
Visualizzare il funzionamento del cervello umano
Visualizzare il funzionamento del cervello umano
Alcune tecniche di recente sviluppo permettono di studiare le basi neuronali della funzione cognitiva nel cervello umano. Due di queste, la tomografia a emissione di positroni e la visualizzazione a risonanza magnetica funzionale, forniscono immagini del cervello durante la realizzazione di compiti specifici, che riffettono l'attività neuronale locale. Questo saggio prende in considerazione le potenzialità spaziali e temporali e i limiti di queste metodiche. Inoltre, esamina i problemi tecnici, biologici e cognitivi che riguardano la comprensione degli obiettivi e dei metodi degli studi di neurovisualizzazione. I progressi nella conoscenza della funzione cognitiva e della funzione cerebrale, resi possibili da questi metodi, sono illustrati con esempi tratti dalla vasta letteratura sulla neurovisualizzazione.
Introduzione
Storicamente lo studio della mente e lo studio del cervello sono stati a lungo separati. La psicologia si avvicinava alla mente come se fosse una scatola nera, un'entità il cui lavorio interno risultava nascosto, ma il cui funzionamento poteva essere esplorato esaminando le trasformazioni fra dati in entrata e dati in uscita e misurando i parametri caratteristici di quelle trasformazioni, come per esempio i tempi di reazione e la precisione nello svolgimento di un compito. Lo studio del cervello, viceversa, era il regno della biologia. Le tecniche classiche utilizzate per studiare l'anatomia e la fisiologia dei tessuti nervosi hanno svelato molti aspetti importanti della struttura e del funzionamento del cervello grazie all'analisi del sistema nervoso di topi, gatti, scimmie e perfino di lumache di mare. Le implicazioni di queste scoperte neurofisiologiche per il funzionamento del cervello umano dovevano sempre essere dedotte, in quanto la natura invasiva delle tecniche a disposizione impediva la loro diretta applicazione allo studio dell'attività cognitiva umana.
Così, per oltre un secolo, gli studi sul comportamento dei cerebrolesi e quelli anatomici del cervello umano post mortem sono stati i metodi neuroscientifici predominanti nell'approccio diretto al problema del funzionamento del cervello umano. Questa metodologia è cambiata radicalmente negli ultimi decenni poiché gli straordinari progressi teorici e tecnologici hanno favorito lo sviluppo di alcune metodiche non invasive in grado di investigare il funzionamento del cervello: le tecniche di neurovisualizzazione (neuroimaging). La vera forza di queste tecniche nel delucidare le più profonde attività del cervello in fase cognitiva si evidenzia soprattutto quando esse vengono combinate con appropriati approcci psicologici.
Poiché gli studi sulle rappresentazioni cerebrali appaiono sempre più frequentemente in letteratura, ponendo importanti problemi a ricercatori di vari campi, diventa sempre più importante avere familiarità con i metodi di neurovisualizzazione. Questo saggio tratta alcune questioni fondamentali per apprezzare le potenzialità di tali tecniche nel chiarimento delle basi neuronali del processo cognitivo e, inoltre, intende fornire materiale di riflessione a chi voglia valutare criticamente gli studi che stanno fiorendo in letteratura. l punti trattati comprendono una descrizione delle tecniche usate per la visualizzazione cerebrale, un'analisi a livello sistemico del funzionamento del cervello e uno schema per valutare la funzione cognitiva a un livello appropriato per gli studi di visualizzazione. Dopo una breve visione d'insieme delle più importanti tecniche di neurovisualizzazione attualmente in uso, verranno esaminate due tecniche che permettono di acquisire informazioni locali collegate alla corrispondente attività neuronale, cioè la tomo grafia a emissione di positroni (PET, Positron Emission Tomography) e la visualizzazione, o imaging, a risonanza magnetica funzionale (fMRl, functional Magnetic Resonance lmaging), e verranno discussi pregi e limiti tecnici e metodologici. Successivamente verranno delineate importanti questioni in grado di stimolare la riflessione sulle facoltà cognitive del cervello. Infine, dopo aver costruito una base per la comprensione delle tecniche di neurovisualizzazione, illustreremo, con esempi tratti dalla letteratura, il modo in cui questo tipo di studi ha contribuito alla nostra conoscenza delle basi neuronali dell' attività cognitiva.
Una rassegna delle varie modalità di neurovisualizzazione
La maggior parte delle tradizionali tecniche neurofisiologiche fornisce informazioni sul funzionamento del sistema nervoso a un livello cellulare o subcellulare. Quasi tutte queste tecniche producono una sorta di rappresentazione visiva del funzionamento neuronale, sia che si tratti di istogrammi delle scariche cellulari, sia di videomicroscopie al rallentatore di cellule in migrazione, oppure del tracciato oscilloscopico di un singolo potenziale d'azione. Tutte potrebbero dunque essere considerate tecniche di neurovisualizzazione. Piuttosto che discutere i confini di questo termine, per gli scopi di questo saggio noi definiamo tecniche di neurovisualizzazione quelle che forniscono informazioni sul funzionamento del sistema nervoso su larga scala, a livello sistemico.
Tutte le tecniche di neurovisualizzazione che saranno qui menzionate misurano, direttamente o indirettamente, una di queste due grandezze: i potenziali elettrici generati dal tessuto nervoso o le variazioni del flusso sanguigno cerebrale. Alcune tecniche, tra le quali l'elettroencefalogramma (EEG), il magnetoencefalogramma (MEG) e certi tipi di visualizzazione ottica, sono sensibili in modo diretto o indiretto alle variazioni dei potenziali elettrici nel tessuto cerebrale provocate dalla depolarizzazione e ripolarizzazione dei neuroni che costituiscono l'attività del cervello. Le altre tecniche, compresa la tomografia computerizzata a emissione di singolo fotone (SPECT, Single Photon Emission Computed Tomography), nonché certi tipi di visualizzazioni ottiche come la spettroscopia nel vicino infrarosso (NlRS, Near InfraRed Spectroscopy) o la PET e la fMRl, misurano, direttamente o indirettamente, le variazioni del flusso sanguigno che accompagnano e riflettono l'attività neuronale.
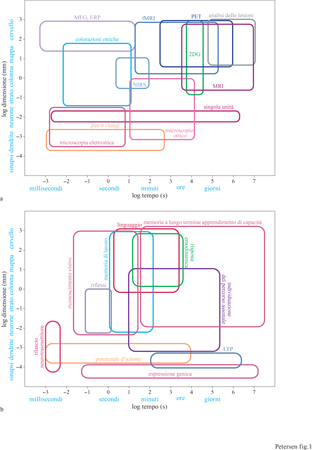
Ciascuna di queste tecniche costituisce un importante tassello per la sua capacità di fornire informazioni riguardo alle caratteristiche spaziali e temporali dell' attività neuronaIe nel cervello integro. La figura (fig. 1) illustra le scale spaziali e temporali alle quali sono sensibili varie tecniche di neurovisualizzazione, nel quadro dei metodi neurofisiologici standard utilizzati per studiare il funzionamento cerebrale. Una discussione dettagliata di ciascuna di queste tecniche esula dallo scopo di questo saggio, ma una breve rassegna sarà sufficiente a delineare il panorama della visualizzazione delle funzioni cerebrali.
Elettroencefalogramma e magnetoencefalogramma
L'elettroencefalogramma utilizza un apparato di elettrodi localizzati sul cranio per rilevare le variazioni dei campi elettrici generate dall'attività neuronale. Analogamente, il magnetoencefalogramma rileva le perturbazioni nel campo magnetico alla superficie del capo provocate dagli effetti induttivi delle variazioni dei campi elettrici che si verificano nel cervello. Entrambe queste tecniche possono essere usate insieme a manipolazioni cognitive allo scopo di studiare gli aspetti del processo cognitivo. l metodi basati sull'elettroencefalogramma, quando usati in tal modo, sono spesso citati come EP (Evoked Potentials, potenziali evocati) o come ERP (Event-Related Potentials, potenziali correlati a eventi). l segnali elettrici o magnetici rilevati da questi metodi forniscono un'immagine in tempo reale dell'attività cerebrale, in termini di tracciati dinamici dei segnali in vari punti del cranio. Tuttavia la localizzazione della fonte è problematica con questo tipo di dati: determinare la collocazione del tessuto neuronale attivo è un problema serio che può dar luogo a un numero pressoché infinito di soluzioni (questo problema è detto problema inverso). Allo scopo di localizzare le fonti bisogna aggiungere ulteriori limitazioni, e relative supposizioni. Così, mentre la risoluzione temporale di queste tecniche è di gran lunga superiore a quella degli altri metodi di visualizzazione del funzionamento del cervello, in molti casi si tratta di tecniche inefficaci per la localizzazione spaziale di segnali multipli. Nuovi e promettenti approcci utilizzano dati provenienti da altre modalità di visualizzazione funzionale per ridurre il problema inverso, specificando il numero di fonti in esame e la loro approssimativa collocazione (George et al., 1995). Tuttavia, a tutt'oggi, metodi basati su principi in grado di integrare queste modalità sono ancora in fase di sviluppo. Attualmente tutti gli sforzi per combinarli si basano su informazioni o supposizioni addizionali che riguardano il funzionamento del sistema nervoso.
Tecniche di visualizzazione ottica
Esistono numerose tecniche di visualizzazione ottica che utilizzano agenti di contrasto esogeni, come coloranti la cui fluorescenza dipende dalla differenza di potenziale, dalla concentrazione di calcio o da altre variabili regolate fisiologicamente. Allo stesso modo esistono tecniche che si basano su fattori endogeni, come la diffusione della luce in risposta al volume cellulare. Gran parte di questi metodi è invasiva o comunque inadatta agli studi sull'uomo, salvo in casi limitati, come in neurochirurgia. Tuttavia la NIRS, che misura attraverso il cranio lo spettro di assorbimento della luce da parte dell'emoglobina ossigenata, dell'emoglobina deossigenata e della citocromo c ossidasi, si dimostra una promettente tecnica di visualizzazione funzionale non invasiva. I principali limiti della tecnica, fino a questo momento, sono costituiti dal fatto che la luce non riesce a penetrare a fondo nei tessuti e dal fatto che il metodo misura risposte emodinamiche relative al flusso sanguigno in siti discreti piuttosto che in tutto il cervello.
La tomografia computerizzata a emissione di singolo fotone
La SPECT misura la distribuzione di un tracciante radioattivo. Le fotocamere SPECT ruotano intorno alla testa rivelando i singoli fotoni emessi dal tracciante in decadimento. Le immagini della distribuzione del tracciante, che riflettono la perfusione del sangue, vengono ricostruite con algoritmi derivati dalla tomo grafia computerizzata (CT, Computed Tomography). Per molti versi simile alla PET, questa tecnica è intrinsecamente di minore sensibilità. Per quanto le capacità spaziali e temporali della SPECT non siano all'altezza di quelle della PET, questa tecnica, grazie al suo basso costo e alla facile disponibilità, si è dimostrata piuttosto valida in sede clinica per studiare il funzionamento neuronale e il legame dei recettori.
La tomografia a emissione di positroni
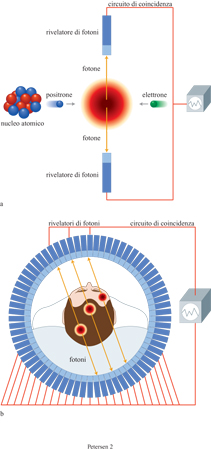
La PET localizza gli eventi di decadimento dei traccianti radioattivi somministrati al soggetto per endovena o tramite inalazione. Nel momento in cui i traccianti decadono, emettono positroni che si annichilano quando collidono con gli elettroni, le loro antiparticelle. Il processo di annichilazione genera fotoni che si muovono in direzioni opposte (fig. 3). I rivelatori PET, sistemati in anelli attorno allo scanner, rivelano questi fotoni. Soltanto le coppie di fotoni registrate da rivelatori opposti in un lasso di tempo molto breve sono considerate provenienti dallo stesso atto di disintegrazione e calcolate perciò come segnali validi. Poiché le coppie di fotoni viaggiano lungo la medesima retta, si considera che la fonte dei fotoni si trovi lungo la retta che passa per i due opposti rivelatori. Questo principio, detto rilevamento per coincidenza (v. figura 3b), fornisce un mezzo molto più efficace per localizzare la fonte dei segnali radioattivi rispetto al rilevamento a fotone singolo che avviene nel caso della SPECT.
Gli eventi di decadimento rivelati e registrati durante una scansione con agenti di contrasto sono molto spesso ricostruiti in un'immagine della distribuzione spaziale dei segnali tramite un algoritmo matematico basato su una retroproiezione filtrata, un algoritrno tomo grafico sviluppato per la prima volta per la CT.
La versatilità della PET come modalità di neurovisualizzazione è dovuta al fatto che si possono usare traccianti specifici per 'etichettare' molte variabili fisiologiche differenti. Tra i marcatori (marker) fisiologici collegati al funzionamento del cervello che la PET può misurare vi sono il metabolismo del glucosio, quello dell'ossigeno, il volume sanguigno, la distribuzione di diversi recettori neurochimici e il flusso sanguigno cerebrale. Le misurazioni di questi marcatori sono importanti per comprendere le basi fisiologiche di vari metodi di neurovisualizzazione, come pure per scopi clinici. Gli studi riguardanti prove di legame dei recettori si sono rivelati di valore incalcolabile in quanto hanno permesso di svelare alcuni 'segreti' farmacologici del cervello, compresa la distribuzione e l'affinità dei recettori e dei siti leganti il farmaco, e la perdita funzionale in una certa classe di recettori associata a sindromi cliniche specifiche. La tecnica più fruttuosa per l'esplorazione del funzionamento del cervello degli esseri umani in relazione all'aspetto cognitivo è quella che fa uso dei paradigmi di attivazione, grazie ai quali l'attività neuronale locale viene dedotta attraverso la misurazione del flusso sanguigno cerebrale a livello locale. Questa tecnica verrà discussa successivamente.
