
Struttura degli odori e sistemi olfattivi
Struttura degli odori e sistemi olfattivi
L'olfatto e il gusto sono i due sensi chimici. Sebbene la differenza sia piuttosto sottile, il termine "gusto" viene in genere riservato alle sensazioni prodotte da contatto diretto, mentre il termine "olfatto" si riferisce a sensazioni prodotte in assenza di contatto diretto. Entrambe le sensazioni sono chimiche perché si basano su cellule sensoriali specializzate che rispondono selettivamente a determinati tipi di molecole. Gli animali, nelle diverse situazioni naturali e in ambienti dove gli odori sono complessi e variano nel tempo, devono elaborare i dati e decidere il loro comportamento tramite il sistema olfattivo. In questo saggio verranno esaminati i meccanismi alla base della diffusione delle sostanze odorifere e alcuni problemi computazionali del sistema olfattivo: la memorizzazione e il riconoscimento di odori, la localizzazione della sorgente, la separazione di odori e, infine, la separazione delle componenti di un odore.
Comportamento e sistema olfattivo
Il ruolo del sistema olfattivo nel comportamento animale dipende fortemente da quanto è lontano un oggetto che emana le sostanze odorifere. Quando questo è abbastanza vicino da dominare la scena sensoriale olfattiva, l'olfatto può essere usato per riconoscerlo. l ratti femmina, per esempio, riconoscono i propri piccoli dal loro odore. L'uomo può riconoscere una rosa o un cavolo bollito dal loro odore, e inoltre ha la capacità, seppur limitata, di distinguere le singole componenti di un odore che sono a lui familiari. Nello stufato possiamo individuare sia l'odore della carne di manzo che l'odore del vino di Borgogna; quando annusiamo il latte inacidito, possiamo sia identificare che si tratta di latte, sia che è presente una seconda componente sgradevole. Sebbene in questo saggio non verrà descritto il senso del gusto, dobbiamo notare che la finezza del senso del gusto nell 'uomo dipende dagli stessi chemiorecettori del naso che sono responsabili dell'olfatto. Infatti, una grave infezione nasale distrugge temporaneamente gran parte del nostro senso del gusto. L'uso dell'olfatto per il riconoscimento di oggetti implica la capacità di imparare a distinguere gli odori e ad associare a certi odori altri aspetti legati all'oggetto che li emana. Per esempio, questa capacità ci potrebbe permettere di associare al profumo di un cibo la possibilità di essere sazi per ore o a un odore il pericolo di essere feriti. Imparare a riconoscere il cibo commestibile è così importante che gli animali superiori si scambiano informazioni a riguardo. Per esempio, un ratto posto in un nuovo ambiente annuserà l'alito degli altri ratti per informarsi su quali cibi siano sicuri da mangiare. Si noti che pulirsi la bocca per un ratto verrebbe considerato un atteggiamento antisociale.
L'olfatto è anche usato per la percezione sensoriale a distanza. Gli esseri umani sono animali altamente 'visivi' e quindi tendono a considerare la vista come il modo migliore per capire quali oggetti li circondino e dove siano situati. Attraverso la vista si possono ottenere queste informazioni senza spostarsi, e questo è possibile anche mediante l'udito, sebbene i pipistrelli e le civette siano molto più bravi. Tuttavia, gli animali che hanno un sistema olfattivo particolarmente sviluppato possono distinguere le diverse sorgenti di cibo disponibili e individuare dove sono situate; inoltre, possono capire la direzione da cui viene un predatore o il luogo dove si trova un possibile partner, rimanendo fermi e annusando l'aria. Le lumache, per esempio, sono in grado di allontanarsi dal loro umido nido sotto una foglia, mangiare su un fungo a due metri di distanza e tornare al nido mediante l'olfatto. I salmoni possono ritrovare il luogo dove sono nati usando l'olfatto.
Inoltre, l'olfatto è molto utile per dirigersi verso oggetti come il cibo o i partner. Quando un amminoacido nutritivo viene posto sul fondo di una provetta, un batterio che si trova abbastanza vicino da rilevarne la presenza si muove verso la sorgente di cibo. Le api vengono guidate dal profumo verso i campi di fiori distanti, le aragoste trovano i pesci morti sul fondo del mare grazie alle sostanze odorifere da loro emesse e, allo stesso modo, i segugi inglesi seguono la pista usando l'olfatto. Tutti questi sono esempi di esplorazione attiva, in cui i movimenti dell'animale sono coordinati dall'odore e, a loro volta, l'intensità e la natura dell'odore dipendono dalla posizione e dai movimenti dell'animale.
La diffusione delle sostanze odorifere
Le molecole che sono emesse da una sorgente odorifera e vengono percepite dal sistema olfattivo devono aver coperto la distanza che separa la sorgente dal rivelatore; il loro moto è il risultato della diffusione e delle correnti d'aria. Quando la sorgente e il rivelatore sono vicini, la sola diffusione potrebbe essere un mezzo adeguato per portare le molecole al rivelatore. Tuttavia, la distanza percorsa per diffusione cresce solo in funzione della radice quadrata del tempo trascorso:
d = √Dt.
