
Ecologia
Ecologia
di Paul Duvigneaud
SOMMARIO: 1. Introduzione e definizioni. □ 2. L'ecologia agli inizi del XX secolo. □ 3. Metabolismo dell'individuo e della popolazione. □ 4. La popolazione: a) numero e densità degli individui; b) natalità, mortalità, crescita; c) equilibrio e fluttuazioni; curve di crescita; d) struttura; e) metabolismo; f) relazioni con lo spazio; area; g) competizione interspeciflca; h) competizione intraspecifica; i) aggregazione; 1) eterogeneità e genetica delle popolazioni: l'ecotipo. □ 5. La comunità: a) definizione; b) relazioni trofiche (alimentari) in seno alle biocenosi; c) produttività della biocenosi; regolazione delle catene trofiche; ecomoni; d) controllo biologico delle popolazioni in seno alle comunità; lotta biologica; la biologia florale; e) struttura nello spazio e nel tempo; stratificazione e periodicità; aggregazione; f) nozione di nicchia ecologica; g) competizione interspecifica o coabitazione; h) dinamismo; i) ampiezza e limiti; l'ecotono; l) la fitosociologia; m) comunità animali; n) microbiocenosi; o) formazioni vegetali del globo; i grandi biomi. □ 6. L'ecosistema: a) descrizione; b) funzionamento dell'ecosistema (metabolismo, produttività e biomassa); c) estensione dell'ecosistema; la biosfera. □ 7. L'ecologia e la società umana. □ Bibliografia.
1. Introduzione e definizioni.
L'ecologia (dal greco οἶκος = casa) è, teoricamente, lo studio dell'habitat degli esseri viventi (organismi). Il nome è stato introdotto da E. Haeckel (v., 1866). In generale, essa comprende le relazioni degli organismi con l'habitat (ambiente vitale circostante), ivi compresi i rapporti degli organismi stessi fra di loro. Con maggiore aderenza alla realtà odierna, l'ecologia può essere definita la scienza degli ecosistemi. Quasi sconosciuta al grande pubblico fino al 1965, essa ha acquistato improvvisamente notevole importanza allorché si è compreso che l'impatto distruttore dell'uomo sul proprio ambiente diviene tale da costituire un pericolo reale, già a breve termine, per l'avvenire della specie umana.
Sembra certo che i primi materiali biologici siano sorti abioticamente nell'oceano primitivo, parecchi miliardi di anni fa, in un ambiente atmosferico fortemente riducente (anaerobiosi) sottoposto a radiazioni ultraviolette di breve lunghezza d'onda e di grande potere sia sintetico, sia distruttivo (v. biofisica; v. vita, origine della).
Nella natura, quale oggi si presenta, le piante verdi sono i soli organismi che possano ancora effettuare la sintesi dei materiali biologici a partire da materia minerale semplice e luce visibile del sole; questi organismi sono perciò detti ‛autotrofi'; gli altri esseri viventi (‛eterotrofi') possono unicamente consumare tali produttori.
La paleoecologia delinea la storia degli ambienti diversissimi che si sono succeduti nel corso delle ere geologiche; un'importante sintesi di essa è l'opera di Ager (v., 1963). Ma fin dal 1926 Brooks aveva tratteggiato una sintesi, divenuta classica, dei ‛climi attraverso le epoche geologiche' (v., 1926).
I materiali biologici, creati sulla superficie del globo dal flusso dell'energia solare e dallo sviluppo dei grandi cicli biogeochimici dell'acqua, del carbonio, dell'azoto, dello zolfo, del fosforo e di altri elementi indispensabili alla vita, si integrano a livelli di organizzazione sempre più complessi quali la cellula, l'individuo con i suoi tessuti e organi, la popolazione, la comunità e infine la biosfera (v. fig. 1).
Una popolazione è un sistema biologico formato da un gruppo di individui della stessa specie, viventi in luoghi, condizioni e periodi determinati.
Una comunità (o biocenosi) è un sistema biologico formato da tutte le popolazioni (quindi da individui di specie diverse) che vivono insieme in luoghi, condizioni ed epoche determinati. Data la sua complessità, la biocenosi può essere suddivisa in fitocenosi, zoocenosi, micocenosi e batteriocenosi, se si considerano separatamente le piante verdi, gli animali, i Funghi e i Batteri, i quali hanno tutti funzioni ben distinte.
È difficile astrarre un sistema biologico dalle condizioni ambientali in cui si trova a funzionare. Questo è particolarmente vero per la biocenosi.
Una biocenosi integrata con il suo ambiente (clima e suolo) forma un ecosistema (la biogeocenosi degli autori sovietici); questo sistema biologico funzionale è una macchina dagli ingranaggi complicati, costituita da flussi, bilanci o cicli di energia e di sostanze chimiche diverse e da catene alimentari anastomizzate a rete. Il suo funzionamento più o meno efficiente porta alla produzione di una maggiore o minore quantità di materia organica. La quantità di materia organica prodotta nell'unità di tempo e di superficie costituisce la produttività; essa, accumulandosi di anno in anno, forma la biomassa dell'ecosistema considerato, che coincide con il peso secco di tutta la materia organica vivente che forma la biocenosi.
Sulla superficie dei continenti, dove domina largamente la biomassa vegetale (fitomassa), gli ecosistemi si riconoscono dalla fisionomia della loro fitocenosi (formazioni vegetali), che costituisce il quadro in cui vivono e si collocano le diverse comunità di organismi eterotrofi. Gli ecosistemi si classificano, secondo la loro fisionomia, in biomi: boschi, praterie, tundre, paludi, semideserti, strati di acqua dolce, ecc.
In seno agli oceani, dove generalmente la biomassa animale (zoomassa) è preponderante, gli ecosistemi sono molto più difficilmente delimitabili, ad eccezione delle coste rocciose, degli atolli e dei banchi corallini.
La superficie del globo terrestre è dunque coperta da un mosaico di ecosistemi, sorta di pellicola vivente che si sovrappone agli strati geologici minerali (litosfera) o che si mescola all'acqua degli oceani (idrosfera) e alla quale V.I. Vernadskij ha dato il nome di biosfera.
Nella biosfera composta da biomi in zonazioni regolari dal polo all'equatore, compare l'uomo: egli inventa l'agricoltura e la domesticazione degli animali; da questo momento in poi la biosfera viene modificata senza sosta: gli ecosistemi naturali sono sempre più frequentemente sostituiti da ecosistemi nuovi, derivati o interamente artificiali (come le città); il prodotto di questa straordinaria trasformazione della biosfera ad opera dell'intelligenza umana, che porta al progresso e al benessere, è stata definita da Vernadskij ‛noosfera'. Questa sfera biologica, all'interno della quale si inserisce l'intelligenza (νόος) dell'uomo, è sottoposta oggi ad abusi pericolosi dovuti all'incremento demografico e allo sviluppo tecnologico galoppante; si parla perciò di ‛tecnosfera'.
La conoscenza dei materiali biologici della cellula e dell'individuo riguarda soprattutto la biochimica, l'anatomofisiologia, l'etologia o scienza del comportamento, e la scienza del condizionamento genetico; invece popolazioni, comunità, ecosistemi e di conseguenza la biosfera e la noosfera formano l'immenso campo dell'ecologia, poiché in questi sistemi biologici si tratta sempre di rapporti degli organismi fra di loro e con il loro ambiente.
2. L'ecologia agli inizi del XX secolo.
L'ecologia è una scienza che esiste da sempre. Un agricoltore esperto, oppure un taglialegna, un pescatore, un cacciatore ugualmente esperti sono stati in tutti i tempi ecologi di valore anche se inconsapevoli. Infatti, scopo principale dell'ecologia è quello di fornire la sintesi di un numero immenso di conoscenze acquisite empiricamente nel mondo e di controllare tramite l'osservazione e l'esperienza le teorie elaborate.
Ricerche di carattere più spiccatamente accademico furono comunque intraprese all'inizio del sec. XIX da specialisti, soprattutto nel campo troppo spesso descrittivo della biogeografia. Dopo i primi incerti tentativi di numerosi precursori, si arrivò alla fondazione vera e propria della geografia delle piante con A. von Humboldt. Nel suo Essai sur la géographie des plantes (1805), egli mescolava alla botanica, alla fisiologia vegetale e all'agricoltura altre scienze che a quell'epoca non avevano con le prime alcuna connessione: la geografia, la climatologia, la chimica, la fisica terrestre, la storia. Nei suoi Prolegomena (1815), in cui trattava De distributione plantarum secundum coeli temperiem et altitudiniem montium, erano fissate definitivamente le basi di una scienza nuova, scienza di sintesi che spiegava la distribuzione delle piante in funzione dei fattori ambientali.
Von Humboldt aveva percorso soprattutto le Ande equatoriali. Il XIX fu il secolo della scoperta biologica del mondo; R. Brown verso l'inizio dell'Ottocento riportò dall'Australia una collezione di 4.000 specie, di cui i nove decimi erano nuove e specifiche di questo continente, dimostrando così, prima di tanti altri, la straordinaria diversità delle flore regionali.
Nel 1872 H.R.A. Grisebach, in Die Vegetation der Erde nach ihrer klimatischen Anordnung poté realizzare una descrizione monumentale della vegetazione dei cinque continenti, delimitando i territori fitogeografici (province) in base alla fisionomia della loro vegetazione dipendente dal clima. Ma Grisebach, pur stabilendo dei rapporti, non spiega niente. Perché una specie si armonizza nella forma, nelle strutture e nel comportamento fisiologico con le diverse componenti del clima in cui vive?
Numerosi autori si dedicano alla fitogeografia fisiologica; che indaga le cause dell'adattamento all'ambiente. Nel 1898 A.F.W. Schimper, con la sua opera Pflanzengeographie auf physiologischer Grundlage, fornisce una sintesi delle conoscenze acquisite nel corso del secolo, mentre fin dal 1882 J. Vesque aveva formulato la nozione di ‛efarmonia' per definire l'armonia tra la struttura morfologica delle piante e il loro ambiente. Infine, nella ormai classica Die Entwicklung der Pflanzengeographie, A. Engler nel 1899 aveva sintetizzato le numerosissime ricerche che avevano condotto a una conoscenza notevolmente approfondita dei territori e degli elementi fitogeografici del mondo.
Mezzo secolo prima, nel 1831, Ch. Darwin si era imbarcato sul Beagle per compiere il giro del mondo. Aveva così constatato che la distribuzione degli animali segue all'incirca quella delle piante. Studiando le isole Galapagos (1835) aveva trovato che in certe tartarughe o pesci, ad esempio, le specie sono strettamente adattate all'ambiente e alle abitudini di vita. Lo studio dei Vertebrati e la scoperta di numerosi fossili di Vertebrati di epoca recente in uno spaccato ovest-est del Sudamerica, gli mostrarono l'adattamento progressivo degli animali al mutare dei climi. Poco più tardi, nel 1858, Darwin contemporaneamente a A.R. Wallace avrebbe avanzato l'ipotesi, dimostratasi estremamente feconda, del trasformismo e dato vita alla teoria dell'evoluzione. Lo stesso Wallace nel 1876 stabilì i fondamenti di un'autentica geografia degli animali e divise il mondo in sei regioni biogeografiche o ‛regni' (v. Wallace, 1876).
La geologia seguiva un'evoluzione parallela che avrebbe permesso ai biologi, fino a quel momento attenti soprattutto al clima, di mettere in evidenza l'enorme importanza del substrato sulla distribuzione degli organismi viventi. I lavori sulle piante dette calcicole o calcifughe o silicicole non si conteranno più.
Le numerose ricerche di Fliche (1874-1888) sulle specie silicicole (Castanea vulgaris, Ulex europaeus, Erica cinerea, Calluna vulgaris, ecc.), caratterizzate dal fatto che non possono frenare l'assorbimento del calcio e lo concentrano in quantità tossiche allorché il suo tenore nel suolo è alto, gettano le basi di una disciplina che conoscerà un notevole sviluppo nella seconda metà del XX secolo, la biogeochimica.
Fra gli organismi e le rocce si sviluppa, per decomposizione fisica e chimica di queste ultime, una stretta zona intermedia, cioè il terreno, ambiente molto complesso al quale colloidi minerali (argilla) e organici (humus) conferiscono proprietà speciali e una enorme importanza. In Russia, V.V. Dokučaev (1846-1903) mostra che il terreno si forma sotto l'azione dei vegetali e degli agenti atmosferici e può essere classificato in zone secondo la latitudine. È la nascita della pedologia, che H. Del Villar proporrà in seguito di chiamare ‛edafologia', per evitare ogni confusione etimologica.
Tutta la pedologia moderna, tanto la chimica, la fisica e la biologia del terreno quanto i processi della loro formazione a partire dalle rocce madri, tanto la classificazione dei terreni quanto le relazioni di questi con la loro vegetazione, si trovano nel Bodenkunde di E. Ramann.
Agli albori del XX secolo, fisiologia e geografia delle piante e degli animali, trasformismo, climatologia e pedologia hanno conosciuto progressi sufficienti per il delinearsi di una scienza di sintesi, che sarà appunto l'ecologia moderna. La sua nascita si può far risalire a J.E.B. Warming, cui si deve, nel 1896, la pubblicazione del volume Lehrbuch der ökologischen Pflanzengeographie: eine Einführung in die Kenntnis der Pflanzenvereine. In quest'opera lo studio dei gruppi vegetali è largamente sviluppato in funzione del loro adattamento morfofisiologico all'ambiente. I principali gruppi trattati sono le Idrofite, le Xerofite, le Alofite e le Mesofite. Prende l'avvio lo studio dei fenomeni di competizione e di successione.
Quasi egualmente importante dell'ecologia di Warming è la ‛bioclimatologia' razionale, nata nel 1901 con l'opera di W. Köppen Versuch einer Klassifikation der Klimate, vorzugsweise nach ihren Beziehungen zur Pflanzenwelt.
Per esempio i climi megatermi, o climi caldi delle pianure tropicali senza stagione fredda e con temperatura del mese meno caldo superiore a 18 °C, comprendono: a) il clima delle liane, più o meno senza periodo secco, con più di due metri di piovosità annua e uno scarto fra la temperatura massima del mese più caldo e la temperatura minima del mese più freddo inferiore a 6 °C; è questo il clima delle foreste sempreverdi, alte e ombrose, a sviluppo ininterrotto, dove dominano i palmizi e gli alberi a grandi foglie lisce, clima adatto alla coltura del cacao, della vaniglia, del pepe, del riso, del cotone, ecc.; b) il clima delle savane tropicali o del baobab, con un periodo secco di almeno due mesi, con meno di due metri di precipitazioni annue e scarto delle temperature suddette che raggiunge i 12 °C; questo clima è adatto alle colture del caffè, del sorgo, della manioca, dei banani, ecc.; anche il riso e il cotone si coltivano bene come nel clima precedente.
Köppen distingue anche i seguenti altri climi: a) xerofili, con vegetazione di steppe e di semideserti, con piante spinose, spesso sfavorevoli all'agricoltura; b) mesotermi (temperati-caldi del tipo mediterraneo), con olivi, brughiere, camelie, fucsie, alte savane di agavi e di aloe; c) microtermi (temperati-freddi), con il Nothofagus nell'emisfero australe, querce e betulle nell'emisfero boreale; d) echistotermi (freddi), dove non possono più vivere gli alberi.
Dopo un periodo di pausa si deve giungere al 1949, anno di pubblicazione del libro di W.C. Allee e altri (v., 1949), e al 1953, data di pubblicazione della I edizione del libro di E. P. Odum, Fundamentals of ecology (v., 19713), perché venga dato l'avvio alla nuova scienza ecologica.
3. Metabolismo dell'individuo e della popolazione.
Il metabolismo di un individuo e quello di una popolazione sono facilmente comparabili, come d'altronde quello di una comunità e di un ecosistema. È comunque necessario definirne in via preliminare i principali meccanismi (v. fig. 2).
In un determinato istante (t1), un individuo o una popolazione possiedono una biomassa data (B) che si può esprimere con il peso vivo (fresco) o il peso di materia secca (MS), trasformabile in chilocalorie (1 g di materia secca corrisponde più o meno a 4,2 kcal).
Il metabolismo è uno scambio complesso di materia tra l'individuo o la popolazione e l'ambiente, e comporta tre fasi (v. fig. 2). La prima di ‛consumazione' (C), per assorbimento diffuso o per ingestione di un nutrimento, esprimibile numericamente in kcal (luce, materia vegetale o animale); la seconda di ‛assimilazione' (A) di una parte dei prodotti consumati, mentre la parte dei prodotti non assimilati (NA) viene eliminata.
È da notare che non tutto il materiale disponibile per il nutrimento degli eterotrofi (ME) viene utilizzato, ma una parte resta non utilizzata (NU).
I materiali organici assimilati si classificano in due gruppi essenziali: 1) alcuni servono da materiale di costruzione necessario a elaborare la materia dell'organismo (anabolismo); piccole molecole si uniscono per formare macromolecole specifiche, che sono raggruppate a loro volta in strutture diverse; questo assemblaggio fu chiamato da A. Lwoff modellamento (patronisation); la materia elaborata forma la produzione netta (PN), che porta a un aumento dei tessuti, cioè della biomassa (T) ; 2) altri sono degradati (catabolismo) e svolgono la funzione di materiale energetico: la loro combustione (respirazione = R) fornisce l'energia necessaria per le sintesi chimiche che portano alla PN è per tutte le funzioni compiute dall'organismo (mantenimento di una certa temperatura, movimento, ecc.).
Ma il catabolismo porta anche alla formazione di rifiuti vari, spesso chiamati prodotti di escrezione: certi animali eliminano l'urina (U), che serve soprattutto a eliminare i cataboliti azotati; nelle piante si formano delle sostanze organiche secondarie complesse (terpeni, steroli, glucosidi, alcaloidi, ecc.), alcune delle quali hanno funzioni etologiche (ecomoni). Si ha dunque:
C = A + NA, e A = PN + R ( + U, se esiste)
C = PN + R + NA ( + U, se esiste), o
PN = C − R − NA ( − U, se esiste).
Teoricamente T (aumento di tessuti di nuova formazione che si aggiungono alla B iniziale) = PN, ma in pratica una parte di PN viene eliminata (El), sia sotto forma di parti morte (Mn) sia sotto forma di parti ingerite da altri organismi consumatori (Cn′); una parte può essere esportata (Esn), lasciando l'ecosistema sotto forma di organismi migranti o sotto forma di organi o prodotti disseminati all'esterno di esso (polline, tronchi di alberi, resina, ecc.). Quindi:
PN = T + Cn′ + Mn + Esn.
Bisogna qui chiarire che l'indice n (nuovo) si riferisce all'eliminazione di materiale captato dopo il tempo t1. L'indice v (vecchio) si riferirà invece all'eliminazione di materiale preesistente, per es. vecchie cortecce d'albero mangiate da animali, o vecchi rami che cadono.
La terza fase del metabolismo è rappresentata dalla separazione o eliminazione nell'ambiente di rifiuti di origine diversa: a) sostanze non assimilate (NA), che corrispondono o a sostanze rigettate tali e quali (nelle piante questo tipo di escrezione consta di sostanze mineralizzate, negli animali, invece, di prodotti rigurgitati) o a una defecazione, prodotti che vengono più o meno modificati dal passaggio nell'apparato digerente senza essere però assimilati; b) cataboliti (escreti): sono il risultato della respirazione (R) e di processi fermentativi; i rifiuti solidi e liquidi dell'organismo, almeno in certi animali, vengono eliminati con le feci, l'urina e il sudore e saranno perciò indicati con FUS (vi confluiscono anche prodotti NA); c) prodotti di secrezione o secreti, variamente utili all'organismo (per es. i succhi digestivi).
Alla fine del periodo di osservazione t2 si ha dunque:
T = C − NA − R − FUS − El.
Se T è positivo c'è una crescita; se T è stazionario c'è un mantenimento della massa corporea; se T è negativo c'è una diminuzione.
L'aumento effettivo di biomassa, cioè quello che nasce meno quello che muore, è:
ΔB = PN − Cn′ − Cv′ − Mn − Mv − Esn − Esv.
Se per ragioni di facilitazioni tecniche poniamo:
Mn + Mv = M
Cn′ + Cv′ = C′
Esn + Esv = Es,
si ha:
ΔB = PN − C′ − M − Es
e si ha anche:
ΔB = T − Cv′ − Mv − Esv.
Si vede che l'accrescimento T non corrisponde all'aumento di biomassa ΔB. La produttività netta può essere aumentata dalla immissione di individui iminigranti (Im).
La formula completa è dunque:
ΔB = PN − C′ − M − Es + Im,
ma nella maggior parte dei casi studiati Im è irrilevante.
4. La popolazione.
La popolazione è un sistema biologico formato da un gruppo di individui della stessa specie, che occupano un dato territorio in un momento determinato.
a) Numero e densità degli individui.
Fra le caratteristiche di gruppo peculiari della popolazione, c'è prima di tutto il numero di individui che la compongono; la densità è il numero di individui per unità di superficie (o di volume) al momento considerato: per esempio 10 capre per ettaro di pascolo, o 5•106 Diatomee per centimetro cubo di acqua di mare. Un altro modo di esprimere la densità è la biomassa, vale a dire il peso di materia secca degli individui per unità di superficie o di volume. Alcune popolazioni sono soggette a esplosioni improvvise (‛invasioni'; v. Elton, 1958); ciò avviene, per esempio, presso numerose popolazioni di Insetti che costituiscono in tal caso un flagello per l'uomo, ma avviene anche presso Vertebrati quali il topo e il lemming. Un caso particolare è quello del Flos aquae, che corrisponde a esplosioni di fitoplancton che colora la superficie dell'acqua con i propri pigmenti. Negli oceani alcuni Dinoflagellati rossi del genere Gonyalax sviluppano delle ‛maree rosse' che provocano eutrofizzazione delle acque e intossicazione di tutti i pesci a causa degli alcaloidi tossici che producono. Queste esplosioni possono essere naturali, ma più spesso derivano dall'azione dell'uomo, il quale introduce delle specie parassite in regioni nuove dove esse non trovano i loro nemici naturali (introduzione del coniglio e della cactacea Opuntia in Australia, che ne è risultata invasa). Questo tipo di introduzione può provocare reazioni a catena, come, per es., l'invasione di vaste aree della Nuova Zelanda da parte di rovi, i cui semi sono stati diffusi da un uccello, lo storno, introdotto in quell'isola dall'uomo. Di tali ‛rotture di equilibrio' solo più tardi si constatano i danni (v. Galoux, 1971).
b) Natalità, mortalità, crescita.