La visualizzazione a risonanza magnetica funzionale
Lo sviluppo della visualizzazione a risonanza magnetica (MRl, Magnetic Resonance Imaging) ha reso possibile visualizzare, in modo tridimensionale e non invasivo, la struttura interna di oggetti composti da materiali chimicamente diversi, con una risoluzione molto più alta rispetto a quella possibile con le tecniche a raggi X come la CT. Fino a poco tempo fa la risonanza magnetica era usata soprattutto in sede clinica allo scopo di visualizzare la struttura di patologie cerebrali come i tumori o gli edemi. Solo negli ultimi anni è divenuto evidente che il suo segnale è in grado di riflettere le variazioni emodinamiche provocate dall'attività neuronale.
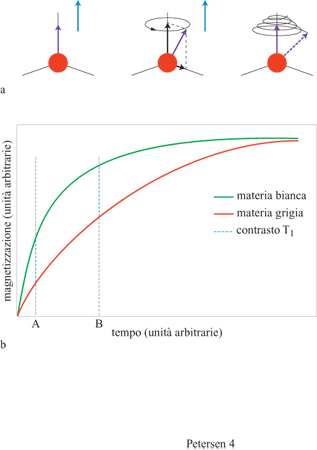
I principi basilari della risonanza magnetica. - Il segnale della risonanza magnetica (MR, Magnetic Resonance; originariamente risonanza magnetica nucleare o NMR, Nuclear Magnetic Resonance) deriva dalle caratteristiche intrinseche dei nuclei dotati di momenti di dipolo magnetico. Tali nuclei, se posti in un campo magnetico esterno, si allineano con esso (fig. 4). l nuclei si possono 'spostare' dall'equilibrio applicando un'altra forza, perpendicolare al campo esterno, un po' come quando si spinge un pendolo in quiete (vale a dire in equilibrio con la forza di gravità) provocandone l'oscillazione. l dipoli eccitati entrano in precessione intorno alla direzione del campo principale con una frequenza caratteristica del tipo di nucleo e proporzionale alla forza del campo, ritornando poi lentamente alloro stato di equilibrio, proprio come l'ampiezza del movimento di un pendolo si riduce progressivamente a zero. Questa precessione o oscillazione dei dipoli magnetici in un campo magnetico costituisce il segnale base dell'MR, il quale induce una corrente elettrica che viene amplificata e misurata.
Le costanti di tempo delle componenti parallela e perpendicolare relative al processo col quale il dipolo ritorna all'equilibrio (tempi di rilassamento) determinano l'intensità relativa del segnale MR in ogni dato momento. Il loro valore dipende dalla composizione del tessuto sottoposto ad analisi: la materia bianca, che presenta un'alta concentrazione di lipidi, presenta costanti di tempo diverse rispetto alla materia grigia. Con una scelta appropriata dei tempi di misurazione delle componenti parallela e perpendicolare e dei tempi di ripetizione del processo di misurazione, è possibile determinare la composizione della fonte del segnale in un dato punto dello spazio.
Quando i dipoli magnetici di specie simili in un campo omogeneo vengono eccitati con un impulso a radiofrequenza, inizialmente oscillano in fase l'uno con l'altro. Queste oscillazioni presto cominciano a uscire di fase a causa delle disomogeneità del campo magnetico, fino ad arrivare a un decadimento del segnale. Lo sfasamento può essere reversibile o irreversibile. Quello reversibile si può compensare invertendo le rotazioni mediante un impulso di riaggiustamento a 180⁰ per creare un'eco e in questo modo ripristinare l'intensità del segnale. Quello irreversibile è dovuto a variazioni microscopiche e casuali del campo magnetico. La costante di tempo con cui il segnale decade a causa di queste microscopiche disomogeneità costituisce un altro parametro caratteristico della composizione del tessuto. Questi parametri, in aggiunta alle componenti parallela e perpendicolare del dipolo durante la fase di rilassamento, sono importanti per la specificazione dell'intensità e della localizzazione di un segnale MR.
La visualizzazione MR Per formare un'immagine. - MR, è necessario che i segnali vengano identificati attraverso specifiche coordinate spaziali. Per fare ciò, le precedenti tecniche devono essere integrate con ulteriori accorgimenti che coinvolgono l'applicazione di gradienti di campo magnetico il cui effetto è di etichettare il segnale proveniente da ciascuna sede con lo stesso contrassegno. Il segnale complesso a variazione temporale che ne risulta viene ricostruito in immagine (segnale a frequenza spaziale) attraverso l'applicazione di una trasformata di Fourier che fa da traduttore fra i segnali nel dominio temporale e nel dominio spaziale. La sequenza dettagliata di impulsi trasmessi ai nuclei, che influenza in modo cruciale il segnale MR ed è essenziale per qualsiasi resoconto completo dell'MR, è troppo complessa per essere trattata qui (DeYoe et al., 1994). Il gran numero di parametri coinvolti in qualsiasi esperimento di visualizzazione MR e l'ampio spettro di valori che si possono scegliere per ciascuno rendono la tecnica molto versatile e stimolante.
Le basi fisiologiche di PET e fMRI: flusso sanguigno e attività neuronale
La capacità di PET e fMRl di fornire dati sull'attività cerebrale locale è dovuta a una fondamentale caratteristica fisiologica del cervello: l'emodinamica cerebrale locale riflette l'attività neuronale. Proprio come il flusso sanguigno nei muscoli aumenta quando essi vengono usati, il flusso sanguigno aumenta in regioni localizzate del cervello che sono coinvolte nell'attività di elaborazione. La relazione tra attività neuronale e flusso sanguigno venne per la prima volta ipotizzata nel 1890 da Ch. Roye Ch. S. Sherrington. Da allora sono state raccolte considerevoli prove a sostegno di quell 'ipotesi, dagli studi sulla perfusione dei cervelli animali sotto stimolazione alle osservazioni intraoperatorie sugli umani (Raichle, 1987). Più recentemente, esperimenti con PET e fMRI hanno confermato e raffinato ulteriormente le nostre conoscenze sulla relazione tra attività cerebrale e flusso sanguigno.
Il flusso sanguigno e il fabbisogno energetico dei neuroni attivi
I neuroni richiedono energia per una varietà di funzioni, comprese la biosintesi, il riassorbimento dei neurotrasmettitori e il mantenimento dei gradi enti ionici. Gran parte dell'energia è utilizzata per sostenere la pompa Na +/K+, piccoli motori molecolari di trasporto che ristabiliscono i gradienti ionici a riposo tra i neuroni e lo spazio extracellulare in seguito alla depolarizzazione. Il fabbisogno di energia della pompa Na+/K+, così come di altre funzioni cellulari, aumenta con l'aumentare dell' attività neuronale. La fonte energetica di questi processi cellulari è l'ATP, molecola sintetizzata nei mitocondri della cellula da glucosio e ossigeno. In assenza di una sintesi continua, le scorte di ATP cellulare si esauriscono rapidamente. Poiché le cellule non immagazzinano un surplus di substrati necessari alla sintesi di ATP, il corpo necessita di un mezzo che rifornisca le cellule dei substrati necessari per soddisfare il fabbisogno di energia.
Il sangue trasporta efficientemente i nutrienti in tutte le parti del corpo, e l'alto rapporto superficie/volume dei capillari consente un efficace trasferimento di materiali alle cellule e dalle cellule. Glucosio e ossigeno, i substrati metabolici per la sintesi di ATP, vengono trasferiti alle cellule attraverso le pareti dei capillari; mentre l'ossigeno diffonde nei tessuti attraverso le pareti dei vasi sanguigni, il glucosio e gli altri elementi nutritivi raggiungono i tessuti attraverso meccanismi di trasporto attivo. Oltre a distribuire questi importanti substrati, il sangue trasporta anche i prodotti metabolici di rifiuto, come il biossido di carbonio. L'anatomia del cervello e il sistema vascolare sono ben strutturati per questo tipo di distribuzione: il neuropilo, dove si trovano gran parte delle sinapsi, presenta la più alta densità capillare, ed è in questa regione che si verifica il più grande incremento del flusso sanguigno durante l'attività neuronale.
I meccanismi esatti attraverso cui viene mediato l'aumento del flusso sanguigno con l'attività neuronale sono sconosciuti. L'aumento del flusso nei capillari potrebbe risultare sia dal reclutamento o dalla dilatazione dei capillari, sia da un aumento della velocità del sangue nei vasi. È stata avanzata l'ipotesi che diversi fattori prodotti durante l'attività neuronale, come ossido di azoto (NO), adenosina e CO2, regolino il flusso, ma la natura precisa della cascata di segnali non è ancora chiara.
Correlazione tra flusso sanguigno e metabolismo: evidenze sperimentali e questioni aperte
Le misurazioni del flusso sanguigno locale a riposo mostrano una forte correlazione tra questo e il metabolismo. La relazione tra flusso sanguigno e metabolismo è però più complicata durante l'attivazione transitoria. P.T. Fox e M.E. Raichle (1984) hanno per primi evidenziato risposte che modificano il flusso sanguigno nella corteccia visiva in presenza di uno stimolo visivo lampeggiante. Quando la frequenza del lampeggiamento veniva progressivamente portata da 2 a 32 Hz, il flusso sanguigno aumentava linearmente da 2 a circa 8 Hz, dopo di che si stabilizzava e declinava leggermente intorno a 32 Hz. La curva della risposta del flusso sanguigno è simile alla curva che descrive le risposte elettriche nella corteccia visiva di fronte alle variazioni della frequenza di lampeggiamento. A partire da questo primo studio PET sull'attivazione, innumerevoli altri esperimenti hanno evidenziato variazioni del flusso sanguigno in risposta a un'attivazione corticale. È dunque ormai accertato che l'attivazione neuronale locale è accompagnata da variazioni locali nel flusso sanguigno cerebrale. In origine si pensava che tali variazioni provocassero un aumento della distribuzione dei substrati metabolici in grado di soddisfare i livelli crescenti del tasso metabolico cerebrale di consumo di glucosio e del tasso metabolico cerebrale di consumo di ossigeno; si pensava cioè che flusso e metabolismo fossero associati a tutti i livelli metabolici. Gli studi PET con fluorodesossiglucosio, composto che consente di quantificare il metabolismo del glucosio, rivelano un aumento del consumo locale di glucosio in presenza di attività neuronale, come nelle aspettative. Gli aumenti del consumo di ossigeno, invece, non corrispondono agli aumenti del metabolismo del glucosio e nel flusso sanguigno. Anche grandi aumenti del flusso sanguigno a livello locale, dell'ordine del 29%, sono accompagnati da modesti aumenti del metabolismo dell'ossigeno, pari a circa il 5% (Fox e Raichle, 1986). La mancata corrispondenza tra gli aumenti del flusso sanguigno e quelli del metabolismo dell'ossigeno indica una diminuzione significativa della frazione di ossigeno estratta a livello locale, vale a dire della quantità relativa di ossigeno estratta dal sangue.
Numerosi studi successivi hanno confermato che non vi è corrispondenza tra il metabolismo ossidativo e il flusso sanguigno durante alti livelli di attività neuronale. Il significato biologico della disparità tra flusso sanguigno e tassi di consumo di ossigeno resta da definire. Alcuni modelli postulano aumentati livelli di attività anaerobica nel corso dell'attività neuronale. Altri suggeriscono che il metabolismo è fondamentalmente aerobico, e che il flusso potrebbe aumentare in modo sproporzionato rispetto al consumo di ossigeno durante la stimolazione neuronale allo scopo di fronteggiare moderati aumenti nel fabbisogno di O2.
Come verrà spiegato in modo più esauriente, questa disparità tra flusso sanguigno cerebrale e metabolismo dell'ossigeno è la base su cui poggia l'MR funzionale. Un aumento del flusso sanguigno a valori di gran lunga superiori a quello necessario per fornire l'ossigeno richiesto per il metabolismo ha come effetto un aumento locale nella concentrazione di emoglobina ossigenata e una relativa diminuzione nella concentrazione della desossiemoglobina. Una variazione nei rapporti tra ossi- e desossiemoglobina determina un cambiamento del segnale MR.
Malgrado le prove incontrovertibili della covariazione di flusso sanguigno e attività neuronale, deve essere ancora compreso molto sulla natura di questa relazione, sui suoi aspetti quantitativi e sui meccanismi fisiologici che stabiliscono il legame. Tante sono le questioni ancora aperte: l'attività neuronale determina un aumento del flusso sanguigno in tutte le regioni del cervello o alcune sono emodinamicamente 'silenti' durante l'attivazione? Qual è l'effetto emodinamico dell'inibizione neuronale? l tempi emodinamici sono simili in tutto il cervello? In quale misura l'area in cui il flusso sanguigno aumenta coincide, dal punto di vista spaziale, con quella dei neuroni attivi? Le risposte a queste e ad altre domande a esse collegate influenzeranno la nostra interpretazione degli studi di neurovisualizzazione con PET e fMRI.
