
Biotecnologie
Biotecnologie
Con l’avvento del ‘Progetto genoma’, da un lato, e lo sviluppo delle metodiche di manipolazione embrionale, dall’altro, le biotecnologie sono entrate prepotentemente nella vita quotidiana di ciascuno di noi. Il nostro genoma viene visto come un pacchetto di programmi in cui sono contenute tutte le istruzioni per costruire il nostro corpo, determinarne il funzionamento e persino modellare la nostra personalità. La disponibilità di tecniche di sequenziamento rapido e a basso costo consentirà, entro breve tempo, di sequenziare l’intero genoma di singoli individui, aprendo così la possibilità di prevedere non solo le malattie che ci colpiranno ma anche, secondo alcuni, di indagare aspetti più profondi della nostra psiche. In realtà, oggi queste prospettive valgono solo per le malattie genetiche che hanno come responsabile un solo gene, per le quali cioè è possibile una diagnosi precisa e una prognosi esatta, mentre per le malattie più complesse, nelle quali sono coinvolti numerosi geni e altrettanti fattori ambientali, siamo ancora al livello di speranze che iniziano adesso a concretizzarsi. Per quanto riguarda l’influenza dei geni sui tratti legati alla personalità e al comportamento morale, non sono ancora stati ottenuti dati certi e solo in futuro si potrà dire se è predominante la componente genetica oppure quella ambientale. Purtroppo gli importanti risultati diagnostici non hanno ancora avuto ricadute terapeutiche. Il limite delle biotecnologie genomiche è proprio questo: siamo in grado di dire chi si ammalerà, ma il più delle volte non possiamo guarire il paziente. È lecito chiedersi a cosa serva essere in grado di prevedere il futuro (sanitario) di una persona se poi non è possibile cambiare l’esito infausto di molte malattie. Da questo punto di vista le biotecnologie sono state finora deludenti, ma è auspicabile, nonché verosimile, che questa situazione possa cambiare in futuro.
Nei trent’anni che ci separano dalla nascita della prima bambina ‘in provetta’ il settore dell’embriologia umana si è ampliato enormemente fino a portare alla produzione delle cellule staminali embrionali e alle possibilità del trasferimento nucleare e della riprogrammazione del genoma cellulare. Questi studi sull’uomo procedono però con grande lentezza, per l’esistenza di precisi limiti di natura etica considerati insuperabili. Tuttavia, la conoscenza sempre più approfondita del nostro DNA spinge a chiedersi se sia lecito offrire per mezzo delle biotecnologie la possibilità di modificare il genoma degli organismi viventi, incluso l’uomo. L’incontro tra le tecnologie basate sull’ingegneria genetica e quelle che consentono la manipolazione embrionale negli animali da esperimento (in particolare nel topo) si è rivelato estremamente proficuo. Nel topo è possibile modificare con precisione geni specifici: questo apre la strada alla possibilità della correzione di difetti genetici, se il gene in oggetto è responsabile di una malattia, o addirittura al miglioramento della specie. Tecnologie che consentano di modificare il genoma di Homo sapiens in maniera eticamente corretta non sono attualmente disponibili, ma potrebbero esserlo in un futuro relativamente lontano. Nel frattempo, tuttavia, queste tecnologie potrebbero rivelarsi utili per nuove forme di terapia basata sulla combinazione di metodiche di ingegneria genetica e di biologia cellulare. La complessità della materia e la rapidità con cui le biotecnologie evolvono rendono tuttavia difficile un giudizio sereno su di esse. Diventa quindi necessario un ampio dibattito sulla loro utilizzazione.
sommario
1. Biotecnologie del genoma. 2. Biotecnologie della manipolazione embrionale. 3. Cellule staminali e clonazione. 4. Biotecnologie della riproduzione umana. 5. Il futuro della specie Homo sapiens. □ Bibliografia.
Biotecnologie del genoma
Spesso le biotecnologie vengono considerate solo sotto l’aspetto delle loro possibili applicazioni industriali e commerciali, in quanto vengono circoscritte alle loro numerose ricadute pratiche. In realtà, le biotecnologie sono innanzitutto tecniche che permettono di conoscere meglio i fenomeni della vita e, in quanto tali, fuoriescono dallo stretto ambito degli specialisti per interessare un pubblico sempre più vasto. Storicamente, è stato il dibattito sulle possibilità di intervento sui viventi, aperte dall’ingegneria genetica, a originare nell’opinione pubblica un sentimento di sospetto di cui l’economista statunitense Jeremy Rifkin è il rappresentante più conosciuto; anche se, per la verità, sin dagli inizi, le applicazioni pratiche delle biotecnologie sono state enormemente positive, come nel caso della produzione di insulina umana (insulina ricombinante). Ma è stato con il Progetto genoma umano che il dibattito sulle biotecnologie e sull’opportunità di una loro utilizzazione ha assunto una diffusione e un interesse planetari.
