
Genetica medica
Genetica medica
La genetica medica è la disciplina che si occupa di studiare le modalità di trasmissione e la patogenesi delle malattie che hanno in tutto o in parte come causa alcune variazioni a livello genetico. In questa specialità medica la conoscenza degli aspetti biologici e di quelli clinici hanno pari dignità e importanza nel trasmettere a chi soffre o è a rischio per una malattia ereditaria informazioni sulla patologia stessa (consulenza genetica). Oltre alle consuete strategie cliniche utilizzate dagli specialisti delle altre discipline mediche, lo sviluppo di questa branca congiuntamente a quello delle tecniche di biologia molecolare e ingegneria genetica ha fornito nuovi strumenti per la diagnosi delle malattie ereditarie (test genetici), che utilizzando l'analisi dei geni stessi o dei loro prodotti molecolari (medicina molecolare) sono in grado di determinare in modo assai preciso l'origine prima di uno stato patologico.
La genetica medica è la scienza delle variazioni biologiche nell'essere umano in relazione allo stato di malattia e di salute. L'assetto particolare e personale delle determinanti genetiche è il fattore principale della variabilità fenotipica negli esseri umani, nonché della predisposizione a eventi patologici. Si può anzi affermare che quasi tutti gli stati patologici hanno una causa o concausa di natura genetica, anche se il peso della determinante ereditaria varia nelle differenti categorie nosologiche, con situazioni nelle quali il malfunzionamento di un singolo gene può essere considerato a tutti gli effetti il responsabile unico della patologia stessa (malattie monogeniche o mendeliane) e altre nelle quali concorrono fattori non genetici (ambientali) e fattori genetici multipli e differenti. Le patologie mendeliane costituiscono sostanzialmente occorrenze rare, mentre le malattie multifattoriali o complesse comprendono la gran parte delle affezioni comuni, quali la tendenza ad accidenti vascolari, malattie metaboliche frequenti come il diabete mellito e l'osteoporosi, nonché molte patologie psichiatriche come la schizofrenia o la depressione maggiore.
Il principio molecolare della variabilità genetica sta nei cambiamenti che possono occorrere nel complesso di informazioni che chiamiamo genoma umano. Il cambiamento di una sequenza di DNA rispetto a un modello che è il più frequente nella popolazione normale (genotipo selvatico) è considerato come una mutazione. Quando una mutazione o variante genetica in una popolazione compare con una frequenza maggiore dell'1% è denominata 'polimorfismo'. Un individuo eredita metà del patrimonio genetico da ciascuno dei genitori e le differenze tra ciò che è stato ereditato dal padre e ciò che è stato ereditato dalla madre ammontano al minimo a una decina di milioni di basi del DNA. Alcuni gruppi di varianti (aplotipi) sono specifici per determinate popolazioni e la loro evoluzione può essere utilizzata per tracciare l'origine delle etnie umane. Le variazioni nella sequenza del DNA che sono causa di malattie genetiche vengono analizzate e studiate in modo differente a seconda della loro tipologia. Le mutazioni più grandi, che interessano singoli cromosomi o parti notevoli di essi (mutazioni cromosomiche), vengono studiate con indagini di citogenetica classica, ovvero con l'individuazione dei cromosomi e delle loro anomalie.
La genetica medica: concetti generali
Le mutazioni più piccole che coinvolgono singole basi del DNA (mutazioni geniche) vengono studiate leggendo direttamente la sequenza del DNA genomico. Le moderne macchine per il sequenziamento automatico del DNA permettono la lettura di migliaia di paia di basi nucleotidiche all'ora. Mutazioni di entità intermedia (mutazioni genomiche) vengono studiate invece con metodiche di citogenetica molecolare, che coniugano tecniche di microscopia classica a tecniche di genetica molecolare, in particolare l'ibridazione con sonde fluorescenti, o FISH. L'attuale tendenza nell'evoluzione tecnologica dell'analisi del DNA è quella di fare il massimo in termini di output, costruendo piattaforme miniaturizzate per l'analisi strutturale e funzionale complessa, che possano essere usate per lo studio simultaneo delle modalità di espressione di migliaia di geni e delle caratteristiche di ampie regioni genomiche (microarrays). Le mutazioni geniche sono dette 'puntiformi' quando coinvolgono poche basi di DNA, e spesso sono dovute a errori di copiatura del DNA stesso da parte delle DNA-polimerasi durante le fasi del ciclo cellulare che precedono i fenomeni di replicazione cellulare di meiosi o mitosi. Quando la variazione nella sequenza del DNA è presente in una cellula della linea germinale, perché insorta durante il processo di formazione dei gameti (mutazione de novo) o perché presente anche in tutte le altre cellule dell'organismo (mutazione costituzionale), questa potrà essere trasmessa alla progenie. Il tasso di mutazioni geniche de novo di origine paterna è maggiore di quello di origine materna e in molti casi aumenta con l'avanzare dell'età paterna, perché molti più cicli di duplicazione del DNA occorrono nella gametogenesi maschile rispetto a quella femminile.
