
Genetica
Genetica
Introduzione
La genetica è il ramo della biologia che studia la trasmissione dei caratteri ereditari, la variazione biologica individuale, l'evoluzione. Per raggiungere questi obiettivi è stato necessario capire anche la struttura fisica e chimica del materiale che forma gli organismi viventi, e in particolare di quello che ha la proprietà di trasmettere l'informazione biologica: il DNA o acido desossiribonucleico. È questa la sostanza base dei geni e delle strutture che li contengono, i cromosomi, e pertanto la genetica è divenuta il nucleo centrale della biologia.
Alla base dell'interesse delle scienze sociali per questo ramo della biologia sta la convinzione che l'origine e la variabilità genetica della nostra specie costituiscano una parte importante della nostra storia sociale, e il riconoscimento che anche i fenomeni culturali, non solo quelli naturali, hanno radici profonde che vanno ricercate lontano nel tempo. Con ciò non si vuole sostenere che la costituzione genetica dei nostri antenati sia stata un fattore determinante nella storia successiva della nostra specie: la realtà è che sappiamo troppo poco, per non dire nulla, delle strutture genetiche che influenzano il comportamento umano. Il motivo d'interesse è un altro: i fenomeni storici e culturali, quindi anche la storia delle istituzioni sociali, hanno spesso un andamento parallelo a quello dei fenomeni di cambiamento genetico. Non riteniamo che i geni determinino storia e cultura; se talora questo determinismo si manifesta, assume forme speciali e molto spesso è apparente. Più frequente e documentabile è il fenomeno opposto: sono la storia e la cultura, e gli istituti e i comportamenti sociali che ne risultano, a influenzare la composizione genetica di una popolazione.
Il termine 'genetica' è stato usato per la prima volta nel 1905 dal biologo inglese William Bateson in una lettera e da lui ufficialmente proposto l'anno successivo, in occasione della terza International Conference on Hybridization and plant breeding organizzata a Londra dalla Royal Horticultural Society, con la seguente definizione: "la genetica è la scienza che studia l'eredità e la variazione cercando di scoprire le leggi che governano le somiglianze e le differenze negli individui che sono in rapporto di discendenza". Lo stesso Bateson precisò che i fenomeni dell'eredità e della variazione sono "implicitamente rilevanti per i problemi teorici propri dell'evoluzionista e del sistematico, oltre a offrire un'applicazione ai problemi pratici dell'allevatore di animali e di piante". Da quell'epoca lo sviluppo della genetica è stato travolgente e si è riflesso in tutte le scienze biologiche e mediche.
La semplice lettura del sommario di questo articolo è sufficiente a indicare il taglio e le inevitabili limitazioni con cui verrà esposta questa disciplina. I primi quattro capitoli sono dedicati a un inquadramento sintetico della materia, mentre gli altri quattro trattano problemi che ci sembrano rilevanti per lo studioso di scienze sociali. Per rendere più esplicita la scelta degli argomenti si è privilegiato ciò che riguarda la genetica dell'uomo normale (non affetto da patologie) e delle popolazioni umane. Non si considereranno perciò la genetica dei virus e dei batteri, la genetica vegetale e la genetica medica. Lo studio della genetica molecolare, oggi in continuo sviluppo, sta alla base di quei processi di manipolazione dei geni che, invadendo impetuosamente ogni campo della genetica, della biologia e della medicina, vengono oggi classificati col nome generico di ingegneria genetica. Non potremo trattare l'argomento in modo sistematico per la complessità delle tecnologie di laboratorio e dei problemi coinvolti, ma è ovvio che le implicazioni sociali ed economiche delle innovazioni che tali tecnologie permettono costituiranno nei prossimi anni realtà con cui dovranno necessariamente misurarsi le nostre stesse strutture sociali, giuridiche e politiche: un esempio di tali implicazioni verrà illustrato nel capitolo conclusivo.
Definizioni e concetti generali
L'informazione genetica è presente nel nucleo di ciascuna cellula di un organismo. È contenuta in strutture chiamate cromosomi, di cui, nella nostra specie, Homo sapiens sapiens, esistono 23 coppie per cellula. Ogni coppia è costituita da un cromosoma trasmesso dal padre e da un cromosoma trasmesso dalla madre, morfologicamente indistinguibili. I cromosomi sono fondamentalmente costituiti da acido desossiribonucleico (DNA), una lunga catena di molecole chiamate nucleotidi o basi. Esistono quattro diversi tipi di nucleotidi indicati dai simboli A, C, G, T. Un segmento di DNA può sembrare una sequenza lineare casuale di nucleotidi, per esempio TAACATGCCAT... . L'ordine dei nucleotidi è in realtà responsabile della successione di eventi biochimici che, partendo dall'informazione contenuta in quell'ordine, inducono la sintesi di un prodotto nella cellula, per esempio una proteina, un enzima, ecc. Tale ordine costituisce l'informazione genetica, così come le lettere dell'alfabeto in successione costituiscono le parole e quindi dei significati ('informazioni') che si possono comunicare. La sequenza dei nucleotidi viene copiata da cellula a cellula all'interno di ogni individuo, in pratica senza errori. Attraverso le cellule sessuali o germinali le catene di DNA dei genitori passano ai figli essenzialmente identiche, e questo è il meccanismo che assicura il mantenimento delle proprietà specie-specifiche dell'uomo. I geni sono segmenti di DNA deputati alla sintesi di prodotti genici. Oggi possiamo studiare direttamente il DNA con metodi chimici, ma fino a poco tempo fa dovevamo limitare il nostro studio al fenotipo, cioè a un prodotto genico che si poteva manifestare nell'aspetto esterno dell'organismo (come il colore azzurro degli occhi, la presenza del gruppo sanguigno Rh positivo, ecc.). La determinazione genetica, cioè il fatto che il carattere fenotipico esaminato sia effettivamente ereditario, si accerta esaminando la sua trasmissione familiare dai genitori ai figli. La nostra specie ha circa 100.000 geni, ma quelli identificati sono circa 2.500. Occorre tuttavia chiarire che la maggior parte dei caratteri che osserviamo non sono strettamente genetici: al più vi è una predisposizione ereditaria (per esempio la tendenza ad avere la pressione alta, un alto livello di colesterolo, forse anche a contrarre l'abitudine al fumo). Vi sono inoltre importanti cambiamenti dell'ambiente di vita, alcuni dei quali possono essere trasmessi per via culturale e socioeconomica. Senza dubbio c'è una componente genetica nel carattere statura, ma anche l'alimentazione esercita un'influenza; abitudini e coercizioni che la riguardano sono trasmesse per via culturale, un'eredità diversa da quella genetica, che non di rado viene con essa confusa. I caratteri più utili per studiare la variabilità e l'evoluzione biologica sono quelli in cui il fenotipo permette di ricostruire con certezza la costituzione genetica dell'individuo; le loro regole di trasmissione sono ben conosciute (leggi di Mendel, v. cap. 3) e le loro frequenze sono per lo più molto stabili nel tempo, ma possono variare da popolazione a popolazione.
