
Udito
Udito
La funzione sensoriale preposta alla percezione del suono è presente in larga parte delle specie animali, dagli Insetti ai Mammiferi, sebbene le strategie di percezione siano differenti. Nella specie umana e negli altri Vertebrati, l'organo alla base delle capacità uditive è l'orecchio. La questione dell'origine evolutiva dell'orecchio dei Vertebrati, ovvero di come esso si sia evoluto dalle forme primitive fino a quelle dei Mammiferi moderni, è aperta e controversa, sebbene la storia evolutiva del sistema uditivo sia stata notevolmente dettagliata rispetto a quella degli altri sistemi sensoriali. Ciò è stato possibile in quanto le strutture recettoriali uditive di Vertebrati estinti erano situate entro una struttura ossea che in molti casi è andata incontro a un processo di fossilizzazione. Lo studio di tali reperti ha fornito numerose informazioni su come l'organo deputato alla percezione uditiva fosse strutturato nel corso dell'evoluzione. Una delle ipotesi più accreditate suggerisce che l'orecchio si sia evoluto da un sistema inizialmente vestibolare, evolutosi a sua volta dalla cosiddetta 'linea laterale' presente in molte specie di Pesci e di Anfibi. Tuttavia, le moderne tecniche anatomiche e neurofisiologiche indicano che, sebbene la linea laterale e l'orecchio possano condividere un antenato comune, si tratta di due sistemi distinti, che non sono in relazione in termini evolutivi. Inoltre, le teorie più recenti sostengono che gli organi di senso responsabili della percezione dei suoni si siano evoluti più volte e in maniera indipendente nel corso della storia dei Vertebrati.
Il suono consiste nella modificazione della pressione di un mezzo elastico solido, liquido o gassoso: nella nostra esperienza generalmente aria. Qualsiasi evento che produce un suono comprime le particelle d'aria con una pressione superiore a quella atmosferica. Tali modificazioni della pressione, note come 'onde sonore', si irradiano nel mezzo di propagazione fino a raggiungere l'organo deputato alla percezione del suono. Il suono più semplice, detto 'tono puro', vibra a un'unica frequenza, data dalla velocità dell'alternanza di compressione ed espansione dell'aria. La frequenza di un dato segnale acustico ne determina la tonalità. Una seconda variabile che definisce un suono è l'ampiezza, o intensità, data dalla distanza percorsa dall'onda sonora in un determinato periodo di tempo e normalmente misurata come pressione. L'ampiezza è interpretata dall'orecchio come volume.
L'ambiente acustico può essere composto di un numero assai elevato di suoni. Tuttavia, la capacità di percepirli è selettiva, e cambia da specie a specie e da individuo a individuo. Essa è determinata dalle caratteristiche fisiologiche dell'organo dell'udito, che definiscono l'intervallo di frequenze percepite e la minima pressione rilevata. Nel caso della specie umana, l'ambito di frequenza percepito è di 0,02÷20 kHz, e la pressione in grado di stimolare una risposta varia da 20×10‒6 a 20 Pa, sebbene negli individui più anziani tali capacità percettive siano ridotte. E' interessante notare come le caratteristiche delle capacità di percezione uditiva siano state disegnate dalla storia evolutiva della specie. Per esempio, nell'uomo, i suoni percepiti con maggiore efficacia includono quelli del pianto del neonato e quelli del linguaggio. In entrambi i casi, è possibile ipotizzare un chiaro vantaggio evolutivo.
L'udito negli invertebrati
I segnali acustici svolgono un ruolo importante nel mediare numerosi comportamenti nel regno animale degli invertebrati. Tra questi, gli Insetti hanno probabilmente evoluto i recettori più elaborati per la ricezione del suono. Numerose specie appartenenti a questa classe sono infatti in grado di ottenere da segnali acustici informazioni riguardanti, per esempio, l'identità e il luogo di provenienza di potenziali predatori, prede, partner o rivali. L'etologia, ovvero lo studio del comportamento, ha mostrato come alcune specie di Insetti sociali, quali grilli e cicale, ma anche il moscerino della frutta (la drosofila), producano suoni che hanno la chiara funzione comunicativa di attrarre i potenziali partner. Altre specie di Insetti, invece, grazie alla funzione uditiva evitano di essere predate. Le falene, per esempio, possono captare gli ultrasuoni emessi dai pipistrelli, loro predatori, e di conseguenza attuano delle strategie comportamentali per cercare di evitarne l'attacco. Inoltre, è stato dimostrato sperimentalmente che anche altri gruppi di Insetti, dai Lepidotteri (le farfalle) ai Ditteri (per es., le mosche), posseggono il senso dell'udito.