Le costanti di diffusione D nell' aria, per molecole di piccole dimensioni, sono di circa 0,01 cm²/s. Gli odori emanati da una sorgente che viene scoperta all'improvviso si diffondono nell'aria percorrendo in un minuto una distanza mediamente inferiore al centimetro. Se siamo molto sensibili all'odore, possiamo individuarlo poco prima, tuttavia questa legge di scala è drastica per odori distanti e ciò rende la diffusione trascurabile a distanze superiori a pochi centimetri. Anche gli spostamenti d'aria che si verificano in un ambiente naturale in cui l'aria sembra ferma, avvengono a una velocità di 0,5 krn/h e trasportano gli odori alla distanza di una decina di metri in un minuto. Per distanze maggiori gli odori vengono trasportati dalle correnti d'aria. Infine, la microturbolenza è responsabile della mescolanza dei flussi d'aria sulla scala del centimetro.
In presenza di spostamenti d'aria normali, un oggetto dotato di un certo odore emana le sostanze odorifere nella direzione verso cui soffia il vento. L'emanazione ha una microstruttura interna dovuta alla turbolenza su piccola scala, e una macrostruttura fluttuante e tortuosa dovuta a turbolenza su grande scala. In generale, gli animali trovano le sorgenti odorifere spostandosi controvento quando l'odore è forte. Il comportamento di una lumaca appartenente alla specie Limax maximus illustra questo punto: in una situazione artificiale in cui un odore attraente viene distribuito uniformemente nell' aria che fluisce dentro una galleria del vento, la lumaca si muove controvento. Limax presenta due organi olfattivi principali, uno su ciascun tentacolo. Quando in un esperimento viene amputato uno dei due organi, l'animale si muove ancora controvento. Invece, in assenza di vento, l'animale con un organo olfattivo amputato gira in tondo seguendo la direzione cui punta l'organo olfattivo restante. Sembra quindi che l'animale sia in grado di confrontare le sensazioni dei due organi olfattivi, e che questo confronto non venga fatto quando il vento non è trascurabile e l'animale sta cercando la sorgente di un odore lontano. Tuttavia, in assenza di vento, il comportamento dell'animale suggerisce che l'odore venga percepito come proveniente dal lato del solo organo olfattivo rimasto.
La distribuzione spaziale delle sostanze odorifere rende vantaggioso muoversi attraverso il vento se l'intensità diminuisce o l'odore viene perso. Il comportamento di un moscerino della frutta che trova una piccola sorgente alla fine della galleria del vento dimostra come esista una strategia più complicata per ritrovare una forte emanazione di odore. Tuttavia, la semplice strategia descritta è quanto basta a un animale così lento come Limax. È curioso notare che mentre si vola non vi sono indizi cinematici o dinamici per determinare la direzione del vento. Un animale che vola deve usare la vista per individuare la componente della velocità perpendicolare alla direzione del volo, oppure deve posarsi da qualche parte per sentire il vento mentre sta fermo. I moscerini della frutta hanno un circuito visivo che è in grado di misurare la deriva nella direzione perpendicolare all'asse del corpo e sulla base di questa determinare la direzione di volo contro vento.
Siccome la direzione del vento in un ambiente all'aperto, in condizioni normali, spesso cambia di angoli fino a 60° su una scala temporale di un paio di minuti, l'animale può 'vedere' in un cono non trascurabile di direzioni, anche rimanendo fermo in un posto. Un cane da caccia di solito punta un uccello che ancora non ha visto, guardando nella direzione del vento che stava soffiando quando aveva sentito l'odore.
Le cellule sensoriali e i loro segnali
Una cellula è un compartimento chiuso delimitato da una sottile membrana. I chemiosensori sono proteine specializzate che si trovano sulla membrana cellulare. Parte della proteina viene esposta al mezzo, tipicamente acquoso, che sta all'esterno della cellula; questa parte contiene un sito attivo che si può legare ad alcune molecole specifiche (ligandi). Il segnale chimico si innesca mediante il legame di un ligando con il sito attivo di un chemiosensore.
I chemiosensori sono piuttosto diffusi tra le cellule animali; alcuni hanno un sito di legame così specializzato da legarsi fortemente solo a un certo tipo di molecole. Le cellule del fegato, per esempio, hanno chemiosensori che si legano all'insulina. Il legame tra le due molecole attiva la via biochimica che converte il glucosio in glicogeno; di conseguenza, il glucosio viene rimosso dal sangue. Il grado di attivazione di questo percorso biochimico viene determinato dalla frazione di chemiosensori che si lega all'insulina, a sua volta determinata dalla concentrazione di insulina nel sangue.
