
Bioenergetica
Bioenergetica
Il giorno in cui iniziai a scrivere questo testo la temperatura esterna nella maggior parte delle praterie canadesi era inferiore a −30 °C con punte di −40 °C, se si considerava il raffreddamento dovuto al vento. Guardando, però, fuori dalla finestra della cucina mi accorsi che intorno alla mangiatoia, appesa a una vecchia pianta rampicante, si era radunato il consueto gruppo di piccoli uccelli: cince, picchi muratore pettobianco, fringuelli e gli ubiquitari passeri che cercavano di farsi largo per raggiungere i semi neri dei girasole. Questa scena così familiare è in realtà una delle più sorprendenti lezioni di bioenergetica, lo studio della fissazione, della conversione e dell'utilizzazione dell'energia da parte dei sistemi viventi. Quei piccoli volatili, la maggior parte dei quali pesa meno di 15 g, mantengono la temperatura corporea interna a 40 °C, tre gradi in più rispetto a quella dei grandi mammiferi. Ciò significa che durante i freddi inverni canadesi si stabilisce un gradiente di temperatura dell'ordine di 80 °C attraverso i tre centimetri circa di spessore del loro minuscolo corpo, ossia tra i loro cuori dal battito veloce e la punta delle loro piume arruffate. Tutto ciò è straordinario, e tutto quello che gli uccelli devono fare è continuare a consumare i semi neri ricchi di olio e perciò di energia (i semi dei girasole, rispetto a quelli dei cereali, contengono circa il 50% in più di materiale digeribile ricco di energia) a una velocità che libera dal loro efficiente metabolismo enzimatico una quantità di calore superiore a quella persa per irraggiamento e convezione dai loro piccoli corpi. Per giungere a questa verità, all'apparenza tanto semplice, è stato necessario però molto tempo. La comprensione dell'equivalenza delle energie e del modo in cui funzionano il metabolismo animale o la fotosintesi vegetale e batterica è diventata possibile molto tempo dopo il calcolo dei tempi e dei luoghi delle eclissi solari.
La bioenergetica è dunque lo studio delle trasformazioni di energia negli organismi viventi e in particolare nelle cellule a livello molecolare. Una delle più importanti acquisizioni della biochimica moderna è stata infatti la scoperta che l'energia fornita dagli alimenti non viene direttamente utilizzata dagli organismi viventi. L'energia delle sostanze nutritive viene liberata in organelli specifici, i mitocondri, e accumulata come energia di legame in particolari molecole, la più importante delle quali è l'adenosintrifosfato (ATP) che si può formare per fosforilazione fotosintetica, utilizzando come fonte di energia la luce, nelle piante, oppure per fosforilazione ossidativa nel processo respiratorio di tutti gli organismi, vegetali e animali. I derivati fosforici ad alto contenuto di energia possono essere utilizzati non solo per la produzione di lavoro meccanico, osmotico o elettrico, ma anche per processi di sintesi allorché due sostanze si combinano tra loro mediante un legame endoergonico la cui formazione richiede un apporto di energia dall'esterno.
Cenni storici
Julius Robert von Mayer, mentre lavorava come medico su una nave olandese diretta a Giava, aveva notato che il sangue dei marinai dei Tropici aveva un colore più brillante. Aveva fornito una spiegazione corretta di tale fenomeno rilevando che nei climi più caldi il metabolismo basale richiedeva meno energia e di conseguenza meno ossigeno. Mayer considerava i muscoli come motori a combustione che ricevono energia dall'ossidazione del sangue; con i suoi calcoli dimostrò che l'energia chimica contenuta nel cibo è sufficiente a fornire l'energia meccanica necessaria per compiere il lavoro muscolare e per mantenere costante la temperatura corporea. Nel 1851 ciò lo condusse alla quantificazione dell'equivalente meccanico del calore e quindi alla formulazione della legge della conservazione dell'energia, che successivamente divenne nota come 'prima legge della termodinamica'. In modo indipendente una quantificazione più accurata dell'equivalenza tra lavoro e calore si ebbe con gli studi di James P. Joule, la cui prima pubblicazione del 1850 indicava un valore che si discostava soltanto dell'1% da quello reale.
Poco dopo, William Thomson (lord Kelvin) descrisse la tendenza universale verso la dissipazione dell'energia meccanica e Rudolf Clausius formalizzò questa intuizione giungendo alla conclusione che il contenuto di energia dell'Universo è fisso e che la sua conversione si traduce in un'inevitabile perdita di calore verso zone a energia inferiore: l'energia cerca una distribuzione uniforme e l'entropia dell'Universo tende al massimo. Questa seconda legge della termodinamica ‒ la tendenza universale verso il disordine e la dissipazione del calore ‒ divenne forse la più influente e fraintesa generalizzazione riguardante il cosmo. Soltanto dopo il chiarimento di questi fondamenti universali la bioenergetica poté fornire un contributo specifico e duraturo in merito all'utilizzazione dell'energia negli esseri umani, nei Mammiferi, negli organismi eterotrofi (che, non potendo sintetizzare le proprie sostanze nutritive, le devono assumere sotto forma di carboidrati, proteine e lipidi dai tessuti animali o vegetali) e negli organismi fotosintetici (autotrofi, in grado di produrre macromolecole organiche complesse da composti inorganici semplici).
La comprensione del metabolismo eterotrofo subì un naturale sviluppo grazie agli studi sulla nutrizione umana, che conobbero una maggiore diffusione e sistematizzazione durante l'ultima decade del XIX secolo. I molti esperimenti calorimetrici condotti dagli scienziati europei e americani durante questo periodo identificarono i precisi equilibri energetici degli organismi viventi e, nel 1902, Max Rubner poté offrire un'eccellente descrizione sistematica del metabolismo umano. Quest'ultima comprendeva quasi tutti i fondamenti attualmente conosciuti dell'energetica umana, incluse la determinazione dei valori di energia dei diversi alimenti, la distinzione tra l'assunzione di cibo e il consumo di energia, la comprensione che tutti i macronutrienti (carboidrati, proteine e lipidi) possono essere trasformati in lavoro (e quindi in calore), la correlazione tra le condizioni ambientali, le circostanze individuali e i tassi metabolici e tra il metabolismo basale e l'area della superficie corporea, insieme al riconoscimento degli effetti dinamici dell'ingestione di cibo (la digestione accelera il metabolismo).
