
udito
udito
L’udito, ossia la capacità di percepire e distinguere una vasta gamma di suoni, è uno dei sensi più importanti per l’uomo. Esso è alla base delle capacità di comunicazione, fornisce una serie continua di informazioni relative all’ambiente circostante e consente di apprezzare la bellezza di una sinfonia o la voce della natura, ma funge anche da sistema di allarme e può contribuire in alcuni casi alla sopravvivenza dell’individuo. I suoni sono prodotti dalle vibrazioni di un corpo, come la corda di una chitarra, la membrana di un tamburo o le corde vocali, e si propagano nell’aria (o in altri mezzi) sotto forma di onde di variazione di pressione. Quando queste raggiungono l’orecchio, l’energia meccanica viene trasmessa tramite una serie di passaggi fino alla coclea, che è l’organo responsabile della trasformazione dei suoni in segnali nervosi. Da qui le informazioni uditive viaggiano lungo le fibre del nervo acustico per raggiungere il sistema nervoso centrale. Alcuni aspetti delle sensazioni uditive, come la localizzazione delle sorgenti sonore, vengono analizzati a livello del tronco dell’encefalo, ma la percezione cosciente dei suoni avviene a seguito dell’attivazione della corteccia cerebrale. [➔ cervello, struttura e funzione del; linguaggio; meccanotrasduzione; musica e cervello; sensazione e percezione] Le onde sonore hanno tre proprietà che le caratterizzano: la frequenza (misurata in hertz), che corrisponde, a livello percettivo, all’altezza del suono (i toni acuti hanno frequenze elevate, quelli gravi frequenze basse); l’ampiezza, in relazione alla quale varia l’intensità del suono (misurata in decibel); il timbro, che dipende dalla complessità dell’onda e si differenzia a seconda della sorgente che emette il suono. L’apparato uditivo umano è sensibile a uno spettro di frequenze acustiche abbastanza ampio, che va da 20 Hz a ca. 20.000 Hz. Molti animali possiedono una sensibilità uditiva superiore a quella dell’uomo: i pipistrelli, le orche e i delfini, per es., sono in grado di captare suoni con frequenze fino a 200.000 Hz (ultrasuoni), mentre elefanti, alligatori e balene sono noti per comunicare tramite infrasuoni (frequenze inferiori a 16 Hz). La capacità dinamica del nostro udito è estesa anche in termini di percezione dell’intensità di un suono: l’escursione tra soglia di udibilità (intensità minima di un suono per essere udito, posta per convenzione a 0 dB) e quella del dolore (intensità minima per avere danni uditivi anche per brevi esposizioni, che è circa 140 dB) è molto grande.