A ogni divisione cellulare il DNA viene replicato, così che le due cellule figlie risultanti contengono l'identica catena di DNA della cellula madre. Con una probabilità molto piccola (dell'ordine di 10-⁸) questo processo di replicazione del DNA avviene con qualche errore, per esempio un nucleotide viene sostituito da un altro dei quattro possibili, oppure vengono aggiunte o cancellate sequenze più o meno estese di nucleotidi. Si parla in questi casi di mutazione. L'effetto di una mutazione sull'organismo dipende dall'alterazione della funzione del gene specifico il cui DNA risulta alterato. Le mutazioni a carico delle cellule non sessuali (somatiche) possono indurre conseguenze gravi (per esempio tumori) nell'organismo in cui si manifestano, ma non vengono trasmesse ai figli. Le mutazioni a carico delle cellule sessuali, quale che sia il loro effetto, vengono invece trasmesse ai figli e sono quelle che interessano la genetica. L'effetto della mutazione è la creazione di un nuovo gene che è in genere leggermente diverso dal vecchio gene: i due tipi vengono chiamati alleli di quel gene. Se l'individuo in cui è avvenuta la mutazione raggiunge lo stadio adulto e ha dei figli, può avvenire che il nuovo allele generato per mutazione venga trasmesso a uno dei figli e così via di generazione in generazione. Può anche avvenire, tuttavia, che il gene mutato venga 'perso' o che, per influenza di forze evolutive che saranno descritte in seguito, aumenti la sua frequenza sino alla sostituzione completa dell'allele mutato. Questa sostituzione di un allele 'vecchio' con un allele 'nuovo' costituisce il processo elementare dell'evoluzione: l'integrazione di questi processi elementari estesa a tutti i geni da una generazione all'altra genera un cambiamento graduale, molto lento nel tempo, della struttura genetica di una popolazione.
La maggior parte delle mutazioni avviene in geni differenti, ma nel tempo lo stesso gene può mutare più volte. In questo modo possono coesistere nella stessa popolazione molti alleli dello stesso gene: si dice allora che il gene è polimorfico oppure che è presente un polimorfismo. I geni che controllano le malattie ereditarie sono polimorfici perché esistono comunemente due alleli dello stesso gene: quello che genera la malattia, molto poco frequente, e quello comune o 'normale' presente nell'individuo che non manifesta la malattia. Esistono tuttavia anche polimorfismi non associati alla manifestazione di una malattia, la cui funzione è ignota: vengono chiamati operativamente (fino a prova del contrario) polimorfismi neutri e sono quelli più comunemente usati per ricostruire la variabilità tra individui e/o di una popolazione rispetto a un'altra. In questo senso i prodotti di tali geni, o le sequenze dei geni stessi, vengono anche denominati marcatori genetici. Quelli più importanti possono classificarsi come segue.
1. Gruppi sanguigni, identificati mediante tecniche immunologiche, che consentono di individuare particolari sostanze che si trovano sulla superficie dei globuli rossi o bianchi (eritrociti o leucociti) del sangue. Esempi di gruppi sanguigni: AB0, Rh, Kell, Duffy.
2. Proteine ed enzimi, identificati mediante tecniche elettroforetiche. Le proteine prodotte dai geni vengono fatte muovere in un campo elettrico (elettroforesi) e dallo studio della loro mobilità, che dipende dalla loro carica elettrica che a sua volta dipende dalla loro struttura chimica, si inducono gli alleli polimorfici dei geni che ne controllano la sintesi. I polimorfismi più studiati sono quelli relativi a proteine o enzimi che si trovano nel plasma o nel siero del sangue o sulla superficie degli eritrociti. Esempi: emoglobina, aptoglobina, fosfatasi acida.
3. Immunoglobuline, identificate mediante tecniche immunologiche. Sono i comuni 'anticorpi' del sistema immunitario, per esempio le gammaglobuline.
4. Polimorfismi del DNA, identificati non più dal loro prodotto ma dallo studio diretto di segmenti di DNA. Oggi (1992) tale studio può essere condotto usando prevalentemente due metodi che fanno ricorso alle nuove tecnologie dell'ingegneria genetica. Il primo metodo viene chiamato 'analisi dei siti di restrizione' e consiste nell'impiego di una batteria di enzimi batterici (enzimi di restrizione) che hanno la proprietà di tagliare le sequenze nucleotidiche del DNA in siti specifici costituiti da 4, 6, o (raramente) più nucleotidi (siti di restrizione). Una mutazione in queste sequenze specifiche può impedire il taglio o può introdurne uno nuovo. I frammenti di DNA che risultano da questa reazione avranno perciò lunghezze diverse a seconda della presenza o meno della mutazione e tale diversità di lunghezza si può identificare mediante elettroforesi. Con questa tecnica è potenzialmente possibile identificare migliaia e forse milioni di polimorfismi; al momento sono stati studiati circa 2.000 polimorfismi di questo tipo. L'altro metodo di identificazione dei polimorfismi del DNA è più diretto e definitivo, e consiste nell'analisi della sua sequenza o almeno di segmenti parziali della stessa. Poiché il numero di nucleotidi nel DNA umano è stimato attorno a circa 3 × 10⁹, questo è anche l'approccio più ambizioso al problema, sul quale sono impegnati molti progetti e risorse finanziarie a livello internazionale. Si noti che l'utilità di sequenziare il DNA dei geni umani è indiscutibile se si vogliono identificare i polimorfismi: a questo scopo sono molto interessanti i tentativi di amplificare e analizzare tracce di DNA in reperti utili per indagini dirette all'identificazione personale (per esempio in sperma, capelli, saliva) o addirittura in reperti fossili antichi non troppo danneggiati. Molto più complesso è tuttavia il problema - fondamentale anche ai fini applicativi - di associare a ciascun polimorfismo, che per definizione può essere diverso da individuo a individuo, la sua funzione biologica. In altre parole, del DNA conosciamo l'alfabeto, potremo identificare la sequenza intera di lettere, ma non sarà facile distinguere una parola dall'altra e soprattutto decifrarne il significato.
