
Complessita biologica
Complessità biologica
I sistemi viventi della biosfera vengono di norma ordinati in una gerarchia di entità, ciascuna delle quali vuole rappresentare un livello di organizzazione della materia vivente. Cellula, organismo pluricellulare, popolazione, comunità ecologica sono oggetti biologici tipicamente graduati secondo un criterio di complessità crescente. Per molti anni la complessità biologica è sembrata essere un attributo, seppure non esclusivo, talmente inerente ai sistemi viventi da venire percepito come un fatto ovvio, come un dato di osservazione evidente, da non meritare un'attenzione teorica particolare. L'assunzione acritica dell'autoevidenza della complessità biologica dei viventi ha avuto come principali conseguenze, da una parte, la sua ontologizzazione e, dall'altra, la mancanza di una definizione chiara ed esaustiva che fosse fondata sui risultati di ricerche specifiche, sperimentali o teoriche. Negli anni Novanta del Novecento, più cautamente, pur rimandando al futuro il compito di formulare una definizione scientifica di complessità biologica, si è cominciato a guardare a essa come a un attributo non tanto dei sistemi viventi in sé stessi, quanto, piuttosto, delle rappresentazioni che dei sistemi viventi dà la scienza. Intrinsecamente complessi sarebbero perciò i modelli della conoscenza biologica e non gli oggetti del mondo biologico.
Tra le tante conseguenze collegabili a tale mutamento di ottica se ne indicano in questa sede soltanto due. In primo luogo, l'attribuire la complessità biologica ai modelli di rappresentazione, sottesi dalla relazione osservatore-osservato, piuttosto che agli oggetti in sé stessi rimuove dall'attività di ricerca le eventuali preclusioni di ordine metafisico, rendendo i sistemi viventi conoscibili anche in via di principio. In secondo luogo, la complessità biologica pone i modelli biologici come in larga misura irriducibili alle spiegazioni classiche nell'ambito delle scienze fisiche, formulate in termini matematici, in cui essa sembra essere invece marginale.
Rimanendo nel solco di una visione tradizionale, epistemologicamente discutibile, ma ancora largamente condivisa sul piano linguistico-comunicativo, che attribuisce la complessità biologica ai sistemi viventi piuttosto che alla modellizzazione che di loro dà la scienza, potremmo dire che il carattere intrinsecamente complesso dei sistemi viventi concorre a fondare l'autonomia della biologia e a mutare i caratteri definitori della conoscenza scientifica. L'idea di scienza basata sul modello proprio delle scienze quantitative, nomologiche e assiomatizzabili, sembra oggi in via di revisione: essa si modifica e si amplia per accogliere quei metodi e quelle procedure propri della conoscenza imperfetta, solitamente non matematizzabile e assai raramente predittiva, che caratterizza le scienze della vita.
Organizzazione
Una proprietà fondamentale di tutti i viventi è quella di essere sistemi organizzati. Possedere un'organizzazione significa presentare un certo insieme di relazioni che assicurano la coerenza interna del sistema. Queste relazioni sono responsabili dell'unitarietà di ogni sistema vivente e della sua tenuta. L'organizzazione dei viventi mostra di avere una natura gerarchica per cui gli organismi unicellulari, quelli pluricellulari, le popolazioni e le biocenosi si presentano come entità fenomeniche caratterizzate da configurazioni strutturali e da dinamiche processuali differenti a ciascun livello e dotate di proprietà né immediatamente deducibili né prevedibili. Oltre che gerarchica, tale organizzazione è anche chiusa, grazie all'innesco di un processo di circolarità in meccanismi altrimenti lineari di causa ed effetto. La logica circolare (ben esemplificata dal rapporto causale: gameti-zigote-gameti) percorre tutti i livelli della gerarchia, dalla cellula all'ecosistema. I sistemi viventi sono sistemi chiusi rispetto alla loro organizzazione interna, ma sono sistemi aperti sull'esterno, con cui scambiano materia, energia, informazione. Ogni organismo è necessariamente connesso con l'ambiente, e tale connessione è a tal punto obbligata che diventa impossibile anche solo concepire l'esistenza di un organismo isolato, oppure isolabile, dal proprio ambiente.
Individualità
I sistemi viventi non sono ripetitivi: l'eterogeneità è la norma. Sia sul piano strutturale sia su quello funzionale, i sistemi viventi sono distinguibili dai sistemi naturali non viventi sulla base della loro individualità. Di norma ogni vivente possiede una propria individualità (un'unicità originariamente dovuta alle proprietà stocastiche della sorgente di variazione) che viene codificata nei geni, costruita epigeneticamente e trasmessa alle generazioni successive. All'interno di ciascun deme (popolazione locale), la somma delle individualità si traduce nell'assetto di variazione genetica mediante cui la popolazione nel suo insieme fronteggia le pressioni dell'ambiente. L'unicità degli individui diventa essenziale per l'adattamento della popolazione a un ambiente perennemente mutevole.