Tra le mutazioni, le sostituzioni di basi (le mutazioni più frequenti) sono quelle nelle quali una o più basi contigue sono sostituite da una o più basi a sequenza differente. Una sostituzione di base può avere tre importanti conseguenze. Cambiare il significato di un codone, l'unità genica che codifica per un amminoacido che a sua volta rappresenta l'unità base dei prodotti proteici (mutazione missenso), trasformare il codone in un codone di terminazione della traduzione (mutazione nonsenso), o non cambiare il significato del codone (mutazione sinonima). Le delezioni invece eliminano una o più basi dalla sequenza selvatica. La maggior parte delle delezioni e delle inserzioni (le inserzioni sono caratterizzate dall'aggiunta di basi nucleotidiche) comprendono meno di cinque nucleotidi e possono essere identificate solamente tramite il sequenziamento del DNA. Se il numero delle basi aggiunte o tolte non è un multiplo di tre, il risultato sarà un'alterazione del quadro di lettura del codice genico e la produzione di una proteina alterata e spesso tronca.
Con l'obiettivo di identificare le mutazioni responsabili di patologie genetiche, l'attenzione dei ricercatori si è per lo più focalizzata sulle regioni del genoma (5% dell'intero genoma) che codificano per sequenze proteiche. Da qui la connotazione delle mutazioni puntiformi in nonsenso e missenso, a seconda della loro influenza sulla sequenza proteica. Tuttavia molte regioni genomiche non sembrano codificare direttamente per strutture proteiche, ma piuttosto servire al controllo dell'espressione genica. Mutazioni in regioni codificanti che sembrano non avere conseguenze sul prodotto proteico (per es., mutazioni sinonime, che non cambiano il codice di lettura genico e quindi la struttura della risultante proteina) possono invece mutare la funzione di regioni regolatorie, che influenzano fenomeni biologici fondamentali, come lo splicing alternativo, ovvero il processo per il quale un singolo gene può decidere di codificare per peptidi di struttura differente.
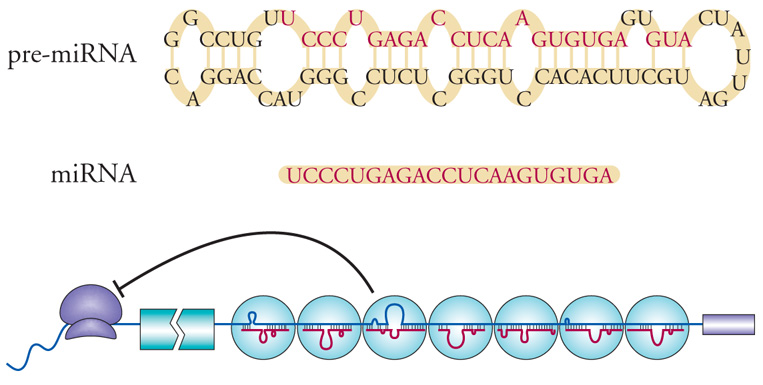
Recentemente è stata descritta una nuova famiglia genica, che rappresenta circa l'1% di tutti i geni nel nostro genoma, che codifica per piccoli RNA non tradotti detti microRNA (miRNA). Fino a oggi sono stati identificati circa 300 miRNA nell'uomo. I miRNA sono in grado di regolare la funzione genica a livello post-trascrizionale. Essi riconoscono regioni specifiche di RNA messaggeri (mRNA) bersaglio e ne impediscono la traduzione in proteina. Si pensa che circa il 30% di tutti i geni sia regolato in questo modo e che ogni miRNA possa riconoscere circa 100 geni diversi come loro bersaglio, mentre ogni singolo gene può essere regolato da differenti miRNA. Sono state anche descritte mutazioni a carico del sistema mRNA/miRNA, capaci di determinare patologie nell'uomo, come il caso di alcuni tumori del sangue o di patologie neuropsichiatriche (fig. 2).