Il Progetto genoma, infatti, si proponeva di decifrare nei dettagli il manuale di istruzioni in grado di costruire l’organismo umano. In termini tecnici, questo significa determinare la sequenza dei nucleotidi (o basi) che costituiscono il nostro DNA. Tuttavia la strada da percorrere per la completa comprensione di tutte le istruzioni contenute in questo manuale non è affatto terminata. In primo luogo, la lingua in cui è scritto ci è nota solo in parte: infatti, mentre alcune piccole sezioni del DNA sono scritte secondo un codice che conosciamo, altre sono ancora incomprensibili. Inoltre, bisogna tener conto del fatto che, nel caso dell’uomo, il DNA deve contenere tutte le istruzioni perché da una singola cellula si possano produrre molti miliardi di cellule che abbiano tra loro rapporti ben precisi, tali da formare tutti gli organi del corpo: la nostra vita comincia da quando il nucleo dell’uovo si fonde con quello dello spermatozoo a formare un unico genoma e, quindi, uno specifico manuale di istruzioni. Da quel momento in poi, la cellula iniziale comincia a dividersi e a moltiplicarsi secondo il codice presente nel DNA stesso. Il fatto che il DNA della farfalla sia diverso da quello dell’elefante spiega perché nascano organismi differenti. Da qui si comprende anche come possano avere origine le malattie genetiche. Esse, infatti, insorgono quando un’istruzione è sbagliata, incompleta o inesatta: una piccola molecola dei 3 miliardi che compongono il DNA viene copiata in maniera errata e il significato dell’istruzione non è più comprensibile.
Biotecnologie sanitarie
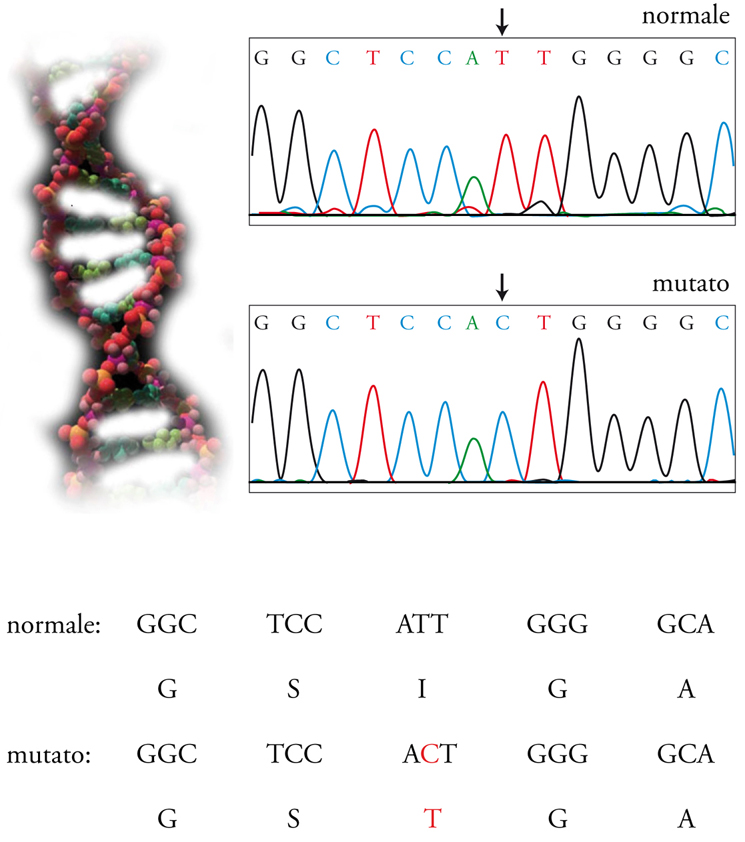
La diagnosi delle malattie ereditarie è una delle ricadute più eclatanti delle biotecnologie del genoma. Oggi sappiamo qual è il gene difettoso nelle malattie genetiche più comuni, come la fibrosi cistica, la sindrome dell’X fragile, la talassemia e così via. Il numero di geni che compongono il DNA è attualmente fissato in poco più di 20.000, ma bisogna notare che, a differenza di quanto si credeva fino ad alcuni anni fa, i geni in senso classico contribuiscono solo in parte al funzionamento del genoma. Questo dato spiega come siano conosciute più di 5000 malattie genetiche. Ancora non è noto il gene responsabile di ciascuna, ma attualmente possiamo fare una diagnosi estremamente precisa di quelle per cui è stato identificato (Tav. I). È possibile diagnosticare i pazienti affetti, i portatori sani e quelli che sono già malati ma spesso anche quelli che si ammaleranno; come pure è possibile formulare la diagnosi quando il feto è ancora nell’utero (diagnosi prenatale) e anche quando è formato da qualche decina di cellule e non si è ancora impiantato nell’utero (diagnosi preimpianto). Tuttavia, ai progressi diagnostici non si sono accompagnati progressi terapeutici paragonabili. La terapia genica, in cui si ponevano grandi speranze negli anni Novanta del Novecento, ha infatti prodotto sinora benefici minimi.
