
Uomo: origine ed evoluzione
Uomo: origine ed evoluzione
di Phillip V. Tobias
Uomo: origine ed evoluzione
sommario: 1. Definizione di uomo. 2. Le caratteristiche di Pongidae. 3. Le caratteristiche di Hominidae: a) caratteristiche morfologiche principali degli Ominidi; b) principali tratti comportamentali degli Ominidi; c) aspetti della biologia molecolare dell'uomo. 4. Strategie adottate nello studio dell'evoluzione umana: a) la strategia impiegata per la fase dell'emergenza degli Ominidi; b) strategie impiegate per la fase dell'evoluzione degli Ominidi; c) strategie impiegate per la fase dell'evoluzione dell'uomo moderno. 5. L'origine di Hominidae. 6. L'evoluzione di Hominidae: a) la scoperta del primo esemplare di Australopithecus; b) il progressivo chiarimento di Australopithecus; c) definizione di Australopithecus; d) le specie di Australopithecus; e) la datazione di Australopithecus; f) caratteri comportamentali di Australopithecus; g) il posto occupato da Australopithecus nella filogenesi degli Ominidi. 7. La comparsa del genere Homo: il più antico Homo, Homo habilis: a) la scoperta dei primitivi Homo in Africa; b) la datazione di Homo habilis; c) alcuni caratteri morfologici distintivi di Homo habilis; d) caratteri comportamentali di Homo habilis; e) il posto di Homo habilis nella filogenesi ominide. 8. Homo erectus: a) la scoperta di Homo erectus; b) collocazione spazio-temporale di Homo erectus; c) alcune caratteristiche morfologiche di Homo erectus; d) caratteri comportamentali di Homo erectus; e) la collocazione di Homo erectus nella filogenesi ominide. 9. La fase della comparsa dell'uomo moderno: a) le forme di transizione erectus-sapiens; b) le più antiche forme identificate come Homo sapiens; c) Neandertal; d) l'uomo di Cro-Magnon; e) Homo sapiens afer; f) il moderno Homo sapiens in Asia; g) riepilogo. 10. Alcuni concetti relativi all'evoluzione degli Ominidi: a) modelli filogenetici; b) evoluzione a mosaico; c) gli ominidi che non hanno avuto successo; d) evoluzione culturale. □ Bibliografia.
1. Definizione di uomo
Il termine ‛uomo', senza ulteriori qualificazioni, è decisamente vago e viene inteso nei modi più diversi. Senza una terminologia più precisa e senza stabilire delle regole per il suo uso, è facile che, in una discussione sull'‛uomo', scienziati, filosofi e non specialisti si fraintendano. Ci sembra perciò necessario, all'inizio di questo articolo, esaminare i diversi usi del termine e definire il senso in cui lo useremo in questo contesto.
Il significato più ampio del termine ‛uomo' e quello più comunemente usato quando si discute l'origine e l'evoluzione dell'uomo è quello impiegato per indicare i membri della famiglia zoologica dell'uomo, che, come suggerito da J. E. Gray (v., 1825 e 1827), è stata denominata Hominidae. Le specie e gli individui appartenenti a tale famiglia sono comunemente noti come Ominidi.
Secondo un altro uso, il termine ‛uomo' è limitato ai membri del genere Homo, un termine latino molto generico che C. von Linné (o Linneo) usò nella decima edizione del Systema naturae (1758) per riferirsi ai vari individui che formano il consorzio degli uomini viventi. Successive ricerche svolte nel XIX e specialmente nel XX secolo hanno mostrato che in Hominidae è stato incluso almeno un altro genere oltre a Homo, per cui se adottiamo il termine ‛uomo' solo per riferirci agli individui appartenenti al genere Homo lo usiamo in un senso più ristretto.
L'uso più restrittivo di questo termine è quello che fa riferimento esclusivamente agli appartenenti alla specie vivente chiamata da Linneo Homo sapiens (1758). In questa accezione il termine è in genere usato da coloro che si occupano delle differenze di comportamento e mentalità tra l'uomo moderno e le attuali scimmie antropomorfe, o di caratteristiche tipicamente umane, come la creatività o la creazione di sistemi etici e filosofici. Tuttavia il termine non viene usato in questa accezione restrittiva dagli studiosi di biologia umana, di paleoantropologia o di archeologia.
Inoltre, in ognuna delle tre accezioni che abbiamo appena ricordato è possibile riscontrare la presenza di diverse sfumature di significato.
In questo articolo l'accezione più ampia del termine ‛uomo' e quella più appropriata: infatti, ci dedicheremo a un gruppo di Primati superiori, i membri della famiglia Hominidae.
I membri di tre gruppi di Primati superiori, cioè l'‛uomo' nel suo significato più ampio, le grandi scimmie antropomorfe (scimpanzé, gorilla e orango) e le piccole scimmie antropomorfe (gibbone e siamango) hanno in comune una tal quantità di caratteristiche morfologiche da giustificare l'opinione, ormai largamente accettata, che appartengano a un unico raggruppamento nella classificazione dei Mammiferi e dei Primati. A questo raggruppamento principale il paleontologo e tassonomista americano G. G. Simpson attribuì la qualifica di ‛superfamiglia', alla quale dette il nome di Hominoidea. Pertanto, il termine ‛Ominoidi' include gibboni, scimmie antropomorfe e uomo, mentre il termine ‛Ominidi' si riferisce soltanto all'uomo in senso lato.
Dal momento che in questo articolo ci occupiamo di Ominidi, è importante sottolineare il fatto che, come ci ha insegnato Linneo 250 anni fa, la classificazione di creature viventi dipende fondamentalmente dalla loro morfologia o struttura corporea. Ciò nel regno animale implica tutti gli aspetti della struttura corporea, sia macroscopica che microscopica, comprende, cioè, non solo ossa e cervello, muscoli e organi, nervi e vasi sanguigni, ma anche tessuti e cellule coi loro minuscoli componenti, compresi i cromosomi. In tempi recenti, sempre nuovi aspetti del fenotipo, cioè i tratti veri e propri dell'organismo, come gruppi sanguigni e serici e una quantità di molecole identificabili, vengono usati come mezzi per una corretta classificazione delle forme viventi. Purtroppo, nello studio dei fossili non ci possiamo servire di quelle prove chiarificatrici che sono i cromosomi, i minuscoli depositari del materiale ereditario, di cui ogni membro di ogni specie vivente possiede un numero definito. Inoltre, lo studioso di evoluzione non si può servire di prove biochimiche dirette, come i gruppi sanguigni, tranne che nei reperti più recenti. Perciò, quando studiamo antichi reperti fossilizzati, siamo obbligati a utilizzare prove di tipo diverso rispetto a quelle impiegate nelle ricerche su forme di vita recenti e moderne. Una descrizione delle varie strategie di ricerca è fornita nel cap. 4.
Per stabilire le affinità esistenti tra forme estinte di Primati superiori, e per individuare la direzione e le caratteristiche dei cambiamenti evolutivi avvenuti lungo periodi di tempo molto lunghi, è necessario avere un idea precisa di che cosa è, e che cosa non è, un ominide. Pertanto, i paleoantropologi si basano su definizioni di Hominidae e di famiglie strettamente collegate, come quelle delle scimmie antropomorfe, sulle quali esiste un accordo generale. Cominceremo quindi esaminando brevemente la famiglia delle scimmie antropomorfe (Pongidae) perché, come ha notato Darwin in The descent of man (1871), lo scimpanzé e il gorilla sono i più stretti parenti dell'uomo.
2. Le caratteristiche di Pongidae
I Pongidae, o Pongidi, comprendono le grandi scimmie antropomorfe: il gorilla e lo scimpanzé africani, l'orango asiatico e altri antichi membri della famiglia ora estinti. Alcuni scienziati, come Simpson, includono tra i Pongidi anche le piccole scimmie antropomorfe (gibbone e siamango), che altri, come G. Heberer, considerano invece abbastanza diverse dalle grandi da giustificare una loro classificazione in una famiglia separata, Hylobatidae. Seguiremo Simpson e molti altri scienziati, e considereremo i gibboni come membri di una sottofamiglia dei Pongidi.
La più recente e più largamente accettata definizione dei Pongidi è stata proposta nel 1955 da sir W. E. Le Gros Clark nel libro The fossil evidence for human evolution. Piccole modifiche vennero apportate nella seconda edizione del libro (1964) e nella terza, curata da B. G. Campbell (1978). Tale definizione è troppo lunga per poter essere citata testualmente, ma va sottolineato il fatto che essa fu espressa in termini non di entità anatomiche apparentemente statiche, ma secondo le linee evolutive generali che distinguono la famiglia, che sono le seguenti: progressive modifiche dello scheletro che si adatta a un tipo particolare di locomozione arboricola detta ‛brachiazione' e si manifesta specialmente nel proporzionale allungamento dell'arto anteriore sia nel suo insieme che nelle sue singole parti (mano, polso, avambraccio, braccio); sviluppo di un forte alluce opponibile alla superficie plantare delle altre dita del piede; relativa riduzione del pollice; mantenimento delle proporzioni del bacino, che sono quelle caratteristiche del bacino dei mammiferi quadrupedi; sviluppo, nei pongidi di maggiori dimensioni, di mandibole massicce associate con creste muscolari pronunziate sul cranio; mantenimento di una posizione arretrata o posteriore dei condili occipitali sulla base del cranio; limitato grado di flessione dell'asse della base del cranio che ne mantiene bassa la volta; ingrandimento degli incisivi e specialmente dei canini che divengono lunghi denti conici che si inseriscono negli spazi (diastemi) dell'opposto mascellare; accentuazione della capacità di taglio del primo premolare inferiore permanente.
Questa lista di caratteristiche non è certo completa; inoltre, paragonando gli antichi pongidi a quelli recenti, è possibile rendersi conto che non tutte le caratteristiche sono manifeste nello stesso grado a ogni stadio evolutivo: alcuni tratti sono comparsi prima di altri, certi si sono sviluppati più rapidamente e altri meno. Tuttavia, da un reperto incompleto, qual è un fossile, è in genere possibile mettere in evidenza un numero di caratteristiche sufficiente a dare un'idea abbastanza chiara dell'appartenenza o meno dell'esemplare ai Pongidi.
Le prove attualmente disponibili indicano senz'altro che gli Ominidi e i Pongidi, specialmente i Pongidi africani, hanno avuto un antenato comune che, in un periodo di tempo compreso tra quattro e dieci milioni di anni fa, dette origine a due linee diverse, appunto gli Ominidi e i Pongidi.
3. Le caratteristiche di Hominidae
Nella ricerca dei criteri in base ai quali può essere caratterizzata la famiglia dell'uomo, possiamo individuare proprietà morfologiche (sulle quali si basano le classiche definizioni zoologiche di Hominidae), aspetti comportamentali e caratteri molecolari. Per lo studioso di paleontologia umana i reperti fossili forniscono scarse informazioni dirette sulla composizione molecolare propria degli Ominidi. Indicazioni un po' più precise sul comportamento dei primi Ominidi provengono dai reperti archeologici, considerati come una rappresentazione fossilizzata del comportamento. La maggior parte delle prove a nostra disposizione riguarda la morfologia degli Ominidi fossili ed è su questo aspetto che concentreremo principalmente la nostra attenzione.
a) Caratteristiche morfologiche principali degli Ominidi
I tratti caratteristici degli Ominidi sono stati enumerati per la prima volta da Gray (v., 1825), ma in seguito sono stati meglio precisati in numerose occasioni, via via che si venivano accumulando nuovi dati e che si procedeva all'analisi di campioni di maggiori dimensioni di Ominidi di tipo diverso.
Nella seconda metà del XX secolo Le Gros Clark (v., 1955) ha proposto una definizione più completa di ciò che si considera un ominide, definizione che è stata leggermente modificata nelle successive edizioni (19642 e 19783) del suo libro The fossil evidence for human evolution. Anche la definizione di Hominidae, come quella di Pongidae, è basata sulle tendenze evolutive che contraddistinguono la famiglia. Le Gros Clark ha elencato 19 precise direzioni, processi o complessi che sono presenti in una parte o in un'altra dello scheletro. Perciò, la loro presenza o assenza, la loro forma o il grado in cui si manifestano possono essere verificati sui resti fossili dello scheletro.
Tra le 19 caratteristiche indicate da Le Gros Clark ricordiamo: modifiche dello scheletro dovute all'adattamento alla postura eretta e alla locomozione bipede; un crescente grado di flessione dell'asse della base del cranio, associato a una crescente altezza della volta cranica; relativo spostamento in avanti dei condili occipitali, cioè delle superfici articolari alla base del cranio tramite le quali il cranio si articola con la vertebra atlante e la colonna vertebrale; sostituzione del margine tagliente del primo premolare inferiore permanente con una struttura bicuspidata (con successiva riduzione secondaria della cuspide linguale della bicuspide); marcata tendenza, in stadi successivi dell'evoluzione, a una riduzione della grandezza dei molari; alterazione progressiva del rapporto tra premolari e molari da una parte, e incisivi e canini dall'altra: i primi si allargano per schiacciare e macinare grazie a un potente apparato masticatore che fornisce una maggiore potenza diretta ai molari, mentre i canini e gli incisivi si rimpiccoliscono; pronunciata e rapida espansione (in alcuni ominidi che si collocano nella fase finale della loro sequenza evolutiva) della capacità cranica, e quindi delle dimensioni del cervello che era contenuto nel cranio; riduzione, negli ominidi di stadi evolutivi più tardi, delle dimensioni della mandibola e dell'area di inserzione dei muscoli masticatori e comparsa di una protuberanza mentoniera o mento osseo.
Questa lista non è assolutamente completa, ma comprende alcune delle principali caratteristiche distintive della famiglia Hominidae. Sappiamo adesso che non tutte queste caratteristiche evolutive fecero la loro comparsa contemporaneamente né il processo evolutivo fu ugualmente veloce. Tuttavia, l'attento esame di fossili di Primati superiori, anche molto antichi e incompleti, in genere riesce a mettere in evidenza la presenza o meno di queste caratteristiche e il riscontro di almeno alcuni di questi elementi distintivi dell'uomo può esser sufficiente a stabilire l'appartenenza del fossile agli Ominidi.
Tuttavia, bisogna ammettere che esistono casi limite la cui interpretazione è controversa. Fossili come Ramapithecus, Oreopithecus e Gigantopithecus mostrano ciascuno una o due o anche più caratteristiche che suggeriscono la presenza di tendenze ominidi, ma esse non sono abbastanza ben definite da permettere di fare una diagnosi relativamente sicura: i tratti rappresentati o sono troppo pochi, o sono espressi in grado minimo, così da rendere impossibile un accordo generale sull'interpretazione dei fossili. In alcuni casi i fossili sono così incompleti che lo studioso non è in grado di determinare se le caratteristiche ominidi avrebbero potuto essere presenti nelle parti mancanti. Ciò ci deve far ricordare che, nel caso che il materiale fossile sia scarso e specialmente quando si tratta di stadi precoci dell'evoluzione degli Ominidi, o quando si ha a che fare con rami ominoidi collaterali, non è possibile asserire in modo netto e inequivocabile che certe caratteristiche sono di natura ominide. In passato, alcuni campioni di questo tipo sono stati correttamente identificati solo dopo la scoperta di altri resti più completi. Così, pochi studiosi includono ancora i membri dei tre generi citati - Ramapithecus, Oreopithecus e Gigantopithecus - tra gli Ominidi, benché in un periodo o in un altro siano state individuate in ognuno di essi caratteristiche considerate proprie degli Ominidi. D'altra parte, l'affermazione che Australopithecus è in realtà un ominide è confermata dalle nuove scoperte e dai nuovi studi effettuati in seguito al rinvenimento del primo esemplare, a opera di R.A. Dart nel 1924.
Per dare al lettore una chiara idea di che cosa si intende per ominide, possiamo individuare sette aspetti principali del modello morfologico ominide (v. tab. I). L'acquisizione di questi sette complessi, in gradi e tempi diversi e attraverso cambiamenti verificatisi con velocità diverse, ha caratterizzato i Primati superiori che si sono diffusi nel corso di quella radiazione evolutiva che va sotto il nome di ominazione.


1. Acquisizione della stazione eretta, abituale o per tempi prolungati, e della locomozione bipede abituale, cioè capacità di stare in piedi, camminare e correre (v. tab. II). Lungi dal rappresentare un gruppo di cambiamenti di scarsa entità, ciò ha implicato: a) alterazioni nella struttura della base cranica e dell'allineamento cranio-vertebrale; b) sviluppo di meccanismi strutturali tali da permettere di scaricare il peso in basso lungo la colonna vertebrale attraverso la parte superiore del sacro fino alle ossa iliache; c) sostanziali modifiche del bacino in modo da rendere possibile il nuovo meccanismo di locomozione senza che ciò pregiudicasse l'espletamento continuo e adeguato dell'altra funzione fondamentale del bacino, cioè il suo uso come canale del parto; d) modifiche della testa e del collo del femore, della lunghezza, curvatura e forma del femore, del meccanismo articolare del ginocchio, della caviglia, del piede e delle dita del piede.
Tutti i resti fossili sembrano indicare che questi elementi vennero sviluppati in stadi molto precoci della storia dell'ominazione.
2. Riassestamento e ristrutturazione dell'arto anteriore, in modo che invece di rimanere un organo per sostenere pesi, per stare in piedi, camminare o correre come nell'andatura quadrupede, o nella brachiazione, o in ambedue, si libera, del tutto o quasi, dagli impegni della locomozione, e può dedicarsi alla manipolazione che diviene sempre più precisa; ciò comporta, fra l'altro, un accorciamento dell'arto nel suo insieme e un allungamento del pollice che sviluppa la proprietà anatomica e funzionale dell'opponibilità.

3. Modifiche della dentatura in senso ominide (v. tab. III). Si verifica una generale riduzione delle dimensioni dei denti, dei mascellari e di tutto l'apparato masticatore; una riduzione differenziale dei canini, che da lunghe zanne divengono denti relativamente piccoli con punte allineate quasi allo stesso livello dei denti vicini nell'arcata dentaria; trasformazioni degli altri denti, riguardanti la forma, struttura e spessore dello smalto della corona, la dimensione assoluta e relativa della corona (compresi cambiamenti nelle dimensioni di certi denti rispetto a quelle di altri), la grandezza della cavità dentaria, numero, forma e struttura delle radici; cambiamenti nello schema di occlusione e di masticazione.
4. Encefalizzazione complessiva. Accrescimento allometrico fortemente positivo del cervello nel suo insieme e contemporaneo aumento della capacità della scatola cranica; in altre parole, la velocità di accrescimento del cervello è di gran lunga maggiore di quella del corpo nel suo insieme.
5. Encefalizzazione selettiva. Sviluppo differenziale di alcune parti del cervello: certe aree aventi funzioni ben precise si ingrandiscono più di altre. In particolare ciò si riscontra nel caso dei lobi frontale e parietale, della parte posteriore della circonvoluzione frontale inferiore (cioè dell'area di Broca) e dell'area di Wernicke. Si riscontra anche lo sviluppo di una asimmetria, che può essere modesta o marcata, di parti speciali degli emisferi cerebrali, in concomitanza con un alto grado di asimmetria funzionale tra l'emisfero sinistro e quello destro (v. emisferi cerebrali: Dominanza cerebrale).

6. Rimodellamento della particolareggiata morfologia del cranio, inclusa la mandibola (v. tab. IV). Tali cambiamenti avvengono in accordo con: a) la posizione della testa su una colonna vertebrale diritta, il che comporta delle modifiche della base e dell'equilibrio del cranio; b) cambiamenti delle dimensioni della dentatura e del modo e della potenza della masticazione; c) accrescimento e rimodellamento del cervello.
7. Sviluppo del linguaggio articolato. È questo un carattere sia funzionale sia comportamentale la cui base strutturale richiede: a) lo sviluppo delle aree del linguaggio nel sistema nervoso centrale; b) cambiamenti periferici riguardanti i canali respiratorio e alimentare in modo da formare il canale vocale.

Non abbiamo detto praticamente niente dei tessuti molli del corpo, come i visceri, i muscoli e i vasi sanguigni. I segni che alcuni muscoli lasciano sulle ossa possono fornire utili informazioni ausiliarie sullo stato e l'anatomia funzionale dei fossili: per esempio i muscoli masticatori e quelli delle mani e dei piedi.
Anche il cervello, come i muscoli, lascia il suo segno sulle ossa. Durante la vita embrionale, fetale e postnatale la forma della scatola cranica si modella accuratamente sulla superficie del cervello in espansione. La parte esterna del cervello, così, lascia la sua impronta sulla parte interna della scatola cranica; in alcune parti, perfino le circonvoluzioni, o giri, e le fessure, o solchi, che le separano lasciano una chiara impronta nell'interno della calotta cranica. Tali impressiones gyrorum, a loro volta, caratterizzano un calco endocranico, artificiale o naturale che sia. È pertanto possibile, se si opera con sufficiente attenzione, leggere alcuni dei segni che originariamente si trovavano sulla superficie del cervello.
Molti studi hanno dimostrato che il calco endocranico riproduce molto fedelmente la forma del cervello. Paragonando i calchi endocranici con i cervelli corrispondenti, Connolly (v., 1950) ha mostrato che i segni delle fessure sono chiaramente visibili in alcune parti dei calchi, specialmente nelle zone più basse; egli, avendo riscontrato anche che tali impronte sull'osso scompaiono in età avanzata, ne deduce che probabilmente l'impronta delle fessure raggiunge il massimo grado di evidenza, sia nelle scimmie antropomorfe sia nell'uomo, in adulti giovani. Questo fatto risulta vantaggioso per lo studio dei reperti fossili, in quanto una larga percentuale delle ossa di ominidi primitivi a nostra disposizione appartiene a individui anatomicamente immaturi. Per esempio, dei circa 160-170 esemplari di australopitecine del Sudafrica a cui possiamo attribuire individualmente un'età, il 48 o 49% sono di individui immaturi, e il 52 o 51% di individui maturi; pertanto una buona percentuale di essi morì in età tale da poter essere ragionevolmente sicuri che avessero crani con impronte ben marcate delle fessure cerebrali, sempre ammettendo che la situazione in relazione a questi primi ominidi fosse la stessa di quella delle attuali scimmie antropomorfe e dell'uomo.