Studi sull'attivazione
Finora abbiamo delineato le basi delle tecniche di neurovisualizzazione funzionale, così come i tipi di segnale che esse rilevano e i rapporti tra questi segnali e l'attività neuronale. Uno degli scopi primari della neurovisualizzazione è attribuire ruoli funzionali a specifiche regioni del cervello nelle quali sono stati individuati i cambiamenti. Attraverso le correlazioni tra attività neuronale e processi cognitivi siamo in grado di migliorare la nostra capacità di determinare le componenti costitutive dei processi cognitivi e, insieme, di aumentare la nostra conoscenza dell'organizzazione cerebrale. Gli studi sull'attivazione, paradigmi per l'applicazione di queste tecniche alla risoluzione di tali problemi, sfruttano il fatto che la risposta emodinamica rilevata con PET e fMRI riflette livelli sotto stanti di attività neuronale. In alcuni casi gli studi sull'attivazione delineano l'anatomia funzionaIe di un compito cognitivo. In altri casi cercano di determinare più in dettaglio il tipo di computo che una regione svolge. Nei paradigmi di attivazione le risposte emodinamiche vengono misurate tra due o più compiti assegnati, nel corso di una serie di scansioni condotte in una singola seduta di sperimentazione. Si ritiene che le differenze a livello regionale tra segnali misurati durante vari compiti riflettano differenze nel grado di attività neuronale locale associata allo svolgimento di quei compiti.
Gli studi PET sull'attivazione
Gli studi PET sull'attivazione usano molto spesso traccianti che incorporano l'isotopo dell'ossigeno ¹⁵O, dotato di un'emivita di 122 s, per misurare le variazioni del flusso sanguigno che si verificano durante lo svolgimento di determinati compiti. La breve emivita permette la registrazione di un numero sufficiente di eventi di decadimento in un periodo limitato, normalmente tra 40 e 60 s. Le caratteristiche di ¹⁵O rendono tale isotopo molto adatto agli studi sull'attivazione. Il tracciante attualmente più usato è acqua marcata con ¹⁵O.
Uno studio PET sull'attivazione normalmente comprende 10÷12 scansioni per soggetto. Questo limite è dovuto alla quantità totale di radiazioni che è possibile somministrare a un soggetto e a considerazioni di natura pratica, legate alla durata della seduta. Allo scopo di ottenere dati chiari, le scansioni sono in genere separate da intervalli di 10÷12 m (5÷6 emivite) in modo da permettere alla radiazione di fondo (noise) di decadere tra una scansione e l'altra. Poiché il movimento nel corso della seduta esplorativa compromette la qualità dei dati, e poiché per il soggetto è difficile restare immobile per lunghi periodi, una tipica seduta raramente dura più di qualche ora. Di conseguenza, può essere realizzato soltanto un numero limitato di scansioni su ciascun soggetto.
Nei circa dieci anni trascorsi dall'inizio degli studi PET sull'attivazione, si è sviluppata una vasta letteratura riguardante un'ampia varietà di aspetti della cognizione, troppo vasta e varia per poterla trattare in un solo saggio. Considerevoli progressi sono stati compiuti nei campi della vista, dell'attenzione, del linguaggio, dell'apprendimento motorio e della memoria, per nominarne solo alcuni. Fino al recente sviluppo della fMRI, la PET non aveva rivali per la capacità di fornire informazioni circa l'organizzazione funzionale del cervello umano su larga scala.
Gli studi fMRI sull'attivazione
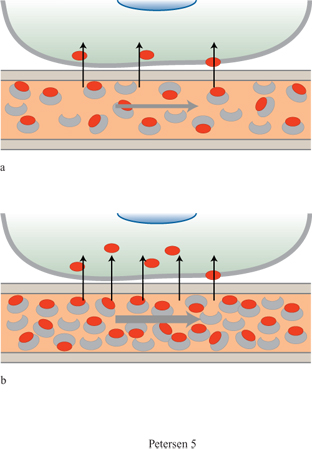
A partire dalle prime dimostrazioni delle potenzialità dell'MR come tecnica per misurare l'attività cerebrale, sono stati sviluppati diversi tipi di sequenze di impulsi sensibili ai parametri correlati al funzionamento del cervello, come la perfusione, il volume sanguigno e l'ossigenazione del sangue. La tecnica BOLD (Blood Oxygenation Level Dependent, dipendente dal livello di ossigenazione del sangue), più frequentemente usata, sfrutta le differenti proprietà magnetiche dell'emoglobina ossigenata e deossigenata e il fatto che le proporzioni relative di queste ultime nel sangue cambiano in seguito all'aumento dell'attività neuronale (fig. 5). Il sangue deossigenato, paramagnetico, genera campi magnetici locali che interrompono l'omogeneità del campo applicato esternamente. Queste disomogeneità provocano uno sfasamento irreversibile degli spin nucleari nel tessuto circostante e una conseguente perdita di segnale da quel tessuto. Quindi, l'aumento della concentrazione di desossiemoglobina determina una perdita del segnale locale fMRI. L'entità dell'aumento di desossiemoglobina dovuto all'attività neuronale dipende dall'aumento di livello del flusso ematico cerebrale regionale e dalla variazione della frazione di ossigeno legato. Poiché con l'attivazione il consumo di O₂ aumenta meno del flusso sanguigno, l'aumento di ossiemoglobina dovuto all'incremento del flusso sanguigno supera di gran lunga la diminuzione causata dalla conversione dell'emoglobina nella forma deossigenata, dovuta al consumo di O₂. Questo porta a un aumento netto nella concentrazione relativa di ossiemoglobina e, di conseguenza, a un aumento del segnale BOLD. Così, forse sorprendentemente, ciò che permette di misurare l'attività cerebrale con l'MR è un eccesso di emoglobina ossigenata nel corso dell'attività neuronale (v. figura 5).
Il rendimento delle tecniche di neurovisualizzazione
Lo sviluppo di PET e fMRl ha dato inizio a una nuova era negli studi sul funzionamento del cervello. Queste efficientissime tecniche hanno suscitato di recente molta attenzione da parte sia di sostenitori entusiasti sia di critici nettamente sfavorevoli a tali applicazioni. Malgrado l'interesse dei media per i metodi di neurovisualizzazione abbia catturato l'immaginazione dell'opinione pubblica, le idee correnti su quanto questi esperimenti sono in grado di cogliere, sulle loro potenzialità e sui loro limiti sono spesso approssimative. Per comprendere le potenzialità di queste tecniche nello svelare la base neuronale dei processi cognitivi è fondamentale capire con quale precisione esse discernano le variazioni del segnale corrispondenti ai cambiamenti negli schemi spaziali o temporali dell'attività neuronale e con quale accuratezza localizzino questi cambiamenti nel cervello.
Segnale e rumore
Il rapporto tra segnale e rumore (SNR, Signal to Noise Ratio) rappresenta il principale limite delle metodologie di neurovisualizzazione. Il vero segnale che riflette le variabili fisiologiche di interesse è parzialmente coperto dal rumore proveniente da varie fonti, tra le quali lo strumento di misurazione, il rumore di fondo e altri processi fisiologici. Le tecniche di neurovisualizzazione possono rivelare qualcosa di importante sul funzionamento del cervello solo quando il segnale è statisticamente separabile dal rumore di fondo; gran parte della sfida e dei limiti che riguardano la neurovisualizzazione dipende dalla necessità di ottenere un SNR accettabile.
Caratteristiche spaziali
Tra i problemi importanti che possono essere affrontati con le tecniche di neurovisualizzazione vi è quello di capire se cambiamenti accertati nell' attività neuronale siano associati a compiti specifici e dove avvengono questi cambiamenti. Le potenzialità e i limiti di queste tecniche stanno nella possibilità di effettuare determinate misure. Ci si potrebbe chiedere, per esempio, quanto vicine possono essere due regioni attive per continuare a distinguerle l'una dall'altra, oppure con quale precisione le attivazioni possono essere individuate nel cervello. Ci si potrebbe chiedere, inoltre, fino a che punto l'intensità e l'estensione del segnale siano in rapporto con l'intensità e l'estensione della sottostante attività neuronale. Questi problemi saranno trattati qui di seguito. In definitiva, il problema principale per gli studi sull' attivazione è capire in che misura la tecnica di neurovisualizzazione rifletta il segnale biologico sottostante. Quando la PET e la fMRl vengono considerate nel contesto della natura dei segnali biologici che esse riflettono, ciascuna ha i suoi vantaggi e i suoi svantaggi; le loro potenzialità spaziali effettive sono paragonabili tra loro più di quanto la loro specificità tecnica possa far pensare.
Risoluzione
Molti ritengono che la risoluzione specifichi le potenzialità spaziali di una tecnica. Il termine risoluzione ha, tuttavia, un preciso significato tecnico: si riferisce a quanto distanti (nello spazio per la risoluzione spaziale, nel tempo per quella temporale) debbano essere due segnali per poter essere distinti come entità separate da una singola misurazione. La risoluzione di una tecnica è influenzata da diversi fattori, che comprendono la strumentazione, la natura del segnale misurato e il rumore tipico di quella tecnologia. La risoluzione si può determinare con test empirici o con l'applicazione di teorie a vari livelli di complessità.
Localizzazione
Per gli studi che si prefiggono di costruire una mappa del cervello, il problema di individuare dove avviene un'attivazione è di primaria importanza. La localizzazione è correlata alla capacità di una tecnica di determinare l'origine spaziale (in questo caso, nel cervello) di un singolo segnale. È un parametro diverso dalla risoluzione e, nei metodi di neurovisualizzazione, l'accuratezza della localizzazione può superare la risoluzione. Oltre all'accuratezza della localizzazione, spesso interessa sapere quanto sia affidabile la localizzazione nel corso di ripetute misurazioni.
Caratteristiche spaziali della PET
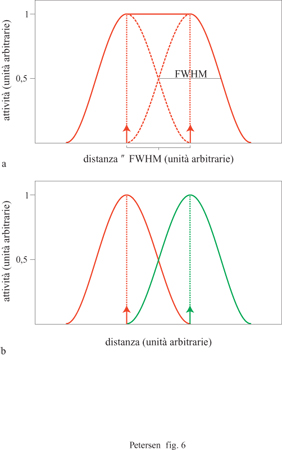
La definizione operativa della risoluzione spaziale della PET corrisponde alla larghezza totale a metà del valore massimo di intensità, o larghezza a metà altezza (FWHM, Full Width Half Maximum) di un'immagine derivata da un punto fonte di radiazione (fig. 6). La struttura dello strumento di misurazione e la fonte del segnale pongono limiti alla risoluzione delle tecniche di neurovisualizzazione: nella PET la risoluzione spaziale intrinseca dipende dall'ampiezza e dalla spaziatura degli elementi rivelatori, dall'energia dell' emissione e dal grado di diffusione dei fotoni, parametri che contribuiscono a offuscare l'immagine acquisita. La risoluzione intrinseca per gli apparecchi PET attualmente disponibili in commercio, come quelli che usano traccianti 15O, è approssimativamente di 4÷5 mm. Strumenti a più alta risoluzione sono in via di sviluppo.
L'SNR della PET dipende dal raggio dello scandaglio dello scanner e dalla sensibilità del rivelatore. Allo scopo di ridurre al minimo l'effetto del rumore ad alta frequenza spaziale, le immagini PET vengono normalmente trattate con un filtro a bassa frequenza prima di ulteriori analisi. La frequenza del filtro, associata con la risoluzione intrinseca dello scanner, determina la risoluzione finale dell'immagine che, per gran parte degli studi PET sull'attivazione, è tra i 10 e i 16 mm. La conseguenza pratica della necessità di avere un SNR accettabile è che nella PET le misure sono mediate su diverse sequenze o soggetti, il che riduce il segnale non sistematico e preserva quello sistematico. Quando i dati sono mediati tra diversi soggetti, la risoluzione spaziale dei dati risultanti è in qualche misura ridotta a causa delle variabilità funzionali e anatomiche.
La localizzazione della fonte di un segnale non è strettamente limitata dalla risoluzione dell'immagine. Infatti, il calcolo del 'centro di massa' di un'attivazione fornisce un'informazione sulla posizione molto più precisa di quanto la risoluzione dell'immagine permetterebbe (Fox et al., 1986). Questo metodo, associato con un'adeguata progettazione dell'esperimento, può aumentare notevolmente le potenzialità spaziali effettive della PET al di là della risoluzione della singola immagine. Sebbene due foci di attività nella stessa immagine si possano distinguere come attivazioni separate quando a separarle è almeno l FWHM, la capacità di distinguere due attivazioni molto vicine nello spazio e presenti in scansioni PET separate non è limitata allo stesso modo. l calcoli sul 'centro di massa' per determinare la collocazione di regioni attivate, individuate da scansioni distinte, possono rivelare spostamenti delle regioni attivate sulla scala di pochi millimetri.
Oltre ai fattori tecnici, le potenzialità spaziali della PET sono limitate da fattori pratici come la regolazione delle radiazioni e da considerazioni di tempo. Poiché le potenzialità spaziali della PET hanno una 'grana' meno fine della scala spaziale della risposta emodinamica, possono esserci limiti nella capacità di riflettere in modo attendibile differenze di piccola scala negli schemi dell'attività neuronale. D'altro canto le immagini PET mediante ¹⁵O mostrano una relazione quantitativa nota con il flusso sanguigno, che è a sua volta correlato con l'attività neuronale, e dunque presentano un importante vantaggio sulla fMRl nell'interpretazione dei risultati.