Progressi importanti nella bioenergetica teorica sono stati compiuti tra le due guerre. Nel 1925 Alfred Lotka formulò la legge della massima energia, secondo la quale il fattore più importante per la crescita, la riproduzione, il mantenimento e l'irradiazione delle specie non è la migliore efficienza di conversione ma il maggior flusso di energia utile, cioè la massima potenza prodotta. Di conseguenza, gli organismi e i sistemi viventi non convertono l'energia con la maggiore efficienza possibile, ma piuttosto a tassi ottimizzati per la massima produzione di energia. Contemporaneamente agli studi di Lotka, Vladimir Ivanovich Vernadsky introdusse il concetto fondamentale di biosfera, il sottile strato in cui si distribuisce la vita sulla Terra, che penetra anche negli strati più profondi dell'idrosfera, in quelli più bassi dell'atmosfera e, come oggi sappiamo, persino a profondità considerevoli nella litosfera.
Nel suo libro pionieristico (pubblicato in Russia nel 1926 e in traduzione francese nel 1929), Vernadsky considerava la biosfera innanzitutto come un'entità attivata dalla radiazione solare, la cui materia raccoglie e ridistribuisce l'energia solare convertendola in energia libera capace di compiere lavoro sulla Terra. Di conseguenza nella visione di Vernadsky il Sole è responsabile dei processi della biosfera quanto dei processi terrestri. Negli scritti dell'ultimo periodo Vernadsky espresse in termini termodinamici la qualità della materia vivente come tendenza all'aumento dell'energia libera della biosfera, mentre i processi non viventi, con l'eccezione del decadimento radioattivo, ne riducono la presenza. Questa distinzione riguardante l'entropia divenne una caratteristica fondamentale delle definizioni di vita biologica proposte per la prima volta da Ludwig von Bertalanffy nel 1940 e successivamente da Erwin Schrödinger nel 1944.
Bertalanffy spiegò l'apparente contraddizione tra la seconda legge della termodinamica e la crescente complessità degli organismi viventi sottolineando come tutti i sistemi viventi, dalle cellule alle civiltà umane, siano sistemi aperti, che riescono a mantenersi in uno stato improbabile di ordine perfetto e di organizzazione crescente in quanto oppongono una resistenza temporanea all'imperativo entropico importando enormi quantità di energia libera; per usare la famosa definizione di Schrödinger: la vita perdura prosperando sulla negentropia.
Altri progressi importanti verificatisi tra le due guerre nella comprensione della bioenergetica includono lo studio dell'efficienza dei trasferimenti energetici tra i livelli trofici e del pattern generale sottostante i diversi tassi di metabolismo basale. Alla fine degli anni Venti Charles Elton fu il primo ecologo a riconoscere che ai livelli superiori delle reti alimentari il numero delle specie diminuisce mentre spesso aumentano le dimensioni corporee. Nel decennio successivo Evelyn Hutchinson ridefinì in termini di produttività il principio stabilito da Elton come piramide di numeri. Il suo lavoro portò direttamente all'approccio trofico-dinamico utilizzato da Raymond Lindeman per i flussi energetici negli ecosistemi. Lindeman è stato il primo biologo a quantificare in termini di efficienza progressiva (l'assimilazione dal livello n al livello n−1) l'inevitabile diminuzione di energia nei trasferimenti tra i livelli trofici successivi di un sistema lacustre. Oggi sappiamo che questa efficienza varia enormemente tra i diversi ecosistemi e i livelli trofici. Studi pionieristici quantitativi sui trasferimenti energetici in ecosistemi acquatici e terrestri rappresentativi sono stati compiuti dai fratelli Eugene e Howard Odum a partire dagli anni Cinquanta.
Rispetto a quanto verificatosi nel campo dell'energetica animale, fino a dopo la Seconda guerra mondiale ben poco era stato chiarito sulla fotosintesi: curiosamente si era arrivati a conoscere la struttura dell'atomo e a scomporlo prima di comprendere il modo in cui le piante verdi utilizzano l'acqua, l'anidride carbonica, i nutrienti e la radiazione solare per produrre nuova massa vegetale. Con la generazione successiva alla Seconda guerra mondiale la bioenergetica progredì a livello sia molecolare sia globale. Per quanto riguarda il livello molecolare, il risultato più importante fu conseguito durante la prima parte degli anni Cinquanta da Melvin Calvin e collaboratori, i quali riuscirono a spiegare la sequenza completa di riduzione fotosintetica che avviene nel ciclo del carbonio e che porta il nome di ciclo di Calvin.
I contributi principali alla comprensione dei processi di conversione dell'energia a livello degli ecosistemi e dei principali biomi provennero dall'International biological programme tra il 1964 e il 1974, che ebbe tra gli obiettivi prioritari quello di studiare la produttività degli ecosistemi terrestri, d'acqua dolce e marini. Attraverso le numerose pubblicazioni che ne derivarono si è raggiunta una migliore conoscenza dei tassi di conversione e di utilizzazione dell'energia nei più importanti ecosistemi terrestri e delle loro variazioni. Durante gli ultimi tre decenni del XX sec. lo studio della bioenergetica globale ha grandemente beneficiato del lancio di satelliti sempre più sofisticati per l'osservazione della Terra, le cui possibili applicazioni vanno dalla mappatura dei cambiamenti che si verificano nelle aree di foresta tropicale pluviale al monitoraggio della densità della clorofilla marina.