Grazie all'attento studio ditali impronte sull'interno della volta cranica o sulla superficie del calco endocranico, i neurologi sono riusciti a identificare quali parti del cervello dei primitivi ominidi fossero molto sviluppate e quali fossero appena rappresentate.
Un carattere morfologico che è impossibile studiare nei fossili è l'assetto cromosomico, il microscopico depositario dei materiali ereditari presente nella maggioranza delle cellule del corpo. Sappiamo che le moderne scimmie antropomorfe, sia africane sia asiatiche, possiedono 48 cromosomi in ogni cellula normale del corpo, mentre l'uomo moderno ne ha 46. Le nuove tecniche per lo studio dei cromosomi hanno messo in evidenza uno schema intricato e caratteristico di bande disposte secondo un modello costante lungo ogni cromosoma, per cui è stato addirittura possibile stabilire quali cromosomi umani sono omologhi a quelli dello scimpanzé e del gorilla e determinare per quali aspetti i cromosomi dell'uomo moderno differiscono da quelli delle scimmie antropomorfe (v. cellula: Fisiologia e genetica: Citogenetica). Purtroppo non abbiamo alcun metodo in base al quale stabilire con esattezza quando si originarono tali differenze, compresa la diversità di numero. Così, per esempio, non siamo in grado di stabilire se Australopithecus possedesse 46 cromosomi come l'uomo moderno o 48 come le attuali grandi scimmie antropomorfe.
Tuttavia, tenendo presenti le sette caratteristiche proprie degli Ominidi che abbiamo ricordato, ci è più facile esaminare e interpretare i reperti fossili; possiamo così delineare quando e come fecero la loro prima comparsa le caratteristiche anatomiche degli Ominidi e come si svilupparono in seguito.
b) Principali tratti comportamentali degli Ominidi
Nell'ambito comportamentale i tratti che distinguono gli Ominidi dai Pongidi sono moltissimi. I traguardi raggiunti dagli Ominidi comprendono tutte le attività complesse, abituali, volontarie e pianificate e il grado di dipendenza da esse ai fini della sopravvivenza quali: raccolta di cibo e caccia; perdita del periodo di estro, sostituito da recettività sessuale durante tutto l'anno; tabù dell'incesto; graduale sviluppo, accanto alla bergsoniana intelligenza concreta, dell'intelligenza razionale, o, secondo un altro tipo di analisi, sviluppo dell'intelligenza operazionale accanto all'intelligenza sensomotoria e preoperazionale di Piaget; acquisizione del linguaggio articolato e della parola; grande sviluppo della capacità di risolvere problemi; simbolizzazione e organizzazione dei simboli in entità culturali coerenti; sviluppo di una cultura sia materiale sia non materiale; flessibilità comportamentale e trasmissione della cultura. La funzionalità e il comportamento ominidi sono contraddistinti da una immaturità molto accentuata alla nascita e per un tempo notevole dopo di essa; infatti c'è un prolungato periodo di infanzia e adolescenza, con ritardata comparsa del menarca (inizio delle mestruazioni), dell'eruzione dei denti e del completamento del processo di crescita e di ossificazione; l'età media si allunga e compare la menopausa; le femmine sviluppano mammelle con caratteri umani.
Scorrendo questa lista, è chiaro che alcuni dei segni distintivi del comportamento ominide possono essere ricavati dallo studio dei reperti paleoantropologici: per esempio, la presenza di segni di cultura materiale, come utensili di pietra e osso, la costruzione di ripari, la raccolta di ocra rossa e di altre terre minerali. La natura dell'intelligenza può essere determinata sulla base di concetti impliciti nei reperti della cultura materiale. Dall'anatomia dei fossili e dalla complessità delle indicazioni di vita culturale possiamo addirittura presumere l'esistenza del linguaggio articolato.
c) Aspetti della biologia molecolare dell'uomo
Un tempo, lo studio dell'anatomia degli ominidi fossili, coadiuvato dalle conoscenze ricavabili dall'anatomia comparata di creature viventi correlate, era in pratica l'unica base su cui poggiavano tutte le nozioni relative all'evoluzione biologica degli Ominidi.
All'inizio degli anni sessanta, un diverso gruppo di studiosi, i biologi molecolari, ha contribuito con nuove conoscenze allo studio del problema delle origini degli Ominidi. Infatti la sistematica e l'evoluzione molecolare si avvalgono di tre approcci diversi: a) la valutazione della ‛distanza immunologica' tra specie e popolazioni diverse; b) la ‛sequenza delle proteine', cioè l'individuazione, resa possibile da nuove e raffinate tecniche biochimiche, della sequenza degli amminoacidi che costituiscono ciascuna proteina presente negli individui di una specie moderna; in certo senso ciascuna proteina ha le sue ‛impronte digitali', ed è così possibile paragonare le proteine corrispondenti in specie diverse, come ad esempio uomo moderno (Homo sapiens) e scimpanzé (Pan troglodytes); c) la sequenza del DNA (acido desossiribonucleico, il depositario delle caratteristiche ereditarie che è la componente autoriproducentesi dei cromosomi e di molti virus) e lo studio del DNA dopo ibridazione tra cellule di specie diverse. Applicando tali metodologie agli ominoidi viventi e basandosi su presupposti teorici è stato possibile durante gli ultimi vent'anni calcolare quando avvenne la divergenza dall'ultimo antenato comune alle scimmie antropomorfe africane e agli Ominidi. Infatti, la maggior parte degli studiosi di evoluzione umana ritiene che sia stata proprio una divergenza verificatasi nella linea evolutiva dell'ominoide africano a dar origine a Hominidae. Sulla base delle più recenti prove molecolari, questa divergenza pare sia avvenuta tra 4 e 7 milioni di anni fa.
Nonostante che alcuni dei presupposti su cui si sono basati gli evoluzionisti molecolari siano stati criticati fin da quando fu proposto per la prima volta il concetto di un ‛orologio molecolare' o strumento per la datazione dell'evoluzione (1962), le ricerche successive, basate su dati ottenuti in indagini molto più ampie di quanto non fosse stato originariamente suggerito per l'‛orologio dell'albumina', hanno progressivamente eliminato le obiezioni e hanno grandemente aumentato la fiducia nei confronti dell'approccio molecolare.
Una delle difficoltà che erano originariamente sorte, in seguito alle ipotesi avanzate in base alla biologia molecolare, era il fatto che la data di origine degli Ominidi risultava essere relativamente tarda (4-7 milioni di anni), mentre l'interpretazione che veniva allora data dei fossili indicava una genesi ominide molto più precoce (15 milioni di anni). Nuove scoperte di fossili e un più attento esame degli esemplari di ominidi più antichi hanno portato, tra il 1975 e il 1982, a modificare l'interpretazione dei fossili. I fossili più antichi, risalenti a 10-20 milioni di anni fa, non sono più considerati Ominidi, mentre fossili di circa 4 milioni di anni, scoperti recentemente, sembrano indicare con sufficiente sicurezza che l'origine degli Ominidi sia più recente.
Pertanto, attualmente si ritiene che le prove ricavabili sia dai fossili sia su base molecolare concordino nell'indicare un' origine relativamente recente (4-7 milioni di anni) di Hominidae. Le difficoltà sorte in un primo tempo con l'approccio molecolare hanno determinato un progressivo raffinamento della biologia molecolare e hanno inoltre portato a riconsiderare la candidatura dei fossili più antichi (per es. Ramapithecus) al titolo di ominide ancestrale.
La maggior parte degli studiosi di evoluzione oggi riconosce il valore del contributo fornito dallo studio delle molecole a quello delle origini degli Ominidi, che è consistito nella possibilità di stabilire il momento in cui gli Ominidi si sono separati dall'antenato che avevano in comune con le scimmie africane.
4. Strategie adottate nello studio dell'evoluzione umana
a) La strategia impiegata per la fase dell'emergenza degli Ominidi
L'emergenza di Hominidae da predecessori non ominidi e la divergenza di Ominidi e Pongidi è stata chiarita grazie a una triplice strategia: lo studio dei reperti fossili e archeologici, la morfologia comparata (compresa la morfologia cromosomica) degli Ominidi e la biologia molecolare comparata.
La controversia tra sostenitori dello studio dei fossili e sostenitori dell'approccio molecolare si è in gran parte risolta nella considerazione che i dati ricavabili dai fossili e le prove basate sulle somiglianze molecolari tra Pongidi viventi e Homo sapiens sono conciliabili. I biologi molecolari hanno attentamente rivisto i loro concetti, le loro supposizioni e i loro dati, mentre i paleontologi hanno sottoposto i fossili a esami più critici. Il risultato è stato una buona sintesi degli approcci paleontologico e molecolare e delle loro conclusioni. Tale sintesi era stata prevista nel simposio Wenner Gren intitolato The role of natural selection in human evolution tenutosi a Burg Wartenstein in Austria nel 1974 (v. Salzano, 1975); fu annunciata in un simposio intitolato Miocene hominoids and new interpretations of ape and human ancestry, organizzato da R. L. Ciochon e R. S. Corruccini nell'ambito dell'VIII congresso della International Primatological Society, tenutosi a Firenze nel luglio del 1980; ed è stata definitivamente sancita a un congresso tenuto alla Pontificia Accademia delle Scienze da dodici studiosi nel maggio 1982, intitolato Recent advances in our knowledge of Primate evolution e presieduto dal prof. C. Chagas, presidente dell'Accademia.
Pertanto, è stata ormai accettata l'idea che fossili, molecole e anatomia comparata possono insieme fornire le basi per lo studio della natura degli Ominidi e per datare le loro origini.
b) Strategie impiegate per la fase dell'evoluzione degli Ominidi
Dopo che gli Ominidi si furono separati dai Pongidi costituendosi a buon diritto come una famiglia, Hominidae, lo studio della loro evoluzione biologica successiva, da circa 4 milioni di anni fa fino a tempi relativamente recenti, è divenuto fondamentalmente uno studio di paleoanatomia. I dati di biologia molecolare comparata servono pochissimo o non servono affatto a sbrogliare la matassa dei modelli evolutivi presenti all'interno dei primi appartenenti a questa famiglia (4-1 milioni di anni fa), mentre assumono un ruolo sempre crescente i dati archeologici ed etologici (comportamentali). I mutamenti biologici che avvennero durante questo periodo, tuttavia, possono in gran parte essere desunti dall'attento studio paleoanatomico dei fossili; tale studio è arricchito dalle testimonianze della cultura materiale che ci illuminano su alcuni importanti aspetti quali dimensione, distribuzione, abitudini alimentari del gruppo, intelligenza tecnica e razionale, capacità di usare un linguaggio, insomma ci illuminano su quello che era il mondo delle idee degli Ominidi durante la loro evoluzione.
c) Strategie impiegate per la fase dell'evoluzione dell'uomo moderno
La storia dell'origine dell'uomo biologicamente moderno e della sua varietà si basa quasi esclusivamente sui dati anatomici o fisici desunti dall'antropologia e sulle variazioni genetiche riscontrabili nelle popolazioni moderne. Lo studio anatomico, in questa terza fase, si basa sulle somiglianze esistenti tra comunità umane viventi e reperti fossili datati con sicurezza, che divengono sempre più numerosi, mentre lo studioso di genetica delle popolazioni umane può usare la frequenza genica per sondare il passato, in quanto può chiarire le interrelazioni genetiche di popolazioni umane recenti. Anche in questa fase, come nella seconda, il ruolo dell'archeologo nel mettere in luce la nascita di schemi comportamentali moderni è importantissimo, mentre le fasi più recenti si possono avvalere degli studi di antropologia sociale, etologia, linguistica ed etnologia.
In definitiva, possiamo individuare tre fasi nell'evoluzione biologica degli Ominidi, per il cui studio è necessario ricorrere a diversi tipi di strategia: 1) per la fase dell'origine ominide (cioè per il periodo che risale a oltre 4 milioni di anni fa): paleoanatomia dei fossili, morfologia comparata (compresa la morfologia cromosomica) e biologia molecolare di Ominidi e non Ominidi viventi; 2) per la fase dell'evoluzione degli Ominidi (da 4 a 1 milione di anni fa): paleoanatomia dei fossili e approccio archeologico; 3) per la fase dell'evoluzione dell'uomo moderno (da 1 milione di anni fa al presente): studio anatomico dei fossili e delle popolazioni umane moderne, studi di biologia molecolare delle popolazioni moderne e utilizzazione delle informazioni ricavabili da archeologia, antropologia sociale, etnologia, linguistica ed etologia.
5. L'origine di Hominidae
Siamo in possesso di un certo numero di fossili ominoidei, risalenti all'Era Terziaria e provenienti da Africa, Europa meridionale e Asia, che secondo diversi studiosi vanno considerati candidati al titolo di Ominidi, o almeno a quello di antenati degli Ominidi. In tre importanti ominoidi del Terziario inferiore, e cioè Gigantopithecus, Oreopithecus e Ramapithecus, sono state riscontrate caratteristiche considerate proprie degli Ominidi; i loro resti risalgono a un periodo compreso tra 14 e 8 milioni di anni fa (tranne Gigantopithecus blacki cinese, al quale è stata assegnata un'età inferiore, cioè compresa nell'ultimo milione di anni). Ciascuno di questi tre generi è stato considerato, in un periodo o in un altro, come facente parte a pieno titolo della famiglia ominide. Ma il ritrovamento di nuovi esemplari, un esame più critico dei fossili precedenti e approfonditi studi di anatomia comparata effettuati su esemplari di grandi dimensioni di scheletri sicuramente appartenenti a Pongidi od Ominidi hanno progressivamente inficiato la validità di tale supposizione, tanto che negli anni ottanta quasi nessuno degli studiosi delle origini degli Ominidi ritiene che questi generi appartengano agli Ominidi secondo la definizione che ne abbiamo data.
È probabile che durante l'Era Terziaria inferiore vi sia stata nel Vecchio Mondo una ricca fauna di Primati superiori, e che molte di queste linee ominoidi stessero sperimentando nuovi modelli morfologici e comportamentali, divenendo, per qualche aspetto e in grado modesto, ‛ominizzate'.
Per esempio, qualche tratto più o meno ominide è manifesto in ognuno di questi tre ominoidi: Oreopithecus (i cui esemplari migliori provengono dai depositi di lignite della Toscana), Gigantopithecus dell'India e Ramapithecus. Praticamente, l'unica modifica in senso ominide comune a tutti e tre, pur con delle differenze, era una riduzione dei canini. Infatti, i canini di Gigantopithecus, pur essendo piccoli e non sporgenti, erano denti conici e appuntiti, mentre quelli ominidi non lo sono. Oreopithecus aveva canini relativamente piccoli, ma premolari e molari molto particolari e diversissimi da quelli di Ominidi e Pongidi. Gigantopithecus aveva rami mandibolari divergenti posteriormente (un carattere pongide), primo premolare inferiore adatto al taglio (tratto pongide), e una combinazione di molari e premolari grandi con incisivi piccoli (un tratto presente in alcuni dei primi ominidi).
Questi e altri esempi mostrano che durante il Terziario era in atto in diversi ominoidi un processo di ominazione, seppure in grado diverso, relativo ad alcune caratteristiche del complesso ominide. In Oreopithecus, ad esempio, il bacino aveva un ileo (parte del bacino) basso e largo come quello degli Ominidi, non alto e stretto come quello dei Pongidi. Tuttavia, un esame più attento ha mostrato che in Oreopithecus l'allargamento dell'osso era dovuto a un'espansione della parte anteriore dell'ileo, non di quella posteriore, come negli Ominidi. Pertanto, questa particolare somiglianza alla struttura ominide era superficiale e dovuta a una causa diversa; le due forme di ileo allargato dovevano corrispondere a modi di locomozione molto diversi, e dobbiamo pertanto escludere che si trattasse di un processo di ominazione dello stesso tipo.
Ulteriori difficoltà sono determinate dall'incompletezza della documentazione fossile. Di Ramapithecus e Gigantopithecus possediamo quasi esclusivamente denti e mandibole, per cui non possiamo sapere niente sulla loro postura e sul modo di locomozione, o sulla dimensione e le caratteristiche del cervello. Tuttavia, anche gli scarsi dati ricavabili sulla loro anatomia mostrano i segni di una ‛evoluzione a mosaico', dato che alcune caratteristiche sono più evolute in senso ominide rispetto ad altre e sembrano rappresentare degli esempi di evoluzione parallela. È ormai opinione generale che nessuno di loro abbia raggiunto il livello orninide; essi, pertanto, non annunciano la nascita della famiglia ominide come tale, anche se devono essere stati strettamente imparentati con quel gruppo ominoide che dette origine agli Ominidi. Attualmente, però, relativamente pochi scienziati ritengono che questi tre generi, o uno di essi, debba essere classificato nella famiglia Hominidae.
I dubbi maggiori sull'eventuale appartenenza agli Orninidi sono stati sollevati a proposito di Ramapithecus; è stata proprio la sua età così antica a scatenare la controversia sulla precocità o tardività della divergenza che ha originato gli Ominidi. Ma dopo ulteriori e attente analisi critiche, al congresso di Firenze del 1980 e a quello Vaticano del 1982 si è concordato che non era più sostenibile considerare Ramapithecus un ominide, mentre era probabile che facesse parte della linea evolutiva che va dai primi ominoidi all'orango.
I due tentativi più recenti di stabilire la data della divergenza Pongidi-Ominidi hanno portato a valutazioni in parte coincidenti, e cioè 4-7 e 4,5-5,5 milioni di anni fa. Attualmente si ritiene pertanto che l'emergenza degli Ominidi debba esser fatta risalire al periodo tra Miocene e Pliocene. La documentazione fossile mette a nostra disposizione un rilevante numero di esemplari che risalgono a questo periodo cruciale (v. tab. VI, nella quale sono elencate alcune delle scoperte, fatte in Africa orientale, di età compresa tra 4 e 10 milioni di anni fa). Tutti gli esemplari sono stati identificati come ominoidi, e alcuni mostrano una particolare somiglianza con gli Ominidi. I resti provenienti da Lukeino, Lothagam e Kanapoi erano così incompleti e mal distinguibili da rivelare assai poco della natura degli ominoidi a cui appartenevano. Uno o alcuni di loro avrebbero potuto appartenere ai primissimi Ominidi, o alla popolazione degli ultimi antenati comuni ai Pongidi e agli Ominidi. Si sa ancora molto poco delle scoperte, annunciate agli inizi degli anni ottanta, avvenute nella Middle Awash River Valley in Etiopia e nelle Samburu Hills nel Kenya settentrionale. Dalle descrizioni preliminari dei fossili di Middle Awash, risulta che il femore era adatto al bipedalismo, mentre è stato valutato che i frammenti del cranio appartenessero a una calotta avente una capacità di circa 400 cc, valore che è vicino a quelli più bassi di Australopithecus africanus del Transvaal. Questi esemplari di Middle Awash provengono da un'area i cui sedimenti, secondo J. Desmond Clark, risalgono ad almeno 6 milioni di anni fa.
Nonostante la loro frammentarietà, alcuni dei fossili citati nella tab. VI potrebbero appartenere a membri della prima popolazione sicuramente ominide, mentre altri all'ultima popolazione di antenati comuni a Pongidi africani e Ominidi. Le scoperte paleontologiche stanno gettando luce proprio sul periodo indicato dalle ricerche biomolecolari.
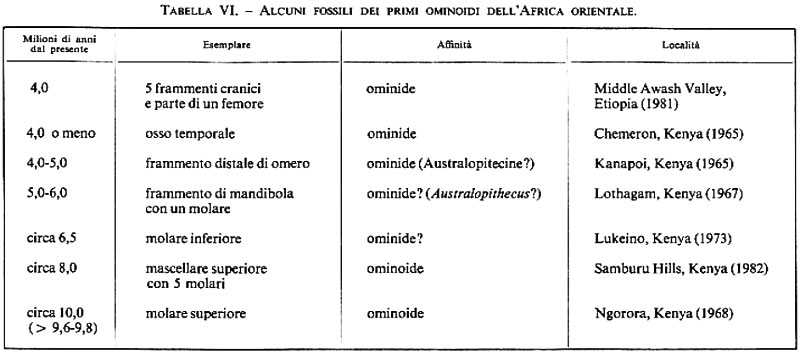
Sulla base delle prove disponibili, sembrerebbe che la famiglia Hominidae abbia avuto origine nel tardo Miocene o all'inizio del Pliocene, cioè non prima di 7 e non dopo 4 milioni di anni dal presente, molto probabilmente nelle fasce equatoriale e tropicale dell'Africa.
6. L'evoluzione di Hominidae
Nel corso del XX secolo sono state scoperte molte centinaia di ominidi fossili. Mentre alla fine del XIX secolo era disponibile solo un'esigua quantità di esemplari di una certa importanza in relazione al problema del ‟posto dell'uomo nella natura" (per usare la felice espressione di Th. H. Huxley), oggi i fossili rilevanti per il chiarimento dell'evoluzione umana sono così numerosi che la sola elencazione dei dettagli di primaria importanza di ogni esemplare riempie un catalogo di molti volumi pubblicato dal British Museum of Natural History nel corso di molti anni a partire dal 1967. Per dare un esempio dello spettacolare ritmo assunto dalle scoperte, specie nella seconda metà del XX secolo, possiamo ricordare il fatto che, mentre il volume relativo all'Africa, pubblicato nel 1967, contava 128 pagine (v. Oakley e Campbell, 1967), già dieci anni dopo si era resa necessaria una seconda edizione del volume che questa volta contava 223 pagine.
Nonostante che in Europa e in Asia siano state fatte molte importanti scoperte, e alcune anche in Australia e in America, il maggior numero di fossili importanti in relazione all'evoluzione della famiglia Hominidae nel periodo tra 4 e 1 milione di anni fa è stato scoperto in Africa. Elenchiamo qui di seguito i nomi dei luoghi in Africa dove sono avvenuti i principali ritrovamenti di ominidi fossili del periodo Plio-pleistocenico con le date delle prime scoperte di ominidi primordiali (per completezza abbiamo incluso anche luoghi come Samburu Hills, Lukeino, Ngorora, anche se gli ominoidi ivi rinvenuti non sono stati ancora identificati con sicurezza).

