
Cellula
Cellula
(IX, p. 666; App. II, i, p. 550; V, i, p. 546)
La fine del 20° sec. registra un netto progresso nella conoscenza dei meccanismi molecolari alla base delle più diverse funzioni delle cellule. La ricerca scientifica in questo campo ha fondamentalmente abbandonato l'approccio descrittivo a favore dello studio integrato della dinamica molecolare dei fenomeni che hanno luogo nelle cellule.
Molti fattori concomitanti hanno reso possibile questo progresso. Il principale è stato la diffusione delle metodologie cosiddette del DNA ricombinante. In breve, esse consentono di isolare, sequenziare, mutare, ed esprimere in singole c. o in organismi, geni oppure segmenti di DNA aventi funzione regolatoria per l'espressione di altri geni. Questo approccio permette di determinare sperimentalmente, nel contesto di c. viventi, la funzione delle proteine codificate dai geni e di decifrare progressivamente le interazioni che esse hanno nell'insieme del metabolismo cellulare. Altro fattore importante è stato il grande progresso della tecnologia strumentale e dell'automazione, che ha investito tutti i settori. Per es., persino in campi assolutamente tradizionali, come quello della microscopia ottica, fondamentali progressi si sono ottenuti grazie ai nuovi microscopi con impiego di raggi laser (per es., microscopio a effetto tunnel; microscopio confocale) cui sono da aggiungere le nuove tecniche di microscopia elettronica (per es., microscopia elettronica a scansione/trasmissione; microscopia elettronica a effetto tunnel; microscopia elettronica analitica).
Dal punto di vista concettuale, la moderna biologia cellulare ha nei fatti messo da parte alcune problematiche tradizionali, come la distinzione fra c. propriamente dette (eucariote) e c. batteriche, e la questione se considerare i virus organismi viventi (v. cellula, IX e App. II). Infatti, è stata prepotentemente dimostrata l'assoluta similarità, a volte la sostanziale eguaglianza, dei processi biologici fondamentali che occorrono in tutti gli organismi, virus e batteri inclusi. Sorprendentemente alla straordinaria varietà di forme, dimensioni e funzioni delle c. di organismi complessi, uomo incluso, corrisponde in paragone una minore differenza a livello molecolare, che spesso è anche di ordine più quantitativo che qualitativo. Quindi, le specifiche caratteristiche degli organismi viventi, lungo tutta la scala tassonomica, dipendono da numerose, quanto apparentemente minori, variazioni e complicazioni di un insieme di meccanismi molecolari fondamentali, su cui l'evoluzione ha lavorato incessantemente. La sfida per la biologia cellulare consiste oggi nel comprendere il funzionamento integrato, a livello di singola c., dei numerosissimi processi che vi occorrono; ma soprattutto nell'affrontare il problema di come e quanto il gran numero di c. e di loro prodotti che insieme formano gli organismi pluricellulari 'dialoghino' fra loro e generino nuove funzioni.
Queste importanti acquisizioni concettuali stanno avendo a loro volta un forte ruolo propulsivo nello sviluppo ulteriore delle conoscenze. Per es., vengono studiati a fondo eucarioti unicellulari come i lieviti, che offrono grandi vantaggi sperimentali per la possibilità di combinare manipolazioni genetiche e molecolari, per poi identificare i corrispondenti prodotti genici e i meccanismi molecolari nelle c. eucariote di mammifero. Ulteriore conseguenza di questi anni di netta espansione delle conoscenze è stato il superamento, nella prassi scientifica, delle tradizionali divisioni fra le diverse discipline che comprendevano la c. come oggetto di studio (biologia cellulare, biologia molecolare, biochimica, citologia, genetica ecc.). Queste divisioni erano basate su un differente approccio culturale e su diverse metodologie sperimentali. Oggi tutte le tradizionali discipline contribuiscono a una nuova 'biologia sperimentale', che si caratterizza fondamentalmente: a) per lo studio parallelo della struttura e delle funzioni delle singole componenti della c. e dei prodotti da essa secreti, e dei meccanismi alla base dell'interazione fra c. di un organismo; b) per l'approccio sperimentale basato sull'uso integrato di tutte le metodologie disponibili.
Il grande sviluppo delle conoscenze di base ha dato ovviamente forte impulso alla ricerca applicata. Questo periodo ha visto crescere e consolidarsi le cosiddette 'biotecnologie', cioè metodologie mirate alla creazione e allo sviluppo di processi biologici utili in settori diversi quali l'agro-alimentare, lo zootecnico, il farmaceutico (v. biotecnologia; ingegneria genetica, in questa Appendice). Particolare preminenza e interesse hanno assunto le biotecnologie mediche, con la speranza di poter risolvere numerose patologie umane, anche attraverso la cosiddetta terapia genica, e di generare una serie di farmaci di totale nuova concezione. Per le evidenti ripercussioni delle biotecnologie, soprattutto quelle mediche legate alla riproduzione umana (v. fecondazione, in questa Appendice), l'aspetto bioetico (v. bioetica, in questa Appendice) ad esse connesso è fortemente sentito anche dalla pubblica opinione.
Negli ultimi anni si sono avuti straordinari progressi nella conoscenza di un grande numero di processi cellulari. I punti cruciali di questa ricerca sono riportati qui di seguito.