Le vie metaboliche
Le due classi fondamentali in cui è suddiviso il metabolismo, autotrofismo ed eterotrofismo, contengono una varietà di affascinanti permutazioni. Combinando i prefissi che identificano le fonti di energia (foto- e chemio) con quelli che si riferiscono ai donatori di elettroni (lito- e organo-), si è in grado di fornire descrizioni accurate di tutte le possibilità metaboliche. Gli organismi che ricavano energia dalla radiazione solare ‒ capaci di convertire l'energia elettromagnetica nei legami fosfato ad alta energia dell'adenosintrifosfato, o ATP (la principale molecola capace di immagazzinare energia) ‒ sono detti 'fotoautotrofi'. Quelli che sfruttano l'energia chimica (sia dei composti inorganici semplici sia delle macromolecole organiche complesse) sono detti 'chemioautotrofi'. Gli organismi che ricavano gli elettroni dagli elementi chimici (idrogeno, zolfo) o dai composti inorganici semplici (acqua, acido solfidrico) sono 'litotrofi', mentre quelli che utilizzano substrati organici complessi sono 'organotrofi' (per es., i batteri e i funghi che decompongono le proteine e la lignina della biomassa morta o gli erbivori che mangiano i carboidrati prodotti dalle piante).
Di conseguenza, tutti gli autotrofi fotosintetici (vegetali terrestri, alghe, fitoplancton, cianobatteri) che ricavano il carbonio dall'anidride carbonica atmosferica sono 'fotolitotrofi', analogamente ai solfobatteri verdi e purpurei, che invece dell'acqua utilizzano l'acido solfidrico come fonte di elettroni. I 'fotoorganotrofi' richiedono energia solare ma utilizzano composti organici: i batteri purpurei non contenenti zolfo possono convertirsi a questa modalità dal fotolitotrofismo anaerobico diventando eterotrofi. La maggior parte dei batteri e tutti i funghi eterotrofi, così come tutti gli animali, è chemiorganotrofa e ottiene l'ATP da reazioni di ossidoriduzione. Gli organismi superiori usano a tal fine substrati organici complessi quali donatori di elettroni e fonti di carbonio, e l'ossigeno come accettore di elettroni. Si comportano allo stesso modo anche batteri comuni come Bacillus e Pseudomonas, ma molti batteri chemioorganotrofi utilizzano nitrati o solfati come accettori di elettroni.
Infine esistono i 'chemiolitotrofi', batteri il cui metabolismo si svolge nella completa assenza di luce e di materia organica: tutto ciò di cui essi hanno bisogno è la presenza di anidride carbonica (CO2) e di un accettore di elettroni (soprattutto l'ossigeno, ma anche nitrati e CO2), insieme a un elemento ossidabile (idrogeno o ferro) o a composti inorganici (acido solfidrico, ammoniaca, nitrati). I batteri nitrificanti che ossidano l'ammoniaca a nitriti e i nitriti a nitrati, i solfobatteri che ossidano i composti solforati e gli archeobatteri metanogeni sono i chemiolitotrofi più importanti della biosfera. Il loro metabolismo risulta indispensabile per il funzionamento continuo dei cicli biogeochimici del carbonio, dell'azoto e dello zolfo.
Forse i chemiolitotrofi più insoliti sono i batteri che vivono in condizioni estreme, in prossimità delle sorgenti idrotermali dei fondali oceanici, e danno origine a ecosistemi bizzarri. Queste sorgenti furono scoperte per la prima volta nel 1977 vicino alle Isole Galápagos; le immersioni successive con batiscafi rivelarono la presenza di emissioni sottomarine in cui le temperature massime dell'acqua erano comprese tra 270 e 380 °C. L'acido solfidrico emesso dalle sorgenti è velenoso per tutti gli organismi superiori, ma i batteri chemioautotrofi lo ossidano facendolo diventare solfato, in una reazione che rilascia energia utile alla loro crescita e forma la base di piramidi trofiche piuttosto complesse, comprendenti vermi tubicoli giganti, granchi e bivalvi. Le sorgenti abissali inoltre consentono la vita dei batteri metanogeni anaerobi, come il Methanococcus jannaschii, il primo archeobatterio di cui nel 1996 è stato sequenziato completamente il genoma e che presenta una temperatura di crescita ottimale a 85 °C.
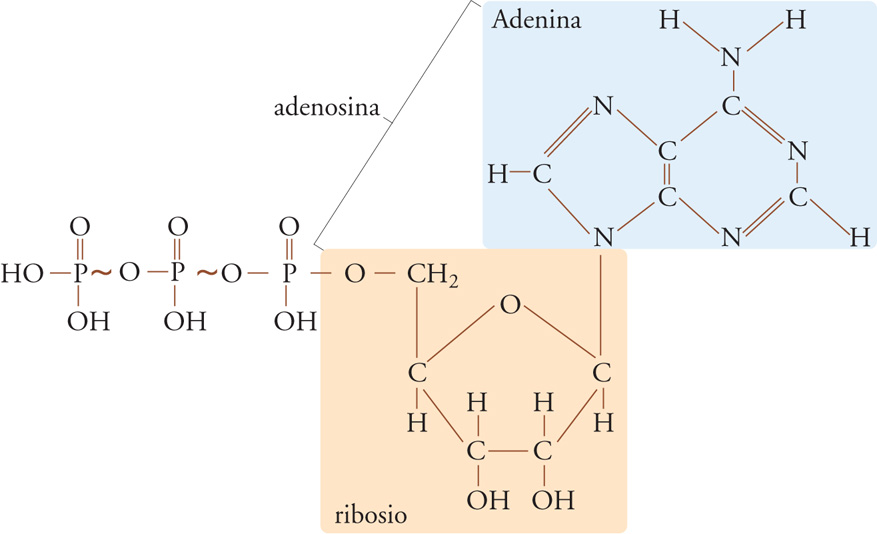
Dal punto di vista energetico c'è una caratteristica chiave condivisa da tutte le forme di vita: l'utilizzazione dell'ATP (che si forma dalla scomposizione del glucosio) per immagazzinare l'energia ricavata dalla scomposizione dei substrati nutritivi (catabolismo cellulare) e impiegarla nella biosintesi dei composti complessi (anabolismo cellulare), nella contrazione muscolare (locomozione) e nel trasporto attivo dei metaboliti (fig. 2). La produzione e la degradazione continua di ATP nelle cellule viventi mostrano un bilancio sorprendente se si considerano le masse totali. Gli esseri umani sintetizzano e degradano circa 3 g di ATP per grammo di peso corporeo secco, producendo e usando giornalmente una quantità di fosfato superiore al proprio peso corporeo. Questo turnover è meno impressionante se paragonato alla respirazione batterica, che può produrre fino a '7 kg di ATP' per grammo di massa batterica secca. L'elevata intensità del metabolismo eterotrofo fa sì che gli organismi viventi superino il Sole in produzione di potenza per unità di massa.