In breve, dall'unica località del 1924 siamo passati alle 22 del 1984. Il ‛decennio d'oro' è stato quello compreso tra il 1965 e il 1974, durante il quale sono state scoperte ben dieci località, sette delle quali in Kenya, due in Etiopia e una in Tanzania. Le località africane nelle quali sono avvenute le scoperte si trovano principalmente in due regioni: in Sudafrica e nella Great Rift Valley, a nord e a sud dell'equatore. Nel primo caso i ritrovamenti sono avvenuti in depositi di calcare dolomitico, nel secondo, principalmente, in depositi sedimentari lacustri e fluviali aperti. Ancor più notevole che non il numero delle località è il numero di individui fossili che tali scoperte hanno portato alla luce.
Nel 1957 i resti fossili identificati come ominidi primordiali provenienti da queste località africane appartenevano a non meno di 126 individui, forse addirittura 163. Anche considerando il valore medio, nel 1957 avevamo a disposizione i resti di circa 145 ominidi fossili.
Venticinque anni dopo erano stati portati alla luce da 19 località africane numerosi resti identificati come certamente o molto probabilmente appartenenti a ominidi. I paleoantropologi che studiavano gli ominidi primordiali africani potevano così disporre di ossa fossili provenienti da un numero di individui compreso tra 405 e 617, con un valore medio di 511.
Pertanto, tra il 1957 e il 1982, il numero di individui identificati come ominidi primordiali sulla base dei fossili africani si è più che triplicato, passando da 145 a 511. Queste cifre non comprendono gli esemplari di ominidi fossili provenienti da altre importanti località africane che risalgono a meno di un milione di anni fa, cioè i fossili di Ternifine in Algeria, di Rabat e Sidi Abderrahman in Marocco, Melka Kontouré in Etiopia, Kanjera in Kenya, Ndutu in Tanzania, Kabwe (Broken Hill) in Zambia, Hopefield e Florisbad in Sudafrica.
Benché il ritmo delle scoperte sia stato altrove meno rapido che in Africa, tuttavia anche in Asia e in Europa sono venuti alla luce nel XX secolo importanti e numerosi ominidi fossili. Nessuno degli esemplari rinvenuti fuori dell'Africa e identificati sicuramente come ominidi, tranne forse uno o due ominidi fossili indonesiani, risale al periodo compreso tra 4 e 1 milioni di anni fa.
Pertanto, il problema delle origini dell'uomo e della sua evoluzione fino a circa un milione di anni fa è risultato essere un problema in larga misura africano, ed è dall'Africa che deve venire la risposta all'antico problema delle origini e dei progenitori dell'uomo. Ciò ha rappresentato una sorpresa per molti studiosi della prima metà del nostro secolo, in quanto l'attenzione generale era rivolta all'Asia sin da quando un chirurgo militare olandese, Eugene Dubois, scopri a Giava nel 1891-1892 un antico ominide dall'aspetto primitivo, Pithecanthropus erectus. Questo reperto giavanese, che presentava fronte bassa e stretta, arcata sopraccigliare estremamente prominente e modesta capacità cranica, aveva, alla fine del XIX secolo, spostato indietro la storia umana a un grado di ominazione addirittura precedente a quello di Neandertal, i cui resti erano stati scoperti in Europa fin dal 1848. Ai fossili indonesiani venne in seguito attribuito il nome di Homo erectus. Le scoperte di Dubois avevano focalizzato l'attenzione sull'Asia come possibile culla dell'umanità e avevano creato, nel primo quarto del XX secolo, una situazione tale per cui tutti si aspettavano che l'‛anello mancante' dovesse essere ritrovato in Asia, a est dell'Eden!
Coloro che si aspettavano di trovare in Asia i più antichi progenitori dell'uomo avevano trascurato un passo molto interessante di The descent of man, che Darwin aveva pubblicato nel 1871. È un'osservazione la cui importanza ai fini della nostra storia è immediata e diretta, e che suona come una profezia: ‟In ogni grande regione del mondo i mammiferi esistenti sono intimamente affini alle specie estinte della stessa regione. È quindi probabile che l'Africa fosse abitata primieramente da scimmie estinte strettamente affini al gorilla e allo scimpanzé; e siccome queste due specie sono ora i più prossimi affini dell'uomo, è in certo modo più probabile che i nostri primi progenitori vivessero nel continente africano che non altrove" (v. Darwin, 18752; tr. it., p. 181). Tuttavia nella frase successiva egli aggiunge: ‟Ma è inutile speculare intorno a ciò''.
Era passato più di mezzo secolo quando nel 1925 R. A. Dart portò la prova che confermava la profezia di Darwin.
a) La scoperta del primo esemplare di Australopithecus
Dall'Africa è venuta la più grande sorpresa per il mondo dell'antropologia, allorché, nel 1925, R. A. Dart, professore di anatomia alla appena fondata scuola medica dell'Università di Witwatersrand in Johannesburg, pubblicò il primo resoconto sul cranio fossile di un bambino. Era stato scoperto alla fine del 1924 vicino al villaggio Tswane di Taung (‛il posto del leone') nella Cape Province settentrionale. La romantica storia del ritrovamento di quel cranio è stata raccontata da Dart stesso nella sua autobiografia, Adventures with the missing link (v. Dart e Craig, 1959), e da R. Broom (v., 1950) nel libro Finding the missing link.
La nuova scoperta, come abbiamo già detto, provocò un terremoto, in quanto la sua localizzazione geografica era del tutto inaspettata: il mondo stava appena abituandosi all'idea che la culla dell'umanità fosse stata l'Asia, quando il bambino di Taung reclamò tale ruolo per l'Africa. Ma la caratteristica più sorprendente della nuova scoperta non era tanto la sua provenienza quanto la sua morfologia. Infatti il cranio aveva canini piccoli come quelli degli Ominidi e molto diversi da quelli dei Pongidi; inoltre la base cranica mostrava chiaramente che i condili occipitali che formano l'articolazione con la colonna vertebrale erano situati molto più anteriormente che nei crani delle scimmie antropomorfe, dal che Dart dedusse che la testa doveva essera stata abbastanza ben bilanciata su una colonna vertebrale praticamente eretta, a differenza di ciò che accade nelle scimmie, la cui testa pende in avanti in quanto la colonna ha una posizione inclinata. Dart aveva quindi messo in evidenza due importanti caratteristiche, la riduzione dei canini e la stazione eretta, che fungono da criteri in base a cui si determina l'appartenenza agli Ominidi. Tuttavia, in un primo momento erano state le caratteristiche del calco endocranico a colpire l'attenzione di Dart; infatti, il cranio di Taung era stato trovato completo di un calco dell'interno della scatola cranica perfettamente conservato che mostrava sulla superficie una quantità di dettagli sulle circonvoluzioni e sui vasi sanguigni. Dart si era in precedenza dedicato a studi sul cervello, per cui fu in grado di individuare sul calco dei segni che interpretò come dovuti alla posizione di una delle scissure cerebrali, il solco lunato, che è tipicamente ominide. Pertanto Dart si convinse che anche da questo punto di vista il bambino di Taung fosse più affine agli Ominidi che ai Pongidi.
Sembrò allora che Dart avesse trovato i resti fossili di un ominide primitivo strettamente correlato alla famiglia dell'uomo. Quando egli pubblicò il primo resoconto su questo esemplare e la sua morfologia, non osò classificarlo come membro della famiglia Hominidae; invece, egli inizialmente ritenne che rappresentasse una nuova famiglia, situata in una posizione intermedia tra le due famiglie ominoidi note, i Pongidi e gli Ominidi, e suggeri di chiamare questa nuova famiglia col nome di Homo-simiadae. Ma i colleghi di Dart non accettarono la proposta di stabilire una nuova famiglia, e in seguito si convenne che il bambino di Taung e altri esemplari di aspetto simile rinvenuti nel Transvaal in Sudafrica dovevano essere considerati membri della famiglia Hominidae. In altre parole, Dart aveva trovato un membro molto primitivo della famiglia dell'uomo.
Che cosa c'era, allora, di così inaspettato nella sua morfologia? La risposta, in breve, è che quello di Taung era il primo ominide con cervello piccolo che veniva scoperto.
Quando consideriamo i tratti anatomici che separano l'uomo attuale dalle grandi scimmie antropomorfe attuali, la principale caratteristica tipicamente umana è la straordinaria ampiezza del cervello. Sia per peso che per volume, il cervello dell'uomo moderno è circa tre volte più grande di quello di maggiori dimensioni riscontrabile nelle grandi scimmie, cioè nel gorilla, nonostante che il peso dell'uomo sia molto inferiore a quello del gorilla (circa la metà). Pertanto, anatomisti e antropologi si erano abituati all'idea che l'uomo fosse un primate superiore con un cervello eccezionalmente grande. I crani fossili umani che erano stati scoperti prima del 1925 avevano confermato tale punto di vista. Tutti i crani neandertaliani trovati in Europa contenevano cervelli grandi, come quelli dell'uomo moderno. Anche l'esemplare di Homo erectus trovato a Giava da Dubois aveva una grande scatola cranica, benché la sua capacità fosse solo doppia (e non tripla) di quella media di un gorilla moderno.
Pertanto fu decisamente una sorpresa constatare che, nonostante le sue molte caratteristiche ominidi, il cranio di Taung aveva una capacità tale da adattarsi a un cervello non più grande di quello delle scimmie antropomorfe. In base all'eruzione dei denti, sappiamo che il bambino di Taung mori a un'età non superiore ai 5 o 6 anni (secondo gli standard attuali); è pertanto ragionevole supporre che cervello e scatola cranica sarebbero cresciuti ancora se fosse vissuto più a lungo. Ma si sa che, sia nell'uomo che nella scimmia antropomorfa, all'‛età dentale' raggiunta dal bambino di Taung, il cervello, che si sviluppa precocemente, ha già raggiunto una dimensione che è il 9o-94% di quella dell'età adulta. Anche calcolando la presunta capacità ‛adulta' del cranio di Taung, le dimensioni rimanevano quelle di una scimmia antropomorfa.
Dart, però, era convinto che l'esemplare di Taung non appartenesse alla famiglia delle scimmie antropomorfe, e lo aveva incluso in una nuova specie e genere che chiamò Australopithecus africanus (la scimmia antropomorfa dell'Africa meridionale). La sua affermazione che la creaturina non era uno scimpanzè aberrante, come sostenevano alcuni dei suoi denigratori, determinò una furiosa controversia nella letteratura scientifica, tanto che per più di 25 anni non fu raggiunto un accordo sul tipo di specie rappresentato dal piccolo fossile. La discussione verteva essenzialmente su questi punti: quel piccolo cervello doveva esser considerato un tratto così importante da impedire che Australopithecus fosse qualificato come ominide, nonostante la preponderanza delle altre caratteristiche ominidi? O l'appartenenza a tale specie doveva fondarsi sulla maggioranza delle caratteristiche - che erano ominidi - anche in assenza della tendenza umana verso un ingrandimento sproporzionato del cervello?
L'accesa discussione si trascinò per decenni, e il problema non si risolse finché la scoperta di molti nuovi fossili e lo studio dettagliato condotto sia su fossili sia su un gran numero di crani e cervelli di scimmie antropomorfe moderne e sull'uomo non dette un'idea più chiara del genere Australopithecus. Alla fine fu riconosciuto che, come aveva supposto Dart, il bambino di Taung differiva per così numerosi aspetti dalle scimmie antropomorfe da rendere impossibile non considerarlo un membro della famiglia degli Ominidi.
Retrospettivamente, possiamo affermare che il grande contributo di Dart è stato quello di forzare il mondo dei paleoantropologi a considerare possibile che un tempo fossero esistiti alcuni membri della famiglia dell'uomo con cervello piccolo, e che questo carattere di per sé non era sufficiente a escludere l'appartenenza di un individuo alla famiglia Hominidae. Inoltre, egli costrinse gli scienziati ad accettare il fatto che non tutte le parti del corpo degli ominidi ancestrali avevano subito il processo di ominazione nello stesso grado o nello stesso tempo; così, mentre Australopithecus africanus poteva esser considerato ominide per i denti, il modo in cui il cranio poggiava sulla colonna vertebrale, la forma del cervello, la presunta postura e andatura, non aveva ancora iniziato il processo di ominazione per ciò che riguardava l'ingrandimento sproporzionato del cervello. Tale schema evolutivo è chiamato ‛evoluzione a mosaico'.
Va detto che anche il tipo di evoluzione a mosaico mostrato dal fossile di Taung era fuori dell'ordinario. Nel primo quarto del XX secolo era diffusa l'idea che l'ingrandimento del cervello doveva aver rappresentato uno dei primi, se non il primo carattere, precedente ai denti o alla postura, a subire il processo di ominazione, e tale punto di vista era stato strenuamente sostenuto dal maestro di Dart, sir Grafton Elliot Smith. Invece, il cranio di Taung mostrava una tendenza del tutto opposta, cioè era ominide per ciò che riguardava denti e postura, ma non aveva ancora un cervello molto sviluppato. Secondo i preconcetti del tempo, la creatura di Taung non rappresentava il giusto tipo di progenitore degli Ominidi; sappiamo ora che non era il cranio di Taung a essere sbagliato, ma quei preconcetti. Lo sviluppo del cervello si è verificato in un punto successivo, e non precedente, della strada che porta all'uomo.
Non è forse esagerato asserire che la scoperta del cranio di Taung e la precisa identificazione del suo stadio evolutivo effettuata da Dart rappresentarono uno dei progressi più significativi e rivoluzionari nell'ambito della ricerca e della comprensione delle origini dell'uomo. Forse bisognerebbe includerla tra le maggiori scoperte scientifiche del nostro secolo. L'opera di scienziati quali R. Broom, J. Robinson, la famiglia Leakey, Ph. V. Tobias, C. Arambourg, Y. Coppens, D. Johanson e dei molti altri che si dedicarono allo studio dell'evoluzione dell'uomo in Africa, potrebbe in certo senso essere considerata come un lavoro di chiarimento di dettagli. La scoperta è di Dart: egli fu l'uomo giusto al momento giusto e nel posto giusto, col necessario bagaglio di conoscenze, perspicacia, fantasia e buona fortuna.
Ma il mondo scientifico fu purtroppo lento a riconoscere l'importanza del contributo di Dart: egli raccolse molte più calunnie che riconoscimenti, per anni dopo la comparsa del suo primo articolo su Australopithecus (v. Dart, 1925). Molte furono le circostanze che giocarono a suo sfavore: il fatto di risiedere a Johannesburg, il nome da lui scelto per la specie, le prevalenti aspettative riguardo sia alla localizzazione geografica sia alla forma anatomica del progenitore putativo, l'incertezza dei metodi di datazione dell'epoca, prima che ne venissero introdotti di più precisi (come i metodi radiometrico e paleomagnetico). Un altro fattore che giocava contro l'accettazione dell'ipotesi di Dart era la giovane età dell'esemplare di Taung; infatti, è ben noto che le caratteristiche di una specie divengono pienamente evidenti in individui che abbiano superato l'adolescenza. Da un bambino di soli 5 o 6 anni non si potevano dedurre con sicurezza le caratteristiche di un individuo adulto e neppure, pertanto, quelle della popolazione a cui apparteneva. Ciò di cui il mondo aveva bisogno per convincersi della propria affinità con quello strano bambino fossile, era un esemplare adulto dello stesso tipo. Ma dovette aspettare ancora quasi una dozzina d'anni prima che ne venisse trovato uno.
b) Il progressivo chiarimento di Australopithecus
Circa cinque anni prima che l'australopitecino di Taung venisse scoperto, erano stati trovati in quella stessa località dei babbuini fossili di un genere estinto, Parapapio. T. R. Jones, nel 1935, e G. W. H. Schepers e W. H. Le Riche, nel 1936, tutti ricercatori nel Dipartimento di Anatomia di Dart, recuperarono in una caverna dolomitica a Sterkfontein vicino a Krugersdorp, non lontano da Johannesburg nel Transvaal, dei babbuini fossili. Schepers e Le Riche informarono Broom della loro scoperta e lo portarono a Sterkfontein; entro otto giorni Broom trovò il primo Australopithecus adulto!
Broom era un medico e paleontologo scozzese che aveva lavorato in Australia alla fine del XIX secolo; all'inizio del XX secolo aveva scoperto a Karoo in Sudafrica molti fossili di rettili simili a mammiferi che avevano chiarito parecchi aspetti dell'origine dei mammiferi a sangue caldo dai loro predecessori rettili. Dopo l'annuncio della scoperta di Dart nel 1925, Broom era stato uno dei suoi pochissimi sostenitori ben informati; anzi, aveva deciso di scoprire un australopitecino adulto. Nel libro The coming of man: was it accident or design?, da lui pubblicato nel 1933, Broom aveva affermato il suo convincimento che ‟Australopithecus è molto vicino al progenitore dell'uomo, e se non un membro dello stesso genere da cui [l'uomo] ha avuto origine, almeno un membro della famiglia". Pertanto, Broom fu uno dei primissimi a classificare la specie rappresentata dal bambino di Taung non come appartenente ai Pongidi, e neppure all'Homo-simiadae di Dart ma alla, famiglia Hominidae. In quello stesso libro Broom aveva pubblicato una ricostruzione che prevedeva la morfologia di un cranio adulto di Australopithecus africanus, che risultò estremamente somigliante agli esemplari adulti da lui scoperti a Sterkfontein dal 1936 in poi.
La scoperta del 1936 forni la prima prova di quale doveva essere stato l'aspetto dell'uomo-scimmia adulto, confermando in sostanza nell'adulto quelle tendenze morfologiche che Dart aveva individuato nel bambino.
Lo strato del deposito di Sterkfontein dal quale era stato estratto il cranio del 1936 risultò molto ricco di fossili, e Broom dedicò ogni sua energia agli scavi e allo studio. Allo scoppio della seconda guerra mondiale, nel 1939, aveva recuperato da questo strato circa 19 frammenti di Australopithecus, che comprendevano crani, mandibole, denti e ossa postcraniali; tale livello fu da T. C. Partridge denominato in seguito Membro 4 della formazione di Sterkfontein, in quanto risultò che sotto di esso si trovavano altri tre Membri che erano perciò più antichi di quello in cui erano stati rinvenuti i frammenti di Australopithecus.
Lo studio di questi fossili convinse Broom che l'uomoscimmia di Sterkfontein era diverso da quello di Taung, e lo chiamò Plesianthropus transvaalensis per sottolineare tale differenza. In seguito, la maggior parte degli studiosi concordò nel considerare i fossili di Sterkfontein come appartenenti alla stessa specie, A. africanus, ma a una sottospecie diversa da quella di Taung. Su queste basi, il nome scientifico completo attribuito alla sottospecie di Taung fu A. africanus africanus, mentre la sottospecie di Sterkfontein fu denominata A. africanus transvaalensis. La denominazione dei fossili determinò una lunga controversia tra i paleoantropologi, ma l'accordo degli studiosi fu maggiore di quanto questa controversia potesse far credere.
Gli anni 1936-1939 rappresentano la prima fase delle scoperte degli Ominidi di Sterkfontein, durante la quale fu scoperto anche, nella vicina Kromdraai, il primo uomoscimmia cosiddetto ‛robusto'. Era evidente che si trattava di un tipo di uomo-scimmia diverso da quello di Sterkfontein: aveva in comune con le altre australopitecine il fatto di avere una piccola scatola cranica, ma ossa e muscoli erano più robusti, e molari e premolari avevano dimensioni maggiori, specie in larghezza. Inoltre, esisteva una marcata discordanza tra la larghezza di molari e premolari e le ridotte dimensioni di canini e incisivi, mentre l'arcata dentaria dei campioni di Sterkfontein mostrava proporzioni relativamente armoniche. Broom chiamò questo nuovo tipo di uomo-scimmia Paranthropus robustus, ma oggi è noto col nome di Australopithecus robustus, cioè viene classificato nello stesso genere ma in una specie differente rispetto ai reperti di Sterkfontein e Taung.
Alcuni anni più tardi, Broom riportò alla luce altri frammenti di uomo-scimmia robusto da una vicina località nota come Swartkrans, e con l'aiuto di J. T. Robinson riuscì a estrarre da quel piccolo deposito un numero maggiore di frammenti di ominidi primordiali di quanti ne siano stati trovati in qualunque altra località sulla Terra.
Mentre non fu difficile ravvisare nell'A. africanus di Sterkfontein il probabile progenitore di Homo, il caso di A. robustus risultò più difficile, in quanto mostrava troppe di quelle caratteristiche che i paleontologi chiamano specializzate o acquisite (o apomorfiche) per poterlo includere nella linea evolutiva di Homo. Questi tratti specializzati o apomorfici facevano pensare a una divergenza rispetto al percorso principale dell'ominazione. Inoltre, tutti i reperti faunistici rinvenuti nei due depositi nei quali furono trovati esemplari di A. robustus, in Transvaal, avevano un'età geologica più recente di quelli da cui provengono i resti del più slanciato A. africanus. Tuttavia, sulla base di criteri morfologici, A. robustus doveva esser considerato un ominide tanto quanto A. africanus: ambedue erano infatti bipedi con cervello e canini piccoli.
Pertanto, tramite Broom, l'Africa aveva introdotto un altro concetto paleoantropologico rivoluzionario, e cioè che non tutti i più antichi ominidi erano antenati di quelli venuti più tardi. Questo era un concetto diffuso in paleontologia in relazione ad altri animali, ma negli studi antropologici non si era mai - o quasi mai - creduto che i più antichi ominidi avessero subito un processo di diversificazione e speciazione ramificato, e neppure che alcune di queste ramificazioni - pure legittimamente ominidi - potessero non aver rappresentato gli antenati di uomini appartenenti al genere Homo. L'accettazione di questo concetto risultò un'amara pillola da inghiottire, e per molti anni ancora vi furono studiosi che includevano in un'unica linea evolutiva che portava all'uomo tutti gli ominidi fossili. Per la maggioranza, tuttavia, i fossili di Kromdraai e Swartkrans erano la prova inequivocabile che i primitivi ominidi si erano divisi, nel loro successivo sviluppo, in almeno due linee evolutive.
Dopo la seconda guerra mondiale, su richiesta del generale J. C. Smuts, Broom riprese a lavorare a Sterkfontein. In questa seconda fase di scavi, dal 1947 al 1949, Broom fu assistito da J. T. Robinson. Furono fatte molte nuove scoperte, compresa quella dello splendido cranio Sts 5 (‛Mrs. Ples'), tratto alla luce nel 1947. Questa fase ebbe termine nel 1949, quando Broom e Robinson rivolsero la loro attenzione al deposito di Swartkrans.