Generazione e mantenimento dell'organizzazione interna delle cellule eucariote animali

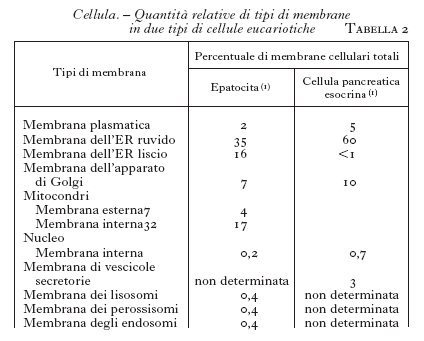
Le c. sono dotate di una complessa architettura, dovuta a una notevole estensione di membrane interne (di norma più del 90% delle membrane totali), non connesse alla membrana plasmatica ma assolutamente simili nella struttura (doppio strato fosfolipidico con proteine intrinseche ed estrinseche) e nelle proprietà chimico-fisiche fondamentali (permeabilità selettiva, fluidità; v. citologia, App. IV). Queste membrane dividono lo spazio intracellulare generando vacuoli (grossolanamente sferici, fino a 1÷2 micron, e quindi con un alto rapporto superficie/volume), tubuli, e piccole vescicole (dai 50 ai 200 nanometri di diametro). In ogni caso si distinguono la membrana e il lume (lo spazio racchiuso nella membrana limitante). Queste strutture occupano circa il 50% del volume totale di una c., mentre l'altro 50% è costituito dal citosol, che è la frazione solubile del citoplasma (tab. 1 e tab. 2). Criteri morfologici e dinamici avevano già consentito di differenziare le membrane cellulari nei diversi organelli (v. citologia, App. IV). Le ricerche più recenti hanno portato alla definizione del concetto di compartimento intracellulare, sulla base di criteri prevalentemente molecolari e funzionali. Ogni compartimento è caratterizzato da un preciso insieme di proteine sia nel lume che nella membrana limitante, ed è quindi distinto dagli altri per i processi metabolici in cui è coinvolto. Anche la composizione lipidica caratterizza i vari compartimenti, ma le differenze sono meno accentuate. Uno o più compartimenti distinti concorrono alla formazione degli organelli (si avverte che i termini organello e compartimento sono spesso erroneamente usati come sinonimi nella letteratura scientifica contemporanea).
La generazione e il mantenimento della compartimentazione intracellulare sono fenomeni molto complessi per vari motivi:
1) una singola c. sintetizza circa 10⁴ diverse proteine, producendone per ciascun tipo numerose copie, per un totale di circa 10¹⁰ copie: la sintesi delle proteine ha luogo nel citosol, ma tutte le copie di una stessa proteina raggiungono, e mantengono, la stessa localizzazione intracellulare finale. La corretta localizzazione è condizione essenziale per l'espressione della funzione della proteina. Sono state descritte patologie umane in cui il difetto molecolare risiede unicamente nel meccanismo di localizzazione: un esempio è fornito dalla malattia da inclusioni cellulari, in cui gli enzimi lisosomiali sono normalmente sintetizzati ma non accumulati nei lisosomi;
2) le proteine non attraversano spontaneamente il doppio strato lipidico delle membrane cellulari, un processo altamente sfavorevole dal punto di vista termodinamico: specifici complessi proteici assolvono il compito di consentire il transito creando nelle membrane 'pori' selettivi (e di norma transitori);
3) le c. non sintetizzano membrane ex novo ma rielaborano e accrescono le preesistenti;
4) continui passaggi di materiale avvengono fra i diversi compartimenti, fra i vari elementi di un singolo compartimento, e fra i compartimenti e il citosol, senza che vada mai perduta l'identità strutturale e funzionale dei singoli compartimenti. Si assiste quindi a un continuo processo di scelta selettiva e di smistamento delle molecole, soprattutto delle proteine. Inoltre, la compartimentazione cellulare non è una rigida e costante caratteristica delle c., bensì il risultato di un equilibrio dinamico: stati fisiologici, come la divisione cellulare, o patologici, come quelli dovuti all'azione di molecole esogene che interferiscono con il metabolismo cellulare, portano a una rapida e profonda ristrutturazione dell'architettura interna delle c. (scomparsa per vescicolazione dell'apparato di Golgi, dell'involucro nucleare e del reticolo endoplasmatico: massiva tubulazione del compartimento endosomiale ecc.);
5) un singolo compartimento può essere rappresentato da strutture fisicamente separate fra loro (v. tab. 1), ma fra i singoli componenti vi è comunque continuo scambio sia di materiale del lume che delle membrane limitanti.