La fotosintesi
Il processo che fornisce energia per la produzione di molecole organiche complesse a partire da composti inorganici semplici e ampiamente disponibili (acqua, anidride carbonica, macro- e micronutrienti) è la conversione energetica più importante che si verifichi nella biosfera. I microrganismi primitivi dipendenti dall'energia idrotermale potevano sostenere soltanto tassi metabolici estremamente bassi. Attingendo alle riserve di idrogeno virtualmente illimitate contenute nell'acqua, l'evoluzione della fotosintesi, che scinde la molecola d'acqua producendo ossigeno, ha reso possibile la fissazione di più di 100 miliardi di tonnellate di carbonio all'anno. Tuttavia, la produzione di nuova fitomassa dipende in modo cruciale dalla presenza di macro- e micronutrienti: è questa nuova massa vegetale che contiene le sostanze nutritive fondamentali necessarie per tutti gli erbivori, ossia zuccheri complessi, acidi grassi e proteine.
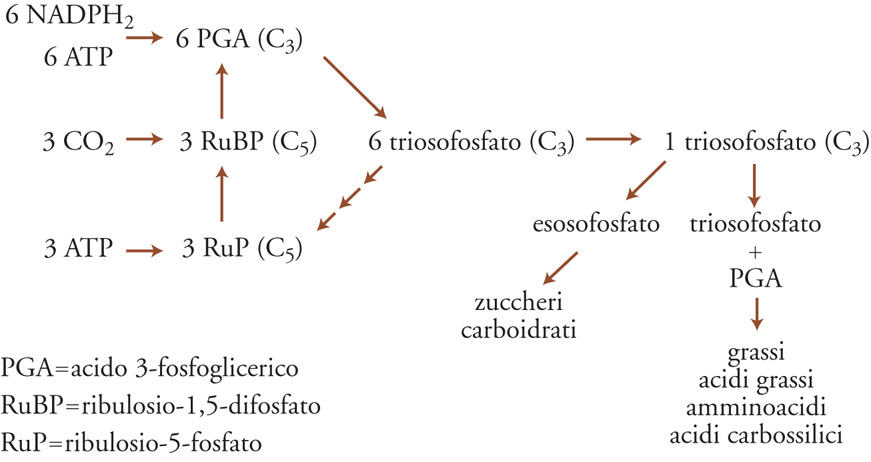
Il processo comincia con l'assorbimento di radiazione solare tramite pigmenti sensibili alla luce (principalmente clorofille) di lunghezza d'onda compresa in due bande ristrette, una tra i 420 e i 450 nm (luce blu), l'altra tra 630 e 690 nm (luce rossa). Ciò significa che nella PAR (Photosynthetic active radiation) è compresa appena meno della metà della radiazione solare totale. La sintesi di fitomassa nella cosiddetta 'via del pentoso fosfato', una via metabolica costituita da molti passaggi enzimatici di carbossilazione, riduzione e rigenerazione, deve essere preceduta dalla formazione di ATP e NADPH (nicotinammideadenindinucleotidefosfato), i due composti che forniscono energia a tutte le reazioni biosintetiche (fig. 3). Il carbonio, ottenuto con la riduzione della CO2, si combina con l'idrogeno dell'acqua e con i micronutrienti per produrre nuova fitomassa. Assumendo che la PAR rappresenti grosso modo il 45% della luce diretta, l'efficienza complessiva della fotosintesi sarebbe di circa il 12%, ma nessuna pianta si avvicina a questo massimo teorico. Tale percentuale varia in modo significativo, a seconda degli ambienti e della disponibilità di nutrienti (soprattutto l'azoto) e di acqua.
La maggior parte della nuova fitomassa viene immagazzinata nei monosaccaridi e nei loro polimeri, in particolare nella cellulosa e nella lignina (un polimero complesso derivante dalla polimerizzazione ossidativa di tre composti fenolici); lipidi a contenuto energetico più elevato sono di solito presenti soltanto nei semi.
Meno di un decennio dopo che Andrew Benson e Calvin chiarirono la sequenza di base del ciclo RPP (Reductive pentose phosphate) della fotosintesi, Hal Hatch e Roger Slack scoprirono che molti vegetali (per es., il grano e la canna da zucchero) all'inizio seguono una via diversa di fissazione del carbonio: a differenza del ciclo RPP, in cui la riduzione dell'anidride carbonica produce acido fosfoglicerico ‒ un composto a tre atomi di carbonio ‒, queste piante sintetizzano ossalacetato (un acido a quattro atomi di carbonio) nelle cellule del mesofillo. L'ossalacetato viene prima ridotto a malato ‒ un altro composto a quattro atomi di carbonio ‒ e quindi spostato nelle cellule della guaina del fascio, dove è rigenerata la CO2, e il piruvato così originato ritorna al mesofillo. La CO2 rigenerata entra quindi nel ciclo di Calvin e il suo carbonio viene incorporato nella nuova fitomassa. Queste piante utilizzano l'acqua in modo molto più efficiente, e grazie all'assenza di fotorespirazione hanno un'efficienza sintetica sostanzialmente più elevata. Inoltre, hanno un vantaggio competitivo nei climi assolati e caldi, svolgendo in modo ottimale le loro funzioni tra i 30 e i 45 °C.