Una terza fase di scavi ebbe inizio a Sterkfontein nel 1956, quando C. K. Brain scoprì degli artefatti litici nella breccia superiore del deposito di Sterkfontein; l'anno seguente anche Robinson tornò a scavare in questa breccia, scoprendo, tra il 1957 e il 1958, oltre ad altri artefatti alcuni resti frammentari di ominidi nella parte superiore del deposito.
La quarta fase di scavi a Sterkfontein iniziò nel 1966 (nel centenario della nascita di Broom), quando Ph. V. Tobias e A. R. Hughes intrapresero uno scavo su larga scala del deposito, cercando di stabilirne la datazione e la stratigrafia e di individuare altri resti ominidi e la loro ecologia. Questo progetto a lungo termine ha prodotto nel 1983 almeno 185 nuovi campioni di ominidi, molti dei quali appartenenti ad A. africanus e provenienti dal Membro 4, ma alcuni anche dal Membro 5, appartenenti a quella che fu poi riconosciuta come una primitiva specie di Homo e associati a utensili litici. Sterkfontein non è l'unica località del Transvaal nella quale è stato trovato A. africanus. Da Makapansgat, circa 320 km a nord di Johannesburg, è stata estratta nel settembre del 1947 la prima serie di campioni di A. africanus transvaalensis. Questi fossili sono stati scoperti da J. Kitching e dai suoi due fratelli, del Bernard Price Institute of the University of the Witwatersrand, e da A. Hughes, del Dart's Anatomy Department. Questi ominidi di Makapansgat erano molto simili a quelli di Sterkfontein, anche se vi erano prove che essi fossero un po' più antichi; anzi, le australopitecine fossili di Makapansgat, fatte risalire a circa 3 milioni di anni fa, sono senz'altro i più antichi fossili ominidi scoperti in Sudafrica fino al 1980.
Swartkrans si trova al di là della valle di Sterkfontein, e là, poco prima della morte di Broom, avvenuta nel 1951, Robinson fece un'altra scoperta storica (1949); infatti, egli dimostrò che in questa caverna, insieme a frammenti di A. robustus, si trovavano frammenti di un tipo più avanzato di ominide, al quale Broom e Robinson dettero il nome di Telanthropus capensis. Alcuni dei frammenti considerati come facenti parte di questo gruppo furono trovati nello stesso livello stratigrafico di A. robustus. In seguito questo ominide più avanzato fu classificato nel genere Homo, e fu studiato in maggior dettaglio da R. J. Clarke che mise in evidenza caratteristiche in certo modo intermedie tra quelle di Homo habilis e Homo erectus.
Così, alla metà del XX secolo, erano state trovate in Sudafrica cinque località, da ciascuna delle quali erano stati estratti fossili attribuiti a un nuovo genere, Australopithecus. Grazie all'accumularsi delle nuove scoperte, ai resoconti pubblicati da Broom, Dart e Robinson, e agli attenti studi comparativi di Le Gros Clark, cominciava a essere accettata l'idea che questo nuovo genere, pur rappresentando una varietà di creature aventi un cervello le cui dimensioni assolute non superavano quelle delle moderne scimmie antropomorfe, doveva essere incluso nella famiglia zoologica Hominidae. Fino a quel momento, la fase australopitecina attraversata dall'umanità nel corso della sua evoluzione era considerata essenzialmente un fenomeno sudafricano. Solo pochi sostenevano che alcuni degli ominoidi fossili trovati in Cina e a Giava fossero correlati ad Australopithecus, ma non si avevano prove convincenti della loro reale appartenenza a questo genere.
Dopo la metà del XX secolo, una serie di scoperte in Africa orientale rivelò la presenza di un'altra forma ancora di australopitecine. Broom aveva ormai fatto accettare l'idea dell'esistenza di un tipo robusto di Australopithecus africano grazie alle sue scoperte a Kromdraai e a Swartkrans, queste ultime effettuate insieme a Robinson. Non era perciò irragionevole aspettarsi che analoghi tipi di uomoscimmia potessero essere trovati in Africa orientale. Ma nessun antropologo era preparato alla scoperta, fatta nel luglio 1959 da Mary Leakey nella gola di Olduvai in Tanzania settentrionale, di un australopiteco con enormi denti e muscolatura potente. Si trattava del cranio di un uomo-scimmia così robusto da far apparire insignificanti quelli di Swartkrans. La sua scoperta era stata prevista nel 1955, quando a Olduvai un assistente della Leakey aveva trovato un gigantesco molare deciduo. La Leakey riscontrò nell'esemplare di Olduvai un altro tipo ancora di australopitecine con cervello piccolo cui fu dato nome di Zinjanthropus boisei (Zinj è un antico nome dell'Africa orientale, e Charles Boise era il finanziatore delle ricerche dei Leakey). Più tardi, lo studio dettagliato effettuato su questo cranio indusse Tobias a includere questa forma dell'Africa orientale nello stesso genere Australopithecus, anche se in una specie separata, Australopithecus boisei. Nonostante la sua evidente somiglianza con A. robustus del Transvaal, per numerosi aspetti risultava più vicino ad A. africanus e per altri differiva da ambedue le specie transvaaliane.
Successivamente, furono trovate a Peninj, vicino al lago Natron in Tanzania settentrionale (1964), e a Omo, verso la parte settentrionale del lago Turkana (allora chiamato lago Rodolfo) in Etiopia (1967), delle mandibole appartenenti ad A. boisei. Crani dello stesso tipo vennero rinvenuti da R. Leakey e dai suoi collaboratori a Ileret e Koobi Fora, a est del lago Turkana, dal 1968 in poi. Inoltre, denti isolati, sempre appartenenti a questa specie, furono trovati in vari strati della Shungura Formation nella Omo Valley, e un cranio fu rinvenuto a Chesowanja, vicino al lago Baringo in Kenya (1970).
Questa ‛popolazione' fossile dell'Africa orientale mostrava che la diversità esistente tra gli ominidi con cervello piccolo era stata ancor più accentuata di quanto fosse risultato dai fossili del Transvaal. La linea evolutiva degli Ominidi avrebbe potuto divergere originando non due ma tre diramazioni, dato che il ramo iper-robusto dell'Africa orientale sembrava diverso da quello robusto del Sudafrica. Inoltre, sia A. robustus del Transvaal, sia A. boisei dell'Africa orientale erano apparsi in tempi relativamente recenti, dato che la maggior parte degli esemplari appartenenti a queste specie aveva un'età compresa tra 2 e 1 milione di anni.
Si comprese così che la fase australopitecina della vita ominide era stata rappresentata non solo in Africa meridionale ma anche in quella orientale. Mentre la specie ‛robusta' del Transvaal aveva dimostrato di avere un corrispettivo in Africa orientale, l'altra forma di australopitecina del Sudafrica - cioè A. africanus, talvolta indicato col nome di australopitecina ‛gracile' - per lungo tempo non fu individuata al di fuori del subcontinente sudafricano. La fase ‛gracile' di ominidi con cervello di piccole dimensioni rimase una sorta di curiosa lacuna nella discendenza rappresentata dai fossili dell'Africa orientale, il cui numero, peraltro, andava rapidamente crescendo.
Dalla fine degli anni sessanta, tuttavia, si cominciò a ritenere che alcuni dei numerosi denti isolati rinvenuti nei membri più antichi della formazione Omo in Etiopia meridionale fossero molto simili a quelli di A. africanus transvaalensis; tali denti sembravano decisamente indicare che anche le australopitecine dell'Africa orientale erano passate attraverso una fase ‛gracile'.
Dal 1973 in poi fu effettuata una serie di spettacolari ritrovamenti a Hadar, nella depressione Afar dell'Etiopia nordorientale. Questo gruppo di reperti di Hadar comprendeva crani e denti, ossa degli arti e una gran quantità di ossa appartenenti a un unico scheletro (denominato ‛Lucy'), e fu portato alla luce dallo statunitense D. C. Johanson e dai suoi collaboratori. Tuttavia, esemplari di ominidi ancora più antichi furono rinvenuti a Laetoli in Tanzania da un gruppo guidato da Mary Leakey. I reperti di Hadar rappresentavano una popolazione molto simile ad A. africanus del Transvaal e furono da alcuni considerati, insieme ai nuovi reperti di Laetoli, come rappresentanti di una nuova specie di Australopithecus primitivo, per la quale venne proposto il nome di A. afarensis (dalla depressione Afar nella quale erano stati rinvenuti); tuttavia Tobias e altri scienziati ritennero che i fossili di Hadar appartenessero alla stessa specie di A. africanus che era stata scoperta in Sudafrica, pur ammettendo che forse potevano far parte di una sottospecie geografica locale. Gli ominidi di Laetoli appartenevano sicuramente a un periodo geologico più antico di quelli di Hadar, e avrebbero potuto rappresentare una fase ancora precedente rispetto ad A. africanus (secondo alcuni) o ad A. afarensis (secondo altri). Ominidi di poco più antichi, datati circa 4 milioni di anni, sono stati estratti all'inizio degli anni ottanta dalla Middle Awash Valley in Etiopia a opera di J. D. Clark e T. D. White.
Così, tra il 1924 e il 1984, l'Africa tropicale e subtropicale ha fornito un gran numero di fossili di creature dalla postura eretta e cervello di piccole dimensioni, con canini ridotti e parecchi caratteri di tipo umano. Questo tipo di primati microencefalici è stato considerato come facente parte della famiglia Hominidae e del genere, ora estinto, Australopithecus.
c) Definizione di Australopithecus
Il termine Australopithecus è stato usato in modi diversi e per definire cose diverse, a seconda delle opinioni dei vari ricercatori. Nella sua accezione più comune, nell'ultimo quarto di questo secolo, Australopithecus comprende tutti quei fossili che in un momento o in un altro sono stati designati come Australopithecus, Plesianthropus, Paranthropus, Praeanthropus, Zinjanthropus e Paraustralopithecus. Nella letteratura scientifica si trovano diverse descrizioni di tale genere, come quella di Robinson (v., 1961) e quella di Le Gros Clark (v., 19642); le ultime sono quelle pubblicate contemporaneamente e indipendentemente da Howell (v., 1978) e da Tobias (v., 1978). Dato che in queste definizioni compare una quantità di dettagli anatomici estremamente tecnici, non vengono qui riportate. I tratti cruciali, tuttavia, dimostrano come Australopithecus si conformi allo schema morfologico ominide che abbiamo definito in precedenza.
Nei frammenti fossili di australopitecine a nostra disposizione erano chiaramente evidenti cinque delle sette caratteristiche morfologiche che abbiamo considerato distintive degli Ominidi (v. cap. 3, § a) e precisamente: 1) basi morfologiche per la postura eretta e la locomozione bipede; 2) segni di una ristrutturazione dell'arto superiore; 3) particolarità dei denti, come canini ridotti e primi premolari inferiori bicuspidati; 4) ingrandimento differenziale dei lobi parietale e temporale degli emisferi cerebrali; 5) pronunciato rimodellamento del cranio in conformità coi cambiamenti della forma del cervello, della dentizione e dell'articolazione cranio-vertebrale. Inoltre, alcuni dati suggeriscono che, nonostante l'assenza della tendenza verso un accrescimento allometrico decisamente positivo delle dimensioni assolute del cervello, il grado di encefalizzazione, o sviluppo cerebrale, in rapporto alle presunte dimensioni corporee era maggiore di quello riscontrato nei Pongidi. Quanto alla settima caratteristica (v. tab. I) che riguarda le basi strutturali del linguaggio articolato, risulta con evidenza uno sviluppo dell'area di Broca (anche se non di quella di Wernicke) ben maggiore che nei Pongidi. Pertanto, in Australopithecus sono presenti tutte le sette principali caratteristiche ominidi, anche se in gradi diversi. Sulla base di questi dati, Australopithecus è stato a buon diritto classificato tra i membri di Hominidae.
d) Le specie di Australopithecus
Pochi sono stati gli argomenti paleontologici che hanno sollevato più controversie dei nomi dati ai fossili. Tanto per cominciare, le prime analisi dei reperti fossili non ammettevano l'idea dell'esistenza di variazioni; quando il numero dei fossili disponibili era ancora limitato, non era naturalmente possibile apprezzare l'ambito delle variazioni esistenti all'interno di una specie. La difficoltà era dovuta anche al fatto che fino alla metà del secolo erano stati fatti ben pochi studi comparati sulla variabilità delle ossa dei Primati superiori viventi, eccettuato l'uomo; si sapeva poco delle variazioni di caratteri osservabili e misurabili in gorilla e scimpanzé viventi. Perciò, due fossili erano facilmente classificati come membri di due specie, o addirittura di due generi diversi sulla base di differenze relativamente piccole. Fu così che i fossili africani aventi un grado di organizzazione strutturale cosiddetta australopitecina vennero classificati in non meno di sette generi diversi, Australopithecus, Plesianthropus, Paranthropus, Meganthropus, Praeanthropus, Zinjanthropus e Paraustralopithecus. In un certo periodo Broom arrivò a classificare le sole australopitecine africane in tre diverse sottofamiglie.
Ma con la scoperta di un numero sempre maggiore di esemplari, si riuscì a determinare meglio la natura e la portata di differenze dovute al sesso, all'età, a variazioni individuali o regionali o temporali. L'analisi di tali ‛popolazioni fossili' si giovò grandemente dei contemporanei studi sulle variazioni riscontrabili all'interno di popolazioni viventi di Primati superiori. Inoltre i metodi statistici divenivano sempre più raffinati, in particolare l'analisi multivariata. Tutto ciò portò a porre l'accento sulla biologia delle popolazioni di ominidi fossili e a semplificare i metodi di classificazione.
Si arrivò quindi a concludere che, come detto in precedenza, tutti i primitivi ominidi caratterizzati dal limitato sviluppo del cervello, prima classificati in sette generi diversi, dovevano rientrare in un genere unico, Australopithecus. Solo un numero limitato di scienziati ritenne di dover continuare a considerare due i generi delle australopitecine. Molte delle discussioni sulla tassonomia dei vari fossili africani erano solo la conseguenza dei diversi punti di vista dei paleoantropologi che dibattevano se la morfologia e le dimensioni di un nuovo esemplare fossero tali da poterlo includere in una specie già esistente e definita. Lo studio più dettagliato dei campioni, un attento paragone con i fossili già noti e la scoperta di un maggior numero di fossili hanno in genere portato a comprendere e spiegare tali differenze. Per un certo periodo, tuttavia, le controversie a questo livello sembravano aver monopolizzato tutte le energie intellettuali ed emotive dei protagonisti. È importante, però, che si sia riusciti a mantenere il senso delle proporzioni riguardo all'importanza relativa, o alla mancanza di importanza, di queste differenze di opinioni. Le principali divergenze, infatti, in genere non erano dovute a differenze dei fossili, ma alla diversa capacità interpretativa di scopritori e descrittori, all'accuratezza delle loro misure, alla loro familiarità con altri fossili coi quali si potevano stabilire dei paragoni e alla loro perspicacia in relazione alla morfologia.
Di conseguenza, si è determinata la tendenza, durata per un lungo periodo e spesso criticata, per cui gli scopritori di fossili esageravano le differenze tra i nuovi campioni che venivano scoperti e quelli trovati in precedenza, e creavano con troppa disinvoltura nuove specie. La storia della paleoantropologia è piena di nomi proposti per queste supposte nuove specie, la maggior parte delle quali, peraltro, venne rapidamente scartata e ne rimane una traccia solo nelle cronache, spesso romantiche e fantasiose, della materia. Talvolta, tali nomi riflettono più la natura e la personalità dello scopritore che non le affinità dei campioni a cui si riferiscono. Questa tendenza a introdurre nuovi nomi potrebbe essere considerata solo un particolare curioso della storia della paleoantropologia, se non avesse troppo spesso creato confusione, mascherato le correlazioni e forse anche ritardato la giusta valutazione delle affinità tra i campioni, rendendo impossibile la vita del più volonteroso studioso della materia.
Naturalmente, poiché i paleontologi si stanno avventurando in età in cui fino a poco tempo fa i campioni erano molto scarsi, per esempio il periodo tra 3,5 e 6 milioni di anni fa, è probabile che verranno individuate nuove specie o addirittura nuovi generi; ma il giudizio finale su come classificare un campione fossile deve basarsi sulla valutazione anatomica, indipendentemente dalla sua età. È certo comunque che lo studio degli ominidi fossili è prima di tutto un problema paleoanatomico; la valutazione della localizzazione e dell'importanza degli ominidi fossili in essa ritrovati necessita inoltre di una buona conoscenza ditafonomia, geocronologia, zoogeografia, paleoecologia, etologia, tassonomia, e , nei casi in cui sia applicabile, anche archeologia.
Dopo questa digressione sulla situazione culturale e storica esistente all'epoca in cui vennero riconosciuti e denominati gli ominidi fossili, possiamo affermare che le specie di Australopithecus riconosciute tali dalla maggior parte degli studiosi fino agli anni ottanta erano comprese tra tre e cinque. Lo schema successivo riassume la situazione e mette in evidenza gli equivalenti.
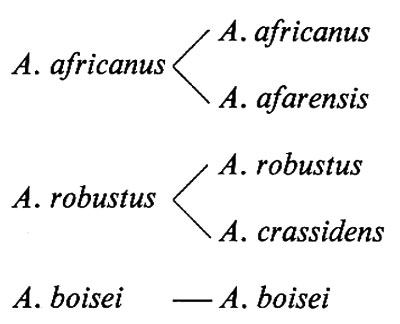
Il metodo di classificazione adottato dipende in larga misura dalle preferenze e dalla comodità, e ora l'uno ora l'altro dei due diviene il più usato a seconda delle circostanze e via via che si scoprono nuovi fossili, vengono pubblicati studi più meticolosi della loro morfologia, e i concetti e gli schemi relativi alla filogenesi si precisano e vengono confermati.
e) La datazione di Australopithecus
Tra le ‛popolazioni' australopitecine ben definite, i fossili di Laetoli vennero considerati fino all'inizio degli anni ottanta i più antichi fossili sicuramente ominidi. La loro età era compresa tra 3,77 e 3,59 milioni di anni dal presente. Ma all'inizio degli anni ottanta, le scoperte effettuate da J. Desmond Clark e T. D. White nella Middle Awash Valley in Etiopia spostarono indietro l'inizio della storia degli ominidi primitivi, presumibilmente australopitecine, e cioè a 4 milioni di anni fa.
Esclusi questi, i fossili più antichi erano quelli di Hadar, risalenti a un periodo compreso tra 3,3 e 2,8 milioni di anni, secondo le stime più recenti, e quelli di Makapansgat, i cui fossili identificati come A. africanus transvaalensis furono datati, sulla base di dati sia paleomagnetici che faunistici, tra 3,32 e 2,90 milioni di anni fa. Un pochino più giovane era la ‛popolazione' australopitecina di Sterkfontein, datata tra 3,0 e 2,5 milioni di anni sulla base di dati relativi alla fauna. Gli ominidi appartenenti ad A. robustus e A. boisei provenienti dall'Africa orientale e meridionale erano chiaramente più giovani (avevano cioè un'età geologica più recente) di quelli di Middle Awash, Laetoli, Hadar, Makapansgat e Sterkfontein. Il progressivo miglioramento dei metodi di datazione ha rivelato che sia A. robustus di Kromdraai e Swartkrans, sia A. boisei dell'Africa orientale comparvero relativamente tardi. La maggior parte degli esemplari di queste specie risale alla prima metà del Pleistocene, cioè a un'epoca compresa tra 2 e i milione di anni fa, e i più antichi al tardo Pliocene, poco più di 2 milioni di anni fa.
f) Caratteri comportamentali di Australopithecus
Accanto alla ricerca di resti fossili di ominidi primitivi, si è svolta una ricerca parallela dei segni di ‛comportamento fossilizzato', cioè di resti della cultura materiale, utensili di pietra o d'osso, e altri reperti archeologici. Dart aveva cercato di individuare i documenti di quella che propose di chiamare ‛cultura osteodontocheratica', che egli riteneva creata da A. africanus. La sua ipotesi che gli ominidi primitivi avessero usato ossa, denti e corna come strumenti era basata in gran parte sulle caratteristiche dei mucchi di ossa trovati nella caverna australopitecina di Makapansgat, nel Transvaal settentrionale. Ma nonostante il suo tentativo energico e vivace di diffondere questo punto di vista, tale ipotesi fu oggetto di sempre maggiori controversie a partire dalla fine degli anni sessanta. C. K. Brain (v., 1981) dimostrò invece che era estremamente probabile che i tratti più distintivi delle montagne d'ossa di Makapansgat fossero opera in gran parte di carnivori, in parte minore di porcospini e altri animali, e forse solo in minima parte di Ominidi.
Così, all'inizio degli anni settanta, era ormai appurato che nonostante vi fossero molte prove che Australopithecus avesse postura eretta, i dati a favore del fatto che possedesse una cultura materiale, o almeno una cultura che utilizzasse strumenti fatti di materiali non deperibili, erano molto scarsi. Tuttavia, specialmente nel terzo quarto del secolo, si era riusciti a stabilire che le grandi scimmie antropomorfe attuali, nonostante i loro piccoli cervelli, erano in grado di elaborare schemi di comportamento più complessi di quanto non si fosse fino allora ritenuto. Tenendo presenti questi dati, come pure l'apparente mancanza di strumenti non deteriorabili nei depositi australopitecini, si ritenne probabile che Australopithecus potesse aver presentato comportamenti almeno altrettanto complessi quanto quelli delle attuali scimmie antropomorfe e che quindi queste creature usassero strumenti fatti per la maggior parte di materiali deperibili che non avevano perciò lasciato alcuna traccia.
g) Il posto occupato da Australopithecus nella filogenesi degli Ominidi
Per determinare la posizione tassonomica e filogenetica di fossili quali quelli rappresentati dai vari campioni di Australopithecus, debbono essere tenuti presenti due caratteri principali, la morfologia e la datazione dei fossili, la cui localizzazione geografica può fornire un utile terzo criterio di valutazione.