Biotecnologie per lo studio dell’evoluzione
Come abbiamo detto, le biotecnologie non vanno giudicate solo per le loro ricadute pratiche. Un aspetto estremamente interessante è quello della genomica comparata: confrontando genomi di specie differenti, possiamo stabilire correlazioni tra le caratteristiche della specie e le istruzioni contenute nel genoma. In tal modo si constata che più lungo è il tempo in cui due specie si sono separate e più numerose sono le differenze a livello di genomi. Un interessante corollario di questi confronti è che è possibile stabilire connessioni di tipo filogenetico, ossia ricostruire alberi evolutivi delle varie specie e mostrarne le origini comuni. Inoltre si può anche cercare di scoprire quali sono le mutazioni responsabili delle differenze tra due specie, come nel caso di Homo sapiens e di Pan troglodytes (lo scimpanzé comune), che viene ritenuta la specie vivente a noi più vicina: il nostro ultimo antenato comune, infatti, è vissuto circa 6-7 milioni di anni fa. Attualmente, visto che anche il genoma dello scimpanzé è stato sequenziato, è possibile confrontarlo con quello umano e stabilire quali sono le differenze: alcune di queste sono certamente responsabili della divergenza che ha portato all’origine delle due specie distinte. Il problema è capire quali sono queste differenze, perché se fossimo diversi anche solo per l’1%, questo significherebbe che abbiamo circa 30 milioni di nucleotidi differenti. Non si tratta, quindi, di un compito semplice, ma la ricerca in questo senso è molto attiva. Per non parlare della possibilità di sequenziare genomi di specie scomparse, come il Mammut (Mammuthus primigenius) e, soprattutto, l’Homo neanderthalensis: entrambe queste specie sono scomparse da un tempo relativamente breve (qualche decina di migliaia di anni) pertanto il loro DNA si è parzialmente conservato, soprattutto nelle zone fredde del pianeta. Si tratta di ricadute forse prive di interesse economico ma certamente di enorme impatto culturale.
Biotecnologie della manipolazione embrionale
Lo studio del genoma, dopo un breve periodo di diffidenza, è stato riconosciuto come un’interessante possibilità per la salute umana. È bene notare come le preoccupazioni iniziali siano state fugate e attualmente si sia raggiunto un certo consenso sul fatto che in esso non vi siano grandi rischi. In realtà, tale studio può avere implicazioni per la privacy dell’individuo, ma due millenni di pratica medica, nonché leggi recenti, fanno sì che questi sia protetto formalmente nella sua sfera privata. A testimoniare questa presa di coscienza, basta ricordare che la legge sulla discriminazione genetica, dopo un’attesa di oltre vent’anni, è stata approvata dalla Camera degli Stati Uniti nel 2008 con una larga maggioranza.
La situazione cambia sensibilmente se si considerano le tecnologie attualmente disponibili per la manipolazione dei gameti e degli embrioni. In questo caso il dissenso è molto più ampio. Sebbene sia probabile che in questo settore le opinioni rimarranno a lungo diverse, è opportuno che potenzialità e limiti di queste tecnologie siano ben chiare.
È stato il topo il modello sperimentale di elezione non solo per i Mammiferi, ma per i Vertebrati in generale, per studiare la riproduzione e lo sviluppo embrionale. Per queste ricerche, avere a disposizione un mammifero è importante e il topo ha in sé numerosi vantaggi: piccole dimensioni, bassi costi, breve tempo di riproduzione e così via. Per questo, molte tecnologie di manipolazione dell’embrione sono state inizialmente impiegate sul topo.
Topi chimerici
Un topo chimerico è un topo formato da cellule che hanno due genomi differenti. Come sappiamo, a eccezione dei gemelli monozigoti, ogni individuo ha un genoma unico. Gli embriologi constatarono che se due embrioni ai primissimi stadi di sviluppo venivano accostati l’uno all’altro (dopo aver eliminato la loro membrana protettiva detta zona pellucida), essi erano in grado di fondersi e di dare origine a un solo embrione, formato da cellule di entrambi gli embrioni originari. Esso, quindi, possiede cellule con due genomi diversi. In omaggio alla chimeramitologica, tali organismi sono stati chiamati chimere intraspecifiche. È anche possibile ottenere chimere interspecifiche se i due embrioni sono di specie differenti, ma comunque abbastanza vicine filogeneticamente.
Tali organismi sono stati utilizzati per vari studi, e in seguito si sono rivelati importanti per la produzione degli animali transgenici detti animali knock-out.