Caratteri mendeliani
In ciascun cromosoma i geni hanno un ordinamento preciso e ciascun gene (segmento di DNA) occupa una posizione specifica, detta locus (plurale: loci). Nelle cellule sessuali (o germinali) della nostra specie (ovociti, presenti nell'ovaio della donna, e spermatociti, presenti nei testicoli dell'uomo) avviene un processo chiamato meiosi, che consiste in una riduzione del numero di cromosomi. Tale processo - necessario perché altrimenti la riproduzione sessuale, che avviene mediante la fecondazione e la fusione di due cellule germinali, raddoppierebbe il numero di cromosomi a ogni fecondazione - venne chiarito solo nel 1930 e si può schematizzare immaginando, per semplicità, di esaminare un organismo con soli 3 cromosomi. Il nucleo della cellula di tale organismo avrà 6 cromosomi, 3 di origine paterna e 3 di origine materna, che possiamo descrivere con il simbolo 1p1m2p2m3p3m (p=paterno, m=materno): sono presenti 3 coppie di cromosomi omologhi. Il meccanismo della meiosi, che avviene esclusivamente nelle cellule sessuali, consiste nella generazione di cellule con nuclei di soli 3 cromosomi, la cui origine paterna o materna è assortita in modo casuale (gameti): in altre parole, il gamete può essere con pari probabilità 1p2p3p, 1p2p3m, 1p2m3p, 1m2p3p, 1p2m3m, 1m2p3m, 1m2m3p, 1m2m3m. Quando il gamete prodotto dall'organismo di sesso maschile (uno degli 8 possibili) si fonde con il gamete prodotto dall'organismo di sesso femminile, si ricostituisce una cellula di 3 coppie di cromosomi da cui, per duplicazioni successive, si svilupperà l'individuo intero.
Grande merito dell'abate Gregor Mendel fu comprendere le conseguenze di questo meccanismo su basi esclusivamente sperimentali, senza conoscere né la struttura cromosomica né il meccanismo della meiosi. I suoi famosi esperimenti sui caratteri presenti nelle piante di pisello (iniziati nel 1856 e presentati - senza essere compresi fino al 1900 - alla Società di Storia Naturale di Brno nel 1865) lo condussero a formulare due principî, o leggi, che stanno alla base della trasmissione dei caratteri genetici. La prima legge, o legge della segregazione, afferma che ogni individuo eredita, per un dato carattere, un allele dal padre e uno dalla madre; i due alleli non vengono modificati o fusi nell'individuo che li eredita, ma si separano (segregano) durante la formazione delle cellule sessuali che daranno poi origine a una nuova generazione di individui e vengono trasmessi in modo indipendente l'uno dall'altro. La seconda legge, o legge dell'assortimento indipendente, afferma che alleli di geni i cui loci sono localizzati su cromosomi diversi si distribuiscono nei gameti in modo indipendente l'uno dall'altro, senza influenze reciproche.
Le implicazioni concettuali e pratiche delle leggi di Mendel, che oggi sembrano così elementari, segnano in realtà un momento critico nel pensiero biologico, forse paragonabile a quello introdotto dal principio di indeterminazione della meccanica quantistica nella fisica del nostro secolo. Per uno strano destino di coincidenze mancate, per ben 45 anni la potenzialità innovativa delle leggi di Mendel non fu compresa. Uno dei problemi della teoria darwiniana della selezione naturale era quello di individuare la fonte della potenzialmente illimitata variazione biologica. Ogni individuo è unico e differente da ogni altro. "Queste differenze individuali assumono per noi la massima importanza perché sono spesso ereditarie [...] e perché forniscono il materiale su cui la selezione della natura può agire, accumulandole" (v. Darwin, 1859; tr. it., p. 56). Ma come si determina questa variazione? Qual è la sua fonte? Su questa domanda Darwin si tormentò tutta la vita senza venire a conoscenza della semplice risposta implicitamente offerta da Mendel: la variazione si genera nelle pressoché infinite combinazioni dei caratteri che passano nei gameti da una generazione all'altra, ed è moltiplicata dalla potenzialità combinatoria del meccanismo della riproduzione sessuale che unisce a caso coppie di gameti.
Lo sviluppo quasi esplosivo della genetica dopo la riscoperta del lavoro di Mendel (attorno al 1900) suggerisce che in esso vi fosse un nucleo innovatore che ha permesso alla disciplina di fare un nuovo balzo in avanti dopo più di trent'anni di speculazioni sbagliate o almeno premature. In uno sforzo di sintesi forse troppo sommario, il contributo più significativo della sua opera può articolarsi in questi punti.
A. Semplicità del disegno sperimentale. I caratteri sono semplici, ben identificati e misurabili; gli organismi scelti, a cominciare dalla pianta del pisello, hanno un tempo di generazione breve, per cui è possibile osservare facilmente più generazioni. Era pensiero comune dei naturalisti contemporanei di Mendel che le leggi dell'ereditarietà fossero diverse per i caratteri discontinui (per esempio il colore degli occhi) e per quelli continui (per esempio l'altezza). A questi ultimi erano soprattutto rivolte le osservazioni degli scienziati che avevano operato prima di Mendel, basate sull'idea che l'evoluzione riguardasse soprattutto i caratteri continui. Fu proprio la difficoltà di comprendere la trasmissione di un carattere continuo a bloccare l'evoluzionismo premendeliano.
B. Natura discreta degli elementi ereditari. I concetti di gene e di allele, sia pure denominati con termini diversi, erano indispensabili per una interpretazione coerente degli esperimenti di Mendel (che non furono pochi: in sette anni di lavoro ibridò più di 30.000 piante). Anche Darwin e De Vries postulavano l'esistenza di elementi (gemmule, pangeni) preposti al controllo dei caratteri ereditari, ma la teoria di Mendel presenta una differenza sostanziale: mentre gli altri autori ipotizzavano l'esistenza di molti determinanti tra loro identici per un dato carattere in ogni cellula, e pensavano che queste repliche venissero trasmesse in molti esemplari alle cellule germinali, Mendel si riferiva chiaramente a un elemento singolo, quello che oggi chiamiamo gene. In tal modo la teoria risulta particolarmente semplice, nel senso che la trasmissione dei caratteri da genitori a figli si manifesta seguendo regole probabilistiche elementari, immediatamente verificabili e universali.