La stragrande maggioranza dei sistemi viventi può essere costituita non solo da entità tra loro intrinsecamente, cioè genotipicamente, diverse, ma può anche essere formata da subunità strutturalmente eterogenee, che hanno quindi comportamenti che seguono leggi differenti l'una dall'altra. È il caso del corredo di proteine enzimatiche di una cellula (che catalizzano particolari reazioni biochimiche regolate da specifici meccanismi di controllo), o di quello dei diversi tipi di cellule costitutive del corpo dei Vertebrati. In sostanza la complessità biologica deve essere riferita a sistemi di tipo individuale, cioè non ripetitivo, per i quali valgono leggi specifiche. Da ciò deriva un'importante conseguenza di ordine teorico generale: la non universalità delle leggi ricavabili dalle descrizioni dei sistemi biologici ha reso estremamente difficile l'applicabilità dei metodi matematici di tipo deterministico ai fenomeni biologici. Un'ulteriore osservazione riguarda il fatto che l'individualità dei sistemi biologici ha una duplice natura: essa è concausa di evoluzione e prodotto storico dell'evoluzione.
Diversità
Differenze di vario ordine e grado sottostanno alla complessità. Indipendentemente dagli effetti di scala, la diversità dei sistemi viventi deriva dalla loro individualità e concorre a definire la loro complessità. Globalmente, la diversità biologica è costituita dall'insieme delle differenze osservabili tra gli esseri viventi. Tali differenze possono essere descritte in termini di quantità, di variazione e di variabilità degli organismi, nonché, semplificando, in rapporto ai geni, alle specie e agli ecosistemi. In particolare, la diversità genetica è rappresentata da tutte le differenze ereditabili esistenti tra gli individui in una popolazione e tra le diverse popolazioni, che sono riconoscibili a livello genico. La diversità genetica è riconducibile, in ultimo, alle differenze di sequenza nelle coppie di basi degli acidi nucleici. Negli organismi dotati di sessualità, le novità genetiche comparse per mutazione possono diffondersi nella popolazione mediante ricombinazione, cioè attraverso un dispositivo capace di generare un immenso numero di differenze.
La struttura e il numero dei cromosomi, come pure la quantità di DNA contenuto in una cellula, sono esempi di diversità genetica. In particolare, la quantità di DNA contenuta in una cellula (dimensione del genoma) permette di confrontare organismi di taxa di livello anche molto superiore a quello di specie. Nei batteri, per esempio, le dimensioni dei genomi variano ampiamente da 6×105 bp a più di 107 bp. Il genoma del micoplasma è formato da circa 400 geni, mentre negli altri batteri il numero di geni varia tra 500 e 8000. La maggioranza degli eucarioti possiede, invece, qualcosa come 50.000 geni e un contenuto di DNA estremamente differente, variabile tra 8,8×106 bp e 6,9×1011 bp. Evolutivamente, la comparsa della sessualità ha provocato un incremento della diversità genetica e della complessità biologica dei sistemi viventi. La sua enorme diffusione tassonomica viene associata a meccanismi di riparazione del DNA e di differenziamento cellulare e viene interpretata come meccanismo base per la produzione di varianti genetiche adattabili ai cambiamenti ambientali per via fenotipica, per l'incremento della cladogenesi e il decremento dei tassi di estinzione. In sostanza la sessualità sembra essere importante perché può agire ed essere favorita a molti livelli di organizzazione dei sistemi viventi, da quello di gene a quello di popolazione-specie.
La diversità tassonomica di specie, oltre alla componente spaziale e geografica, ha una componente temporale. La diversità oggi osservabile è il risultato di un processo storico lunghissimo iniziato tra 3900 e 3400 milioni di anni fa con la comparsa della vita sul nostro pianeta. Tale processo di produzione delle differenze tra i viventi ha natura irreversibile. La ricostruzione paleontologica dimostra che la diversità macrotassonomica (intesa come ricchezza di tipi organizzativi) raggiunse il suo apice nel Cambriano, e che la diversità di famiglie e di specie è andata aumentando dal Cambriano al Pleistocene, con poche battute d'arresto in corrispondenza delle fasi, brevi e isolate, di estinzione di massa. Le differenze ecologiche, e quindi la complessità ecologica degli ecosistemi, sono le più difficili da definire perché di norma le comunità e gli ecosistemi sono entità dai contorni sfumati, perché gli ecosistemi non sono realtà esclusivamente biologiche ma includono anche la componente abiotica e, infine, perché esiste un fattore di scala spazio-temporale che influenza pesantemente l'osservazione, la misurazione e l'interpretazione delle differenze sia intra- sia interecosistemiche.