I più antichi esemplari di Australopithecus rappresentati nella documentazione fossile risalgono all'ultima parte del Terziario, più precisamente alla seconda metà del Pliocene, cioè a 4-3 milioni di anni fa. Dato che sono noti diversi depositi fossiliferi risalenti a questo periodo, è probabile che durante l'ultimo ventennio di questo secolo troveremo che le radici del genere Australopithecus - forse addirittura della stessa famiglia Hominidae - risalgono alla prima parte del Pliocene (da 5 a 4 milioni di anni fa) o perfino agli ultimi eoni dell'era precedente, il Miocene (oltre 5 milioni di anni dal presente).
Dei fossili australopitecini trovati nell'ultimo quarto del secolo - e come abbiamo visto si tratta di alcune centinaia di individui - quelli più antichi (risalenti al periodo tra 4 e 2,5 milioni di anni fa) sono essenzialmente di tipo gracile, molto simili ad Australopithecus africanus. Possiamo pertanto concludere che A. africanus (e A. afarensis, se questa è una specie diversa) rappresenta il genere Australopithecus quale si espresse nel Pliocene.
I fossili più tardi, del periodo a cavallo tra Pliocene e Pleistocene e della prima metà del Pleistocene, cioè quelli risalenti al periodo che va da un po' più di 2 a circa i milione di anni fa, appartengono fondamentalmente alle specie a struttura corporea più massiccia, robusta (in Transvaal) e iper-robusta (in Africa orientale).
Nel gruppo più antico sono incluse popolazioni i cui rappresentanti possono a buon diritto essere ritenuti i progenitori sia delle posteriori australopitecine robuste e iper-robuste, sia di altre forme di Ominidi, anch'esse posteriori, classificate nel genere Homo. D'altra parte, le specie più tarde, A. robustus e A. boisei, sembra siano state super-specializzate e non aver avuto alcun ruolo quali antenati di Homo. In realtà, queste australopitecine più tarde non si ritrovano nella documentazione fossile risalente al periodo posteriore a circa un milione di anni fa, e probabilmente si estinsero. Nell'ultimo quarto del secolo i paleoantropologi hanno cominciato a concordare nel ritenere che una popolazione primitiva di australopitecine gracili, come A. africanus del Transvaal e il suo equivalente dell'Africa orientale, da alcuni ritenuto appartenente alla diversa specie A. afarensis, sia stata probabilmente la più antica specie di Ominidi conosciuta, progenitrice di tutti, o quasi tutti, i successivi Ominidi. Una divergenza (dadogenesi) nelle linee evolutive di queste australopitecine gracili, verificatasi 2,5-2,25 milioni di anni fa, dette origine a due o forse tre ramificazioni; da una di queste derivò la prima specie di Homo, Homo habilis, mentre l'altra, o le altre, svilupparono i caratteri tipici di A. robustus e A. boisei. Secondo questa interpretazione della documentazione fossile, quest'ultima linea evolutiva sopravvisse in Africa per circa un milione e mezzo di anni ancora, e fu perciò contemporanea della linea Homo che si stava sviluppando. Perciò, per tutta la prima metà del Pleistocene coesistettero due o tre specie di Ominidi; il ramo robusto scomparve circa un milione di anni fa, mentre il ramo Homo è durato fino a oggi.
In conclusione, la morfologia di Australopithecus ha per molti versi colmato lo iato morfologico tra l'uomo del genere Homo e le attuali grandi scimmie antropomorfe dell'Africa. Può pertanto esser considerato l'‛anello mancante' ipotizzato nel XIX secolo, confermando la profezia fatta da Darwin nel 1871: ‟E in certo modo più probabile che i nostri primi progenitori vivessero nel continente africano che non altrove".
7. La comparsa del genere Homo: il più antico Homo, Homo habilis
L'Africa ha provocato per due volte una rivoluzione nel nostro modo di pensare riguardo all'evoluzione umana: la prima fu determinata dalla scoperta, a partire dal 1925, della fase di ominazione nota col nome di Australopithecus. Come abbiamo già detto, la morfologia di questo genere abituò il mondo della paleoantropologia all'idea che l'appartenenza alla famiglia Hominidae era compatibile col fatto di possedere un cervello non più grande di quello degli attuali Pongidi. La seconda rivoluzione fu determinata dalla scoperta che alcuni ominidi, la cui morfologia era tale da includerli a buon diritto nel genere Homo, erano vissuti in Africa in un periodo molto più antico di quanto non si pensasse. Anzi, le prove indicavano che alcuni di questi primitivi membri del genere Homo erano stati contemporanei di alcune forme di Australopithecus sopravvissute più a lungo, cioè A. robustus in Sudafrica e A. boisei in Africa orientale.
Nonostante che molti studiosi fossero propensi a individuare nei fossili di Olduvai i più antichi membri di Homo, la scoperta della presenza di un vero uomo in Africa risale ai ritrovamenti fatti da Broom e Robinson a Swartkrans nel Transvaal verso la metà del secolo.
Fino ad allora, anzi, fino al 1961, era quasi universalmente accettato che i più antichi rappresentanti del genere Homo fossero da individuare nei fossili trovati in Asia e nell'Africa nordoccidentale e orientale, considerati appartenenti alla specie Homo erectus. Essi comprendevano fossili in precedenza designati coi nomi di Pithecanthropus, Sinanthropus e Atlanthropus.
Si determinò pertanto una sorta di rivoluzione quando si cominciarono a scoprire in Africa fossili risalenti a un periodo anteriore a quello di Homo erectus, che mostravano pochi dei tratti caratteristici di questa specie e tuttavia erano identificabili come membri del genere Homo.
a) La scoperta dei primitivi Homo in Africa
Le prime indicazioni dell'esistenza di un esemplare di Homo in un deposito di fossili di Australopithecus si ebbero due anni prima della morte di Broom. Il 29 aprile del 1949, mentre Broom si trovava negli Stati Uniti, il suo assistente Robinson portò alla luce, a Swartkrans, una mandibola che fu catalogata come SK 15. Broom e Robinson (v., 1949) si limitarono a dire che questa apparteneva ‟a quello che evidentemente è un nuovo tipo di uomo", che proposero di chiamare Telanthropus capensis. In seguito, A. Simonetta (v., 1957) e Robinson (v., 1961) proposero di considerare questa mandibola, un'altra catalogata come SK45 e pochi altri frammenti, trovati a Swartkrans, come appartenenti a Homo erectus. Così, questa fu la prima scoperta fatta in Africa di un primitivo membro del genere Homo. In seguito, si riuscì a dimostrare che la mandibola SK 45 (ma non la SK 15) e alcune altre ossa di Swartkrans erano contemporanee degli esemplari di Australopithecus robustus trovati nella stessa caverna. Successivamente, al Membro 1 della formazione di Swartkrans, nel quale erano stati trovati i due ominidi contemporanei, fu attribuita un'età compresa tra 2 e 1,5 milioni di anni.
Un secondo gruppo di frammenti di Homo primitivo provenne dal Membro 5 della formazione di Sterkfontein. Gli scavi del 1957 e del 1958 avevano portato alla luce degli utensili litici, proprio come a Swartkrans, e anche alcuni frammenti ominidi di denti e mandibole. Nonostante che Robinson li considerasse frammenti di australopitecine, Tobias ritenne che per alcuni versi potessero appartenere a Homo. I successivi ritrovamenti nel Membro 5 di Sterkfontein, effettuati da A. R. Hughes e Tobias, confermarono la presenza in questo strato di un primitivo appartenente al genere Homo (mentre resti di A. africanus transvaalensis erano stati rinvenuti nel sottostante e più antico Membro 4 di Sterkfontein).
Il terzo gruppo di prove della presenza di un primitivo Homo in Africa venne dalle scoperte effettuate dai Leakey a partire dal 1959. Dai livelli I e II della sequenza stratigrafica di Olduvai cominciarono a esser portati alla luce dei frammenti la cui classificazione risultò difficile. Il primo gruppo di questi frammenti, l'ominide 4 di Olduvai, comprendeva uno stretto molare inserito in un frammento di mandibola e un esile premolare, forse associato al precedente. Ciò avvenne prima della scoperta del famoso cranio di A. boisei (‛Zinjanthropus') effettuata da Mary Leakey nel luglio del 1959. La frammentarietà dell'ominide 4 di Olduvai era tale che nell'eccitazione determinata dal rinvenimento dell'esemplare di A. boisei, OH 5, si sottovalutò l'importanza di quei frammenti. Tuttavia, in un secondo tempo, un secondo gruppo di esili denti (OH 6) venne ad aggiungersi a quegli enigmatici frammenti. Si trattava chiaramente di un contemporaneo di A. boisei con molari e premolari più piccoli e relativamente più stretti; ma la natura esatta della popolazione di cui OH 4 e OH 6 erano rappresentanti fu per un certo tempo incerta.
Nel 1960 fu trovato a Olduvai OH 7, che in seguito fu considerato l'esemplare tipo di Homo habilis. Esso comprendeva una serie di ossa giovanili, cioè una mandibola, un paio di ossa parietali e alcune ossa della mano. Fu subito chiaro che le ossa parietali avevano dimensioni maggiori in tutti i sensi (tranne lo spessore) rispetto a qualunque osso parietale di Australopithecus rinvenuto nell'Africa meridionale e orientale. Tobias, a cui i Leakey avevano affidato il compito di descrivere A . boisei trovato a Olduvai, fu convocato da Johannesburg a Nairobi, dove, coadiuvato dai Leakey stessi e da Shirley Coryndon, effettuò la prima ricostruzione del diametro biparietale del cranio. Le sue dimensioni erano sorprendenti. Le misurazioni effettuate su tutti i parietali australopitecini disponibili in Sudafrica mostravano che quelli di OH 7 erano più grandi di quelli appartenenti a tutti gli ominidi primitivi africani trovati fino a quel momento. Sembrava pertanto ragionevole supporre che anche il cervello una volta contenuto in quei parietali avesse dimensioni maggiori di quelli dei tre tipi di Australopithecus rappresentati fino ad allora nelle collezioni.
Le differenze con A. africanus messe in evidenza in seguito ad accurate misurazioni riguardavano le grandi dimensioni dei parietali, e quindi la capacità complessiva del cranio, e il relativo restringimento dei premolari e del primo molare permanente; tali differenze rendevano la sua struttura più simile a quella dei moderni ominidi. Per quattro anni non fu presa alcuna decisione definitiva su quali fossero gli affini di questo nuovo ominide di Olduvai; durante questo periodo vennero eseguite accurate misurazioni comparate, e Tobias mise a punto un metodo per calcolare la presunta capacità cranica totale. Fu così ottenuto lo sbalorditivo valore di 674 cc, che rappresentava un aumento del 50% rispetto alla capacità media degli esemplari sudafricani di A. africanus. Non si trattava di un semplice spostamento verso i valori più alti della gamma di variabilità, ma di un aumento della capacità cranica tale da farlo collocare in un diverso ordine di grandezza. Numerosi altri fossili di Olduvai, compresi tre begli esemplari chiamati fantasiosamente Cenerentola (OH 13), George (OH 16) e Twiggv (OH 24), confermarono la presenza di questa combinazione di caratteristiche e di parecchi altri aspetti di fine morfologia che avevano caratterizzato una intera popolazione di ominidi primitivi. Anche a East Turkana e a Omo vennero trovati in seguito frammenti dello stesso tipo.
Nel frattempo, gli studi condotti a Londra da J. R. Napier, P. Davis e M. Day avevano messo in evidenza numerose caratteristiche del genere Homo nelle ossa di arti trovate a Olduvai, specialmente nelle giovanili ossa della mano che erano strettamente correlate ai parietali e alla mandibola di OH 7.
Le evidenti differenze presenti nei denti e nelle ossa degli arti e lo sbalorditivo aumento del 50% nelle dimensioni cerebrali indussero alla fine L. S. B. Leakey, Tobias e Napier (v., 1964) a concludere che il gruppo di frammenti ritrovati nel livello I e nel livello II inferiore della gola di Olduvai appartenevano a un'unica specie di Homo e non ad Australopithecus. Inoltre, questa specie era chiaramente diversa dalle due specie di Homo note in precedenza, e cioè Homo erectus e Homo sapiens. Su suggerimento di Dart, gli autori denominarono questa nuova specie Homo habilis, non tanto a causa del cervello più grande e della mano più ominizzata, quanto perché i frammenti scheletrici erano stati trovati insieme ad artefatti dell'industria olduviana.
L'inserimento della nuova specie nel genere Homo implicava solo una lieve modifica della definizione di Homo proposta da Le Gros Clark (v., 19642), si doveva solo, cioè, allargare l'ambito delle capacità craniche che si considerava potessero essere presenti all'interno di questo genere.
Altri esemplari di primitivi Homo furono identificati quando R. J. Clarke stabili una connessione anatomica tra un mascellare e una parte cranio-facciale di Swartkrans, riuscendo a formare un cranio composito identificato come SK 847, che Clarke e Howell (v., 1972) riconobbero appartenere a Homo. Lo studio estremamente dettagliato effettuato da Clarke (v., 1977) su questo cranio, pur confermando la sua affinità con Homo, non riuscì a determinare a quale specie esso appartenesse, se dovesse cioè essere classificato come H. habilis, H. erectus o come una forma di transizione tra i due.
La scoperta, sempre in Sudafrica, di un quarto probabile membro del genere Homo, molto simile a Homo habilis dell'Africa orientale, fu effettuata da A. R. Hughes a Sterkfontein nell'agosto del 1976. Questo esemplare, catalogato come Stw 53, proveniva da quello stesso strato, detto Membro 5, dal quale C. K. Brain e Robinson avevano estratto alcuni utensili litici.
Così, alla fine del terzo quarto del secolo, si era entrati in possesso di importanti dati che provavano che nell'Africa sia meridionale che orientale erano vissute già alla fine del Pliocene (circa 2 milioni di anni fa) delle creature appartenenti al genere Homo, e che queste erano state contemporanee di altre popolazioni di Australopithecus sopravvissute più a lungo.
Nonostante che inizialmente vi fosse una certa riluttanza a includere questi fossili primitivi di Homo nella nuova specie Homo habilis, all'inizio dell'ultimo quarto del secolo questa ipotesi era stata largamente accettata.
b) La datazione di Homo habilis
Via via che venivano reperiti nuovi esemplari considerati appartenenti alla specie Homo habilis, divenne chiaro come questi dovessero essere cercati tra i resti fossili di Olduvai (livello I e livello II inferiore), in alcuni strati di Koobi Fora e Ileret, a est del lago Turkana, e nei livelli superiori della sequenza di Omo Valley, oltre che nel Membro 5 di Sterkfontein e forse nel Membro 1 di Swartkrans. A tutti questi resti di H. habilis fu assegnata un'età compresa tra 2,3 e 1,6 milioni di anni; in altre parole, sembravano più antichi anche dei più antichi fossili considerati appartenenti a H. erectus, la cui presenza era documentata da un milione e mezzo di anni fa in poi. Per tale ragione alcuni studiosi indicano colloquialmente questi primi resti di Homo col nome di Homo pre-erectus. Come gruppo, questo taxon fece la sua comparsa poco prima della fine del Pliocene, si diffuse nel periodo a cavallo tra Pliocene e Pleistocene e perdurò per diverse centinaia di migliaia di anni durante il Pleistocene.
c) Alcuni caratteri morfologici distintivi di Homo habilis
Il dato fondamentale su cui si basò l'identificazione di Homo habilis con una creatura più ominizzata che non A. africanus fu la dimostrazione, fornita da Tobias, della maggiore capacità cranica - e quindi del maggior volume cerebrale - dell'esemplare OH 7, che presentava ossa parietali allargate. Da allora, la validità del criterio che si basa sulla capacità cranica per attribuire un esemplare a H. habilis è stata verificata da R. Holloway, D. Pilbeam e Tobias su questo e su molti altri campioni. Holloway ha nuovamente valutato la capacità cranica dei fossili di A. africanus del Transvaal, e sulla base dei risultati ottenuti su 6 crani è stato possibile calcolare una capacità media di 442 cc; la capacità media ottenuta per 6 campioni di H. habilis dell'Africa orientale è stata di 645 cc, con un aumento del 46% rispetto alla media ottenuta per A. africanus.
Il volume cerebrale è ovviamente correlato alle dimensioni corporee. Lo studioso di anatomia comparata francese G. Cuvier fu il primo, all'inizio del XIX secolo, a introdurre il concetto di ‛peso cerebrale relativo', cioè a esprimere il peso del cervello come una frazione del peso corporeo. Studi più recenti, condotti da O. Snell, E. Dubois, R. Bauchot, H. Stephan e E. Thenius, hanno dimostrato che una specie i cui membri sono in media più pesanti ha in genere volume cerebrale maggiore rispetto a una con peso corporeo inferiore. Il volume cerebrale di A. africanus, in termini assoluti, non era molto maggiore di quello delle attuali scimmie antropomorfe. Sulla base dei dati ottenuti sulle ossa degli arti e sulla colonna vertebrale degli ominidi fossili, è probabile che le dimensioni corporee di A. africanus fossero inferiori a quelle delle attuali scimmie antropomorfe; in questo caso, il loro volume cerebrale relativo risulterebbe un po' inferiore a quello di Australopithecus.
Tuttavia, la decisa tendenza ominide a un aumento cerebrale, sia assoluto che relativo, non era ancora evidente allo stadio di Australopithecus. Un apprezzabile ingrandimento delle dimensioni assolute e relative del cervello si manifestò più tardi, con l'avvento del genere Homo; infatti le ossa postcraniali di H. habilis rivelano che i membri di questa specie erano piccoli, praticamente pigmoidi, e tuttavia il loro cervello aveva un volume del 46% maggiore rispetto a quello di A. africanus. Questo è certo uno dei tratti paleoanatomici più appariscenti di Homo habilis. Naturalmente, all'interno di questa specie, come di qualunque altra specie, estinta o attuale, esiste un certo margine di variabilità individuale, ed è noto che il volume cerebrale si modifica anche a seconda dell'età e del sesso. Cosi la capacità cranica dei sei crani attribuiti a H. habilis (quattro di Olduvai e due di East Turkana) variava, nei valori ‛adulti', tra 513 e 770-775 cc; la media calcolata da Tobias per tre presunti maschi di H. habilis è stata di 695 cc, e per tre presunte femmine di 594 cc. L'indice di differenza dovuta al sesso tra questi due piccoli gruppi di campioni è risultato paragonabile a quello degli ominoidi moderni.
Un secondo gruppo di tratti principali che contraddistingue H. habilis comprende la forma del calco endocranico e gli ingrandimenti selettivi di certe aree degli emisferi cerebrali in confronto ad altre. È stato dimostrato che le parti inferiori dei lobi frontali e parietali avevano subito un aumento selettivo in H. habilis rispetto ai calchi endocranici di A. africanus e ai cervelli delle grandi scimmie antropomorfe attuali. Queste due regioni sono le sedi dell'area di Broca e dell'area di Wernicke, rispettivamente, e sono preposte al linguaggio articolato. Questo dato, insieme alle testimonianze della cultura, ha suggerito a Tobias l'ipotesi che H. habilis possedesse non solo le basi neurologiche del linguaggio articolato, ma anche, vista la sua cultura materiale relativamente complessa, che utilizzasse una rudimentale forma di linguaggio. Dati di questo tipo, cioè neuroanatomici e culturali, non sono stati trovati per A. africanus.
Un terzo gruppo di caratteri tipici di H. habilis è quello relativo ai denti. Nell'insieme, essi avevano dimensioni assolute più ridotte di quelli di A. africanus; i premolari e i primi molari permanenti erano più stretti, così che la corona aveva una forma più allungata di quella, più massiccia, riscontrabile nelle australopitecine. La riduzione differenziale dei denti lungo l'arcata dentaria riscontrata in H. habilis deve probabilmente esser messa in relazione con un diverso schema di occlusione e di attrito tra denti inferiori e superiori rispetto a quello osservabile in scimmie antropomorfe e Australopithecus. Sono state messe in evidenza anche molte altre differenze relative ai denti.
Differenze sono state notate anche in relazione alle ossa della mano e forse a quelle del piede, che risultano più orninizzate in H. habilis che in Australopithecus. I dati a questo proposito sono rimasti a lungo incerti a causa della difficoltà di associare delle ossa isolate di arti con crani e denti ben identificati (l'identificazione delle specie fossili, infatti, si basa principalmente sulle caratteristiche del cranio, della mandibola e dei denti); tuttavia, la base cranica di H. habilis dimostra che in questa specie la testa poggia con miglior equilibrio su una colonna vertebrale più eretta che non in A. africanus, il che significa che l'adattamento anatomico alla postura eretta e alla locomozione bipede era più avanzato nella direzione degli adattamenti scheletrici presenti nell'uomo moderno.
In breve, H. habilis era più simile all'uomo moderno di A. africanus, specie per quanto riguarda le dimensioni e la forma del cervello, i denti e l'adattamento alla postura eretta.
d) Caratteri comportamentali di Homo habilis
In tutti i siti nei quali sono stati scoperti resti di Homo habilis, sia nell'Africa orientale che in quella meridionale, sono stati trovati anche strumenti litici. Nella sequenza della gola di Olduvai, strumenti litici e resti fossili di H. habilis erano presenti negli stessi strati fertili, cioè nei membri inferiori, e anche a Sterkfontein, dove sono stati riscontrati nel Membro 5; invece nello strato immediatamente sottostante, il Membro 4, contenente resti di A. africanus, non è stata riscontrata alcuna traccia di H. habilis nè di artefatti litici. La presenza di strumenti litici è chiaramente associata a resti di H. habilis.
Mary D. Leakey, che è stata la più importante scienziata che ha condotto degli scavi nella gola di Olduvai e un'autorità per ciò che riguarda la sua sequenza archeologica, ha fornito prove inequivocabili a favore dell'ipotesi che sia stato proprio H. habilis l'artefice dell'antichissima e primitiva cultura che va sotto il nome di industria olduviana. Questa scienziata ha portato alla luce resti di H. habilis direttamente associati a strumenti olduviani da almeno 6 località di Olduvai, 5 situate nello strato I, e una nella parte inferiore dello strato II medio. La sua opera ha decisamente confermato l'ipotesi che coloro che produssero strumenti in modo sistematico a Olduvai appartenevano alla specie H. habilis.
Inoltre, la data della prima comparsa di H. habilis nella documentazione paleontologica (circa 2,3 milioni di anni fa) corrisponde quasi esattamente a quella della prima comparsa di strumenti litici nella documentazione archeologica africana.