Malattie genetiche
Se consideriamo le cause di morbilità nella popolazione generale, più del 75% degli individui sarà nella propria vita colpito da patologie con importante componente genetica, il 5% prima dei 24 anni. Di queste patologie lo 0,4% saranno legate a disordini cromosomici, il 2% a patologie monogeniche, mentre le malattie cosiddette 'complesse' rappresenteranno il gruppo di gran lunga più importante.
Eredità mendeliana. Le malattie causate in parte preponderante dalla mutazione di un solo gene, dette 'malattie mendeliane', vengono distinte in patologie autosomiche e patologie legate ai cromosomi del sesso a seconda che la posizione (locus) dei geni, le cui mutazioni sono responsabili della malattia, sia su un autosoma o su un cromosoma del sesso (in particolare il cromosoma X). A loro volta queste patologie vengono distinte in malattie recessive e malattie dominanti. In realtà la classica definizione delle patologie monogeniche, come patologie nelle quali tutti i fenomeni clinici sarebbero secondari alla mutazione di un singolo gene, è solamente una semplificazione. Esistono fattori genetici e non genetici in grado di variare gli effetti patologici delle singole mutazioni (geni e fattori modificatori), cosicché anche in una stessa famiglia, nella quale venga trasmessa la stessa mutazione genetica, i diversi portatori del difetto genetico possono mostrare caratteristiche cliniche differenti.


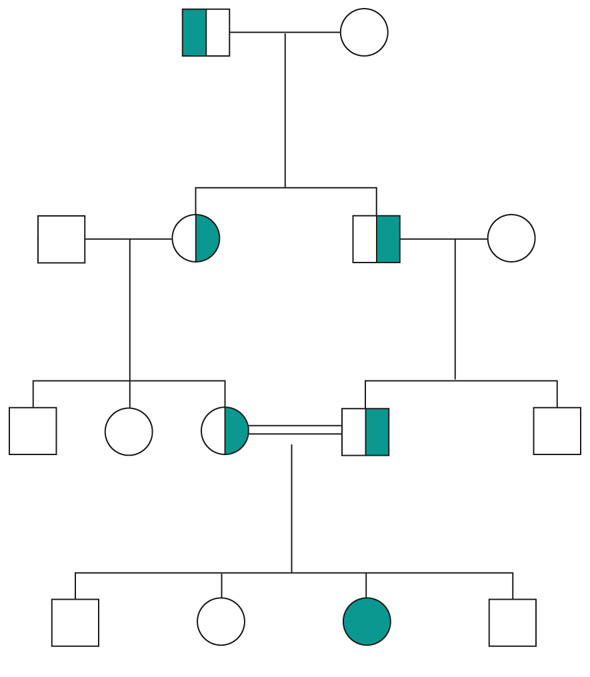
Ogni gene posizionato sugli autosomi (geni autosomici) esiste nel genoma umano in duplice copia, una copia di origine materna e una di origine paterna. Le due copie del gene (alleli) possono essere uguali (e l'individuo è detto omozigote per quel gene) o differenti (eterozigote). La distribuzione degli alleli a un singolo locus è detta genotipo, ed è il substrato molecolare per la determinazione delle caratteristiche individuali (fenotipo). I principî di dominanza determinano il modo nel quale il rapporto tra i due alleli a un singolo locus determina il fenotipo. Un allele è detto dominante (A) su un altro recessivo (a), quando il fenotipo determinato dal genotipo omozigote AA e da quello eterozigote Aa sono identici. Nelle condizioni autosomiche dominanti, un individuo è affetto se è portatore di almeno uno dei due alleli della coppia genica mutato (eterozigote). Nelle singole famiglie, la malattia colpisce entrambi i sessi in proporzioni simili e può essere trasmessa alle generazioni successive da genitori di ambedue i sessi (fig. 3). Un paziente ha generalmente un genitore affetto e il 50% della prole rischia di essere affetta. Una patologia autosomico-dominante può tuttavia colpire un individuo i cui genitori non sono affetti. Ciò può avvenire più frequentemente per la comparsa di una nuova mutazione durante la formazione dei gameti in uno dei genitori, oppure perché un genitore che costitutivamente non reca la mutazione ha alcune delle cellule della linea germinale mutate (mosaico germinale). È anche possibile che uno dei genitori sia portatore della mutazione ma non esprima il fenotipo patologico, un fenomeno che è detto 'difetto di penetranza' della mutazione (fig. 4). Quando una mutazione è completamente penetrante, tutti i portatori della mutazione saranno affetti.