Caratteristiche spaziali della fMRI
In teoria non c'è un limite inferiore alle capacità della fMRl di risolvere l'informazione spaziale, poiché con l'aumento dell'intensità dei gradienti di campo magnetico e del tempo di visualizzazione si possono distinguere strutture sempre più piccole. Tuttavia, i magneti adatti per studiare il cervello umano normalmente producono campi di intensità fino a 4 tesla (più spesso 1,5 tesla) e, per mantenere la durata della scansione entro un lasso di tempo ragionevole, le immagini del cervello umano vengono raramente spinte al di sotto della soglia del millimetro. Di solito, quando si riportano i dati fMRl, si registra la dimensione lineare del voxel di acquisizione (il volume di tessuto su cui viene di volta in volta focalizzata la sorgente); questa è una buona approssimazione della risoluzione spaziale dell'immagine.
L'SNR della fMRl è proporzionale al segnale intrinseco (comprendente l'intensità del campo), al volume del voxel e alla radice quadrata del tempo di visualizzazione. Di conseguenza vi è un nesso diretto tra la dimensione lineare del voxel, cioè la risoluzione spaziale, e il tempo di visualizzazione. Ridurre la dimensione lineare del voxel alla metà (vale a dire il voxel di un fattore 8) richiede un aumento di 64 volte del tempo di visualizzazione per mantenere lo stesso SNR. Poiché la modulazione dell'intensità del segnale BOLD durante l'attivazione funzionale è relativamente piccola, solitamente dal 2 al 5% al di sopra del segnale in stato di quiete, 'l'arte' dell'MR consiste nel massimizzare il segnale rispetto al rumore. Le dimensioni del voxel scelte negli esperimenti fMRl riflettono la necessità di questo bilanciamento e non la risoluzione spaziale della tecnica in sé. Per migliorare l'SNR, i dati vengono spesso mediati su diverse sequenze condotte su uno stesso soggetto. Occasionalmente i dati vengono mediati su diversi soggetti, come nella PET, per aumentare ulteriormente l'SNR, ma di nuovo, a causa della variabilità anatomica e funzionale tra i soggetti, ciò avviene a scapito della risoluzione spaziale dell'immagine finale.
L'accuratezza di localizzazione della fMRl resta una questione aperta; in qualche misura essa dipende dalla sequenza di impulsi utilizzata: alcune sequenze vengono concepite per evidenziare le vene di deflusso, le quali possono trovarsi ad alcuni millimetri dal sito dell'attività neuronale che provoca i cambiamenti di ossigenazione, altre sono studiate per mirare ai letti capillari del tessuto attivo in modo più specifico. Inoltre, una scansione con una piccola dimensione del voxel dirige la tecnica verso l'identificazione di piccole regioni con grande variazione di ossigenazione, che di nuovo possono corrispondere alle localizzazioni delle vene di deflusso, non del neuropilo. Queste misurazioni potrebbero fornire un'idea fuorviante sulla localizzazione e sull'intensità dell'attività neuronale sotto stante. Per tutti i motivi qui sopra discussi, i voxel nella gran parte degli studi fMRl misurano almeno alcuni millimetri di lato.
Altre implicazioni biologiche possono influenzare in modo significativo i limiti delle potenzialità spaziali effettive della fMRl. Per esempio, la fMRl è molto sensibile ai moti biologici, come la respirazione e la pulsazione dei tessuti collegati al cuore, che possono creare seri artefatti nel segnale. Il problema più importante è che ancora non si comprende appieno la natura delle componenti fisiologiche che contribuiscono al segnale MR. È chiaro, tuttavia, che le potenzialità spaziali della fMRl approssimano o superano la scala della risposta che misurano. Spingere la risoluzione spaziale molto al di là della scala spaziale del segnale da misurare porta scarsi vantaggi e qualche svantaggio; perciò il problema di ciò che viene misurato dalla fMRI diviene di fondamentale importanza. Quasi certamente la potenzialità spaziale effettiva della fMRl è, al suo meglio, nella scala di specificità della risposta emodinamica, vale a dire a livello dei millimetri (Malonek e Grinvald, 1996; Woolsey et al., 1996) e non al livello dei singoli neuroni.
Intensità ed estensione spaziale
Oltre a sapere dove ha origine un segnale (localizzazione) e se proviene da una singola fonte (risoluzione) è interessante conoscere la forza della risposta biologica sotto stante e quanto tessuto cerebrale dà origine al segnale. È stato dimostrato, almeno in alcuni casi, che l'intensità del segnale PET e fMRI varia con l'intensità dello stimolo o con la variazione di altri parametri che influenzano l'attività neuronale (Fox e Raichle, 1984; Kwong et al., 1992). Alcuni studi suggeriscono, tuttavia, che la PET possieda un intervallo dinamico maggiore rispetto alla fMRI. Quindi la PET sarebbe una tecnica più utile per stabilire il grado di attività neuronale dall' intensità della risposta. La situazione si ribalta, però, quando si tratta di determinare l'estensione spaziale dell'attivazione. Poiché l' ampiezza della risposta misurata nella PET è collegata alla sua intensità, la PET non è adatta a determinare a livello millimetrico l'estensione spaziale della risposta biologica sottostante. Studi recenti suggeriscono che la fMRl rifletta con maggior precisione l'estensione spaziale del segnale rispetto alla PET.
Caratteristiche temporali
La caratterizzazione delle potenzialità temporali dei metodi di neurovisualizzazione non è meno complessa della loro caratterizzazione spaziale. Né nella PET né nella fMRl la strumentazione in sé è un fattore che limita la risoluzione temporale. Tuttavia in nessuna delle due modalità di visualizzazione l'informazione portata dalle misurazioni in brevi intervalli temporali riflette il funzionamento del cervello in tempo reale (millesimi di secondo). La PET fornisce utili informazioni nel corso di una singola scansione, in genere dai 40 ai 60 s per misurare l'attivazione con ¹⁵O. La risoluzione temporale della fMRl può essere manipolata modificando il protocollo di sperimentazione, ma per scopi pratici oscilla tra un secondo e alcune decine di secondi. Come avviene con la risoluzione spaziale, la capacità dei metodi di neurovisualizzazione di risolvere gli eventi nella dimensione temporale dipende da numerosi fattori, collegati alla strumentazione, all'SNR, e ai fattori biologici.
Caratteristiche temporali della PET
La PET è spesso criticata per la sua bassa risoluzione temporale, ma questa non è la conseguenza di una strumentazione poco sensibile. l macchinari PET sono estremamente sensibili alle differenze temporali, nell'ordine dei nanosecondi per determinare le coincidenze nel rilevamento fotonico. Le ragioni della bassa risoluzione temporale delle scansioni PET sull'attivazione sono di due tipi: sebbene l'informazione sul decadimento dei positroni sia raccolta in tempo reale, essa è di tipo probabilistico, e quindi è necessario accumulare un gran numero di coincidenze perché possa riflettere con precisione lo schema del flusso sanguigno; la nostra capacità di interpretare i dati dipende dai modelli cinetici del tracciante nel flusso sanguigno e dalla sua distribuzione. L'accumulo di dati nel corso di decine di secondi è organizzato per produrre stime quantitative del flusso sanguigno, ed effettuare misurazioni per periodi molto più lunghi o più corti del tempo di rilascio iniziale del tracciante riduce l'interpretabilità della scansione.
Poiché la PET utilizza in genere compiti impegnativi e media su periodi di 40÷60 s, essa non è seriamente influenzata dalla nostra mancanza di comprensione delle dinamiche temporali della risposta emodinamica sotto stante nella scala dei secondi. Tuttavia, la sua risoluzione temporale produce altre complicazioni che influenzano l'interpretazione dei dati. Normalmente i compiti vengono ripetuti più volte nel corso di una scansione, ma durante quel lasso di tempo possono verificarsi numerosi processi cognitivi, previsti e imprevisti. Quindi, una scansione PET fornisce una misura del flusso sanguigno cerebrale a livello regionale, integrato lungo l'intera durata della misurazione, che può quindi comprendere molti processi diversi da quelli che il ricercatore è interessato a esaminare o pensa di esaminare. Si tratta di un aspetto da tenere in considerazione quando si interpretano i dati PET.
Caratteristiche temporali della fMRI
Con l'invenzione della fMRl ecoplanare (EPl, EcoPlanarImaging) e il contemporaneo sviluppo della strumentazione che consente una variazione rapidissima dei gradi enti di campo magnetico, una singola immagine MR può essere acquisita in meno di 100 ms e i dati sull'intero cervello in 1÷2 s. Tuttavia la qualità del rapporto tra il segnale e il rumore di una singola immagine è troppo scadente perché una singola scansione EPI sia di per se stessa utile. l dati fMRl a singolo schema sono difficili da interpretare così come i dati raccolti su brevi porzioni di tempo con la PET. Di norma, i segnali fMRl vengono mediati su intervalli di tempo che vanno dai 10 s ai 30 s. Di conseguenza i dati fMRI sono soggetti agli stessi problemi interpretativi dei dati PET.
Sebbene i dettagli delle caratteristiche temporali della risposta emodinamica non siano ancora chiari, si sa che una variazione rilevabile del flusso sanguigno si prolunga per diversi secondi oltre l'attività neuronale che l'ha generato, e che sono necessari alcuni secondi affrnché il segnale raggiunga il massimo e poi decada. Quindi, ancora una volta, sebbene la fMRl possa misurare variazioni di flusso sanguigno quasi in tempo reale, ciò che essa registra corrisponde a un'attività neuronale svoltasi diversi secondi prima. Le caratteristiche temporali generali del ritardo emodinamico possono essere tenute in considerazione mediante alcuni metodi di analisi, ma i tempi di ristagno possono differire da una regione corticale all'altra e le costanti di tempo per la generazione e la caduta del segnale possono variare a seconda dei diversi tipi di stimolo. In ogni caso, i dati attuali suggeriscono che la determinazione del momento iniziale e della durata della risposta emodinamica misurata in una data area in seguito a un particolare stimolo è piuttosto attendibile; ciò consente di trarre alcune conclusioni sulle proprietà temporali delle risposte misurate.
La capacità della fMRI di acquisire dati in brevi periodi di tempo offre importanti vantaggi. Poiché la fMRl non è invasiva e non utilizza sostanze radioattive, in teoria non vi è alcun limite al numero di volte in cui un soggetto può essere sottoposto alle scansioni. Ciò offre un sostanziale vantaggio sulla PET per quanto riguarda l' ottenimento di un buon rapporto tra segnale e rumore, e consente la progettazione di nuovi esperimenti che mirano agli aspetti temporali dell'emodinamica. Le scansioni fMRI, per esempio, possono essere archiviate post hoc e analizzate in gruppi separati, rendendo possibili esperimenti in cui diversi tipi di prove siano usati contemporaneamente, diversamente dagli schemi bloccati della PET (Buckner et al., 1996). Le metodiche a prove miste sono spesso più efficaci per esplorare la funzione cognitiva.
Gli studi sulla visualizzazione ottica (Malonek e Grinvald, 1996) suggeriscono che un iniziale decremento dell' emoglobina ossigenata precede un aumento del flusso sanguigno, ed è più strettamente associato alle caratteristiche spaziali e temporali dell'attività neuronale di quanto non sia il successivo aumento di flusso sanguigno. Un rapido decremento iniziale nel segnale MR (circa 500 ms) è stato rilevato sia con la modalità spettroscopica sia con quella di visualizzazione (Menon et al., 1995), aprendo la possibilità di registrare questi sottili cambiamenti emodinamici. Se segnali di questo tipo potessero essere rilevati in modo affidabile, la capacità della fMRI di riflettere le dinamiche temporali e la localizzazione spaziale dell' attività neuronale sarebbe notevolmente incrementata. Comunque, dati interessanti e convincenti riguardanti la rilevabilità e la costanza di questo effetto devono essere ancora presentati.
Una visione sistemica del cervello
Oltre a comprendere le potenzialità e i limiti delle tecniche sperimentali usate per studiare le basi cerebrali della funzione cognitiva, è anche necessario avere un'idea del cervello in se stesso, a un livello sistemico, decisivo per capire le prospettive della neurovisualizzazione. Avendo la sola conoscenza del cervello a livello cellulare, per esempio, di contro a una prospettiva sistemica, si potrebbe accettare l'idea errata che la risoluzione dei metodi di neurovisualizzazione sia insufficiente per osservarne il funzionamento. Il ragionamento è il seguente: la risoluzione spaziale e temporale di PET e fMRl è nell'ordine dei millimetri e dei secondi. Ma la cognizione è mediata dai neuroni, la cui scala spaziale è nell'ordine dei micron e la cui attività si misura in millesimi di secondo. Anche un compito cognitivo complesso, come riconoscere un volto o costruire una frase, si svolge nell'ordine delle centinaia di millesimi di secondo. Se i metodi di neurovisualizzazione non possono risolvere dimensioni spaziali e temporali di queste scale, come potranno fornirci informazioni interessanti sulle basi cerebrali della cognizione? l limiti di questa considerazione diventeranno evidenti con un'adeguata comprensione della base neuronale della cognizione a livello sistemico.