Altri attributi di gruppo della popolazione sono il tasso di natalità e il tasso di mortalità, cioè il numero di individui che nascono o muoiono in una popolazione nell'unità di tempo (anno), espresso in per cento (%) o in per mille (‰). Così il tasso di natalità dell'uomo è attualmente intorno al 34‰, e il tasso di mortalità intorno al 15‰.
Il tasso di accrescimento annuale è la differenza fra il tasso di natalità e quello di mortalità; per l'uomo è dunque oggi 34‰ − 15‰ = 19‰, o 1,9%. Questo porta ad una esplosione demografica dell'umanità tale che la popolazione mondiale raddoppia ogni 37 anni. È importante rilevare che il 19‰ è un valore medio: mentre nei paesi sviluppati il tasso di accrescimento è intorno al 10‰, in quelli in via di sviluppo può raggiungere il 35‰.
Poiché la Terra ha sempre la stessa estensione, molti pensano che sia urgente prendere in esame misure che tendano a un accrescimento zero (v. demografia).
c) Equilibrio e fluttuazioni; curve di crescita.
Le grandi densità di popolazione e le esplosioni sporadiche mostrano come le specie abbiano un potenziale biotico (capacità di riproduzione e moltiplicazione) enorme. Ogni specie in cui questo potenziale non incontrasse ostacoli potrebbe, attraverso una crescita esponenziale, coprire in poco tempo l'intero pianeta. Si pensi ai ratti, che hanno un potenziale biotico di tre gravidanze di sette piccoli ogni anno (600.000 ratti in tre anni da una sola coppia). Per fortuna si stabilisce un certo stato di equilibrio. Il realizzarsi del potenziale biotico viene frenato e in seguito fermato dalla resistenza dell'ambiente: mancanza di spazio e di nutrimento, autoinquinamento dell'ambiente e, soprattutto, competizione con le altre specie (competizione interspecifica). Elton (v., 1927) ritiene che i nemici siano più importanti della mancanza di nutrimento.
La resistenza opposta al potenziale biotico, la cui realizzazione sarebbe rappresentata da una curva esponenziale, dà luogo a una curva a S (curva logistica di Verhulst), con un plateau che nelle popolazioni naturali corrisponde a una popolazione ottimale, mentre in numerose popolazioni antropogene corrisponde in realtà a una popolazione massimale (di sopravvivenza biologica). A partire dal plateau la popolazione può evolvere in direzioni differenti: a) mantenersi allo stesso livello; b) aumentare lentamente, se si sviluppa un adattamento all'ambiente (v. fig. 3, curva 1); c) declinare e talvolta estinguersi; d) fluttuare regolarmente o irregolarmente (v. fig. 3, curve 2-4).
Sono ben note le fluttuazioni stagionali: per es., in primavera abbiamo il periodo della riproduzione, in cui la popolazione ha un potenziale biotico massimale, dopo di che si manifesta la resistenza dell'ambiente: piccoli mal curati, lotta per lo spazio e il cibo, predatori, malattie, climi sfavorevoli; ciò riconduce progressivamente la popolazione a un livello di base che si manterrà fino al successivo periodo di riproduzione.
Di maggiore entità sono le fluttuazioni cicliche; sono note a tutti le fluttuazioni annuali delle vespe, delle farfalle, delle pulci, della frutta; causa primaria è l'instabilità dell'ambiente (inverni rigidi, periodi di siccità, cicloni tropicali). Le fluttuazioni periodiche sono particolarmente curiose: oscillazioni cicliche ogni 9-10 anni della lepre polare e del suo predatore, la lince; ogni 4 anni della volpe polare e del lemming, ecc. Si è tentato senza grande successo di trovare una spiegazione a questo fenomeno attribuendolo alle oscillazioni climatiche dovute in particolare alle macchie solari. Oggi si ritiene che le oscillazioni cicliche siano legate al funzionamento stesso dell'ecosistema (ipotesi di Schulz del ciclo nutritivo).
Le fluttuazioni repentine (v. fig. 3, curva 4), con improvviso aumento della popolazione, sono particolarmente importanti quando si tratta di parassiti o di devastatori, che possono costituire un autentico flagello.
d) Struttura.
La distribuzione dei sessi o delle età (piramide delle età), la speranza di vita alla nascita o nelle diverse età, costituiscono caratteristiche strutturali della popolazione.
e) Metabolismo.
Si è visto nella fig. 1 che la popolazione ha un metabolismo; è percorsa da un flusso di energia e di sostanze elementari, presenta un bilancio idrico, possiede una produttività, ecc.
f) Relazioni con lo spazio; area.
La popolazione ha un certo limite e occupa una certa area, il che implica rilevanti relazioni spaziali che si possono studiare ponendosi da due punti di vista diversi.
1. Problemi dinamici. Certi individui si spostano, modificando continuamente la distribuzione spaziale della popolazione. Questo spostamento costituisce la dispersione (dispersal), che nelle piante avviene secondo numerosissime modalità; la dispersione ha in generale un carattere passivo: solo una parte dell'individuo (spora) viene disseminata dal vento, dall'acqua, dagli animali o dall'uomo. Presso gli animali la dispersione è più spesso attiva e gli individui nella loro interezza si spostano volando, nuotando e muovendosi sul terreno.
Perché intervenga una reale modificazione della distribuzione della popolazione è necessario che la dispersione (o disseminazione) venga seguita dall'insediamento: i botanici usano anche talvolta il termine di ‛ecesi' (F.E. Clements). L'insediamento di una popolazione comincia in genere con l'insediamento di un individuo, o di alcuni individui (necessità della coppia nelle specie con individui unisessuati), che in seguito si moltiplicano (insediamento della specie).
In questo modo c'è un'estensione (spreading) della popolazione, che può avvenire sia in maniera diffusa e vaga (propagazione), sia al contrario in una maniera precisa fra due o più punti ben delimitati geograficamente e più o meno lontani fra di loro (migrazione). A seconda del senso della migrazione, si parla di emigrazione o di immigrazione, o di migrazione di andata e ritorno (v. migrazione).
L'immigrazione massiccia di una popolazione in una regione nuova (‛esplosione') può divenire una invasione se questa popolazione esercita effetti negativi su determinate popolazioni autoctone della regione invasa.
L'estensione di una popolazione, favorita dai mezzi di dispersione o di disseminazione, è frenata o limitata da numerosi fattori ambientali. Si parla in questo caso di fattori limitanti (luce, temperatura, acqua, composizione chimica del mezzo ambiente, ecc.). Per certi valori di questi fattori l'ecesi non può avvenire e la popolazione non si può stabilire in quell'ambito.
2. Problemi statici; geografia botanica e zoologica. Ogni specie ha un'area globale composta da più aree particolari, di cui è necessario dunque fare una classificazione. Lo studio delle aree (areografia) permette di definire le condizioni bioclimatologiche dell'esistenza delle specie e di dividere il mondo in territori biogeografici.
L'area - o areale - di una specie, limitata climaticamente, fornisce notizie importanti sulla possibilità di acclimatazione della specie considerata in una regione nuova.
Il XX secolo ha visto un'enorme moltiplicazione di carte di distribuzione di specie animali e vegetali.
I fattori limitanti (autoecologia). Il mezzo abiotico nel quale vivono gli organismi, o ambiente, ha due componenti: il substrato e il clima.
Il substrato primigenio fu l'acqua, probabilmente dolce e certamente calda, degli oceani primitivi. Con l'estendersi della vita ai continenti, il substrato terrestre diviene la roccia madre geologica, più o meno decomposta e alterata in superficie in una massa più molle alla quale si mescolano dei detriti organici: il terreno. I fattori ambientali dovuti al terreno si definiscono ‛fattori edafici' (da ἔδαϕος, terreno).
Il clima è in gran parte condizionato dall'impatto dei raggi solari sull'atmosfera terrestre, mentre i differenti fattori climatici che ne risultano e che agiscono sugli individui sono: energia, luce, temperatura, acqua, vento, ecc. Questi sono modificati sulla superficie terrestre dalla topografia del terreno e soprattutto dalla pendenza e dall'esposizione, dando luogo a mesoclimi e microclimi.
Si sa bene che nell'emisfero boreale i pendii esposti a mezzogiorno ricevono una maggiore irradiazione solare che non le pianure o i pendii esposti a settentrione, cosicché è stato possibile piantare dei vigneti sul versante sud di certe vallate dell'Europa centrale.
I climi cambiano con l'altitudine, che determina ‛piani climatici' ai quali corrispondono piani di vegetazione.
1. Bioclimatologia. Nel 1953 Bagnouls e Gaussen proposero di definire le condizioni bioclimatiche di una regione in base all'entità della stagione secca. Oggi si è stabilito che un mese secco è quello in cui il totale mensile delle precipitazioni espresso in millimetri è uguale o minore al doppio della temperatura media espressa in gradi centigradi:
P≤2t.
Dai ‛diagrammi di Gaussen', che riportano comparativamente le variazioni mensili di P e di t, si può ricavare l'entità (durata e intensità) della stagione secca.
2. Mesologia. Certi fattori ambientali (‛fattori mesologici') limitano l'estensione di una determinata popolazione in situazioni di concorrenza con popolazioni meglio adattate.
Una legge fondamentale per questo tipo di studio è la ‛legge del minimo' (formulata da J. von Liebig nel 1840): per occupare un dato biotopo, una popolazione ha bisogno di una certa quantità di bioelementi nutritivi necessari al suo sviluppo; l'elemento il cui tenore si avvicina di più al valore minimo critico tende a limitare lo sviluppo della popolazione. Questa legge deve essere completata tenendo presente che deve essere rispettato un certo equilibrio fra alcuni elementi importanti e che la quantità di bioelementi necessari è spesso funzione di fattori fisici dell'ambiente. Va inoltre tenuta presente la ‛legge della tolleranza' (v. Shelford, 1913), secondo la quale lo sviluppo di una popolazione in un certo biotopo è controllato dalla deficienza o dall'eccesso qualitativi o quantitativi di diversi fattori ambientali, chimici e fisici, deficienza o eccesso che si avvicinano ai limiti di tolleranza della popolazione. Quindi l'eccesso di un fattore può essere dannoso (legge del massimo) così come lo è la sua deficienza, considerando la natura della popolazione esaminata.
In natura l'abbondanza di un fattore pur favorevole alla popolazione può causarne la scomparsa poiché dà inizio a una successione di eventi nella quale domineranno le popolazioni meglio adattate (per es., eliminazione di una popolazione di prateria da parte di popolazioni di foresta).
Anche i fattori mesologici sono stati oggetto di numerosissimi studi nel XX secolo.
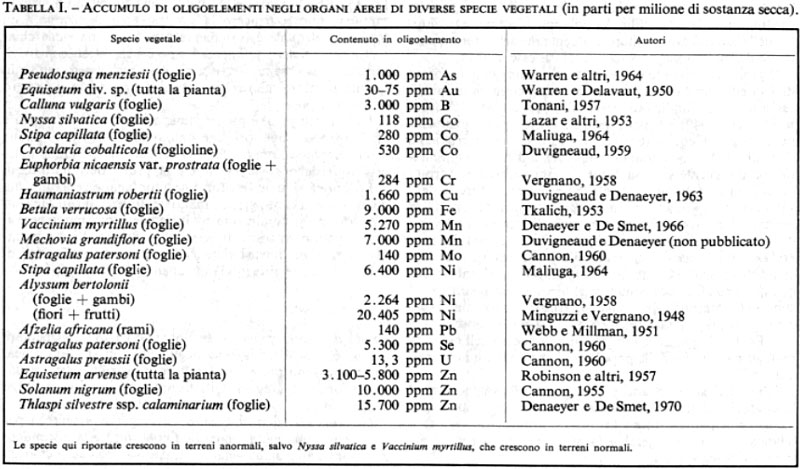
3. Biogeochimica. I componenti chimici del terreno sono spesso fattori limitanti di grande importanza. A causa del loro carattere più spettacolare, i più studiati sono i terreni anormali e la vegetazione che li ricopre.
I terreni anormali sono terreni la cui composizione chimica sembra costituire un fattore limitante essenziale per la vita delle piante. I tre principali tipi sono rappresentati: a) dai terreni a forte concentrazione salina (terreni salini o alcalini); b) dai terreni non equilibrati per difetto di uno o più elementi indispensabili, come per esempio terreni deficienti di P e Co in Australia, terreni poveri di K, terreni carenti di Mo (la deficienza di un elemento nutritivo può anche corrispondere a un eccesso di un altro: eccesso di Mg in rapporto al Ca nei terreni serpentinici); c) dai terreni non equilibrati per eccesso di uno o più elementi. Questi ultimi si distinguono in: 1) terreni anormalmente ricchi in polielementi: gessosi (eccesso di Ca e S), calcarei (eccesso di Ca), dolomitici (eccesso di Mg), serpentinici (eccesso di Mg); 2) terreni ricchi di metalli pesanti tossici: serpentinici (Ni, Cr, Co), calaminiferi (Zn, Pb), cupriferi o cobaltiferi (Cu, Co, Ni), manganiferi (Mn), seleniferi (Se); 3) terreni radioattivi (U).
Per quanto riguarda la vegetazione particolare sopra accennata, essa può essere distinta in piante indicatrici e piante accumulatrici. Le prime indicano con la loro presenza condizioni chimiche particolari del suolo. Esclusive o semplicemente preferenti, sono generalmente adattate geneticamente al loro substrato; può trattarsi di ecotipi puramente fisiologici o di specie nettamente differenziate. Tale aspetto genetico dell'adattamento può essere messo in evidenza tramite il trapianto di queste piante da un ambiente all'altro, con la misura dell'indice di tolleranza o mediante lo studio degli isoenzimi. Le piante indicatrici sono considerate ‛specialiste' per un particolare suolo (selenofite, cuprofite, zincofite, ecc.); fanno eccezione le erbacce (weeds) che invadono qualsiasi tipo di terreno, anche il più contaminato, e sopportano l'esposizione a radiazioni ionizzanti di alta intensità: queste specie sono definite ‛generaliste'.
Le piante accumulatrici si distinguono in piante accumulatrici propriamente dette e in piante concentratrici. Le prime assorbono in quantità massicce gli elementi presenti in eccesso nel suolo e li accumulano nei loro tessuti, esse cioè tollerano nelle loro cellule quantitativi anormali di uno o più elementi, tossici per le piante non adattate. La tab. I mostra che questo potere di accumulo esiste presso numerose specie vegetali e per gli elementi più diversi.
Le piante concentratrici possono accumulare nelle loro foglie uno o più elementi la cui concentrazione nel suolo non è elevata. Questo tipo di accumulo viene detto ‛concentrazione' e si osserva nei terreni più diversi per es. il Vaccinium myrtillus, pianta che si stabilisce nei terreni ricchi di humus acido delle foreste dell'Europa centrale, può accumulare più di 5.000 ppm di Mn nelle sue foglie (v. Denaeyer e De Smet, 1966).
I concentratori più straordinari sono alcuni Astragalus degli Stati Uniti (particolarmente Astragalus racemosus), le cui radici hanno il potere di estrarre il selenio immobilizzato sotto forma molto poco solubile in una roccia madre scistosa sottostante e di concentrano nei loro organi aerei; questi dunque convertono il Se minerale poco solubile in una forma organica idrosolubile, che ritorna al suolo e si accumula in superficie in forma assorbibile e tossica per la vegetazione selvatica e per le piante coltivate, che quindi regrediscono.
Fra gli animali si hanno numerosi esempi di specie concentratrici: uno dei più tipici è dato dalle Ascidie, che accumulano nelle proprie cellule sanguigne il vanadio.
4. Ecomorfologia. È lo studio dell'influenza dei fattori ambientali sulla struttura morfologica e anatomica dei vegetali. Sono stati proposti diversi tipi di classificazioni di eco-strutture in rapporto ad alcuni fattori limitanti (luce, temperatura, pressione osmotica, ecc.). Il sistema più usato è quello appresso schematizzato, che distingue i tipi biologici in base all'adattamento delle piante alla cattiva stagione e alla natura e al grado di protezione di cui godono i germogli persistenti.
Fanerofite: specie a germogli persistenti situati in alto; Camefite: specie a germogli persistenti situati a poca distanza dal suolo; Emicriptofite: specie a germogli persistenti situati all'altezza del suolo; Criptofite: specie a germogli persistenti situati sottoterra (geofite) o sott'acqua (idrofite); Terofite: piante annuali il cui sviluppo si compie in una sola stagione favorevole e che non passano la cattiva stagione se non allo stadio di seme. C. C. Raunkiaer (v., 1934) ha caratterizzato i climi delle regioni con il tipo biologico dominante: clima equatoriale, Fanerofite; clima temperato dell'Europa occidentale, Emicriptofite; clima sahariano, Terofite (da ϑέρος, bella stagione).
Se le specie si equilibrano, tanto nella morfologia quanto nella fisiologia, con le condizioni del loro ambiente, si deve ammettere che specie differenti, che vivono in uno stesso clima, debbano avere una tendenza a rassomigliarsi vegetativamente, a qualsiasi famiglia appartengano. È questo il fenomeno della convergenza, che si osserva molto spesso, come accade per es. con le piante cactiformi delle regioni semidesertiche dell'America tropicale (Cactaceae), dell'Africa orientale (Euphorbiaceae) e dell'Africa meridionale (Asclepiadaceae).
L'Africa meridionale presenta da NE a SO e da E a O tutte le successioni dei climi, dal clima tropicale umido fino al clima desertico primario. La regione non ha subito cataclismi geologici dopo l'era terziaria e la sua flora attuale è l'espressione di un'evoluzione ecologica che si è andata sviluppando secondo un gradiente di aridità crescente di specie sempre più adattate. A partire dall'albero equatoriale sempre verde e a larghe foglie indivise, l'evoluzione è avvenuta nella seguente direzione: a) riduzione delle dimensioni della pianta: b) aumento della ramificazione; c) riduzione delle dimensioni delle foglie, o produzione di foglie composte; d) caducità del fogliame; e) ramificazione più densa della nervatura delle foglie; f) sviluppo della sclerocaulia o della sclerofillia; g) sviluppo di spine; h) sviluppo della succulenza; i) aumento generale dello xerofitismo (cuticola, peli, fibre, produzione di oli essenziali).
Il punto culminante dell'evoluzione ecologica è la forma di vita effimera (‛effimerofite'); le piante effimere (annuali) resistono alle condizioni desertiche più estreme. A. Aubreville (v., 1949) ha riscontrato lo stesso tipo di evoluzione a nord dell'equatore e ha proposto lo stesso gradiente di aridità crescente andando dalla Costa d'Avorio al Sahara, con il seguente sistema di formazioni vegetali: a) foresta densa umida pseudoequatoriale (rain forest); b) foresta densa umida a foglie caduche (deciduous rain forest); c) foresta mista umida e secca (insieme di specie di foresta densa umida e di foresta chiara secca); d) foresta chiara secca a Isoberlinia-Uapaca (specie macrofille); e) foresta chiara secca a sottobosco di Combretacee; f) macchia a Combretacee con qualche albero dominante; g) foresta chiara con specie spinose (soprattutto Acacia microphylla); h) macchia di specie spinose; i) steppa subdesertica con specie spinose.
5. Ecofisiologia. La fisiologia consiste spesso nell'osservare degli individui o delle popolazioni in condizioni controllate.
I botanici si sono anche preoccupati di trasportare il laboratorio di fisiologia vegetale sul campo. I due argomenti principali di studio in questo ambito sono stati: il bilancio idrico dei vegetali e la fissazione del carbonio attraverso la fotosintesi.
g) Competizione interspecifica.
È il fattore limitante più importante: i giardini botanici e zoologici mostrano che numerosissime specie possono vivere nelle più diverse regioni se si sopprime la competizione con le specie autoctone meglio adattate. Il problema sarà trattato nel cap. 5, È g.
h) Competizione intraspecifica.
La competizione fra individui è una caratteristica universale delle popolazioni e si manifesta ogni volta che, da parte dei singoli, la domanda di spazio, luce, acqua o alimenti superi la disponibilità dell'ambiente.
Negli animali, la competizione dovuta a mancanza di spazio può essere accompagnata da reazioni psicologiche che risultano da una coabitazione troppo stretta.
Nelle popolazioni vegetali, si constata che alcuni individui prosperano e divengono dominanti, mentre altri, che presentano uno sviluppo notevolmente più lento, declinano, dovendosi contentare di ciò che i primi lasciano loro a disposizione, e soccombono se non vi è abbastanza per la vita di tutti. Nelle foreste, una tale competizione porta a un insieme di alberi dominanti e di alberi dominati, anche della stessa età; gli uni sono alti e robusti, gli altri sono bassi e deboli.
La vittoria nella competizione può essere dovuta alla fortuna (primo arrivato, primo a germinare, meglio sviluppato in condizioni di microclima favorevoli), ma molto più spesso questa vittoria è dovuta a una migliore costituzione genetica (sopravvivenza dei meglio adattati).
Nelle popolazioni di piante coltivate, nelle quali gli individui sono piantati a distanze uguali, la competizione porta generalmente non a una diminuzione del numero degli individui, ma a una riduzione simultanea della produzione di tutti gli individui. Così la competizione presso i vegetali è la risultante di due fenomeni: tendenza a una riduzione della vitalità degli individui, tendenza all'espansione e alla dominanza di individui più fortunati o meglio dotati geneticamente.
Sembra che il senso della proprietà sia molto sviluppato presso certi animali; un individuo, una coppia o un gruppo familiare può restringere la propria area di azione a un territorio al quale è fortemente attaccato, che viene chiamato home range; se l'home range è ben difeso contro gli intrusi, lo si chiama ‛territorio': esso può essere difeso in quanto territorio alimentare, zona di riproduzione, ovvero luogo di riposo.
La ‛territorialità' è più pronunciata presso i Vertebrati e gli Artropodi superiori, che hanno modi di riproduzione assai complessi comprendenti la costruzione del nido, la deposizione delle uova, la cura e la protezione dei piccoli.
I proprietari di un territorio ne allontanano generalmente gli intrusi con canti, grida e manovre diverse. Il rumore è un meccanismo generalmente molto efficace per mantenere il clan separato, si tratti di Scimmie o di Uccelli. Le battaglie sono rare; intrusi potenziali evitano generalmente di entrare in un territorio già occupato (v. etologia).
i) Aggregazione.
Grado di aggregazione. - In una popolazione elementare è raro che la distribuzione degli individui sia omogenea. Infatti possono presentarsi tre tipi di distribuzione: casuale, uniforme, aggregata. Quest'ultimo tipo è di gran lunga il più comune.