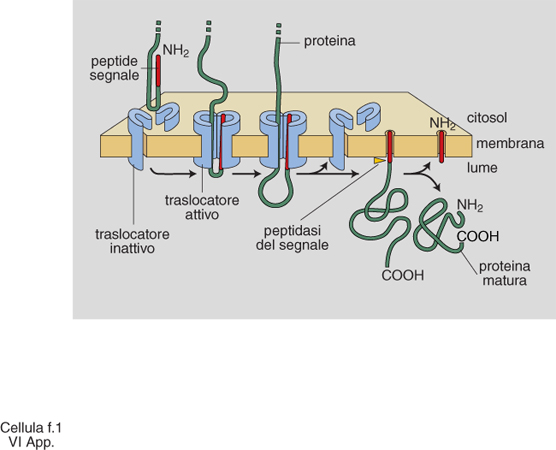
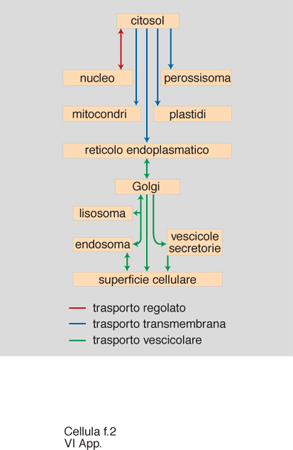
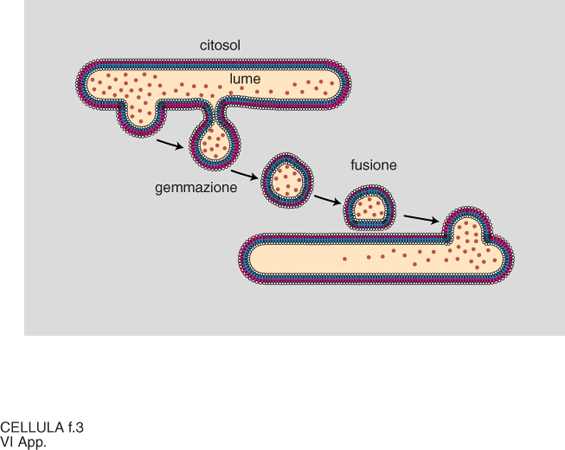
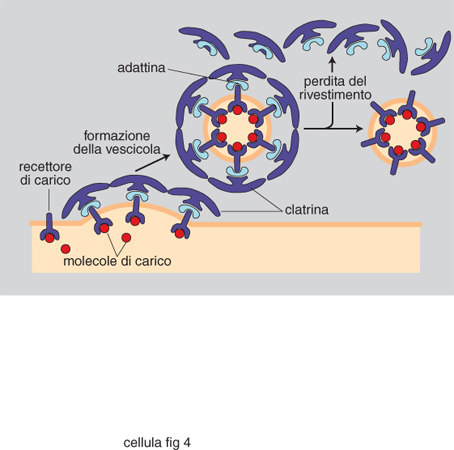
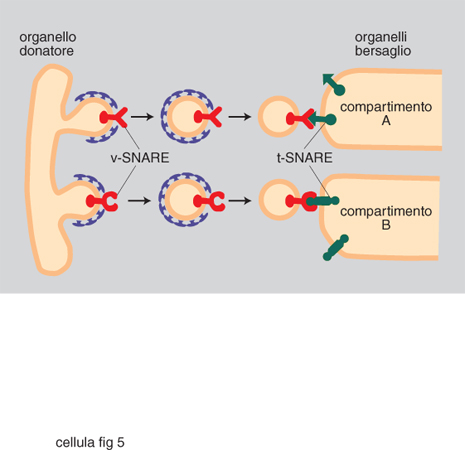
Lo scenario descritto è emerso in parallelo alla comprensione dei principi generali alla base della generazione e del mantenimento della compartimentazione cellulare. Sono stati individuati segnali di localizzazione (tab. 3), costituiti da una successione continua o discontinua di amminoacidi delle proteine da localizzare, e i complessi macromolecolari della c., costituiti da un insieme di specifiche proteine, deputati a riconoscere i segnali e a mettere in atto la localizzazione stessa (fig. 1). Aiuta a comprendere la dimensione di questi processi l'osservazione che la sola analisi genetica in lieviti ha già individuato circa 70 geni che codificano le proteine degli apparati di localizzazione, un numero certamente destinato a crescere. Il 'traffico' delle proteine nelle c. può essere riassunto come indicato in fig. 2. Se ne distinguono tre tipi fondamentali, a seconda che gli eventi siano fra citosol e nucleo, fra citosol e organelli, o fra i compartimenti delle vie di secrezione ed endocitosi, funzionalmente interconnesse. Nell'ultimo caso, il traffico è essenzialmente mediato da vescicole trasportatrici, che si distaccano per gemmazione del compartimento di partenza, attraversano il citosol (si pensa interagendo con filamenti del citoscheletro per raggiungere il compartimento di destinazione), e rilasciano il contenuto del lume e della membrana limitante al compartimento di destinazione con un processo di fusione delle membrane (fig. 3, fig. 4 e fig. 5). Tutti i diversi momenti del traffico, vescicolare e non, sono strettamente controllati per garantirne la specificità.
La comprensione dei meccanismi responsabili della specificità è ancora parziale, ma il senso generale è emerso chiaramente. La chiave di volta del processo risiede nel riconoscimento sterico fra proteine (un processo concettualmente simile al riconoscimento enzima/substrato), che innesca l'assemblaggio o l'attivazione temporanea del complesso macromolecolare deputato alla localizzazione. Il traffico intracellulare richiede energia, che viene fornita dalla c. attraverso l'idrolisi di nucleosidi trifosfati. Particolare rilevanza negli eventi di traffico hanno le proteine che legano e consumano GTP (guanosintrifosfato). Fra di esse, ben caratterizzata è la famiglia delle proteine Rab, composta da più di trenta membri, tutte coinvolte in diversi momenti del traffico vescicolare. Queste proteine esercitano un ulteriore controllo sulla fedeltà del processo e ne dettano il 'tempo', fungendo da veri e propri 'interruttori' molecolari attraverso l'utilizzo del GTP.