Oltre agli studi etologici, anche l'analisi morfologica ha dimostrato come nella classe degli Insetti esistano diverse strutture anatomiche deputate alla ricezione dei suoni, la più semplice delle quali è probabilmente l'organo cordotonale. Esso è formato da due o più scolopidi, i quali a loro volta consistono in alcune cellule di rivestimento e in una cellula gangliare, che è provvista di un pelo in grado di vibrare e che è connessa, mediante una fibra nervosa, ai gangli del sistema nervoso centrale. È interessante notare come tali strutture siano morfologicamente molto simili ai recettori tattili, suggerendo che la percezione del suono possa costituire un affinamento del senso tattile. Gli organi cordotonali si sono evoluti in diverse forme, la più nota delle quali è probabilmente l'organo di Johnston, situato sul secondo articolo delle antenne di molte specie di Insetti. Nelle zanzare, questi apparati permettono il riconoscimento specie-specifico della frequenza del battito delle ali delle femmine. Nelle api, l'organo di Johnston svolge un ruolo nella percezione dei segnali acustici che vengono emessi durante la danza con cui gli individui comunicano ai conspecifici la posizione delle fonti di cibo.
Un secondo tipo di recettore presente negli Insetti, come cavallette e cicale, è l'organo timpanico, che è caratterizzato dalla capacità di percepire i cambiamenti della pressione dell'aria dovuti a segnali acustici. Gli organi timpanici, localizzati in parti diverse del corpo (come arti, metatorace e addome, a seconda delle specie), sono costituiti da una sottile membrana, detta 'timpano', che è posta sulla superficie esterna del corpo e tesa sopra una cavità piena d'aria. Le deformazioni della superficie della membrana sono rilevate da un gruppo di scolopidi che sono in comunicazione con il sistema nervoso centrale.
L'intervallo di frequenze rilevato dagli organi timpanici è ampio, di 0,3÷20 kHz nelle cavallette, sebbene la sensibilità sia più elevata per le frequenze più alte. Altre specie di Insetti sono in grado di percepire anche ultrasuoni, ovvero suoni che superano la soglia uditiva umana dei 20 kHz. Le locuste, per esempio, sono in grado di percepire suoni della frequenza di 30 kHz, che corrisponde alla frequenza di emissione dei segnali prodotti dal loro stesso apparato stridulante.
L'udito nei Vertebrati
Tutti i Vertebrati posseggono abilità uditive che in generale sono notevolmente più raffinate di quelle degli invertebrati, per quanto riguarda sia la capacità di discriminazione sia la quantità d'informazioni raccolta da ciascun segnale sonoro. Queste abilità sono dovute non solo alla maggiore complessità degli organi deputati alla percezione dei segnali acustici, ma anche alla più raffinata capacità di elaborare tali segnali da parte del sistema nervoso centrale.
Nei Pesci, le onde sonore, che in questo caso consistono in modificazioni della pressione del mezzo acquatico, sono percepite mediante due sistemi: l'orecchio e la linea laterale. Sebbene in questa classe di animali non sia presente un orecchio esterno, le vibrazioni sonore sono trasmesse attraverso le ossa del cranio fino a raggiungere un orecchio interno. In alcune specie, la vescica natatoria funge da struttura risonante, amplificando il segnale acustico che viene trasmesso all'orecchio interno da apposite strutture anatomiche. Nei Pesci ossei (Teleostei), l'organo uditivo è collocato sul retro del cranio, in una capsula ossea. L'orecchio interno è costituito dal labirinto, il quale è formato da 3 canali semicircolari, perpendicolari gli uni agli altri e collegati a una struttura sacciforme, detta 'utricolo', che è collegata a sua volta, nella parte inferiore, a una seconda struttura sacciforme, detta 'sacculo'. Adiacente al sacculo vi è una struttura incurvata, la lagena, provvista di un addensamento di cellule sensoriali, o macula, le cui ciglia sono connesse a una struttura calcarea densa, detta 'otolito'. Le onde sonore si propagano attraverso il mezzo acquatico e fanno vibrare in modo uniforme tutti i tessuti tranne l'otolito, generando in tal modo una differenza tra i movimenti delle cellule della macula e quelli dell'otolito: questa differenza viene trasdotta in segnale elettrico, e viaggia lungo le fibre nervose fino al sistema nervoso centrale per dare forma alla sensazione uditiva.