Il cervello e le unità funzionali localizzate
Il cervello dei mammiferi è un organo fortemente strutturato, con numerose regioni distinte che si possono identificare dal punto di vista anatomico, fisiologico e funzionale. La suddivisione della corteccia in base alle differenze anatomiche nella microarchitettura corticale ha anticipato lo sviluppo dei metodi fisiologici per esplorare il cervello. Nel corso degli ultimi quarant'anni gli studi sulle proprietà fisiologiche delle cellule nella corteccia cerebrale dei mammiferi hanno dimostrato che regioni corticali diverse sovrintendono all'elaborazione di tipi di informazione diversi, come il movimento, il colore, la forma e la collocazione degli stimoli visivi. È possibile dimostrare che queste distinzioni che riguardano la funzione si accordano in molti casi con confini strutturali ben identificati. Il cervello è dunque un organo formato da molteplici sotto unità funzionali collegate tra loro, ed è questo principio di localizzazione a rendere possibile l'uso delle tecniche di neurovisualizzazione per lo studio dei processi cognitivi. A partire dalla metà degli anni Ottanta, gli studi PET e fMRI sugli esseri umani hanno dimostrato in modo convincente che la corteccia umana, come quella dei primati non umani, è suddivisa in aree funzionali specifiche (Fox et al., 1986; Zeki et al., 1991; Sereno et al., 1995; Tootell et al., 1995; De Yoe et al., 1996; Engel et al., 1997). Oltre alla specificità funzionale, molte aree funzionali sono fortemente organizzate dal punto di vista topografico. Un esperimento progettato intelligentemente consente di sfruttare le tecniche di neurovisualizzazione per esplorare l'organizzazione topo grafica all'interno di alcune di queste aree (Fox et al., 1986; Sereno et al., 1995; DeYoe et al., 1996; Engel et al., 1997).
All'interno di molte mappe esiste una divisione del lavoro tra moduli più piccoli che elaborano determinati tipi di informazione. Cellule specializzate in alcuni processi dell'elaborazione del colore, per esempio, sono distribuite attraverso la corteccia visiva primaria in strutture 'a macchia'. Le cellule sensibili all'orientamento sono sistemate in colonne secondo un orientamento specifico. Mentre questo livello di organizzazione resta al di là dell' attuale risoluzione spaziale delle tecniche di visualizzazione, sarà forse possibile studiarlo con i futuri sviluppi tecnologici. Al momento, comunque, è possibile esplorare la fisiologia di queste classi di neuroni visualizzando, durante la variazione parametrica dello stimolo, le dimensioni a cui essi sono sensibili. Numerosi studi, per esempio, hanno dimostrato l'esistenza di diverse sensibilità di contrasto nelle regioni della corteccia visiva (Tootell et al., 1995; Boynton et al., 1996).
Tuttavia, è vero in larga misura che la funzione cognitiva è distribuita, e ciò determina implicazioni significative per la neurovisualizzazione. L'elaborazione è distribuita in un senso macroscopico, poiché differenti regioni del cervello sono responsabili dell'elaborazione di diversi aspetti di un compito, e per l'esecuzione di quest'ultimo sono necessarie molte regioni discrete che lavorino in concerto fra loro. Anche i compiti cognitivi più elementari hanno molteplici componenti, i cui correlati neuronali possono agire in parallelo o in serie in molte parti diverse del cervello. PET e fMRl, raccogliendo i dati su tutto il cervello, sono le uniche tecniche adatte a esplorare la distribuzione delle componenti funzionali entro quest'organo. L'elaborazione neuronale può essere vista come distribuita anche a livello microscopico, poiché reti di molteplici neuroni sono in molti casi necessarie per elaborare le complesse funzioni richieste da molti compiti.
All'interno di una regione corticale le reti locali responsabili di attività specifiche possono estendersi su una regione vasta diversi millimetri. Ciò non impedisce l'uso della neurovisualizzazione per lo studio di queste regioni, ma è importante riconoscere le implicazioni dell' elaborazione locale. In primo luogo, il ruolo svolto dai singoli neuroni non è analizzabile attualmente con PET o fMRI. Inoltre, diverse popolazioni di neuroni possono occupare la stessa piccola regione di tessuto ma essere preposte ad attività di elaborazione differenti, come nel caso della corteccia visiva primaria. Risposte emodinamiche simili possono, quindi, essere ottenute da stimoli e compiti diversi. In tali circostanze non è possibile distinguere le popolazioni neuronali l'una dall'altra. L'attivazione di popolazioni neuronali sovrapposte può limitare la capacità di queste tecniche di effettuare distinzioni funzionali molto sottili. Per questo motivo, tali tecniche non saranno, probabilmente, mai utili per 'leggere il pensiero' o per altri compiti che richiedano un'unica attribuzione a schemi di attività.
Una prospettiva sistemica del cervello rivela che l'attività neuronale durante la realizzazione di compiti cognitivi è localizzata fortemente a livello delle regioni corticali, per quanto queste regioni possano essere ampiamente distribuite nel cervello. Nel determinare quali regioni siano attive, e durante quali compiti, stiamo iniziando a capire come il cervello suddivida compiti cognitivi complessi in componenti più semplici. Gli esperimenti di neurovisualizzazione hanno fornito delle informazioni attendibili e riproducibili sui centri dell' attività neuronale durante compiti che vanno dalla vista al linguaggio, alla memoria e all'abilità di apprendimento. Ciò dimostra che il cervello non è un organo omogeneo, nel quale tutti i processi cognitivi impiegano gli stessi gruppi di neuroni, ma è piuttosto una struttura altamente organizzata con unità funzionali specifiche.
Implicazioni della variabilità individuale
Dato che il cervello è suddiviso in sotto unità funzionali, è interessante chiedersi quanto esse siano stereotipate. Studi funzionali e anatomici hanno dimostrato che l' organizzazione del cervello è simile nella maggior parte degli individui normali, ma dettagli nelle dimensioni, nella forma e nell' esatta dislocazione delle aree funzionali e dei riferimenti anatomici variano approssimativamente nell' ordine di un centimetro. Questa variabilità, anche se troppo ridotta per invalidare gli approcci di neurovisualizzazione funzionale, ha però implicazioni che riguardano l'accuratezza e la precisione con cui le regioni funzionali possono essere localizzate mediante PET e fMRI.
Allo scopo di ottenere delle misure affidabili nelle regioni in cui il segnale associato a compiti particolari è piuttosto debole, vengono utilizzate tecniche statistiche per migliorare l'SNR. Nella PET vengono mediate diverse scansioni (tipicamente 6 o più) relative allo stesso tipo di compito, condotte, nella gran parte dei casi, su un certo numero di soggetti diversi. Il fatto di lavorare con diverse anatomie introduce una certa variabilità. Inoltre, l'esecuzione di una media intersoggettiva impone di trovare un mezzo per sistemare ciascuna scansione in uno spazio anatomico uniforme che per convenzione è il cosiddetto atlante di Tailarach (Tailarach e Toumoux, 1988). Gli algoritmi attualmente a disposizione per registrare le scansioni in uno schema standard hanno una precisione di circa 3 mm. Questi fattori abbassano la precisione con cui la PET può collocare segnali in determinate aree del cervello. Tuttavia, dal momento che gli effetti primari delle differenze individuali sono nell'ordine delle dimensioni del filtro usato per trattare i dati grezzi, la risoluzione non risulta troppo ridotta. L'alta riproducibilità dei centri di attivazione in studi e soggetti diversi attesta la notevole coerenza dell'organizzazione cerebrale in individui diversi (Hunton et al., 1996). Ciò significa che la variabilità intersoggettiva rappresenta soltanto un disturbo di poco conto di fronte all'elevato grado di coerenza.
Viceversa, la fMRI può in qualche misura evitare il problema della variabilità individuale poiché interi studi possono essere realizzati su un singolo soggetto. Tuttavia, rimane la necessità di collocare le attivazioni in uno spazio comune allo scopo di confrontare individui e studi diversi e costruire un modello globale di funzionamento del cervello. La variabilità individuale è un fattore di complicazione rispetto a questo scopo, ma si stanno sviluppando metodi migliori che utilizzano sia i punti di riferimento anatomici sia quelli funzionali allo scopo di registrare i singoli cervelli in un atlante comune.
La conoscenza dei processi cognitivi come ausilio nella progettazione degli esperimenti
La conoscenza delle potenzialità e dei limiti degli strumenti di misurazione e la comprensione della biologia del cervello sono di fondamentale importanza, ma, allo scopo di progettare o valutare gli studi di visualizzazione funzionale, è necessario anche conoscere le basi degli approcci cognitivi. La progettazione e l'analisi degli esperimenti rappresentano le sfide più grandi di questa impresa. Qui di seguito verranno discussi alcuni problemi essenziali per la progettazione di adeguati esperimenti di neurovisualizzazione e per l'interpretazione dei loro risultati.
Negli studi sull'attivazione vengono utilizzati tre tipi di compiti: i compiti bersaglio, comprendenti un compito o un sottocompito di interesse; i compiti di comparazione, simili ai precedenti, ma che manipolano o mantengono costante una componente di interesse; i compiti di base, di livello più basso, usati per rappresentare uno stato inattivo. Le differenze nell'attivazione regionale tra scansioni di compiti attivi o tra scansioni di compiti attivi e di base riflettono differenze nelle richieste associate al compito. La selezione dei compiti da sottoporre a scansione è estremamente importante poiché influenza l'interpretazione e il risultato dello studio. La scelta delle condizioni di base e di comparazione è in parte governata dalla scomposizione funzionale del compito di interesse primario.
La scomposizione funzionale
Compiti cognitivi complessi sono quasi sempre costituiti da numerose sottooperazioni. In gran parte degli esperimenti sull'attivazione, per esempio, un soggetto riceve istruzioni, percepisce stimoli, svolge determinate operazioni cognitive e risponde apertamente in un modo prescritto. La scomposizione funzionale si riferisce alla scomposizione concettuale di un compito nelle sue componenti e rappresenta una 'incisione' nell'integrità di un compito, lungo le sue giunture funzionali. Una scomposizione funzionale attenta e appropriata, sebbene attiri raramente l'attenzione, è alla base del successo di un esperimento di neurovisualizzazione.
Di solito uno studio sull'attivazione è ispirato dall'interesse verso un particolare tipo di processo cognitivo. Mentre è ben evidente che un'ipotetica scomposizione dei processi che avvengono durante un periodo di scansione è necessaria per interpretare i risultati degli esperimenti di neurovisualizzazione, è meno frequentemente riconosciuto che, negli esperimenti migliori, particolari della stessa modalità sperimentale emergono da successivi ripetuti momenti di scomposizione funzionale. Poiché la scomposizione funzionale ha conseguenze su quasi tutti gli aspetti dell'impostazione sperimentale, qui di seguito discuteremo in modo abbastanza approfondito i passaggi della progettazione di un esperimento di visualizzazione che sono collegati o che si basano sulla scomposizione funzionale.
Scomposizione funzionale e impostazione dell'esperimento. - Solitamente i processi cognitivi che gli studi di neurovisualizzazione prendono in esame sono quelli di interesse generale e le procedure scelte per arrivare a questi processi sono spesso quelle che sono state studiate a fondo con altri metodi empirici. Molti studi di psicologia cognitiva, per esempio, si sono indirizzati verso la scomposizione di procedure complesse in componenti più semplici ed essenziali e verso la comprensione di come queste componenti interagiscano con compiti specifici. Se è disponibile la relativa documentazione, di carattere psicologico o neuropsicologico, riguardante la natura del compito, questa può essere usata per indirizzare la scomposizione funzionale in un esperimento di visualizzazione.
Molto di rado è possibile scomporre completamente un processo in senso funzionale sulla base di dati empirici. In assenza di prove empiriche indipendenti, come si può affrontare il problema della scomposizione funzionale? Qui le tecniche di una corretta impostazione sperimentale sono di vitale importanza per compensare la mancanza di una scomposizione funzionale complessiva. Compiti di comparazione devono essere strettamente associati al compito bersaglio su molti fronti allo scopo di cercare di manipolare specifiche richieste del compito e di mantenere costanti, per quanto possibile, le caratteristiche del compito non rilevanti per il problema in questione. Questo approccio sperimentale evita l'implicita assunzione di una scomposizione funzionale in quelle aree in cui è sconosciuta.
Il metodo più diretto per affrontare l'esperimento consiste nel caratterizzare un paradigma del compito bersaglio tenendo conto delle informazioni introdotte, delle necessità di elaborazione e dei risultati. L'esperimento ideale mantiene due di queste tre componenti fissate nei compiti di comparazione e manipola il più basso numero possibile di variabili per volta. Così, si potrebbero porre domande del tipo: qual è la modalità sensoria degli stimoli usati? Qual è la modalità della risposta? Che tipo di informazione è manipolata (linguistica, visuale, acustica, ecc.)? Che tipi di rappresentazioni cognitive è probabile siano coinvolte e che tipi di operazioni cognitive invocate? Ci sono particolari modelli cognitivi che siano stati suggeriti per lo svolgimento di un compito? Poiché così tante rilevanti operazioni cognitive non sono completamente specificate, un protocollo sperimentale semplice ed efficiente, che vari il minor numero possibile di componenti non direttamente rilevanti per il compito in atto, sarà il più interpretabile. In questo modo differenze nell'attivazione tra le condizioni di scansione potranno essere attribuite senza ambiguità alle variabili manipolate. In modo analogo, i parametri mantenuti costanti tra i compiti potranno essere usati per stabilire i ruoli funzionali delle regioni fortemente attive tra le scansioni.