La produzione primaria lorda annua delle piante terrestri è nell'ordine dei 100 miliardi di t di carbonio e, nei migliori modelli disponibili di produzione primaria netta, l'intervallo dei valori possibili è compreso tra 45 e 60 miliardi di t, con la maggior parte della fotosintesi attribuibile alle foreste tropicali e temperate. Le stime globali della produzione primaria netta negli oceani sono quasi identiche (35-50 miliardi di t) ma, a causa delle piccole dimensioni del fitoplancton e della sua breve vita, la fitomassa fissa dell'oceano è soltanto una piccolissima frazione di quella terrestre.
Mentre sulle terre emerse la maggior parte della produttività e dell'accumulo di biomassa si concentra nei grandi alberi, i produttori principali degli oceani sono rappresentati da minuscoli organismi unicellulari in balia delle correnti. In un ambito di dimensioni comprese tra meno di 2 e 200 μm di diametro sono incluse percentuali variabili di archeobatteri, eubatteri, cianobatteri e protisti eucarioti. Il cianobatterio unicellulare Prochlorococcus, scoperto soltanto nel 1988, è il più piccolo e insieme il più abbondante organismo fotosintetico che popoli gli oceani del globo, dove il suo contributo alla produzione primaria netta totale nelle acque povere di nutrienti è compreso tra il 30 e l'80%. Inoltre, mentre la lunghezza della vita media di tutte le piante terrestri è di circa 10 anni (o molti decenni per la maggior parte delle specie arboree), le cellule del fitoplancton vivono soltanto da uno a cinque giorni.
Poiché i nutrienti affondano costantemente nelle profondità al di sotto della zona eufotica (lo strato più superficiale degli oceani, in cui penetra la radiazione solare e che nelle acque più chiare può avere uno spessore superiore ai 100 m), la fotosintesi in ambiente marino è spesso limitata dalla disponibilità di azoto; spesso vi è anche carenza di fosforo e di altri micronutrienti chiave, soprattutto ferro e silicio. Questo è il motivo per cui la superficie degli oceani aperti e gli strati d'acqua immediatamente sottostanti sono tra gli ambienti meno produttivi del pianeta. Elevati rapporti tra superficie e volume e il lento affondamento delle cellule fitoplanctoniche attraverso lo strato eufotico sono ovvi adattamenti per massimizzare i tassi di assorbimento dei nutrienti. La produttività del fitoplancton può innalzarsi a valori molto elevati soltanto dove l'upwelling (la risalita dalle acque oceaniche di profondità) arricchisce gli strati superficiali importando acqua carica di nutrienti (al largo delle coste occidentali delle Americhe, dell'Africa, dell'India e intorno all'Antartide). Altre zone altamente produttive si associano ad acque costiere poco profonde, in particolare dove esse sono arricchite dai deflussi di nutrienti di origine continentale.
Il metabolismo animale
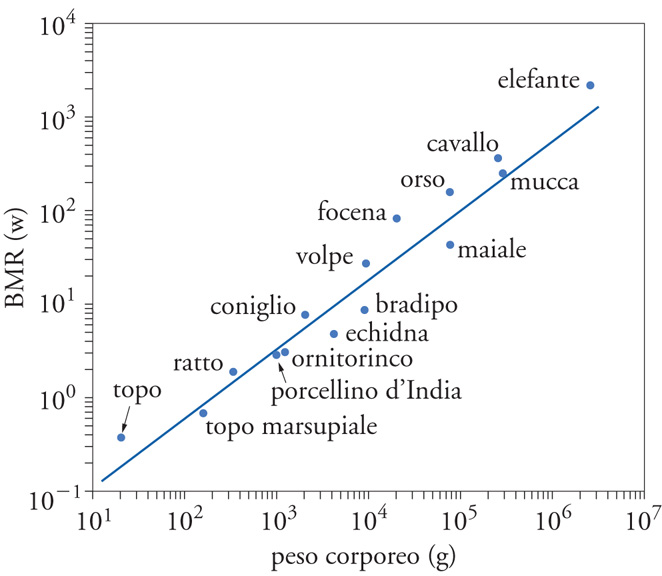
A dispetto della straordinaria diversità di dimensioni, forme, nicchie, modalità di alimentazione e comportamenti, tutti i Vertebrati e gli invertebrati, cioè tutti gli organismi che comunemente definiamo animali, condividono la stessa tipologia fondamentale di metabolismo. L'universalità delle proprietà chiave del metabolismo animale fu dimostrata per la prima volta nel 1932 da Max Kleiber nella forma di un semplice grafico su doppia scala logaritmica in cui il peso corporeo di diversi Mammiferi era posto in funzione del loro BMR (Basal metabolic rate) (fig. 4).
Il BMR misura il dispendio energetico a riposo in uno stato successivo ai processi di assorbimento (la digestione aumenta il metabolismo) e in ambienti termicamente neutri e, quindi, veicola informazioni fondamentali sulle necessità nutrizionali degli animali, rendendo possibili affascinanti comparazioni intra- e interspecifiche. Kleiber era giunto alla conclusione che il BMR di animali diversi dipendesse dalla loro massa. All'insieme originario di pochi dati, che andavano dai ratti ai bovini, vennero aggiunti i calcoli riguardanti altre specie e il grafico fu esteso a un ambito di pesi corporei compresi tra quello del topo e quello dell'elefante. Questi studi confermarono l'esistenza di una relazione costante tra massa e metabolismo basale, per cui quest'ultimo è proporzionale alla massa dell'organismo elevata alla potenza 0,75. La generalizzazione di Kleiber è divenuta una delle più importanti e note leggi della bioenergetica.