C. Sconfitta definitiva della teoria dell'eredità per fusione. La conclusione di Mendel, secondo cui ogni carattere si presenta in una cellula uovo fecondata (chiamata oggi zigote) con due e solo due fattori, uno derivato dal padre e l'altro derivato dalla madre, i quali possono essere differenti, fu la nuova idea che rivoluzionò la genetica. Infatti, se i fattori (oggi geni) derivati dal padre e dalla madre sono differenti, non si vede la ragione per cui dovrebbero fondersi, come sosteneva la maggior parte dei naturalisti contemporanei di Mendel; è più naturale supporre una loro separazione durante la formazione delle cellule germinali o addirittura anche dopo la fecondazione. Tale idea, rivelatasi poi esatta, andava contro l'apparenza nel caso dei caratteri continui. Si pensi al già menzionato carattere 'statura' nelle numerose famiglie con un genitore di alta e un genitore di bassa statura e figli di statura intermedia: è naturale, in tal caso, pensare a una 'fusione' del carattere come se, nella fecondazione, due fattori si mescolassero per dar luogo a un risultato 'medio'. Solo molti decenni più tardi fu compreso che: 1) i caratteri continui, quando trasmessi per via genetica, sono il risultato dell'azione simultanea di molti geni; 2) la statura in particolare non è un carattere mendeliano semplice, nel senso che alla sua espressione, oltre che una componente genetica con trasmissione mendeliana, contribuiscono certamente fattori ambientali, come un'alimentazione che migliora di generazione in generazione, ecc.
D. Esistenza di più forme dello stesso gene (alleli). l fatto che un determinato carattere (fenotipo) sia il risultato dell'azione di due alleli dello stesso gene permette di calcolare in modo preciso la trasmissione (segregazione) di quel carattere nei figli. Se un gene ha quattro alleli, a, b, c, d, e due individui con coppie di alleli rispettivamente ab e cd si uniscono, ai loro figli verranno trasmesse le coppie di alleli ac o ad o bc o bd ciascuno con probabilità 1/4. Merito particolare di Mendel fu anche quello di ipotizzare che l'espressione di certi alleli può dominare su quella di altri, nel senso, per esempio, che il fenotipo generato dalla coppia di alleli ab può essere identico a quello della coppia aa (a domina su b). Esempio tipico è il fenotipo 'colore degli occhi', dove il colore scuro domina sul colore azzurro. Due individui con il fenotipo 'occhi scuri', determinato in entrambi dalla coppia ab, possono avere sia figli con occhi scuri (coppie di alleli aa o ab), con probabilità 3/4, sia figli con occhi azzurri (coppie di alleli bb), con probabilità 1/4. Il rapporto 3/4:1/4 = 3:1 viene chiamato rapporto di segregazione.
Le frequenze geniche: evoluzione nel tempo e distribuzione nello spazio
Si consideri il gene che determina il gruppo sanguigno Rh positivo o negativo. Ogni individuo può essere identificato come positivo o negativo a seconda del risultato di un test immunologico: se si esamina un campione di individui, si troveranno individui positivi e individui negativi. La loro distribuzione percentuale è tanto più precisa quanto più esteso è il campione e in genere differisce da campione a campione. Se, per esempio, si considera un campione sufficientemente numeroso degli abitanti della Sardegna, si trova che la frequenza degli individui Rh negativi è del 4%; ma lo stesso tipo di indagine compiuto sulla popolazione basca, che vive nella regione dei Pirenei tra Francia e Spagna, rivelerebbe che fino a 30 Baschi su 100 hanno il gruppo sanguigno Rh negativo. Questi sono i due valori estremi nella variazione delle frequenze del gene Rh in Europa. I geni che esistono in più forme riconoscibilmente diverse (come positivo e negativo per l'Rh) mostrano in genere variazioni della frequenza di almeno due di esse in popolazioni diverse: anche popolazioni vicine sono raramente identiche fra loro. La differenza di frequenze dello stesso gene in due popolazioni indica la distanza genetica fra di esse; di solito si usano formule più elaborate che la semplice differenza tra le frequenze, ma è importante soprattutto prendere in considerazione geni diversi e calcolare distanze genetiche medie. La variabilità genetica tra popolazioni, tradotta quantitativamente nelle reciproche distanze genetiche fra le popolazioni prese a due a due, costituisce il 'documento' di base sul quale lavora chi studia l'evoluzione biologica delle popolazioni umane.
Rispetto al documento storico o archeologico, quello 'genetico' ha un difetto e un pregio. Il difetto è quello di riferirsi alle popolazioni oggi esistenti, e perciò di rappresentare il risultato sincronico di un processo evolutivo la cui sequenza diacronica va ricostruita a ritroso, senza possibilità di verificare sperimentalmente le varie fasi del processo stesso (oggi abbiamo la possibilità di studiare i geni di cadaveri antichi, ma permangono problemi tecnici da risolvere). Il pregio deriva da una proprietà importante del cambiamento biologico: la sua estrema lentezza. Il processo fondamentale di cambiamento biologico di tutti gli organismi viventi, la mutazione, consiste, come si è detto, in un cambiamento casuale del messaggio chimico costituito dal DNA che forma un gene. La mutazione avviene in un individuo ed è trasmissibile ai suoi discendenti, ma, siccome ogni individuo ha un numero limitato di discendenti immediati, devono passare molte generazioni prima che si diffonda a un numero di individui sufficiente perché sia facilmente osservabile in una popolazione. La trasmissione dei mutamenti sociali e culturali (come per esempio il cambiamento di dieta) avviene ovviamente secondo altri meccanismi; questa trasmissione non riguarda esclusivamente i figli dei portatori del mutamento, come avviene invece nelle mutazioni biologiche, e i mutamenti possono diffondersi rapidamente in una popolazione come un'epidemia. I 'documenti' genetici possono quindi fornirci - è questo il loro pregio - informazioni su eventi più antichi, di cui non esiste memoria storica.
Le frequenze geniche variano nello spazio e nel tempo. Vedremo che per un meccanismo di natura statistica, noto col nome di 'deriva genetica', in una popolazione reale le proporzioni dei suoi geni variano nel tempo, in altre parole la popolazione 'evolve' dal punto di vista biologico in modo tanto più significativo quanto più piccolo è il numero di individui che la compongono. Poiché le popolazioni si distribuiscono in gruppi più o meno isolati tra loro (l'isolamento dipende dalla distanza) e ognuno di questi gruppi evolverà in modo più o meno indipendente, dalla differenziazione nel tempo di ognuno di essi discenderà anche una differenziazione nello spazio: storia e geografia dei geni sono strettamente correlate.Si è già parlato della mutazione: essa costituisce il meccanismo fondamentale del cambiamento genetico generando nuovi alleli e, nel caso in cui venga duplicato per accidente un intero gene, anche nuovi loci. Senza mutazioni non vi sarebbe evoluzione. Si tratta, tuttavia, di un evento molto raro, che nell'uomo, a seconda dei geni, capita in una su 10⁴-10⁶ cellule germinali (spermatozoi e uova) per gene per generazione, il che significa che la sua possibilità di diffondersi in una popolazione è minima, se non vi sono altri meccanismi in grado di aumentarne la frequenza.