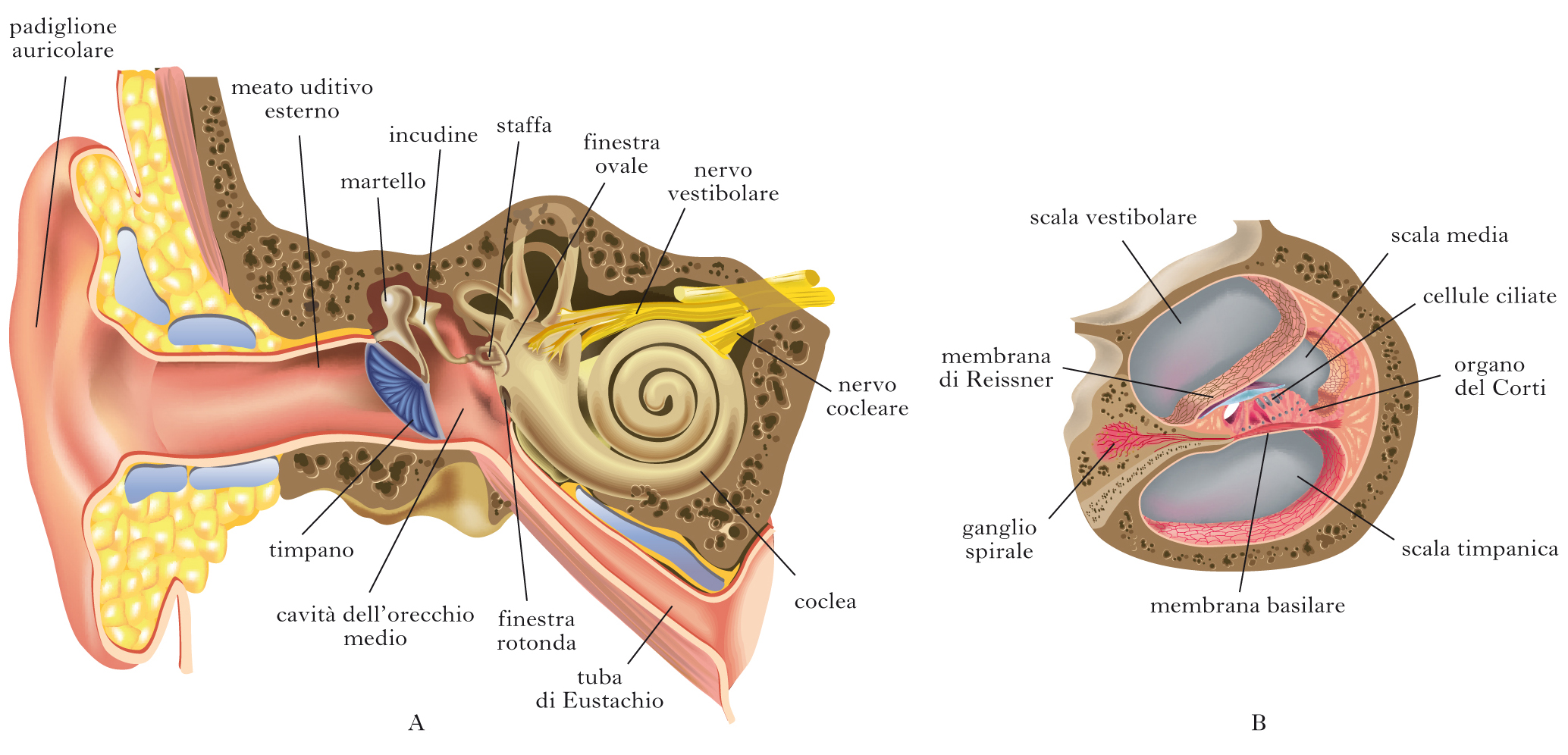
L’orecchio
L’orecchio è costituito da tre parti, ognuna delle quali svolge una funzione diversa. Il padiglione auricolare e il condotto uditivo, che compongono l’orecchio esterno, convogliano l’energia sonora sulla membrana timpanica. Le vibrazioni di questo sottile diaframma innescano il movimento di tre piccoli ossicini, collegati tra loro (martello, incudine e staffa), che costituiscono l’orecchio medio, e in questo modo vengono trasmesse alle strutture dell’orecchio interno. La coclea, così chiamata per la sua forma a spirale che la rende simile al guscio di una lumaca, è la porzione dell’orecchio interno devoluta alla funzione uditiva. Essa è suddivisa in tre concamerazioni, riempite di un liquido con una composizione peculiare: osservandola in sezione trasversale, si distinguono dall’alto verso il basso la scala vestibolare, la scala media e la scala timpanica. Alla base della coclea, la scala vestibolare comunica con l’orecchio medio per mezzo della finestra ovale, una membrana simile al timpano, su cui insiste il piede della staffa; la scala timpanica parimenti possiede un’apertura basale, detta finestra rotonda, chiusa da un rivestimento elastico. Queste due cavità sono in connessione tra loro all’apice della coclea, mentre la scala media è un dotto indipendente, delimitato da due lamine membranose; in partic., essa è separata dalla sottostante scala timpanica dalla membrana basilare. La struttura in cui hanno luogo i processi di trasduzione dei segnali acustici è l’organo del Corti, appoggiato sulla membrana basilare e costituito da diversi tipi cellulari, tra cui le cellule ciliate, che sono i recettori sensoriali primari. L’azione della staffa sulla finestra ovale genera variazioni di pressione che si propagano nel liquido cocleare e si riflettono, in ultima analisi, in deflessioni della membrana basilare e nella depolarizzazione delle cellule ciliate.