Topi transgenici
I topi transgenici sono topi in cui un gene estraneo è stato introdotto mediante iniezione in un embrione allo stadio di una cellula. I primi esperimenti di ingegneria genetica avevano mostrato che un gene poteva essere inserito in un batterio e venir prodotto da esso: in tal modo si era ottenuta l’insulina umana, una pietra miliare nella storia delle biotecnologie. Questo risultato aveva suscitato una grande eccitazione e aveva fatto pensare che fosse facile inserire un gene in una cellula di mammifero sia per scopi di ricerca sia per scopi medici (in questo secondo caso si parla di terapia genica). In realtà, i progressi furono assai più lenti di quanto si fosse sperato.
Tuttavia, vi fu un settore in cui le cose funzionarono al di là delle più rosee previsioni. Alcuni embriologi si chiesero se fosse possibile introdurre un gene in un embrione allo stadio di una cellula. L’esperimento sembrava abbastanza difficile per vari motivi, tra cui il fatto che le tecniche per realizzare tale introduzione, oltre a essere poco efficienti, non erano adatte per essere applicate agli embrioni. Si pensò, così, di iniettare una piccolissima quantità di DNA in soluzione direttamente nel nucleo di un embrione tramite un sottile capillare di vetro che bucava l’embrione. Sembrava piuttosto improbabile che l’esperimento potesse funzionare perché, anche senza tener conto del danneggiamento dell’embrione, il DNA avrebbe dovuto integrarsi nel suo genoma, per di più in un momento delicatissimo per il futuro sviluppo. Invece funzionò: il gene venne integrato nel genoma della cellula ricevente e mantenuto durante tutte le divisioni cellulari, produsse la proteina pertinente e, soprattutto, fu trasmesso alla progenie. In altre parole, il gene veniva considerato come un gene endogeno.
L’importanza di questi animali per gli studi biologici è stata enorme e ha aperto la strada alla produzione di ulteriori modificazioni del genoma.
Cellule staminali embrionali di topo
Le cellule che formano gli embrioni ai primi stadi di sviluppo vengono chiamate blastomeri. Negli anni Settanta del secolo scorso, queste cellule furono al centro di molti studi di embriologia sperimentale. Tra l’altro, si scoprì che l’uso di blastomeri forniva un altro metodo per ottenere chimere: infatti, se blastomeri isolati singolarmente da una blastocisti (donatrice) venivano introdotti in una blastocisti integra (ricevente), essi venivano incorporati da quest’ultima e contribuivano così alla formazione dell’intero organismo. Nel tentativo di coltivare in provetta tali cellule, si erano ottenute linee tumorali che, se iniettate nel topo, avevano la particolare proprietà di produrre tumori che contenevano cellule di vari tessuti, cioè di formare teratomi. Si tratta di tumori, presenti anche nell’uomo, che hanno origine da cellule molto indifferenziate, attualmente considerate cellule totipotenti, ossia l’equivalente di cellule staminali embrionali (CSE), che a loro volta sarebbero identiche ai blastomeri. Nel corso dei loro studi, alcuni embriologi iniettarono cellule di teratomi murini in blastocisti, ottenendo topi parzialmente chimerici: in alcuni casi questi avrebbero dato origine a progenie sana derivata dalle cellule donatrici cancerose. Questo risultato sorprendente è tuttavia stato contestato e attualmente non si è sicuri della sua effettiva realizzazione.
Tuttavia, da questi tentativi è chiaro che a metà degli anni Settanta, più o meno nel periodo in cui si avviava la ricerca che avrebbe portato agli animali transgenici, l’interesse per i blastomeri era molto diffuso. La disponibilità di blastomeri non tumorali coltivati in provetta avrebbe ampliato le conoscenze su questo tipo di cellule e avrebbe consentito la loro manipolazione. Nel 1981 due ricercatori riuscirono a ottenere indipendentemente linee stabilizzate di cellule che, se iniettate in una blastocisti, potevano contribuire alla formazione di tutti i tessuti dell’organismo, gameti compresi: questo indicava che le cellule erano realmente totipotenti dal momento che colonizzavano anche la linea germinale. Ne discendeva, quindi, che qualsiasi modificazione introdotta in queste cellule – per esempio l’inserimento di un gene esogeno – sarebbe stata trasmessa anche a quella parte di progenie di un animale chimerico che fosse derivato dalle cellule manipolate e reintrodotte in una blastocisti. Esse potevano fornire un altro metodo per ottenere animali transgenici.