Instabilità dinamica e funzioni motorie dei microtubuli
I microtubuli sono strutture cilindriche cave delimitate da 13 protofilamenti, ciascuno dei quali è costituito dalla successione regolare di dimeri di molecole di tubulina alfa e beta (v. cellula: Citoscheletro, App. V). Nelle c. animali i microtubuli convergono verso una regione circoscritta, posta di solito in prossimità del nucleo, definita come centro di organizzazione dei microtubuli o centrosoma, e identificabile per la presenza dei centrioli e di una quantità variabile di materiale amorfo pericentriolare. Studi non più recentissimi avevano già dimostrato che proprio in questa regione prendono origine quasi tutti i microtubuli della cellula. Per convenzione si definisce estremità - (meno) del microtubulo quella che si trova in prossimità del centrosoma ed estremità + (più) quella più distante.
Il maggior interesse in questo campo di ricerca è attualmente legato alle seguenti problematiche: comprensione dei meccanismi molecolari che presiedono alla formazione dei microtubuli; caratterizzazione del comportamento dinamico dei microtubuli; approfondimento del loro ruolo funzionale, soprattutto durante il processo di mitosi (v. oltre), in cui i microtubuli si riorganizzano in modo spettacolare a formare il fuso mitotico.
La polimerizzazione della tubulina, e quindi l'accrescimento del microtubulo, dipende dalla disponibilità di guanosintrifosfato (GTP) e di ioni Mg++.
Tuttavia esperimenti di polimerizzazione delle tubuline condotti in vitro hanno mostrato che in queste condizioni il processo è abbastanza lento a meno che non vengano aggiunte anche proteine citoplasmatiche. Difatti alla costituzione del sistema dei microtubuli partecipano diversi tipi di proteine, complessivamente indicate come MAP (proteine associate ai microtubuli); è interessante notare come c. di tipo differente contengano MAP differenti, così come all'interno dello stesso tipo cellulare coesistano tipi differenti di MAP. L'esempio più eclatante è la c. nervosa, dove ai microtubuli presenti nei prolungamenti dendritici è associata la MAP nota come MAP₂, mentre ai microtubuli dell'assone si associa la MAP nota come proteina τ.

Il centrosoma appare come la sede in cui si realizzano gli eventi molecolari che consentono la 'nascita' di nuovi microtubuli. Il centrosoma può essere isolato dalla c., e in questa maniera recentemente si è potuto procedere alla caratterizzazione del materiale proteico pericentriolare associato ai centrioli. In questo modo si è giunti a un'interessante scoperta: il centrosoma contiene tra i suoi vari costituenti (molti dei quali devono essere ancora identificati) molecole di una particolare forma di tubulina, la tubulina gamma, assai simile ma non identica alla tubulina alfa e beta. La tubulina gamma non entra nella costituzione del microtubulo, ma la sua presenza nel centrosoma risulta indispensabile perché si inneschi la polimerizzazione (fig. 6). Tra i punti che attendono un chiarimento rimane il significato dei centrioli nel centrosoma; le c. vegetali, per es., non hanno centrioli eppure sono capaci di formare microtubuli.
Il comportamento dinamico dei microtubuli è stato rivelato da osservazioni cinematografiche. È emersa una serie di proprietà che si compendiano nel principio della instabilità dinamica dei microtubuli. Secondo questo principio, in primo luogo la maggior parte dei microtubuli è sottoposta a fasi di lento accrescimento seguite da improvviso e brusco accorciamento; questi fenomeni sono correlati rispettivamente all'aggiunta o alla sottrazione di molecole di tubulina all'estremità + del microtubulo. In secondo luogo, il processo non è sincronizzato: questo comporta che mentre alcuni microtubuli sono in fase di accrescimento, altri collassano, e comunque in ogni momento è possibile osservare nuovi microtubuli che si formano dal centrosoma. Il terzo punto importante è che accanto a questa classe di microtubuli altamente dinamici esiste un'altra componente, per lo più minoritaria, di microtubuli molto stabili che non manifestano la tendenza a collassare. Questa classe di microtubuli diventa altamente rappresentata in strutture come cilia, flagelli e gli stessi centrioli.