Mappa tonotopica
I suoni elementari, detti toni puri, sono quelli rappresentabili come un’onda sinusoidale di una certa frequenza. Nel mondo che ci circonda, tuttavia, gli stimoli acustici sono suoni complessi, prodotti dalla sovrapposizione di più suoni semplici. Una delle caratteristiche primarie della percezione uditiva è che il sistema uditivo funziona come un analizzatore di frequenze che scompone i suoni complessi nelle loro componenti pure. Le proprietà meccaniche della membrana basilare costituiscono l’elemento chiave alla base di questo processo: essa, infatti, è più larga ed elastica alla sua estremità apicale e più stretta e rigida alla base (la parte più prossima all’orecchio medio). Ne deriva che le vibrazioni indotte dalla stimolazione acustica non sono uniformi per tutta la sua lunghezza, ma presentano escursioni di ampiezza massima in punti particolari, strettamente correlati alla frequenza delle onde sonore. All’apice della coclea la membrana basilare risponde preferenzialmente alle frequenze più basse udibili, nella parte opposta sono le frequenze elevate a determinare i movimenti più ampi; le frequenze intermedie sono rappresentate in sequenza tra queste estremità. Questa organizzazione è detta mappa tonotopica. Gli stimoli sonori evocano un movimento complesso della membrana basilare, in cui le principali frequenze che compongono il suono determinano un’escursione approssimativamente proporzionale alla loro intensità nei punti della membrana sintonizzati su di esse. Il movimento della membrana basilare viene trasdotto nella depolarizzazione delle cellule ciliate localizzate in posizioni corrispondenti dell’organo del Corti, dando inizio al processo di codificazione delle frequenze e delle intensità dei suoni (➔ meccanotrasduzione).
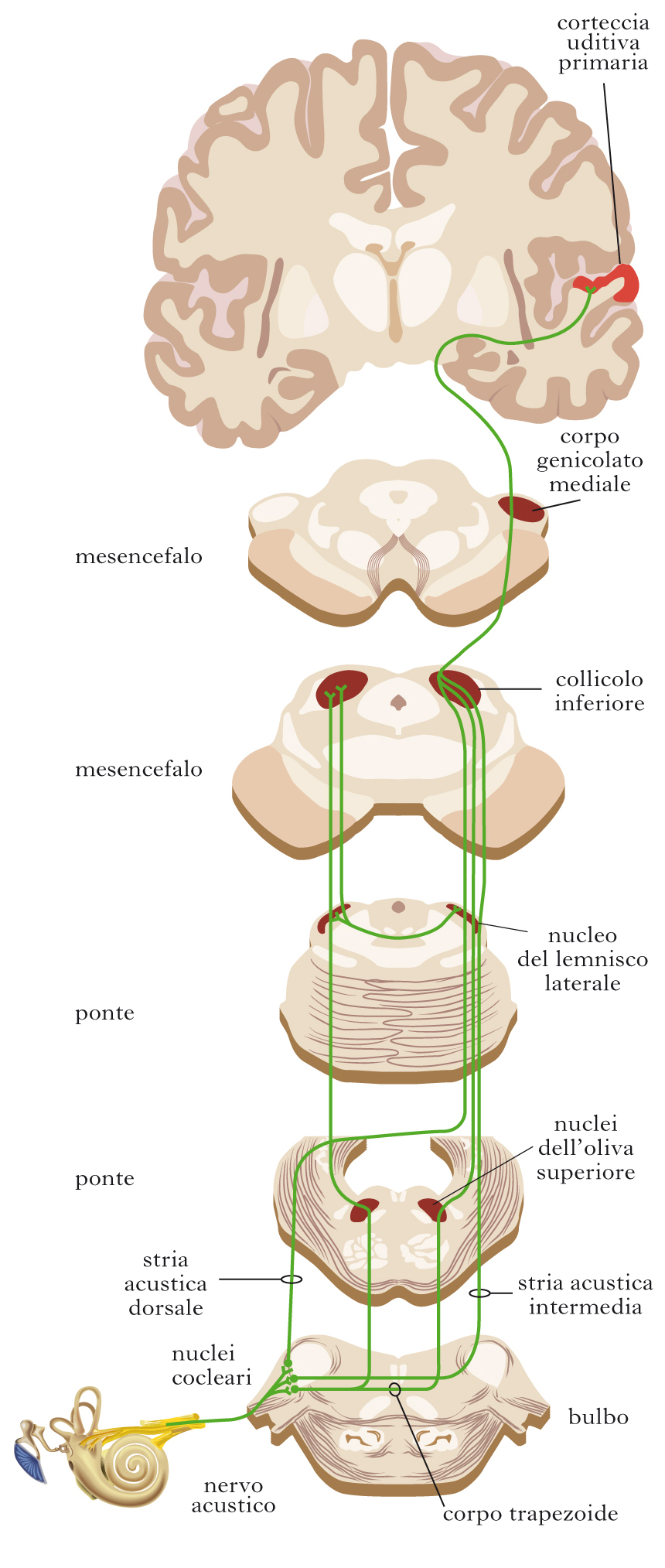
Il sistema uditivo centrale
Data la sua complessità, qui verranno prese in considerazione solo le strutture principali del sistema uditivo. Le informazioni acustiche vengono trasmesse al cervello dalle cellule ciliate tramite i neuroni del ganglio cocleare (situato all’interno del sottile strato osseo che ricopre la coclea), i cui assoni si riuniscono a formare il nervo cocleare, un ramo del nervo statoacustico. Ogni neurone prende contatto con una sola cellula ciliata e questo fa sì che l’organizzazione tonotopica dell’organo del Corti venga mantenuta nelle vie uditive. Essa è presente in tutte le strutture del sistema uditivo qui esaminate. Le fibre della branca cocleare del nervo statoacustico vanno a terminare nei nuclei cocleari del tronco dell’encefalo, i cui neuroni inviano i propri assoni ad