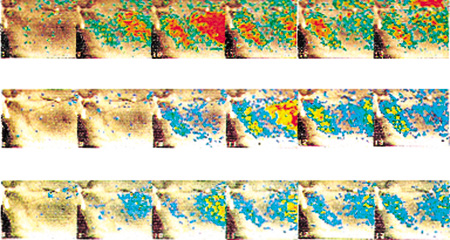
Lo stesso tipo di meccanismo cellulare è responsabile della risposta dei neuroni sensoriali olfattivi alle sostanze odorifere. Le cellule sensoriali sono neuroni specializzati che presentano chemiorecettori sulla superficie dei loro dendriti. Come per la maggior parte degli altri neuroni, quando vengono attivati, generano potenziali d'azione, cioè i segnali elettrochimici di breve durata (circa 2 ms) con i quali i neuroni comunicano l'uno con l'altro. È sufficiente posizionare due sonde di un oscilloscopio sensibile vicino a un neurone per rilevare i potenziali d'azione generati da quel neurone. In figura (fig. 1) viene mostrata la risposta del bulbo olfattivo di salamandra sottoposto a diverse sostanze odorifere.
Queste ultime generano una raffica di potenziali d'azione in una cellula la cui attività elettrica è normalmente molto più bassa; tale risposta sembra essere diversa per diverse sostanze odorifere. La biochimica che dall'evento di legame porta alla generazione di potenziali d'azione è complessa. Il legame tra il ligando e il chemiorecettore modifica la conformazione strutturale di quest'ultimo e avvia l'attività catalitica di un enzima all'interno della cellula. Di conseguenza viene attivata una cascata di processi chimici che alla fine determina l'apertura o la chiusura dei canali ionici attraverso cui fluisce la corrente entro la membrana della cellula nervosa. Il processo è molto simile a quello che avviene nell' occhio e che converte l'assorbimento della luce in un segnale nervoso. La differenza principale è che in un caso il cambiamento iniziale della conformazione della proteina viene generato dal legame delligando, e nell'altro dal cambiamento conformazionale della porzione cromo fora del recettore di luce, quando questa viene assorbita.
Alcuni chemiorecettori olfattivi sono altamente specifici: si legano a una particolare molecola molto di più di quanto riescano a legarsi a qualsiasi altra; si dice che sono, perciò, calibrati con precisione per rispondere a quel tipo di molecola. La loro risposta può essere interpretata come una misura della concentrazione di quella particolare specie chimica alla quale si legano selettivamente. Altri chemiorecettori si legano con la stessa facilità a una varietà di molecole diverse, in qualche caso nemmeno simili tra loro, e per questo si dice che sono poco calibrati. Gli insetti e gli animali superiori hanno due sistemi olfattivi: uno, il sistema specifico, riguarda soprattutto odori istintivi, e l'altro, il sistema generalista, riguarda l'elaborazione di odori in un contesto più generale. La capacità di un ratto appena nato di trovare la mammella della madre si basa sull'odore e sul sistema specifico. Di solito quest'ultimo ha recettori ben calibrati, come nel caso del feromone del sesso che permette alla falena maschio di trovare una femmina a un chilometro di distanza.
Nella visione dei colori, dato che ci sono solo tre tipi di recettori (i coni rossi, i coni verdi e i coni blu), lo spazio dei colori ha tre dimensioni. In analogia alla visione dei colori alcuni ricercatori hanno provato a misurare il numero di dimensioni dello spazio olfattivo studiando la risposta dell'uomo agli odori e le impressioni di somiglianza e differenza nel caso si confrontino odori diversi. Questa linea di ricerca non ha portato a risultati certi, tuttavia in genere produceva numeri superiori a 10. l profili delle cellule recettrici indicherebbero che lo stesso numero sarebbe valido per la salamandra. Recenti studi di genetica molecolare indicano che nel topo vi sono circa 400 proteine diverse che funzionano da chemiorecettori olfattivi. Alcune prove sperimentali (sebbene meno convincenti) indicano che una qualsiasi cellula sensoriale presenta un solo tipo di proteine chemiorecettrici. Se questo fosse vero, nel topo esisterebbero 400 tipi diversi di cellule che corrispondono ad altrettanti recettori olfattivi. Ciò porterebbe ad affermare che lo spazio degli odori di un topo ha 400 dimensioni. In realtà, si sa ancora relativamente poco su quanto siano diversi questi chemiorecettori, o se siano raggruppabili in un piccolo numero di famiglie che presentano profili di risposta simili. Nel caso del gusto, possiamo associare alla lingua dell'uomo uno spazio a quattro dimensioni che corrispondono al gusto amaro, dolce, acido e salato. Inoltre, è noto che mentre lo zucchero ha un gusto dolce per la maggior parte delle persone, una determinata sostanza avrà un gusto dolce per alcune persone ma non per altre. Percepire quella sostanza come dolce è il risultato di una semplice eredità mendeliana, e le persone che non lo sentono dolce probabilmente non sono in grado di sintetizzare un tipo di proteina chemiorecettrice presente in altre persone. Quindi, mentre la psicofisica del gusto viene ben descritta da uno spazio del gusto quadri dimensionale, esistono più di quattro tipi di recettori. Di conseguenza possiamo solo concludere che allo stato attuale il numero di dimensioni dello spazio degli odori deve essere specifico per ogni animale e deve variare nell'intervallo che va da 10 a 1000.