Oltre alla mutazione, i meccanismi evolutivi che possono cambiare le frequenze alleliche sono la migrazione, la selezione naturale e la deriva genetica casuale.
La migrazione ha un effetto analogo a quello della mutazione, in quanto i nuovi individui immigrati possono introdurre geni nuovi, o comunque alterare la distribuzione di frequenze degli alleli nella popolazione che li riceve. Gli alleli appena introdotti potranno anche essere trasmessi alle generazioni successive. In questo senso la migrazione tende a diminuire le differenze genetiche tra popolazioni: se due o più popolazioni hanno frequenze geniche diverse, lo scambio migratorio reciproco tende a rendere le loro frequenze geniche più simili e, se continua per molte generazioni, tende a renderle uguali. L'effetto genetico della migrazione dipende allora in modo critico dal rapporto numerico tra le due popolazioni, un dato demografico che purtroppo si può ricavare raramente e indirettamente dalle fonti archeologiche e storiche, e solo per ordini di grandezza. Conviene comunque distinguere qualitativamente due casi. Il primo si verifica quando lo spostamento riguarda individui, famiglie o piccole comunità, avviene per lo più su piccole distanze e causa una continua mescolanza all'interno delle popolazioni. Il secondo riguarda invece spostamenti di grandi masse di popolazione, che avvengono per lo più su grandi distanze, con episodi di espansione e successivi insediamenti su terre spesso disabitate. È stato questo secondo tipo di migrazione umana, che si può più propriamente definire 'colonizzazione', il maggiore responsabile dei grandi cambiamenti genetici che hanno caratterizzato popoli interi o addirittura continenti.
La selezione naturale è il meccanismo attraverso il quale i fenotipi aumentano o diminuiscono la loro frequenza a seconda della maggiore o minore capacità di adattamento all'ambiente naturale degli individui che li portano. Fu intuizione rivoluzionaria di Darwin aver posto a fondamento dell'evoluzione delle specie il meccanismo dell'adattamento biologico per selezione naturale. In senso tecnico maggiore o minore adattamento (fitness darwiniana) significa capacità di generare figli che possano raggiungere l'età fertile in numero maggiore o minore rispetto alla media dei figli degli individui che vivono nello stesso ambiente. Si tratta di un processo per tentativi ed errori, di cui si può prevedere l'andamento sulla base esclusiva di variabili demografiche. L'ambiente seleziona i tipi più 'adatti', quelli cioè che rivelano un'accresciuta fertilità o una probabilità maggiore di sopravvivenza all'età fertile. In questo modo aumenterà la frequenza degli alleli che determinano tipi più 'adatti' all'ambiente particolare, e il processo di selezione naturale continuerà una generazione dopo l'altra fino a quando altre condizioni ambientali selezioneranno altri tipi genetici.
Per meglio illustrare questo meccanismo evolutivo conviene fare un esempio. L'incapacità di digerire il latte fresco costituisce un disturbo abbastanza comune nelle popolazioni europee: è di natura ereditaria e dipende dalla mancanza di un enzima, la lattasi, che permette di metabolizzare il lattosio, uno zucchero contenuto nel latte fresco. Comunemente la lattasi è presente nel neonato fino allo svezzamento, ma nell'individuo adulto se ne trovano dosi minime: la tolleranza al lattosio (il gene prende il nome di 'persistenza della lattasi') ha tuttavia una frequenza tra il 50 e il 100% nelle popolazioni europee che usano il latte fresco nella loro alimentazione. Si suppone che la mutazione che permette alla lattasi di persistere nell'individuo adulto abbia avuto modo di affermarsi e diffondersi con l'introduzione in Europa dell'agricoltura, quando l'addomesticamento di bovini e ovini ha reso disponibile, e forse in certe condizioni necessario per qualche popolazione, l'uso del latte fresco quale risorsa alimentare. In determinate condizioni ambientali gli individui tolleranti al lattosio sono stati avvantaggiati rispetto a quelli intolleranti, e il gene della persistenza della lattasi si è così diffuso fino a raggiungere le frequenze che si osservano oggi. Si noti che tale gene è inutile in popolazioni la cui dieta è priva di latte fresco: in effetti, in tali casi, la sua frequenza è pressoché nulla. Se è lecito datare l'origine della mutazione della persistenza della lattasi all'origine dell'agricoltura nel Neolitico, 10.000 anni fa, è possibile avere un'idea quantitativa del vantaggio biologico, o coefficiente di fitness, che devono avere gli individui con quel fenotipo rispetto a coloro che ne sono privi: è dell'ordine dell'1%, ovvero, se si paragona un gruppo di genitori senza quel fenotipo con un gruppo di ugual numero di genitori con quel fenotipo, e il primo ha generato 100 figli, il secondo avrà dato origine a 101 figli. Questa differenza, che sembra piccola, accumulata di generazione in generazione, può modificare la struttura genetica di una intera popolazione; la selezione naturale è in effetti un meccanismo evolutivo che, a differenza della mutazione, è in grado di trasformare la nostra specie in tempi relativamente brevi, entro l'arco d'esistenza di Homo sapiens. Poiché la selezione è specifica rispetto a un determinato ambiente, non solo l'evoluzione temporale, ma anche la distribuzione geografica delle frequenze geniche ne verrà influenzata.
Esistono molte forme di selezione naturale. Alla mutazione della persistenza della lattasi si associa l'espressione 'selettivamente vantaggiosa', per far notare che l'allele che conferisce vantaggio a chi lo porta in un determinato ambiente tende ad aumentare la sua frequenza nel tempo (in quell'ambiente) fino a raggiungere il 100%. Analogamente, esistono mutazioni selettivamente svantaggiose (chiamate anche 'deleterie'). Tutte le patologie genetiche sono, in genere, selettivamente svantaggiose, anche se dal punto di vista della genetica il vantaggio o lo svantaggio si misura dalla possibilità o meno di avere figli che raggiungono l'età fertile, per cui le patologie che insorgono dopo tale età hanno poca o nulla rilevanza.