Nei vegetali, si riproducano essi vegetativamente o per semi, esiste una netta tendenza a concentrare la discendenza intorno alla pianta madre; lo stesso succede presso gli animali, i quali depongono i piccoli o le uova sempre nello stesso luogo e generalmente in gruppo; in questo caso può esserci una tendenza sociale al gruppo (congregazione).
Gli aggregati possono essere di dimensioni molto variabili e possono, a loro volta, essere distribuiti: a caso, uniformemente, in aggregati.
In fitosociologia, il grado di aggregazione (chiamato talvolta grado di socialità) può essere valutato tramite una scala di cinque gradini: a) individui isolati; b) gruppo ristretto; c) gruppo molto esteso; d) popolazioni estese lasse; e) popolazioni estese serrate.
In natura, l'aggregazione degli individui in seno alle popolazioni è dovuta a cause molto diverse: a) al tipo di riproduzione della specie; b) a differenze locali nell'ambiente, per es. a piccole discontinuità nel biotopo; c) a fattori climatici, per es. l'aggregazione di animali per resistere a un periodo freddo; d) a fattori biotici: gli individui si aggregano in luoghi ove essi sono protetti contro certi pericoli del mondo esterno; e) alla concorrenza di popolazioni di altre specie, in densa aggregazione, che producono nel biotopo una discontinuità di nicchie ecologiche; f) all'attrazione sociale presso gli animali.
L'aggregazione può aumentare la competizione fra gli individui per il cibo, lo spazio, l'acqua, ecc., ma questo inconveniente può essere controbilanciato da un aumento della sopravvivenza del gruppo. In effetti, gli individui raggruppati possono avere una minore mortalità rispetto agli individui isolati, durante periodi climatici sfavorevoli, o quando sono attaccati da altri organismi: la superficie e- sposta all'ambiente è minore in proporzione alla massa e il gruppo può modificare favorevolmente il microclima o il microhabitat.
È presso gli animali che si trovano i maggiori valori di sopravvivenza dovuti all'aggregazione degli individui. Un'aggregazione troppo forte, però, può portare talvolta a una sovrappopolazione pericolosa. Esiste dunque per ogni specie un grado di aggregazione ottimale che corrisponde a una crescita e a una sopravvivenza ottimali della popolazione, in condiziom date. Questo è il principio di W.C. Allee, secondo il quale la mancanza o, al contrario, l'eccesso di aggregazione possono essere fattori limitanti (v. Allee, 1931).
Congregazione. Società animali. - Gli animali si raggruppano generalmente per soddisfare esigenze fondamentali: possono raccogliersi intorno a una sorgente di nutrimento o di acqua; possono formare gruppi migratori alla ricerca di cibo; se si sentono deboli possono unirsi con animali più forti, per evitare l'attacco dei predatori; possono vivere da parassiti su piante o animali. Possono anche avere un forte istinto sociale che li porta a vivere strettamente raggruppati. Questo fenomeno di aggregazione volontaria, che conferisce alle popolazioni animali certi vantaggi sugli individui isolati, può essere chiamato ‛congregazione'.
L'adattamento sociale, che dà al gruppo un'unità di comportamento, gli permette di perfezionare l'adattamento per selezione naturale. Nei casi più evoluti le relazioni di cooperazione fra individui portano a una divisione del lavoro, con formazione di specialisti. In questa maniera da una popolazione non organizzata si passa a una popolazione più o meno altamente organizzata, che viene chiamata società (v. associazioni biologiche).
l) Eterogeneità e genetica delle popolazioni: l'ecotipo.
Un altro attributo del gruppo è l'eterogeneità della popolazione per certi caratteri morfologici o fisiologici degli individui che la compongono. Questa variabilità della popolazione è in rapporto alla variabilità genetica individuale. La variabilità in seno alla popolazione è il punto di partenza per l'evoluzione che avverrà per sopravvivenza dei più adatti in un ambiente che cambia.
La selezione naturale delle variazioni ereditarie può giungere fino alla comparsa di nuove specie, per trasformazione di una differenza quantitativa in una differenza qualitativa; il trasformismo si effettua in una direzione determinata, seguendo dei phyla prestabiliti.
All'interno della sua area, la specie sviluppa le sue popolazioni con un numero variato di individui che vivono in un luogo (biotopo), condizionati da fattori dell'ambiente circostante. Questi biotopi differenziati sono un'importante causa della discontinuità nei caratteri morfofisiologici medi delle popolazioni che li abitano; ogni biotopo raggruppa gli individui che gli sono meglio adattati per conformazione degli organi e abitudini di vita.
Da tutto questo risulta che popolazioni della stessa specie, ma appartenenti a biotopi diversi, si differenziano fra di loro per strutture e funzioni, e questo può essere attribuito a due cause distinte: a) l'accomodamento, facoltà che possiede una specie fortemente plastica di armonizzarsi alle condizioni ambientali, sviluppando dei caratteri non ereditari chiamati ‛modificazioni'; le popolazioni così ottenute sono dette ‛ecofeni'; b) l'adattamento, differenziamento di razze distinte, geneticamente adattate alle particolari condizioni ambientali, tramite lo sviluppo di mutazioni di carattere ereditario; le popolazioni così sviluppate sono degli ‛ecotipi' (v. adattamento).
Per decidere se si tratta di un adattamento genotipico ereditario, o di un semplice accomodamento dovuto alla plasticità fenotipica, bisogna trapiantare le specie da esaminare in ambienti omogenei e non variabili: gli adattamenti sussistono, gli accomodamenti scompaiono.
J. Clausen, D. D. Keck e W. M. Hiesey (v., 1940) hanno studiato il differenziamento ecotipico di diversi elementi della flora indigena della California, in un'area (transect) lunga 320 km che si estende dall'Oceano Pacifico fino alla Sierra Nevada. Le piante sono state trapiantate e studiate in tre giardini sperimentali situati a differenti altitudini, costituenti degli habitat standard (v. fig. 4). Si constatò, per esempio, che la specie Potentilla glandulosa si era differenziata in quattro ecotipi che differivano fra loro per numerosi caratteri sia fisiologici sia morfologici; i quattro ecotipi hanno valore di sottospecie. La sottospecie glandulosa è l'ecotipo costiero; la sottospecie reflexa occupa le colline ai piedi della Sierra e più in alto si trova nelle zone rocciose secche; la sottospecie hanseni si incontra nei pascoli verso i 1.500 m di altitudine; la sottospecie nevadensis occupa le zone alpine verso i 3.300 m. Questi quattro ecotipi sottoposti a trapianti reciproci mostrano che ciascuno è il meglio adattato al suo ambiente originario.
Il gruppo di specie Achillea borealis-lanulosa rappresenta un caso diverso, in quanto colonizza il transect mediante una serie continua di ecotipi (v. fig. 4); tali specie formano un notevole ventaglio di razze aventi periodicità stagionali che corrispondono a quelle dei loro diversi ambienti; esse crescono tutto l'anno nel clima sempre favorevole della pianura costiera; dormono d'estate nel clima mediterraneo, d'inverno nel clima alpino, ove sopravvivono grazie allo stelo corto.
In un ambito non vegetale, un caso classico è quello studiato da H.B.D. Kettlewell, riguardante il melanismo industriale delle farfalle (v. mimetismo): nelle regioni industriali l'inquinamento atmosferico annerisce i tronchi degli alberi distruggendone i licheni, che hanno colore grigio, e coprendoli di fuliggine. Fu constatato che 16 specie di farfalle ripartite in 12 generi svilupparono forme melaniche, poiché il loro substrato di riposo sono i tronchi degli alberi, e che, se esse non avessero presentato una forma di mimetismo cromatico nei confronti dell'ambiente, sarebbero state facile preda degli uccelli.
5. La comunità.
a) Definizione.
La comunità (o biocenosi, v. Moebius, 1877) è un sistema biologico formato da popolazioni che vivono entro un dato biotipo in un determinato periodo; benché composta da piante, da animali, da batteri, da funghi e da altri organismi viventi, tuttavia costituisce un raggguppamento di aspetto relativamente uniforme e di composizione floristica e faunistica determinata. Le popolazioni che formano una data comunità biotica vivono insieme in maniera ordinata e coordinata e non come organismi distribuiti a caso e indipendenti gli uni dagli altri (v. Clements, 1905 e 1916; v. Shelford, 1913).
b) Relazioni trofiche (alimentari) in seno alle biocenosi.
Gli organismi che elaborano materia organica a partire dai soli elementi minerali forniti dall'ambiente abiotico sono detti produttori. Costituiscono il punto di partenza delle catene trofiche, sequenze coordinate di organismi nel corso delle quali gli uni mangiano gli altri, prima di essere a loro volta mangiati. Queste catene trofiche legano fra loro il livello di produzione con più livelli di consumo, di riduzione e di trasformazione, durante i quali la materia dei cadaveri o dei rifiuti finisce per essere mineralizzata e ritornare al substrato e all'ambiente sotto forma di sali minerali o di elementi semplici abiotici (v. fig. 5).
Livelli di produzione primaria. - I produttori primari sono gli organismi che per fotosintesi, o più raramente per chemiosintesi, accumulano energia potenziale sotto forma di materia organica elaborata (glucidi, lipidi, protidi, vitamine) a partire da CO2, da H2O e da elementi minerali forniti dal mezzo abiotico. Questi sono per lo più le piante verdi micro- e macroscopiche.
Il livello di produttività primaria può essere costituito da una fitocenosi complessa o, al contrario, dalla popolazione di una sola specie dominante.
Livelli di consumo. - I consumatori si nutrono direttamente o indirettamente delle materie organiche prodotte dai produttori, liberando CO2 tramite la respirazione.
1. Consumatori di materia vivente. I consumatori di primo ordine (C1) si nutrono direttamente di produttori; essi sono gli animali erbivori (fitofagi); i consumatori di secondo ordine (C2) sono i carnivori, che si nutrono di erbivori (carnivori del primo livello); i consumatori di terzo ordine (C3) sono i carnivori che si nutrono di altri carnivori (carnivori del secondo livello). I consumatori di secondo e terzo ordine sono generalmente predatori che catturano la loro preda direttamente volando, correndo o nuotando. Alcuni poi sono parassiti e da sedentari sfruttano l'ospite parassitato, senza distruggerlo brutalmente.
2. Diversivori. Numerosi animali possono avere un'alimentazione variata ed essere allo stesso tempo erbivori e carnivori. L'uomo è il più completo diversivoro (onnivoro).
3. Consumatori di materia organica morta. A seconda che tale materia organica sia ancora più o meno fresca (cadaveri) o, al contrario, alterata, e ancora a seconda che essa sia di origine vegetale o animale, si differenziano in: a) necrofagi (sciacalli, avvoltoi, iene), animali consumatori di cadaveri di animali freschi o poco decomposti; spesso completano il ‛lavoro' dei carnivori ed è pertanto difficile distinguerli dai consumatori di secondo o terzo ordine; b) detritivori, i quali, a loro volta, si dividono in: saprofagi, animali consumatori di resti di piante (Lombrichi), o di resti di animali o vegetali molto alterati (xilofagi); saprofiti, piante consumatrici di resti vegetali (Batteri, Funghi, Fanerogame decolorate); coprofagi e coprofite, animali e piante consumatori di escrementi.
Livelli di trasformazione e di mineralizzazione. - I trasformatori decompongono la materia organica morta, rimaneggiata o no dai detritivori. Sono un'immensa popolazione di batteri e funghi che riducono infine i cadaveri o le deiezioni di tutti i tipi allo stato inorganico liberando CO2, NH3, H2S e sali.
Certi mineralizzatori (chemiotrofi) ossidano a sostanze più assimilabili dalle piante l'azoto e lo zolfo esistenti in forma ridotta. Certi batteri (solfatanti) trasformano i solfuri in solfati; i batteri della nitrificazione assicurano l'ossidazione dell'ammoniaca a nitrati.
Livelli di fissazione dell'azoto. - I fissatori di azoto (FN) hanno una posizione particolare; la loro nutrizione azotata avviene a partire da azoto molecolare N2. Ma come sorgente di carbonio e di energia devono utilizzare materie organiche più elaborate, che essi prendono dai detriti o dalla materia vivente delle radici o delle foglie di certi produttori. Questi organismi dunque sono eterotrofi per quello che riguarda il carbonio e autotrofi per ciò che concerne l'azoto.
Catene trofiche. - Si distinguono tre tipi di catene trofiche: a) le catene di predatori, che vanno dai produttori agli erbivori che si cibano di loro e che a loro volta saranno mangiati da piccoli carnivori, i quali saranno a loro volta mangiati da carnivori più grossi e così di seguito; a parte i grandi Cetacei, consumatori diretti di plancton, queste catene sono generalmente molto più lunghe nell'ambiente acquatico che non in quello terrestre; b) le catene di parassiti, che vanno dagli organismi di grandi dimensioni a quelli di dimensioni sempre più piccole; ci sono parassiti dei parassiti (iperparassiti; Pa2), per es. batteri o virus che infestano altri parassiti; c) le catene dei detritivori, che vanno dalla materia organica morta (detrito) agli organismi di piccole dimensioni; si è sempre più messo in evidenza l'importanza del micelio dei Funghi, che forma una sorta di ‛prateria' sul detrito, prateria abitata da un gran numero di animali micofagi.
Rete trofica. - Uno stesso produttore può servire da alimento a erbivori diversi e questi a loro volta a vari carnivori. Da ciò risulta nel bioma una molteplicità complessa di catene trofiche che si anastomizzano in una rete trofica (v. fig. 5).
Piramidi ecologiche. - In una catena di predazione, il numero degli individui, e in ogni caso la biomassa, diminuiscono da un livello trofico al seguente. Si possono così costruire delle piramidi il cui vertice è occupato da un carnivoro che controlla l'insieme (v. fig. 6).
c) Produttività della biocenosi; regolazione delle catene trofiche; ecomoni.
La fotosintesi permette di produrre nelle piante verdi una certa quantità di materia organica (assimilazione = produttività lorda PL), una parte della quale è respirata (RA), mentre il resto (produttività primaria netta PN1) serve a elaborare nuovi tessuti vegetali e nuove sostanze. L'equazione fondamentale è dunque:
PL = PN1 + RA oppure
PN1 = PL − RA.
Una parte più o meno rilevante della produzione pri maria PN1 serve ad alimentare le diverse catene di consumatori e di trasformatori ed è trasformata nella sostanza di questi (produttività secondaria PN2, terziaria PN3, ecc.); questa trasformazione esige una considerevole energia respiratoria RE (respirazione degli eterotrofi), ugualmente sviluppata a partire da PN1.
La produttività netta della biocenosi, cioè dell'ecosistema, è dunque:
PNE = PL − (RA + RE).
Quindi il funzionamento di una rete trofica porta a una produzione e a una ripartizione molto complesse, nella biomassa della biocenosi, dei materiali organici primari: protidi, lipidi, glucidi, acidi nucleici, ecc.
Ma al di fuori di questi materiali biologici primari, le diverse specie che compongono una comunità formano anche delle sostanze dette secondarie (glucosidi, alcaloidi, steroli, terpeni, ecc.), la cui funzione di ‛coadiuvanti etologici' non era stata ben compresa prima degli anni sessanta. Ilarkin ha creato nel 1960 il termine di ‛ecomoni' per le sostanze responsabili delle interazioni chimiche fra organismi della stessa specie o di specie diverse.
Effetti chimici intraspecifici. Le autotossine sono costituite da veleni, rifiuti, ecc., sostanze tossiche o inibitrici per gli individui della popolazione che li produce. Sono soprattutto importanti gli autoinibitori adattativi, che limitano la popolazione nel numero, in modo da evitare una densità eccessiva pregiudizievole.
I ‛feromoni' sono messaggeri chimici fra individui della stessa specie, che mettono in moto il comportamento sessuale o sono segnali di organizzazione o di difesa della società; agiscono in bassissime concentrazioni. Essi si distinguono in: a) feromoni sessuali (afrodisiaci e/o attrattivi); b) feromoni di riconoscimento sociale; c) feromoni di allarme e di difesa; d) feromoni di marcamento di territori o di piste.
Effetti chimici interspecifici (‛allelochimica'). - Gli ‛allomoni' danno un vantaggio adattativo all'organismo che li produce; si dividono in: a) repellenti (forniscono una difesa respingendo un attacco o un'infezione); b) sostanze di fuga (facilitano la fuga senza essere dei repellenti veri e propri, come per es. l'inchiostro dei Cefalopodi); c) sopprimenti (inibiscono o sopprimono altre specie competitive; comprendono gli antibiotici, le sostanze che frenano la germinazione di altre specie, e sostanze ormonali analoghe all'ormone giovanile degl'Insetti (v. ormoni negli invertebrati); d) veleni (avvelenano la preda); e) induttori (modificano la crescita di un'altra specie alla quale sono associati: formazione delle galle); f) antidoti e anticorpi (neutralizzano gli effetti di un veleno o di un altro agente aggressivo); g) attraenti (si distinguono in: richiami chimici, che attirano la preda verso il suo predatore; richiami per l'impollinazione).
I ‛kairomoni' danno un vantaggio adattativo all'organismo che li riceve e si possono classificare in: a) attraenti (segnalano la localizzazione del cibo); b) induttori (stimolano lo sviluppo adattativo dell'organismo ricevente); c) segnali di avvertimento (di pericolo o di tossicità, danno un vantaggio agli organismi che li ricevono); d) stimolanti (del tipo degli ormoni, vantaggiosi per l'organismo che li riceve, in quanto ne favoriscono la crescita).
I ‛deprimenti' inibiscono o intossicano senza vantaggio per il produttore (tossine batteriche).
I ‛cooperomoni' danno vantaggio adattativo sia al produttore sia a chi li riceve (impollinazione dei fiori da parte degli animali che vi trovano il cibo).
d) Controllo biologico delle popolazioni in seno alle comunità; lotta biologica; la biologia fiorale.
Predazione e parassitismo. - Le diverse popolazioni che formano una comunità sono unite da legami trofici che quasi sempre sono ricollegabili a fenomeni di predazione o di parassitismo.
Quando esiste una certa specificità fra la preda e il suo predatore, o fra l'ospite e il suo parassita, c'è il controllo di una popolazione sull'altra e il fenomeno può svilupparsi a catena. Venendo a mancare la fonte di nutrimento (preda e ospite del parassita), anche il predatore e il parassita muoiono. Predazione e parassitismo possono avere dunque anche degli effetti negativi sulle popolazioni che ne sono interessate. Queste relazioni sono state studiate ed espresse sotto forma matematica da A. J. Lotka (1925) e V. Volterra (1926) che le hanno rappresentate con equazioni (equazioni di Lotka-Volterra).
Generalmente gli effetti negativi sono poco importanti se le popolazioni che interagiscono hanno presentato un'evoluzione comune, in una biocenosi relativamente stabile: la selezione naturale tende a realizzare un equilibrio (equilibrio naturale) fra le popolazioni scelte per formare la comunità. Una forte interazione nociva (con rottura di equilibrio) si osserverà invece nelle biocenosi che subiscono un brusco cambiamento: casi di biocenosi modificate dall'azione dell'uomo in epoche recenti. L'introduzione accidentale o volontaria da parte dell'uomo di predatori o parassiti in regioni nuove per essi, dove trovino condizioni ottimali di sviluppo, provoca invasioni spettacolari accompagnate da rottura degli equilibri preesistenti: valga per tutti l'esempio dei vigneti europei distrutti dalla fillossera introdotta dall'America.
Spesso gli organismi sono tre, come nel caso di numerosi parassiti delle piante e dell'uomo che hanno bisogno di due ospiti distinti per compiere il loro ciclo vitale. Tale è il caso, per es., del plasmodio della malaria, che si alterna fra la zanzara e l'uomo.
La lotta biologica. - La lotta biologica consiste nel far combattere un flagello indesiderabile da parte di uno dei suoi nemici naturali, piuttosto che combatterlo con pesticidi chimici spesso pericolosi per l'ambiente perché agiscono alla cieca e possono intossicare le catene trofiche e provocare rotture di equilibrio. Per esempio: la cactacea del genere Opuntia era stata introdotta dall'America in Australia nel 1840; nel 1920 ayeva invaso 200.000 km2 del territorio australiano. Si raggiunse una riduzione del 75-90% tramite l'introduzione della farfalla Cactoblastis cactorum, il cui bruco si nutre di Opuntia; è stato così creato un equilibrio fra le due specie, che si sono andate sviluppando in proporzioni ragionevoli.
È contro gli insetti parassiti di colture e piantagioni che la lotta biologica assume aspetti particolarmente importanti (v. lotta biologica).
Oltre alla creazione di razze resistenti della pianta coltivata, si utilizzano i nemici più vari dei suoi parassiti nocivi (predatori come Insetti, Uccelli, Roditori, entomofagi; animali parassiti come Insetti, Nematodi, Batteri, virus e Funghi).
Un'altra forma di lotta biologica è data dall'uso di armi chimiche di origine biologica: gli attrattivi sessuali, che permettono di catturare e di distruggere in massa i maschi; l'ormone giovanile o ecdisone che blocca lo sviluppo o la riproduzione dell'insetto attaccato.
Infine, un'altra tecnica consiste nell'introdurre nella popolazione dell'insetto parassita maschi sterilizzati, di modo che una parte considerevole di femmine depone uova non fertili.
È ben nota l'utilizzazione di coccinelle contro cocciniglie: dal 1888, l'azione della cocciniglia Icera purcasi, che parassitava gli agrumi della California, fu combattuta e vinta con l'introduzione della coccinella Novius cardinalis, parassita degli agrumi e predatore proveniente dall'Australia.
Esempi di lotta microbiologica sono l'impiego del virus della mixomatosi contro l'eccessivo moltiplicarsi, in Australia, del coniglio selvatico, e quello di preparazioni di Bacillus thuringiensis contro numerose specie di bruchi parassiti.
Il problema della lotta biologica è in effetti molto complesso; l'arma biologica deve essere specifica, sufficientemente efficace e persistente, e deve avere il suo massimo di attività nel momento preciso in cui è da combattere il nemico. Deve integrarsi nell'ecosistema trovandovi delle stazioni-rifugio, all'occorrenza predisposte dall'uomo, dove ne sia assicurata la sopravvivenza. Deve giungere a un controllo permanente, cioè a uno stato di equilibrio (in suo favore) con il nemico.
La biologia fiorale. - I rapporti mutualistici tra i fiori e gli animali che compiono l'impollinazione procurandosi il cibo hanno un'importante azione positiva nel mantenimento delle popolazioni cui si riferiscono.
Un gran numero di lavori specialmente in questi ultimi decenni ha mostrato l'importanza ecologica e genetica dei meccanismi dell'impollinazione (Pechoutre, 1909; Von Frisch, 1954; Kügler, 1955; Werth, 1956).