Topi knock-out e knock-in
Tuttavia, entrambe le metodiche per ottenere animali transgenici (come pure una terza, basata su vettori virali che non ebbe grande diffusione) non consentivano di inserire un gene in un punto preciso del genoma: l’integrazione del gene avveniva in modo casuale. In alcune situazioni questo fatto costituiva una limitazione. Uno dei limiti era il fatto che il gene inserito in un punto che non corrispondeva alla sua posizione originaria, subiva delle interferenze nella sua regolazione e questo alterava la sua funzione fisiologica e quindi le conclusioni che se ne potevano trarre. Inoltre, se fosse stato possibile inserire un frammento genico in modo mirato, sarebbe stato possibile anche aggiustare un gene non funzionante: questo, nel caso dell’uomo, avrebbe significato la correzione di una malattia genetica. In effetti, proprio in quegli anni, si susseguivano continue scoperte di geni responsabili di malattie umane. Tuttavia, nel topo, per ragioni tecniche, la possibilità di riparare un gene era meno facile che non quella di distruggerlo. Che senso può avere distruggere un gene? In primo luogo, avere a disposizione un organismo in cui un gene è stato distrutto significa comprendere quale è la sua funzione: essa viene studiata ‘per difetto’. In secondo luogo, se il gene che viene distrutto nel topo è il corrispettivo di un gene che provoca una malattia genetica umana, allora quel topo diventa un modello di quella malattia umana: su quel topo possiamo effettuare esperimenti che, per motivi etici, non possono essere eseguiti nell’uomo e possiamo anche testare terapie per vedere se esse funzionano almeno nell’animale.
Ottenere questa inserzione mirata (denominata ricombinazione omologa) non è cosa facile: nel lievito, per esempio, essa è piuttosto frequente, ma nelle cellule di mammifero avviene molto raramente. Tuttavia Mario Capecchi, che studiava la ricombinazione omologa nelle cellule dei Mammiferi, comprese che, avendo a disposizione milioni di cellule staminali in provetta, il problema non era la frequenza del fenomeno, ma la sua capacità di selezionarlo. L’idea della selezione non era nuova, in quanto è alla base di tutti gli esperimenti di trasferimento genico, ma egli introdusse alcune modifiche che gli permisero di identificare le cellule (cloni) in cui la modificazione era avvenuta in maniera precisa distinguendole dalla maggioranza in cui essa era avvenuta in maniera casuale. Inserendo le cellule staminali così modificate (ne bastano pochissime) in una blastocisti normale, egli ottenne un topo chimerico, la cui progenie, essendo derivata dalle cellule modificate, era essa stessa modificata. Nascono così gli animali knock-out, ovvero topi in cui un gene predefinito è stato inattivato. Attualmente, i topi così prodotti sono migliaia ed essi si sono rivelati insostituibili nello studio di numerose malattie, tra cui il cancro. Per i suoi lavori, Mario Capecchi ha ottenuto il Nobel nel 2007.
Con lo stesso principio, grazie a ulteriori affinamenti tecnici, è stato possibile, anziché inattivare, ‘correggere’ topi con geni alterati. Mentre la produzione di topi knock-out richiede solo che una qualsiasi grossolana alterazione del gene venga introdotta, la correzione del difetto è necessariamente più fine. Tale tecnologia è denominata knock-in e non è utilizzata solo per aggiustare i geni alterati, ma soprattutto per introdurre modificazioni puntiformi in geni specifici. Di qui l’idea di utilizzarla anche nell’uomo per curare le malattie genetiche a livello di embrione. Le difficoltà sono attualmente di tipo tecnico: il passaggio attraverso un organismo chimerico nell’uomo renderebbe la cosa non realizzabile. Ma se in futuro fosse possibile controllare la correzione a livello di gamete, la cosa potrebbe diventare tecnicamente possibile: avremmo in questo caso una terapia genica ‘germinale’ in cui la correzione verrebbe tramandata alla progenie.
Cellule staminali e clonazione
Per quasi vent’anni, le cellule staminali embrionali murine sono state utilizzate soprattutto per produrre animali knock-out. Era noto da tempo che esse erano totipotenti e che potevano essere spinte a differenziarsi in vitro in vari tessuti, ma questa loro proprietà non era molto studiata. Le cose cambiarono quando, nel 1998, James A. Thomson e i suoi colleghi dell’Università del Wisconsin comunicarono di aver prodotto CSE umane.
Cellule staminali embrionali umane
La possibilità di produrre CSE umane era stata riconosciuta da tempo, ma molti ricercatori evitarono di dedicarsi ad analisi di questo tipo perché consci che ciò avrebbe sollevato problematiche etiche, visto che per ottenere linee di CSE è necessario (almeno per ora) distruggere embrioni. Inoltre, la legislazione statuniten-se non consentiva di lavorare su embrioni umani confondi governativi e, in effetti, Thomson utilizzò fondiprivati.