Di molti geni non si conosce il ruolo selettivo. Molti geni sono selettivamente neutri, nel senso che, almeno per quel che ne sappiamo, non conferiscono a chi li porta né vantaggio né svantaggio. Ovviamente un gene selettivamente neutro in un dato ambiente e in un certo periodo di tempo può diventare vantaggioso o svantaggioso in un altro ambiente e in un altro periodo di tempo e viceversa. Tale relatività storica e geografica rende la selezione naturale un meccanismo estremamente elusivo e poco misurabile: può aumentare, ma anche diminuire, la variabilità genetica intra- o tra popolazioni, e l'unico modo per caratterizzarlo rispetto agli altri meccanismi consiste nel fatto che è gene-specifico, modifica cioè la frequenza dei vari geni in un modo differenziato che dipende dal contesto storico (evoluzione), geografico (ambiente) e fenotipico degli individui sui quali agisce.
La deriva genetica, o deriva genetica casuale, è un meccanismo evolutivo che dipende dalle dimensioni finite delle popolazioni e consiste in una fluttuazione delle frequenze geniche da una generazione all'altra, fluttuazione che è tanto più ampia quanto più piccole sono le dimensioni della popolazione. Si considerino in una popolazione due alleli allo stesso locus, A e B, con pari frequenza del 50%, e si immagini per semplicità che ogni individuo porti un solo gene di quel locus invece di due (uno di origine paterna e uno di origine materna). Ora, per rendere l'esempio più chiaro anche se poco realistico, si immagini che in quella popolazione i possibili genitori abbiano potuto generare solo tre figli. Vi sono quattro possibilità: 1) tutti e tre i figli sono A; 2) tutti e tre i figli sono B; 3) 2 figli sono A e uno è B; 4) 1 figlio è A e 2 figli sono B. Le ultime due sono più probabili perché costituiscono le combinazioni più vicine alla distribuzione della popolazione dei genitori di partenza. Tuttavia le frequenze di A e B cambiano dalla generazione dei genitori a quella dei figli, perché da una distribuzione di A e B del 50% nella popolazione dei genitori, nel campione dei figli la distribuzione di A e B può essere 2/3 e 1/3 o viceversa. In entrambi i casi le frequenze cambiano da una generazione all'altra, e se invece di 3 figli ce ne fosse solo 1 (riduzione della popolazione), questo potrebbe essere solo o A o B e il cambiamento sarebbe ancora più drastico: la frequenza dell'allele A sarebbe cresciuta dal 50 al 100% o diminuita allo 0% (e viceversa per B).
Un effetto di queste oscillazioni delle frequenze geniche è quello di eliminare (alla lunga) tutti gli alleli di una popolazione tranne uno. Infatti, se un allele raggiunge per caso la frequenza dello 0%, da quel momento in poi è eliminato e solo una mutazione (o la migrazione da un'altra popolazione) può nuovamente reintrodurlo nella popolazione. L'effetto della deriva genetica è tanto più rilevante quanto più piccole sono le dimensioni della popolazione considerata. È lecito allora supporre che questo meccanismo evolutivo abbia avuto molta importanza nei primi stadi dell'evoluzione umana, quando la distribuzione della nostra specie, sia in Europa sia negli altri continenti, era frammentata in molti gruppi di dimensioni ridotte. Se nelle grandi città di oggi l'effetto della deriva genetica deve essere minimo, negli agglomerati con pochi abitanti può risultare ancora notevole. I comuni di montagna ne costituiscono un esempio. È capitato a molti di notare il numero estremamente limitato dei cognomi delle famiglie che vi abitano. Pur non essendo ovviamente il prodotto di un gene, il cognome ne segue le regole di trasmissione lungo la linea maschile, nel senso che viene trasmesso di padre in figlio esattamente come un gene. Quando le dimensioni della popolazione sono piccole o si riducono per cause di forza maggiore (guerre, migrazioni, malattie, ecc.) il numero dei cognomi, che si possono formalmente paragonare a degli alleli, si riduce per deriva. Da una generazione all'altra alcuni cognomi scompaiono e da quel momento, anche se la popolazione tende nuovamente a crescere, il numero dei cognomi rimane quello della generazione precedente.
L'analogia coi cognomi ci aiuta a comprendere l'effetto della deriva genetica sulla variabilità entro e tra popolazioni, che è esattamente contrario a quello della migrazione. La deriva genetica, infatti, tende a far diminuire la variabilità all'interno di una popolazione, in quanto il numero dei cognomi (=alleli) diminuisce e gli individui tendono ad avere lo stesso cognome (=allele) e quindi a essere più simili tra loro, ma d'altra parte fa aumentare la variabilità tra le popolazioni, poiché, se il numero dei cognomi (=alleli) diminuisce all'interno di due popolazioni e tra esse non vi è contatto, è più probabile che i cognomi (=alleli) siano diversi nelle due popolazioni, in quanto ognuna si è evoluta in modo indipendente dall'altra e in ognuna alcuni dei cognomi (=alleli) si sono persi in modo casuale.
I quattro meccanismi che si sono descritti per sommi capi interagiscono nella loro evoluzione temporale e il risultato del loro effetto congiunto si traduce nella distribuzione geografica delle frequenze dei vari geni che osserviamo oggi nelle diverse popolazioni. Per dare un quadro sintetico di tale variabilità, è opportuno fare riferimento a un modello evolutivo, anche se logicamente tale modello deriva dall'analisi dei dati anziché costituirne una premessa. A grandi linee potrebbe essere il seguente.
1. I dati archeologici testimoniano che i primi insediamenti della nostra specie hanno coinvolto piccoli gruppi distribuiti su larghe superfici con pochi scambi tra loro. Per esempio, nel periodo tra l'ingresso di Homo sapiens sapiens in Europa (circa 40.000 anni fa) e l'inizio del Neolitico (10.000 anni fa), la deriva genetica ha avuto modo di produrre in Europa le maggiori differenziazioni genetiche.
2. In questo panorama di estrema differenziazione delle frequenze geniche hanno avuto luogo, in momenti critici della storia generalmente associati a innovazioni tecnologiche di grande rilevanza per la produzione del cibo o per la conquista territoriale, movimenti di grandi masse di popolazione, colonizzazioni, nuovi insediamenti, ecc. L'effetto di tali migrazioni, di segno contrario a quello della deriva genetica, è quello di smussare le differenze genetiche e creare gradienti continui di cambiamento. Questi gradienti, se le popolazioni non fossero soggette a nuove migrazioni, sarebbero destinati a rimanere stabili nel tempo, tanto che ancor oggi potremmo osservarne le tracce.