Nel progettare i compiti di comparazione occorre tenere presente che variazioni apparentemente piccole nei tratti superficiali tra i compiti possono avere effetti sorprendenti sugli schemi di attivazione (Fox e Raichle, 1984). Parametri come la frequenza di presentazione visiva, la frequenza delle risposte motorie, la frequenza o familiarità di parola e così via possono influire notevolmente sull' emodinamica e confondere l'interpretazione dei risultati sperimentali. Di conseguenza è importante cercare di adattare per quanto possibile questi parametri alle condizioni sperimentali, a meno di verificame esplicitamente gli effetti. Per esempio, nell' esaminare le differenze di elaborazione tra parole ad alta e bassa frequenza, le parole dovrebbero essere presentate nella stessa misura, essere della stessa lunghezza e dello stesso numero di sillabe, e l'ideale sarebbe che fossero il più possibile corrispondenti anche nella composizione fonetica. Un diagramma funzionale delle informazioni in entrata, delle ipotetiche fasi intermedie di elaborazione e dei risultati può essere un'utile guida per sviluppare un modello sperimentale complessivo e per valutare la scelta appropriata degli stimoli e dei compiti.
Scelta dei compiti di base. - Molti studi sull'attivazione sono volti a rispondere alla domanda: quali regioni del cervello sono coinvolte nel compito X? Per rispondere a questa domanda l'ideale sarebbe confrontare i livelli di attività durante un compito X con i livelli di attività quando il cervello 'non sta facendo nulla' o è a riposo. Ciò è impossibile, in quanto il cervello non è mai veramente a riposo, dovendo presiedere alle funzioni omeostatiche, regolare il livello di attenzione o di allerta, eccetera. È però possibile individuare condizioni che prevedono un'elaborazione cognitiva minima. Alcuni studi prescrivono di dire al soggetto di 'pensare al nulla'. Poiché è piuttosto difficile che questo avvenga, più spesso si scelgono come compiti di base dei compiti semplici che tuttavia richiedono un minimo di attenzione, come mantenere la vista fissa su un punto al centro del campo visivo. L'impegno su un compito preciso aumenta la probabilità che si realizzino scansioni di base uniformi nei diversi soggetti, e rende meno probabile che il soggetto si impegni in altri processi cognitivi più difficili da determinare. Naturalmente resta sempre la possibilità che i soggetti si impegnino in processi estranei durante lo svolgimento di un compito specifico.
I compiti di base vengono solitamente scelti in modo tale che essi non condividano importanti caratteristiche o esigenze con il compito di interesse. Bisogna sempre tenere a mente che il compito scelto come presumibilmente semplice potrebbe in realtà non essere tale, o potrebbe avere in comune con il compito di interesse alcune componenti, senza che lo sperimentatore ne sia consapevole. Molti gruppi utilizzano compiti di base standard in tutti i loro studi, indipendentemente dall'obiettivo dello studio. Ciò rende più facile trovare degli schemi comuni di attivazione in molti studi diversi e apre la possibilità a future metaanalisi. A prescindere dalla scelta del compito di base, è di primaria importanza essere consapevoli delle operazioni cognitive richieste da quel compito, così come delle altre in cui il soggetto potrebbe impegnarsi.
Scelta dei compiti di comparazione. - I compiti di comparazione sono compiti più complessi, spesso di alto livello, scelti o perché differiscono in qualche modo specifico dal compito primario di interesse (nel qual caso sono a volte chiamati compiti di contrasto), o perché condividono con esso importanti caratteristiche, oppure per entrambi i motivi. Una scomposizione funzionale del compito primario di interesse può identificare caratteristiche funzionali che dovrebbero essere presenti o assenti nel potenziale compito di comparazione allo scopo di verificare un'ipotesi specifica sul ruolo funzionale di una o più aree oppure una particolare scomposizione funzionale. Si possono così sviluppare compiti che soddisfino questi criteri. La scelta dei compiti di comparazione o di contrasto è una discriminante fondamentale per l'effettiva pregnanza e validità di uno studio. Un'attenta scelta dei compiti può far molto per svelare il ruolo funzionale di una o più regioni del cervello, mentre una loro scelta inappropriata può produrre uno studio che fallisce nel centrare qualsiasi specifico problema cognitivo.
La misura del comportamento: un controllo essenziale. - Uno degli aspetti più spesso trascurati nell'impostazione di un esperimento di neurovisualizzazione è quello relativo al controllo del fatto che i soggetti svolgano effettivamente il compito che il ricercatore crede stiano svolgendo. È questo un requisito minimale per uno studio ben progettato. Per tale motivo è necessario acquisire misure comportamentali nel corso delle sedute esplorative. Dati come i tempi di reazione, i parametri di precisione, la frequenza dell'azione, possono essere usati per valutare lo svolgimento del compito e le strategie usate che hanno un impatto significativo sull' interpretazione dei risultati della neurovisualizzazione. Per esempio, la difficoltà del compito, gli effetti relativi all'attenzione, o gli effetti dell'innesco o della pratica possono influenzare le risposte emodinamiche locali, e possono essere valutati analizzando le differenze dei tempi di reazione. Le misurazioni comportamentali aiutano a stabilire che i processi cognitivi che il ricercatore si prefigge di verificare siano quelli che effettivamente egli sta investigando. Con analisi sofisticate essi si possono correlare ai risultati dell'attivazione per determinare gli effetti della pratica, la difficoltà, ecc. È quasi superfluo rilevare che una conduzione fortemente comportamentale del compito e dei metodi di valutazione del compito stesso è fondamentale per impostare correttamente ed eseguire con successo un esperimento.
In certe condizioni, specialmente quelle della fMRI, in cui il movimento facciale che accompagna le risposte verbali può creare serie interferenze, non è possibile misurare il comportamento palese durante la scansione. Si possono però impiegare delle strategie per valutare se il compito viene svolto nel modo previsto. Il ricordo degli oggetti, per esempio, può essere condizionato manipolando il modo in cui essi vengono codificati: un compito a codificazione profonda, come un giudizio semantico, produce un ricordo migliore rispetto a un compito a codificazione debole, come un giudizio ortografico. Si possono presentare ai soggetti alcuni oggetti e chiedere loro di svolgere segretamente dei compiti che comportano una manipolazione in profondità dell'elaborazione. Al termine della seduta sarà possibile stabilire se abbiano o meno svolto questi compiti, come richiesto, attraverso dei test sulla memoria.
Nessuna scomposizione funzionale è inattaccabile. - Per quanto una data scomposizione funzionale possa sembrare completa e giustificata, bisogna ricordare che il suo ruolo è quello di avanzare un'ipotesi di lavoro, una soltanto tra le numerose alternative possibili. La validità di una particolare scomposizione funzionale si può sempre mettere in discussione, specialmente se le prove empiriche ottenute dall' esperimento o da altre fonti non si conformano al modello ipotizzato. Questa cautela è di fondamentale importanza e verrà ricordata anche nel seguito.
Sottrazione
Nell'analisi dei dati sperimentali il confronto diretto tra i compiti bersaglio e i compiti di comparazione rivela le regioni attivate in modo diverso nei due casi. In alternativa, ciascun compito di contrasto può essere confrontato con gli altri tenendo fermo un compito di base comune che rivela sia le analogie sia le differenze tra i compiti. Entrambi questi approcci si basano su un procedimento di sottrazione, il metodo più semplice, più diretto e anche più frequentemente usato per confrontare l'attivazione cerebrale durante due stati diversi. Ma questo metodo è anche quello più spesso frainteso.
La maggior parte degli esperimenti di neurovisualizzazione utilizza la sottrazione per individuare le regioni di variazione emodinamica tra due diversi compiti. Le immagini mediate e normalizzate di una situazione di base o di comparazione vengono sottratte dalle immagini mediate e normalizzate di un compito bersaglio. Le regioni a intensità positiva nell'immagine risultante riflettono le regioni cerebrali maggiormente attive nel compito bersaglio; le regioni a intensità negativa riflettono le regioni maggiormente attive nel compito di base o di comparazione. Le differenze nell'attivazione locale tra le diverse condizioni di scansione si possono attribuire alle differenze tra i compiti svolti. Le implicazioni della sottrazione di immagine non possono andare oltre, in quanto la sottrazione è soltanto un mezzo in grado di rivelare le differenze di attività tra due condizioni diverse.
Il fraintendimento nasce perché le implicazioni della sottrazione di immagine vengono spesso fuse con quelle associate ai metodi di sottrazione negli studi sui tempi di reazione, che assumono la pura additività. Gli studi sui tempi di reazione sono infatti basati sulla premessa che passaggi di elaborazione aggiuntivi richiedono tempi aggiuntivi di elaborazione, in modo additivo: la sottrazione dei tempi di reazione del processo A dai tempi di reazione dei processi A+B dà come risultato il tempo di elaborazione del processo B. Le interazioni tra i processi o la delezione dei processi non vengono prese in considerazione. Nella neurovisualizzazione, un'analisi dei compiti nella quale questi sono concepiti come complementari, con ciascuno collegato all'altro ma progressivamente più complesso, si basa su un'assunzione simile. Tale interpretazione implica che sottraendo l'attivazione collegata a un compito dall'attivazione collegata a un compito successivo e più complesso sia possibile isolare le regioni cerebrali responsabili delle richieste aggiuntive. Con una scomposizione funzionale appropriata un tale approccio potrebbe dimostrarsi illuminante, ma una varietà di interazioni complesse tra le zone cerebrali o tra le richieste dei compiti potrebbe invalidare l'interpretazione.
L'uso della sottrazione di immagine di per sé non implica alcun impegno per quanto concerne la relazione fra due o più compiti organizzati gerarchicamente. Le critiche rivolte alla sottrazione di immagine sono mal indirizzate, in quanto esse dovrebbero mirare piuttosto alla particolare strategia interpretativa nella quale si presume che i compiti cognitivi siano collegati. Un tale fraintendimento deriva dalla confusione tra metodi analitici e interpretazione dei risultati.
Implicazioni pratiche. - Poiché i dati sull'attivazione sono sempre analizzati in riferimento a una situazione di comparazione, quei processi che sono sempre attivi, e quindi presenti sia nelle scansioni attive che in quelle di base, vengono eliminati nel corso dell'analisi. Si potrebbe pensare che questo sia un grave svantaggio della tecnica di sottrazione. Tuttavia, se un processo è sempre attivo e la sua attività non può essere manipolata, nessuna tecnica sarà in grado di determinare il suo funzionamento, dal momento che non sarà possibile fare alcuna correlazione con un'altra variabile.
Messe di fronte a questo scenario, le tecniche di neurovisualizzazione non sono meno efficaci di altre neurotecniche. In realtà, pochi processi cognitivi di interesse, seppure esistono, risultano essere costantemente attivi a un singolo livello. L'individuazione dei processi cognitivi e di quelli neuronali è solitamente portata a termine attraverso manipolazioni: come cambia il comportamento se un soggetto è sottoposto allo stimolo A piuttosto che a quello B? L'attività di questo particolare neurone aumenta in presenza dello stimolo C? Finché il livello di attività di un processo può essere manipolato, le differenze nell'entità delle attivazioni saranno evidenti e potranno essere individuate con le tecniche di sottrazione.
Le insidie dell'attribuzione funzionale
Dato che la neurovisualizzazione cerca di attribuire ruoli funzionali a regioni cerebrali specifiche o a gruppi di regioni, non è sorprendente che le pubblicazioni sulla neurovisualizzazione spesso affermino, in modo più o meno esplicito, che un certo studio dimostra che l'area X attiva la funzione Y. È raro che ciò sia vero. È virtualmente impossibile interpretare la rilevanza funzionale di una qualsiasi regione attivata sulla base di una o di alcune scansioni. Quanto si può dire con sicurezza è che il compito complesso A, realizzato con i parametri usati nell'esperimento, induce un'attività nella regione X e che tale compito può essere provvisoriamente scomposto in senso funzionale nei sottocompiti W, Y, Z, ecc. Al massimo, un esperimento di neurovisualizzazione può fornire forti indizi che quella regione sia coinvolta in un particolare processo. Nessuno studio singolo può dimostrare la funzione di una particolare regione cerebrale. Perché si possa con forza stabilire un'attribuzione funzionale è necessario un processo esteso e interattivo che defrniamo triangolazione funzionale.
L'importanza della triangolazione funzionale
Non diversamente dalla triangolazione nelle rilevazioni, la defrnizione del panorama anatomico-funzionale richiede molteplici misurazioni nel corso di una varietà di compiti, e un continuo affrnamento dell'interpretazione del ruolo che una particolare area svolge nella cognizione. L'osservazione dell' attivazione di una specifica zona del cervello in una vasta gamma di compiti caratterizzati da una stessa sottocomponente funzionale costituisce una buona prova per l'attribuzione funzionale. L'assenza di quella attivazione in compiti simili privi di quella componente funzionale rafforza ancor più l'inferenza. La sfida della triangolazione funzionale risiede nell 'individuazione di compiti appropriati per verificare le ipotesi funzionali. Dal momento che lo 'spazio' della funzione cognitiva è multidimensionale, possono essere necessarie molte misurazioni per accumulare prove sufficienti a triangolare la funzione in una qualche area, con la clausola che qualsiasi attribuzione funzionale è incrementale e aperta a revisioni di fronte a eventuali dati incoerenti. Si deve notare che la triangolazione funzionale si basa fortemente sulla scomposizione funzionale e sulla modalità sperimentale seguita, ed è quindi soggetta a errori derivanti da errori in questi due processi. Se possibile, si dovrebbero utilizzare, per l'interpretazione dei dati, anche risultati provenienti da tecniche diverse dalla neurovisualizzazione. In effetti gran parte delle pietre di paragone per le nostre interpretazioni, come l'individuazione e la determinazione delle funzioni fondamentali delle aree sensorie primarie, derivano da studi sul comportamento in seguito a lesione negli esseri umani e da studi fisiologici sui primati non umam.