All'inizio dell'ultimo quarto del secolo H. habilis era ormai concordemente ritenuto l'artefice di strumenti litici costruiti secondo schemi intenzionali e regolari e che, inoltre, si evolvevano col tempo. Comportamenti analoghi caratterizzavano anche le altre specie di Homo, cioè H. erectus e H. sapiens. Questi supposti tratti comportamentali, insieme ai criteri morfologici, fornivano un forte sostegno all'ipotesi che le popolazioni habilis fossero affini a Homo piuttosto che ad Australopithecus.
e) Il posto di Homo habilis nella filogenesi ominide
Tutti i caratteri morfologici di H. habilis suggerivano una situazione quale ci si sarebbe potuti aspettare in membri successivi, più ominizzati, appartenenti alla stessa linea evolutiva dei fossili di Laetoli e di Hadar, e nella quale rientravano gli esemplari di A. africanus di Makapansgat e Sterkfontein. Anche la collocazione temporale di H. habilis, da 2,3 a 1,6 milioni di anni, era in buon accordo col concetto che questa specie fosse derivata da A. africanus transvaalensis o dai suoi equivalenti dell'Africa orientale.
Quando L. S. B. Leakey, Tobias e Napier annunciarono che i fossili di Olduvai, diversi da A. boisei, appartenevano a una nuova specie di Homo, cioè H. habilis, due furono i fatti che risultarono più sorprendenti: il primo fu l'età dei fossili di Olduvai, e il secondo fu la loro stessa esistenza come popolazione separata. Infatti fino agli anni sessanta, e per alcuni studiosi anche in seguito, coloro che avevano accettato l'idea che A. africanus rappresentasse il più probabile progenitore di Homo ritenevano che i confini di popolazione della specie A. africanus fossero largamente sovrapposti a quelli della specie H. erectus. Se questo dato fosse risultato vero, vi sarebbe stato tra questi due taxa ancestrali uno ‛spazio morfologico' insufficiente all'inserimento di un'altra specie. Tuttavia, non appena venne analizzata la ‛distanza biologica' tra A. africanus e H. erectus risultò che per taluni aspetti critici, quali la capacità cranica e le dimensioni dei denti, il mutamento era relativamente grande e significativo. Fu quindi possibile dimostrare, sulla base di dati paleoanatomici, che tra le due specie vi era in effetti spazio sufficiente per inserire un'altra specie sulla stessa linea evolutiva. Anche gli studi del fattore tempo mostrarono che l'intervallo tra l'esemplare più recente di A. africanus del Transvaal e quello più antico di H. erectus era sufficiente, dato che corrispondeva a un periodo di almeno un milione di anni. L'eminente studioso americano di evoluzione G. G. Simpson calcolò che l'intervallo di tempo minimo necessario allo stabilirsi di una nuova specie era di 500.000 anni; fu perciò possibile sostenere che non solo la distanza biologica, ma anche la distanza temporale era sufficiente a farvi rientrare H. habilis come specie separata, situata nella parte iniziale della linea evolutiva di Homo.
L'altro aspetto sorprendente delle scoperte dei Leakey fu che gli esemplari che potevano essere attribuiti a Homo avessero potuto esistere addirittura 1,8 milioni di anni fa, che è la data alla quale possono esser fatti risalire i più antichi fossili di H. habilis trovati nella sequenza di Olduvai. Arrotondando un po' le cifre, possiamo dire che i più antichi fossili di H. habilis trovati a Omo, East Turkana, Olduvai e Sterkfontein hanno un'età di 2 milioni di anni. La scoperta dell'antichità del genere Homo fu uno dei meriti più rilevanti della famiglia Leakey. A ciò bisognerebbe aggiungere il fatto che il genere non ebbe inizio con H. erectus, il quale aveva un cervello relativamente grande, bensì col più modesto H. habilis, il quale peraltro riuscì vincitore nella competizione con le contemporanee australopitecine robuste, sopravvissute per lungo tempo.
All'inizio degli anni ottanta H. habilis è riuscito a conquistarsi un posto sicuro nella filogenesi ominide. La specie è stata considerata come uno dei derivati delle australopitecine gracili, che ebbero origine quando la linea evolutiva ominide subì una divisione cladogenetica, poco dopo 2,5 milioni di anni fa. La comparsa di H. habilis ha segnato l'inizio di una nuova fase dell'evoluzione degli Ominidi, caratterizzata dallo sviluppo di un cervello di maggiori dimensioni, dall'uso di strumenti e dalla dipendenza dalla cultura. Il processo di ominazione, divenuto evidente per la prima volta con Australopithecus, fa un salto in avanti nella direzione dell'uomo moderno; ha così origine la linea evolutiva del genere Homo. Se è vero che il progenitore di H. habilis fu A. africanus (col suo equivalente dell'Africa orientale) è altrettanto vero che la specie da esso derivata fu quella di H. erectus.
8. Homo erectus
Una delle specie estinte appartenenti alla famiglia Hominidae è Homo erectus (chiamato in precedenza Pithecanthropus). I membri di questa specie differivano da parecchi punti di vista dall'uomo moderno, che è stato classificato come una specie diversa di Primati, Homo sapiens. La storia di Homo sapiens risale a 100-200.000 anni fa, al massimo 300.000, a seconda della definizione di H. sapiens che viene adottata e a seconda che i vari fossili risalenti a questo periodo siano considerati gli ultimi rappresentanti di H. erectus o i primi di H. sapiens.
La documentazione fossile, tuttavia, ha prodotto resti di un altro tipo ancora di uomo, precedente a questo periodo e classificato come H. erectus. Questi resti sono stati trovati in estese zone del Vecchio Mondo, dove si può ritenere che questa specie abbia vissuto per più di un milione di anni (da circa 1,5 a circa 0,3 o 0,2 milioni di anni fa). La posizione spaziale e temporale di H. erectus è tale da farlo configurare come il diretto progenitore di H. sapiens. Proprio questa è la ragione dell'importanza particolare che gli viene attribuita nell'evoluzione generale degli Ominidi, in quanto sembra il più probabile predecessore immediato dell'uomo moderno. All'inizio di questo periodo, circa 1,5 milioni di anni fa, H. erectus si sarebbe evoluto dal poco anteriore H. habilis, come è confermato dai molti fossili provenienti dall'Africa meridionale e orientale, che mostra no caratteristiche di transizione tra H. habilis e H. erectus.
a) La scoperta di Homo erectus
E. H. Haeckel è stato uno dei grandi sostenitori di Darwin in Germania, tanto che Darwin stesso riteneva che il successo della teoria dell'evoluzione in Germania fosse dovuto in gran parte all'entusiastico appoggio di Haeckel. Nei suoi scritti Haeckel ipotizzò l'esistenza di una creatura ancestrale più antica e dall'aspetto più arcaico dell'uomo di Neandertal, i cui resti avevano cominciato a essere portati alla luce in Europa dal 1848 in poi. A questo ipotetico ominide ancestrale egli dette il nome di Pithecanthropus ancor prima che venisse scoperto, considerandolo una sorta di anello mancante che avrebbe dovuto colmare il vuoto presente nella catena evolutiva tra scimmia antropomorfa e uomo.
Anni dopo, tra il 1891 e il 1892, un giovane chirurgo militare olandese, E. Dubois, scoprì i resti di una creatura simile a quella descritta da Haeckel: si trattava di una calotta cranica, di un femore e di alcuni denti rinvenuti a Trinil, Giava. Egli rimase colpito dalla bassa volta cranica, con fronte stretta e sfuggente, arcate sopraorbitane prominenti e un deciso restringimento retro-orbitario del cranio: per queste caratteristiche, concluse Dubois, il nuovo cranio si trovava in una posizione intermedia tra quella dell'uomo e quella delle scimmie antropomorfe attuali. Egli riesumò il vecchio termine di Haeckel e dette a questo reperto il nome di Pithecanthropus (letteralmente uomo-scimmia). Vicino al cranio, Dubois trovò un femore chiaramente ominide, lungo e diritto, e così simile a un moderno femore umano che egli ritenne che il suo possessore doveva aver camminato in posizione eretta. Egli suppose che il femore fosse appartenuto allo stesso individuo, o almeno a un ominide dello stesso tipo di quello del cranio, facendo di questi pochi resti giavanesi il prototipo di una nuova specie, Pithecanthropus erectus. Le caratteristiche primitive del suo cranio e la capacità cranica relativamente modesta facevano arretrare la storia umana a un grado di ominazione più antico di qualunque altro noto fino ad allora.
Oggi, questa creatura viene chiamata Homo erectus. Resti di questa specie sono stati trovati in Indonesia, Cina, parte dell'Europa e Africa settentrionale e orientale. È stata proprio la scoperta di Dubois a far concentrare l'attenzione del mondo accademico sull'Asia, ritenuta la culla dell'umanità, e per un certo tempo la previsione di Darwin relativa all'Africa fu dimenticata. Tuttavia, nonostante che a quel tempo l'Africa non venisse considerata come possibile luogo di origine dell'umanità, la scoperta di Dubois aprì un nuovo capitolo nella storia degli Ominidi; infatti aveva messo in luce una fase della storia umana più antica e dall'aspetto più arcaico di qualunque altra nota fino allora. Questa scoperta costrinse gli antropologi a trovare un posto nella famiglia Hominidae anche al Pithecanthropus.
b) Collocazione spazio-temporale di Homo erectus
Le scoperte di fossili umani effettuate in seguito resero necessario stabilire una nuova e diversa specie. In un primo periodo le scoperte furono fatte principalmente in Asia; fossili simili a quelli di Dubois vennero trovati a Trinil, Kedung Brubus, Mojokerto, Sangiran e Sambungmachan in Indonesia. Un'altra serie di scoperte avvenne in Cina, specialmente nella famosa caverna chiamata Chukut'ien vicino a Pechino, ma sfortunatamente tutti i resti rinvenuti in questa località andarono persi durante la guerra cino-giapponese, nel 1941. Nel periodo successivo alla seconda guerra mondiale nuovi resti di H. erectus vennero scoperti nella stessa caverna, e anche in altre due località del distretto di Lantian, nella provincia di Shanhsi, e nel distretto di Hsik'ang, provincia di Anbwei.
Alla fine della seconda guerra mondiale la bizzarra localizzazione delle prime scoperte sembrava suggerire che H. erectus (ancora chiamato Pithecanthropus) fosse una particolare espressione asiatica di uomo primitivo; poi, le nuove scoperte africane cominciarono a modificare questo punto di vista, e iniziò a farsi strada l'idea che H. erectus avrebbe potuto esistere anche in Europa.
Le scoperte africane di esemplari considerati appartenenti a H. erectus furono fatte a Ternifine, a est di Mascara in Algeria, a Sidi'abd ar-Rahman, Thomas Quarry, Salé e Rabat in Marocco, forse a Yayo (Koro Toro) nella Repubblica del Ciad, a Omo e Melka Kontouré in Etiopia, a East Turkana e forse Baringo in Kenya, a Olduvai in Tanzania e probabilmente a Swartkrans (Membro 2) in Sudafrica.
Parecchi fossili europei furono da molti scienziati considerati appartenenti a H. erectus, anche se altri studiosi, e specialmente F. C. Howell, ritenevano che facessero parte di H. sapiens, in una sua fase iniziale (sottospecie indeterminata) (v. Howell, 1978). Questi esemplari includevano la mandibola di Mauer, in Germania, l'osso occipitale e alcuni denti isolati di Vértesszöllös, in Ungheria, il cranio di Petralona, in Grecia nordorientale, i fossili di Arago, di Montmaurin e forse Lazaret (in Francia) e i frammenti di cranio di Bilzingsleben in Germania. Howell richiamò l'attenzione sul fatto che questi fossili europei si scostavano in parte dalla caratteristica morfologia di H. erectus, e li considerò come espressioni molto primitive di H. sapiens. Se questa ipotesi risultasse corretta, dovremo probabilmente ammettere che H. sapiens abbia avuto origine molto prima di 100 o 200.000 anni fa, e che esso sia esistito in Europa e forse nell'Africa settentrionale quando in alcune parti dell'Africa e dell'Asia si trovava ancora H. erectus. Secondo questa ipotesi, la fase H. erectus non è rappresentata tra i numerosi fossili di ominidi europei. Questi stessi fossili europei potrebbero anche essere considerati rappresentanti di una variante geografica, o sottospecie, di H. erectus. E già stata ammessa l'esistenza di numerose di tali sottospecie, come quella di Giava, quella cinese, quella dell'Africa nordoccidentale e quella dell'Africa orientale.
Così, all'inizio degli anni ottanta non era ancora stata raggiunta una opinione unanime su come e quando fosse avvenuta la transizione da H. erectus a H. sapiens. Quasi tutti gli studiosi di questo campo ritenevano che H. erectus fosse stato il predecessore di H. sapiens, ma c'era qualche divergenza di opinione riguardo alla classificazione di certi fossili, specie quelli europei. Tali differenze erano in genere dovute alla diversità delle definizioni date delle due specie, alle differenti opinioni riguardo alle implicazioni della morfologia e ai diversi concetti di variabilità intraspecifica. Era difficile stabilire se i fossili avessero caratteristiche intermedie o di transizione tra le due specie, perché spesso, a seconda di come veniva interpretata la loro evoluzione, alcune popolazioni della specie più antica potevano divenire i primi membri della specie successiva. Se c'era stata continuità tra i due successivi gruppi di popolazioni, dove si doveva tracciare la linea di demarcazione tra le due specie? Il problema divenne ancor più complesso da un punto di vista metodologico, via via che la documentazione fossile si faceva più ricca. Tanto più numerosi erano i fossili disponibili, tanto più probabile era che vi fossero delle forme intermedie. Proprio questa osservazione fornì la conferma che vi era stato un processo evolutivo da una forma nell'altra, e allo stesso tempo rese più difficile la classificazione e la sistematizzazione. Il problema è molto meno arduo per le specie viventi e contemporanee, tra le quali i confini sono in genere ben definiti. Nelle specie estinte, invece, specialmente quando vi sia la prova che esse si siano succedute l'una dopo l'altra, come è il caso di H. habilis, H. erectus e H. sapiens, questi confini sono molto meno netti. Non ci si deve meravigliare, pertanto, se in una sequenza come questa si sono trovati esemplari con caratteristiche intermedie tra H. habilis e H. erectus (come il fossile SK 847 di Swartkrans) e altri che appaiono come forme di transizione tra H. erectus e H. sapiens (come i fossili di Petralona e Arago).
Ai fossili generalmente considerati appartenenti a H. erectus viene attribuita un'età compresa tra un milione e mezzo e almeno 500.000 o forse 300.000 o anche 200.000 anni.
c) Alcune caratteristiche morfologiche di Homo erectus
La maggior parte delle caratteristiche più appariscenti che distinguono H. erectus riguardano crani e denti. La capacità cranica media di H. erectus era maggiore di quella di A. africanus e di H. habilis, ma inferiore a quella di H. sapiens. Abbiamo visto che era di 442 cc nel caso di A. africanus, e 645 cc per H. habilis. La capacità cranica media di H. erectus, senza tener conto di differenze geografiche o cronologiche, era di 941 cc, con un aumento di circa il 46% rispetto a H. habilis. L'ambito di valori riscontrato nei fossili disponibili di H. erectus va da circa 725 a 1.225 cc, e da alcune valutazioni risulta che vi fosse una differenza di capacità media tra gli esemplari giavanesi e quelli cinesi; infatti, per H. erectus di Giava è stato calcolato un valore medio di circa 883 cc, mentre era di circa 1.043 per gli esemplari cinesi trovati a Chukut'ien. In teoria, le diverse dimensioni cerebrali tra i due esemplari asiatici di H. erectus potrebbero esser state la conseguenza di un ulteriore incremento evolutivo verificatosi in popolazioni di H. erectus successive, come quelle cinesi. Altrimenti, potrebbero essere interpretate come una differenza nelle caratteristiche medie presenti in due diverse razze o sottospecie di H. erectus. La capacità cranica media dell'uomo moderno, appartenente alla specie H. sapiens, è 1.350 cc, cioè circa il 43% in più rispetto alla media relativa a H. erectus. Secondo G. P. Rightmire, in genere i più antichi membri di H. erectus avevano una inferiore capacità media (826 cc gli esemplari risalenti a 1,5 milioni di anni), quelli successivi maggiore (924 cc quelli di 800.000 anni fa, e 1.012 cc quelli di 600.000 anni fa); questi valori sono stati ottenuti senza considerare l'origine geografica dei fossili.
Vi sono dati che indicano che le dimensioni corporee di H. erectus erano superiori a quelle di H. habilis e circa uguali a quelle dell'uomo moderno. Pertanto, l'aumento della capacità cranica media dovrebbe, almeno in parte, essere considerato una conseguenza delle maggiori dimensioni corporee di H. erectus rispetto a H. habilis, e in parte della progressiva encefalizzazione verificatasi durante l'evoluzione di Homo.
La scatola cranica di H. erectus era bassa, con pareti restringentisi verso l'alto, e le ossa della volta erano estremamente ispessite; il rilievo sopraorbitario (torus) era molto pronunciato, e vi era una sporgenza ossea molto spessa nella parte posteriore del cranio. La fronte era sfuggente, e la parte anteriore del cranio, immediatamente dietro al torus sopraorbitario, presentava un restringimento da lato a lato. Il naso era largo, i mascellari e il palato erano ampi e piuttosto prominenti. I denti erano nell'insieme più larghi di quelli di H. sapiens, ma più piccoli di quelli di Australopithecus e H. habilis; gli incisivi e i canini erano particolarmente larghi per un ominide, alcuni addirittura più grandi di quelli di Australopithecus. Alcuni dei canini più grandi trovati in alcuni crani di H. erectus presentavano apici moderatamente sporgenti oltre la corona dei denti adiacenti. Le ossa degli arti e il bacino di H. erectus, pur se abbastanza simili a quelli di H. sapiens, mostrano alcune differenze anatomiche. Ciò nonostante, nell'insieme indicano che la postura e il tipo di locomozione degli ominidi a cui appartenevano non erano funzionalmente molto diversi da quelli dell'uomo moderno.
Sarebbe tuttavia errato ritenere che la transizione da H. habilis a H. erectus e H. sapiens sia stata graduale, continua. Vi è infatti un gruppo di caratteristiche che mostra come H. erectus si sia specializzato seguendo una linea di sviluppo diversa da quella che va dal suo presunto predecessore (H. habilis) al suo successore (H. sapiens); si tratta dell'ispessimento delle ossa del cranio e del particolare sviluppo delle arcate sopraorbitarie e del torus occipitale. Tali caratteristiche non si riscontrano nelle attuali scimmie antropomorfe, australopitecine o H. habilis, né nell'attuale H. sapiens. Questa combinazione di caratteristiche, che sono considerate ‛specializzate', ha suggerito ad alcuni scienziati l'ipotesi che H. erectus abbia rappresentato un ramo evolutivo che non può essere considerato quello da cui ha avuto origine H. sapiens. Contro questo punto di vista bisogna considerare: primo, che i fossili disponibili ci dicono relativamente poco della variabilità di queste caratteristiche; secondo, che nessun dato o ipotesi ragionevole è riuscito a spiegare in modo soddisfacente le condizioni fisiologiche, ecologiche o climatiche che possono aver determinato questo ispessimento del cranio, o l'effetto che l'incremento cerebrale, che è stato una caratteristica cosi rilevante agli effetti dell'avanzamento evolutivo degli individui all'inizio del Pleistocene medio, può aver avuto sullo sviluppo del cranio; e, terzo, il fatto stesso che le caratteristiche intermedie presenti in diversi dei crani più tardi, tanto che alcuni li considerano appartenenti a H. erectus e altri a H. sapiens avvalorano l'ipotesi che alcune popolazioni di H. erectus si siano evolute nelle più antiche popolazioni di H. sapiens. In effetti, i primi tipi di H. sapiens, appartenenti alle cosiddette sottospecie Neandertal e Rodesiana, presentano ancora caratteristiche simili a quelle di H. erectus.
Pertanto, questi tratti ‛specializzati' acquisiti di per sé non sono sufficienti a escludere che H. erectus sia stato il progenitore dei primi H. sapiens, anche se rimane da spiegare perché l'ispessimento delle ossa craniche e lo sviluppo di creste localizzate sul davanti e sul retro del cranio siano divenuti tratti tanto caratteristici di H. erectus. Per molti altri rispetti (denti, encefalizzazione e tipo di locomozione) la struttura corporea di H. erectus corrisponde perfettamente a ciò che si sarebbe potuto prevedere per una forma intermedia tra Australopithecus e H. habilis da un lato, e i primi H. sapiens dall'altro.
d) Caratteri comportamentali di Homo erectus
I siti in cui sono state effettuate le scoperte ci hanno aiutato a chiarire quali avrebbero potuto essere i tratti comportamentali di H. erectus. A Chukut'ien i resti di H. erectus sono stati rinvenuti in una caverna, ma di per sé questo fatto non indica che questa fosse una popolazione di cavernicoli. Tuttavia, la presenza di ossa animali carbonizzate, di semi, di focolari e carbone, di strumenti d'osso e di pietra, indicano che H. erectus ha vissuto a lungo a Chukut'ien come troglodita o cavernicolo. I resti di Lantian, Trinil, Sangiran, Mojokerto, come pure quelli di Ternifine e Olduvai, sono stati trovati tutti in località aperte, talvolta in ghiaie o argille fluviali, talvolta in arenarie fluviali o in conglomerati e rocce vulcaniche, o nell'invaso di laghi. Tali siti suggeriscono che H. erectus abbia vissuto anche in accampamenti all'aperto lungo le rive di corsi d'acqua o di laghi; anzi, la vicinanza all'acqua dev'essere stata decisiva per la sopravvivenza dell'uomo. I resti dell'uomo di Vértesszöllös mostrano come egli vivesse in terreni bassi e fangosi che si erano depositati insieme a minerali intorno alle sorgenti di un affluente del Danubio. Questi presunti accampamenti all'aperto contenevano una quantità di strumenti e schegge di pietra, ossa spezzate e in parte bruciate di animali probabilmente cacciati per procurarsi cibo, e tracce di qualcosa che appare essere stato un focolare.
Così, sia Chukut'ien sia Vértesszöllös indicano che i primi individui del Pleistocene medio erano in grado di utilizzare il fuoco, il che probabilmente facilitò la loro vita come cavernicoli e il loro trasferimento in climi più freddi. Anzi, proprio questo fattore avrebbe potuto favorire le migrazioni degli antichi uomini dai caldi tropici e subtropici (dove sembra abbiano avuto origine) verso le regioni fredde, spesso ghiacciate, dell'Europa e dell'Asia preistoriche.