Nel 1970 è stata proposta da G. L. Stebbins una teoria dell'evoluzione delle piante da fiore per adattamento agli animali impollinatori e anche agli agenti fisici, come il vento (anemogamia) e l'acqua (idrogamia).
Simbiosi a livello delle radici; noduli fissatori di azoto e micorrize. - In certe circostanze, si vedono apparire sulle radici di piante superiori delle protuberanze che risultano da fenomeni di simbiosi: sono i noduli fissatori d'azoto e le micorrize.
Nelle Leguminose, le radici sono invase da ‛cordoni' costituiti da batteri del genere Rhizobium. Questi cordoni inducono l'ipertrofia, sotto forma di noduli, di certi tessuti della radice dentro le cui cellule si liberano i batteri. All'interno di queste cellule invase si compie, con l'intervento di un pigmento rosso del tipo dell'emoglobina, un'intensa fissazione di azoto atmosferico. La vita dei noduli è effimera: essi presto si decompongono e mettono in libertà nel terreno grandi quantità di azoto, che entrano nella catena di nitrificazione.
Le micorrize sono prodotte da funghi simbionti; possono essere endotrofe ed ectotrofe. Le prime, poco deformate o semplicemente ‛tubercolizzate', sono penetrate dal micelio, che sviluppa nelle cellule corticali delle vescicole caratteristiche (sono tipiche della famiglia delle Orchidacee). Le ectotrofe sono deformazioni spesso coralloidi circondate dal micelio del fungo (caratterizzano le radici degli alberi in difficoltà alimentari). I vantaggi procurati da questi ultimi simbionti sono l'assorbimento delle sostanze minerali (soprattutto N e P bloccati nell'humus) e l'assorbimento diretto delle sostanze organiche elaborate dal fungo stesso (per es., l'acido glutammico, che il boleto cede al pino). D'altra parte anche il fungo trae vantaggio da questo tipo di vita, poiché riceve dall'albero alcuni materiali energetici che gli sono necessari (glucidi) e che è incapace di sintetizzare. I due simbionti si scambiano anche alcuni fattori di crescita. L'argomento delle micorrize ha un'importanza considerevole in silvicoltura.
e) Struttura nello spazio e nel tempo; stratificazione e periodicità; aggregazione.
In una biocenosi, gli organismi sono distribuiti nello spazio in maniera da utilizzare il più efficacemente possibile le condizioni che sono loro offerte dall'ambiente. La posizione di certe popolazioni è dettata dalla ricerca di protezione contro un fattore ambientale svantaggioso o contro un nemico (piante sciafile, animali arboricoli). Altri rapporti corologici si stabiliscono fra specie in competizione per la luce, il cibo o l'acqua.
Tutto ciò porta a una strutturazione spaziale in strati orizzontali più o meno numerosi, aerei e sotterranei. In una foresta, la stratificazione aerea (strato arborescente, arbustivo, erbaceo, muschioso) corrisponde a un'utilizzazione razionale dell'energia solare; la stratificazione degli organi sotterranei risponde a uno sfruttamento razionale delle ricchezze alimentari e idriche del suolo.
Nelle biocenosi acquatiche si ritrova questa stessa stratificazione ispirata alla luce, alla temperatura, alla tensione di ossigeno, ecc. Infine la strutturazione nello spazio si completa con la strutturazione nel tempo o periodicità (fenofasi).
La periodicità, così netta a livello delle fitocenosi, per esempio il periodo della fogliazione, quello della fruttificazione ecc., esiste anche a livello delle zoocenosi; numerosi animali componenti una comunità non hanno mai l'occasione di incontrarsi poiché esplicano le loro attività in momenti diversi. Le alternanze possono essere relative alle stagioni o alle diverse ore del giorno. Citiamo l'esempio degli uccelli migratori, il cui soggiorno corrisponde all'enorme moltiplicazione estiva delle popolazioni di Insetti, che dipende direttamente dall'aumento della temperatura e indirettamente dallo sviluppo fenofasico della vegetazione.
Nella foresta è facile distinguere specie diurne (talpe, sparvieri, farfalle, formiche, api, ecc.) e specie notturne (uccelli notturni, pipistrelli, falene, ecc.).
Nella biocenosi, gli strati sono essi stessi strutturati in base alla diversità delle popolazioni che li compongono e al grado di aggregazione che presentano. La diversità è diventata in questi ultimi anni uno dei capitoli più importanti dell'ecologia, poiché sembra andare di pari passo con la stabilità delle biocenosi e con il mantenimento degli equilibri naturali.
f) Nozione di nicchia ecologica.
Ch. Elton, nel 1926, creò il termine ‛nicchia ecologica', che designa sia lo spazio occupato da una popolazione nella biocenosi sia la funzione che essa vi compie. La nicchia di un animale è data dal suo posto particolare nell'ambiente e dalle sue relazioni nei confronti del cibo e dei nemici. Essa spesso può essere definita in base alla dimensione dell'animale e alle sue abitudini alimentari. Esistono specie che presentano una nicchia ecologica molto ristretta (‛specialisti', come per es. gli Insetti che si nutrono di polline, o i Colibrì bottinatori di nettare); ed esistono specie che presentano una nicchia molto ampia (‛generiche', di cui l'uomo, onnivoro, è l'esempio migliore).
Una biocenosi è un intrico e un incastro di nicchie ecologiche diverse; la tendenza a che ogni nicchia sia occupata da un'unica popolazione determina una selezione naturale. Come i vari continenti e gli oceani hanno la loro flora e la loro fauna particolari, delle biocenosi analoghe (isocenosi) hanno le loro nicchie ecologiche occupate da popolazioni differenti che appartengono a taxa completamente diversi (convergenza): il bisonte, il canguro e l'antilope saiga (animali da pascolo) occupano la stessa nicchia ecologica, rispettivamente nelle steppe americane, australiane e asiatiche.
Il concetto di nicchia ecologica può essere esteso a una parte o a tutto l'insieme della biosfera e comporta allora delle zone di adattamento, molto diverse ecologicamente; il passaggio di un gruppo da una zona all'altra porterebbe a un adattamento generale, responsabile della ‛macroevoluzione'. Ne è un esempio l'evoluzione dei cavalli, che furono inizialmente piccoli animali brucatori a piedi digitati, prima di passare nella steppa e divenirvi grossi animali pascolanti, con zoccoli.
Allorché un gruppo può stabilirsi in una zona nuova, vi scopre un insieme di nicchie ecologiche diverse, di modo che l'evoluzione vi si presenta come un ‛irradiamento adattativo'. Il caso dei Marsupiali, che in Australia imitano nelle loro abitudini e nelle loro forme i Mammiferi dell'emisfero boreale, è particolarmente probante (v. fig. 7).
g) Competizione interspecifica o coabitazione.
In una biocenosi si instaura una competizione serrata fra una specie e il suo predatore, tale però da mantenere in equilibrio sia la preda sia il predatore, il quale ha soprattutto la funzione di eliminare i malati e i peggio adattati della specie di cui si nutre.
Esistono anche rapporti più generali di competizione o di coabitazione fra specie dello stesso livello trofico, adattate alla stessa nicchia ecologica dal punto di vista delle necessità di luce, temperatura, sostanze nutritive, ecc. Si tratta, dunque, di interazioni molto complesse che dipendono dalla densità di popolazione.
L'indipendenza fra specie esiste solo quando la densità è estremamente scarsa (‛pionieri' dispersi su una superficie nuda). La lotta fra i candidati all'occupazione di una nicchia, che determina una vera selezione, l'una ‛selettiva' l'altra ‛eliminatoria', evolve verso due direzioni:
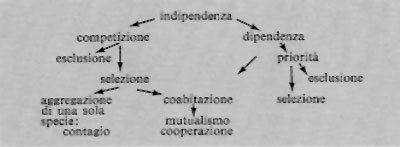
La dipendenza è la relazione fra popolazioni con forme di vita differenti, nella quale una riceve un beneficio, non reciproco, dall'associazione con l'altra (liane, epifite, animali che nidificano fra i rami degli alberi, saprofiti legati a certe specie determinate, ecc.). Essa è dunque legata alla stratificazione e porta alle ‛sociomorfosi'.
La competizione è la concorrenza che si stabilisce fra tutte le specie capaci di occupare un certo territorio, ma di cui solo una parte sarà ammessa a installarvisi, mentre le altre ne saranno escluse.
In una biocenosi equilibrata, esiste in teoria una sola popolazione per nicchia ecologica, come conseguenza del fenomeno darwiniano della selezione del più adatto nella lotta per l'esistenza (principio di G.F. Gause).
Se si tratta di vegetali la competizione, dopo l'eliminazione dei meno resistenti, porta spesso alla coabitazione fra varie specie che vivono nelle stesse condizioni di vita formando un ‛gruppo socioecologico'; i fattori decisivi del successo sono, come ha mostrato Ellenberg, la costituzione morfologica e la costituzione fisiologica.
Bisogna distinguere bene: la coabitazione può essere considerata come l'occupazione di nicchie ecologiche distinte nella stessa biocenosi; se certe risorse sono presenti in sovrabbondanza in una nicchia ecologica, le possono utilizzare nello stesso tempo molte specie. Queste hanno spesso la stessa forma di vita e costituiscono delle ‛specie omologhe', che da una parte possono coabitare ma dall'altra possono sostituirsi le une alle altre, senza modificazioni di rilievo del manto vegetale. Si parla di mutualismo (cooperazione) allorché gli organismi dipendenti o coabitanti contribuiscono al benessere reciproco
Nella competizione trionfano i più adatti ai fattori limitanti dell'ambiente. Ciò nonostante è raro che l'optimum fisiologico (ottenuto in coltura pura) coincida con l'optimum ecologico in seno alla biocenosi (v. fig. 8); a causa della competizione, l'optimum ecologico è spostato verso il minimo o il massimo, o verso entrambi, formando una curva a due picchi (v. fig. 8, curva B).
Gli effetti della competizione possono essere modificati dal fatto che una specie modifica l'ambiente in cui vive: essa lo rende più favorevole a un competitore, che finirà per prendere il sopravvento. Per es. le piante casmofite, colonizzatrici dei terreni rocciosi, si insinuano nei crepacci e fra le rocce, da cui per sgretolamento ricavano un po' di terra, che trattengono attorno alle radici; ben presto questo stesso terreno sarà invaso da altre specie il cui dinamismo eclisserà i primi colonizzatori, e cosi di seguito. Si è così iniziata e sviluppata una successione. La successione si sviluppa naturalmente anche quando delle piante di una certa altezza non possono resistere a piante più alte, che sottraggono loro la luce necessaria alla fotosintesi.
h) Dinamismo.
Una fitocenosi si modifica nel corso del tempo. Se si parte da un biotopo vergine, roccia nuda, isola vulcanica, duna di sabbia o più semplicemente colture abbandonate (old field, negli Stati Uniti), vi si osserva una successione primaria di comunità sempre più complesse, aventi dimensioni sempre più grandi e biomassa sempre più imponente. Attraverso stadi intermedi più o meno numerosi si passa dalla fitocenosi pioniera a una fitocenosi terminale più o meno fluttuante, ma nondimeno fissa e presentante il massimo di biomassa in rapporto al clima, al periodo e al terreno considerati. Una biocenosi in equilibrio con il clima viene chiamata ‛climax'. Se si aggiungono gli animali al climax, ne risulta un ‛bioma'.
Il climax si mantiene per tutto il tempo che dura il clima al quale corrisponde, cioè talvolta anche milioni di anni (praterie americane); allorché intervengono grandi cambiamenti climatici, o semplicemente si esercita l'azione umana, il climax è sostituito da un nuovo climax o da una vegetazione antropogena.
Il climax (o il bioma) è la biocenosi più stabile e anche la più diversificata di una successione dinamica, in certe condizioni climatiche (v. fig. 9). (Si vedano a questo proposito i contributi di Clements).
Una successione secondaria si sviluppa allorché una vegetazione primaria è distrutta o profondamente modificata dal diboscamento, dal dissodamento, dagli incendi, dai denti degli animali domestici; si giunge poi ad un climax che, molto spesso, non corrisponde al climax ideale, poiché il terreno, più o meno degradato in partenza, non è più capace di evolversi in suolo di climax. Questo climax ‛potenziale' (‛plesioclimax', secondo Gaussen) è il solo che si possa concepire in una regione molto popolata o fortemente modificata dall'uomo.
i) Ampiezza e limiti; l'ecotono.
Il senso dato al termine biocenosi dipende dall'ampiezza dell'unità considerata; nel caso di una foresta, per esempio, il termine di biocenosi si applica tanto alle popolazioni che colonizzano un semplice ceppo, quanto alle comunità costituenti la foresta nel suo insieme; si devono dunque considerare delle ‛biocenosi maggiori' e delle ‛biocenosi minori'.
Una biocenosi maggiore ha dimensioni e organizzazione sufficienti a renderla relativamente indipendente dalle biocenosi adiacenti; integrata nel suo ambiente, essa sopravvive finché le è fornita in sufficienza l'energia solare.
In natura la zona di contatto fra due biocenosi differenti costituisce un ‛ecotono' dove gli effetti dei legami possono essere assai notevoli (flora e fauna speciali e particolarmente ricchi). Talvolta biocenosi adiacenti si fondono progressivamente l'una con l'altra, (ecotono largo) senza che si abbiano fra di esse dei limiti ben definiti (continuum).
l) La fitosociologia.
Il paesaggio di una certa regione è composto da un mosaico o da una zonazione di fitocenosi, che si riconoscono per la loro fisionomia particolare (formazioni).
Una formazione è un raggruppamento vegetale la cui fisionomia è determinata dal prevalere di uno o più tipi di forme vitali. Negli studi fitosociologici si usano unità più piccole: l'associazione vegetale, fitocenosi dalla composizione floristica determinata che presenta una fisionomia uniforme e cresce in condizioni ambientali ugualmente uniformi. L'associazione vegetale, in cui ogni specie ricerca ‟il suo profitto esclusivo, è - come dice Ch. Flahault - l'espressione della concorrenza vitale e dell'adattamento all'ambiente di un gruppo di specie". ‟La lotta per l'occupazione del suolo - asserisce P. Jaccard - ecco il fattore sociale per eccellenza...". L'idea di una coabitazione di specie, con legami di dipendenza o di mutualismo, viene generalmente a completare quella di un semplice avvicinamento fortuito. Si arriva a pensare che l'associazione vegetale agisca come un blocco unico nei confronti dei fenomeni competitivi e dinamici.
In natura, una certa associazione deve formarsi in tutte quelle aree in cui i fattori ambientali le sono favorevoli (v. associazioni biologiche).
Tendenze della fitosociologia. - Diverse tendenze si sono sviluppate nella fitosociologia. Si può addirittura parlare di scuole di fitosociologia. Secondo la Scuola di Uppsala (di cui Du Rietz è il maggior rappresentante), l'associazione è basata sulla costanza-dominanza; è una fitocenosi di composizione floristica realmente stabile e omogenea, in cui ogni strato è caratterizzato dalla presenza di costanti- dominanti. Un tale concetto fa dell'associazione un insieme estremamente rigido, quasi matematico.
Secondo la Scuola di Zurigo-Montpellier di Flahault e Braun-Blanquet, il carattere più importante è la fedeltà: l'associazione, che è più o meno stabile, è caratterizzata da specie che le sono fedeli.
I gradi di fedeltà sono i seguenti: 5) specie esclusive; 4) specie elettive; 3) specie preferenti; 2) specie accessorie; 1) specie accidentali. Le specie da 5 a 3 sono dette fedeli, le altre indifferenti, presentando una grande possibilità di diffusione. In ogni caso, si tratta di specie rare, che vivono in un ambiente particolare (terreni avvelenati da metalli pesanti, o asfissianti, ecc.).
Un'associazione è abitualmente designata con il nome generico latino di una specie dominante o caratteristica, seguita dal genitivo del nome specifico, per es.: Fagetum silvaticae, Bidentatum tripartiti.
Si accennerà adesso alle scuole dinamiste, e in particolare a quelle francesi (rappresentate da Kunholz-Lordat, da Gaussen e da Rey) e a quelle anglosassoni, il cui esponente principale è Clements (v., 1907). Questi considera il climax come un organismo complesso e inseparabilmente legato al suo clima; esso costituisce la principale unità di vegetazione e forma la base della classificazione naturale dei gruppi vegetali. Nel sistema di Clements l'unità di vegetazione che corrisponde al climax generale di un territorio è la ‛formazione', per esempio quella a Quercus-Fagus (foresta caducifoglia) delle praterie dell'Ovest; essa è caratterizzata da specie predominanti che si ritrovano in tutta la sua estensione: otto specie di Quercus e quattro specie di Carya. Seguendo i principali tipi di climi regionali, la formazione è allora divisa in associazioni, caratterizzate dalla dominanza locale di due o più specie nello strato superiore. Cosi la formazione a Quercus-Fagus comprende: l'associazione a Quercus-Castanea dentula; l'associazione a Fagus-Acer saccharum; l'associazione a Quercus-Carya (v. fig. 9).
Mentre Clements considera la comunità come un organismo biologico, secondo Gleason, capostipite della scuola americana moderna, in natura le popolazioni delle diverse specie si sviluppano e reagiscono l'una indipendentemente dall'altra: l'associazione non è un'unità di vegetazione, ma la coabitazione per pura coincidenza di popolazioni selezionate dall'ambiente. Da tale ipotesi individualistica della vegetazione si è sviluppato, a partire dal 1950, il concetto di continuità della vegetazione (continuum), che si contrappone alla nozione classica di associazione implicante una discontinuità del tappeto vegetale. Più di recente i due concetti si sono riavvicinati, dando luogo alla nozione dialettica di ‛discontinuità nella continuità', che mette tutti d'accordo ed è la base di rilevamenti fitosociologici più completi, riguardanti territori nella loro globalità.
m) Comunità animali.
Le comunità animali meglio studiate sono le comunità marine e quelle di acqua dolce. È d'altronde in queste comunità che la biomassa animale può dominare largamente la biomassa vegetale, poiché questa è prevalentemente costituita da fitoplancton a ricambio veloce.
Inoltre le condizioni dell'ambiente acquatico sono molto più facili da studiare che non quelle dell'ambiente terrestre; i numerosi studi compiuti sulle comunità acquatiche ben integrate nel loro ambiente circostante hanno dato inizio alla moderna ecologia basata sullo studio degli ecosistemi (v. Lindeman, 1942).
Le comunità animali terrestri sono state oggetto di molti studi per lo più a sé stanti (v. Allee e altri, 1949); è tuttavia difficile isolare queste comunità dal quadro generale delle comunità vegetali che forniscono loro il cibo.
Delle rassegne molto importanti sulle comunità di Vertebrati terrestri sono state pubblicate recentemente (v. Balogh, 1958). Temi principali sono: gli insetti defolianti delle colture e delle foreste, i grandi erbivori delle praterie, la fauna del suolo (pedofauna detritivora), che assicura il ricambio della materia organica.
Il ruolo della pedofauna nella fertilità dei terreni tropicali è stato oggetto di studio approfondito da parte di Maldague.
n) Microbiocenosi.
La microbiologia del suolo, iniziata nel secolo passato, fu codificata all'inizio del XX secolo da Löhnis e in seguito sviluppata da Waksman (v., 1927 e 1952).
Un oggetto recente dello studio delle microbiocenosi è la rizosfera, accumulo di batteri e funghi intorno alle radici delle piante superiori, la cui importanza per la vita di queste ultime si dimostra ogni giorno di più. Le azioni antagoniste dei microrganismi del suolo, che realizzano un equilibrio in seno alle loro comunità, sono esplicate tramite gli antibiotici, un gran numero dei quali è stato oggi isolato. Certi antibiotici degli Actinomiceti (streptomicina, clortetraciclina, ossitetraciclina, cloramfenicolo) hanno trovato un'enorme applicazione pratica in medicina (v. chemioterapia antibatterica). I funghi hanno dunque un'importanza enorme, tanto che senza di essi nella biosfera la vita delle comunità vegetali e animali sarebbe impossibile.
Alcuni autori isolano le specie di batteri (v. Burges, 1967), li contano e li pesano: in media circa 1 g di batteri per m2 di suolo (cioè da uno a parecchi miliardi per g di terreno), da 1 a 10 g di funghi per m2 di suolo (fino a 10 km di micelio per g di terreno).
o) Formazioni vegetali del globo; i grandi biomi.
Alle grandi zone climatiche che si succedono, allineate da est a ovest, dai poli all'equatore, corrispondono grandi tipi zonali di vegetazione, che sono le grandi formazioni vegetali del pianeta e che sui continenti hanno una loro fisionomia, determinata dalla dominanza di certe forme di vita.
Queste formazioni vegetali, chiamate ‛biomi' (Clements, 1936; Odum, 1954) sono il quadro della vita e della evoluzione di innumerevoli popolazioni di batteri e di animali; la popolazione umana, in rapido incremento, marca sempre di più le biocenosi maggiori, per la designazione delle quali ci si è messi d'accordo verso il 1970, nell'ambito dell'IBP (International Biological Program).
Viene oggi universalmente usato un sistema di classificazione proposto da Rübel (v., 1930), leggermente modificato, basato sulle conoscenze delle piante che si avevano fin dall'inizio del secolo. Ecco una classificazione preliminare molto generale: 1) lignosa: formazioni composte da piante più o meno lignificate; a) silvae: formazioni forestali; b) fruticeta: formazioni arbustive; c) suffruticeta: formazioni di sottoarbusti del tipo lande a brughiera; 2) herbosa: formazioni composte da piante erbacee, terrestri (terri-herbosa) o acquatiche (aqui-herbosa); 3) deserta: formazioni semidesertiche di piante basse, molto metamorfosate (xeromorfiche), all'inizio molto lontane le une dalle altre, in modo da utilizzare efficacemente le condizioni ambientali assai ostili. Una classificazione più particolareggiata è la seguente.
Pluviisilvae: foreste fitte equatoriali ombrofite di pluviilignides sempervirentes o semicaducifoglie (rain forest), poco metamorfosate ma ricche in sociomorfosi diverse (liane, fanerogame, epifite, piante marantoidi; oloparassite di radici, funghi seminicoli, ecc.).
Hiemisilvae: foreste chiare e savane boscose tropicali, caducifoglie durante la stagione secca piuttosto breve; relativamente poco metamorfosate.
Laurisilvae: foreste sempreverdi di laurilignidi (tipo lauro) dei climi temperati o subtropicali umidi, dove l'influenza dell'oceano o delle nuvole è molto forte; abbondanti le Gimnosperme, specialmente a grandi foglie (Podocarpus e Araucaria) e le Pteridofite; abbondanti le Briofite sia sul terreno che sui tronchi e sui rami degli alberi (foreste di Formosa, di Ceylon, foresta temperata del Giappone).