L'importanza della triangolazione funzionale può essere ulteriormente delineata considerando un caso particolarmente problematico di attribuzione funzionale: l'elaborazione automatica. Si afferma a volte che certi tipi di stimoli si servono automaticamente di certi percorsi di elaborazione e che quindi questi processi sono sempre attivi in determinati contesti. Per esempio, è stato sostenuto che l'analisi semantica procede automaticamente al manifestarsi di una parola, a prescindere dal fatto che il soggetto vi presti consapevolmente attenzione o acceda al significato della parola. Poiché la sottrazione elimina l'evidenza di un processo presente sia nella scansione bersaglio che in quella di base, qualsiasi scansione di base che impieghi parole maschererà la presenza di attivazioni associate a un'elaborazione semantica automatica. Con un'appropriata triangolazione funzionale (che implica un' appropriata scelta delle condizioni di base e di contrasto) anche questi processi possono essere osservati con la neurovisualizzazione: nel compito di base o di contrasto devono essere inseriti stimoli simili a quelli che si ritiene reclutino questi processi automatici, ma che differiscano da essi per il fatto di non permettere a tali processi di svolgersi. Così, per esempio, le regioni che potrebbero essere coinvolte in un'analisi semantica automatica sono quelle attivate dal manifestarsi passivo di parole reali ma non dal manifestarsi passivo di pseudoparole (sequenze pronunciabili e percepibili come simili a parole che non hanno un significato associato) o di parole in una lingua non familiare, ecc. Se si osservasse che queste stesse regioni vengono attivate in tutti i compiti che contemplano la presentazione di parole, si otterrebbe un'utile prova a sostegno della tesi dell'elaborazione automatica. La principale difficoltà di questo approccio è che le regioni candidate all'elaborazione automatica in questione non possono essere funzionalmente dissociate dagli altri processi che necessariamente differiscono tra situazioni di comparazione e situazioni bersaglio. In questo caso, per esempio, le attivazioni osservate in presenza di una parola, ma non di una pseudoparola, potrebbero corrispondere sia a un'analisi semantica sia a un altro processo che può sempre accompagnare la lettura, come l'accesso lessicale. Si deve anche ricordare che spesso le richieste del compito si possono manipolare, anche quando il compito non si può del tutto eliminare. Gli effetti di questa manipolazione si possono osservare nei risultanti dati sull'attivazione.
Anche utilizzando un approccio di triangolazione funzionaIe, resta la possibilità che una singola regione sia responsabile di alcuni o perfino tanti diversi tipi di operazioni cognitive. Questa conclusione non riflette necessariamente un'incoerenza biologica o interpretativa. È risaputo che in certe aree cerebrali, come la corteccia visiva, differenti classi di cellule in una piccola regione sono responsabili di differenti operazioni. E questo accade probabilmente in tutto il cervello. Poiché il flusso sanguigno viene misurato (e forse regolato) a livello regionale (nell'ordine dei millimetri) e non a livello cellulare (nell'ordine dei micron), l'attivazione di differenti popolazioni di neuroni nella stessa area può portare a schemi di flusso sanguigno identici, almeno data la risoluzione e la sensibilità delle attuali tecniche di neurovisualizzazione. Quindi l'attivazione della stessa regione in compiti diversi non implica necessariamente l'operatività di identiche componenti funzionali. L'analisi dei dati sperimentali l metodi di neurovisualizzazione presentano un rumore intrinseco, e allo stato attuale c'è scarso consenso riguardo ai corretti metodi di analisi statistica dei dati. L'analisi dei dati risultanti da PET e fMRI cerca di individuare attivazioni regionali e, come accade in gran parte delle ricerche scientifiche, di determinare la significatività statistica delle variazioni misurate. In questo paragrafo discutiamo brevemente il fondamentale problema dell'analisi dei dati della neurovisualizzazione, e indichiamo i punti che meritano una particolare considerazione quando si valutano i metodi di analisi.
Comparazioni multiple
La maggiore difficoltà nel determinare la significatività statistica delle variazioni di segnale osservate è il problema delle comparazioni multiple. Una tipica immagine del cervello umano contiene migliaia di voxel. La soglia di significatività comunemente usata nei metodi statistici standard (p <0,05) indica che una misurazione sopra la soglia su venti è un falso positivo. Dal momento che le statistiche sulla neurovisualizzazione vengono realizzate su migliaia di voxel, misure significative non corrette genereranno molti falsi positivi. Per fronteggiare questo rischio, devono essere impiegate contromisure appropriate. l metodi studiati per affrontare questo problema comprendono l'aggiustamento delle soglie di significatività con il metodo di Bonferroni o con altre correzioni, una varietà di passaggi di riduzione dei dati, come la suddivisione dei dati in gruppi di verifica a generazione indipendente di ipotesi, o l'assunzione che un certo numero di voxel contigui debba superare la soglia di significatività.
Sottrazione versus correlazione
La significatività dei dati PET è determinata mediante test statistici su un certo numero di immagini indipendenti calcolate come differenza tra un compito bersaglio e un compito di base. Sia l'attendibilità della misurazione che l'entità dei cambiamenti influenzano il grado calcolato di significatività statistica. Poiché i dati fMRI includono una componente temporale, essi possono essere analizzati sia con una tecnica di sottrazione, nella quale la media delle misure di controllo sia sottratta dalla media delle misure attivate, sia attraverso la correlazione dei tempi di attivazione con un segnale modello a variazione temporale (quando il segnale modello è una funzione a forma rettangolare, la correlazione è matematicamente equivalente alla sottrazione). Sia le analisi a sottrazione dei dati sia quelle a correlazione possono impiegare uno shift di fase per tenere conto del prolungarsi del segnale emodinamico, con notevole miglioramento del segnale. Il vantaggio della sottrazione rispetto alla correlazione è che i dati indicanti l'intensità delle risposte non vengono scartati. L'intensità delle risposte è un elemento significativo dell'attivazione cerebrale e un importante complemento per la localizzazione delle risposte. Il vantaggio della correlazione è che la durata temporale di riferimento può essere costruita in modo da includere i tempi di inizio e di caduta del segnale che si avvicinano maggiormente alle variabili biologiche rilevanti, fornendo in alcuni casi un miglior rapporto tra segnale e rumore. È possibile sfruttare il meglio dei due metodi facendo un'analisi iniziale basata sulla correlazione e registrando successivamente le variazioni di intensità delle regioni significativamente attive.
La scelta dei test statistici
Un fattore fondamentale che contribuisce alla difficoltà di analizzare i dati di neurovisualizzazione è che il rumore dell'immagine di attivazione non è distribuito in modo statisticamente normale e non esiste a tutt'oggi una sua caratterizzazione generale. Per questo motivo, non è stato possibile determinare le procedure statistiche più adatte ad analizzare questi dati. Attualmente gran parte degli studi impiega statistiche parametriche; tuttavia, un crescente numero di ricercatori analizza i dati fMRl con statistiche non parametriche, come la statistica di Kolmogorov-Smirnov e il test di Wilcoxon a somma media. La progettazione di metodi statistici per l'analisi di dati di visualizzazione è un campo di ricerca in espansione. Si deve anche dire, tuttavia, che, malgrado la varietà degli approcci attualmente impiegati, in prima approssimazione i diversi metodi statistici forniscono risultati molto simili.
Il contributo della visualizzazione alla conoscenza del funzionamento del cervello
Avendo ormai acquisito in un certo senso i limiti cui sono soggetti i metodi di neurovisualizzazione, il lettore si potrebbe chiedere quali progressi siano stati compiuti grazie a queste tecniche. La neurovisualizzazione è stata a volte criticata per aver fornito informazioni che avrebbero potuto essere ottenute con altre neurotecniche. Critiche più aspre sostengono che i dati di neurovisualizzazione non sono state in grado di rivelare nulla che non sapessimo già. Nel contestare queste critiche illustreremo alcuni progressi ottenuti con tali tecniche.
È un caso piuttosto raro in qualunque campo che si ottenga un insieme correlato di informazioni con un approccio specifico e solo con quello. In questo senso le tecniche di neurovisualizzazione non sono privilegiate rispetto ad altre tecniche scientifiche. Mentre è certo che gran parte della conoscenza che abbiamo acquisito attraverso gli studi di neurovisualizzazione avrebbe potuto esserlo attraverso l'esame di una popolazione di pazienti con lesioni, o attraverso l'esame elettro fisiologico intraoperatorio di soggetti epilettici, o con altre tecniche sperimentali, bisogna sottolineare che i metodi di neurovisualizzazione offrono, in molti casi, un mezzo più pratico per studiare particolari problemi cognitivi. Utilizzando un numero ridotto di soggetti umani normali, gli esperimenti di neurovisualizzazione possono affrontare problemi cognitivi che altrimenti necessiterebbero di pazienti con lesioni fortemente specifiche, pazienti ai quali pochissimi ricercatori potrebbero avere accesso. Inoltre, dedurre un funzionamento normale da studi su pazienti con lesioni può essere problematico per diverse ragioni. Il danno alla regione deputata alla facoltà cognitiva in questione, o comprendente le fibre nervose che passano attraverso l'area lesionata, può influenzare il comportamento e complicare il compito di localizzare la funzione. Per di più, il danno cerebrale potrebbe avere effetti non specifici o secondari sui processi cognitivi che potrebbero non riflettere con precisione il ruolo della regione danneggiata nella cognizione. Per queste e altre ragioni l'uso di tecniche di neurovisualizzazione su soggetti normali è spesso più adatto per esplorare la cognizione umana. Per indicare più concretamente i tipi di contributi che la neurovisualizzazione è in grado di fornire, daremo alcuni esempi di studi che hanno accresciuto le nostre conoscenze.
Conferme sull'uomo di quanto già noto dagli studi sui primati
Numerosi studi su aree cognitive differenti, dagli studi sul sistema visivo a quelli sulla memoria, hanno confermato ipotesi formulate sulla base dei risultati degli studi sulle lesioni e sulla fisiologia dei primati, associando caratteristiche regioni del cervello a specifici ruoli cognitivi. Come è stato fatto in modo esauriente per le scimmie, è stata disegnata la corteccia visiva umana tenendo conto dell'eccentricità e dell'angolo visivo, ed è stata suddivisa in diverse regioni definite in senso funzionale (Fox et al., 1986; DeY oe et al., 1994; Sereno et al., 1995; Tootell et al., 1995; Engel et al., 1997). Inoltre, le funzioni di contrasto e di sensibilità al colore dei neuroni nella corteccia visiva umana sono state studiate in modo parametrico (Boynton et al., 1996). l risultati sono molto simili a quelli ottenuti con le tecniche fisiologiche nelle scimmie. Analogamente, le regioni nella corteccia prefrontale dorso laterale bilaterale, associate alla memoria operativa nelle scimmie, sono state confermate come sede della memoria operativa dagli studi di neurovisualizzazione effettuati sull'uomo (Petrides et al., 1993; Courtney et al., 1997).
Ottenimento di informazioni non disponibili dagli studi sugli animali
In aggiunta a quanto ipotizziamo sulla base della neurofisiologia dei primati, esiste la possibilità che i ruoli funzionali di certe aree del cervello non possano essere del tutto confrontabili tra l'uomo e i primati non umani. L'uomo, per esempio, possiede capacità cognitive che mancano ai primati non umani, come quella di usare il linguaggio; perciò certe regioni cerebrali devono essere state adattate a compiere queste funzioni. Gli studi di M. Petrides e collaboratori (1993) sulla memoria verbale operativa nell'uomo hanno dimostrato che le regioni della corteccia prefrontale dorsolaterale, omologhe a quelle che servono la memoria operativa nelle scimmie, sono coinvolte nel mantenimento dell'informazione verbale a breve termine. Inoltre, un gran numero di studi (Petersen et al., 1988; 1989; Petrides et al., 1993; Raichle et al., 1994; Buckner et al., 1996) ha accumulato prove del fatto che una regione nella corteccia prefrontale sinistra, vicina all'opercolo, è deputata alla realizzazione di compiti linguistici. Attualmente si stanno progettando studi mirati per isolare in modo più specifico il ruolo funzionale di quest'area nel linguaggio. Dunque, gli studi di neurovisualizzazione sull 'uomo consentono di esplorare la base neuronale delle funzioni cognitive specificamente umane.