Altre volte la patologia ha un'espressività variabile e alcuni individui esprimono il difetto genetico solo a livelli subclinici. Espressività variabile e difetto di penetranza sono i tipici fenomeni determinati da modificatori genetici o ambientali, e indicano come anche le malattie che generalmente vengono considerate monogeniche sono in realtà il risultato dell'azione di diverse componenti, stante un contributo importante della singola mutazione genetica. Mentre alcune forme cliniche mendeliane sono invariabilmente dovute a mutazioni di un gene singolo, altre mostrano il fenomeno della eterogeneità genetica (di locus), dove la stessa malattia può essere causata da singole mutazioni ma in geni differenti. Per esempio, più di 40 geni possono essere responsabili del fenotipo clinico della retinite pigmentosa. Questo fenomeno, oltre a essere di impaccio nella scelta del test genetico a cui sottoporre i pazienti con queste forme patologiche, sta rivoluzionando la nosologia di molte specialità mediche. Quando mutazioni differenti dello stesso gene danno origine a quadri clinici simili si parla di eterogeneità allelica. Questo fenomeno è conosciuto da tempo, e alcuni geni possono mutare in migliaia di modi differenti, e anche questa è una difficoltà quando si prende in considerazione la possibilità di un test genetico.
Il fenomeno opposto a quello della eterogeneità genetica di locus è quello della serie allelica. Qui mutazioni differenti dello stesso gene danno luogo a malattie diverse o perché regioni differenti dello stesso gene hanno distinte specificità funzionali, oppure perché il gene è espresso in diversi tessuti. Nonostante la diversità tra i Mammiferi e altre specie animali, come Vermi o Insetti, il numero di geni nei vari genomi non è molto differente. Il genoma umano contiene solo il doppio dei geni di un semplice verme. Tuttavia negli animali più complessi ogni gene è in grado di codificare per più prodotti (per di più proteici). Ovvero, i genomi evolvono non solo moltiplicando e cambiando i geni presenti, ma anche utilizzandoli in modo alternativo. Nelle patologie a trasmissione autosomica recessiva gli individui sono affetti quando sono omozigoti per alleli mutati e sono generalmente figli di individui sani eterozigoti portatori di una mutazione genica. Anche in questo caso la patologia colpisce in modo identico i due sessi, e il 25% dei figli di due portatori sarà malato, il 50% sarà portatore sano e un altro 25% sarà di sani e non portatori. Quando nella popolazione generale lo stato di portatore è raro, l'occorrenza di un individuo malato per una malattia recessiva autosomica deve sempre fare considerare la possibilità che i genitori siano consanguinei. Maggiore è il grado di consanguineità, maggiore la condivisione del genoma e quindi la possibilità di generare figli omozigoti per mutazioni per i quali i genitori siano portatori sani, mutazioni che hanno ereditato da un avo comune (fig. 5). Questo è il motivo per il quale il rischio di patologia malformativa e di ritardo mentale è superiore nei matrimoni tra consanguinei, rispetto al rischio generale.