Spinisilvae e Spinifruticeta: foreste tropicali caducifoglie durante la stagione secca molto lunga, molto ricche di ecomorfosi di adattamento al secco: spine, forte ritenzione di acqua, tronchi riserva (alberi bottiglia del tipo baobab), ecc.
Durisilvae e Durifruticeta: foreste, macchie e chaparral sclerofille, sempreverdi, molto ricche in ecomorfosi di adattamento al clima mediterraneo: foglie sclerosate, piccole, dure, a bordi spesso spinosi.
Aestisilvae: foreste temperate di piante caducifoglie, molto poco metamorfosate, che perdono le loro foglie durante la stagione fredda (inverno); sociomorfosi poco abbondanti.
Aciculisilvae: foreste soprattutto boreali (taiga), talvolta di montagna, caratterizzate da Conifere ad aghi; nel sottobosco sono presenti le Ericacee suffruticose e le Briofite.
Ericisuffruticeta: lande suffruticose di regioni artiche (tundra), boreali o temperate, che si estendono fino alle regioni mediterranee dove esse si trasformano progressivamente in macchie (Durifruticeta); più o meno intensamente metamorfosate; spesso ricche di Licheni.
Rosulifruticeta: formazioni particolari di altitudini elevate sotto il clima equatoriale, basse foreste di arbusti a tronco corto terminanti con una infiorescenza circondata da una rosetta di grandi foglie (Senecio e Lobelia, piante arborescenti di alta montagna dell'Africa equatoriale, Espeletia delle Ande, ecc.).
Molliprata (Sempervirentiherbosa): formazioni erbose sotto i climi temperati, sufficientemente umidi tutto l'anno; rigidità di foglie delle ‛Graminoidi' dovuta alla loro turgescenza; malgrado un periodo di riposo invernale, sembrano sempreverdi; poco metamorfosate. Molto propagate dall'uomo sotto forma di ‛praterie permanenti' utilizzate per gli allevamenti.
Duriprata: formazioni. erbose tipiche dei climi più o meno aridi, con foglie rigide a causa dell'abbondanza dei tessuti meccanici e di una cuticola spessa e riccamente silicizzata; ecomorfosi di adattamento al lungo periodo secco, sclerofillia, organi sotterranei di riserva di acqua, trofofillia, ecc. Si dividono generalmente in: steppe (steppiprata), formazioni basse per niente o poco arbustive; savane (savaniprata), formazioni alte più o meno densamente arbustive o arboree.
Le steppe propriamente dette si sviluppano in climi temperati o subtropicali a carattere continentale; esse formano terreni ricchi di humus del tipo černozëm (steppa russa, prateria americana, pampa).
Le savane si sono estese a opera del fuoco in immense regioni intertropicali; si possono dividere in: a) alte savane fresche ad arbusti a larghe foglie; b) basse savane secche ad arbusti spinosi con foglie piccole (Acacia); c) steppe tropicali, formazioni basse, per niente o poco arbustive.
Altherbosa: formazioni interzonali di erbe molto alte in zone a clima favorevole, proprie di ambienti in cui l'acqua è abbondante (paludi a Phragmites in tutte le latitudini, paludi a Papyrus nelle regioni intertropicali, ecc.).
Siccideserta: semideserti caratterizzati dalla mancanza di acqua liquida utilizzabile. Alcuni sono caldi, come il Sahara, il Sind e i deserti d'Australia; altri sono freddi d'inverno, come gli altopiani del Tibet e il Great basin degli Stati Uniti.
Frigorideserta: semideserti caratterizzati da gelate sfavorevoli alla vegetazione (tundra propriamente detta, vegetazione alpina).
I tipi edafici dei semideserti sono molto diversi: halodeserta, mobilideserta, petrideserta, rupideserta, saxideserta, ecc.
È difficile inserire in queste diverse formazioni vegetali gli animali e i Batteri importanti che vi corrispondono, per farne dei biomi.
Nella fig. 10 abbiamo rappresentato la zonazione polo- equatore delle principali formazioni precedentemente menzionate, con alcuni animali tipici.
In uno studio di insieme sulla produttività della biosfera, l'International Biological Program ha diviso i continenti in cinque biomi principali, rappresentati nella fig. 11 con la loro ampiezza in correlazione con la temperatura e le precipitazioni medie annue; conviene aggiungere naturalmente le acque dolci e le paludi a erbe alte. (Per quanto riguarda la distribuzione della vita nel mare e nei laghi, v. oceanografia biologica, idrobiologia).
La cartografia fitosociologica. - Le nozioni di ‛formazione' e ‛associazione' sono basilari per un tipo di cartografia della vegetazione indispensabile per lo studio dei terreni e per la comprensione e la risoluzione di grandi problemi di ambiente che coinvolgono tutta la biosfera.
Rodin e Basilevich (v., 1967 e 1968) hanno portato su carte i valori di produttività e i cicli biogeochimici diversi che caratterizzano ogni regione.
Il sistema di Gaussen (v., 1954), basato sulle serie di vegetazione, è stato applicato a una carta ecologica del mondo in corso di pubblicazione da parte dell'UNESCO.
6. L'ecosistema.
a) Descrizione.
La biocenosi dipende strettamente dai fattori abiotici dell'ambiente in cui vive. Questi fattori sono di ordine climatico ed edafico. Quando sono in quantità sufficiente, essi assicurano il funzionamento della rete trofica della biocenosi, permettendo sintesi, flussi energetici e conversioni di materia organica che portano alla produzione di una certa quantità e qualità di biomassa nei diversi livelli trofici. Perciò ogni biocenosi (v. flg. 5) forma con il substrato che l'alimenta (edafotopo) e con il clima nel quale vive (climatopo) un sistema funzionale, o ‛ecosistema', che è la più grande unità biologica funzionale: ‛unità' poiché funziona a circuito chiuso, la più ‛grande' poiché include sia gli organismi sia il loro ambiente abiotico, che s'influenzano reciprocamente e sono reciprocamente necessari per il mantenimento armonioso della vita.
L'ecosistema - termine creato da A.G. Tansley (v., 1935), ma definito da R.L. Lindeman (v., 1942) e da V.N. Sukatchev - si compone dunque: a) della fitocenosi autotrofa (livello di produzione) e di diverse zoocenosi, micocenosi e microbiocenosi eterotrofe raggruppate a livello di consumatori e trasformatori di ordine diverso (rete trofica); b) dell'ecotopo, insieme dei fattori dell'ambiente olistico raggruppati in climatopo ed edafotopo, da cui può essere separato un idrotopo, poiché il fattore acqua è a cavallo fra il clima e il suolo negli ecosistemi terrestri e costituisce l'elemento essenziale dell'ambiente abiotico negli ecosistemi acquatici; c) di tutte le relazioni e interazioni degli organismi fra di loro e con l'ecotopo (catene trofiche, legami corologici, aggregazioni, ecc.). Esso presenta un vero metabolismo, come una macchina estremamente complicata con gli ingranaggi più o meno regolati, in cui il flusso energetico, il bilancio idrico e il ritorno in ciclo dei bioelementi azionano le catene trofiche; l'ecosistema ha come risultato la produzione di materiali biologici primari molto diversi e di innumerevoli sostanze organiche secondarie (ecomoni) regolatrici del metabolismo. La velocità di produzione costituisce la produttività, che può essere studiata dal punto di vista quantitativo e qualitativo.
La biomassa così prodotta, che si esprime in peso di materia secca (MS) o meglio in chilocalorie (kcal) e che può trasformarsi poi in materia organica morta (necromassa, humus), comprende una mineralomassa (ceneri).
b) Funzionamento dell'ecosistema (metabolismo, produttività e biomassa).
Senza entrare in dettagli, prendiamo l'esempio di tre ecosistemi molto diversi, ma molto importanti nel mondo moderno: il campo di grano, la foresta, la prateria permanente. L'unità scelta è la kcal.
Il campo di grano. - Lo schema della fig. 12 (v. Ničiporovič, 1968) illustra il funzionamento (flusso di energia e ciclo del carbonio) di un ecosistema ‛campo di frumento' preso a una latitudine media in URSS. Per semplificare sono state rappresentate solo le radiazioni solari luminose fotosinteticamente attive (RFA), tralasciando il vicino infrarosso, che costituisce circa la metà della radiazione globale e che svolge un ruolo molto importante nella traspirazione. Durante un periodo di vegetazione attiva di circa quattro mesi, le RFA hanno un valore di circa 2,2•109 kcal/ha; 0,3•109 kcal/ha sono state riflesse o trasmesse al suolo. Le reazioni essenziali in seno all'ecosistema sono lo stabilirsi di una produttività lorda PL e di una produttività primaria netta PN1. Si è visto che PL è uguale a PN1 + R, dove R è la respirazione.
Le equazioni chimiche della fotosintesi (PL = Φ) e di R sono date nella stessa fig. 12.
Nelle foglie, 16 t di H2O sono trasformate, per fotolisi, in 14 t di O2, liberate nell'atmosfera, e 2 t di H, utilizzate per ridurre 20 t di CO2 in 14 t di materia organica (PL), con liberazione di 8 t di H2O. Prima di compiere questo grandioso lavoro fotochimico le piante assorbono all'incirca 1,3•109 kcal di luce solare, la magior parte della quale si trasforma in calore latente di evaporazione (principalmente sotto forma di traspirazione) di un'enorme quantità di acqua. Questa traspirazione permette l'assorbimento, necessario per l'elaborazione della materia organica, di circa 1 t di nutrienti minerali (N, P, K, S, Ca, ecc.) attinti dal suolo attraverso le radici. Le restanti 0,6•109 kcal/ha sono messe a disposizione dei tessuti fotosintetici delle piante, con elaborazione di 14 t di glucidi (produttività lorda).
Nel corso dello sviluppo della fitocenosi fino alla sua maturità e per fornire l'energia di crescita necessaria (adenosintrifosfato, ATP), i processi respiratori (R) utilizzano 4 di quelle 14 t di glucidi che, combinate con 4 t di O2, liberano, oltre all'energia di reazione, 6 t di CO2 e 2 t di H2O. Ne risulta una produttività primaria netta (PN1) di 10 t/ha.
Si vede che il rendimento è modesto; 2,2•109 kcal di RFA danno 40•106 kcal di materia secca di frumento, con un rendimento dell'1,8%; questa cifra è d'altra parte relativamente elevata, poiché il rendimento per le messi è generalmente tra lo 0,5 e l'1,5%.
Si vede così che, per ettaro, si liberano 14 t di O2 (che provengono dall'acqua), di cui 4 t sono riassorbite per attività respiratoria; le altre 10 sono trattenute dall'atmosfera per turbolenza e saranno poi utilizzate dalla respirazione di altri organismi (piante e animali, uomo compreso).
La foresta caducifoglia temperata (Aestisilvae). - La fig. 13 rappresenta l'esempio di un Querceto-Carpineto in Belgio (v. Duvigneaud, 1967) di biomassa (B) pari a 350 t/ha, di superficie fogliare (SF) pari a 6 ha/ha e in piena crescita.
Questo ecosistema si complica per il fatto che la fitocenosi è in aumento. Ogni anno, un incremento T (PN1 apparente, cioè PN1 diminuita delle perdite per caduta di lettiera e consumo da parte di erbivori) si aggiunge a una biomassa di organismi perenni.
Questo incremento, aggiunto sotto forma di legno, è particolarmente utile. È generalmente espresso in m3 dai forestali e in peso secco o kcal dai biologi e dagli ecologi.
La produttività primaria netta (PN1) è in questo caso di 16 t/ha/anno (64•106 kcal), un po' maggiore di quella di un campo di grano. Il rendimento fotosintetico delle RFA è del 2,3%.
Di 5 t di lettiera che cadono ogni anno, 4 sono respirate dai decompositori e 1 t forma humus (ΔH), che si aggiunge alle 150 t di humus (H) preesistente.
La prateria permanente (Molliprata). - Questo ecosistema (sempervirentiherbosa, molliprata) è largamente propagato e migliorato da parte dell'uomo per l'allevamento del bestiame.
Ridotto ai suoi elementi essenziali (sole-erba-vacche-decompositori, v. fig. 14), illustra in maniera molto semplice i problemi della produttività primaria e secondaria. L'esempio della figura è quello di un prato verde seminaturale in Gran Bretagna, in cui la densità di bovini è piccola (1,2/ha) e in cui questi pascolano liberamente un'erba non concimata. I valori sono stati rapportati a una superficie di 1 m2.
Si vede che le 400.000 kcal di energia luminosa assorbite annualmente da i m2 di prateria servono a elaborare 2.500 kcal di erba; di questa 1,2 vacche non utilizzano che 730 kcal per elaborare appena 40 kcal di loro materia (produttività secondaria netta): 244 kcal sono in effetti utilizzate per la respirazione (soprattutto per gli spostamenti e i movimenti della testa) e 446 kcal sono espulse sotto forma di feci o di urina.
Questo può tradursi approssimativamente come segue: su 1 ha di prateria e in 180 giorni all'anno (periodo di pascolo), circa 5,5 t di erba (MS) sono fornite a 1,2 vacche di 350 kg che producono 180 kg di bovini vivi, corrispondenti a circa 100 kg di carne fresca.
Si vede l'immenso spreco che fa la vacca della materia vegetale: solo il 29% di questa è consumata e, inoltre, di questo 29% solo il 5% serve alla produttività secondaria netta, il resto è perduto sotto forma di feci e di urina, oppure per una respirazione intensa dovuta ai movimenti e agli spostamenti dell'animale. 840 kcal sono ingerite da altri erbivori, molluschi, larve di ditteri, i quali per una biomassa molto minore devono mangiare molto più dei Mammiferi, dato il rapporto molto più grande della loro superficie rispetto al peso; 930 kcal di parti morte sono messe, con le 446 kcal di escrementi, a disposizione dei microrganismi decompositori.
Una produttività di 4 t di radici per ettaro fornisce ai decompositori altre 1.700 kcal/m2.
Questo esempio illustra il fatto che gli ecosistemi sono poco redditizi e che l'allungamento di una catena trofica e il passaggio da un livello trofico al seguente avvengono con una perdita di produttivita considerevole.
In questo caso il rendimento fotosintetico annuale della fitocenosi è dell'1%; quello della vacca nei confronti della fitocenosi è dell'1,6% e nei confronti dell'energia solare è dello 0,01%. Abbiamo tuttavia considerato il caso sfavorevole di un ecosistema vicino a un ecosistema naturale. Ma in caso di condizioni ottimali questi rendimenti si possono quadruplicare.
c) Estensione dell'ecosistema; la biosfera.
Sul terreno è spesso molto difficile assegnare a un ecosistema un limite definito in una foresta o in una prateria ed è difficile definirne l'estensione (non parliamo poi delle difficoltà che presenta a questo riguardo l'ambiente acquatico, in particolare l'oceano).
In numerosi casi, niente impedisce di dare al concetto di ecosistema il suo senso etimologico di sistema ecologico: un paesaggio definito oppure una regione determinata possono costituire dei sistemi ecologici funzionali particolarmente efficienti. Negli elementi di un paesaggio gli ecosistemi elementari sono uniti da rapporti collaterali molto stretti: un ecosistema acquatico, fiume o lago, può dipendere profondamente, per i sali minerali e la materia organica di cui si alimenta, dagli ecosistemi terrestri che lo costeggiano; questi sono reciprocamente influenzati, dal punto di vista dei fattori temperatura e acqua, dalla vicinanza di uno strato liquido.
La biosfera, composta dall'integrazione di tutti gli ecosistemi mondiali, non è altro che un gigantesco ecosistema (globo terrestre) in cui tutte le parti sono interagenti.
Le relazioni collaterali fra ecosistemi che danno luogo a una catena non costituiscono solo un insieme di relazioni fisiche e chimiche dovute alla topografia, ma si realizzano anche attraverso organismi diversi: trasporto di spore o di animali, che può avvenire su territori molto vasti.
Bilancio energetico. - L'energia solare che alimenta l'ecosistema, sotto forma di luce e di calore, è in parte riflessa, in parte assorbita dagli organismi viventi (essenzialmente dalle foglie), in parte trasmessa al suolo. Il bilancio energetico si suddivide in due parti (v. fig. 15).
1. La radiazione luminosa o energia a breve lunghezza d'onda (λ compresa fra 0,28 e 3 μ) è
Qb = S + D − a (S + D),
dove S è la ‛radiazione solare diretta', D è la radiazione del cielo o ‛radiazione solare diffusa' dall'atmosfera e dalle nuvole.
Queste radiazioni comprendono:
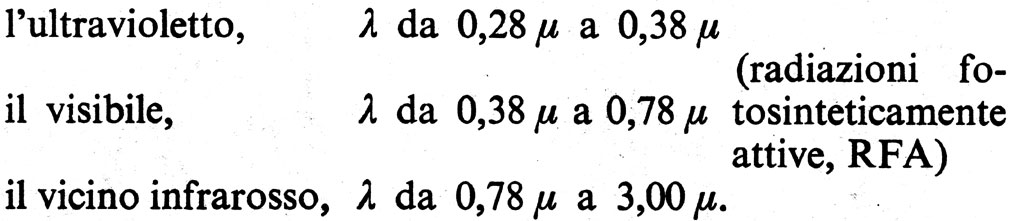
La somma S + D, su una superficie orizzontale, viene chiamata ‛radiazione globale' (G). Di questa radiazione globale, una parte viene riflessa (coefficiente di ‛albedo' a) allorché essa colpisce delle superfici naturali. Una prateria verde può riflettere il 10-15% della luce visibile; una foresta ‛nera' composta da Conifere, il 5-10%. Il vicino infrarosso è invece riflesso in quantità maggiori (circa il 40%).
Generalmente si ammette che per una vegetazione densa (coltura) in regioni temperate venga riflesso il 17% della radiazione solare, mentre il 70% viene assorbito dalle foglie e il rimanente 13% viene trasmesso.
2. La radiazione termica o energia a lunga lunghezza d'onda (lontano infrarosso per il quale λ va da 3μ a 100μ) è
Ql = A − aNA − T.
L'atmosfera terrestre invia verso la Terra una sua propria radiazione A (Gegenstrahlung), la cui intensità dipende dalla temperatura e dal tenore dell'aria in vapor acqueo e in CO2; questa radiazione diffusa infrarossa, che sta fra 3 e 100μ, è a sua volta riflessa dalla superficie terrestre con un coefficiente aN che è del 4% per le foglie o gli aghi degli alberi e per l'acqua, ma giunge all'11% per la sabbia chiara.
La radiazione (T) di ogni porzione della superficie terrestre è funzione della sua temperatura e varia dunque fortemente nel corso della giornata.
Quasi sempre, tanto di giorno che di notte, la radiazione terrestre è maggiore della radiazione dell'atmosfera, di modo che il bilancio è negativo e il suolo irradia calore verso l'atmosfera. Il bilancio dell'irraggiamento totale si esprime dunque attraverso la formula:
Q = Qb + Ql =
S + D − a(S + D) + A − aNA − T
= G − aG + A − aNA − T.
Nel corso di una intera giornata, o nel corso di un intero anno, Q è generalmente positivo, poiché l'intensità della radiazione solare incidente di breve lunghezza d'onda è maggiore della radiazione termica di lunga lunghezza d'onda riemessa dalla Terra. Ne dovrebbe derivare un raffreddamento progressivo dell'atmosfera e un riscaldamento progressivo della superficie terrestre, ove la vita diventerebbe ben presto impossibile. E tuttavia viene realizzato un equilibrio termico che mostra la relativa costanza dei climi durante lunghi periodi. Questo equilibrio è dovuto da una parte a un trasferimento di energia dalla superficie terrestre verso l'atmosfera per conduzione e convezione (calore sensibile: K), dall'altra a un'utilizzazione dell'eccesso di calore, da parte delle superfici vegetali e terrestri, per vaporizzazione (evapotraspirazione) dell'acqua (V).
Al di fuori di questi due fattori che equilibrano l'eccesso di energia che si trova sulla superficie terrestre, ve ne sono degli altri, meno importanti quantitativamente ma molto più importanti qualitativamente, come l'assorbimento di calore, particolarmente di giorno, da parte del suolo e del substrato (QG) e soprattutto l'utilizzazione dell'energia luminosa nella sintesi clorofilliana compiuta dalle piante verdi (aPG = PN1).
In fin dei conti si ha:
Q + V + K + QG + aPG = 0;
questi parametri possono essere sia positivi sia negativi. Ne esistono altri (v. fig. 15), ma sono di minore importanza.
Bilancio idrico (v. Penman, 1963). - Gli ecosistemi terrestri sono per lo più alimentati, per quanto riguarda l'acqua, dalle precipitazioni atmosferiche. L'autoalimentazione, con acqua proveniente dallo stesso ecosistema, è di scarso rilievo: l'acqua evapotraspirata è trascinata nell'atmosfera dalla turbolenza e dai venti orizzontali. D'altro canto, una parte dell'acqua che cade sull'ecosistema si perde nel suolo e alimenta le falde liquide e i corsi d'acqua. Non si può, dunque, parlare di un ciclo dell'acqua in seno all'ecosistema, ma di un bilancio idrico.
Il bilancio dell'acqua di un ecosistema (v. fig. 16) comporta l'acqua che penetra nell'ecosistema (apporto), quella che viene ritenuta (contenuto) e quella che ne esce (perdita).
Gli apporti di acqua si realizzano in tre forme ben note: di vapore, liquido e solido. L'acqua proviene in gran parte dalle precipitazioni atmosferiche (Pr) (caduta di pioggia, brina, neve, grandine e nevischio). Acqua liquida può inoltre entrare nell'ecosistema per ascensione capillare (AC) da una falda profonda.
Talvolta la nebbia Ne e le nuvole Nu possono essere una sorgente molto cospicua di acqua per l'ecosistema. Il vapor d'acqua dell'aria può condensarsi sotto forma di rugiada (R). Nell'ecosistema, inoltre, può penetrare acqua per trasferimenti laterali (nebbia, Ne; scorrimento superficiale, SS; drenaggio orizzontale, DO).
Così l'equazione generale degli apporti è:
Apporti = Pr + Ne + Nu + R + SS + DO.