Spesso la diversità ecologica viene articolata su più livelli gerarchici, da quello di biocenosi a quello di ecosistema, di paesaggio, anche se vi sono dubbi che a ognuno di questi livelli corrisponda effettivamente un'entità strutturalmente e funzionalmente autonoma. Tipicamente la diversità ecologica viene caratterizzata con riferimento a tre principali componenti. La diversità alfa (α) e la diversità gamma (γ) riguardano il numero di taxa presenti, rispettivamente, alla scala locale e a quella regionale; la diversità β rappresenta l'intensità del cambiamento in specie osservate in zone di transizione tra due tipi diversi di sistemi lungo un gradiente fisiografico. La diversità beta (β) misura, attraverso indici, il livello di somiglianza tra aree prossime, la diversità ecologica tra biotopi contigui. La più semplice relazione tra queste tre componenti è tale per cui la diversità β è uguale al rapporto tra la diversità γ e la diversità α.
Il fatto che le specie di una comunità non abbiano tutte lo stesso numero di individui, ma presentino differenti abbondanze, è uno degli aspetti più cospicui degli ecosistemi. Per lungo tempo si è creduto che la complessità biologica strutturale degli ecosistemi potesse essere ridotta, o almeno sintetizzata, in un indice quantitativo che ne misurasse la diversità di specie, ma indagini più approfondite hanno messo in luce che, così intesa, la diversità non è una proprietà esclusiva delle biocenosi, ma di qualsiasi collezione di tipi eterogenei caratterizzata da abbondanze differenti. C'è invece concordanza sul fatto che nel tempo la diversità complessiva di una biocenosi cresca (anche nel caso in cui il numero assoluto di specie rimanga costante), a mano a mano che le abbondanze delle specie tendono a equivalersi. La complessità ecologica ha anche manifestazioni funzionali coincidenti con i tipi di relazioni che intercorrono tra gli organismi di un ecosistema e i suoi processi. Sono processi ecosistemici lo stoccaggio e il flusso di materiali e di energia, il mantenimento della stabilità ecologica, la ripartizione delle risorse tra le specie, nonché molti comportamenti delle specie (tra i tanti, quelli collegati alla competizione, alla predazione, al parassitismo, all'impollinazione). La diversità ecologica funzionale riguarda i vari modi con cui si possono realizzare le funzioni ecologiche degli ecosistemi: a ognuno di questi modi corrisponderà il tipo di organizzazione proprio di uno specifico ecosistema.
Relazionalità
Dall'azione di forze si passa alle relazioni attraverso segnali. La possibilità, comprovata per certi sistemi non viventi (per es., il sistema fisico di Benard o la reazione di Belousov e Zhapotinski), di passare spontaneamente da condizioni di tipo caotico a stati di tipo ordinato, ha indotto molti ricercatori a ritenere che processi analoghi fossero responsabili della complessità biologica dei sistemi viventi e del suo incremento. L'analogia tra i sistemi complessi di tipo fisico o chimico e quelli biologici è però solo superficiale. In effetti, l'azione delle forze fisiche esterne che tendono a strutturare le unità caotiche di un sistema chimico molecolare viene a mancare nei sistemi biologici. Qui, invece, l'elemento di ordine, capace di organizzare le componenti eterogenee del sistema, è rappresentato da relazioni che promuovono l'interconnessione tra le parti. Tali relazioni hanno la forma di un segnale. Attraverso segnali le cellule di una coltura batterica o le componenti di un sistema pluricellulare trasferiscono informazioni dall'una all'altra, coordinando i processi interni con quanto accade all'esterno di ciascuna di esse. A livello degli organismi pluricellulari, il sistema costituito dal segnale e dal suo recettore non solo assicura l'integrazione tra le varie cellule, per esempio durante i processi di sviluppo, ma negli animali può anche consentire l'integrazione sociale degli individui, vuoi attraverso l'azione degli ormoni, vuoi attraverso quella dei neurotrasmettitori.
In tutti i gruppi di organismi si assiste a una diversificazione della coppia segnale-recettore. Tale diversificazione evolutiva produce sia una complicazione del sistema, sia una sua divisione in compartimenti, con la localizzazione, in alcune regioni del corpo, delle cellule contenenti i differenti recettori. Inoltre, se da una parte l'evoluzione filogenetica della coppia segnale-recettore ha la funzione di rendere più efficiente e affidabile l'integrazione tra gli individui (con ciò aumentando la diversità interna, intrasistemica, dei sistemi viventi), dall'altra essa serve a mantenere isolati sistemi tra loro incompatibili e in competizione (favorendo così la diversità esterna, intersistemica). I segnali coinvolti nel comportamento, e responsabili della produzione delle barriere interspecifiche di isolamento riproduttivo precopula, esemplificano puntualmente la funzione di mantenimento e incremento della diversità intersistemica e illustrano, a un livello assai elevato di complessità biologica, quale sia la natura delle 'forze' che determinano la dinamica dei sistemi viventi. Negli animali dotati di un sistema nervoso e di un cervello sufficientemente complesso, il sistema segnale-recettore è ancora all'opera nel consentire l'apprendimento e dunque la generazione di più sofisticati meccanismi di produzione di diversità e di complessità. Infine, riferendosi a un altro ambito fenomenologico, è sempre la capacità relazionale dei sistemi viventi che è chiamata in causa nell'elaborazione epigenetica dell'informazione genomica nei processi di riconoscimento immunitario, nella morfogenesi, nell'ontogenesi del comportamento.