Nelle società occidentali, le malattie degenerative che coinvolgono organi come il midollo osseo, il fegato, il rene, il cervello, il muscolo, costituiscono un problema enorme. Esse, per ora, possono essere trattate con il trapianto (con l’esclusione del cervello), ma vi sono due grossi problemi che limitano questo approccio: la scarsità di organi e il rigetto. Le CSE sono totipotenti, pertanto possono teoricamente differenziarsi in qualsiasi tessuto: esse, quindi, potrebbero risolvere il primo problema. Naturalmente bisogna trovare le condizioni giuste per indirizzare le cellule verso un tessuto piuttosto che verso un altro e studi in provetta in questo sensosono in corso in tutto il mondo. Tuttavia, gli ostacoli sono molti poiché è necessario formare tessuti perfettamente funzionanti: per esempio, per il morbo di Parkinson – che è una tra le malattie che sembrano più facilmente aggredibili con questo approccio – è necessario ottenere cellule nervose di un tipo specifico, cioè neuroni dopaminergici; ma questa non è l’unica difficoltà. Bisogna poi reinserire tali neuroni nel posto giusto, ossia nel cervello, e fare in modo che essi possano ripristinare le connessioni che avevano i neuroni precedenti, cosa al momento assolutamente fuori dalla nostra portata, poiché non possiamo indirizzarli in alcun modo e il risultato è vincolato alla loro capacità di riuscirci da soli. Inoltre, molti organi hanno una struttura tridimensionale complessa, come il rene, che ha una struttura interna difficilissima da ricostruire ed è al di sopra delle nostre attuali possibilità. Altri tessuti come il midollo osseo, sembrano più facili da sostituire con cellule nuove. È pertanto possibile che a medio termine le CSE possano produrre risultati per alcuni organi e per alcuni tessuti, ma non per altri. Infine, ci sono gli aspetti etici, che qui possono solo venir accennati: per molti, distruggere embrioni per ottenere CSE umane è una prospettiva inaccettabile e in vari Paesi sussistono impedimenti – anche di carattere legislativo – alla ricerca in tal senso.
Cellule staminali dell’adulto
Questi impedimenti non sussistono, invece, per quelle che vengono chiamate cellule staminali adulte. Ingenere, i nostri tessuti non sono statici, ma contengono cellule che muoiono e che vengono rimpiazzate da altre cellule: questo significa che in ogni tessuto vi sono cellule staminali, in grado cioè di dare origine a tutte le altre che compongono l’organo. In effetti, da oltre 40 anni si trapiantano cellule staminali ematopoietiche in quella procedura che una volta veniva semplicemente chiamata trapianto di midollo (osseo). Attualmente, però, questo settore si è ampliato notevolmente e siamo in grado di ottenere ampie porzioni di cute o di ricreare cornee a partire dalle cellule stesse del paziente: in questi casi, le cellule staminali possono essere fornite dal paziente stesso e quindi il problema del rigetto non esiste.
Anche i tessuti dei muscoli e del cervello contengono cellule staminali che potrebbero essere utilizzate per curare malattie degenerative di questi organi, tuttavia il più delle volte le cellule del paziente non possono essere utilizzate in quanto difettose: nelle malattie ereditarie, infatti, proprio il difetto genetico è la causa delle degenerazioni. È quindi necessario usare cellule di donatori le quali solitamente devono affrontare il problema del rigetto.
Clonazione
Di tutte le biotecnologie, quelle legate alla clonazione sono certamente le più affascinanti e le più temibili. Il loro fascino è innegabile, perché l’idea di poter ricostruire un uomo o una donna perduti sembra in qualche modo realizzare il sogno dell’immortalità. Naturalmente si tratta di un’illusione, visto che ciascun individuo è determinato non solo dal proprio genoma ma anche dall’insieme di esperienze accumulate durante la vita.
La pecora ‘Dolly’, il primo mammifero clonato, è uno dei pochi esempi in cui una scoperta rilevante è stata ottentuta con l’impiego di un mammifero diverso dal topo. Venuta alla luce nel 1996 e scomparsa nel 2003 prima di raggiungere i 7 anni di età, essa ha rappresentato una sorpresa non solo per l’opinione pubblica ma anche per gli addetti ai lavori. Si sapeva che ogni cellula di un organismo contiene lo stesso DNA delle cellule embrionali, ma si riteneva che durante lo sviluppo del feto accadesse qualcosa nella conformazione del DNA e delle proteine che lo avvolgono a formare la cromatina, che impediva un percorso a ritroso. Dolly ha dimostrato che non è così e che, almeno in alcuni casi, il nucleo di una cellula adulta può essere ‘riprogrammato’, cioè può essere riportato al punto di partenza, allo stato del nucleo dello zigote, e da lì ripartire a formare un intero organismo. I meccanismi di questa riprogrammazione sono quasi completamente ignoti, ma costituiscono un settore di ricerca attivissima.
Attualmente le specie clonate sono una ventina e non vi è quindi alcun dubbio che il sistema funzioni. A questo punto è inevitabile chiedersi se la clonazione possa funzionare anche nell’uomo. In linea di principio sembrerebbe che non vi siano ostacoli tecnici; malgrado ciò, la clonazione umana non è attualmente sull’agenda di alcun ricercatore del settore, perché sono troppe le difficoltà da superare e i problemi etici da affrontare. Dolly fu l’unico successo su 277 tentativi e l’attuale efficienza della procedura, anche se leggermente migliorata, è ancora molto bassa: molte gravidanze vengono perdute e molti nati sono ‘difettosi’. Un tale risultato nell’uomo sarebbe inaccettabile. Anche se non si può escludere a priori che qualcuno possa impiantare un laboratorio di embriologia sperimentale su una piattaforma in acque extraterritoriali, questo scenario, tuttavia, è per ora alquanto improbabile.