I segni della cultura di H. erectus comprendono gli strumenti trovati negli stessi depositi in cui sono state rinvenute le sue ossa. In alcune aree, come Chukut'ien e Vértesszöllös, tali artefatti comprendevano strumenti per tagliare, ricavati da ciottoli di pietra, e anche schegge litiche e artefatti ossei. In altre aree, per esempio a Ternifine, sono state trovate asce bifacciali a mano e raschiatoi di un tipo che gli archeologi associano con la primitiva industria acheuleana. Insomma, in aree diverse del mondo, popolazioni della stessa specie di Ominidi sono state trovate insieme a materiale culturale in pietra molto differente. Questo fatto ci fa ricordare che gli aspetti ereditari o razziali della struttura corporea non sono inestricabilmente connessi con manifestazioni di tipo culturale come le tradizioni relative alla costruzione di strumenti; sarebbe stato avventato e sbagliato il tentativo di risalire all'aspetto fisico di una popolazione dagli strumenti litici che era in grado di fabbricare.
Insieme ai frammenti di H. erectus sono state trovate numerose ossa di animali non ominidi, molte delle quali sembrano rotte di proposito, cotte e anche lavorate. Ciò sembra confermare che H. erectus era un cacciatore e si cibava di carne fresca. La sua dieta era composta, oltre che dalla carne, da vegetali, come foglie carnose, frutta, bacche e radici; per esempio, nella caverna di Chukut'ien è stata trovata una raccolta di semi di Celtis.
Sembra non possano esserci dubbi sul fatto che H. erectus fosse onnivoro (come lo è oggi H. sapiens). Questa è la dieta più opportuna e H. erectus è stato probabilmente il primo dei grandi opportunisti: proprio questa caratteristica lo ha dotato della sua grande flessibilità e adattabilità evolutiva.
Tra le popolazioni di H. erectus non troviamo segni evidenti di sepoltura rituale dei morti. Non sono state trovate sepolture complete né tombe o corredi funebri insieme o vicino alle ossa. Quanto ad altri segni di cerimonie magiche o religiose, è probabile che venisse praticato il cannibalismo rituale. Le ossa umane di Chukut'ien risultavano spezzate nello stesso modo di quelle animali. Tuttavia, nonostante che tra le migliaia di frammenti di ossa non ominidi trovati nella caverna fossero rappresentate tutte le parti dello scheletro, la stragrande maggioranza dei resti dei più di 40 ominidi trovati nella caverna era composta di crani, e solo in piccola parte da ossa degli arti. Sembra pertanto che le teste siano state deliberatamente prescelte, come accade ancor oggi tra i cacciatori di teste. Ciò fa pensare all'esistenza di un culto di questo tipo tra gli abitanti di Chukut'ien.
In praticamente tutti i crani di H. erectus, la base risultava spezzata nella regione vicino al foramen magnum (l'apertura attraverso la quale il cervello si prolunga nel midollo spinale). Anche ominidi più tardi, come i neandertaliani del Monte Circeo, alcune popolazioni dell'Età del Bronzo in Germania e alcune popolazioni moderne del Pacifico, praticavano intenzionalmente una mutilazione della base cranica. Sulla base di questa analogia, alcuni scienziati, come A. C. Blanc, hanno ritenuto non solo che la frattura delle base cranica fosse praticata da H. erectus intenzionalmente, ma che si trattasse di una mutilazione rituale, probabilmente connessa con la manducazione rituale del cervello. Se questa interpretazione è giusta, la caccia di teste, la mutilazione rituale della base cranica e forse il cannibalismo rituale rappresenterebbero le prime espressioni di riti non solo di H. erectus, ma dell'intera storia dell'evoluzione ominide.
È indubbio, comunque, che H. erectus fosse in possesso di una cultura complessa, ben superiore a qualunque altra evidenziabile nei depositi fossiliferi dei più antichi ominidi.
e) La collocazione di Homo erectus nella filogenesi ominide
Secondo l'opinione della maggior parte degli studiosi di fossili umani, Homo erectus è stato l'immediato progenitore di H. sapiens. Taluni hanno ritenuto le relazioni tra i due taxa così strette da suggerire che H. erectus debba essere incluso nella specie H. sapiens in qualità di sottospecie primitiva, H. sapiens erectus, contrapposta alla successiva specie H. sapiens sapiens. All'inizio degli anni ottanta, tuttavia, secondo l'opinione prevalente le basi morfologiche erano sufficienti a mantenere separate le specie dei due gruppi di Ominidi.
Molti ritenevano estremamente probabile anche il fatto che H. habilis si fosse evoluto in H. erectus, e pertanto H. habilis, H. erectus e H. sapiens venivano considerate tre specie successive (o cronospecie) della stessa linea evolutiva. Vi era buon accordo anche circa il periodo durante il quale era avvenuta la transizione tra le prime due (1,6-1,5 milioni di anni fa), mentre le valutazioni del periodo di transizione fra le altre due, cioè H. erectus e H. sapiens, davano risultati compresi tra 500.000 e 100.000 anni fa, a seconda di come i vari paleoantropologi interpretavano la morfologia di certi fossili, e delle definizioni date delle due specie.
Nonostante le diverse opinioni degli studiosi riguardo a questi dettagli, in genere tutti concordavano su quale fosse il posto occupato da H. erectus nella linea evolutiva che porta all'uomo moderno.
9. La fase della comparsa dell'uomo moderno
Appartengono a questa fase numerosi esemplari considerati di transizione tra H. erectus e le prime sottospecie di H. sapiens; tre importanti sottospecie, note col nome di Homo sapiens soloensis, Homo sapiens rhodesiensis e Homo sapiens neanderthalensis; alcune sottospecie più tarde di H. sapiens, gli immediati precursori dell'attuale uomo. In relazione a certi periodi di tempo, la documentazione fossile è così ricca che abbiamo l'imbarazzo della scelta. Durante gli stadi più tardi di questo periodo, due fattori hanno grandemente aumentato le probabilità che i fossili si preservassero, e cioè la diffusa abitudine di vivere in caverne e quella di seppellire i morti. Abbiamo perciò centinaia di fossili che risalgono a questa fase durante la quale fece la sua prima comparsa l'uomo moderno; gli esemplari sono così numerosi che sarebbe non solo inutile, ma addirittura controproducente ai fini della chiarezza enumerarli singolarmente.
Possiamo dire che all'inizio dell'ultimo quarto del secolo era ormai generalmente accettata l'idea che H. sapiens si fosse evoluto da H. erectus, anche se bisognava ancora trovare una definizione di H. sapiens chiara e di soddisfazione per tutti, e stabilire dove e quando fosse avvenuta la differenziazione della specie. Tuttavia, la documentazione fossile comprende alcuni resti di ominidi provenienti da Africa, Europa e Asia che sembrano chiarire le origini di H. sapiens.
Nel cercare una definizione di H. sapiens siamo portati a far riferimento alle popolazioni viventi. Infatti, quando il grande naturalista e sistematico svedese C. von Linné stabili la prima classificazione degli esseri umani viventi, per descrivere le caratteristiche del genere Homo nel quale li aveva inclusi scrisse semplicemente ‟Nosce te ipsum" (Systema Naturae, 1735).
Nella seconda metà del XX secolo apparvero due utili definizioni di H. sapiens: la prima fu quella di Le Gros Clark (v., 19642) e comprendeva una lista di tratti distintivi essenziali. Alla luce di successivi studi effettuati sui primi H. sapiens, come ha giustamente fatto rilevare Howell (v., 1978, p. 201), tale definizione risultava così restrittiva da escludere ‟le precedenti popolazioni del Pleistocene superiore, che tuttavia presentano alcune caratteristiche che ricordano quelle della specie".
La seconda definizione è stata quella di Howell, anche se preceduta dall'affermazione che ‟non si cerca di dare una definizione onnicomprensiva della specie sapiens" (ibid.). Howell fece un lungo elenco di tratti evolutivi e caratteristiche derivate, sia comportamentali che morfologiche, evidenti nelle popolazioni di H. sapiens sapiens risalenti all'epoca a cavallo tra Pleistocene superiore e Recente, paragonandoli con quelli di H. erectus. Questa definizione è troppo lunga e troppo tecnica per essere riportata in questo articolo; rimandiamo perciò il lettore interessato alla pubblicazione originale.
a) Le forme di transizione erectus-sapiens
Alcuni fossili del Vecchio Mondo sono risultati per lungo tempo difficili da interpretare, e sono stati da qualcuno considerati membri di H. erectus, da altri di H. sapiens. In effetti presentano caratteristiche che possono essere considerate di transizione tra le due specie.
I fossili europei comprendevano i crani di Swanscombe e di Steinheim, e molti di quelli compresi nella definizione di ‛H. erectus europeo' (come la mandibola di Mauer in Germania, il cranio di Petralona in Grecia, i resti di Vértesszöllös in Ungheria, i fossili di La Chaise in Francia). In genere, presentavano cervelli più grandi e denti più piccoli di quelli di H. erectus, e tratti specializzati di erectus (come ispessimento delle ossa craniche e torus sopraorbitario e occipitale) meno pronunciati. La maggior parte di questi esemplari risale al Pleistocene medio, a un periodo, cioè, compreso tra 300.000 e 100.000 anni fa.
Molti degli esemplari rivenuti nel Maghreb, in Africa nordoccidentale, presentano caratteristiche di transizione erectus-sapiens: tra questi comprendiamo il cranio di Sālē, e, secondo Howell, il secondo ominide della Thomas Quarry. Anche questi fossili, pur se con qualche incertezza, sono stati fatti risalire al Pleistocene medio. Le caratteristiche della mandibola di Baringo e del cranio di Ndutu, in Africa orientale, sono state giudicate intermedie; si è ritenuto che la prima risalga al Pleistocene medio, e il secondo forse a 500.000 anni fa.
Questa grande quantità di fossili africani ed europei, oltre ai numerosi esemplari di forme intermedie trovati in Cina, ha fatto ritenere che all'inizio del Pleistocene medio o alla fine del Pleistocene inferiore abbiano cominciato a manifestarsi numerose divergenze dalla morfologia tipicamente erectus in popolazioni per altri aspetti erectus. Questi differenziamenti anatomici avvennero generalmente in direzione sapiens. La combinazione di caratteristiche erectus e sapiens in queste popolazioni potrebbe essere considerata come la prima manifestazione della differenziazione in senso sapiens di una popolazione fondamentalmente erectus.
b) Le più antiche forme identificate come Homo sapiens
Oltre alle popolazioni di transizione appena ricordate, quattro sono state le sottospecie in cui si è individuata la piena espressione della specie H. sapiens nelle sue forme più antiche: H. sapiens rhodesiensis, dell'Africa meridionale, H. sapiens soloensis, di Giava, H. sapiens neanderthalensis, di Europa, Africa e area litoranea del Mediterraneo, e H. sapiens afer, dell'Africa subsahariana. Le prime tre possedevano crani con pesante torus sopraorbitario, torus occipitale massiccio, e spessore della calotta variabile da modesto a pronunciato; queste caratteristiche sono considerate di derivazione H. erectus. La capacità cranica media di H. rhodesiensis (oltre 1.250 cc) e di H. soloensis (1.151 cc) era maggiore di quella riscontrata negli esemplari di H. erectus, ma non così grande come quella del moderno H. sapiens. Oltre alla maggiore espansione cerebrale rispetto a H. erectus, i crani di queste tre sottospecie mostrano un minor grado di assottigliamento verso l'alto delle pareti della scatola cranica, specie se guardati dal di dietro, e un minor grado di appiattimento della parte superiore del cranio.
Per ciò che riguarda il taxon rhodesiensis, siamo in possesso di due crani molto ben conservati, provenienti uno da Kabwe (già Broken Hill, Zambia) e l'altro da Hopefield, Saldanha Bay (Sudafrica). Un frammento di mandibola, trovato nella Caverna di Hearths, Makapansgat (Transvaal settentrionale), è stato considerato della stessa sottospecie e anche alcuni frammenti cranici provenienti dal lago Eyasi in Tanzania potrebbero essere appartenuti a rhodesiensis.
Appartengono alla sottospecie soloensis alcuni crani rinvenuti a Ngandong (Giava).
c) Neandertal
Appartengono a questo gruppo più di 100 fossili provenienti da Europa, Asia occidentale e area litorale del Mediterraneo. Il nome significa ‛Valle del Neander' e si deve al fatto che il primo esemplare descritto è stato trovato nella valle del fiume Neander, in Germania, nel 1856. In un primo tempo si pensò che questi resti appartenessero a un genere diverso, in quanto si ritennero ‛estremamente primitivi' e ben diversi dagli odierni esseri umani. A quel tempo non erano ancora stati trovati resti di ominidi più antichi e più primitivi. In un secondo momento vennero considerati come una specie diversa del genere Homo, e finalmente, nell'ultimo quarto del XX secolo, furono ritenuti una sottospecie estinta della specie H. sapiens, alla quale venne dato il nome di H. sapiens neanderthalensis, che divenne la meglio conosciuta delle tre più antiche sottospecie di H. sapiens. Questa popolazione aveva cervello di grande dimensione (diversi esemplari avevano una capacità cranica addirittura superiore a quella media del moderno H. sapiens); tuttavia, i crani avevano mantenuto una serie di caratteristiche arcaiche che sembravano derivare da un progenitore erectus, come fronte sfuggente, arcata sopracciliare prominente, calotta cranica con sommità piuttosto appiattita, torus occipitale, spessore delle ossa craniche maggiore di quello del moderno H. sapiens, mento osseo scarsamente sviluppato, palato e mascellare superiore allargati, e un certo rigonfiamento di quest'ultimo osso. Lo scheletro postcraniale, nonostante la struttura tozza e massiccia e le larghe superfici articolari delle ossa degli arti, era in pratica molto simile a quello dell'uomo moderno.
I fossili considerati neanderthalensis sono particolarmente numerosi in Europa; ne sono infatti stati trovati in Francia, Belgio, Germania, Gibilterra, Iugoslavia, Italia, Portogallo, Spagna, Unione Sovietica, Israele e Iraq. Una mezza dozzina di esemplari che alcuni studiosi ritengono appartenere a questa sottospecie sono stati trovati nel litorale mediterraneo meridionale, per esempio a Haua Fteah, in Cirenaica, e Jebel Irhoud, in Marocco, anche se quest'ultimo cranio mostra dei tratti più sviluppati o più simili a quelli dell'uomo moderno di quelli riscontrabili nella forma classica. Essi vissero nel periodo compreso all'incirca tra 100.000 e 40.000 anni fa.
Questa sottospecie è caratterizzata da una vita culturale ricca e complessa, comprendente abitudini quali vivere in caverne, uso del fuoco, tecniche culturali e industrie litiche progredite, sepoltura dei morti e pratiche rituali a essa associata.
Howell ha messo in relazione lo sviluppo e la piena espressione delle caratteristiche neandertaliane con l'ecologia glaciale o periglaciale dell'Europa occidentale: secondo la sua opinione, in questo ambiente difficile si svilupparono le caratteristiche neandertaliane classiche, non altrettanto evidenti al di fuori dell'area glaciale dell'emisfero settentrionale.
Alla pari delle altre tre più antiche sottospecie di H. sapiens, la neanderthalensis pare si sia sviluppata da un progenitore erectus. Numerosi esemplari con caratteristiche intermedie dimostrano che in Europa e in Medio Oriente almeno una parte dell'eredità genetica neandertaliana si continua nel successivo H. sapiens sapiens. Esemplari di tipo intermedio sono stati trovati nell'Europa centrale (Cecoslovacchia) e sul monte Carmelo, in Israele. Mentre nell'Europa occidentale sembra che la forma più avanzata di uomo neandertaliano sia stata sostituita dall'attuale forma di uomo (H. sapiens sapiens) in modo piuttosto improvviso, in Europa centrale si è verificata una transizione più graduale da forme di tipo neandertaliano a forme più moderne.
L'uomo di Neandertal è essenzialmente limitato al periodo tra la fine del Pleistocene medio e il Pleistocene superiore, ed è stato sostituito, circa 40.000 anni fa, dalle prime forme europee di uomo moderno.
d) L'uomo di Cro-Magnon
La prima popolazione sapiens moderna comparsa in Europa e nel litorale Mediterraneo è spesso indicata col nome di ‛razza di Cro-Magnon': Cro-Magnon è infatti il nome di un riparo roccioso nel villaggio di Les Eyzies de Tayac, in Dordogna (Francia), dove nel 1868 furono trovati i resti fossili di un ‛antico uomo' insieme a quelli di altri individui. Il deposito dal quale questi vennero estratti si ritiene risalga alla terza fase (o stadio) dell'ultima delle grandi glaciazioni dell'emisfero settentrionale, detta Würm III; la fase interglaciale che la precedette è chiamata Eem, e il periodo a cavallo tra le due (Eem-Würm) risale con tutta probabilità a 75.000 anni fa. Questa data corrisponde all'acme evolutivo dei neandertaliani. La fase Würm III risale a un periodo più tardo del Pleistocene e l'ambito di tempo al quale può esser fatta risalire la popolazione Cro-Magnon di H. sapiens sapiens è compreso tra circa 35.000 e 10.000 anni fa.
Dal 1868 sono stati scoperti numerosi altri esemplari di uomini di questo tipo, che sembrano i diretti progenitori del moderno ramo caucasoide dell'umanità. Dove esattamente abbiano fatto la loro comparsa non è noto, anche se alcuni hanno avanzato l'ipotesi che avrebbe senz'altro potuto trattarsi dell'Europa sudorientale e del litorale mediterraneo orientale. La struttura scheletrica dell'uomo di Cro-Magnon era quasi identica a quella dell'uomo moderno, e non presentava alcuna di quelle specializzazioni ossee tipiche dei neandertaliani, quali platicefalia, torus occipitale, torus sopraorbitario, ‛rigonfiamento' del mascellare superiore, ossa degli arti con estremità molto allargate. La mancanza di questi tratti caratteristici e la presenza di un mento osseo inserito sulla mandibola, la fronte prominente e quasi verticale, ecc., dimostrano che la morfologia dell'uomo di Cro-Magnon era decisamente evoluta nella direzione dell'uomo moderno.
Anche il progresso culturale era estremamente evidente. La documentazione archeologica è ricca di strumenti di pietra, osso e legno differenziati e di complessità molto maggiore. Uno sviluppo notevolissimo è testimoniato dalla comparsa di espressioni artistiche. Anche se risulta che già da lungo tempo gli ominidi del Pleistocene raccoglievano terre colorate, specie l'ocra rossa, solo con l'uomo di Cro-Magnon si hanno le prime e copiose manifestazioni di espressioni artistiche quali pitture, graffiti e sculture; naturalmente, anche le pratiche rituali come la sepoltura dei morti, presenti già fra i neandertaliani, vengono mantenute.
Per gran parte del XX secolo si è ritenuto che quella di Cro-Magnon fosse la più antica popolazione di Homo sapiens anatomicamente moderna; tuttavia, a partire dall'ultimo quarto del secolo si sono venuti accumulando dati che sembrano indicare la presenza in Africa di una forma ancora più antica di uomo moderno.
e) Homo sapiens afer
Benché Linneo abbia dato una definizione del genere Homo nella prima edizione del Systema naturae (1735), solo nelle edizioni successive dette il nome di Homo sapiens all'uomo moderno, dando anche una classificazione dell'uomo attuale. La sua quadripartizione dell'umanità in varietà diverse nei quattro quarti del globo a seconda del colore della pelle (quella americana, rossa; quella europea, bianca; quella asiatica, bruna; quella africana, nera), non ha tuttavia resistito alla prova del tempo. È comunque interessante notare che una delle categorie da lui proposte, Homo sapiens afer, è stata fatta rivivere da L. H. Wells (v., 1969 e 1972) quale sottospecie comprendente tutti i resti umani provenienti dal Sudafrica, risalenti al tardo Pleistocene. Questa sottospecie avrebbe dovuto comprendere quelle popolazioni ritenute il comune progenitore delle popolazioni Khoisan e Negra dell'attuale Africa subsahariana; Tobias indica questa popolazione ancestrale comune col nome ‛proto-negriforme'.
Si ritiene che diversi resti umani appartengano a questa popolazione africana ancestrale: tra questi sono compresi quelli di Border Cave, nel Natal, di Tuinplaas, Springbok Flats, in Transvaal, di Fish Hoek, Matjes River e Klasies River Mouth Cave, sulla costa meridionale del Capo. Anche il cranio di Florisbad, vicino a Bloemfontein (Orange Free State), potrebbe appartenere a questa stessa sottospecie, pur mantenendo alcuni tratti morfologici che ricordano H. sapiens rhodesiensis. Altri resti riferibili a questa sottospecie sono stati scoperti in Zambia, Kenya ed Etiopia meridionale (v. Howell, 1978). La morfologia di questi fossili subsahariani appare essenzialmente moderna. All'inizio dell'ultimo quarto del secolo, alcuni studiosi hanno sostenuto che parecchi esemplari di questi ominidi africani, in particolare quelli di Border Cave e di Klasies River Mouth Cave, avessero un'età superiore ai 100.000 anni; se ciò risultasse vero, significherebbe che proprio in Africa hanno avuto origine i più antichi Homo sapiens di tipo moderno, che risalirebbero a decine di migliaia di anni prima dei fossili anatomicamente moderni trovati in Europa o in Asia. La modernità anatomica di questi esemplari è fuori di dubbio, ma permane qualche incertezza circa la loro datazione. È innegabile tuttavia che da diversi siti dell'Africa meridionale siano emerse prove che dimostrerebbero come alcune forme moderne di uomo risalgano addirittura al Pleistocene superiore o perfino al Pleistocene medio.
Un'altra di queste forme apparentemente moderne di Homo sapiens è stata rinvenuta nella formazione Kibish del Bacino del fiume Omo, in Etiopia sudoccidentale. Anche se le affinità morfologiche di questi resti sono state interpretate in maniera contrastante, l'analisi effettuata da C. Stringer (v., 1974) proverebbe che i crani di Kibish, nonostante siano molto dissimili fra loro, sarebbero decisamente diversi da H. sapiens soloensis e H. sapiens rhodesiensis e senz'altro più simili a rappresentanti di H. sapiens di un periodo più tardo del Pleistocene (v. Howell, 1978). È possibile che la variazione intraspecifica sia sufficiente a comprenderli nella sottospecie H. sapiens afer, oppure potrebbero rappresentare un'altra sottospecie di H. sapiens non ancora identificata. Anche per i crani di Kibish è stata proposta una datazione di oltre 100.000 anni, che non è stata, però, ancora confermata.