La seconda struttura sensoriale di cui sono dotati i Pesci è la linea laterale. Tale dispositivo permette di rilevare suoni di frequenza molto bassa, al di sotto dei 200 Hz, sebbene la sua funzione primaria sia quella di rilevare gli spostamenti compiuti dall'animale in seguito ai suoi stessi movimenti. Si tratta di un organo indispensabile affinché i Pesci che vivono in branco possano mantenerne coesa la formazione. Una particolare forma di tale organo è rappresentata dal canale della linea laterale, da cui si è ipotizzato possano essere derivati i canali semicircolari interni al corpo.
I Pesci hanno capacità uditive meno raffinate rispetto agli altri Vertebrati, potendo percepire unicamente suoni prodotti in un intervallo di frequenze di 0,16÷6 kHz circa. Le specie di Pesci che hanno un udito più sviluppato comprendono, tra gli altri, i Ciprinidi che, come le altre specie appartenenti al gruppo tassonomico degli Otophysi (Cypriniformes, Characiformes, Siluriformes e Gymnotiformes), sono provvisti dell'apparato di Weber. Questo consiste in una modificazione delle prime quattro o cinque vertebre, le quali, mettendo in contatto la vescica natatoria con l'orecchio, rendono più efficiente la trasmissione del suono.
Nel passaggio evolutivo dai Pesci agli Anfibi, oltre alla lagena si sviluppa un'ulteriore struttura sacciforme, detta 'papilla basilare', che nei Rettili e negli Uccelli si allunga e nei Mammiferi prende la forma di un canale spiraliforme e diviene la 'coclea'. Negli Anfibi Anuri, al contrario dei Pesci, l'orecchio è aperto all'esterno, ed è costituito da un apparato interno e da uno medio. Negli Anfibi l'orecchio medio consta di un solo ossicino, la columella, mentre nei Rettili e negli Uccelli consta della staffa e dell'extracolumella. In queste ultime due classi di animali è presente anche un orecchio esterno, benché non caratterizzato da un padiglione auricolare come nei Mammiferi.
L'orecchio dei Mammiferi
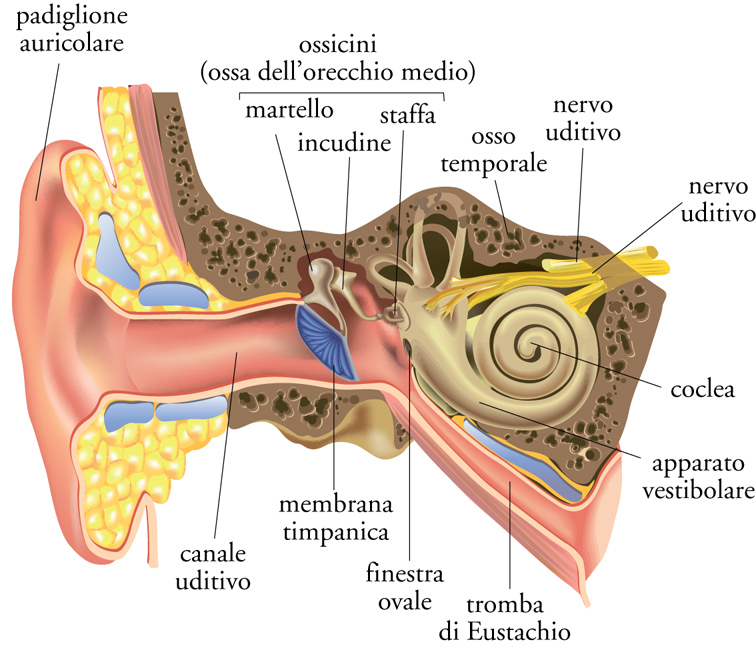
È interessante notare come le più importanti scoperte sull'anatomia e la fisiologia dell'udito compiute fino al XIX sec. siano state fatte quasi tutte in Italia. Per esempio, le descrizioni e i disegni delle strutture dell'orecchio fatti da Vesalio nel XVI sec. costituiscono un capolavoro dello studio anatomico del corpo umano. A partire dal XX sec., numerosi ricercatori europei e statunitensi hanno fornito un rilevante contributo al chiarimento della neurofisiologia dell'udito, e tra questi si è distinto l'ungherese Georg von Békésy, che per le sue ricerche è stato insignito del Premio Nobel. L'organo dell'udito dei Mammiferi può essere convenientemente suddiviso in tre parti: orecchio esterno, orecchio medio e orecchio interno.