Sono oggi disponibili centinaia di BMR per specie che vanno dagli organismi unicellulari alle balene e tutti, attraverso diciotto ordini di grandezza, confermano la legge di Kleiber. Esistono comunque variazioni che escono da questa norma e che sono legate in genere a diversi adattamenti all'ambiente. Per esempio, al fine di termoregolare i loro corpi nelle acque fredde, le foche e le balene hanno un BMR pari al doppio di quello degli altri animali della loro taglia, mentre i BMR specificatamente bassi dei Mammiferi deserticoli riflettono un adattamento alle periodiche carenze di cibo e alla ricorrente o cronica scarsità d'acqua. E se non sorprende che i bradipi abbiano valori di BMR inferiori rispetto a quelli indicati dai loro pesi corporei, i maiali si discostano dai valori attesi ancora di più e, di conseguenza, diventano efficienti produttori di carne.
L'origine della legge di Kleiber non è ancora stata chiarita. Le spiegazioni proposte vanno dalla considerazione delle richieste meccaniche dei corpi animali ai vincoli geometrici e fisici della rete di vasi di cui necessitano i corpi viventi per distribuire le risorse e rimuovere le sostanze di rifiuto. Le loro proprietà richiedono che il metabolismo dell'intero organismo sia proporzionale alla massa dell'organismo elevata alla potenza 0,75; la legge di Kleiber sarebbe applicabile quindi a tutte le forme di vita. Tuttavia altri scienziati sono giunti alla conclusione che questo pattern di proporzionalità, riscontrabile trasversalmente in tutte le specie, sia un semplice sottoprodotto della selezione evolutiva che modella le dimensioni corporee all'interno delle specie. La ricerca per dare una spiegazione alla legge del coefficiente 0,75 continua.
La termoregolazione negli animali
Gli animali condividono anche due limitazioni energetiche fondamentali. Innanzitutto, in quanto eterotrofi, devono nutrirsi di biomassa contenente tutte le sostanze nutritive essenziali per la crescita e la riproduzione e perciò sono o esclusivamente erbivori oppure carnivori od onnivori in vario grado. In secondo luogo, i limiti termici di tutta la vita macroscopica sono piuttosto ristretti perché le proteine corporee iniziano a denaturarsi quando la temperatura si innalza al di sopra dei 45 °C, mentre al di sotto di 0 °C l'acqua intracellulare congela e i cristalli di ghiaccio rompono le cellule. A questo punto, si pongono due quesiti importanti: in che modo gli animali riescano a sopravvivere in ambienti che vanno dai ghiacci polari battuti dal vento con minime invernali di −70 °C ai deserti subtropicali con temperature massime giornaliere di 50 °C; e come gli eterotrofi affrontino le oscillazioni giornaliere della temperatura che possono superare i 30 °C e che normalmente sono comprese tra 10 e 15°C.
Ovviamente gli animali non hanno la possibilità di sopravvivere alle temperature estreme, come fanno molti microrganismi che sono in grado di entrare, dopo una quasi completa disidratazione, in uno stato di vita latente, simile alla morte, non soltanto per settimane ma anche per anni, decenni e oltre. Uno dei fatti più affascinanti della bioenergetica è che l'evoluzione ha prodotto due risposte molto diverse alla sfida termica, entrambe di grande successo nella loro specificità. Tutti gli ectotermi, cioè tutti gli invertebrati (che per numero sono dominati ampiamente dagli artropodi, soprattutto insetti), i pesci, gli anfibi e i rettili regolano la loro temperatura corporea passivamente, scegliendo i microambienti opportuni. Le salamandre vivono bene con temperature di appena 10 °C, mentre le lucertole deserticole preferiscono temperature di 35-40 °C.
Gli organismi di questo tipo non necessitano di un buon isolamento del corpo e possono farcela anche con un metabolismo basale basso. La combinazione di questi due fattori però indica che la dimensione del corpo è il determinante critico della loro temperatura corporea e quindi dei loro periodi di attività. Gli ectotermi di maggiori dimensioni (anaconde, coccodrilli, varani di Komodo) possono mantenere la temperatura corporea all'interno di un ambito di valori relativamente ristretto e la loro considerevole inerzia termica fa sì che possano avere periodi più lunghi di attività giornaliera, mentre la loro massa corporea rende possibili ampi intervalli tra un pasto e il successivo. Per contro i minuscoli insetti non possono innalzare la loro temperatura al di sopra di quella ambientale pur essendo in grado di farla variare abbastanza facilmente muovendosi; se vogliono restare attivi, però, devono mangiare molto spesso.
Alcuni ectotermi, tuttavia, hanno modalità che permettono loro di scendere al di sotto dei limiti di temperatura di cui abbiamo parlato o di superarli. Alcuni insetti possono diventare veri endotermi per brevi periodi prima di iniziare il volo, quando riscaldano i muscoli con il meccanismo del brivido. Alcune falene notturne possono raggiungere una temperatura toracica di 30 °C anche a temperature esterne vicine al punto di congelamento, ma un simile risultato richiede una tale quantità di energia che questi insetti devono trascorrere più del 90% del tempo restando inattivi sotto strati isolanti di foglie e neve. Alcuni pesci antartici sopravvivono nelle acque più fredde della Terra sintetizzando almeno otto differenti glicopeptidi, composti proteici antigelo che si adsorbono a minuscoli cristalli di ghiaccio inibendone la crescita nei tessuti corporei. Di conseguenza questi pesci possono nuotare in acque piene di ghiaccio a −2 °C. Le proteine antigelo (insieme a proteine che fanno da nuclei di cristallizzazione del ghiaccio iniziando la formazione di minuscoli cristalli extracellulari) vengono utilizzate anche da alcune tartarughe, anfibi e bruchi, che possono congelarsi conservando intatte le strutture intracellulari necessarie per la vita.