Va segnalato, comunque, che all’inizio del 2007 è apparso un editoriale sulla rivista “Nature”, che per la prima volta considerava la possibilità che la procedura di clonazione nella nostra specie potesse essere utilizzata in casi di infertilità. Dal momento che la rivista può essere considerata come l’espressione di una larga parte della comunità scientifica, questo editoriale potrebbe rappresentare l’indizio di una svolta rispetto alla condanna della clonazione riproduttiva che prima di allora era stata praticamente unanime.
Clonazione terapeutica
Il processo di clonazione utilizzato nel caso di Dolly viene denominato ‘trasferimento di un nucleo di cellula somatica in oocita enucleato’ o più semplicemente ‘trasferimento nucleare’ (nuclear transfer). L’oocita (uovo) è una cellula molto particolare che contiene nel proprio citoplasma tutti i fattori necessari affinché il nuovo nucleo formato dalla fusione dei due pronuclei (dell’uovo stesso e dello spermatozoo) possa iniziare a replicarsi e ad attivare l’espressione di un gran numero di geni. Si ritiene che questi stessi fattori siano in grado di attivare anche il nucleo che viene in esso trasferito artificialmente mediante la procedura del nuclear transfer. Se noi conoscessimo alla perfezione questi fattori e come essi agiscono, potremmo forse riuscire a riprogrammare il nucleo senza bisogno di trasferirlo in un oocita, e questo non solo rappresenterebbe un interessante progresso tecnologico, ma eliminerebbe qualsiasi problema etico legato al sistema attuale di ottenimento delle CSE umane. Qualche risultato in questo senso è stato raggiunto recentemente e vi è oggi grande entusiasmo al riguardo.
Biotecnologie della riproduzione umana
Nell’uomo nessuna di queste tecnologie è stata utilizzata, se si fa eccezione per qualche tentativo di clonazione terapeutica. Ma nell’uomo da decenni sono diffuse pratiche di fecondazione assistita che hanno portato nel 1978 alla nascita di Louise Brown, la prima bambina concepita in provetta. Da allora si stima che circa 3 milioni di bambini siano nati mediante le tecnologie che vanno sotto il nome di fecondazione assistita. Nel caso Brown, un ovulo della madre venne asportato e messo in provetta assieme agli spermatozoi del padre: l’embrione formatosi venne reinserito nell’utero della madre; in questo caso, quindi, sia i gameti sia l’utero in cui la gravidanza ebbe luogo appartenevano ai genitori del bambino. Ma esistono anche esempi di altro tipo: gameti di donatori, uteri in affitto, mamme-nonne e così via.
Attualmente si parla di fecondazione omologa per definire i casi come quelli di Louise Brown, in cui i figli sono geneticamente figli di entrambi i genitori e la gravidanza viene portata avanti dalla madre genetica. Si parla di fecondazione eterologa quando almeno un gamete non appartiene alla coppia: generalmente la ragione per ricorrere a un donatore di gameti si trova nel fatto che uno dei due partner è sterile, ma vi sono anche casi in cui le donne ricorrono a questa pratica per motivi diversi (per es., donne single ecc.). In realtà, la fecondazione eterologa ha preceduto di parecchio la nascita di Louise Brown, perché l’inseminazione artificiale (cioè l’inoculo di sperma di donatore in una donna) è pratica più antica, per via della sua semplicità. Si parla invece di madri surrogate quando la gravidanza di un embrione viene portata avanti da una donna che non ha contribuito geneticamente all’embrione: questo può capitare quando la donna ha subito interventi di asportazione dell’utero. Per quanto riguarda la clonazione, nessun essere umano è mai stato clonato, anche se periodicamente sono state fatte circolare false notizie a questo riguardo.
Tutte queste possibilità hanno dato origine a un certo numero di controversie e talvolta si è dovuto ricorrere al giudice per dirimere delicate questioni tra le persone coinvolte. Spesso i giudici hanno dovuto emettere sentenze in settori nuovi e delicati. Attualmente la materia tende a essere maggiormente regolata che negli anni passati, anche se la legislazione può variare a seconda dello Stato in cui viene emanata e le polemiche non possono dirsi del tutto placate.