Orecchio esterno. È costituito dal padiglione auricolare, ovvero dalla struttura che comunemente viene chiamata orecchio, e dal canale che conduce al timpano. Il padiglione è presente unicamente nei Mammiferi e gioca un ruolo cruciale nella percezione del suono in quanto la sua superficie, caratterizzata da aree concave e convesse, modifica le caratteristiche acustiche del segnale, amplificandone o attenuandone selettivamente l'energia in specifici intervalli di frequenza. Per tale motivo, la combinazione di elementi che costituiscono questa struttura è assai varia nel mondo animale; i Chirotteri (e in particolare i Microchirotteri) rappresentano forse l'esempio più noto della incredibile varietà e originalità di forme che può assumere il padiglione auricolare. Al fine di percepire meglio i suoni, molte specie animali sono anche in grado di muoverlo e direzionarlo verso la sorgente sonora. Nella specie umana, il padiglione ha una conformazione tale da amplificare i suoni in un intervallo di frequenza di 2÷5 kHz circa, ovvero quelli corrispondenti alla gamma importante per il linguaggio parlato.
Orecchio medio. La membrana timpanica (timpano) separa l'orecchio esterno da quello medio. Quest'ultimo è costituito da un insieme di strutture ossee e muscolari. In particolare, tre ossicini, chiamati 'martello', 'incudine' e 'staffa' (le più piccole ossa del corpo), mettono in comunicazione la membrana timpanica con la finestra ovale, trasducendo l'energia della pressione esercitata dal segnale sonoro. Poiché la superficie della membrana timpanica è notevolmente più estesa di quella della finestra ovale, tale struttura produce un'amplificazione dell'energia meccanica della pressione sonora, al fine di mettere in movimento il fluido che si trova nella coclea dell'orecchio interno. I due muscoli dell'orecchio medio sono il tensore del timpano, che è collegato al martello, e lo stapedio, che è collegato alla staffa. Essi svolgono la funzione di ridurre l'energia del segnale acustico, anche al fine di proteggere le delicate strutture dell'orecchio interno da rumori troppo forti e quindi potenzialmente dannosi. In particolare, ne provocano la contrazione i suoni che superano di circa 80÷100 dB la soglia uditiva. Tale contrazione avviene anche per favorire la percezione selettiva di una specifica gamma di frequenze: agendo come un filtro, essa permette infatti di bloccare i suoni con frequenze inferiori a circa 1 kHz.
Orecchio interno. Questa parte ha la funzione di convertire l'energia meccanica dei suoni in attività neuronale. Essa è costituita dalla coclea e dall'apparato vestibolare. Quest'ultimo svolge un ruolo che non concerne l'udito, ovvero quello di rilevare informazioni relative alla posizione e al movimento del corpo nello spazio, al fine di mantenere la postura e permettere la coordinazione del movimento, mentre la coclea è la sede in cui i suoni sono trasformati in impulsi nervosi. La coclea è caratterizzata da una struttura a spirale, la cui base è adiacente alla membrana della finestra ovale, che rappresenta il confine tra l'orecchio medio e quello interno. La parte più distale è invece chiamata apice. L'interno della coclea è suddiviso in tre parti: il canale vestibolare, il canale del timpano e il dotto cocleare; quest'ultimo contiene la membrana basilare, una struttura elastica che separa il dotto cocleare dal canale timpanico e che vibra in risposta ai suoni. Fra la membrana basilare e il dotto cocleare si trova l'organo del Corti, che prende il nome da Alfonso Corti, che lo scoprì e ne studiò la funzione a metà del XIX secolo. All'interno di tale organo sono presenti caratteristiche cellule ciliate, e le terminazioni delle fibre del nervo acustico che prendono contatto con la base di queste cellule mediante sinapsi.