All'altro estremo dello spettro troviamo il verme Alvinella pompejana, una creatura dall'aspetto irsuto lunga 6cm, che forma colonie ai lati delle sorgenti idrotermali attive dei fondali oceanici. Recenti misurazioni sulla East Pacific Rise hanno mostrato che la temperatura media all'interno dei tubi in cui questi vermi vivono era di 69 °C, con frequenti punte massime di 81 °C. Dato che la temperatura all'apertura del tubo è in media di 22 °C, in tutta la lunghezza del piccolo corpo dell'animale si viene a creare un impressionante gradiente di 60 °C. A. pompejana è l'unico animale la cui tolleranza alle alte temperature si avvicina alla resistenza al calore mostrata dai microbi.
La grande quantità di ectotermi e la rilevante varietà delle loro nicchie provano che i loro adattamenti energetici sono stati un notevole successo dell'evoluzione. Tuttavia, i microambienti trasportabili e costanti degli endotermi (animali a sangue caldo) hanno fornito loro un forte margine competitivo negli ecosistemi favorevoli consentendone la vita anche nelle parti più inospitali della biosfera. L'endotermia, la condizione in cui la temperatura interna è mantenuta fra i 36 e i 40 °C nella maggior parte dei Mammiferi e fra i 38 e i 42 °C negli Uccelli, è un compromesso evolutivo che bilancia i benefici del mantenimento di una temperatura interna stabile su valori vicini a quelli dell'optimum biochimico con i rischi di dissipazione del calore e con i costi energetici del metabolismo e dei sistemi di isolamento termico.
Una termoregolazione a meno di 30 °C avrebbe richiesto un'energia minore ma un raffreddamento per evaporazione molto più elevato. Con una temperatura della superficie corporea spesso inferiore a quella dell'aria circostante, non ci sarebbero state perdite di calore per conduzione o convezione e le alte velocità di evaporazione avrebbero aumentato i rischi di disidratazione. Il raffreddamento causato dalla maggiore traspirazione non avrebbe funzionato con il notevole isolamento termico rappresentato dalla pelliccia e ciò avrebbe limitato la diffusione degli endotermi ai soli ambienti freddi. La minore temperatura interna, inoltre, avrebbe ridotto l'efficienza delle reazioni biochimiche. Anche in questo caso la dimensione degli organismi è un elemento rilevante: i piccoli endotermi sono più svantaggiati a causa della loro maggiore perdita di calore per convezione e della loro necessità di rapide risposte metaboliche a diminuzioni drastiche della temperatura.
Se un solo agosto freddo può essere fatale per un colibrì di 10 g, nessun animale ibernante che pesi più di 5kg ha necessità di ridurre la propria temperatura corporea più di qualche grado. Le piume e la pelliccia sono materiali isolanti eccezionali: anche a −30 °C la pelle dei Mammiferi artici ha una temperatura paragonabile a quella di un uomo con vestiti adeguati. Per gli endotermi acquatici la situazione si complica in modo particolare, in quanto la velocità di conduzione termica dell'acqua è venti volte superiore a quella dell'aria. Uno spesso strato di grasso non è sufficiente per i Mammiferi acquatici più piccoli: il loro metabolismo basale deve essere raddoppiato o triplicato rispetto a quello dei Mammiferi terrestri di taglia simile e, di conseguenza, questi animali devono anche trovare e digerire una quantità di cibo fino a tre volte superiore.
Per gli animali deserticoli affrontare il problema del calore è altrettanto difficile: essi sono esposti a una radiazione solare la cui potenza supera il loro metabolismo di un ordine di grandezza. La sudorazione e il respiro ansante costituiscono le due migliori risposte attive che richiedono entrambe un'attenta gestione dell'equilibrio idrico. Molti animali deserticoli di piccole dimensioni sopravvivono con cibo secco senza assumere acqua liquida, evacuano urina e feci concentrate e si rifugiano in buche scavate nel terreno. I cammelli si alimentano con piante rinsecchite, tollerano lunghi periodi senza bere, possono perdere fino al 40% della propria massa corporea e rapidamente riacquistare la normale idratazione bevendo l'equivalente del 30% del proprio peso corporeo in 20-30 minuti.
Il legame tra endotermia e successo evolutivo di Uccelli e Mammiferi è ovvio. Anche se disporre di un microambiente trasportabile pressoché costante ha richiesto molti adattamenti nell'alimentazione e nel comportamento per mantenere tassi metabolici molto elevati, l'endotermia ha conferito un evidente vantaggio competitivo aprendo alla colonizzazione da parte delle specie endotermiche anche le zone più inospitali della biosfera. Non vi è alcun dubbio che senza l'endotermia la nostra specie non avrebbe avuto successo; del resto per la vita umana come noi la conosciamo l'ectotermia non sarebbe stata concepibile.
Energetica umana
Come negli altri Mammiferi, il BMR dell'uomo è proporzionale alla potenza 0,75 della massa corporea: in una donna del peso di 50 kg il BMR è di poco superiore ai 60 W e la sua richiesta energetica di cibo è paragonabile all'alimentazione elettrica richiesta da una lampadina di debole potenza. Variazioni complesse dei BMR sono determinate dalla taglia corporea, dalla composizione (ossia la parte dei tessuti che metabolizza, sempre inferiore nelle donne) e dall'età. La relazione tra il BMR e il peso corporeo è ben rilevata, per entrambi i sessi e per tutte le classi di età, da semplici equazioni lineari, il cui potere predittivo è alto, però, soltanto per i bambini e gli adolescenti. Per quanto concerne gli uomini di età compresa fra i 30 e i 60 anni, il gruppo che costituisce la maggior parte della popolazione economicamente attiva, la correlazione spiega soltanto un terzo circa della varianza. Queste equazioni, basate su misurazioni condotte nell'ambito delle popolazioni occidentali, hanno la tendenza a sovrastimare il BMR delle popolazioni dei Paesi asiatici e africani a basso reddito.