Tendenze evolutive e complessità
L'ordinamento di complessità biologica crescente, cioè cellula procariotica, cellula eucariotica, organismo pluricellulare, popolazione, comunità, ecosistema, di solito si accompagna all'idea che nella storia della vita sul pianeta sia riconoscibile una tendenza storica verso la crescita della complessità biologica dei sistemi viventi. Per quanto suggestiva, questa convinzione si dimostra errata, perché se è indiscutibile che le forme organizzative più semplici hanno preceduto di norma quelle più complesse, tuttavia la coesistenza temporale di forme poco complesse accanto a forme molto complesse testimonia non la sostituzione di sistemi meno complessi con sistemi più complessi, ma la coesistenza di sistemi a vario grado di complessità. Inoltre è anche noto che nel corso dell'evoluzione vi sono stati non pochi casi in cui i sistemi viventi sono andati incontro a processi di drastica semplificazione di alcune subunità, dunque a una riduzione della loro complessità biologica generale. È il caso, assai popolare, della semplificazione anatomica dell'occhio, fino alla sua scomparsa totale e definitiva negli animali ciechi e anoftalmi che vivono negli ambienti cavernicoli. Un'evoluzione regressiva analoga è presentata dai parassiti intestinali nei quali si assiste, tra l'altro, alla perdita dell'apparato digerente.
Informazione e complessità
Nei sistemi viventi l'informazione è conservata e organizzata negli acidi nucleici che specificano in maniera non lineare i vari fenotipi, i quali, a loro volta, manifestano complessità biologica gerarchizzata. Nel decennio successivo al 1953, dopo che James Watson e Francis Crick ebbero scoperto la struttura a doppia elica del DNA, alcuni ricercatori formularono in maniera rigorosa il problema dell'esistenza di un 'codice genetico' (l'insieme delle regole con cui una sequenza di nucleotidi di un gene viene tradotta nella sequenza di amminoacidi di una proteina). La prossimità temporale tra la scoperta dell'esistenza di un tale codice e l'elaborazione teorica della nozione di informazione, sviluppata nel contesto di problemi tecnologici di ingegneria delle comunicazioni a opera di Claude E. Shannon e Warren Weaver nel 1949, concorsero a suggerire l'accostamento tra l'informazione biologica e l'informazione quale è intesa nella teoria omonima. Precisamente, vennero cercate correlazioni e analogie sulla misura del contenuto di informazione di sequenze (normali e mutanti per una o più variazioni) e sulla misura dell'entropia di informazione.
In questo approccio, le sequenze nucleotidiche vengono considerate sequenze di simboli dell'alfabeto a quattro lettere proprie degli acidi nucleici. La formazione di una data sequenza Ik di lunghezza n avviene allora con probabilità pk=4−n, e il contenuto di informazione, espresso in bit, della sequenza è Ik=2n, indipendentemente dalla particolare sequenza considerata. In effetti l'informazione di Shannon considera solo la componente statistica dell'informazione, senza riferimento al suo contenuto, dunque al significato, che è invece estremamente importante per i sistemi viventi, dato che è il significato del messaggio a determinare le conseguenze dell'informazione erogata. Approcci successivi dovuti a Andrei Nikolaevich Kolmogorov e Gregory J. Chaitin considerarono la struttura interna delle sequenze. Nella teoria algoritmica dell'informazione il contenuto informativo di un messaggio è pari alla lunghezza del più piccolo programma di computer che, una volta eseguito, è in grado di produrre l'oggetto. Ne deriva che messaggi periodici o molto ridondanti possiedono basso rapporto di informazione algoritmica/complessità biologica, mentre sequenze casuali hanno un alto rapporto di informazione algoritmica/complessità biologica, non potendo in alcun modo essere create attraverso un programma che sia più breve delle sequenze stesse.
Tuttavia il fatto che una sequenza casuale sia dotata di alta complessità biologica contraddice la realtà biologica, nella quale i sistemi altamente complessi non hanno strutture e comportamenti governati dal caso, ma sono invece altamente organizzati al loro interno. Anche se la nozione di complessità biologica algoritmica, essendo interessata alla struttura interna del messaggio, risponde meglio alle esigenze dei biologi, tuttavia non consente di affrontare il problema cruciale del significato. Quanto sia importante questo punto è chiaro considerando, per esempio, che i geni strutturali dell'uomo e dello scimpanzé sono identici al 99% e che basta solo una frazione piccola, ma altamente significativa, di geni diversi per determinare differenze biologiche di grande rilievo. Le differenze di sequenza nel DNA tra uomo e scimpanzé consistono prevalentemente nell'uso del codice per quanto riguarda le sequenze tradotte e di variazioni nelle sequenze non trascritte. La distanza genetica media tra le due specie, stimata su circa 50 geni strutturali, è risultata essere addirittura inferiore a quella tra specie sorelle, che sono per definizione anatomicamente poco distinguibili. Tali piccole differenze, capaci però di generare diversità organizzative tra le due specie, vanno quasi certamente ricondotte a cambiamenti a carico di pochi geni regolatori.