Il termine anosmia descrive l'incapacità di percepire odori, ovvero lo stato di 'cecità' olfattiva. Un tipo di anosmia prevede l'incapacità di percepire certi odori ed è l'equivalente olfattivo della cecità per un colore specifico. Anosmie ereditabili altamente specifiche indicano che almeno alcuni dei nostri recettori devono essere ben calibrati per percepire solo certi odori. Per esempio, l'androstenone è virtualmente inodore per circa un terzo delle persone, ha un profumo lieve e gradevole per un altro terzo, e un odore forte e sgradevole per l'altro terzo, sebbene le persone dei tre tipi sono indistinguibili quando si prendono in considerazione altri odori.
I tre problemi computazionali dell'olfatto
L'etologia degli animali macrosmatici descrive il comportamento in varie situazioni naturali in presenza di ambienti dove gli odori sono complessi e variano nel tempo. La risposta delle cellule sensoriali agli odori descrive l'informazione in base alla quale l'animale deve 'elaborare' i dati disponibili e decidere cosa fare nelle diverse situazioni. l tre problemi più semplici che riguardano il sistema olfattivo e che possono essere isolati in questo comportamento sono la memorizzazione e il riconoscimento di odori, la separazione di odori e la separazione delle componenti. In primo luogo, come può un animale ricordare le informazioni sul pattem di eccitazione dei neuroni sensoriali in modo che gli odori che si ripresentano possano essere riconosciuti indipendentemente dall'intensità e che la loro intensità possa essere valutata? Quando gli odori di oggetti diversi vengono mescolati dalle turbolenze presenti in un normale flusso d'aria, come può l'animale separarli e ottenere l'informazione su dove i diversi oggetti si trovino? Quando alcuni odori noti vengono mescolati, come fa l'animale a identificare gli odori componenti?
Memorizzazione e riconoscimento di odori
L'aspetto algoritmico della memorizzazione e del riconoscimento di odori è concettualmente il più semplice di questi problemi ed è particolarmente rilevante per i comportamenti che si osservano quando un solo odore domina la scena olfattiva. La maggior parte degli odori naturali consiste in un miscuglio di diversi tipi di molecole. A un certo livello di intensità, un odore complesso b può essere descritto dall'insieme delle concentrazioni delle molecole di specie i che lo compongono Nbi. Al variare dell'intensità dello stimolo, ogni componente aumenta (o diminuisce) perché moltiplicata per uno stesso fattore comune. Risulta utile descrivere lo stimolo come il risultato di due fattori, l'intensità l e le componenti normalizzate nbi:
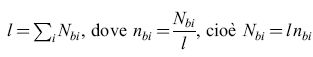
Le nbi sono normalizzate, cioè rappresentano le concentrazioni relative delle diverse molecole, e l descrive l'intensità globale dell' odore. Idealmente una data qualità di un odore è descritta dal pattem di nbi, che rimane inalterato quando l'intensità globale l cambia. Quando viene presentato uno stimolo descritto da un insieme di concentrazioni di molecole di specie j, un rivelatore ideale di qualità olfattive Nsi, risponderebbe in modo affermativo alla domanda sulla presenza dell'odore b se e solo se per un certo valore di l si ha che Nsj ≈ l nbj per tutti i valori di j. l criteri per stabilire quanto si debbano rispettare queste relazioni di proporzionalità per confermare l'avvenuto riconoscimento dipenderanno dal contesto biologico. Questo tipo di elaborazione è stato chiamato appaiamento analogico.
L'identificazione del colore della luce potrebbe essere descritta in modo simile. Idealmente un animale potrebbe avere migliaia di recettori di luce molto specifici, ognuno dei quali risponderebbe a una specifica lunghezza d'onda. In questo caso il confronto tra un nuovo stimolo e un colore conosciuto costituirebbe un problema di appaiamento analogico in migliaia di dimensioni. Tuttavia, la nostra visione dei colori usa solo tre recettori poco calibrati. Il confronto tra colori che il nostro cervello effettua ha ancora la forma di un appaiamento analogico, ma solo in tre variabili.
Gli studi sulle cellule sensoriali dei vertebrati nei sistemi generalisti, descritti nel paragrafo precedente, mostrano che ciascun tipo di molecola stimola molte cellule sensoriali differenti, alcune più fortemente, altre meno. Allo stesso tempo, ogni cellula viene attivata da diversi tipi di molecole. Il pattem di attivazione relativa che si ottiene osservando una popolazione che contiene diversi tipi di cellule sensoriali determina la qualità dell'odore nel sistema olfattivo generali sta e indica se la sostanza odorifera sia costituita da un miscuglio chimico o da un singolo tipo di molecole. La riduzione del problema ideale (un recettore per ciascun tipo di molecola) a un insieme di alcune centinaia di tipi di cellule recettrici poco calibrate è simile alla riduzione del problema che opera il sistema visivo quando utilizza solo tre classi di recettori. l sistemi olfattivi generalisti degli animali superiori sembrano risolvere il problema computazionale:
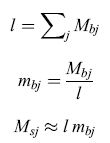
esattamente come descritto in precedenza, con la sola differenza che l'indice j adesso si riferisce ai canali effettivi di input invece che ai tipi di molecole e le componenti M all'attività dei vari canali di input. Il problema che deve affrontare il sistema nervoso viene reso più difficile di quanto possa sembrare dal fatto che le singole cellule sensoriali sono non lineari e che la frazione di recettori con un ligando legato ha una dipendenza non lineare dalle concentrazioni molecolari. Tuttavia, dobbiamo notare che il sistema visivo deve affrontare le stesse difficoltà nel contesto del problema del colore.
