C. (1957) Pleiotropy, natural selection, and the evolution of senescence. Evolution, Il, 398-411.
WILLIAMS, O.C. (1966) Natural selection, the cost of reproduction, and a refinement of Lack's principle. Am. Nat., 100, 687-690.
YOUNG, T.P. (1990) Evolution of semelparity in Monnt Kenya lobelias. Evol., Ecol., 4, 157-172.
Bibliografia generale
CHARLESWORTH, B. Evolution in age-structured populations, 2a ed. Cambridge-New York, Cambridge University Press, 1994.
ROFF, D.A. The evolution of life histories: theory and analysis. New York, Chapman & Hall, 1992.
ROSE, M. The evolution of aging. Oxford, Oxford University Press, 1991.
STEARNS, S.C. The evolution of life histories. Oxford-New York, Oxford University Press, 1992.
WILLIAMS, O.C. Adaptation and natural selection. Princeton, Princeton University Press, 1966.