altri centri cerebrali attraverso tre vie principali: la stria acustica dorsale, la stria acustica intermedia e il corpo trapezoide. Il fascio più cospicuo di fibre ascendenti è il corpo trapezoide, che veicola i segnali elettrici ai nuclei dell’oliva superiore, parimenti localizzati nel tronco encefalico. Le prime interazioni tra le informazioni che provengono dall’orecchio destro e da quello sinistro hanno luogo a livello di questa struttura, per la decussazione (incrocio da una parte all’altra del sistema nervoso) di una parte delle fibre che appartengono al corpo trapezoide. Gli assoni dei neuroni olivari, insieme ad assoni provenienti dai nuclei cocleari (stria acustica dorsale e intermedia), proiettano, attraverso il lemnisco laterale, al collicolo inferiore del mesencefalo. Da qui le vie ascendenti del sistema uditivo si estendono al corpo genicolato mediale del talamo. Anche se ci si riferisce spesso alle componenti sottocorticali del sistema uditivo come a semplici nuclei di collegamento, esse fanno molto di più che trasmettere passivamente l’informazione dal nervo cocleare alla corteccia uditiva: è stato dimostrato, infatti, che i nuclei del tronco dell’encefalo rivestono una grande importanza nella localizzazione delle sorgenti dei suoni. Per quanto riguarda il collicolo inferiore e i nuclei talamici, invece, il significato funzionale di queste regioni resta ancora incerto, ma è stato osservato che si tratta di strutture di straordinaria complessità che ricevono afferenze anche da altri sistemi sensoriali (in partic., da quello visivo) ed è quindi possibile che fungano da stazioni di integrazione delle diverse modalità sensoriali. Le fibre che hanno origine nel nucleo genicolato mediale del talamo terminano a livello della corteccia cerebrale. La proiezione talamica più cospicua raggiunge l’area uditiva primaria, che corrisponde all’area 41 di Brodmann ed è localizzata sulla superficie laterale del lobo temporale; questa regione, tuttavia, è circondata da numerose altre aree che rispondono agli stimoli acustici e si pensa siano devolute all’analisi di aspetti diversi delle informazioni uditive. Uno degli aspetti più importanti della percezione uditiva dell’uomo risiede nel suo ruolo nell’analisi del linguaggio (➔). Le conoscenze riguardanti le modalità di elaborazione dei suoni che costituiscono il linguaggio sono molto limitate, anche se lo studio delle lesioni cerebrali che producono afasie selettive e l’introduzione delle tecniche di visualizzazione dell’attività nervosa, in partic. la risonanza magnetica funzionale, hanno iniziato a fornire informazioni sulle aree coinvolte.