Scelta tra modelli psicologici specifici
Nella psicologia cognitiva si verificano spesso situazioni in cui due o più teorie contrastanti spiegano ugualmente bene un fenomeno. In alcuni casi, la neurovisualizzazione può essere usata per fornire prove che aiutino a scegliere una teoria piuttosto che un'altra. Gli esperimenti di neurovisualizzazione, per esempio, hanno fornito importanti evidenze che permettono di scegliere tra due modelli psicologici, prima indifferenziabili, riguardo al ruolo che svolge l'attenzione spaziale nei compiti di ricerca visiva. Gli studi psicologici sui paradigmi della ricerca visiva riportano curve con un aumento del tempo medio di scoperta dell'obiettivo in presenza di un aumento di fattori distraenti in concomitanza con i compiti di ricerca visiva. È stato ipotizzato che queste curve risultino o dal decrescere dell' efficienza della ricerca parallela con più fattori distraenti o dal tempo aggiuntivo richiesto per realizzare cambi seriali di attenzione rispetto agli oggetti a disposizione.
l soli studi comportamentali non erano in grado di distinguere tra i due modelli. Con una serie di esperimenti PET, M. Corbetta e collaboratori hanno individuato le aree nella corteccia parietale superiore che sono coinvolte nei cambiamenti di attenzione spaziale. In uno studio successivo (Corbetta et al., 1995), questi autori hanno scoperto che queste stesse regioni nella corteccia parietale superiore erano fortemente attivate nel corso di un compito di ricerca sulla congiunzione, e ciò ha fornito una prova evidente che il cervello impiega una strategia di ricerca seriale per questi compiti di congiunzione, e non una ricerca parallela.
Prove a sostegno di ipotesi generali sulla cognizione
l miglioramenti nella precisione e nella velocità con cui si svolge un compito dopo un certo periodo di pratica sono stati attribuiti o al miglioramento, con la pratica, dell'efficienza operativa del sistema neuronale, o a un cambiamento nei circuiti neuronali che svolgono compiti ben noti o praticati. È probabile che un'analisi attenta della letteratura sulle lesioni avrebbe potuto consentire agli scienziati di discriminare tra queste due possibilità, ma il tipo di cambiamento sotteso agli effetti della pratica è divenuto chiaro soltanto quando tali effetti sono stati esaminati con l'uso dei metodi di neurovisualizzazione. Numerosi studi sia sull'apprendimento motorio che su quello cognitivo hanno dimostrato che differenti circuiti del cervello si attivano in relazione all'automaticità di un compito, e che questi percorsi neuronali possono cambiare nel giro di minuti. M.E. Raichle e collaboratori (1994) hanno dimostrato che un compito di risposta verbale non preparato attiva sicuramente regioni del cingolo anteriore prefrontale sinistro e delle cortecce temporali posteriori sinistre. Dopo un certo allenamento, le misure del comportamento cambiano: le regioni menzionate precedentemente diminuiscono la loro attivazione, e la corteccia silviana insulare mostra un'attivazione più alta; lo schema dell'attività non è distinguibile da quello osservato in un semplice compito di ripetizione di parole. Di fronte a un nuovo stimolo, gli schemi di attivazione si riconvertono in quelli di un compito non preparato. Risultati simili, che mostrano un cambiamento nell'architettura neuronale sottesa allo svolgimento di compiti semplici e sperimentati, sono stati forniti di recente a proposito dell'apprendimento motorio e visuomotorio (Doyon et al., 1996). Ulteriori esplorazioni dei tempi di cambiamento degli schemi e delle correlazioni con le variabili di comportamento getteranno nuova luce sui cambiamenti dei percorsi di elaborazione che accompagnano le realizzazioni più complesse.
Risultati imprevisti e nuove problematiche
Oltre ad aiutare a far luce su problemi che la psicologia e gli studi sulle lesioni non erano stati in grado di chiarire, la neurovisualizzazione ha fornito alcuni risultati del tutto imprevisti, ma sicuramente attendibili, le cui implicazioni per la teoria neuronale e psicologica restano oscure. Uno di questi è la scoperta decisamente importante che una regione nella corteccia prefrontale dorso laterale destra è coinvolta nel recupero di informazioni durante compiti di memoria episodica (Tulving et al., 1994). Una così chiara dissociazione del recupero della memoria episodica da altri tipi di recupero della memoria non era stata mai anticipata prima degli studi di neurovisualizzazione e, sebbene le implicazioni funzionali e cognitive di questa scoperta debbano ancora essere determinate, è probabile che possano influenzare in modo significativo la nostra conoscenza dell'organizzazione funzionale dei processi della memona.
Conclusioni
Numerosi e importanti progressi nella comprensione delle basi neuronali della funzione cognitiva sono stati già compiuti grazie alle tecniche di neurovisualizzazione, e altri senza dubbio verranno, dal momento che la nostra conoscenza continua a crescere e le tecniche usate per investigare la cognizione continuano a migliorare. Gli studi PET e fMRI sull'attivazione rappresentano veramente un matrimonio tra neuroscienza e psicologia, e forniscono sia una migliore comprensione della cognizione e degli approcci cognitivi sia una migliore conoscenza del cervello. È necessario conoscere entrambi questi domini per poter apprezzare le qualità e i limiti di queste tecniche e per potersi impegnare in modo critico nell'eccitante impresa di visualizzare il funzionamento del cervello umano.
Bibliografia citata
BOYNTON, G.M., ENGEL, S.A., GLOVER, G.H., HEEGER, D.J. (1996) Linear systems analysis of functional magnetic resonance imaging in human VI. J. Neurosci., 16, 4207-4221.
BUCKNER, R.L., BANDETTINI, P.A., O'CRAVEN, K.M., SAVOY, R.L., PETERSEN, S.E., RAICHLE, M.E., ROSEN, B.R. (1996) Detection of cortical activation during averaged single trials of a cognitive task using functional magnetic resonance imaging. Proc. NatI. Acad. Sci. USA, 93, 14878-14883.
CORBETTA, M., SHULMAN, G.L., MIEZIN, F.M., PETERSEN, S.E. (1995) Superior parietal cortex activation during spatial attention shifts and visual feature conjunction. Science, 270, 802-805.
COURTNEY, S.M., UNGERLEIDER, L.G., KEIL, K., HAXBY, J.V. (1997) Transient and sustained activity in a distributed neural system for human working memory. Nature, 386, 608-611.
DEYOE, E.A., BANDETTINI, P.A., NEITZ, J., MILLER, D., WINANS, P. (1994) Functional magnetic resonance imaging (FMRI) of the human brain. J. Neurosci. Methods, 54,171-187.
DEYOE, E.A., CARMAN, G.J., BANDETTINI, P.A., GLICKMAN, S., WIESER, J., Cox, R., MILLER, D., NEITZ, J. (1996) Mapping striate and extrastriate visual areas in human cerebral cortex. Proc. NatI. Acad. Sci. USA, 93, 2382-2386.
DOYON, J., OWEN, A.M., PETRIDES, M., SZIKLAS, V., EVANs, A.C. (1996) Functional anatomy of visuomotor skill leaming in human subjects examined with positron emission tomography. Euro. J. Neurosci., 8, 637-648.
ENGEL, S.A., GLOVER, G.H., WANDELL, B.A. (1997) Retinotopic organization in human visual cortex and the spatial precision of functional MRI. Cereb. Cortex, 7, 181-192.
FOX, P.T., MINTUN, M.A., RAICHLE, M.E., MIEZIN, F.M., ALLMAN, J.M, V AN ESSEN, D.C. (1986) Mapping human visual cortex with positron emission tomography. Nature, 323, 806-809.
FOX, P.T., RAICHLE, M.E. (1984) Stimulus rate dependence of regional cerebral blood flow in human striate cortex, demonstrated by positron emission tomography. J. Neurophysiol., 51, 1109-1120.
FOX, P.T., RAICHLE, M.E. (1986) Focal physiological uncoupling of cerebral blood flow and oxidative metabolism during somatosensory stimulation in human subjects. Proc. NatI. Acad. Sci. USA, 83, 1140-1144.
GEORGE, J.S., AINE, C.J., MosHER, J.C., SCHMIDT, D.M., RANKEN, D.M., SCHLITT, H.A., WOOD, C.C., LEWINE, J.D., SANDERS, J.A., BELLIVEAU, J.W. (1995) Mapping function in the human brain with magnetoencephalography, anatomical magnetic resonance imaging, and functional magneti c resonance imaging. J. Clinical Neurophys., 12, 406-431.
HUNTON, D.L., MIEZIN, F.M., BUCKNER, R.L., VAN MIER, H.I., RAICHLE, M.E., PETERSEN, S.E. (1996) An assessment offunction-anatomical variability in neuroimaging studies. Human Brain Mapping, 4, 122-139.
KWONG, K.K. et al. (1992) Dynamic magnetic resonance imaging of human brain activity during primary sensory stimulation. Proc. NatI. Acad. Sci. USA, 89, 5675-5679.
MALONEK, D., GRINVALD, A. (1996) Interactions between electrical activity and cortical microcirculation revealed by imaging spectroscopy: implications for functional brain mapping. Science, 272, 551-554.
MENON, R.S., OGAWA, S., HU, X., STRUPP, J.P., ANDERSON, P., UGURBIL, K. (1995) BOLD based functional MRI at 4 Tesla includes a capillary bed contribution: echo-planar imaging correlates with previous optical imaging using intrinsic signals. Magn. Reson. Med., 33, 453-459.
PETERSEN, S.E., FOX, P.T., POSNER, M.I., MINTUN, M., RAICHLE, M.E. (1988) Positron emission tomographic studies of the processing of single words. J. Cog. Neurosci., l, 153-170.
PETERSEN, S.E., FOX, P.T., POSNER, M.I., MINTUN, M., RAICHLE, M.E. (1989) Positron emission tomographic studies of the cortical anatomy of single-word processing. Nature, 331, 585-589.
PETRIDES, M., ALIVISATOS, B., MEYER, E., EVANS, A.C. (1993) Functional activation of the human frontal cortex during the performance of verbal working memory tasks. Proc. NatI. Acad. Sci. USA, 90, 878-882.
RAICHLE, M.E. (1987) Circulatory and metabolic correlates of brain function in normal humans. In Handbook of physiology: a criticaI, comprehensive presentation of physiological knowledge and concepts. The nervous system, vol. V: Higher Junctions of the brain, a c. di Mountcastle V.B., Plum F., Bethesda, American Physiological Society, e Baltimora, Williams & Wilkins, pp. 643-674.
RAICHLE, M.E., FIEZ, J.A., VIDEEN, T.O., MACLEoD, A.-M., PARDO, J.V., Fox, P.T., PETERSEN, S.E. (1994) Practice-related changes in human brain functional anatomy during nonmotor leaming. Cereb. Cortex, 4, 8-26.
SERENO, M.I., DALE, A.M., REPPAS, J.B., KWONG, K.K., BELLIVEAU, J.W., BRADY, T.J., ROSE N, B.R., TOOTELL, R.B. (1995) Borders of multiple visual areas in humans revealed by functional magnetic resonance imaging. Science, 268, 889-893.
TAILARACH, J., TOURNOUX, P. (1988) Co-planar stereotaxic atIas of the human brain. New York, Thieme MedicaI Publishers, Inc.
TOOTELL, R.B., REPPAS, J.B., KWONG, K.K., MALAcH, R., BORN, R.T., BRADY, T.J., ROSEN, B.R., BELLIVEAU, J.W. (1995) Functional analysis of human MT and related visual cortical areas using magnetic resonance imaging. J. Neurosci., 15, 3215-3230.
TULVING, E., KAPUR, S., MARKOWITSCH, H.J., CRAIK, F.I., HABIB, R., HOULE, S. (1994) Neuroanatomical correlates of retrieval in episodic memory: auditory sentence recognition. Proc. NatI. Acad. Sci. USA, 91, 2012-2015.
WOOLSEY, T.A., ROVAINEN, C.M., COX, S.B., HENEGAR, M.H., LIANG, G.E., LIU, D., MOSKALENKO, Y.E., SUI, J., WEI, L. (1996) Neuronal units linked to microvascular modules in cerebral cortex: response elements for imaging the brain. Cereb. Cortex, 6, 647-660.
ZEKI, S., WATSON, J.D., LUECK, C.J., FRISTON, K.J., KENNARD, C., FRACKOWIAK, R.S. (1991) A direct demonstration of functional specialization in human visual cortex. J. Neurosci., Il, 641-649.
Bibliografia generale
DEYOE, E.A., BANDETTINI, P.A., NEITZ, J., MILLER, D., WINANS, P. (1994) Functional magnetic resonance imaging (FMRI) of the human brain. J. Neurosci. Methods, 54,171-187.
GEORGE, J.S., AINE, C.J., MosHER, J.C., SCHMIDT, D.M., RANKEN, D.M., SCHLITT, H.A., WOOD, C.C., LEWINE, J.D., SANDERS, J.A., BELLIVEAU, J.W. (1995) Mapping function in the human brain with magnetoencephalography, anatomical magnetic resonance imaging, and functional magnetic resonance imaging. J. Clinical Neurophys., 12, 406-43l.
POSNER, M.I., RAICHLE, M.E. Images of Mind. New York, Scientific Arnerican Library, 1994.
RAICHLE, M.E. Circulatory and metabolic correlates of brain function in normal humans. In Handbook of physiology: a criticaI, comprehensive presentation of physiological knowledge and concepts. The nervous system, voI. V: Higher functions of the brain, a c. di Mountcastle V.B., Plum F., Bethesda, Arnerican Physiological Society, e Baltimora, Williams & Wilkins, 1987, pp. 643-674.
ROLAND, P.E. Brain activation. New York, Wiley-Liss, 1993.