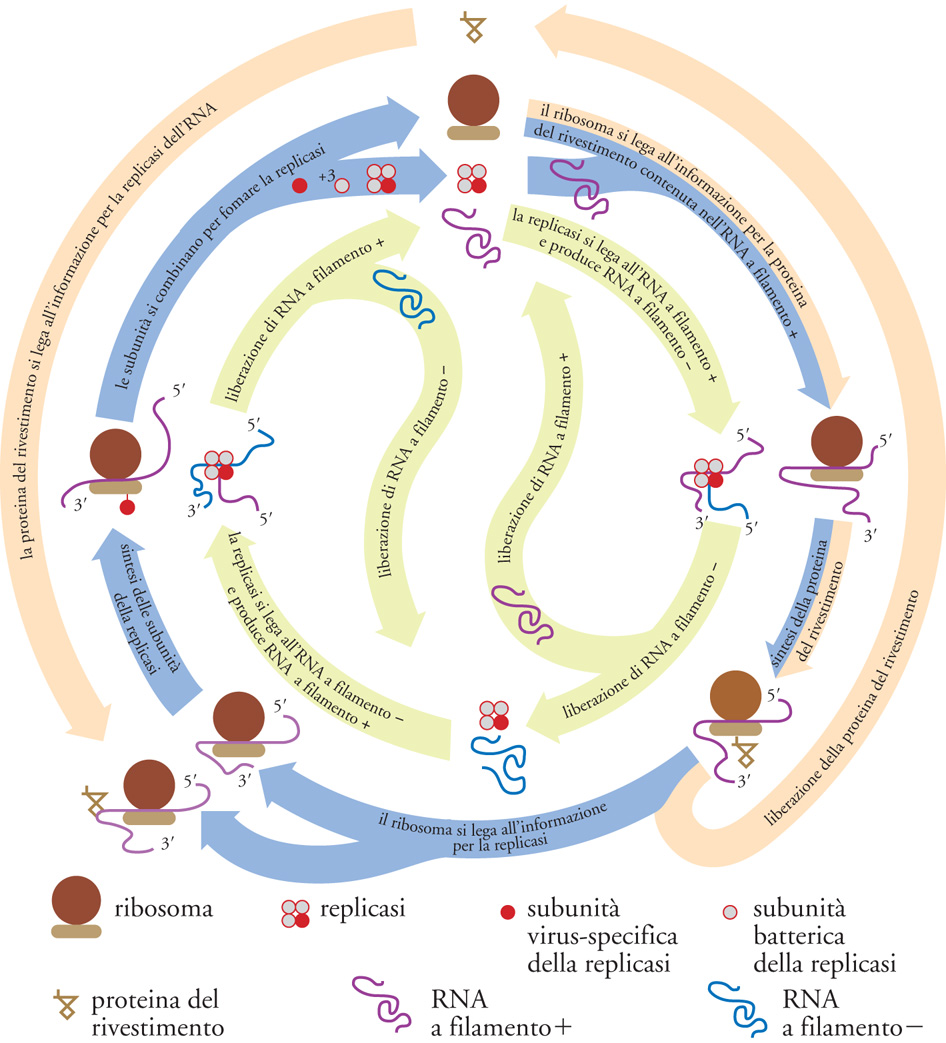
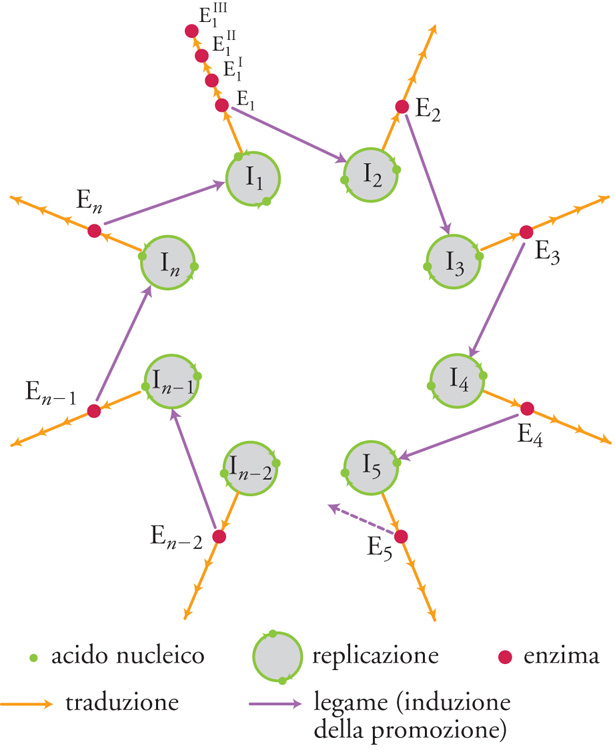
Il passaggio critico nella gerarchia della complessità biologica corrisponde ovviamente all'incremento di questa nei sistemi viventi, particolarmente intenso durante le transizioni evolutive più importanti (per es., il passaggio da cellula procariotica a cellula eucariotica, da sistema unicellulare a sistema pluricellulare, da individuo a colonia). Pur in mancanza di una teoria generale dell'incremento di complessità biologica, questo fenomeno ha iniziato ad avere una sua spiegazione circoscrivendo l'analisi a sistemi molto semplici come i virus. A cavallo tra la fine degli anni Settanta e i primi anni Ottanta del Novecento Manfred Eigen e Peter Schuster hanno proposto un modello teorico, detto 'dell'iperciclo', di notevole interesse euristico, in cui sono specificate le condizioni minime che permettono di conservare e accumulare informazione biologica: un passo indispensabile per l'aumento di complessità biologica di un sistema. L'iperciclo di Eigen e Schuster è un concatenamento chiuso di cicli di reazioni in cui enzimi e acidi nucleici promuovono reciprocamente le loro sintesi (figg. 5 e 6). Dato che le molecole replicanti sono dipendenti le une dalle altre (c'è cooperazione), cambia la dinamica di popolazione e gli individui molecolari non competono (come invece accade con la replicazione indipendente delle molecole, che provoca competizione e produzione di 'quasispecie': una distribuzione di individui mutanti centrata su una o più sequenze dominanti, analoga al tipo selvatico della genetica classica), ma si integrano a un livello più elevato.