Altri resti di possibili ominidi africani, antichi ma simili al moderno H. sapiens, sono stati trovati a Kanjera nel Kenya occidentale; la loro datazione non è tuttavia sicura, benché K. P. Oakley (v., 1974) abbia messo in evidenza come tali frammenti cranici umani siano notevolmente più giovani della fauna presente a Kanjera nel Pleistocene medio.
All'inizio degli anni ottanta, quindi, il mondo della paleoantropologia non aveva ancora trovato un accordo sulla datazione da attribuire ai più antichi reperti africani di uomo anatomicamente moderno, nè l'idea di una possibile origine africana dell'uomo anatomicamente moderno aveva trovato posto nelle teorie relative alla comparsa del moderno H. sapiens. Analogamente, devono ancora essere chiariti i rapporti genetici e filogenetici tra H. sapiens afer e H. sapiens sapiens. Possiamo solo dire che le più recenti scoperte africane potrebbero rimettere in discussione l'ipotesi eurocentrica e mediterraneocentrica dell'origine del moderno H. sapiens.
f) Il moderno Homo sapiens in Asia
Abbiamo già ricordato la presenza di resti neandertaliani in Iraq e in Israele. Oltre a questi, sono stati scoperti due gruppi di forme essenzialmente moderne di uomo, la cui età può esser fatta risalire a quella delle più antiche forme di H. sapiens sapiens europee. Dalla caverna di Skhul, sul monte Carmelo in Israele, sono stati portati alla luce dei crani estremamente simili alle forme europee moderne, e a essi è stata attribuita un'età di 35-40.000 anni; sarebbero perciò un po' più antichi dei più antichi esemplari Cro-Magnon dell'Europa occidentale.
Anche il cranio della caverna di Niah, a Sarawak nel Borneo settentrionale, avrebbe pressappoco la stessa età. Anch'esso ha una forma simile al moderno sapiens e vi si possono riscontrare delle somiglianze con le popolazioni australiane, specie quelle abitanti un tempo in Tasmania.
Queste forme moderne scoperte in Israele e nel Borneo hanno comunque un'età di 40.000 anni, risalgono cioè al Pleistocene superiore; è stato sostenuto, invece, che in Cina sono stati trovati degli esemplari di Homo sapiens risalenti addirittura al Pleistocene medio. Il meglio conservato di questi fossili è il cranio di Dali, e un altro buon esemplare è quello del distretto Maba. Tuttavia questi fossili possono essere considerati gli equivalenti morfologici di forme primitive di H. sapiens, quali soloensis, rhodesiensis o neanderthalensis, e non una primitiva espressione di H. sapiens anatomicamente moderno.
g) Riepilogo
La comparsa, circa 35-40.000 anni fa, di una forma moderna di H. sapiens è stata dimostrata in tutte e tre le grandi regioni del Vecchio Mondo, cioè Europa, Africa e Asia. Inoltre, un piccolo ma crescente numero di dati provenienti dall'Africa subsahariana sembra indicare che una forma essenzialmente moderna di H. sapiens avrebbe potuto aver origine in Africa ancora prima di 40.000 anni fa, forse addirittura 100.000 (data ritenuta il confine tra Pleistocene medio e superiore). La datazione, peraltro non ancora confermata, attribuita ai fossili di Kibish, Border Cave e Klasies River Mouth ha fatto avanzare l'ipotesi che forme anatomicamente moderne di H. sapiens avrebbero potuto aver origine in Africa già nell'ultimo periodo del Pleistocene medio.
Molto rimane tuttavia da chiarire riguardo al tempo e al luogo della prima comparsa dell'uomo moderno. Le ricerche di genetica delle popolazioni, basate sulla frequenza di specifici marcatori genici in varie popolazioni, hanno mostrato che le popolazioni europee moderne (caucasiformi) e africane (negriformi) hanno una maggiore affinità genetica di quanto sia le une che le altre presentino rispetto a quelle asiatiche (mongoliformi). Ciò potrebbe voler dire che l'uomo africano e quello europeo hanno avuto un comune progenitore in tempi più recenti che non le popolazioni afro-europee e asiatiche. È senz'altro sorprendente, come ha fatto rilevare Wu Rukang (v., 1982), che gli ominidi fossili cinesi, dai più antichi esemplari di H. erectus all'attuale Homo sapiens - come gli esemplari di Chukut'ien -, mostrino tutti delle caratteristiche decisamente mongoliformi, come incisivi superiori a forma di pala, zigomi sporgenti, ossa nasali orientate in senso più perpendicolare al massiccio facciale, ‛cresta' sagittale dell'osso frontale e presenza di ossa wormiane. Ciò sembrerebbe indicare che buona parte dell'eredità genetica dei popoli mongoliformi deriva da popolazioni ancestrali situate nelle stesse zone geografiche, anche se i movimenti di popolazioni e il flusso genetico devono aver giocato un ruolo importante, come è dimostrato dal fatto che anche tra i caucasiformi e i negriformi si rileva la presenza di incisivi superiori a forma di pala, seppure con una frequenza inferiore a quella riscontrabile tra i mongoliformi.
Sulla base dei dati disponibili attualmente, gli Ominidi sono considerati come un gruppo di Primati fondamentalmente africano, la cui età è compresa tra circa 5 e circa 1,5 milioni di anni dal presente (che sono le età di Australopithecus e Homo habilis). In un periodo di tempo compreso tra 1,5 e 0,5 milioni di anni fa essi si diffusero dall'Africa, in cui presumiamo abbiano avuto origine, in vaste zone dell'Europa e dell'Asia (stadi evolutivi Homo erectus e prime forme di Homo sapiens); infine, durante gli ultimi 100.000 anni, l'uomo è comparso anche nelle due ultime grandi aree abitabili della terra, America e Australia (stadio evolutivo Homo sapiens).
Il passaggio dell'uomo in America avvenne attraverso una striscia di terra, la Beringia, situata tra la Siberia e l'Alaska (dove attualmente si trova lo stretto di Bering), in un periodo durante il quale il livello del mare era più basso di quanto non sia adesso. La popolazione che migrava era mongoliforme ed è probabile che tale attraversamento sia avvenuto più volte. Sono stati trovati diversi fossili che rappresentano questi primi abitanti del Nuovo Mondo. Per entrare in Australia dall'Asia sudorientale e dal grande arcipelago Indonesiano, invece, l'uomo (e molti altri mammiferi terrestri) doveva attraversare una grande estensione di acque profonde nota come ‛linea di Wallace' (dal nome di A. R. Wallace, che è considerato, insieme a Darwin, il co-scopritore della teoria della selezione naturale). Quando il livello del mare era più basso, la linea di Wallace separava due grandi estensioni di terra, l'altopiano sottomarino della Sonda, che univa le Filippine, Borneo, Giava e Sumatra al continente asiatico, e l'altopiano sottomarino di Sahul, che connetteva la Nuova Guinea e l'Australia. Oggi questa linea corre tra Bali e Lombok, Borneo e Celebes, Mindanao e le isole di Sangi e Tahuna. Per attraversare questa linea e raggiungere l'Australia, l'uomo deve aver in qualche modo passato il mare; del resto era sufficiente un numero relativamente piccolo di individui e poche traversate per fondare la popolazione degli aborigeni australiani. Rappresentanti fossili di questi antichi immigrati australiani sono stati trovati a Talgai, Cohuna, Keilor, Kow Swamp e Lake Mungo.
10. Alcuni concetti relativi all'evoluzione degli Ominidi
Molti dei concetti applicabili all'evoluzione degli esseri viventi sono stati usati con successo nell'analisi dell'evoluzione degli Ominidi.
a) Modelli filogenetici
La scoperta di un sempre maggior numero di fossili di Ominidi e la loro datazione, effettuata con metodi di crescente accuratezza, hanno rivelato l'esistenza di almeno due modelli filogenetici nell'evoluzione umana. Il primo è chiamato evoluzione filetica, e indica il progressivo differenziamento che si produce lungo una determinata genealogia (la specie A dà origine alla specie B, e questa alla specie C). E questo il modello che sembra riscontrabile nella genealogia di Homo: Homo habilis → Homo erectus → Homo sapiens. Questo stesso modello potrebbe essere stato operante anche nella genealogia dell'antico Australopithecus gracile (sia che consideriamo questo taxon fossile un'antica sottospecie di A. africanus, sia che lo consideriamo una specie diversa, A. afarensis) dalla quale si sarebbe originato il successivo A. africanus.
Il secondo modello filogenetico è chiamato cladistico, o evoluzione ramificata. È esemplificato dalla grande divergenza (o cladogenesi) verificatasi circa 2,5-2,0 milioni di anni fa nella genealogia degli Ominidi, che ha dato origine alle due o tre linee evolutive delle australopitecine robuste e di Homo. Secondo alcuni scienziati, questa particolare divergenza avrebbe potuto essere più complessa, avrebbe potuto, cioè, dare origine non a due, ma a tre o anche quattro linee evolutive: A. robustus in Africa meridionale, A. boisei, in Africa orientale, A. africanus in una forma immutata o scarsamente mutata perdurante fino a una data molto posteriore (come quella suggerita da T. C. Partridge e K. Butzer per il cranio di Taung), e Homo habilis.
Secondo una teoria in auge negli anni settanta, i principali mutamenti evolutivi capaci di originare nuove specie sono avvenuti ai punti di cladogenesi (chiamati in questa teoria interpunzioni). Però, gli intervalli compresi tra due successive interpunzioni erano caratterizzati da periodi di equilibrio durante i quali si verificavano pochi o nessuno di quei mutamenti evolutivi che determinavano la speciazione (cioè la formazione di nuove specie). Pertanto questa teoria, che è in genere associata ai nomi di S. J. Gould e N. Eldredge (v., 1977; v. Eldredge e Gould, 1972), è nota col nome di punctuated equilibrium (equilibrio con interpunzioni). È interessante rilevare che Darwin stesso nella quarta edizione di The origin of species (1866) e nelle successive ammetteva la possibilità che nel corso dell'evoluzione lunghi periodi di equilibrio precedessero e seguissero periodi di cambiamenti più rapidi. Egli affermava di non ritenere che il processo di cambiamento ‟proceda continuativamente; è molto più probabile che ogni forma permanga inalterata per lunghi periodi e che poi nuovamente si modifichi" (v. Darwin, 18726; tr. it., p. 128).
Tuttavia, una differenza importante tra la teoria darwiniana e quella dell'equilibrio con interpunzioni è che secondo Darwin le interpunzioni erano punti di rapida modificazione della specie, ma tali modificazioni potevano assumere la forma (per usare dei termini moderni) di speciazione sia filetica (graduale) sia cladistica (ramificata). A seconda delle circostanze, poteva venir adottato l'uno o l'altro dei due modelli evolutivi. Per Eldredge e Gould, invece, le interpunzioni sono fasi di modificazioni accentuate, necessariamente associate a eventi cladistici o di ramificazione, cioè alla divergenza evolutiva di una specie in due o più specie. L'interpretazione data da Darwin degli eventi evolutivi ammetteva un maggior numero di possibili modelli di modificazione; la teoria di Eldredge e Gould limita il numero dei modelli filogenetici e, in particolare, praticamente esclude la possibilità che nella speciazione intervengano modificazioni graduali o di tipo filetico.
Coloro che hanno criticato la teoria di Eldredge e Gould hanno richiamato l'attenzione sul fatto che tale modello, specialmente se riferito ai mammiferi terrestri e agli Ominidi, non sembra caratterizzarne per intero la storia, per cui bisogna ritenere che anche il gradualismo filetico abbia avuto luogo. In particolare, per ciò che riguarda gli Ominidi, la ricostruzione della loro filogenesi che ha incontrato maggior favore alla fine degli anni settanta ha messo in evidenza una importante interpunzione verificatasi circa 2,5-2,0 milioni di anni fa. Questa fu seguita, nella linea evolutiva che porta a Homo sapiens, da una lunga fase di gradualismo filetico che ha portato questa linea a passare attraverso tre stadi successivi (habilis, erectus, sapiens) senza che si sia potuta rilevare la presenza di ramificazioni. D'altra parte, nel periodo precedente ai 2,5 milioni di anni fa avrebbe benissimo potuto esserci stata una lunga fase di equilibrio evolutivo, con relativamente pochi cambiamenti biologici significativi degli antichi ominidi australopitecini. Pertanto, nella storia della filogenesi ominide, quale è stata ricostruita, sembra siano intervenuti sia modelli di evoluzione con interpunzioni, sia di gradualismo filetico.
b) Evoluzione a mosaico
Abbiamo dunque mostrato come i cambiamenti evolutivi nella linea ominide siano avvenuti con velocità diverse e in tempi diversi; lo stesso si può dire riguardo alle diverse parti del corpo e ai diversi complessi strutturali-funzionali. Così, la postura eretta e il sistema di locomozione si sono evoluti durante uno stadio molto precoce della storia ominide, mentre il cervello ha raggiunto tardi le attuali dimensioni assolute. A questo concetto, al fatto cioè che le varie parti del corpo si siano evolute in tempi diversi e con diversa velocità, viene dato il nome di ‛evoluzione a mosaico'.
c) Gli ominidi che non hanno avuto successo
Dato che si è verificata una ramificazione della linea evolutiva ominide verso la fine del Pliocene, è chiaro che non tutti gli ominidi più antichi sono stati progenitori dei successivi e dell'uomo moderno. Per esempio, Australopithecus robustus e A. boisei si trovano su rami collaterali dell'albero che rappresenta la famiglia dell'uomo e non sono stati progenitori di Homo.
d) Evoluzione culturale
Una importante caratteristica della documentazione paleoantropologica è che i cambiamenti anatomici e i presunti cambiamenti funzionali hanno proceduto di pari passo, specie durante il Pleistocene, con comportamenti e modi di vita di crescente complessità. Importanti traguardi nella storia della cultura sono: l'acquisizione di abitudini venatorie (invece che di sola raccolta); l'uso della pietra come materiale da cui ricavare utensili; l'uso del fuoco e il suo controllo; lo sviluppo dell'arte e della rappresentazione simbolica; la nascita di pratiche rituali; l'acquisizione del linguaggio articolato e la sua evoluzione. L'uomo è divenuto l'animale culturale per eccellenza.
(L'Autore desidera ringraziare per la collaborazione prestata il prof. B. Merry, e inoltre K. V. Tangney, K. Copley, M. Heldan, A. R. Hughes, P. Faugust, C. Richards, A. Webb e J. Bunning).
Bibliografia
Blanc, A. C., Il sacro presso i primitivi, Roma 1945.
Blanc, A. C., Some evidence for the ideologies of early man, in Social life of early man (a cura di S. L. Washburn), Chicago 1961.
Brain, C. K., The hunters or the hunted? An introduction to African cave taphonomy, Chicago-London 1981.
Broom, R., The coming of man: was it accident or design?, London 1933.
Broom, R., Finding the missing link, London 1950.
Broom, R., Robinson, J. T., A new type of fossil man, in ‟Nature", 1949, CLXIV, pp. 322-323.
Broom, R., Robinson, J. T., Swartkrans ape-man Paranthropus crassidens, in ‟Transvaal museum memoirs", 1952, VI, pp. 1-123.
Butzer, K. W., Palaeo-ecology of South African australopithecines: Taung revisited, in ‟Current anthropology", 1974, XV, pp. 367-382.
Chagas, C. (a cura di), Pontifical Academy of Sciences working group on recent advances in the evolution of Primates. Summary, in Pontificiae Academiae Scientiarum scripta varia, vol. L, Città del Vaticano 1983, pp. 197-204.
Clark, W. E. Le Gros, The fossil evidence for human evolution, Chicago 1955, 19642, 19783.
Clarke, R. J., The cranium of the Swartkrans hominid SK 847 and its relevance to human origins, Johannesburg 1977.
Clarke, R. J., Howell, F. C., Affinities of the Swartkrans 847 hominid cranium, in ‟American journal of physical anthropology", 1972, XXXVII, pp. 319-336.
Connolly, C. J., External morphology of the Primate brain, Springfield 1950.
Dart, R. A., Australopithecus africanus: the man-ape of South Africa, in ‟Nature", 1925, CXV, pp. 196-199.
Dart, R. A., The osteodontokeratic culture of Australopitecus prometheus, in ‟Transvaal museum memoirs", 1957, X, pp. 1-105.
Dart, R. A., Craig, D., Adventures with the missing link, New York 1959.
Darwin, Ch., The origin of species by means of natural selection, London 18726 (tr. it.: L'origine delle specie, Torino 1959).
Darwin, Ch., The descent of man and selection in relation to sex, London 18752 (tr. it.: L'origine dell'uomo, Roma 1972).
Dubois, E., Pithecanthropus erectus, eine Menschenähnliche Übergangsform von Java, Batavia 1894.
Eldredge, N., Gould, S. J., Punctuated equilibria: an alternative to phyletic gradualism, in Models in paleobiology, San Francisco 1972, pp. 82-115.
Gould, S. J., Eldredge, N., Punctuated equilibria: the tempo and mode of evolution reconsidered, in ‟Paleobiology", 1977, III, pp. 115-151.
Gray, J. E., Outline of an attempt at the disposition of the Mammalia into tribes and families with a list of the genera apparently appertaining to each tribe, in ‟Annals of philosophy" (New Series), 1825, X, pp. 337-344.
Gray, J. E., Synopsis of the species of the class Mammalia, as arranged by Cuvier and other naturalist, in The animal kingdom, di G. Cuvier, vol. V, London 1827, p. 391.
Heberer, G. (a cura di), Menschliche Abstammungslehre: Fortschritte der Anthropogenie 1863-1964, Stuttgart 1965.
Howell, F. C., Hominidae, in Evolution of African Mammals (a cura di V. J. Maglio e H. B. S. Cooke), Cambridge, Mass., 1978, pp. 154-248.
Hughes, A. R., Tobis, Ph. V., A fossil skull probably of the genus Homo from Sterkfontein, Transvaal, in ‟Nature", 1977, CCLXV, p. 310-312.
Huxley, Th. H., Evidence as to man's place in nature and other essays, London 1854-1863 (tr. it.: Il posto dell'uomo nella natura e altri scritti, Milano 19612).
Johanson, D. C. e altri, Pliocene hominid fossils from Hadar, Ethiophia, in ‟American journal of physical anthropology", 1982, LVII, pp. 373, 719.
Johanson., D.C., Edey, M., Lucy: the beginning of human kind, London 1981 (tr. it.: Lucy, le origini dell'umanità, Milano 1981).
Leakey, L. S. B. e altri, Olduvai Gorge, vol. I, A preliminary report on the geology and fauna, 1951-1961, Cambridge 1965.
Leakey, L. S. B., Tobias, Ph. V., Napier, J. R., A new species of the genus Homo from Olduvai Gorge, in ‟Nature", 1964, CCII, pp. 7-9.
Leakey, M. D., Olduvai Gorge, vol. III, Excavations in bed I and II, 1960-1963, Cambridge 1972.
Leakey, M. D. e altri, Fossil hominids from the Laetoli beds, in ‟Nature", 1976, CCLXII, pp. 460-466.
Linnaeus, C., Systema naturae sive regna tria naturae systematice proposita per classes, ordines, genera et species, Leyden 1735.
Linnaeus, C., Systema naturae, animalia, Stockholm 175810.
Oakley, K. P., Revised dating of the Kanjera hominids, in ‟Journal of human evolution", 1974, III, pp. 257-258.
Oakley, K. P., Campbell, B. G., Catalogue of fossil hominids. I. Africa, London 1967.
Partridge, T. C., Geomorphological dating of cave opening at Makapansgat, Sterkfontein, Swartkrans and Taung, in ‟Nature", 1973, CCXLVI, pp. 75-79.
Robinson, J. T., The australopithecines and their bearing on the origin of man of stone tool-making, in ‟South African journal of science", 1961, LVII, pp. 3-16.
Salzano, F. M. (a cura di), The role of natural selection in human evolution, Amsterdam 1975.
Sergi, S., Il cranio del secondo paleantropo di Saccopastore, in ‟Paleontographia italica", 1948, XLII, pp. 25-164.
Sergi, S., Il cranio neandertliano del Monte Circeo (Circeo I), Roma 1974.
Simonetta, A., Catalogo e sinonimia annotata degli ominidi fossili ed attuali, 1758-1955, in ‟Atti della Società toscana di scienze naturali", 1957, pp. 53-112.
Simpson, G. G., A new classification of Mammals, in ‟Bulletin of the American Museum of Natural History", 1931, LIX, pp. 259-293.
Simpson, G. G., The principles of classification and classification of Mammals, in ‟Bulletin of the American Museum of Natural History", 1945, LXXXV, pp. 1-350.
Stringer, C. B., Population relationships of later Pleistocene hominids: a multivariate study of available crania, in ‟Journal of archeological science", 1974, I, pp. 317-342.
Tobias, Ph. V., Australopithecus. Homo habilis, tool-using and tool-making, in ‟South African archaeological bulletin", 1965, XX, pp. 167-192.
Tobias, Ph. V., Olduvai Gorge, vol. II, The cranium and maxillary dentition of Australopithecus (Zinjanthropus) boisei (a cura di L. S. B. Leakey), Cambridge 1967.
Tobias, Ph. V., The place of Australopithecus africanus in hominid evolution, in Recent advances in primatology (a cura di D. J. Chivers e K. A. Joysey), vol. III, London 1978, pp. 373-394.
Tobias, Ph. V., Hughes, A. R., The new Witwatersrand University excavation at Sterkfontein, in ‟South African archaeological bulletin", 1969, XXIV, pp. 158-169.
Wells, L. H., Homo sapiens afer Linn. - Content and earliest representatives, in ‟South African archaeological bulletin", 1969, XXIV, pp. 172-173.
Wells, L. H., Late stone age and middle stone age tool-makers, in ‟South African archaeological bulletin", 1972, XXVII, pp. 5-9.
Wu Rukang, Contribution to symposium on hominid evolution, Edmonton 1982.