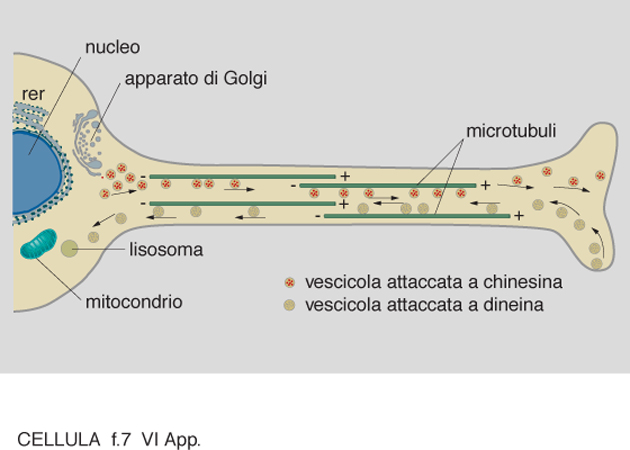
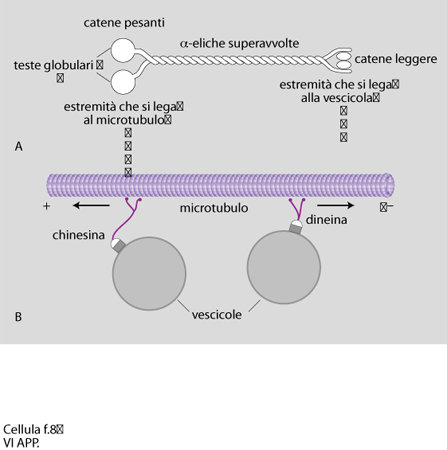
Una delle ipotesi più stimolanti riguardo al possibile ruolo funzionale dei microtubuli aveva previsto la possibilità che essi venissero utilizzati a modo di 'binari' per il trasporto di strutture vescicolari e per il movimento degli organelli. Lo studio di come vescicole ricoperte di membrana fossero trasportate avanti e indietro lungo i microtubuli presenti nei lunghi prolungamenti delle c. nervose non solo ha confermato questa ipotesi, ma ha portato all'identificazione dei cosiddetti 'motori microtubulari' (fig. 7). Si tratta di complessi proteici che in virtù della loro capacità di idrolizzare l'ATP (adenosintrifosfato) sono in grado di convertire energia chimica in energia meccanica. I motori, associandosi ai microtubuli da un lato e agli elementi da trasportare dall'altro, consentono lo scorrere di questi ultimi lungo i microtubuli. Sono stati così inizialmente caratterizzati due complessi polipeptidici chiamati chinesina e dineina citoplasmatica, il primo in grado di far muovere le vescicole dal corpo cellulare verso la periferia, il secondo capace di produrre un moto retrogrado inverso dalla periferia verso il corpo cellulare. Uno schema molto semplificato di come possano funzionare le molecole di chinesina e di dineina è mostrato in fig. 8. Ulteriori ricerche promosse da questa scoperta hanno portato a interessanti nuovi risultati: la dimostrazione della presenza di chinesina e dineina in moltissimi tipi di c. e la scoperta che esistono molte altre molecole che presentano analogie strutturali e funzionali con la chinesina. A oggi si conoscono una trentina di queste molecole, definite KRP (Kinesin Related Proteins), identificate in eucarioti sia unicellulari che pluricellulari. Tutti i membri di questa famiglia hanno in comune una sequenza di 340 amminoacidi che costituisce la regione che nella molecola prende contatto con il microtubulo, e che funziona da 'motore' in virtù dell'attività enzimatica ATPasica ad essa associata. Questi dati hanno messo in evidenza la rilevanza del traffico bidirezionale di materiali basato sulle funzioni motorie dei microtubuli.
Gran parte dei futuri sforzi in questo campo sarà volta sicuramente a identificare le funzioni di ciascuno dei membri della famiglia delle proteine chinesina-simili. In questa direzione un aspetto di grande rilevanza è stato la dimostrazione della partecipazione dei motori cellulari ai molteplici eventi che caratterizzano la divisione cellulare. Proteine simili alla chinesina e alla dineina citoplasmatica sono state identificate in associazione non solo con i microtubuli del fuso mitotico, ma anche con la cromatina dei cromosomi. Queste molecole, capaci di generare forze di movimento, sarebbero quindi in grado di dirigere sia il moto dei cromosomi sia lo scorrimento reciproco dei microtubuli che si osserva in mitosi. Si apre dunque una nuova frontiera in cui la divisione cellulare può venire reinterpretata e analizzata in termini di rapporti dinamici tra microtubuli e cromosomi mediati dai motori cellulari.
Il controllo del ciclo cellulare
Negli organismi pluricellulari il processo di proliferazione cellulare provvede allo sviluppo dell'organismo durante la vita embrionale nonché alla crescita e conservazione dei tessuti nei soggetti adulti. A sviluppo ultimato, nell'uomo, la capacità di replicazione è ristretta ad alcuni tessuti le cui c. sono sottoposte a usura o rapido ricambio (epidermide, mucosa intestinale, midollo osseo). L'attività proliferativa è rigidamente controllata: un aspetto eclatante della trasformazione neoplastica risiede proprio nello stravolgimento del controllo della moltiplicazione cellulare. Lo svolgimento dell'attività proliferativa richiede che la c. che va a dividersi per produrre due c. figlie provveda: 1) a replicare il proprio materiale genetico (DNA) in due copie identiche, una per ciascuna delle c. figlie; 2) a generare un apparato in grado di separare tali copie, organizzate in forma di cromatina, e di ripartirle nelle due nuove c.; 3) a duplicare e ripartire correttamente gli organelli citoplasmatici.