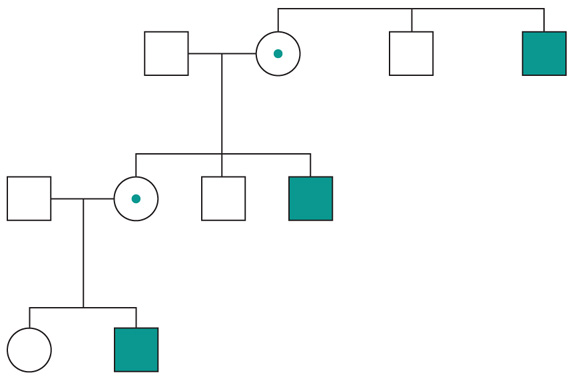
Altre patologie monogeniche sono dovute a mutazioni di geni presenti sul cromosoma X (patologie legate alla X, fig. 6). Questi geni sono presenti in duplice copia nelle femmine (che recano nelle loro cellule due cromosomi X, uno di origine paterna e uno materna) e in singola copia nei maschi emizigoti, che hanno un solo cromosoma X di origine materna e un piccolo cromosoma Y di origine paterna (che determina il sesso maschile, tramite l'azione di un piccolo gene, SRY, che è il determinante umano del testicolo). Gli individui affetti da malattie recessive legate alla X sono quasi esclusivamente maschi, con la mutazione del gene sul cromosoma X ereditato da una madre eterozigote portatrice sana oppure il risultato di un evento de novo durante la gametogenesi materna. Una portatrice avrà un rischio del 50% di avere figli maschi affetti, e del 50% di avere figlie femmine portatrici. Se un padre è affetto, tutte le figlie femmine saranno portatrici e tutti i figli maschi saranno sani, dato che un figlio maschio eredita dal padre affetto il cromosoma Y e non quello X.
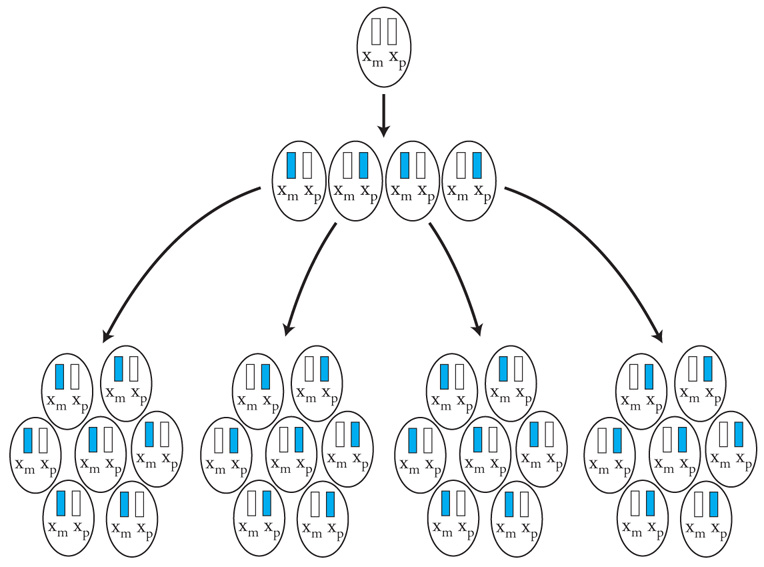
Lo sbilanciamento genico tra i due sessi per quanto riguarda il cromosoma X (due X nella femmina e una nel maschio) viene aggiustato da un'inattivazione quasi completa di uno dei due cromosomi X nella femmina (inattivazione del cromosoma X). L'inattivazione funzionale di un cromosoma X nella femmina è casuale, in modo tale che in un individuo femmina alcune cellule inattivano la X di origine paterna e altre quella di origine materna (fig. 7). L'inattivazione è legata a un cambiamento chimico del DNA e delle proteine associate, ed è in qualche modo provocata dall'azione di un gene (Xist), che è attivo sul cromosoma X inattivo e inattivo su quello attivo.
Eredità mitocondriale. Non tutti i geni nell'uomo sono presenti nel nucleo delle cellule. Un organulo cellulare, il mitocondrio, possiede un proprio piccolo genoma. I mitocondri originano probabilmente come batteri a metabolismo ossidativo endosimbionti di cellule eucariotiche primordiali, inglobati dagli eucarioti quando l'atmosfera del pianeta, in origine riducente, divenne ossidante a causa dell'accumulo di ossigeno. Questo rimasuglio del genoma originario del batterio inglobato nelle cellule contiene 37 geni, che hanno a che fare con il metabolismo respiratorio e con le funzioni di replicazione del DNA, trascrizione e traduzione all'interno del mitocondrio. Mutazioni a carico dei geni presenti nel genoma mitocondriale sono responsabili di alcune patologie umane. L'eredità di patologie dovute a mutazioni del genoma mitocondriale hanno alcune caratteristiche particolari. Un individuo eredita i mitocondri quasi esclusivamente dalla madre. Pertanto la trasmissione di queste patologie avviene sempre in modo matrilineare e i maschi affetti non trasmetteranno la malattia alla prole.