Percezione dell’intensità e discriminazione delle frequenze
Mentre la percezione dell’intensità dei suoni è semplicemente correlata ai livelli di attività che essi inducono nelle fibre del nervo acustico, l’analisi delle frequenze che compongono gli stimoli sonori avviene in maniera leggermente più complessa. È stato osservato che ogni assone risponde in maniera preferenziale alla stimolazione da parte di una particolare frequenza acustica, che costituisce la frequenza caratteristica di una certa fibra del nervo statoacustico, e la sua localizzazione all’interno del nervo è in rapporto con la rispettiva frequenza caratteristica. Si configura in questo modo un codice di posizione, secondo cui il sistema nervoso è in grado di discriminare le frequenze in relazione ai siti di attivazione neuronale indotti da un determinato stimolo. È stato osservato che anche il pattern di attività delle fibre del nervo statoacustico può codificare le informazioni relative alla tonalità dello stimolo: infatti, le fibre scaricano con una frequenza che è correlata con quella dello stimolo. In questo caso si parla di codice di frequenza. La codificazione in frequenza riveste una particolare importanza quando l’intensità dei suoni è tanto elevata da saturare la risposta dei neuroni che hanno come frequenze caratteristiche quelle che compongono i suddetti segnali. Le singole fibre, infatti, pur essendo sintonizzate su una frequenza preferita, rispondono anche a stimoli di frequenza leggermente maggiore o minore, quando essi sono presenti a intensità elevate. Nel caso di suoni particolarmente intensi, dunque, rispondono fibre aventi frequenze caratteristiche diverse, ma l’andamento temporale della loro frequenza di scarica fornisce ugualmente informazioni circa la frequenza dello stimolo. La discriminazione degli stimoli acustici ha, dunque, inizio nelle strutture periferiche del sistema uditivo, ma a livello corticale la sintesi delle informazioni, e probabilmente ulteriori livelli di analisi, determinano l’instaurarsi della percezione cosciente.
Localizzazione dei suoni
L’apparato uditivo è in grado di localizzare le sorgenti da cui provengono le onde sonore in maniera molto precisa, in partic. rispetto all’asse azimutale (ossia l’asse orizzontale, rispetto a cui si distinguono i suoni che provengono da destra e da sinistra). Questa possibilità deriva in gran parte dall’analisi dei ritardi delle informazioni acustiche. I suoni generati da una sorgente localizzata, per es. alla sinistra della testa, raggiungono l’orecchio sinistro prima di quello destro: il tempo che intercorre tra il rilevamento del suono da parte delle due orecchie è detto ritardo interaurale. È chiaro che si tratta di tempi molto piccoli, ma specializzazioni neuronali devolute a questa funzione fanno sì che la sensibilità ai ritardi interaurali sia nell’ordine dei 10 μs, che corrisponde a una capacità di localizzare la provenienza dei suoni con l’approssimazione di qualche grado. Questa discriminazione temporale non dipende da un’analisi complessa, ma fa semplicemente assegnamento sul ritardo con cui i potenziali d’azione che devono segnalare la presenza dei suoni arrivano ai nuclei del complesso dell’oliva superiore, e in partic. al nucleo olivare mediale. Per continuare con l’esempio, il suono che arriva all’orecchio sinistro viene trasformato in energia elettrica dall’organo del Corti e questa viene trasmessa, tramite il nervo statoacustico e i nuclei cocleari, fino al nucleo olivare mediale; lo stesso suono dà origine a una serie simile di eventi anche nell’orecchio destro, solo con qualche istante di ritardo. I neuroni dell’oliva superiore ricevono afferenze da entrambe le orecchie e possono, quindi, fungere da ri levatori di coincidenza, attivandosi solo quando i segnali provenienti dalle due parti li raggiungono simultaneamente. Questa struttura presenta un’organizzazione delle connessioni neurali provenienti dalle due orecchie tale per cui a una particolare posizione all’interno del nucleo corrisponde