Nel modello dell'iperciclo la transizione al gradino superiore avviene in più fasi: i replicatori indipendenti competono per le risorse e ottimizzano la propria fitness individuale; l'interdipendenza riproduttiva riduce, fino a eliminarla, la competizione tra replicatori; la riproduzione fra replicatori cooperanti ha come effetto la formazione di un'unità funzionale dove sono integrate le diverse funzioni di fenotipi differenti; la compartimentalizzazione dei replicatori crea, attraverso l'integrazione spaziale, una nuova unità di selezione; l'integrazione dei replicatori produce una nuova classe di individui replicatori passibili di evoluzione autonoma: i genotipi dei replicatori inizialmente autonomi formano un genoma unico, più grande. Tra l'altro, il modello prevede l'interazione con individui molecolari 'parassiti', che traggono vantaggio temporaneo dalla nuova organizzazione senza integrarvisi, e individua nell'abbondanza di risorse la condizione più favorevole all'integrazione. L'incremento della complessità biologica di un sistema può avvenire anche per altre vie. La duplicazione genica e genomica, per esempio, è un meccanismo che consente ai geni duplicati di sviluppare eventuali nuove funzioni. In condizioni ambientali favorevoli (risorse abbondanti e di basso costo energetico), le varianti nuove con genoma accresciuto competono con successo contro le varianti più piccole ma ottimizzate. Le condizioni da soddisfare perché vi sia incremento di informazione e di complessità biologica nell'evoluzione, riguardano la componente relazionale e sono: l'esistenza di un meccanismo per la 'simbiosi' (come nel caso della produzione della cellula eucariote, che può essere vista anche come una cellula 'pluricellulare') e l'esistenza di un meccanismo per l'epigenesi.
Selezione naturale
In larga misura l'evoluzione dei sistemi viventi è la storia dei passaggi da un livello di complessità biologica all'altro, ovvero, secondo molti ricercatori, da un'unità di selezione all'altra. L'avvio del processo evolutivo ha coinciso con l'originarsi di molecole autoreplicantesi; queste vengono poi incorporate in unità cellulari assai ben delimitate, capaci successivamente di integrare al loro interno i genomi di organelli dotati di autoreplicabilità, e poi in grado di sfruttare (come popolazioni, non come individui), grazie alla sessualità, l'enorme vantaggio della ricombinazione genetica, con l'incorporazione e la rapida diffusione delle combinazioni più vantaggiose. A questo punto del processo evolutivo, con la comparsa del sesso e delle specie, la selezione naturale assume la forma, che è più familiare ai biologi, di riproduzione differenziale di genotipi. Il modello esplicativo elaborato con la 'teoria sintetica dell'evoluzione' è appunto centrato sulla riproduzione differenziale di varianti individuali all'interno di una variazione intrapopolazionale di caratteri ereditabili. Tuttavia, è opinione diffusa che questo modello esplicativo vada ora modificato per accogliere le moderne acquisizioni della biologia a livello molecolare, cellulare e dello sviluppo. I dati sperimentali portati da queste discipline hanno cominciato a chiarire, individuandone in molti casi i meccanismi, le fasi del processo di incremento di complessità biologica precedente la comparsa del livello di popolazione-specie.