In seguito a un'onda sonora, la staffa vibra avanti e indietro, esercitando una pressione variabile sulla finestra ovale e quindi sul liquido incomprimibile presente nel canale vestibolare e nel canale timpanico. Tale pressione provoca a sua volta movimenti oscillatori della membrana basilare, stimolando le cellule ciliate presenti nell'organo del Corti. È interessante notare come la struttura della membrana basilare, più larga verso la base e più stretta verso l'apice della coclea, sia tale che, a seconda della frequenza del segnale acustico percepito, solo una parte specifica della membrana, e quindi della popolazione di cellule ciliate, venga stimolata. In particolare, i segnali ad alta frequenza stimolano in misura maggiore la membrana basilare nella regione in cui essa è stretta, mentre quelli a bassa frequenza la stimolano nella regione in cui essa è più larga. Una conseguenza dell'esistenza di tale meccanismo, che rende possibile una prima analisi sofisticata del segnale acustico percepito, è che le caratteristiche specie-specifiche della forma della membrana basilare determinano l'ambito di frequenze che ciascuna specie è in grado di rilevare. Studi recenti hanno mostrato come ulteriori meccanismi di tipo elettromeccanico contribuiscano, nei Mammiferi e più in particolare nella specie umana, a dare forma alla raffinata potenzialità di sintonizzare e quindi di percepire frequenze differenti.
Verso la fine del XX sec., sono stati caratterizzati i processi che permettono agli eventi meccanici che si verificano nella coclea di essere trasdotti nell'eccitazione delle fibre nervose acustiche. È stato osservato come i movimenti del liquido nella coclea che producono vibrazioni della membrana basilare stirino i fasci di ciglia delle cellule dell'organo del Corti. Anche spostamenti minimi delle ciglia determinano un effetto sull'apertura e chiusura dei canali ionici delle loro cellule. Le modificazioni dei canali ionici provocano l'eccitazione delle cellule ciliate e, conseguentemente, dei neuroni che costituiscono le fibre afferenti.
Da ciascuna coclea emergono circa 40.000 fibre uditive, che costituiscono la porzione uditiva dell'ottavo nervo cranico. Ciascuna fibra uditiva ramifica e innerva i nuclei cocleari dorsali e ventrali. Questi proiettano a loro volta al nucleo olivare superiore nel tronco encefalico. Tale struttura riceve informazioni sia dall'orecchio sinistro sia da quello destro, e perciò rappresenta la prima stazione di integrazione per l'ascolto binaurale. Successivamente, l'informazione arriva al collicolo inferiore, che costituisce il centro uditivo del mesencefalo, per poi raggiungere il nucleo genicolato attraverso il talamo, e infine la corteccia uditiva situata nella circonvoluzione temporale superiore vicino alla scissura di Silvio. Nella specie umana, e anche in altri Mammiferi, lungo il percorso dalla coclea alla corteccia, i neuroni che costituiscono le vie uditive presentano un alto grado di organizzazione, detta 'tonotopica', che ne riflette la sensibilità alle diverse frequenze. Per esempio, le cellule sensibili alle alte frequenze sono vicine tra loro e distanti da quelle sensibili alle basse frequenze. (fig. 1)
La percezione del suono nello spazio
Gli organi deputati alla percezione dei segnali acustici nei Vertebrati sono generalmente due. Sebbene l'ascolto con un solo orecchio, detto 'monoaurale', sia sufficiente per la localizzazione della posizione della sorgente sonora nel caso in cui si abbia la possibilità di ruotare il capo, l'utilizzo di entrambe le orecchie permette di eseguire lo stesso compito con maggiore facilità e accuratezza. In particolare, l'efficienza superiore dell'ascolto binaurale è dovuta alla possibilità di cogliere una differenza tra le due orecchie, concernente sia il tempo sia l'intensità con cui il suono viene percepito. A volte la differenza di intensità è dovuta al fatto che la testa proietta un'ombra sonora, schermando una delle due orecchie. Tale effetto è presente solo per i segnali ad alta frequenza, la cui onda sonora, essendo corta, non riesce ad 'aggirare' la testa, come fa invece quella dei suoni a bassa frequenza. Infatti, l'identificazione della sorgente dei suoni a bassa frequenza è più difficile, e si basa principalmente sulle differenze nel tempo di arrivo del segnale alle due orecchie.