Una parte delle acque di precipitazione, di nebbia e di rugiada è intercettata dalla fitocenosi e restituita all'atmosfera (intercettazione, It); l'acqua che attraversa il foglia- me raggiunge l'edafotopo per vie diverse. Una parte giunge direttamente al suolo senza incontrare ostacoli: è la penetrazione diretta (PeD); un'altra parte passa sugli organi aerei della fitocenosi e sgocciola fino al livello del suolo: è l'acqua di sgocciolamento (Sg); un'altra parte ancora scivola lungo rami, tronchi, steli, foglie: è l'acqua di scolo (Sc) o stem flow.
La penetrazione totale (PeT) che forma le precipitazioni nette (PrN) utilizzabili dalla biocenosi può dunque essere espressa sotto la forma:
PeT = PrN = PeD + Sg + Sc.
Dal punto di vista tecnico è difficile misurare separatamente PeD e Sg; PeD + Sg formano il throughfall degli autori inglesi, penetrazione attraverso il fogliame o PeF.
Si ha cioè:
PeF = PeD + Sg.
Il contenuto in acqua corrisponde alla quantità totale di acqua assunta dall'edafotopo (HT) e dalla biocenosi (costituita da fitocenosi, zoocenosi e micocenosi: HB = HP + HZ + HM), più il vapore acqueo dell'aria interna all'ecosistema (HA).
Si ha dunque:
H = HT + HB + HA.
L'acqua di idratazione della fitomassa (HP), che si può chiamare idromassa e che si confonde quasi con HB, è poco rilevante in rapporto alle precipitazioni: circa 100 t/ha in una foresta e 2 t/ha in una prateria, laddove la precipitazione è di circa 10.000 t/ha/anno (1.000 mm). Le fluttuazioni giornaliere e stagionali di HP sono notevoli e raggiungono il 30% in una foresta temperata.
Le perdite di acqua avvengono sotto forma di liquido o di vapore. L'acqua delle precipitazioni nette (PrN) si infiltra nel suolo (If) o vi scivola sopra (SS). L'acqua perduta per scivolamento è d'altro canto tanto più abbondante quanto maggiore è la perdita e quanto più la vegetazione è ridotta; il suolo nudo è oggetto di una forte erosione. L'infiltrazione si esprime con la formula: If = PeN − SS.
L'acqua di infiltrazione contribuisce prima di tutto a ricostituire interamente la riserva idrica dell'edafotopo. Allorché il suolo raggiunge la sua capacità massima di ritenzione, l'acqua di infiltrazione scola in profondità (drenaggio verticale = DV) alimentando le falde o ‛aquafer' profonde o muovendosi orizzontalmente fino a una zona di assorbimento (drenaggio orizzontale = DO). Questo scolamento è tanto più accentuato quanto più il clima è piovoso e quanto più il suolo è permeabile (più leggero o più sabbioso); esso invece è tanto più ridotto quanto più il suolo è ricco di radici e quanto più presenta un'elevata traspirazione. Questo scolamento porta in soluzione gli elementi biogeni e i colloidi del suolo.
L'acqua dello scorrimento e del drenaggio costituisce l'acqua corrente (streamflow) dei ruscelli e dei fiumi, cioè quella che alimenta gli ecosistemi acquatici e di lì gli ecosistemi urbani e industriali.
La quantità di acqua che lascia l'ecosistema sotto forma di un flusso di vapore dipende dalla quantità di energia disponibile, dalle superfici di scambio e dalla resistenza offerta dai diversi organi. Essa comprende l'intercettazione (It), la traspirazione (Tr) e l'evaporazione dal suolo (ES). Il flusso globale di vapore che lascia l'ecosistema è l'evapotraspirazione reale (ETR).
ETR = It + Tr + ES.
La quantità di acqua traspirata dalle piante è grande, tanto più quanto migliore è la loro alimentazione in acqua. Una betulla lascia evaporare ogni giorno 15 l di acqua, un faggio 100 l, un tiglio 200 l, un ettaro di foresta da 20.000 a 50.000 l. Un ettaro di betulle, la cui massa fogliare è appena di 4.940 kg, fa evaporare più acqua (47.000 l al giorno) di un ettaro di Conifere, la cui massa fogliare raggiunge i 31.000 kg (traspirazione di 43.000 l al giorno). La traspirazione di un tappeto vegetale va dalle 2.000 alle 6.000 t di acqua per ettaro e per anno, secondo il tipo di coltura e il clima. L'acqua traspirata è un fattore di produttività molto importante. Il coefficiente di traspirazione è la massa di acqua traspirata per unità di massa di materia secca elaborata durante una stagione. Questo coefficiente è enorme e varia da 300 a 1.000 a seconda delle specie. Le differenze dipendono soprattutto dal clima: le piante delle zone aride consumano una quantità d'acqua doppia di quelle delle regioni umide per produrre la stessa quantità di materia secca.
Nelle regioni aride alcune piante a radici lunghe possono andare a cercare l'acqua anche a grande profondità: queste ‛freatofite' sono al contempo degli indicatori di strati liquidi e organismi nocivi, perché tendono ad asciugare la zona.
L'evaporazione è naturalmente funzione del clima ed è stata molto studiata dai meteorologi; essa è molto meno accentuata nei tappeti vegetali che non nell'aria libera, a causa della funzione schermo delle piante.
L'evapotraspirazione, che è la somma dell'acqua traspirata delle piante e di quella evaporata dal suolo, può essere stimata, nelle regioni temperate, in una quantità compresa fra le 3.000 e le 7.000 t/ha/anno. Le perdite di acqua possono esprimersi così:
Perdite = SS + DV + It + Tr + ES.
L'importanza del bilancio idrico dipende da quanto detto fino ad ora. Si può parlare di funzione schermo degli ecosistemi nei confronti delle precipitazioni, che interessa due livelli: la fitocenosi (schermo verde) e il suolo (schermo bruno). L'acqua che questo schermo ‛lascia passare' per scorrimento e infiltrazione è quella che alimenta i corsi d'acqua e le falde acquifere superficiali e profonde. L'acqua immessa di nuovo nell'atmosfera è responsabile della produttività primaria, ma attraverso ciò è di fatto responsabile anche della produttività secondaria.
L'evaporazione è anche una inutile perdita di acqua; essa è tanto maggiore quanto meno il suolo è coperto di vegetazione; si cerca di ridurla coprendo il suolo con sostanze poco permeabili (mulching).
La fig. 17 illustra il ciclo dell'acqua nella Repubblica Federale Tedesca.
L'edafotopo. - Il terreno è il substrato delle comunità terrestri come l'acqua lo è per le comunità acquatiche.
Il terreno ha un'importanza così grande per l'agricoltura che il suo studio è divenuto una scienza autonoma, la pedologia; Dokučaev, fondatore della pedologia, l'ha orientata fin dai suoi inizi in un senso tale che non è possibile dissociarla completamente dallo studio degli ecosistemi.
Il terreno forma l'edafotopo dell'ecosistema. È lo strato superficiale della litosfera, più o meno tenero e friabile, nel quale le piante sono radicate. Risulta dalla decomposizione meccanica e chimica della roccia madre geologica, sotto l'azione dei fattori esterni, clima e vegetazione.
È ben conosciuto il seguente triangolo di interrelazioni:
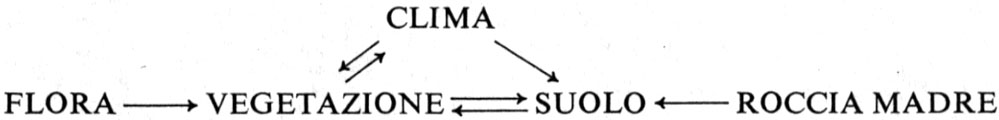
Per lo sviluppo considerevole del loro sistema radicale, le piante superiori sfruttano al massimo le condizioni offerte dal terreno. Alle radici si mescolano numerosi organismi viventi: batteri, funghi e animali, che costituiscono la pedoflora e la pedofauna. Numerosi microrganismi si agglomerano al livello delle radici, formando la rizosfera. Cadaveri ed escrementi, più o meno degradati dai microrganismi decompositori, aggiungono al terreno una materia organica estremamente complessa: l'humus.
Il terreno è dunque una entità estremamente complessa e profondamente viva e diversificata, un ecosistema sui generis. L'edafotopo ne costituisce la parte abiotica, composta da una frazione minerale e una frazione organica.
La frazione minerale del terreno si forma a partire dalla roccia madre tramite tre processi fondamentali: a) disintegrazione meccanica; b) decomposizione (alterazione) chimica; c) dissoluzione. L'importanza di questi processi dipende dalla qualità della roccia madre.
La disintegrazione meccanica è prodotta: 1) da fluttuazioni di temperatura: l'alternanza di freddo e caldo produce differenze di dilatazione o di contrazione, in superficie e in profondità, dei diversi minerali che costituiscono la roccia, determinando così una sua frammentazione in pezzi sempre più piccoli; 2) dall'azione cuneo dell'acqua, che gela negli interstizi e nelle fessure, e da quella delle radici che vi si insinuano e si accrescono; 3) dall'azione dei ghaccio in movimento (nel fondo dei ghiacciai); 4) dall'azione abrasiva di particelle trasportate dal vento e dalle onde.
Così la roccia madre (RM) è progressivamente disintegrata in particelle sempre più piccole (regoliti) che formano lo scheletro del terreno. Facendo passare queste particelle su setacci di maglie diverse, si può separarle in categorie di grandezza determinata:

I terreni composti da prodotti grossolani di una disgregazione puramente fisica (deserti, regioni alpine e artiche) sono dei terreni scheletrici: litoterreni se costituiti di ghiaia, regoterreni se costituiti di sabbia fine.
La decomposizione chimica segue la disgregazione meccanica ed è tanto più attiva quanto più sono piccole le particelle. Essa avviene in fase acquosa per ossidazione o per azione idrolitica dell'acqua, rinforzata dalla presenza di CO2 disciolto e di acidi minerali od organici formati dalle radici.
Avvengono due fenomeni: sparizione di certi minerali primari, neoformazione di minerali secondari.
I risultati generali della decomposizione delle rocce sono: 1) una desilicificazione: perdita di silicio; 2) una dealcalinizzazione: perdita di cationi; 3) una formazione di sostanze nuove, i minerali argillosi (silice-allumina).
Le particelle di argilla (micelle) sono delle scagliette esagonali dotate di proprietà colloidali, capaci in particolare di assorbire i cationi biogeni, proteggendoli così dal dilavamento. L'argilla dunque è un elemento di fertilità del suolo. Vi sono due grandi gruppi di minerali argillosi: le montmorilloniti e le caoliniti; queste ultime, avendo una più debole carica elettrica, hanno una minore capacità di trattenere i cationi. Esse si formano soprattutto nei climi tropicali e sono responsabili della scarsa fertilità dei relativi terreni.
Alla frazione minerale del terreno bisogna aggiungere una frazione organica: l'humus. Questa sostanza proviene dalla decomposizione della lettiera, massa di materia organica morta proveniente dalle parti morte delle piante e dai cadaveri degli animali, che si accumula ogni anno sulla superficie del suolo. Si distinguono diversi tipi di humus: torba (formatasi in anaerobiosi), mor (in ambiente povero di ossigeno), moder (in ambiente con contenuto medio), mull (in ambiente ricco d'ossigeno). Le sue particelle colloidali adsorbono i cationi biogeni ancor più energicamente delle argille.
Gli elementi minerali che si trovano in abbondanza nel terreno allo stadio di cationi o di anioni sono, escluso il titanio, necessari alla vita delle piante e degli animali e sono perciò chiamati elementi biogeni. La frazione maggiore di essi si trova imprigionata nello scheletro del terreno, da cui le piante l'assorbono lentamente. Inoltre nel suolo stesso sono presenti tutti gli altri elementi del sistema di Mendeleev. È poi noto che il potassio e il fosforo sono fissati nel suolo, fra fogli di micelle di argilla o sotto forma di composti insolubili. L'azoto, infine, non è contenuto nelle rocce madri e deve essere preso dall'atmosfera.
Il pH del suolo ha una grandissima importanza; le piante vengono classificate in oxilofite o acidofite (localizzate in suoli acidi), neutrofite (intermedie) e basofite (in suoli alcalini). Negli estuari, dune, falesie, deserti, il suolo è spesso salato, con una pressione osmotica molto elevata. L'ecologia delle alofite ha avuto negli ultimi decenni un immenso sviluppo, che è alla base della valorizzazione delle terre aride, la cui superficie ricopre circa ¼ dei continenti.
Si è visto che il terreno è un miscuglio molto complesso di frammenti più o meno sottili di roccia e di micelle colloidali di argilla e di humus. Il contenuto del terreno in questi diversi elementi ne determina la trama: un terreno ricco di sabbia è un terreno leggero, ben aerato; un terreno argilloso è pesante, difficile da lavorare, male aerato.
Le proprietà dei colloidi fanno in modo che le particelle del terreno si leghino in aggregati più o meno grossi, cementati dall'argilla e dall'humus, mentre la loro coesione è assicurata dalle radichette delle piante. Il grado di aggregazione del suolo in grumi più o meno grossi ne indica la struttura. Una buona aggregazione è dovuta alla flocculazione dei suoi colloidi. Una peptizzazione dei colloidi porta a un suolo omogeneamente compatto, non aerato e pertanto non adatto alla vita delle piante e degli animali. Trama e struttura concorrono alla porosità del suolo e quindi al suo stato di aerazione e al suo regime idrico. Un suolo poco poroso è detto asfissiante.
L'infiltrazione di acqua nel suolo è generalmente verticale. Questo movimento (percolazione) può trascinare materia disciolta o particelle solide, dando vita, talvolta, alla formazione di strati sovrapposti, che con gli strati umiferi in superficie formano il profilo del suolo (v. fig. 18). Gli strati principali, dall'alto verso il basso, si succedono nella maniera seguente.
1. Strati olorganici, formati da materia organica più o meno decomposta che si trasforma lentamente in humus; vi si distinguono: a) uno strato A00 (L): lettiera di organi morti non decomposti; b) uno strato A0: humus, formato talvolta da due sottostrati ben visibili che sono lo strato di fermentazione (F) e lo stratQ umificato (mar) ma senza incorporazione di materia minerale (H).
2. Strato misto inorganico A1, più o meno scuro, dove l'humus è incorporato nella materia minerale (moder, mul) mediante processi biologici.
3. Strati inorganici, più chiari o vivamente colorati, in cui la materia organica è quasi completamente assente. Vi si possono distinguere uno strato A2 di dilavamento o eluviazione e uno strato B di arricchimento o di illuviazione. Quest'ultimo può presentare diversi sottostrati differentetemente colorati, per es. uno colorato in nero per humus (Bh) e uno colorato in rosso-bruno per ossidi di ferro (Bf).
4. Strato da cui ha origine il terreno, costituito da un sottostrato di origine diretta, sassoso (C1), e dalla roccia madre (C2).
Comunque fra tutti questi strati esistono spesso delle transizioni; d'altra parte alcuni possono mancare oppure essere invertiti.
La maggior parte dei terreni mondiali rientrano nelle grandi categorie proposte da Kubiena (v., 1953).
I cicli biogeochimici. - I grandi cicli biogeochimici della biosfera sono in effetti la somma di cicli particolari che si svolgono a livelli di ciascun ecosistema preso isolatamente. I cicli non funzionano sempre in maniera regolare e presentano dei punti di stagnazione laddove le materie organiche si accumulano. Dalla velocità e dall'intensità dei cicli dipende la produttività degli ecosistemi e di conseguenza quella della biosfera.
1. Ciclo del carbonio. Questo ciclo (v. fig. 19) si confonde in parte con il flusso energetico. Le sorgenti di carbonio in natura sono tanto numerose quanto varie. Nell'atmosfera troviamo il carbonio sotto forma di composto gassoso: CO2 (0,03%); lo stesso diossido di carbonio (anidride carbonica) è presente allo stato disciolto nell'acqua. Captato dalle piante il CO2 è trasformato, tramite la fotosintesi, in glucidi e altri composti (v. fotosintesi).
L'idrogeno necessario è fornito dalla fotolisi dell'acqua, il cui ossigeno ritorna nell'atmosfera. Le diverse sostanze così elaborate servono come alimento e come materiale da costruzione alle piante verdi e agli animali consumatori. D'altra parte tutti gli organismi respirano, cioè assumono ossigeno ed emettono carbonio nell'atmosfera sotto forma di CO2. Allorché sopraggiunge la morte, i cadaveri vengono decomposti e mineralizzati da ‛biotrasformatori' di tutti i tipi, che formano catene lungo le quali il carbonio è rimesso in circolo sotto forma di CO3 (respirazione del suolo).
In certe condizioni (bassa temperatura, forte acidità) i cadaveri e i residui vegetali accumulati producono un rallentamento di questo ciclo: vengono trasformati in nuove materie organiche che diventeranno l'humus. Talvolta la catena dei decompositori non funziona per mancanza di aria o per eccessiva acidità, cosicché le materie organiche si accumulano sotto forma di torba e si ha una stagnazione del ciclo.
2. Ciclo dell'ossigeno. Questo ciclo è strettamente legato a quello del carbonio. Tutto l'ossigeno atmosferico, e di conseguenza quello che, portato per turbolenza dall'aria, impregna attivamente tutti i compartimenti dell'ecosistema, è prodotto attualmente dalla fotosintesi dei produttori primari (piante verdi). L'equilibrio (21% di ossigeno nell'aria) è dovuto al fatto che una quantità di carbonio organico stechiometricamente uguale a quella dell'ossigeno prodotto è riutilizzata dalla respirazione dei consumatori, dei trasformatori e anche dei produttori. Questa riossidazione può essere prodotta anche dal fuoco.
3. Ciclo dell'azoto. Malgrado la sua grande complessità, questo ciclo (v. fig. 20) funziona bene e rapidamente. L'aria, che contiene circa l'80% di azoto, è la grande riserva e la valvola di sicurezza del sistema; essa alimenta costantemente il ciclo in molte maniere. Innanzitutto le scariche elettriche che si producono durante i temporali formano, dall'N2 e dall'O2 dell'aria, ossidi di azoto, che sono dalla pioggia trascinati nel terreno, dandovi luogo a nitriti e nitrati, i quali vengono poi utilizzati dalle piante. Ma è per azione dei microrganismi azotofissatori che la maggior parte dell'azoto dell'aria è fissato sotto forma di protidi. Molto spesso si tratta di batteri che agiscono in aerobiosi (Azotobacter) o in anaerobiosi (Clostridium), i cui cadaveri arricchiscono il terreno di N organico, che viene rapidamente mineralizzato.
I batteri più efficaci sono quelli che vivono in simbiosi con le Leguminose, in noduli che si sviluppano sulle radici di queste piante. In presenza di molibdeno il Rhizobium assimila enormi quantità di N2; l'azoto organico così formato arricchisce considerevolmente gli organi aerei delle piante ospiti e viene liberato nella rizosfera. Anche altri batteri vivono in simbiosi con piante superiori e fissano l'azoto, ad esempio gli Actinomiceti delle radici dell'Alnus. Infine nell'acqua e sulla terra umida vi sono le Alghe blu, Cianoficee, che fissano direttamente l'azoto dell'aria.
Tali diverse fonti di azoto portano quest'ultimo a contatto delle radici, generalmente sotto forma di nitrati, che sono utilizzati dalle piante per costruire le loro proteine. Queste sono alla base dell'alimentazione azotata degli animali. Proteine vegetali e animali sono il nutrimento anche degli organismi parassiti. Quando sopravviene la morte, i decompositori fanno rapidamente ritornare l'azoto allo stato minerale; gruppi diversi di bioriduttori sono specializzati ciascuno in una parte di lavoro; la catena termina con l'azione degli organismi ammonizzanti, che producono ammoniaca. L'NH3 può allora entrare nel ciclo della nitrificazione: Nitrosomonas la ossida a nitrito, e Nitrobacter ossida rapidamente i nitriti a nitrati. Ammoniaca e nitrati sono quindi messi a disposizione delle piante verdi e il ciclo ricomincia.
D'altra parte l'azoto è costantemente reso all'aria da parte dei batteri denitrificanti, che decompongono i nitrati in N2 e lo restituiscono all'atmosfera. La maggior parte delle piante verdi non può assumere l'azoto, con le radici, se non sotto forma di anioni nitrici NO−3 o di cationi ammonio NH+4, la cui abbondanza relativa dipende dall'attività dei batteri nitrificanti. D'altronde le piante verdi non possono che assorbire l'azoto minerale (mineralizzazione netta) che i microrganismi del suolo lasciano loro.
La velocità di mineralizzazione netta fissa la quantità di N assorbita dalle radici e, attraverso questa, la produttività degli ecosistemi. Nella maggior parte degli ecosistemi il ciclo dell'azoto è molto economico e soltanto quantità molto piccole di azoto vanno perdute, ad es. per dilavamento.
4. Ciclo dello zolfo. Lo zolfo che esiste nei terreni normali proviene o direttamente dalla roccia madre che conteneva pirite (FeS2) o calcopirite (CuFeS2), oppure dalla decomposizione della materia organica. Esistono però territori, talvolta molto estesi, in cui la roccia madre è costituita da gesso (CaSO4•2H2O); questi sono composti da ecosistemi semidesertici se il clima è molto arido. Lo zolfo si trova nelle piante incorporato principalmente nelle proteine ed è da esse restituito al suolo quando muoiono.
Numerosi microrganismi (Proteus, Serratia, Penicillum, ecc.) riducono lo zolfo organico a H2S; questo puà essere ossidato a solfato (da Thiobacillus, Beggiatoa, Chromatium, ecc.); i solfati sono assunti dalle radici delle piante e ciò assicura la continuità del ciclo. I solfati possono anche essere ridotti a solfuri dal Desulfovibrio.
5. Ciclo del fosforo. È un ciclo semplice ma aperto (v. fig. 21). La sorgente iniziale di fosforo è l'erosione delle rocce madri, ma alcune di queste ne contengono molto poco, cosa che favorisce l'origine di un deserto (Australia).
Un fenomeno molto ben conosciuto è la fissazione del fosforo nel suolo, sotto una forma non assimilabile dalle piante. Questa fissazione avviene sotto forma di fosfati insolubili; in un mezzo molto acido si formano fosfati di ferro, alluminio e manganese, insolubili. In un mezzo alcalino un eccesso di calcio precipita un fosfato tricalcico complesso (apatite).
L'organomassa del suolo esplica un ruolo di biotampone, poiché il fosforo vi si trova principalmente sotto forma organica (fitina, fosfolipidi); se essa è abbondante e il suo ricambio è rapido, grazie a popolazioni microbiche attive, il suolo è costantemente rialimentato in fosforo assimilabile che non ha il tempo di fissarsi.