un gruppo di neuroni che danno risposta massimale a un ben definito ritardo interaurale. Il nucleo contiene, quindi, una mappa dei ritardi interaurali, che corrisponde a una mappa della localizzazione dei suoni lungo l’asse azimutale. Anche in questo caso le informazioni vengono trasdotte alla corteccia uditiva tramite le successive stazioni di ritrasmissione secondo un codice di posizione. Il sistema uditivo si avvale, però, di un’ulteriore modalità di elaborazione degli stimoli per valutare l’origine dei suoni, in cui impiega elementi collegati alla loro intensità. A causa dei fenomeni di assorbimento dei suoni da parte dei tessuti della testa, un suono che raggiunge l’orecchio più vicino alla sua sorgente è di intensità maggiore rispetto a quello che viene percepito dall’orecchio del lato opposto. In questo meccanismo svolge un ruolo essenziale un altro nucleo del complesso dell’oliva superiore, ossia il nucleo laterale, i cui neuroni rispondono preferenzialmente quando l’intensità dello stimolo sonoro che arriva a un orecchio è maggiore di una certa entità di quella recepita dall’altro orecchio. Anche questo nucleo è organizzato secondo una mappa che prevede le possibili localizzazioni delle sorgenti acustiche lungo l’asse orizzontale, in questo caso codificate dalle differenze di intensità del suono percepite dalle due orecchie.
Riconoscimento dell’identità della sorgente sonora
Un’altra funzione fondamentale della percezione uditiva è riconoscere l’identità delle sorgenti dei suoni, e quindi il loro significato. Il nostro sistema uditivo subisce un bombardamento pressoché continuo di informazioni acustiche, che si traduce in un pattern di attività in costante cambiamento negli assoni che compongono il nervo statoacustico. È evidente che la scomposizione delle frequenze degli stimoli sonori e la percezione della loro intensità non sono sufficienti per classificare un suono come il cinguettio di un uccellino piuttosto che come la melodia di un flauto. Questo stadio di elaborazione delle informazioni uditive è certamente a carico dei circuiti della corteccia cerebrale, ma si è ancora lontani dal comprendere quali siano i meccanismi neurali che operano a questo livello. Le conoscenze relative ai tipi di analisi compiute dalla corteccia uditiva si limitano per ora all’identificazione di gruppi di neuroni che rispondono a caratteristiche complesse degli stimoli, come l’inizio o la fine di un suono (o entrambi), le variazioni di tonalità, le variazioni di intensità, o a stimoli che contengono molte frequenze, ma sono necessari ulteriori studi per comprendere come la codifica delle proprietà complesse dei suoni si traduca nella percezione e nella classificazione di questi ultimi.
Orecchio assoluto
La capacità di riconoscere l’altezza di una nota, anche dopo averla ascoltata una sola volta, si chiama orecchio assoluto. È stato osservato che questa abilità deriva da un intenso addestramento nella pratica musicale; non tutti i musicisti, tuttavia, possono vantare tale attitudine, per cui è possibile che esistano predisposizioni innate (➔ musica e cervello).
Sordità
Nella maggior parte dei casi la sordità dipende dalla perdita delle cellule ciliate della coclea, che si può verificare sia nei bambini sia negli individui adulti. La sordità precoce può essere congenita ed essere conseguenza di patologie genetiche o problemi prenatali, o acquisita durante l’infanzia per cause infettive, tossiche o traumatiche. Per quanto riguarda la sordità che insorge in età più tardiva, le possibili cause includono infezioni, traumi cranici, l’assunzione di farmaci ototossici, problemi circolatori, cause neurologiche. Le conseguenze di questo deficit funzionale sono gravi; per questo motivo sono importanti i progressi registrati negli ultimi anni nel campo del trattamento della sordità, in partic. con lo sviluppo di apparecchi acustici amplifilicatori e protesi cocleari che in molti casi sono in grado di compensare (almeno parzialmente) l’insufficienza sensoriale. Laura Baroncelli