Per l'ascolto binaurale, le informazioni provenienti dalle due orecchie vengono integrate nel sistema nervoso centrale a partire dal tronco encefalico, a livello del complesso olivare superiore. Studi approfonditi, finalizzati alla comprensione dei substrati neurali della capacità di localizzare una sorgente sonora, sono stati condotti sul barbagianni (Tyto alba) mediante un approccio integrato comportamentale, fisiologico e anatomico. Questa specie è infatti assai abile nell'identificazione della posizione di sorgenti sonore, dato che la sua sopravvivenza dipende dalla caccia notturna, in volo, di prede quali piccoli Mammiferi. Il barbagianni può localizzare una sorgente sonora con una precisione di circa 1-2 gradi sia in azimut sia in elevazione, nel campo di frequenze tra 3 e 9 kHz. È stato dimostrato come queste sue abilità siano associate a una struttura del sistema nervoso assai raffinata, che consiste in rappresentazioni neuronali dello spazio acustico. In particolare, nella regione del collicolo inferiore, i neuroni sono organizzati in modo da rispondere a una rappresentazione sferica dello spazio acustico, con i suoni provenienti da una particolare direzione che stimolano selettivamente alcuni neuroni e i suoni provenienti da sorgenti vicine che stimolano i neuroni vicini. Inoltre, questa mappa acustica si combina con quella visiva del tetto ottico, integrando l'informazione proveniente dai due sensi. Ogni neurone del tetto ottico risponde infatti a stimoli visivi o acustici provenienti da una particolare posizione nello spazio, detta 'campo ricettivo'. Tale mappa spaziale fornisce l'informazione su cui si basano le risposte motorie del barbagianni in relazione alla posizione di una preda che produce suoni.
La sordità e i disturbi uditivi
I deficit della funzione uditiva sono denominati 'sordità' o 'ipoacusie'. La sordità e i disturbi a essa associati costituiscono un serio problema medico e sociale, che colpisce circa il 10% della popolazione mondiale. Tale percentuale è più alta nelle nazioni povere a causa della mancanza di igiene, prevenzione e terapie. La sordità si può presentare con diversi livelli di gravità: da una riduzione della soglia uditiva selettiva e parziale che può rendere problematica la comprensione del linguaggio (riduzione della sensibilità di circa 40 dB) fino alla perdita totale della funzione uditiva (riduzione della sensibilità di oltre 90-100 dB). Una sordità grave presente fin dalle prime fasi dello sviluppo, dovuta per esempio a cause genetiche, può avere conseguenze sulle abilità linguistiche dell'individuo una volta adulto. Infatti si impara a parlare ascoltando, e la macanza di questa possibilità può avere pesanti ripercussioni sullo sviluppo del linguaggio.
In base alla porzione del sistema uditivo che è colpita dalla patologia e ai relativi problemi funzionali, le sordità vengono suddivise schematicamente in tre categorie principali: (a) la sordità di trasmissione o conduzione, causata da alterazioni che riguardano l'orecchio esterno e/o quello medio, e normalmente dovuta a malformazioni, traumi o infiammazioni dell'apparato di trasmissione dei suoni. Gli ossicini dell'orecchio medio rappresentano il sito più frequentemente affetto. Queste patologie provocano una diminuzione limitata delle capacità uditive, che in genere non supera i 50÷55 dB e che riguarda maggiormente l'intervallo di frequenze più alto; (b) la sordità neurosensoriale, generalmente dovuta a una perdita di funzionalità dell'orecchio interno, e sicuramente la più frequente (rappresenta circa il 70-80% dei casi di ipocusia). Gli individui affetti da questo tipo di perdita uditiva sentono in modo impreciso e non riconoscono i suoni. Tra le cause si annovera l'esposizione a farmaci, e in particolare ad antibiotici quali la streptomicina. Tali sostanze ototossiche (che danneggiano l'orecchio) entrano nelle cellule ciliate passando attraverso i canali ionici e attaccano i mitocondri, compromettendo a volte irrimediabilmente la funzionalità cellulare. Generalmente, la regione basale della coclea è la più vulnerabile agli antibiotici e ad altri medicinali, quali per esempio l'aspirina, e di conseguenza la perdita uditiva riguarda primariamente le alte frequenze. La sordità neurosensoriale può essere causata anche dall'esposizione a un rumore improvviso e particolarmente intenso, come un'esplosione, oppure da un'esposizione prolungata e continua ad ambienti rumorosi, quali le discoteche. In queste condizioni, le cellule ciliate sono traumatizzate da un prolungato e ripetuto affaticamento cocleare indotto dal rumore. Una strategia per risolvere il danno alla coclea, recentemente sviluppata con successo e sempre più diffusa, è il trapianto cocleare, in cui viene inserita al posto della coclea danneggiata una protesi elettronica che ne svolge la funzione. Altri approcci terapeutici sono in via di sviluppo. Per esempio, è stato osservato come gli Uccelli siano in grado di rigenerare le cellule ciliate perdute in seguito a traumi o esposizioni ad agenti tossici e, recentemente, risultati simili sono stati ottenuti sui Mammiferi. Sebbene una terapia non sia ancora stata messa a punto, le ricerche finalizzate a identificare strategie per stimolare la rigenerazione delle cellule ciliate come rimedio alla sordità nella specie umana sono molto promettenti; (c) la sordità centrale si presenta quando, nonostante le vibrazioni sonore siano correttamente recepite dall'orecchio e i relativi impulsi vengano generati, le vie uditive non sono in grado di trasmettere i segnali o il sistema nervoso centrale non elabora correttamente l'informazione. La neuropatia uditiva rappresenta una delle forme più note di sordità centrale. I soggetti che ne sono affetti presentano normali otoemissioni acustiche, ma la trasmissione del segnale dall'orecchio interno al cervello è compromessa. Altri esempi di sordità centrale sono rappresentati dall'agnosia uditiva, in cui è presente un'incapacità di identificare i suoni pur in presenza di una soglia uditiva normale, e l'agnosia uditiva verbale, che consiste nell'incapacità di decodificare il codice fonologico.