3. L'effetto congiunto di deriva genetica e migrazione, la prima inducendo variabilità genetica, la seconda rimuovendola, tende dunque a creare una situazione di relativa stabilità nei periodi in cui non avvengono nuovi movimenti migratori su larga scala. Intervenendo su tale 'scenario' genetico, la selezione naturale può modificarlo, ma purtroppo non sappiamo in quale direzione ciò avvenga. È lecito supporre, tuttavia, che molti geni non ne rimangano influenzati, essendo selettivamente neutrali, e che i pochi che ne potrebbero subire gli effetti li subiscano in modo diverso l'uno dall'altro. Poiché il problema più delicato nell'interpretare la variabilità geografica delle frequenze geniche consiste proprio nel verificare la validità di quest'ultima ipotesi, si è cercato di separare l'effetto congiunto di deriva genetica e migrazione da quello della selezione naturale, elaborando metodi di analisi in grado di 'filtrare' una singola pressione evolutiva, e di conservare per quanto possibile l'informazione globale fornita da tutti i geni esaminati.
Variabilità genetica e cambiamento socioculturale
La variabilità comune a molti geni è quella verosimilmente legata all'effetto congiunto di deriva genetica e migrazione piuttosto che a pressioni selettive, e dovrebbe pertanto essere quella che più direttamente può darci informazioni sulla nostra storia di migrazioni e insediamenti passati. La tecnica di elaborazione statistica adottata a questo scopo prende il nome di analisi delle componenti principali.
L'idea centrale è di operare una trasformazione delle variabili originali nel numero minimo di variabili possibili pur conservando la variabilità dei dati originali. Nella nostra analisi dei dati genetici dell'Europa sono state sufficienti 10 nuove variabili (chiamate componenti principali) per riprodurre il 90% della variabilità di 95 frequenze geniche. Ognuna di queste componenti si può considerare come una qualsiasi variabile continua e in quanto tale può essere rappresentata graficamente su una carta geografica. A tale tipo di carta geografica si è dato il nome di mappa sintetica.
Da un punto di vista pratico solo le prime componenti principali spiegano una proporzione sufficientemente informativa della variabilità dei dati originali. La prima componente spiega la quantità più alta di variazione, le successive spiegano proporzioni di variabilità progressivamente più piccole, che al di sotto di una certa soglia riproducono fluttuazioni statistiche casuali.
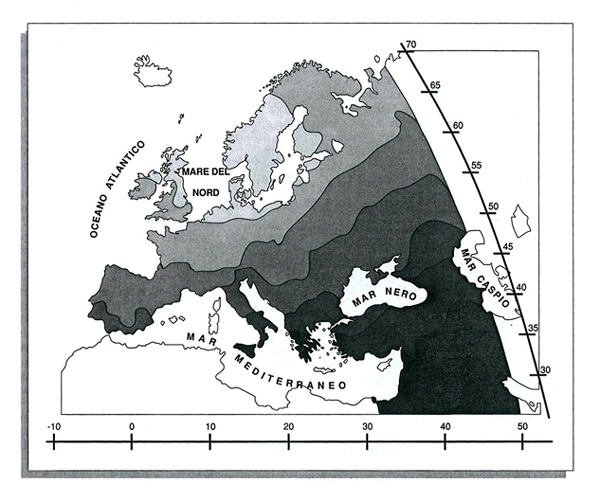
La mappa sintetica dei valori della prima componente principale, calcolata dalle frequenze osservate in Europa di 95 geni, è riprodotta nella fig. 1. La rappresentazione sintetizza il 28,1% della variabilità delle frequenze geniche originali. In realtà è un fatto piuttosto straordinario che con una variabile sola si possa riassumere il 28% della variabilità contenuta in 95 variabili indipendenti. La spiegazione consiste semplicemente nel fatto che vi è stato un avvenimento storico importante che ha determinato il cambiamento di queste variabili in modo parallelo.
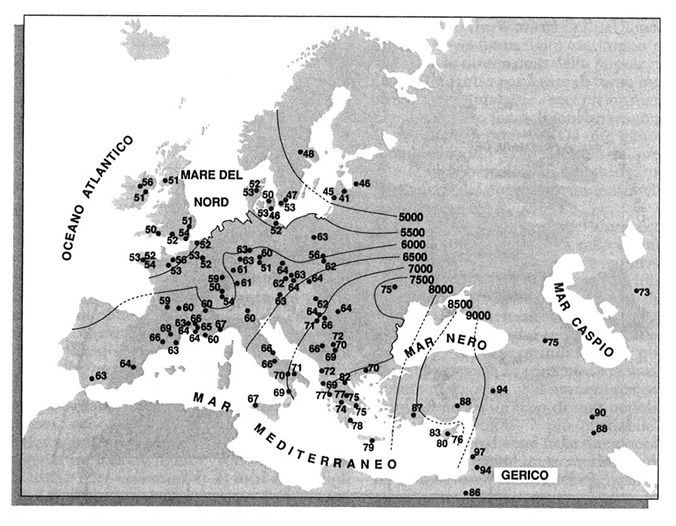
La mappa della fig. 1 dimostra un gradiente molto chiaro con un estremo nel Medio Oriente e l'altro a Nord Est, in corrispondenza di Irlanda, Gran Bretagna settentrionale e Scandinavia. Una mappa molto simile, ma meno precisa perché ricostruita su un numero minore di frequenze geniche (38), è stata pubblicata già nel 1978 in un lavoro (v. Menozzi e altri, 1978) in cui si notava la corrispondenza tra il gradiente che si osserva nella fig. 1 e la distribuzione delle datazioni archeologiche che indicano la coltivazione dei cereali introdotta dagli agricoltori neolitici in Medio Oriente e poi diffusa, nell'arco di 4.000 anni, in tutta l'Europa. La mappa di tali datazioni è riportata nella fig. 2.
La correlazione tra le due mappe è molto elevata (circa dell'80%), e non è certamente casuale. È stato simulato il processo di transizione da un'economia basata su caccia e raccolta del cibo a un'economia agricola con espansione degli agricoltori dal Medio Oriente verso l'Europa nordoccidentale, e si è trovata una precisa corrispondenza tra la distribuzione delle componenti principali dei dati simulati e la distribuzione di quelle delle frequenze geniche reali (v. Rendine e altri, 1986).