Animali macrosmatici come i ratti o i cani da caccia hanno una straordinaria capacità di distinguere odori molto simili. Tramite manipolazioni genetiche è possibile avere due ceppi di topo che differiscono per un solo gene. Se questo gene appartiene al gruppo di istocompatibilità, le urine dei due tipi di topo vengono facilmente distinte da ratti addestrati, sebbene il prodotto del gene sia esso stesso una proteina inodore e non abbia nulla a che fare con i processi metabolici che producono le componenti dominanti delle urine. Il riconoscimento degli odori coinvolge necessariamente due aspetti: uno riguarda la memorizzazione della conoscenza della qualità degli odori percepiti in passato, l'altro riguarda il confronto tra gli odori conosciuti e le qualità degli odori passati al fine di identificare l'odore presente e dame almeno una stima qualitativa dell'intensità.
La localizzazione della sorgente e la separazione di odori
Il problema della localizzazione della sorgente e della separazione di odori viene affrontato quando un animale con sistema olfattivo particolarmente sviluppato percepisce odori da sorgenti distanti. L'odore di una piccola quantità di cibo viene trasportato dal vento e, a causa della natura turbolenta del flusso d'aria, viaggia erraticamente cambiando continuamente direzione. In media, l'animale può percepire l'odore quando il vento sta soffiando dalla direzione del cibo. Se il cibo si trova molto vicino, la correlazione è molto alta, ma esso non viene percepito affatto se il vento soffia in altre direzioni.
Siccome la direzione del vento cambia nel tempo, la sostanza odorifera percorrerà una traiettoria tortuosa su lunghe distanze. Vi sarà un intervallo di direzioni del vento per le quali l'odore può essere percepito, e non sarà sempre avvertito dall'animale quando il vento soffia in una data direzione. La correlazione tra la presenza dell'odore e la direzione del vento decresce con la distanza dalla sorgente. Una sorgente odorifera estesa ha un tipo diverso di correlazione rispetto a una piccola. Quindi, quando vi è una sola sorgente odorifera, un animale fermo può ottenere informazioni sulla direzione e la distanza della sorgente. Se sono presenti due oggetti in due direzioni ben separate, l'animale può stabilire quale dei due sia più lontano.
Separazione delle componenti di un odore
Quando due sorgenti odorifere sono situate nella stessa direzione, allora si presenta un problema computazionale aggiuntivo. Per un sistema generali sta, gli odori vengono descritti dal pattem di attivazione che l'odore provoca tramite un insieme di recettori. In generale, a un odore P corrisponde un pattem e a un odore Q un altro. Supponiamo che in un esperimento i due odori siano completamente mescolati. Quando il miscuglio viene percepito, genera un pattem di eccitazione PQ nelle cellule sensoriali. È possibile che l'animale possa riconoscere che la sorgente odorifera è composita e contiene un miscuglio di P e di Q? Se entrambi gli odori sono sconosciuti, non vi è alcun modo di ottenere una separazione in un sistema generali sta. Non riusciamo a percepire che un profumo elegante possa essere il prodotto del miscuglio di vari odori, ognuno dei quali a sua volta è un odore complesso. Analogamente, una lumaca può essere addestrata a evitare un odore composito PQ senza che per questo mostri ripugnanza per P o Q presi separatamente. Questo problema sembrerebbe rendere molto difficile la percezione a distanza, dal momento che in un ambiente naturale l'aria che proviene da una sorgente odorifera si mescola sempre, a causa delle turbolenze, con quella che proviene da altre sorgenti che si trovano sul percorso. Tuttavia, sono presenti due sorgenti di informazioni addizionali che aiutano l'elaborazione del segnale olfattivo: le fluttuazioni del rapporto di mescolanza e la conoscenza di uno o di entrambi gli odori.
La possibilità di utilizzare le fluttuazioni per separare gli odori mescolati viene dal fatto che per una piccola sorgente il mescolamento dovuto alla turbolenza diluisce l'odore e lo può mescolare ad altri pacchetti di aria che contengono altre sostanze odorifere, ma tutti i contributi della piccola sorgente al pattem di eccitazione dei recettori si muoveranno insieme, in un cammino casuale che dipende dai movimenti turbolenti nell'aria. A questi contributi si potrebbero mescolare gli odori di un altro oggetto, situato in un posto diverso. Le componenti di un altro oggetto fluttueranno insieme, ma i contributi di due oggetti diversi fluttueranno in modo indipendente e non saranno correlati. Grazie a questa indipendenza diventa possibile elaborare lo stimolo olfattivo sulla base delle fluttuazioni e raggruppare insieme i contributi di ciascun oggetto indipendente.