La percezione di infrasuoni e ultrasuoni
I suoni emessi e percepiti dagli animali possono andare oltre l'intervallo di frequenze che la specie umana è in grado di percepire, di 0,02÷20 kHz. I suoni al di sotto e al di sopra di tale intervallo sono detti, rispettivamente, 'infrasuoni' e 'ultrasuoni'.
Gli infrasuoni vengono normalmente prodotti e percepiti da specie terrestri e acquatiche di grandi dimensioni, tra cui gli elefanti (Elephas maximus, Loxodonta africana). Anche le balene comunicano con infrasuoni, che possono essere percepiti a centinaia di chilometri di distanza grazie all'eccellente capacità di propagazione, dovuta all'alta densità, del mezzo acquatico rispetto all'aria.
Lazzaro Spallanzani, naturalista, biologo e fisiologo, alla fine del Settecento aveva osservato l'abilità dei pipistrelli di evitare gli ostacoli al buio, e con una serie di rimarchevoli esperimenti aveva dedotto che essa non dipende dall'integrità della funzione visiva. Solamente negli anni Trenta del XX sec. i progressi della tecnologia hanno permesso di individuare gli ultrasuoni emessi da questi animali. I pipistrelli (Chiroptera) sono in grado di percepire l'ambiente circostante e di localizzare la preda utilizzando l'eco delle loro stesse emissioni vocali ad alta frequenza. La cosiddetta ecolocazione costituisce un sistema estremamente efficace per sondare l'ambiente, e i Chirotteri ne danno una dimostrazione brillante riuscendo a catturare fino a 5000 insetti in una sola notte. L'apparato uditivo di questi animali è molto sofisticato: esso è caratterizzato da grandi padiglioni auricolari finemente strutturati e consente un'efficiente ricezione degli echi riflessi al fine di ricostruire, mediante un'analisi dettagliata di questi ultimi, la forma e la posizione dell'ostacolo incontrato. A livello del sistema nervoso centrale, i Chirotteri presentano una serie di strutture e di vie neuronali altamente specializzate per l'analisi dell'ambiente acustico.
In particolare, i pipistrelli utilizzano tre diverse strategie uditive per ecolocalizzare l'ostacolo in ciascuna delle tre dimensioni. L'individuazione sul piano orizzontale è realizzata in base alla differenza tra le due orecchie nella percezione dell'eco. Infatti, la ricezione binaurale permette il calcolo della posizione nello spazio dell'oggetto-bersaglio rispetto all'azimut. La localizzazione sul piano verticale avviene mediante l'uso delle parti più esterne delle orecchie, chiamate pinnae. In molte specie, esse possono essere ruotate, normalmente in modo alternato (una in avanti e l'altra all'indietro e viceversa), in relazione con l'emissione del segnale. Tale accorgimento consente all'animale di percepire specifiche modificazioni dell'eco riflesso che sono indice dell'elevazione dell'oggetto-bersaglio. Nelle specie che non possono muovere le pinnae, la localizzazione verticale si avvale dell'analisi delle alterazioni spettrografiche prodotte dalla riflessione del segnale. La terza dimensione, ovvero la distanza, è individuata mediante la valutazione del tempo che il segnale acustico impiega per incontrare l'oggetto e ritornare alle orecchie dell'emettitore. Inoltre, l'integrazione di diverse misure di distanza e l'analisi dello spostamento Doppler ‒ cioè la variazione di frequenza del segnale dovuta alla differenza di velocità tra il pipistrello, che emette il segnale e ne analizza l'eco riflesso, e il bersaglio ‒ forniscono informazioni sulla velocità relativa della preda. L'analisi delle caratteristiche più fini dello spettrogramma fornisce ulteriori informazioni che permettono, per esempio, di discriminare una potenziale preda da un oggetto generico, o di individuare il tipo di preda. Ciascuna specie di insetto batte infatti le ali, durante il volo, con una cadenza regolare, che genera una periodica modulazione in frequenza nello spettro dell'eco riflesso. Il ritmo del battito d'ali, e conseguentemente la tipologia della modulazione in frequenza, sono specie-specifiche e quindi ‒ come un''impronta sonora' ‒ consentono al pipistrello di identificare il tipo di preda. L'importanza di tale parametro e, più in generale, dell'analisi fine dello spettrogramma dell'eco riflesso nel processo di ecolocazione, è confermata dal fatto che, durante la caccia, il chirottero ignora qualunque insetto che non sia in volo.