Eredità multifattoriale. La gran parte delle malattie genetiche ha modelli di trasmissione più complessi (caratteri multifattoriali) ed è determinata dall'interazione di più fattori genetici e di fattori ambientali. In queste patologie viene spesso riscontrata una certa aggregazione familiare, che tuttavia non può essere spiegata dai modelli mendeliani. Molte di queste patologie hanno una predilezione di sesso (per es., il sesso femminile nell'artrite reumatoide) e i rischi di ricorrenza in una famiglia decrescono in modo deciso con il diminuire del grado di parentela. Gran parte della ricerca genetica molecolare si sta occupando dello studio dei geni responsabili dell'aumentata o diminuita suscettibilità a queste patologie, tramite l'analisi della distribuzione nelle popolazioni di varianti genetiche comuni e la loro possibile associazione alle patologie complesse. Accanto ai classici modelli di patologie monogeniche e patologie multifattoriali stanno emergendo altre modalità di trasmissione ereditaria come quella digenica o trigenica, dove due o tre mutazioni in geni differenti sono necessarie per determinare un effetto patologico.
La consulenza genetica e i test genetici
Nell'attività del genetista medico il momento fondamentale è la consulenza genetica. La consulenza genetica è il processo per il quale pazienti o parenti, a rischio per una malattia che possa essere ereditaria, sono informati sulle conseguenze del disturbo, la probabilità di svilupparlo o trasmetterlo e sui mezzi per prevenirlo, evitarlo o migliorare la condizione clinica. Obiettivo della consulenza genetica è quello di aiutare il consultante, la coppia o la famiglia a comprendere le informazioni mediche, inclusa la diagnosi, la prognosi e le terapie disponibili; rendersi conto del contributo ereditario alla malattia e del rischio di ricorrenza; prendere le decisioni che sembrano appropriate in rapporto al rischio di ricorrenza, ai progetti familiari, agli standard etici e religiosi e agire in accordo con queste decisioni; ottenere il miglior possibile adattamento alla malattia in un soggetto affetto o a rischio di ricorrenza. Durante la consulenza il genetista medico traccia un albero genealogico della famiglia (pedigree), pone una diagnosi clinica, stima i rischi di occorrenza e di ricorrenza della patologia, informa sulle caratteristiche clinico-genetiche della stessa e consiglia eventuali test genetici, atti a verificare o scartare l'ipotesi clinica di patologie ereditarie.
Il test genetico è detto 'test diagnostico' quando viene fatto su persone che hanno o possono avere un particolare disordine. Un test è presintomatico, se la sua positività in un individuo sano indica il futuro sviluppo quasi inevitabile della malattia. Un terzo tipo di test è quello predittivo, che copre un ampio range di situazioni, il risultato del quale determina un aumentato o ridotto rischio di una malattia, ma con un grado di certezza non completo. I test genetici possono essere eseguiti prima della nascita (test prenatali) su tessuto o materiale fetale, come i villi coriali, le cellule del liquido amniotico, il liquido stesso, o il sangue, o dopo la nascita (in generale sul sangue o su altri liquidi organici). Vi sono test genetici che vengono eseguiti su particolari gruppi di individui (screening genetici). Un test di screening può essere applicato a tutta la popolazione (per es., i test neonatali), a sottopopolazioni particolari (per es., lo screening per la malattia di Tay-Sachs negli ebrei aschenaziti, dove ha un'incidenza notevolmente superiore alle altre etnie), o come test a cascata sui parenti di un paziente affetto da una patologia genetica, che fossero essi stessi a rischio di sviluppare o trasmettere la malattia. Test particolarmente utili sono quelli di citogenetica.
La citogenetica
La citogenetica è quella branca della genetica che studia i cromosomi, la loro struttura, funzione e alterazioni. Le 23 paia di cromosomi possono essere identificate in considerazione di alcune caratteristiche particolari, che sono la loro lunghezza, la posizione di una costrizione che contiene una struttura particolare detta 'centromero', e, come risultato di alcune metodiche di colorazione, dal pattern di formazione di bande chiare e scure lungo il cromosoma stesso (bandeggio cromosomico) la cui distribuzione è tipica per ogni cromosoma.