Sebbene questa procedura sembri difficile e dia l'impressione di essere una soluzione troppo astratta, lo stesso tipo di problema può essere facilmente illustrato per il sistema visivo. Si schizzi a caso un foglio con gocce di inchiostro in modo che la pagina appaia come una struttura a chiazze senza segni particolari, e poi se ne facciano due fotocopie. Quindi si ritagli da una delle due copie una figura semplice come il profilo di un cane e si stenda il ritaglio sull'altra copia. In pratica il profilo della figura sarà invisibile dal momento che copre un disegno casuale con un altro disegno casuale equivalente. Se ora si agita la figura ritagliata, si potrà immediatamente percepire il profilo che scomparirà di nuovo non appena si smette di agitare la figura ritagliata. L'idea che sta alla base del funzionamento sia del sistema olfattivo che di quello visivo è che ciò che fluttua (si muove) insieme è un oggetto singolo. Non sappiamo come il sistema visivo sia in grado di fare questa operazione di raggruppamento, tuttavia vediamo che lo fa senza pensarci troppo e con una rapidità incredibile. Il sistema uditivo è in grado di effettuare un raggruppamento simile. Anche se non è stata studiata in dettaglio, la capacità degli animali macrosmatici di capire il loro ambiente è assai meno misteriosa se pensiamo che lo stesso tipo di elaborazione è così evidente in altri sistemi sensoriali.
L'uomo ha la capacità di separare gli odori nelle loro componenti quando questi ultimi sono già conosciuti, anche quando gli odori sono completamente mescolati. l profumieri spesso diventano degli esperti nell'arte di riconoscere un certo numero di sostanze, e questo permette a un'industria di scomporre e poi copiare un profumo che si è rivelato un successo commerciale per un'altra. Analogamente i bravi cuochi sono in grado di individuare i diversi ingredienti di un piatto preparato da altre persone. Questa separazione in componenti non include informazioni dettagliate sulle quantità relative coinvolte, che vengono poi trovate negli esperimenti.
Il processo di adattamento è un potente aiuto per questa analisi. La nostra sensibilità alla maggior parte degli odori diminuisce in seguito a una esposizione prolungata. Se ci muoviamo dentro una cucina dove vi è un forte odore di cavolo che cuoce, dopo un minuto non ci rendiamo più conto della presenza di un odore così forte pur essendo ancora sensibili ad altri odori. Alcuni odori sono così effimeri che la nostra reazione a essi termina in mezzo secondo, e quando annusiamo una seconda volta non li ritroviamo più. L'adattamento differenziato per le diverse componenti degli odori dimostra che lo stesso odore non sarà percepito necessariamente allo stesso modo dopo pochi secondi e gli esperti che conoscono questo fenomeno lo possono sfruttare per separare le varie componenti. Il processo di adattamento non dovrebbe essere confuso con il cambiamento fisico nel tempo dell'odore, che pure costituisce un problema per i profumieri e aiuta a separare le varie componenti. Le diverse componenti chimiche di un odore hanno diverse velocità di evaporazione su una superficie e scompaiono in tempi diversi. Quindi, un profumo spruzzato sulla pelle cambierà nel tempo poiché variano le concentrazioni relative delle sue componenti. Questo costituisce un problema per il profumiere che vorrebbe produrre un profumo che duri a lungo e non cambi troppo nel tempo. Nello stesso tempo costituisce un aiuto per scomporre un profumo, perché fornisce effettivamente all'analista miscugli con diverse quantità relative delle componenti.
L'adattamento è un fenomeno complesso principalmente studiato negli esperimenti che prevedono l'uso di coppie di odori. Nel protocollo sperimentale più semplice l'odore a viene presentato per pochi secondi e successivamente viene misurato il suo effetto sul livello di soglia per l'odore b. È interessante notare che gli effetti non sono sempre inibitori: spesso l'effetto è di aumentare la sensibilità all'odore b.
La capacità di separare le miscele nelle loro componenti è utile per gli animali che vivono nel loro habitat naturale. Per esempio, l'intensità dell'odore di trifoglio in fiore confrontata con l'intensità dell'odore delle piantine senza fiore è importante per un'ape che deve valutare se un leggero odore dolce valga la pena di un volo o se la presenza di fiori è trascurabile. Tuttavia, nonostante sia semplice immaginare situazioni in cui l'intensità delle componenti è importante per determinare il comportamento, questo settore dell'etologia è stato sostanzialmente trascurato.
Anatomia del sistema olfattivo

Il sistema olfattivo dei vertebrati e degli invertebrati è caratterizzato innanzitutto da neuroni che presentano sulle loro membrane proteine chiamate chemiorecettori. Gli assoni di queste cellule trasportano i segnali dei potenziali d'azione dalle cellule olfattive alla prima area di elaborazione (fig. 2). Quest'ultima, nei vertebrati, è rappresentata dal bulbo olfattivo da cui partono gli assoni che arrivano alla corteccia prepiriforme (corteccia olfattiva), la prima area di elaborazione corticale del sistema olfattivo. Quest'area fa parte della paleocorteccia, distinta dalla neocorteccia che occupa la maggior parte del volume del cervello dell'uomo. La paleocorteccia presenta meno strati di neuroni e un'architettura più semplice rispetto alla neocorteccia. Di tutte le regioni della paleocorteccia, la corteccia olfattiva ha l'architettura più semplice, che sembra sia divenuta più complessa in aree come l'ippocampo. Uno dei motivi per cui i ricercatori studiano il sistema olfattivo è che potrebbe rivelare alcuni principi di funzionamento di altre aree della paleocorteccia. Negli invertebrati, come la lumaca e la locusta, le prime connessioni dalle cellule sensoriali sono situate in una struttura conosciuta come il lobo dell'antenna.