I BMR sono peraltro estremamente variabili da persona a persona: un individuo può vivere con la metà dell'energia di cui necessita un altro e mantenersi lo stesso in salute. Pur mancando ancora una spiegazione esauriente di tale enorme variabilità individuale, sappiamo per certo che in termini relativi i reni sono gli organi metabolicamente più attivi, seguiti dal cuore, dal cervello e dal fegato. In termini assoluti, il fegato negli adulti e il cervello nei bambini utilizzano la parte più cospiscua del BMR (almeno un quinto). Il cervello di un neonato, che corrisponde a un decimo della massa corporea totale, richiede tipicamente più del 40% di BMR; anche negli adulti, quando i muscoli sono ben sviluppati, i quattro organi metabolicamente più attivi gravano per i due terzi sul BMR.
È noto inoltre l'andamento tipico del BMR durante il corso della vita: comincia con un breve periodo in cui si mantiene a un valore massimo compreso tra 2,3 W/kg circa (rispetto al peso corporeo) alla nascita e 2,7 W/kg circa nei tre-sei mesi successivi, segue quindi un rapido decremento fino a circa la metà del valore massimo al diciottesimo anno di età, prima di stabilizzarsi nuovamente per i successivi quattro decenni. Il declino riprende intorno all'età di 60 anni per arrivare intorno a 1 W/kg a 70 anni. Il BMR non va confuso con le necessità minime per la sopravvivenza, poiché la risposta metabolica al cibo e l'energia necessaria per mantenere l'igiene personale basilare lo aumentano anche nelle persone sedentarie di almeno un 15-20%.
L'adattamento energetico più notevole dell'uomo è forse la risposta al calore. L'organismo umano risponde anche al freddo, tramite la vasocostrizione: per esempio, gli aborigeni australiani che vivevano nudi potevano dormire senza elevare il loro BMR riducendo l'afflusso sanguigno nella pelle e nelle estremità. Tale adattamento naturalmente è inadeguato nei climi freddi, dove si deve far ricorso a soluzioni artificiali (vestiti, ripari, fuoco). Ovviamente si cerca riparo anche nei climi caldi e ci si veste in maniera adeguata (oggi sempre di più cercando rifugio in luoghi dotati di aria condizionata), ma si fa affidamento anche sugli aggiustamenti particolarmente efficaci della nostra termoregolazione.
Le risposte iniziali al calore prevedono la dilatazione dei vasi sanguigni periferici e lo spostamento di altro sangue dalle vene interne a quelle più superficiali. Quindi, quando la temperatura della pelle si avvicina ai 35 °C, di solito iniziamo a sudare e, quando svolgiamo un lavoro pesante, questa risposta può essere considerevolmente più efficace che in altre specie di mammiferi. Un cavallo può traspirare a una velocità di 100 g/m2 all'ora e un cammello a 250 g/m2, mentre un uomo può arrivare a una media di 500 g/m2 all'ora. Senza sudare una persona perderebbe in media 12 W per m2 di superficie corporea; all'incirca la stessa quantità si perde con la respirazione e la diffusione attraverso la pelle (in totale appena 20 W/m2). Negli adulti invece la traspirazione di 500 g/m2 all'ora equivale a una perdita di calore compresa addirittura tra 550 e 625 W.
Gli individui con migliore acclimatazione possono traspirare fino a 1100 g/m2 per ora, equivalenti a 1390 W. Ciò è sufficiente per prevenire pericolosi aumenti della temperatura interna del corpo anche in individui sottoposti a lavoro intenso, ma in questo caso è necessaria un'adeguata reidratazione. Una parziale disidratazione temporanea è comune durante il lavoro intenso o le lunghe corse e non causa problemi finché il deficit idrico si risolve entro il giorno successivo. La capacità di sudare molto è codificata geneticamente ed è conservata anche dalle popolazioni che vivono da millenni in climi più freddi. Gli Europei, la cui risposta iniziale al calore elevato è un pericoloso innalzamento della temperatura interna e del battito cardiaco prossimo al massimo tollerabile, riescono a uguagliare la velocità di sudorazione di popolazioni ben acclimatate native dei tropici nel giro di soli 10 giorni. La nostra abilità di far fronte al surriscaldamento sudando deve essere vista, insieme all'andatura bipede, alla mancanza di una pelliccia, al grande cervello e alla capacità di utilizzare un linguaggio simbolico, come uno dei tratti chiave che definiscono la nostra specie.
Bibliografia
Brown, West 2000: Scaling in biology, edited by James H. Brown and Geoffroy B. West, Oxford, Oxford University Press, 2000.
Calvin 1989: Calvin, Melvin, Forty years of photosynthesis and related activities, "Photosynthesis research", 21, 1989, pp. 3-16.
Garby, Larsen 1995: Garby, Lars - Larsen, Poul S., Bioenergetics: its thermodynamic foundation, New York, Cambridge University Press, 1995.
Kleiber 1961: Kleiber, Max, The fire of life, New York, Wiley, 1961.
Lowen 1994: Lowen, Alexander, Bioenergetics, New York, Penguin/Arkana, 1994.
Nicholls, Ferguson 2002: Nicholls, David G. - Ferguson, Stuart J., Bioenergetics, San Diego, Academic Press, 2002.
Smil 1991: Smil, Vaclav, General energetics: energy in the biosphere and civilization, New York, Wiley, 1991.
Smil 2002: Smil, Vaclav, The earth's biosphere: evolution, dynamics, and change, Cambridge (Mass.), MIT Press, 2002.