Nel campo della genetica medica, la citogenetica studia le deviazioni nell'assetto cromosomico di un individuo (aberrazioni cromosomiche) rispetto alla situazione di normalità tipica della popolazione di riferimento sana. Le aberrazioni hanno una frequenza significativa in situazioni come poliabortività, infertilità, malformazioni congenite, ritardo mentale. La maggior parte delle anomalie cromosomiche esercita il suo effetto aumentando o diminuendo la quantità di materiale genetico, che può variare da interi cromosomi a piccole porzioni di essi. Recentemente sono state sviluppate metodiche di analisi capaci di identificare anche piccole alterazioni. Accanto a casi evidenti di sbilanciamento strutturale dei cromosomi, sono stati decritti anche casi di sbilanciamenti di tipo funzionale. Per esempio, se un individuo con un assetto cromosomico normale (quindi con un genoma bilanciato) ha ereditato i due elementi di una coppia cromosomica da uno dei genitori, anziché un cromosoma da ciascun genitore (disomia uniparentale), può mostrare uno stato di grave patologia (fig. 8). Ciò è probabilmente dovuto al fatto che alcuni alleli vengono espressi in modo differenziale in base all'origine parentale, ovvero se trasmesso dalla madre (imprinting paterno) o dal padre (imprinting materno). Questo significa che, dei due alleli di una coppia, solo uno viene espresso a seconda se l'imprinting è paterno o materno. Nel caso di disomia uniparentale per una regione contenente geni con imprinting, la mancanza del complemento genico di un genitore può avere quindi le stesse conseguenze dell'assenza del gene stesso.
I progressi delle biotecnologie applicate alla patologia umana ci spinge alla sistematizzazione di una medicina individualizzata alle caratteristiche genetiche del singolo. Ne è testimone il fiorire di studi in campo farmacogenetico. La farmacogenetica è lo studio della risposta individuale a un farmaco, e mette in relazione un dato profilo genetico con le modalità di questa risposta o con la comparsa di fenomeni avversi. Meno entusiasmanti sono i risultati degli studi ormai più che ventennali sulla possibilità di una terapia mirata alla sostituzione genetica (terapia genica). Infine, bisogna notare come la genetica medica presenti numerose problematiche di carattere etico, dato che la cognizione delle proprie caratteristiche genetiche può, per esempio, influenzare le scelte riproduttive (interruzione di gravidanza, fecondazione medicalmente assistita, rinuncia alla procreazione). Diversamente dalle altre discipline cliniche, è determinante il fatto che in questa branca medica la distribuzione del carico morale delle decisioni, che devono essere assunte in presenza di un rischio o di una certezza di patologia ereditaria spesso pregiudizievole per la qualità di vita del paziente e della sua famiglia, non ricade esclusivamente su chi fornisce la diagnosi, ma anche sulla persona che richiede la consulenza, nonché, in senso lato, sulla società e sulle sue scelte di ordinamento.
Bibliografia
Antonarakis 2003: Antonarakis, Stylianos E., Mutations in human disease: nature and consequences, in: Emery and Rimoin's principles and practice of medical genetics, edited by David L. Rimoin e altri, London, Churchill Livingstone, I, 2003, pp. 83-103.
Connor 2003: Chromosomal disorders, edited by Michael Connor, in: Emery and Rimoin's principles and practice of medical genetics, edited by David L. Rimoin e altri, London, Churchill Livingstone, I, 2003, pp. 1129-1230.
Cook 2003: Cook, Jackie, Mendelian inheritance, in: Emery and Rimoin's principles and practice of medical genetics, edited by David L. Rimoin e altri, London, Churchill Livingstone, I, 2003, pp. 104-124.
Korenberg 2003: Korenberg, Julie R., Chromosomal basis of inheritance, in: Emery and Rimoin's principles and practice of medical genetics, edited by David L. Rimoin e altri, London, Churchill Livingstone, I, 2003, pp. 149-173.
Lewin 1994: Lewin, Benjamin, Genes V, New York, Oxford University Press, 1994.
Sturtevant 1965: Sturtevant, Alfred H., A history of genetics, New York, Harper & Row, 1965.
Wallace 1992: Wallace, Douglas C., Diseases of the mitochondrial DNA, "Annual review of biochemistry", 61, 1992, pp. 1175-1212.
Wallace 2003: Wallace, Douglas C., Mitochondrial genes in degenerative disease, cancer and aging, in: Emery and Rimoin's principles and practice of medical genetics, edited by David L. Rimoin e altri, London, Churchill Livingstone, I, 2003, pp. 299-409.