Il bulbo olfattivo dei vertebrati e la sua elettrotisiologia

Il bulbo olfattivo è caratterizzato da quattro tipi principali di neuroni (fig. 3). I più importanti sono le cellule mitrali. Ogni cellula mitrale presenta la sua arborizzazione dendritica principale in una struttura globulare chiamata glomerulo. L'assone di ciascuna cellula sensoriale si connette a un singolo glomerulo. Nel ratto, sono presenti circa 2000 glomeruli e ben 5.10⁷ cellule recettrici, e quindi circa 2000 connessioni che arrivano a ogni glomerulo. Studi recenti di biologia molecolare mostrano che solo una cellula recettrice su 1000 contiene una proteina specifica che si comporta da recettore.
I numeri sono tali da indurci a pensare che forse esistono 2000 classi di cellule recettrici, che ogni classe contiene solo un tipo di recettore e che vi sono uno o due glomeruli che ricevono input da tutti i recettori di una certa classe. Sfortunatamente ancora non si sa se in una determinata cellula sia presente una sola proteina recettrice, quindi, questo semplice modello potrebbe non essere corretto. In corrispondenza dei glomeruli vi è una struttura di diramazione dal momento che ciascun glomerulo contiene i dendriti di circa 25 cellule mitrali. Tutte le cellule descritte finora producono neurotrasmettitori in genere eccitatori; il circuito appena descritto porterebbe a uno schema in cui l'eccitazione passa dai neuroni olfattivi alle cellule mitrali i cui assoni trasferiscono l'eccitazione alla corteccia piriforme. Tuttavia, il circuito è più complicato per la presenza di cellule inibitorie. Inoltre, un gruppo di cellule periglomerulari, situate attorno a ciascun glomerulo, ricevono input dagli assoni delle cellule sensoriali. Queste cellule si connettono mediante sinapsi bidirezionali ai dendriti delle cellule mitrali che forniscono un percorso attraverso il quale il segnale, che entra nel bulbo olfattivo sull' assone di una cellula sensoriale, può in seguito inibire la risposta delle cellule mitrali. Il bulbo olfattivo contiene anche piccole cellule inibitorie chiamate granuli che sono circa dieci volte più numerose delle cellule mitrali. I granuli ricevono un input sia dalle connessioni collaterali degli assoni delle cellule mitrali, sia dalle sinapsi dendritiche bidirezionali che si connettono alle cellule mitrali. Queste cellule inibitorie forniscono il meccanismo mediante il quale ogni cellula mitrale può inibire indirettamente la risposta di un'altra cellula mitrale o di se stessa.
Le cellule eccitatorie chiamate cellule a pennacchio sono il quarto tipo principale di cellula del bulbo olfattivo. Gli input che ricevono le cellule a pennacchio sono qualitativamente simili a quelli delle cellule mitrali, come del resto il loro circuito di inibizione. Tuttavia, mentre le cellule mitrali inviano prevalentemente il loro output alla corteccia piriforme, le cellule a pennacchio spesso inviano gli assoni verso altre regioni del cervello, come il nucleo olfattivo anteriore e la corteccia piriforme anteriore. Tali cellule forniscono anche il collegamento tra i bulbi olfattivi destro e sinistro.
Allo stato attuale non è disponibile una descrizione generale soddisfacente delle funzioni computazionali o dell'algoritmo del bulbo olfattivo. Anche il suo semplice circuito è troppo complicato e con troppe incognite per una simulazione che abbia senso. La situazione è resa ancora più complicata dal fatto che vi è un elevato numero di assoni che portano segnali al bulbo olfattivo da altre parti del cervello, corteccia prepiriforme inclusa. Tuttavia, nella maggior parte dei casi, dobbiamo notare che i primi centri di elaborazione olfattiva di tutti gli animali sono sede di oscillazioni elettriche. Dopo aver percepito un odore, il bulbo olfattivo dei ratti o dell 'uomo comincia ad avere delle oscillazioni di 40÷50 Hz. Il lobo dell'antenna e il corpo a forma di fungo della locusta presentano oscillazioni di 20 Hz e il lobo procerebrale della lumaca oscilla a 0,5 Hz. È ovvio pensare che queste oscillazioni, facilmente prodotte dai circuiti eccitatori-inibitori presenti in questi organismi, giochino un ruolo importante nell'elaborazione del segnale olfattivo.
Sistemi specializzati
Per la maggior parte degli organismi si possono trovare due classi differenti di odori. La prima riguarda l'ambiente generale, in cui si possono presentare nuove combinazioni di odori, possono verificarsi nuove situazioni e l'apprendimento può essere specifico per un individuo e per una situazione. Il sistema olfattivo deve essere preparato a trattare nuovi odori sconosciuti, a imparare nuovi odori e ad adattarsi a nuove circostanze. La seconda classe di odori comprende quelli che sono specifici per una specie, gli odori (o analogamente i sapori) dei cibi più importanti, gli odori di sostanze nocive o quelli di altri organismi della stessa specie.