Gli ultrasuoni non sono utilizzati unicamente per individuare ostacoli o possibile prede, ma anche per non farsi individuare. Infatti, l'emissione di suoni rappresenta un pericolo, in quanto questi possono attirare l'attenzione di un predatore. Al fine di evitare ciò, diverse specie di Roditori prediligono le frequenze che sono al di sopra della soglia uditiva dei loro potenziali predatori. Gli ultrasuoni prodotti dai topi e dai ratti, per esempio, variano in un intervallo di 20÷70 kHz. I neonati di molte specie di Roditori emettono ultrasuoni per comunicare con la madre. Le alte frequenze si disperdono infatti molto rapidamente nell'aria ed è facile individuare la direzione dalla quale provengono. Mediante tali segnali acustici, il neonato può dunque richiamare l'attenzione della madre evitando nel contempo di attirare possibili predatori.
Bibliografia
Clark 1991: Clark, William, Noise exposure from leisure activities: a review, "Journal of the Acoustical Society of America", 90, 1991, pp. 175-181.
Corti 1851: Corti, Alfonso, Recherches sur l'organe de Corti de l'ouïe des mammifères, ‟Zeitschrift für wissenschaftliche Zoologie", 3, 1851, pp. 109-169.
Griffin 1965: Griffin, Donald R. - Friend, J.H. - Webster F.A., Target discrimination by the echolocation of bats, "Journal of experimental zoology", 158, 1965, pp. 155-168.
Hauser 1996: Hauser, Marc D., The evolution of communication, Cambridge (Mass.), MIT Press, 1996.
Hudspeth 1989: Hudspeth, A. James, How the ear's works work, "Nature", 341, 1989, pp. 397-404.
Hudspeth 1997: Hudspeth, A. James, Mechanical amplification of stimuli by hair cells, "Current opinion in neurobiology", 7, 1997, pp. 480-486.
Hudspeth 2000: Hudspeth, A. James e altri, Putting ion channels to work: mechanoelectrical transduction, adaptation, and amplification by hair cells, "Proceedings of the National Academy of Sciences USA", 97, 2000, pp. 11765-11772.
Knudsen 1981: Knudsen, Eric I., The hearing of the barn owl, "Scientific American", 245, 1981, pp. 82-91.
Konishi 1993: Konishi, Masakaru, Listening with two ears, "Scientific American", 268, 1993, pp. 34-41.
Lawrence, Simmons 1982: Lawrence, Beatrice D. - Simmons, James A., Echolocation in bats: the external ear and perception of the vertical positions of targets, "Science", 218, 1982, pp. 481-483.
Lefebvre 1993: Lefebvre, Philippe e altri, Retinoic acid stimulates regeneration of mammalian auditory hair cells, "Science", 260, 1993, pp. 692-693.
Moss, Zagaeski 1994: Moss, Cynthia F. - Zagaeski, Mark, Acoustic information available to bats using frequency modulated sonar sounds for the perception of insect prey, "Journal of the Acoustical Society of America", 95, 1994, pp. 2745-2756.
Von Békésy 1960: von Békésy, Georg, Experiments in hearing, New York-London, McGraw-Hill, 1960.
Webster 1992: The evolutionary biology of hearing, edited by Douglas B. Webster, Arthur N. Popper, Richard R. Fay, New York, Springer, 1992.
Wever 1974: Wever, Ernest G., The evolution of vertebrate hearing, in: Handbook of sensory physiology, edited by Wolf D. Keidel, William D. Neff, Berlin, Springer, 1974, pp. 423-454.