Sebbene tutte queste conoscenze mancassero all'epoca della formulazione della teoria sintetica (1936-1947), e siano continuate a mancare ancora per alcuni decenni, molti biologi, sulla base di risultati di ricerche condotte con metodi tradizionali, avevano sollevato già negli anni Sessanta la questione della probabile esistenza di livelli di azione multipli della selezione naturale, sia al di sopra sia al di sotto del livello individuale. Il quadro generale che sta dietro all'idea della molteplicità delle unità di selezione è che al nascere di un nuovo livello organizzativo si verifica un cambiamento nel bersaglio e nella forma della selezione. In un primo momento la selezione viene esercitata soltanto dall'ambiente esterno; con la nascita del nuovo livello strutturale (se esso contiene fisicamente il livello strutturale più antico) l'ambiente esterno agisce direttamente solo sull'unità di questo nuovo livello: la selezione sull'unità gerarchicamente inferiore non è più esercitata dallo stesso ambiente di prima (quello che precedentemente era esterno) ma dall'unità neoformata, che pertanto funzionerà come ambiente per l'unità più vecchia e gerarchicamente inferiore. Questa concezione prevede un controllo verticale, dall'alto, dell'unità maggiore sull'ambito delle possibili variazioni dell'unità minore. E anche se, in effetti, i cambiamenti dell'unità di livello inferiore possono arrivare a influenzare la replica dell'unità del livello immediatamente superiore, tale azione dal basso è consentita nella misura in cui questi cambiamenti non sono svantaggiosi per l'unità maggiore che interagisce con l'ambiente esterno. In questa visione, all'incremento di complessità biologica è associato lo slittamento della selezione dal vecchio al nuovo livello di organizzazione, che quindi perde gradi di libertà trasformandosi in vincolo interno al sistema.
Un esempio convincente di incremento della complessità biologica strutturale con importanti effetti di regolazione funzionale è offerto dal fenomeno del linkage disequilibrium (disequilibrio da associazione). Si è constatato che geni strettamente associati sul cromosoma tendono a trovarsi in uno stato di disequilibrio da associazione. In questo caso l'organizzazione del cromosoma riduce la probabilità che, dentro i gameti, alleli di siti variabili si trovino associati a caso: tale organizzazione rappresenta un vincolo per questo fenomeno. Questo controllo può essere esercitato dall'alto, come risultato di una storia evolutiva con una forte componente deterministica: è il caso delle inversioni paracentriche polimorfiche del terzo cromosoma di Drosophila pseudoobscura. Tali inversioni cromosomiche, ben note ai genetisti, opponendosi ai normali meccanismi di ricombinazione, rappresentano una forma di organizzazione spaziale del cromosoma, selettivamente premiata, nella quale si trovano riuniti alleli che di per sé possono non fornire elevatissimi vantaggi, ma che nella forma associata interagiscono tra loro conferendo un vantaggio selettivo all'individuo portatore.
L'inversione paracentrica conferisce vantaggio al gruppo perché, riducendo la frequenza dei genomi ricombinanti, protegge gli assortimenti aploidi vantaggiosi. Se l'inversione fornisce un vantaggio selettivo al suo portatore, non è necessario che la selezione su ogni singolo locus genico sia forte (con bassa ricombinazione, infatti, il coefficiente di accoppiamento tra geni può essere molto grande, senza che vi siano grosse differenze negli effetti medi degli alleli ai loci separati, detti 'fitness marginali'). Fenomeni come quello descritto dimostrano che i singoli geni non evolvono indipendentemente. Da qui, ancora una volta, l'impossibilità di prevedere la dinamica evolutiva di singoli geni se questi sono organizzati in un'unità evolutiva di ordine superiore. Nel caso del coadattamento genetico, quindi, l'unità di selezione non coincide con singoli geni, ma con più ampie frazioni di genoma. Più in generale, nei sistemi viventi l'unità di selezione dovrebbe essere identificabile, almeno in linea di principio, a diversi livelli purché essa si replichi e conservi la propria identità col passare delle generazioni.
Complessità biologica e storia
Uno degli aspetti più ricchi di conseguenze, sebbene immediatamente non appariscente, dei sistemi viventi è quello di essere sistemi non universalmente necessitati, nel senso che le loro strutture e i loro comportamenti non sono descrivibili con leggi universali, bensì con enunciati validi caso per caso, o tutt'al più validi per una casistica assai limitata. Da cosa derivi questa caratteristica è facilmente comprensibile, considerando i seguenti esempi quantitativi che illustrano un peculiare aspetto dei sistemi viventi: l'enorme discrepanza tra quanto è teoricamente possibile su base combinatoria e quanto invece si è storicamente realizzato.
Diversità molecolare
Una proteina di piccolo peso molecolare è costituita da circa 300 amminoacidi, ognuno dei quali è codificato da tre nucleotidi; perciò i geni che specificano la proteina sono costituiti da circa 1000 nucleotidi. All'interno di ciascuna catena, ogni nucleotide rappresenta una sola tra quattro possibilità. Il numero di tutte le possibili catene è allora eguale a 41000, ovvero circa 10600. Il numero di sequenze amminoacidiche immaginabili è quindi 10600, ma solo una frazione piccolissima possiede la stabilità e l'attività enzimatica necessarie a garantire il funzionamento cellulare. La probabilità che l'ottimalità di queste sequenze amminoacidiche sia nata per caso è praticamente zero ed è necessario ipotizzare l'esistenza di una causa, di un fattore deterministico sotto forma di selezione. Inoltre, considerando anche che la vita sul pianeta ha meno di 4 miliardi di anni, un lasso di tempo breve rispetto alla potenziale produzione di combinazioni di amminoacidi, si deve concludere che deve esservi stata un'azione selettiva particolarmente intensa.