Il futuro della specie Homo sapiens
L’applicazione delle biotecnologie della manipolazione embrionale combinate con quelle dell’ingegneria genetica e della genomica aprono la prospettiva stimolante, e nel contempo angosciante, della modificazione genetica di Homo sapiens. Siamo in grado di modificare a piacere il genoma del topo e di clonare animali e non vi sono aspetti tecnici così complessi da impedire in linea di principio di modificare, in un futuro a medio termine, anche il genoma della nostra specie. Molti pensano che vi siano ottime ragioni per non farlo, mentre altri si chiedono che cosa realmente ci vieti di procedere in questo senso. Certamente ci vorranno ottime argomentazioni perché si raggiunga un consenso sulla liceità della modificazione del genoma: la prima applicazione potrebbe essere nella lotta alle malattie, mediante una sorta di ‘vaccinazione’ genetica. Ma, successivamente, le motivazioni potrebbero diventare più sfumate ed estendersi a cambiamenti per certi aspetti superflui o addirittura discriminatori. Sarà necessaria un’attenta valutazione delle conseguenze sociali ed etiche di queste azioni, ma è probabile che fra qualche secolo le concezioni in proposito saranno molto diverse da quelle attuali, in cui ogni novità viene accolta con grande sospetto. Mentre sarebbe ingenuo ritenere, basandosi su una presunta totale libertà della ricerca, che ogni innovazione sia lecita, è tuttavia necessario valutare ogni azione umana sulla base di quello che essa può comportare in termini di progresso della società.
Bibliografia
Bazzi, Vezzoni 2003: Bazzi, Adriana - Vezzoni, Paolo, Biotecnologie della vita quotidiana, Roma-Bari, Laterza, 2003.
Beja-Pereira 2003: Beja-Pereira, Albano e altri, Gene-culture coevolution between cattle milk protein genes and human lactase gene, “Nature genetics”, 34, 2003, pp. 311-313.
Capecchi 1989: Capecchi, Mario R., Altering the genome by homologous recombination, “Science”, 244, 1989, pp. 1288-1292.
Cavalli Sforza 2000: Cavalli Sforza, Luigi L. - Menozzi, Paolo - Piazza, Alberto, Storia e geografia dei geni umani, Adelphi, Milano, 2000 (trad. ingl.: The history and geography ofhuman genes, Princeton,: Princeton University Press,1994).
Chimpanzee Sequencing and Analysis Consortium 2005: Initial sequence of the chimpanzee genome and comparison with the human genome, “Nature”, 437, 2005, pp. 69-87.
Dulbecco, Vezzoni 1998: Il Progetto Genoma, a cura di Renato Dulbecco, Paolo Vezzoni, “Le Scienze. Quaderni”, 100/D, 1998.
Dolly’s legacy, “Nature”, 445, 2007, p. 795.
Evans, Kaufman 1981: Evans, Martin J. - Kaufman, Matthew H., Establishment in culture of pluripotential cells from mouse embryos, “Nature”, 292, 1981, pp. 154-156.
Gallavotti 2004: Gallavotti, Andrea e altri, The role of barren stalk1 in the architecture of maize, “Nature”, 432, 2004, pp. 630-635.
Garrigan, Hammer 2006: Garrigan, Daniel - Hammer, Michael F., Reconstructing human origins in the genomic era, “Nature reviews genetics”, 7, 2006, pp. 669-680.
Green 2006: Green, Richard E. e altri, Analysis of one million base pairs of Neanderthal DNA, “Nature”, 444, 2006,pp. 330-336.
Holden 2007: Holden, Constance, Long-awaited genetic nondiscrimination bill headed for easy passage, “Science”, 316, 2007, p. 676.
Martin 1981: Martin, Gail R., Isolation of a pluripotent cell line from early mouse embryos cultured in medium conditioned by teratocarcinoma stem cells, “Proceedings of the National Academy of Sciences USA”, 78, 1981, pp. 7634-7638.
Okita 2007: Okita, Keisuke - Ichisaka, Tomoko - Yamanaka, Shinya, Generation of germline-competent induced pluripotent stem cells, “Nature”, 448, 2007, pp. 313-317.
Poinar 2006: Poinar, Hendrik N. e altri, Metagenomics topaleogenomics: large-scale sequencing of mammoth DNA,“Science”, 311, 2006, pp. 392-394.
Rifkin 1998: Rifkin, Jeremy, The biotech century, London, Gollancz, 1998 (trad. it.: Il secolo biotech, Milano, Baldini & Castoldi, 1999).
Thomson 1998: Thomson, James A. e altri, Embryonic stem cell lines derived from human blastocysts, “Science”, 282, 1998, pp. 1145-1147
Vezzoni 2003: Vezzoni, Paolo, Si può clonare un essere umano?, Roma-Bari, Laterza, 2003.
Vezzoni 2006: Vezzoni, Paolo, Il futuro e il passato dell’uomo, Milano, Bruno Mondadori, 2006.
Vezzoni 1997: Vezzoni, Paolo - Sacco, Maria Grazia - Villa, Anna, Animali chimerici, transgenici, knockout e clonati, in: Clonazione: problemi etici e prospettive scientifiche,a cura di Renato Dulbecco, Paolo Vezzoni e altri, Milano, Le Scienze, 1997.
Wilmut 1997: Wilmut, Ian e altri, Viable offspring derived from fetal and adult mammalian cells, “Nature”, 385, 1997,pp. 810-813.