L'odore della propria specie e le differenze di odore che distinguono i sessi sono importanti per la maggior parte degli organismi terrestri. Il sistema di feromoni che determina l'attrazione sessuale degli insetti è un esempio particolarmente semplice. Gli insetti che non formano colonie escono dalle uova e sono liberi. Una conoscenza innata delle sostanze odorifere prodotte dall'altro sesso della stessa specie consente di localizzare e identificare con successo un potenziale partner. In effetti, il corteggiamento e la ricerca del partner negli animali che vengono allevati in assenza di adulti della stessa specie devono essere geneticamente o epigeneticamente precostituite.
Gli odori dell'ambiente e del cibo possono anche essere speciali. Alcuni buoni odori semplici come, per esempio, l'odore aromatico della frutta per un onnivoro, e quello di alcune sostanze chimiche dannose come il solfuro di idrogeno possono essere così stabili e importanti da giustificare l'evoluzione di un apparato specifico per individuarli e classificarli. Le risposte precostituite forniscono anche un quadro di riferimento nell'ambito del quale gli organismi possono associare input sensoriali prima sconosciuti. Se si prende un animale che abbia una risposta positiva precostituita alla somministrazione di zucchero e gli si dà acqua e zucchero, che è inodore, in associazione a un odore neutro come, per esempio, quello del benzene, l'animale imparerà rapidamente a muoversi verso la sorgente di quell'odore. Si pensa che per la maggior parte dei mammiferi il sistema olfattivo accessorio sia di solito la sede principale della sensibilità precostituita a odori speciali. Il sistema specializzato dei mammiferi è caratterizzato dall' organo vomeronasale che gioca il ruolo di epitelio sensoriale per il sistema accessorio.
Il bulbo olfattivo accessorio riceve il suo input sensoriale diretto dall'organo vomeronasale. Mentre molti primati hanno un organo vomeronasale evidente, si discute ancora se l'uomo abbia una struttura vomeronasale e, in caso affermativo, ci si chiede se sia caratterizzato da neuroni sensoriali olfattivi. Tuttavia, l'organizzazione del cervello raramente è così semplice da assegnare una sola funzione a una specifica regione del cervello. Quindi, l'eventuale mancanza di questo organo non significa che per l'uomo non esistano odori attraenti o sgradevoli precostituiti. Per esempio, l'odore delle uova marce (solfuro di idrogeno) sembra essere naturalmente sgradevole per l'uomo e probabilmente non è una sorpresa che l'odore caratteristico della carne o delle uova in decomposizione sia divenuto un avvertimento che quel cibo non va mangiato. Molto dipende dalla robustezza del nostro apparato digerente! Per l'aragosta i pesci morti sono un possibile pasto e un piccolo polipeptide di quattro amminoacidi, che spesso viene generato da un muscolo in decomposizione, è un odore attraente per questo organismo. De gustibus non est disputandum.
Bibliografia generale
BUCK, L.B. (1996) Information coding in the vertebrate olfactory system. Annu. Rev. Neurosci., 19, 517-544. ERMENTROUT, B., FLORES, J., GELPERIN, A. (1998) Minimal model of oscillations and waves in the Limax olfactory lobe with the tests of the model's predictive power. J. Neurophysiol., 79, 2677-2689.
GETCHELL, T.V., DOTY, R.L., BARTOSHUK, L.M., SNOW, J.B. Smell and taste in health and disease. New York, Raven Press, 1991.
HOPFIELD, J.J. (1991) Olfactory computation and object perception. Proc. Natl. Acad. Sci. USA, 88, 6462-6466. HOPFIELD, J.J. (1995) Pattem recognition computation using action potential timing for stimulus representation. Nature, 376, 33-36.
HOPFIELD, J.J. (1996) Transforming neural computations and representing time. Proc. Natl. Acad. Sci. USA, 93, 15.440-15.444.
LAING, D.G., DOTY, R.L., BREIPOHL, W., a c. di, The human sense of smell. Berlino-New York, Springer Verlag, 1991.
LAWLESS, H.T. Olfactory psychophysics. In Tasting and smelling, a c. di Beauchamp G.K., Bartoshuk L., Academic Press, San Diego, pp. 125-221, 1997.
MacDONALD, D.W., MULLER-SCHWARZE, D., NATYNCZUK, S.E., a c. di, Chemical signals in vertebrates 5. Oxford-New York, Oxford University Press, 1990.
MARGOLIS, F.L., GETCHELL, T.V. Molecular neurobiology oJthe olfactory system: molecular, membranous, and cytological studies. New York, Plenum Press, 1988.
SERBY, M.J., CHOBOR, K.L., a c. di, The science of olfaction. New York, Springer Verlag, 1992.
SINGER, A.G., BEAUCHAMP, G.K., YAMAZAKI, K. (1997) Volatile signals of the major histocompatibility complex in male mouse urine. Proc. Natl. Acad. Sci. USA, 94, 2210-2214.
WEHR, M., LAURENT, G. (1996) Odour encoding by temporal sequences of firing in oscillating neural assemblies. Nature, 384, 162-166.