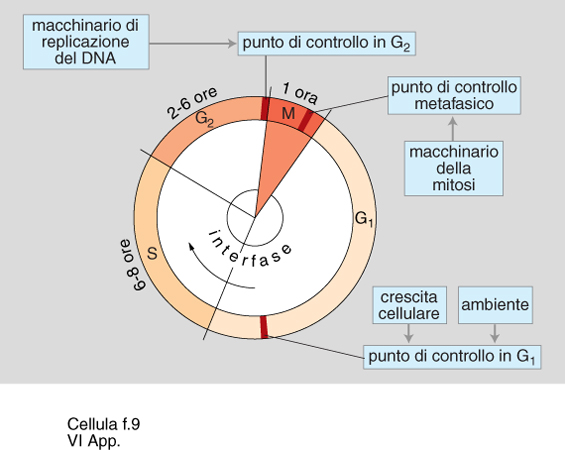
Il ciclo cellulare è costituito da quella serie di eventi che si susseguono dall'inizio di una divisione cellulare fino all'inizio della divisione successiva (v. citologia: Riproduzione cellulare, App. IV). Il tempo necessario a completare il ciclo cellulare è indicato come tempo di duplicazione. Popolazioni cellulari differenti, anche all'interno dello stesso organismo, possono avere ciclo cellulare con tempi di duplicazione differenti. Nelle c. di mammifero di solito la durata del ciclo oscilla tra 16 e 24 ore. Durante la mitosi, che è la fase in cui si manifestano le più vistose modificazioni morfologiche delle c. (v. citologia, App. II), si realizza la divisione fisica della c. madre nelle c. figlie. L'interfase precede la mitosi, ed è in questo periodo che la massa cellulare si accresce, il materiale genetico viene duplicato e vengono sintetizzate numerose diverse proteine necessarie allo svolgimento delle varie fasi del ciclo replicativo. La duplicazione del materiale genetico (fase S) non occupa che una limitata porzione dell'interfase. Essa è preceduta da una fase pre-sintetica G₁ e da una fase post-sintetica G₂ che precede la mitosi (v. citologia: Riproduzione cellulare, App. IV). La proliferazione cellulare si ottiene dunque dal ciclico ripetersi di queste quattro fasi: mitosi - G₁ - fase S - G₂. Lo studio delle modalità di duplicazione delle c. eucariotiche più semplici, come quelle di lievito, ha consentito di evidenziare il fatto che le c. sono capaci di eseguire una sorta di monitoraggio sul corretto svolgimento del processo replicativo, predisponendo veri e propri punti di controllo (check points), a livello dei quali si può arrestare il ciclo cellulare se esso non sta procedendo correttamente (fig. 9).
Così per es., se l'accrescimento della c. non ha portato al raggiungimento di dimensioni adeguate, il ciclo può fermarsi in G₁; se il materiale genetico è stato incompletamente o erroneamente duplicato il ciclo può fermarsi in G₂. In ogni caso non si darà esecuzione alla divisione cellulare.
Studi genetici condotti con c. mutanti di lievito, incapaci di svolgere cicli replicativi normali, hanno consentito di decifrare una parte degli strumenti molecolari che le c. utilizzano nel controllo del ciclo cellulare. Recenti studi indicano inoltre che questi meccanismi sono comuni a tutti gli eucarioti.

In particolare si è visto che tra i componenti chiave si collocano specifiche protein-chinasi, enzimi che catalizzano l'aggiunta di gruppi fosfato a specifici amminoacidi delle proteine. Questi enzimi si attivano grazie al loro legame con una proteina regolatrice chiamata ciclina: dunque, una maniera con cui le c. possono modulare l'attività di queste chinasi (definite appunto chinasi ciclina-dipendenti) è quella di far variare in modo ciclico e regolato la concentrazione delle cicline. Nelle c. di alcuni tipi di lievito (eucarioti unicellulari) un complesso ciclina/chinasi viene attivato alla transizione G₂-M e un analogo complesso viene attivato nella transizione G₁-S (fig. 10). Uno degli aspetti più attuali dello studio sul controllo del ciclo cellulare è volto all'identificazione dei possibili substrati del complesso ciclina-chinasi, in modo da poter interpretare molecolarmente alcuni aspetti morfologici da tempo descritti come peculiari della divisione cellulare.
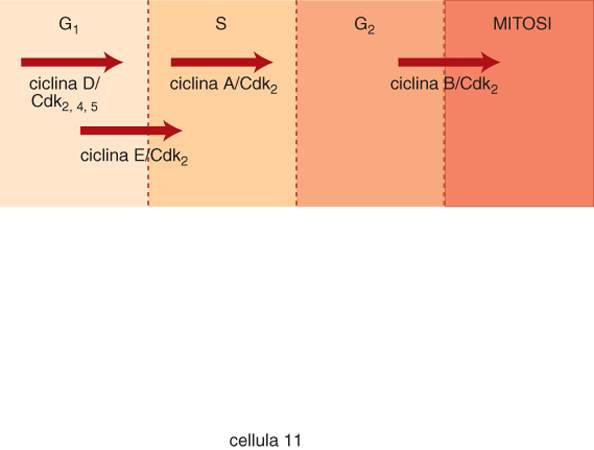
Nei mammiferi il ciclo cellulare è più complesso: lo si evince anche dal fatto che sono state individuate svariate chinasi del ciclo cellulare e numerose cicline in grado di formare complessi attivi durante le varie fasi del ciclo (fig. 11). Molti di questi agiscono in G₁ ed è probabile che siano responsabili tra l'altro dell'attivazione del complesso macchinario molecolare, enzimatico e non, necessario per dare l'avvio alla duplicazione del DNA. Quando la c. sta per passare dalla fase G₂ alla mitosi si forma un complesso attivo chinasi-ciclina definito fattore promuovente la mitosi (MPF). MPF è presente in tutte le c. eucariotiche, ed è in grado di fosforilare, tra l'altro, alcune proteine associate alla membrana del nucleo e altre associate ai microtubuli. È verosimile che la dissoluzione della membrana nucleare e la costituzione del fuso mitotico osservate nella profase della mitosi (v. citologia: Riproduzione cellulare, App. IV) possano essere conseguenza di queste reazioni di fosforilazione.