Negli ambienti acquatici il fosforo diviene un oligoelemento ed è per questo che la produttività degli ecosistemi che li caratterizzano è spesso ridotta. Ciò nonostante, l'uomo ha arricchito recentemente gli ecosistemi acquatici di fosforo, riversandovi dei detergenti molto ricchi di fosfati. Questo però a lungo andare può portare a un rovesciamento della situazione.
6. Ciclo dei cationi biogeni. Gli esseri viventi non possono vivere se non contengono quantità sufficienti di alcuni cationi. K, Ca, Mg e talvolta Na appartengono al gruppo degli ‛elementi maggiori' poiché sono necessari in grandi quantità; mentre B, Zn, Cu, Co, Mo sono oligonutrienti, perché servono solo in quantità minori; Fe e Mn occupano invece una posizione intermedia (mesonutrienti). È importante notare che alcuni di questi elementi sono necessari solo agli animali, altri invece solo alle piante.
Sulla terraferma la sorgente di cationi biogeni è principalmente il terreno, che li riceve dalle rocce madri, decomposte; è importante anche l'apporto della pioggia. Questi elementi vengono assorbiti dalle radici, poi distribuiti ai diversi organi delle piante, ma tendono ad accumularsi nelle foglie. Servono così come alimentazione agli erbivori ed entrano nella catena trofica dei consumatori dei diversi ordini.
La mineralizzazione dei rifiuti e dei cadaveri porta gli elementi nel suolo, al livello delle radici, e il ciclo sembra così poter continuare in eterno. Nelle foreste può aver luogo un rallentamento temporaneo per accumulazione nel legno e nelle scorze degli alberi. Un blocco del ciclo, invece, avviene allorché la lettiera si decompone lentamente e l'humus si accumula sulla superficie del terreno.
Nei climi umidi il ciclo può essere profondamente disturbato dal dilavamento del terreno da parte delle piogge; queste trascinano verso la falda acquifera e i corsi d'acqua quantità di elementi talvolta molto considerevoli. Questa situazione può diventare particolarmente grave nei climi tropicali, nei quali l'equilibrio è più difficile da mantenere, a causa del fortissimo dilavamento provocato dalle acque a carattere torrenziale, e a causa dello scarso potere assorbente del terreno (poco humus, caolino).
Il cammino dei bioelementi segue due vie principali (v. Ovington, 1962): a) ciclo biologico, o ciclo chiuso; b) ciclo geochimico, o ciclo aperto verso il mondo esterno.
Il ciclo biologico (v. fig. 22) è la circolazione degli elementi biogeni in seno all'ecosistema. Il ciclo biologico annuale comprende: a) assorbimento, da parte delle radici, degli elementi chimici del suolo; b) restituzione al suolo di una parte di questi elementi per caduta di lettiera, per acqua di lavaggio della fitocenosi, per acqua di sgocciolamento, per secrezioni radicolari, per radici morte; c) ritenzione, negli organi perenni della fitocenosi, dell'altra frazione degli elementi assorbiti.
L'assorbimento, che grossolanamente corrisponde ai bisogni della biocenosi, è naturalmente la somma degli elementi ritenuti e degli elementi restituiti (assorbimento = ritenzione + restituzione). Gli elementi ritenuti si sommano ogni anno e formano la mineralomassa della fitocenosi (v. Duvigneaud, 1968).
Il ciclo geochimico è una sorta di flusso che proviene dal mondo esterno e che si interseca con il ciclo biologico. Comprende l'entrata (input) e l'uscita (output) di elementi minerali. L'entrata comprende: a) apporto da parte delle precipitazioni: polvere, pioggia, neve, ecc.; b) apporto di azoto per fissazione di N2 dell'aria da parte dei microrganismi specializzati dell'ecosistema; c) apporto di elementi chimici diversi per decomposizione della roccia madre: alcune radici penetrano in profondità nella roccia, dissolvendone certi elementi che poi trasmettono agli organi aerei, donde tornano al suolo per caduta di lettiera. L'uscita comprende: a) perdita di elementi a opera delle acque di drenaggio; b) perdita di elementi a opera dell'uomo. C'è una perdita massiccia di elementi al momento della raccolta delle produzioni primarie o secondarie e del loro trasporto al di fuori dell'ecosistema. Quest'ultimo argomento è stato oggetto di molti studi per l'agricoltura; si sa oggi che certe piante impoveriscono il suolo più di altre, e ciò è molto importante per lo sfruttamento agricolo del terreno.
Flusso di energia e produttività degli ecosistemi (metabolismo). - La biomassa (B) è teoricamente la quantità di organismi presenti per unità di superficie dell'ecosistema al momento dell'osservazione; la si valuta in numero di individui (densità) o la si misura in peso o in contenuto di energia (cal). Essa comprende una mineralomassa (ceneri) e una organomassa (massa di materia organica di individui viventi):
biomassa = mineralomassa + organomassa.
A questa biomassa si aggiungono i cadaveri che, a livello delle fitocenosi terrestri, sono soprattutto organi morti ancora attaccati alle piante viventi: si può parlare allora di necromassa (N). Una parte degli organi morti si stacca e cade al suolo. Al livello del suolo si accumula materia morta, formata dalla lettiera (L) degli organi morti che si decompone più o meno rapidamente in una massa nera, lhumus (H), a cui si aggiunge la lettiera delle radici morte nel suolo.
Per alcuni autori, la biomassa è l'insieme di tutta la materia organica, vivente o morta, compresa nell'ecosistema. È preferibile allora parlare di materia organica totale dell'ecosistema (MOT):
MOT = B + N + L + H.
La biomassa vegetale (fitomassa) è più facile da misurare della biomassa animale (zoomassa). La tab. II (v. p. 267) dà dei valori di fitomassa per alcuni grandi tipi di ecosistemi.
La velocità di produzione della biomassa è la produttività. La produttività primaria (v. fig. 23) è la velocità con la quale l'energia viene immagazzinata dall'attività fotosintetica sotto forma di materia organica, che può costituire un incremento, essere utilizzata come alimento dai consumatori, o semplicemente morire ed essere lasciata ai decompositori.
La fitocenosi produttrice dell'ecosistema si compone: a) di un sistema fotosintetico costituito da organi assimilatori, che in un ecosistema terrestre sono principalmente le foglie (F); b) di un sistema non fotosintetico, costituito da radici, organi di riserva, ecc. (NF). Si definisce produttività lorda (PL) il prodotto della fotosintesi totale del sistema fotosintetico (assimilazione totale), comprendente anche gli assimilati che saranno poi bruciati durante la ‛respirazione di mantenimento' del sistema (RMF = respirazione di mantenimento delle foglie). La quantità di materia organica assimilata che può essere esportata verso il sistema non fotosintetico,
PL − RMF = PS,
è la produttività in sovrappiù. Questa, traslocata dalle foglie verso il sistema non fotosintetico, deve assicurare il mantenimento degli organi esistenti, la costruzione di organi nuovi e l'elaborazione e l'immagazzinamento dei materiali di riserva.
Il mantenimento degli organi esistenti esige energia respiratoria; una parte di PS è pertanto consumata sotto forma di respirazione di mantenimento del sistema non fotosintetico (RMNF).
Sussiste perciò la relazione
PS − RMNF = PNA.
PNA, produttività netta di assimilazione, non sarà incorporata interamente nella costruzione di organi nuovi o di materie di riserva nuove; una parte fornirà l'energia respiratoria. Questa respirazione di costruzione (RC) deve essere dedotta da PNA; in conclusione avremo:
PN1 = PNA − RC,
dove PN1 è la produttività primaria netta (o fotosintesi apparente), cioè la somma, che si può pesare, di tutti i tessuti formati durante l'unità di tempo scelta e di tutti i materiali nuovi immagazzinati in tutti gli organi. Essa è dunque la velocità di immagazzinamento, nella fitocenosi, della materia organica formata in sovrappiù rispetto a quella consumata per respirazione:
PN1 = PL − (RMF + RMNF + RC).
Se si designa con RA (respirazione autotrofa) l'insieme delle perdite avute a opera della respirazione della fitocenosi produttrice:
RA = RMF + RMNF + RC,
la formula diventa più semplicemente:
PN1 = PL − RA.
Si usa il termine di produttività secondaria per la biomassa prodotta dai consumatori o dai decompositori. Si usa misurarla dall'intensità respiratoria.
Il flusso di energia che attraversa un livello trofico è l'assimilazione totale a quel livello, cioè la somma della biomassa prodotta e delle sostanze respiratorie che sono servite per produrla. È ancora, se ci si riferisce al metabolismo di una popolazione, la differenza fra ciò che viene consumato e quello che non è stato assimilato:
A = C − NA.
La fig. 24 schematizza il passaggio dell'energia attraverso quattro livelli di un ecosistema ideale, la biocenosi del quale sarà ridotta a una comunità vegetale produttrice e alle popolazioni di consumatori di prima classe (erbivori), di consumatori di seconda classe (carnivori) e di decompositori (batteri). Si può supporre che ogni consumatore si nutra interamente del solo alimento offertogli da questa catena (che costituisce uno schema semplificato). Il flusso energetico presenterà un rapido decrescere, poiché in natura il rendimento di produttività, quando si passa da un livello trofico al successivo, è scarso.
L'approvvigionamento di energia dell'ecosistema si può considerare proveniente dalla radiazione solare totale (cioè λ compreso fra 0,3μ e 3μ), o dalla radiazione fotosinteticamente attiva RFA (cioè λ compreso fra 0,39μ e 0,76μ), la cui intensità è circa il 50% della precedente, ma di cui solo la metà è veramente attiva, poiché la luce verde è poco assorbita dalla clorofilla.
A livello dell'ecosistema si può distinguere:
SG = irraggiamento globale G = S + D (v. sopra)

Sp = radiazione visibile che penetra nella fitocenosi (Sv − albedo)
Sa = energia luminosa assorbita dai tessuti forniti di clorofilla: è la consumazione propriamente detta (C1)
Sf = energia luminosa fissata nella fotosintesi: A1 = PL.
La produttività primaria netta (PN1) è divisa fra il contributo annuale all'aumento della biomassa (nuovi tessuti o materia di riserva) e un supporto per gli eterotrofi (ME2), composto da una frazione C2 ingerita dagli erbivori e da una frazione L1 lasciata dai decompositori, cui si aggiunge una frazione non utilizzata (NU2):
PN1 = T1 + C2 + L1 + NU2 + Es1,
dove Es1 è il materiale esportato (legno, fieno, ecc.), generalmente trascurabile.
Nell'esempio complicato di una foresta caducifoglia, l'incremento T1 è molto complesso e si compone dell'accrescimento dei tronchi, dei rami e delle radici degli alberi, dell'accrescimento delle piante erbacee e della produzione di semi.
C2 può variare in maniera considerevole da un anno all'altro (invasioni di bruchi, ecc.).
L1 si compone di organi vegetali morti e più specialmente della caduta della lettiera di foglie, di legno morto e di radici prodotte e morte durante l'anno.
La lettiera totale (Lt) può essere divisa in due parti: la lettiera nuova che corrisponde a quella prodotta dagli organi formati durante il tempo di osservazione (generalmente un anno) e che può essere simboleggiata con L1n, e la lettiera vecchia che corrisponde a quella prodotta dagli organi formati prima del periodo di osservazione (L1v). Solo L1n appartiene al flusso di energia.
L'attività degli erbivori sottrae una parte (ME2) di PN1, una frazione della quale NU2 non viene utilizzata; della frazione consumata, C2, una parte A2 viene assimilata e una parte NA2 viene eliminata sotto forma di feci. A, è in parte respirata e perduta sotto forma di CO2 (R2), oppure è eliminata sotto forma di urina (U2); il resto serve a elaborare PN2, produttività secondaria dell'erbivoro:
A2 = PN2 + R2 + U2
PN2 = A2 − (R2 + U2)
PN2 = C2 − (NA2 + R2 + U2).
PN2 comprende un aumento di peso (T2) degli individui (vecchi o giovani), ma anche la materia che è servita ad alimentare successivamente carnivori (ME3), o a produrre dei cadaveri o organi morti (L2) lasciati ai decompositori. E ancora si ha:
A3 = PN3 + R3 + U3;
PN3 (produttività secondaria dei carnivori) = A3 − (R3 + U3), e così via.
La materia organica lasciata ai decompositori (Lt) è di norma totalmente respirata (R4 = respirazione del suolo, per la massima parte) nel caso di una biocenosi in equilibrio; nel caso di una fitocenosi in progressione o di certi ecosistemi a suolo torboso, una parte di questa materia organica è fissata sotto forma di humus (H), la cui quantità aumenta di anno in anno.
Lo schema della fig. 24 esprime un fatto ben conosciuto: gli esseri viventi sono dei cattivi trasformatori di energia; il flusso energetico diminuisce fortemente lungo una catena trofica, poiché in essa le trasformazioni avvengono con cattivo rendimento.
La produttività netta di una biocenosi PNE è allo stesso tempo quella dell'ecosistema:
PNE = PL − (RA + RH)
PNE = PN1 − RH,
dove
RH = R2 + R3 + R4.
PNE è l'aumento netto di energia immagazzinata nel sistema; è l'energia contenuta nell'incremento di tessuti vivi e di materia organica morta (humus):
PNE = T + ΔH.
In un ecosistema relativamente stabilizzato, la materia organica del suolo e i detritivori che se ne nutrono sono in equilibrio; ciò viene sottolineato dal fatto che la quantità di lettiera e di humus rimane costante da un anno all'altro. RH, valore complesso che corrisponde alla respirazione di animali, funghi e batteri, può dunque essere considerata corrispondente alla massa di tutti gli organi consumati e morti.
Il climax corrisponde a uno stadio in cui nell'ecosistema vita e morte sono in equilibrio: la materia che nasce è uguale a quella che muore. In un ecosistema che non è giunto al suo stadio di climax, la vita prevale sulla morte e la dominanza relativa della materia che nasce su quella che muore è tanto più forte quanto più l'ecosistema è giovane.
L'ecosistema in azione. - Finora abbiamo parlato di un ecosistema prossimo a una condizione di stabilità. Ma un ecosistema è quasi sempre in evoluzione dinamica, che può essere considerata come un aumento progressivo di biomassa fino allo stato di climax, nel quale essa diviene costante e massima (v. fig. 25A).
I diversi aspetti della produttività in funzione dell'età possono essere dedotti dai diagrammi della fig. 25; si può notare che la produttività primaria netta è massima per una popolazione relativamente giovane, poi diminuisce progressivamente con l'età. Questo è dovuto al fatto che la biomassa fogliare resta costante, mentre quella delle parti legnose che non fotosintetizzano è in continuo aumento.
Esistono notevoli differenze fra un ecosistema giovane, in pieno sforzo dinamico, e un ecosistema maturo. La biocenosi giovane ha una produttività elevata, poiché è grande il rapporto PL/B; avvicinandosi alla maturità, aumenta B e il rapporto PL/B diminuisce. Arrivati al climax, ciò che nasce è uguale a ciò che muore e PNE = o. Nei boschi ove l'uomo coltiva alcuni appezzamenti, la PN1 è mantenuta al livello più alto (v. fig. 25C).
La struttura si modifica parallelamente: a) le catene trofiche, lineari e basate soprattutto sulla predazione, divengono reticolate e basate sui detriti; b) la diversità, inizialmente debole, diviene sempre più marcata; c) la stratificazione e l'eterogeneità spaziale, inizialmente poco organizzate, si organizzano sempre più. In una successione, una produzione di quantità caratterizza dunque l'ecosistema giovane, mentre una produzione di qualità è tipica dell'ecosistema maturo.
Una biocenosi matura è molto diversificata; in essa sono molto importanti i fenomeni di mutualismo, parassitismo, predazione, commensalismo, ecc., mentre diviene sempre più efficiente il controllo a feedback.
Stratificazione ed eterogeneità spaziale si aggiungono alla varietà specifica, per creare la diversità in seno all'ecosistema. Una tendenza importante in una successione è l'aumento della diversità biochimica. La relazione di causa ed effetto fra diversità e stabilità è così importante che le fu dedicato un intero congresso nel 1969 a Brookhaven (v. Woodwell e Smith, 1969).
Si prova soprattutto questa relazione con un aumento inverso. Nel caso di regressione; il passaggio da diversità a monotonia si accompagna con il passaggio dalla stabilità all'instabilità; quando si ha il passaggio da un ecosistema forestale più o meno vicino al climax a un ecosistema di degradazione (landa erbosa) e a un ecosistema agricolo, si osserva quanto segue: a) nell'ecosistema forestale il numero degli stadi della rete trofica è così grande e la struttura così complessa che delle perturbazioni in un qualche punto del sistema sono diluite nella massa e non provocano grandi effetti: il funzionamento normale dell'ecosistema si mantiene tale di anno in anno; b) nel sistema degradato, la catena dei predatori è spesso troncata, la produttività primaria fluttua fortemente, e così pure le popolazioni di erbivori e di carnivori, poco diversificate; nelle annate difficili, può venire consumata una gran parte della produzione primaria, e ciò accentua le fluttuazioni delle popolazioni, alcune delle quali vanno incontro a carestie; c) nei sistemi agricoli, il numero delle popolazioni vegetali è fortemente ridotto, ma la loro produzione è grande; il numero delle specie animali si riduce anch'esso, ma abbonda di specie a riproduzione rapida. Questi ecosistemi totalmente instabili possono mantenersi solo con l'introduzione di energia, nelle forme più svariate, da parte dell'uomo.
Il fatto che la diversità biotica rafforzi la stabilità dell'ambiente giustifica, in una politica di mantenimento e di sistemazione di un territorio dove vive l'uomo, la preservazione dei boschi, delle campagne, dei corsi d'acqua e di tutti i biotopi senza valore economico, anche se una tale conservazione può portare a una riduzione della produzione alimentare ed economica.
L'uomo che cerca di ottenere un massimo di produttività dall'ambiente vi fa sviluppare degli ecosistemi giovani (monocolture) o ringiovanisce gli ecosistemi maturi senza lasciarli evolvere verso il loro climax.
L'uomo però non vive solo di alimenti: gli è necessaria un'atmosfera ben equilibrata in ossigeno e diossido di carbonio; gli è necessaria dell'acqua pura; gli serve un quadro di vita bello e sano. Molte risorse essenziali sono in questo senso fornite dagli ecosistemi maturi, anche se poco produttivi. L'uomo ha dunque bisogno sia di ecosistemi giovani (molto produttivi) sia di ecosistemi maturi (poco produttivi). Un ecosistema intermedio, che aggiunga all'alta produttività di un ecosistema agricolo tutti i vantaggi di un ecosistema poco produttivo, costituisce una situazione di compromesso. Esempi di questo tipo sono gli estuari dei fiumi e le risaie.
Produttività primaria degli ecosistemi terrestri e sue zonazioni. - È inutile insistere sull'importanza, per gli uomini, della produttività primaria del tappeto vegetale, le basi del cui studio furono gettate nel 1932 da Boysen-Jensen (v., 1932).
Il vero impulso che portò a una presa di coscienza dei fini moderni dell'ecologia fu dato in Europa, nel 1960, da Walter e Lieth in un colloquio organizzato a Stoccarda. Presto si creò, nel 1963, l'IBP, International Biological Program, che aveva un tema ambizioso: produttività della biosfera e benessere dell'umanità.
Lo studio della produttività degli ecosistemi terrestri e acquatici doveva essere accompagnato dallo studio dei processi di produzione, che sono di due tipi (v. Duvigneaud, 1968): a) estrinseci: fattori del clima e del suolo (energia, acqua, materie minerali del suolo e dell'atmosfera); b) intrinseci: fattori ecofisiologici (fotosintesi, fissazione dell'azoto, utilizzazione dell'acqua). Per raggiungere questi scopi era necessario standardizzare i metodi, e a questo fine l'IBP pubblicò una serie di manuali.
L'energia solare intercettata annualmente dalla Terra è dell'ordine di 5•1020 kcal (5,8•1017 chilowattore, il che, a latitudini medie, rappresenta un impatto di 9-10•109 kcal/ha/anno. Ci si può chiedere con quale efficacia quest'energia sia utilizzata dai diversi tipi di ecosistemi. Ne abbiamo dato qualche esempio nel cap. 6, Èb, con le figg. 12, 13 e 14; la produttività primaria netta di un campo di grano, di una foresta a foglie caduche e di una prateria permanente sono rispettivamente di 10, 10 e 16 t MS/ha/anno.
Prima della venuta dell'uomo, i biomi naturali seguivano l'andamento generale dei climi, in zonazioni orizzontali parallele dai poli all'equatore, perturbate qua e là dai massicci montani e dal frastaglio dei continenti. Si può osservare una sorta di continuum nord-sud di grandi gruppi di ecosistemi: tundra, taiga, foresta temperata caducifoglia, foresta di lauri, steppe, semideserti, savane tropicali, foreste tropicali caducifoglie, foreste equatoriali (v. fig. 10). Ecosistemi a zone si succedevano anche nel senso delle altitudini, disponendosi a gradino sui versanti delle montagne.
All'incirca fino ai tropici la temperatura è il principale fattore limitante; nelle regioni intertropicali, il fattore più importante è la distribuzione delle precipitazioni nel corso dell'anno. I diversi fattori vengono poi modificati dall'esposizione al sole e, nel caso delle montagne, dalla pendenza di queste. Gli ecosistemi possono anche sconfinare dalle loro zone normali e costituire così degli ecosistemi extrazonali con le zone vicine. Pertanto il principio di zonalità dei suoli, stabilito nel secolo scorso da Dokučaev per la Russia e che si applica alle formazioni vegetali, è applicabile anche agli ecosistemi.
Nella tundra i resti vegetali si accumulano sotto forma di una lettiera, costantemente satura di acqua, poco favorevole alla vita animale; la pedofauna, poco abbondante, è concentrata nello strato muschioso e le sue possibilità di migrazione verticale sono limitate dalla temperatura troppo bassa e da una umidità eccessiva. Nella taiga del nord la pedofauna è più variata, ma la sua densità è bassa ed è limitata principalmente alla lettiera superficiale. Nella taiga del sud la pedofauna è più ricca, il suolo è secco in estate e permette la penetrazione degli animali; lo strato umifero può raggiungere uno spessore di 10-15 cm. Nella zona delle foreste miste il suolo è più caldo; una pedofauna abbondante e multiforme mischia la materia organica al suolo fino a grande profondità: lo strato umifero è di 30-40 cm. Nella zona delle steppe, lo strato di humus raggiunge il metro di spessore; in primavera, quando il suolo è ancora umido, numerosi invertebrati sono attivi, in prossimità della superficie; in estate, quando il suolo si secca, questi animali penetrano più in profondità per continuare le loro attività, e così si forma uno strato umifero enorme. Nei deserti la mancanza di acqua impedisce a una pedofauna molto ridotta di consumare resti vegetali durante gran parte dell'anno. In superficie vi è una mancanza assoluta di humus.