Polimorfismo
Partendo da 6000 geni strutturali (numero medio stimato per Drosophila) e ammettendo verosimilmente che la metà sia polimorfica, avremo 3000 geni. Assumendo una media di 2 alleli per locus elettroforetico, avremo 3 possibili combinazioni alleliche per locus, che per tutti i loci dà un numero totale di combinazioni alleliche pari a 33000 (63000 nel caso diploide). Questa è una cifra strabiliante, non solo assolutamente superiore al totale dei genotipi aploidi già prodotti e producibili in futuro, ma anche di gran lunga superiore al numero stimato di tutte le particelle elementari del nucleo atomico (protoni, elettroni, ecc.) presenti nell'universo (ca. 1079). La massa di questi genotipi supererebbe, se tutti fossero prodotti, la massa dei nucleoni. Invece, di tutte le possibili combinazioni genotipiche solo alcune effettivamente si sono realizzate, e solo poche altre si realizzeranno; ciò rappresenta una chiara evidenza della natura relativa e contingente dei sistemi biologici. È questa contingenza a fondare la natura storica dei viventi: la complessità biologica osservabile deriva da vincoli strutturali e funzionali preesistenti, che a loro volta determinano la dinamica futura dei sistemi. Non tutti i cammini evolutivi sono dunque possibili, ma solo quelli compatibili con il regime di vincoli esterni e interni al sistema. I sistemi viventi sono dunque entità storicamente determinate e la biologia assume anch'essa carattere storico ogni volta che si pone problemi che vanno oltre il comportamento delle molecole.
Ci si potrebbe chiedere infine se sia opportuna un'indagine 'al di là delle molecole' e dei loro comportamenti. La risposta è affermativa, perché la natura dei vincoli che determinano le condizioni di possibilità dei sistemi viventi è esplicitabile solo in termini evolutivi. La ricerca ha iniziato a fare luce sulla natura di questi vincoli. Si tratta dei vincoli adattativi, sincronici, che strutturano il collegamento con i fattori ambientali, e dei vincoli di sviluppo che specificano diacronicamente il modo d'essere di ciascun individuo e di ciascuna specie. La natura di questi vincoli legittima, inoltre, gli interrogativi teleonomici sui perché di certe manifestazioni dei viventi. Il nucleo delle risposte alle domande sui perché non è niente altro che la descrizione della comparsa e dell'affermazione nel tempo dei vincoli capaci di conferire identità agli organismi, oppure di trasformarla. Le domande sui perché non sembrano riducibili a domande sul come. Piuttosto si hanno spiegazioni di due ordini distinti: una funzionale, finalizzata all'esplicitazione delle cause prossime, dei meccanismi in gioco; l'altra tesa a elaborare una spiegazione che è la storia delle vicende responsabili del fenomeno indagato. Sul piano della generalizzazione, questa storia funziona come modello esemplare, esplicativo per tutta la serie di eventi assimilabili a quello studiato.
Bibliografia
Ageno 1986: Ageno, Mario, Le radici della biologia, Milano, Feltrinelli, 1986.
Bonner 1988: Bonner, John T., The evolution of complexity by means of natural selection, Princeton (N.J.), Princeton University Press, 1988.
Buss 1987: Buss, Leo W., The evolution of individuality, Princeton (N.J.), Princeton University Press, 1987.
Eigen, Schuster 1979; Eigen, Manfred - Schuster, Peter, The hypercycle. A principle of natural self-organization, Berlin-New York, Springer, 1979.
Heywood 1995: Global biodiversity assessment, edited by Vernon H. Heywood, Cambridge-New York, Cambridge University Press, 1995.
Maynard Smith, Szathmáry 1995: Maynard Smith, John - Szathmáry, Eörs, The major transitions in evolution, Oxford-New York, Freeman/Spektrum, 1995.
Mayr 1997: Mayr, Ernst, This is biology: the science of the living world, Cambridge (Mass.)-London, Belknap Press of Harvard University Press, 1997 (trad. it.: Il modello biologico, Milano, McGraw-Hill, 1998).
Rizzotti 1996: Defining life. The central problem in theoretical biology, edited by Martino Rizzotti, Padova, University of Padova, 1996.
Schulze, Mooney 1993: Biodiversity and ecosystem function, edited by Ernst-Detlef Schulze, Harold A. Mooney, Berlin-New York, Springer, 1993.
Signor 1990: Signor, Philip W., The geologic history of diversity, "Annual review of ecology and systematics", 21, 1990, pp. 509-39.