Il controllo del ciclo cellulare è fondato su un programma genetico interno ma è dipendente da segnali esterni. Si tratta per lo più di controlli di tipo positivo, esercitato cioè da sostanze capaci di indurre l'entrata o la progressione nel ciclo delle cellule. Per es., fattori di crescita e altre sostanze di tipo ormonale agiscono, spesso a concentrazioni bassissime e con grande specificità, su c. bersaglio dotate di recettori specifici sulla loro superficie, e ne modulano l'attività proliferativa. Lo studio sui meccanismi di trasduzione dei segnali elicitati da queste molecole all'interno della c. fino al nucleo, sede della duplicazione del DNA, è di grande attualità. Per contro, il fatto che la maggior parte delle c. dell'organismo adulto non si dividano, e che anche c. proliferanti possano astenersi dal dividersi per periodi più o meno lunghi, depone per l'esistenza di meccanismi di controllo negativo. In questo ambito rientra l'identificazione delle funzioni di geni cosiddetti oncosoppressori, in quanto la loro espressione tenderebbe a inibire la comparsa di tumori. L'approfondimento dei meccanismi che controllano la proliferazione cellulare rappresenta un tema arduo ma anche una sfida avvincente e necessaria, come ci ricorda la gravità delle malattie neoplastiche in cui l'attività proliferativa si pone fuori controllo, svincolata dalle esigenze generali dell'organismo (v. oncogeni, App. V e in questa Appendice).
Apoptosi
L'equilibrio tra processi di proliferazione e morte cellulare consente agli organismi pluricellulari di provvedere al loro sviluppo durante l'embriogenesi e di mantenere l'omeostasi di tessuti e organi nell'individuo adulto. Esiste pertanto una 'morte cellulare programmata' la cui esecuzione è controllata e fondata su un meccanismo attivato dalla c. stessa. Essa si manifesta attraverso una precisa e costante sequenza di aspetti morfologici, alla cui base vi è un processo che è rimasto fondamentalmente conservato, attraverso l'evoluzione, dai nematodi fino all'uomo. Il termine apoptosi, derivato dalla parola greca che indica distacco, caduta (in particolare delle foglie di un albero), viene spesso usato per definire questo processo. L'apoptosi va pertanto distinta da altre forme patologiche di morte cellulare che non sono parte di un programma di sviluppo dell'organismo e che occorrono con differenti meccanismi. Le forme di 'suicidio cellulare' che caratterizzano l'apoptosi sono ampiamente diffuse in natura sia tra gli invertebrati che tra i vertebrati. Come si è detto, nella maggior parte dei casi l'apoptosi rappresenta un evento del tutto fisiologico e previsto. Nei nematodi, per es., c. nervose, muscolari, epiteliali, intestinali e delle gonadi vanno incontro ad apoptosi durante il normale sviluppo. Nei vertebrati, l'apoptosi è stata osservata in pressoché ogni tessuto (e studiata con particolare attenzione nello sviluppo del sistema nervoso e del sistema immunitario); nell'uomo, in condizioni sia fisiologiche che patologiche. L'apoptosi potrebbe provvedere a rimuovere c. prodotte in eccesso, c. che abbiano esaurito la propria funzione, c. potenzialmente dannose per l'organismo, c. che si sono sviluppate in modo improprio.
Il processo di apoptosi è oggi al centro di imponenti sforzi della ricerca volti a definirne i molteplici aspetti morfologici, biochimici e genetici. Gli eventi morfologici che caratterizzano l'apoptosi differiscono da quelli della necrosi cellulare in cui le c. muoiono come conseguenza di ingiurie subite principalmente a livello della membrana plasmatica. Tipicamente nell'apoptosi si osserva una sequenza di modificazioni che comprende: 1) la condensazione della cromatina del nucleo a formare numerose masse di materiale granulare addossate in corrispondenza della periferia del nucleo; 2) la formazione di numerose e vistose protrusioni a livello della membrana plasmatica; 3) la rottura del nucleo e la formazione di masse di cromatina delimitate da membrana nucleare. Queste segregano all'interno delle grosse protuberanze che si formano sulla superficie cellulare: a seguito del loro distacco dal corpo cellulare vengono a trovarsi confinate in strutture, definite 'corpi apoptotici', delimitate da una membrana derivata da quella plasmatica. Oltre alla cromatina i corpi apoptotici possono contenere anche organelli citoplasmatici ammassati ma ancora ben conservati. La formazione dei corpi apoptotici caratterizza l'evento terminale dell'apoptosi, la quale interessa spesso intere popolazioni cellulari (è emblematica per es. la perdita della coda dei girini), ma può talora interessare un numero limitato di c. nell'ambito di un'ampia popolazione di c. in normali condizioni. Queste ultime sono in grado di introdurre nel proprio citoplasma i corpi apoptotici, attraverso il meccanismo della fagocitosi (v. cellula: Endocitosi, App. V), e questi vengono degradati all'interno dei vacuoli lisosomali (fig. 12). Il processo è molto rapido e non resta traccia delle c. morte.