Questa zonalità si applica altresì alla produttività primaria netta (PN1), stimata anche dalla biomassa (B) degli ecosistemi giunti al climax.
Si può inoltre notare che gli ecosistemi più produttivi sono le foreste equatoriali e le foreste temperate caducifoglie. In effetti gli ecosistemi alluvionali e di palude, che non abbiamo trattato, sono anch'essi altamente produttivi. Malgrado la scarsa biomassa, le steppe sono relativamente produttive, ma la produttività è soprattutto a livello di radici.
Si vede anche che la produttività lorda degli ecosistemi di foreste tropicali è molto elevata, ma che una gran parte di questa viene perduta sotto forma di respirazione, di modo che la produttività netta non è più elevata di quella di una zona temperata.
In questo quadro di vegetazione, brulicante di vita animale e batterica, appare l'uomo; vive inizialmente di frutti spontanei e di caccia, poi scopre l'agricoltura e l'allevamento; con il ferro, il fuoco e i denti degli animali domestici trasforma gli ecosistemi naturali in tre categorie nettamente distinte: a) la foresta (silva), fonte di legname dagli svariati usi e territorio di caccia; b) il campo (ager), luogo di coltura di piante alimentari e tessili; c) il prato (saltus), luogo di pascolo delle mandrie (v. KuhnholtzLordat, 1938).
La zonazione naturale degli ecosistemi terrestri, a limiti spesso progressivi, fa posto a un mosaico di ecosistemi più o meno antropogeni (la selva originaria è sostituita da piantagioni monospecifiche) che si susseguono in modo brusco gli uni agli altri.
Alcuni ecosistemi naturali sussistono là dove la loro utilizzazione è difficile: dune; paludi, torbiere, rocce, ecc. divengono luoghi di conservazione delle flore e delle faune primitive. Nell'artico, dove il clima è sfavorevole all'agricoltura, la tundra copre ancora oggi superfici molto estese.
Così l'uomo va modificando la biosfera primitiva, sforzandosi di migliorare la sua produttività quantitativa e qualitativa. Si può immaginare una biosfera ideale, una sorta di paradiso terrestre risultante dai progressi dell'agricoltura e dalla valorizzazione delle risorse naturali attraverso la scienza, l'ingegneria e la tecnologia: è quella che è stata chiamata noosfera da Vernadskij.
Ma purtroppo in numerose regioni del globo e malgrado i progressi crescenti dell'ecologia agricola, diboscamenti sconsiderati o un'agricoltura mal condotta portano, per l'erosione e il disseccamento che provocano, a perdite di biomassa e di produttività; questo processo, attraverso la trasformazione in savana o in steppa, può giungere fin alla desertificazione finale; una gran parte del Sahara ha questa origine.
Nel mondo moderno una nuova forma di ecosistema assume importanza: la città (urbs) e i suoi annessi (centri industriali, autostrade, aeroporti, complessi sportivi, ecc.) che sviluppano il loro ‛suolo-supporto', a detrimento della ‛terra-nutrice' dei paesaggi naturali. L'uomo diventa sempre più un malato di civilizzazione, distrugge tutto ciò che non è utile e produttivo. Così il caos urbanistico si sostituisce ai gruppi architettonici storici, il caos turistico annienta i paesaggi naturali tipici. Negli ambiti urbani e rurali si è lontani dalla noosfera auspicata da Vernadskij e si preferisce parlare di tecnosfera.
Gli esempi citati precedentemente mostrano che in regioni temperate un ecosistema che funzioni bene ha una efficacia fotosintetica annua di 0,5-1% nei confronti dell'energia globale del sole e di 1-2% nei confronti delle RFA.
I rendimenti rispetto alle RFA sono funzione della proporzione di organi vegetali utilizzati dall'uomo:
grano: 0,6%
patate: 1,2%
erba: 2,0%
barbabietole: 2-3%.
Diversi autori hanno cercato di stabilire l'efficacia foto- sintetica massimale (Emax) per una regione determinata, nel caso in cui l'alimentazione in acqua e sostanze minerali fosse ottimale.
Per Ničiporovič (v., 1968) questa efficacia massima deve essere valutata intorno al 4,5% delle RFA durante il periodo di vegetazione attiva, fase fogliare, con temperatura media dell'aria superiore a 5-10°. Si ottiene così (v. fig. 26): a) per le alte latitudini (65-70° N) e per RFA annue di 1,5•109 kcal/ha, un periodo di vegetazione da 2 a 3 mesi e una produttività netta ottimale di 10-15 t/ha; b) per latitudini medie (35-50° N) e per RFA di 4-5•109 kcal/ha, un periodo di vegetazione da 4 a 9 mesi e una produttività netta ottimale di 30-50 t/ha; c) per le zone tropicali, le RFA si elevano a 10•109 kcal/ha, il periodo di vegetazione è di 343-365 giorni e la produttività ottimale di 100-120 t/ha. Si tratta naturalmente di ecosistemi erbacei in cui il sistema non fotosintetico ridotto non consuma la parte più grande di PL.
Se si paragonano le produttività massime calcolate con quelle realizzate, si scopre che sussistono ampie possibilità per aumentare la produttività fotosintetica della biosfera.
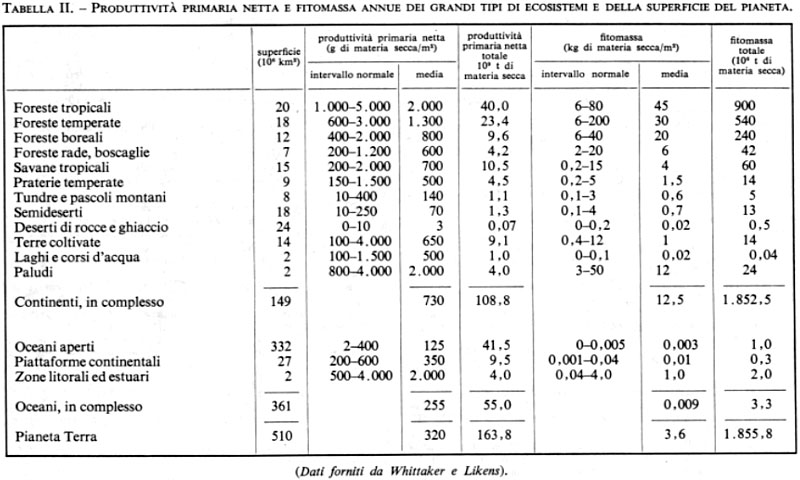
Nella tab. II è mostrata la produttività primaria per ettaro e la produttività primaria totale dell'insieme di ecosistemi (terrestri, di acqua dolce e marini) della biosfera; se ne può ricavare l'efficacia fotosintetica seguente per i grandi biomi terrestri:
silva: 0,33%
ager: 0,25%
saltus: 0,10%
tundra: 0,02%
deserto: 0,01%.
Questa produttività molto bassa dei continenti (circa 110•109 t/anno) è certo dovuta a fenomeni di degradazione antropogena di tutti i tipi; Rodin e Basilevich hanno mostrato che la produttività primaria potenziale dei continenti sarebbe (se non ci fosse l'azione umana) di 220•109 t/anno.
Ma l'intenzione degli ecofisiologi di realizzare, con la modernizzazione dei metodi e delle tecniche, un'efficacia di circa il 4,5% potrebbe portare, con la mobilitazione di immensi mezzi finanziari, a una produttività annuale dei continenti di almeno 1.000•109 t, e quindi una quantità molto più grande dell'attuale sarebbe messa a disposizione degli uomini.
7. L'ecologia e la società umana.
Si può calcolare che l'uomo disponga annualmente per la propria alimentazione di 2,6•1015 kcal, di cui gli 8/10 sono d'origine vegetale; e di 74,5•106 t di proteine, di cui meno di 1/3 di origine animale. Questa disponibilità alimentare deve servire a quasi 4 miliardi di individui (3,7•109 nel 1972). Se si considera il fabbisogno medio giornaliero di un individuo sufficientemente nutrito pari a 2.400 kcal e a 60 g di proteine, le necessità di una popolazione così numerosa assommano a quasi 1013 kcal e a oltre 2•105 t di proteine al giorno.
Attualmente la popolazione è anche più numerosa e in continuo accrescimento (v. demografia). Inoltre la ripartizione dei viveri non è affatto uniforme fra le varie regioni della terra e la quantità delle proteine animali è insufficiente, come risulta dai dati della FAO. Circa i 2/3 degli esseri umani attualmente viventi sono sottoalimentati (v. alimentazione).
Poiché la biosfera è un sistema finito e quindi le sue risorse sono limitate, si pongono a questo proposito molti gravi problemi che assillano la società umana. L'ecologia può dare qualche indicazione per la soluzione di alcuni di essi. Questi contributi sono principalmente di tre tipi: a) miglioramento delle risorse della biosfera; b) funzione protettrice degli ambienti biologici; c) formazione di una coscienza ecologica nell'opinione pubblica.
Il miglioramento delle risorse naturali può aver luogo per diverse vie: 1) perfezionamento dell'ecologia agraria sulle basi della fisiologia vegetale (v. botanica), della fitogeografia e della pedologia; 2) miglioramento genetico delle piante e degli animali per produrre varietà adatte alle diverse condizioni di coltura, per es. adottando la pratica dell'ibridazione, come è avvenuto per il granoturco (v. botanica); 3) miglioramento dei pascoli; 4) valorizzazione dei terreni incolti, in particolare delle zone aride (per le quali esiste un apposito programma dell'UNESCO); 5) lotta antiparassitaria e controllo dei pesticidi (v. lotta biologica e pesticidi); 6) sfruttamento di possibili nuove risorse, in particolare quelle del mare (v. oceanografia biologica).
La politica demografica, tendente a frenare l'incremento della popolazione mondiale, esula dai compiti dell'ecologia, ma deve tener conto dei dati che questa scienza può fornire (v. demografia).
La conservazione delle risorse naturali e la protezione degli ambienti biologici è oggi uno dei rami più importanti dell'ecologia applicata (v. natura, protezione della).
La formazione di una coscienza ecologica è, allo stato attuale, una funzione della massima importanza; essa deve essere affidata soprattutto all'insegnamento scolastico e alla diffusione delle nozioni ecologiche e di protezione dell'ambiente tramite i cosiddetti mass media (stampa a grande diffusione, radio, televisione, cinematografo, ecc.).
La gravità delle alterazioni indiscriminate degli ambienti naturali, determinate dagli sviluppi della tecnologia, dal miglioramento delle condizioni di vita delle popolazioni, dall'incremento demografico, è tale da giustificare le più vive apprensioni per il prossimo futuro dell'umanità. Perciò lo studio dell'ecologia e l'applicazione dei suoi risultati ai problemi concreti si impongono oggi su scala mondiale.
Bibliografia.
Ager, D. V., Principles of paleoecology, New York 1963.
Allee, W. C., Animal aggregations. A study in general sociology, Chicago 1931.
Allee, W. C., Emerson, A. E., Park, O., Park, T., Schmidt, K. P., Principles of animal ecology, Philadelphia-London 1949.
Andrewartha, H. G., Introduction to the study of animal populations, Chicago 1961.
Aubreville, A., Climats, forêts et désertification de l'Afrique tropicale, Paris 1949.
Balogh, J., Lebensgemeinschaften der Landtiere, Berlin 1958.
Bates, M., The forest and the sea. A study of the economy of nature and the ecology of man, London 1961.
Bates, M., Animal worlds, London 1963.
Bodenheimer, F. S., Précis d'écologie animale, Paris 1955.
Boysen-Jensen, P., Die Stoffproduktion der Pflanzen, Jena 1932.
Braun-Blanquet, J., Die Pflanzenwelt der Plessuralpen, Chur 1917.
Braun-Blanquet, J., Eine pflanzengeographische Exkursion durch Unterengadin und in den schweizerischen Nationalpark, Zürich 1918.
Braun-Blanquet, J., L'origine et le développement des flores dans le massif central de France, Paris-Zürich 1923.
Braun-Blanquet, J., Vegetations-Entwicklung und Bodenbildung in der alpinen Stufe der Zentralalpen, Zürich 1923.
Braun-Blanquet, J., Pflanzensoziologie, Zürich 1927.
Brooks, C. E. P., Climate through the ages. A study of the climatic factors and their variations, London 1926.
Burges, A., Micro-organisms in the soil, London 1967.
Burges, A., Raw, R. (a cura di), Soil biology, London-New York 1967.
Clarke, G. L., Elements of ecology, New York 1965.
Clausen, J., Keck, D. D., Hiesey, W. M., Experimental studies on the nature of species, Washington 1940.
Clements, F. E., Research methods in ecology, Lincoln 1905.
Clements, F. E., Plant physiology and ecology, London 1907.
Clements, F. E., Plant succession. An analysis of the development of vegetations, Washington 1916.
Clements, F. E., Climatic cycles and human populations in the Great Plains, Washington 1938.
Clements, F. E., Shelford, V. E., Bioecology, New York-London 1939.
Denaeyer M. E., De Smet, S., Recherches sur l'écosystème forêt. La chênaie mélangée calcicole de Virelles-Blaimont. Bilan annuel des apports d'éléments minéraux par les eaux de précipitation sous couvert forestier dans la forêt caducifoliée mélangée de Blaimont, in ‟Bulletin de la Société Botanique de Belgique", 1966, IC, pp. 345-375.
Dreux, Ph., Précis d'écologie, Paris 1974.
Dussart, B., Limnologie, Paris 1966.
Duvigneaud, P. (a cura di), Documentation. L'écologie, science moderne de synthèse, vol. II, Ecosystème et biosphère, Bruxelles 1962, 19672.
Duvigneaud, P., La productivité primaire des écosystèmes terrestres, in Problèmes de productivité biologique (a cura di M. Lamotte e F. Bourlière), Paris 1967, pp. 37-92.
Duvigneaud, P., Recherches sur l'écosystème forêt. La chênaie-frênaie à coudrier du bois de Wève. Aperçu sur la biomasse, la productivité et le cycle des éléments biogènes, in ‟Bulletin de la Société Botanique de Belgique", 1968a, CI, pp. 111-127.
Duvigneaud, P., Recherches sur l'écosystème forêt. La chênaie mêlangée calcicole de Virelles-Blaimont. Bisect biogéochimique et composition des nappes foliaires en polyéléments biogènes, in ‟Bulletin de la Société Botanique de Belgique", 1968b, CI, pp. 129-139.
Duvigneaud, P. (a cura di), Productivity of forest ecosystems, Paris 1971.
Duvigneaud P. e altri, Recherches sur l'écosystème forêt. La chênaie mélangée calcicole de Virelles-Blaimont. Biomasse, productivité et cycle des polyéléments biogènes dans un écosystème ‛chênaie mélangée' (Querceto-carpinetum calcicolum). Essai de phytogéochimie forestière, in ‟Memoires de l'Institut des Sciences Naturelles de Belgiques", 1967.
Eckardt, F. E. (a cura di), Functioning of terrestrial ecosystems at the primary production level. Proceedings of the Copenhagen Symposium, UNESCO, Paris 1968.
Ellenberg, H., Mueller-Dombois, D., Tentative physiognomic-ecological classification of plant formations of the earth, in ‟Bericht über das Geobotanische Forschungs-Institut Rübel", 1967, XXXVII, pp. 21-55.
Elton, C. S., Animal ecology, London 1927.
Elton, C. S., The ecology of invasions by animals and plants, London 1958.
Elton, C. S., The ecology of animals, London 1966.
Farb, B., L'écologie, Paris 1965.
Galoux, A., Flux et transferts d'énergie au niveau des écosystèmes forestiers, in Productivity of forest ecosystems (a cura di P. Duvigneaud), Paris 1971, pp. 21-40.
Gaussen, H., Le dynamisme des biocénoses végétales, in ‟L'année biologique", 1951, XXVII, pp. 89-102.
Gaussen, H., Expression des milieux par des formules écologiques. Leur représentation cartographique. Colloque sur les régions écologiques du globe, Paris 1954.
Gaussen, H., Bagnouls, F., L'indice xérothermique, in ‟Association des Géographes Français. Bulletin", 1952, CCXXII-CCXXIII, pp. 10-16.
Haeckel, E., Generelle Morphologie der Organismen, Berlin 1866.
Jaccard, P., Lois de distribution florale dans la zone alpine, in ‟Bulletin de la Societé Vaudoise des Sciences Naturelles", 1902, XXXVIII, 144, pp. 122-123.
Kettlewell, H. B. D., Further selection experiments on industrial melanism in the Lipidoptera, in ‟Heredity", 1956, X, pp. 287-301.
Kettlewell, H. B. D., The phenomenon of industrial melanism in Lepidoptera, in ‟Annual review of entomology", 1961, VI, pp. 245-262.
Knight, C. B., Basic concepts of ecology, New York 1965.
Kubiena, W. L., The soils of Europe, London 1953.
Kühnelt, W., Ecologie générale concernant particulièrement le règne animal, Paris 1969.
Kuhnholtz-Lordat, G., La terre incendiée, Nimes 1938.
Lamotte, M., Bourlière, F. (a cura di), Problèmes de productivité biologique, Paris 1967.
Lemée, G., Précis de biogéographie, Paris 1967.
Lewis, T., Taylor, L. R., Introduction to experimental ecology, London 1967 (tr. it.: Introduzione all'ecologia sperimentale, Milano 1973).
Liebig, J. von, Die organische Chemie in ihrer Anwendung auf Agrikultur und Physiologie, Braunschweig 1840.
Lieth, H., Die Stoffproduktion der Pflanzendecke, Stuttgart 1962.
Likens, G. E., Bormann, F. H., Pierce, R. S., Fisher, D. W., Nutrient hydrologic cycle interaction in small forested watershed-ecosystems, in Productivity of forest ecosystems (a cura di P. Duvigneaud), Paris 1971, pp. 553-563.
Lindeman, R. L., The trophic-dynamic aspect of ecology, in ‟Ecology", 1942, XXIII, pp. 399-418.
Löhnis, F., Studies upon the life cycles of the Bacteria, Washington 1922.
Lotka, A. J., Elements of physical biology, Baltimore 1925.
Macan, T. T., Freshwater ecology, London 1963.
Macfadyen, A., Animal ecology. Aims and methods, London 1957, 19632.
Maldague, M. E., Aspects faunistiques de la fertilité des sols forestiers et spécialement des sols forestiers équatoriaux, Louvain 1967.
Marcuzzi, G., Ecologia animale, Milano 1968.
Margalef, R., Ecologia, Barcellona 1976.
Maynar-Smith, J., Models in ecology, Cambridge 1974 (tr. it.: L'ecologia e i suoi modelli, Milano 1974).
Moebius, K., Die Auster und die Austernwirtschaft, Berlin 1877.
Moore, H. B., Marine ecology, London 1958.
Ničiporovič, A. A., Evaluation of productivity by study of photosynthesis as a function of illumination, in Functioning of terrestrial ecosystems at the primary production level. Proceedings of the Copenhagen Symposium (a cura di F. E. Eckardt), UNESCO, Paris 1968, pp. 261-270.
Odum, E. P., Fundamentals of ecology, Philadelphia-London 19713 (tr. it.: Principî di ecologia, Padova 1973).
Ovington, J. D., Quantitative ecology and the woodland ecosystem concept, in ‟Advances in ecological research", 1962, I, pp. 103-192.
Ovington, J. D., The better use of the world's fauna for food, London 1963.
Ozenda, P., Biogéographie végétale, Paris 1964.
Penman, H. L., Vegetation and hydrology, Farnham 1963.
Pérès, J.-M., Devèze, L., Océanographie biologique et biologie marine, 2 voll., Paris 1961-1963.
Raunkiaer, C. C., The life forms of plants and statistical plant geography, Oxford 1934.
Reid, K., Lauwerys, J. A., Joffe, J., Tucker, A., Man, nature and ecology, London 19742 (tr. it.: Uomo, natura, ecologia, Milano 1974).
Rodin, L. E., Basilevičh, N. I., Production and mineral cycling in terrestrial vegetation, Edinburgh-London 1967.
Rodin, L. E., Basilevičh, N. I., World distribution of plant biomass, in Functioning of terrestrial ecosystems at the primary production level. Proceedings of the Copenhagen Symposium (a cura di F. E. Eckardt), UNESCO, Paris 1968, pp. 45-52.
Rübel, E., Pflanzengesellschaften der Erde, Bern-Berlin 1930.
Rübel, E., Pflanzen soziologischer Aufbau, Halle 1936.
Sacchi, C. F., Testard, P., Ecologie animale: organismes et milieu, Paris 1971.
Shelford, V. E., Animal communities in temperate America, Chicago 1913.
Simonetta, A., Ecologia, Torino 1975.
Stebbins, G. L., Processes of organic evolution, Englewood Cliffs, N. J., 1966.
Tansley, A. G., The use and abuse of vegetational concepts and terms, in ‟Ecology", 1935, XVI, pp. 284-307.
Vernadskij, V. I., The biosphere and the noosphere, in ‟American scientist", 1945, XXXIII, pp. 1-12.
Volterra, V., Variazioni e fluttuazioni del numero d'individui in specie animali conviventi, in ‟Memorie dell'Accademia dei Lincei" (serie 6), 1926, II, pp. 31-113.
Volterra, V., Leçons sur la théorie mathématique de la lutte pour la vie, Paris 1930.
Waksman, S. A., Microbiology, London-Baltimore 1927.
Waksman, S. A., Soil microbiology, New York-London 1952.
Waksman, S. A., Starkey, R. I., The soil and the microbe, New York 1931.
Wallace, A. R., The geographical distribution of animals, 2 voll., London 1876.
Wallace B., Srb, A. M., Adaptation, Englewood Cliffs, N. J., 1961.
Walter, H., Die Vegetation der Erde in öko-physiologischer Betrachtung; vol. I, Die tropischen und subtropischen Zonen; vol. II, Die gemässigten und arktischen Zonen, Stuttgart 1964.
Whittaker, R. H., Woodwell, G. M., Measurement of net primary production of forests, in Productivity of forest ecosystems (a cura di P. Duvigneaud), Paris 1971, pp. 159-175.
Williams, C. B., Patterns in the balance of nature and related problems in quantitative ecology, London 1964.
Woodbury, A. M., Principles of general ecology, New York 1954.
Woodwell, G. M., Smith, H. H., Diversity and stability in ecological systems. Brookhaven Symposia in Biology, Brookhaven 1969.
Woodwell, G. M., Whittaker, R. H., Primary production in terrestrial ecosystems, in ‟American zoologist", 1968, pp. 19-30.