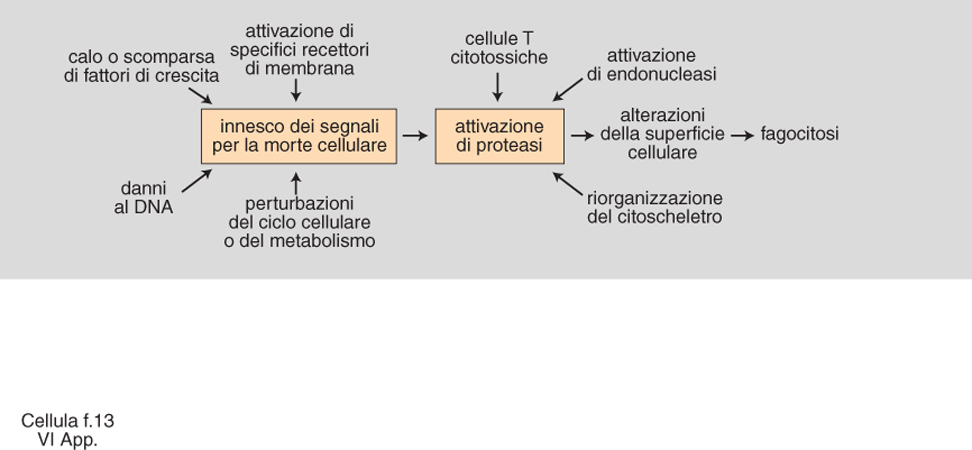
Una delle cause fisiologicamente più importanti in grado di promuovere l'apoptosi è la modificazione delle concentrazioni di ormoni e/o fattori di crescita. In alcuni casi anche la trasformazione neoplastica o l'infezione delle c. da parte di certi virus si sono dimostrate capaci di indurre nelle c. interessate il processo di apoptosi, prevenendo, nel primo caso, la formazione o l'eventuale accrescimento dei tumori, nel secondo, la propagazione dell'infezione alle c. non ancora infettate. L'apoptosi può anche essere indotta sperimentalmente e studiata utilizzando le colture in vitro. Lo spettro degli agenti causali in grado di promuovere l'apoptosi è ampio: si tratta di agenti sia fisici sia chimici, talora anche di farmaci. Un evento che accompagna frequentemente il processo di apoptosi è la frammentazione della molecola di DNA contenuta nei cromosomi. I frammenti sono prodotti dall'attivazione di enzimi con attività DNAsica che agiscono a livello delle regioni più accessibili della molecola generando frammenti costituiti all'incirca da 180 paia di basi (o da multipli di questo numero). Tale evento è irreversibile, e va inteso come punto di non ritorno nel processo. Studi recenti hanno evidenziato come nella c. apoptotica si rendano attive svariate proteine enzimatiche con attività proteolitica. Molte di queste agiscono in cascata, autoattivandosi e/o attivando altre proteasi. Gli enzimi di questo tipo sono stati chiamati caspasi. Ogni differente caspasi è definita da un numero (caspasi 1, caspasi 2 ecc.); a oggi si conoscono circa 10 caspasi. Il problema biologico più interessante da indagare è l'individuazione dei substrati intracellulari di questi enzimi: così facendo si potrà giungere a ulteriori chiarimenti dei processi molecolari dell'apoptosi. Per gli eventi che caratterizzano l'apoptosi v. .
Gli studi genetici hanno proceduto di pari passo con quelli morfologici e biochimici. Il modello di riferimento in questo caso è stato il nematode Caenorhabditis elegans, in cui il disegno complessivo di divisioni e morti cellulari, che ne caratterizza lo sviluppo, è stato determinato con l'osservazione diretta dell'animale che è trasparente. In altre parole si sa esattamente quante c., e in quale parte dell'organismo in sviluppo, andranno incontro ad apoptosi (131 su 1090 nello sviluppo di un adulto ermafrodita). Questa osservazione conferma che si tratta di un processo programmato e regolato. Attraverso lo studio di individui portatori di mutazioni sono stati identificati i principali geni che presiedono alla morte cellulare programmata in Caenorhabditis elegans. Essi sono ricordati con la sigla ced (cell death). In particolare, il gene ced 3, che codifica per un enzima proteolitico analogo alle caspasi, sembra svolgere un ruolo chiave. Nei mammiferi esistono almeno 6 differenti geni con omologia a ced 3 di Caenorhabditis elegans, i quali codificano per enzimi che sono inizialmente sintetizzati come precursori inattivi. Che cosa possa dare avvio all'attivazione sequenziale di questi enzimi non è ancora noto. Nel nematode è il prodotto codificato dal gene ced 4 che sembra avere la capacità di attivare l'enzima codificato da ced 3. Al contrario, la capacità di ced 3 di causare l'apoptosi può essere bloccata dalla presenza del prodotto del gene ced 9. Presumibilmente ced 9 è attivo nella maggior parte delle c. normali impedendo in tal modo l'apoptosi. L'omologo nei mammiferi di ced 9 è stato identificato nel gene bcl-2.
bibliografia
B. Alberts et al., Molecular biology of the cell, New York 1983, 1994³ (trad. it. Bologna 1984, 1995³).
D.M. Glover, C. Gonzalez, J.W. Raff, Il centrosoma, in Le Scienze, 1993, 300, pp. 38-44.
A. Quattrone, S. Capaccioli, I geni della morte cellulare programmata, in Le Scienze, 1995, 326, pp. 42-50.
S.L. Wolfe, An introduction to cell and molecular biology, Belmont 1995 (trad. it. Napoli 1996).
T. Alescio, C. Garbi, L. Alescio-Zonta, Biologia generale e molecolare della cellula, Padova 19962.
J. E. Rothmann, L. Orci, La gemmazione di vescicole nelle cellule viventi, in Le Scienze, 1996, 333, pp. 38-44.
Alberts et al., Essential cell biology, New York 1998 (trad. it. Bologna 1999).