La mappa sintetica della fig. 1 è di estremo interesse per la struttura genetica delle popolazioni europee. Dimostra infatti che: 1) la diffusione dell'agricoltura in Europa nel Neolitico non fu esclusivamente una diffusione delle tecniche agricole senza spostamento di individui, ma fu anche spostamento degli agricoltori, probabilmente spinti da un'esplosione demografica indotta dalle stesse innovazioni tecnologiche; 2) il processo, iniziatosi 10.000 anni fa, nell'arco di 4.000 anni ha sostituito i cacciatori-raccoglitori con gli agricoltori (i nuovi insediamenti che ne sono conseguiti sono stati così rilevanti, per la struttura delle popolazioni europee, che ancor oggi possiamo identificarne le tracce genetiche; la diffusione della nuova tecnologia ha costituito, per così dire, la 'struttura portante' delle differenze genetiche osservate in Europa).
L'introduzione dell'agricoltura ha quindi innescato una rivoluzione culturale che ha coinvolto usi e costumi, regimi economici e lente espansioni demografiche il cui segno è ancor oggi riconoscibile. La mappa che sintetizza la distribuzione di tutti i geni conosciuti oggi nel mondo, anche se di lettura più articolata, non è in disaccordo con l'idea che i luoghi dove hanno avuto origine nuove tecnologie di produzione del cibo sono anche quelli da cui si sono diffuse migrazioni di individui, oltre che di idee innovative.
Variabilità genetica e cambiamento linguistico
Che lingua parlavano gli agricoltori neolitici mediorientali che si sono sparsi per l'Europa? L'archeologo Colin Renfrew (v., 1987) ha prospettato l'ipotesi che essi parlassero lingue indoeuropee e che le abbiano diffuse in Europa. Questa ipotesi è in accordo con quella prospettata dal linguista Dolgopolsky (v., 1988), il quale ritiene che le lingue indoeuropee abbiano avuto origine in Anatolia, ma è in disaccordo con l'opinione di molti altri linguisti. In realtà è molto difficile ricostruire una lingua parlata diecimila anni fa, nella totale assenza di documenti scritti. La scrittura ha poco più di cinquemila anni e la velocità di trasformazione delle lingue, essendo molto elevata, rende assai difficile ricostruire lingue molto antiche. Le ipotesi fatte finora sulla zona di origine delle lingue indoeuropee sono numerose e diverse (v. Mallory, 1989); l'ipotesi prospettata dall'archeologa Marija Gimbutas (v., 1991), che ha incontrato un certo successo, colloca tale zona in un'area della Russia meridionale sopra il Mar Nero e il Caucaso. A cominciare da circa cinquemila anni fa (all'inizio dell'età del bronzo), in quest'area si è sviluppata una civiltà di pastori nomadi che hanno addomesticato il cavallo, un animale diffuso nella steppa che si estende quasi ininterrotta dalla Romania alla Manciuria. La civiltà basata sullo sfruttamento del cavallo è stata denominata cultura kurgan, dal nome delle tombe a monticello molto comuni nella pianura meridionale russa. Il cavallo, inizialmente usato come fonte di cibo, è poi divenuto di grande importanza come animale da trasporto, e ha permesso una rivoluzione della tecnica militare (v. Diamond, 1991). A partire da quattromila anni fa i popoli del Medio Oriente e dell'Asia occidentale e meridionale, dove si erano saldamente affermati imperi e civiltà agricole molto importanti, si sono trovati esposti alle incursioni e alla conquista di pastori nomadi della Russia meridionale. Questi pastori parlavano lingue indoeuropee, e le hanno imposte in Iran, Afghanistan, Pakistan e India. È probabile, anche se non è accettato da tutti gli archeologi (in particolare dallo stesso Renfrew), che le migrazioni indoeuropee dei pastori della Russia meridionale si siano dirette anche verso occidente, verso la Grecia e l'Italia. È anche possibile che discendenti di questi pastori, rimasti per esempio in Ucraina e nella Russia meridionale fino all'epoca classica (come gli Sciti di cui parla a lungo Erodoto), abbiano dato origine ad altri popoli, successivamente protagonisti delle espansioni di cui oggi registriamo le tracce genetiche.
Oltre che dell'espansione degli agricoltori neolitici dall'Anatolia, che leggiamo nella rappresentazione sintetica della prima componente principale delle frequenze geniche (v. fig. 1), abbiamo prove genetiche di altre espansioni, anche se purtroppo l'analisi non ci permette di datarle. In particolare la mappa sintetica di un'altra componente genetica importante riproduce un'espansione a partire dalla zona dei kurgan, e confermerebbe quindi l'ipotesi della Gimbutas. Si deve però chiarire che non è necessario scegliere fra l'ipotesi di Renfrew e Dolgopolsky (i quali per ragioni profondamente diverse pongono le origini indoeuropee in Anatolia) e l'ipotesi della Gimbutas, perché possono essere vere entrambe, anche se riferite a tempi diversi. L'argomento di Renfrew - un popolo che si espande porta con sé la propria lingua - è storicamente ben documentato, specie se questo popolo si espande in una regione a bassa densità demografica. L'espansione dell'agricoltura ha favorito l'insediamento iniziale di gruppi neolitici anche nelle steppe euroasiatiche, da cui potrebbe essere derivato lo sviluppo della pastorizia e della civiltà dei kurgan. Con l'età del bronzo e con l'uso militare del cavallo sono potute avvenire altre espansioni di pastori nomadi, tre o quattromila anni dopo l'inizio dell'espansione dell'agricoltura. I nomadi parlavano lingue indoeuropee, certo diverse da quelle originali, dato il tempo intercorso fra le due espansioni, e le diffusero su una parte dei territori su cui si erano insediati i neolitici, sostituendo forse alcune delle lingue parlate in precedenza dagli agricoltori. Sia l'espansione neolitica sia quella dei pastori nomadi sono potute avvenire perché determinate da una serie di innovazioni importanti che hanno permesso l'aumento del cibo disponibile, e quindi sia la crescita demografica che la migrazione. L'espansione dei pastori nomadi ha coinvolto un numero minore di persone rispetto a quella neolitica, anche perché è avvenuta in regioni che avevano già un'alta densità di popolamento, ma ha avuto l'appoggio di un'organizzazione sociale e di una potenza militare relativamente avanzate.
Se fossimo in grado di disegnare in un grafico le tappe della nostra civiltà, tracceremmo una linea che illustrerebbe dapprima la capacità di disegnare, poi quella di rappresentare i nostri comportamenti in simboli astratti, per giungere infine all'elaborazione di un sistema di scrittura in grado di riprodurre tutte le sfumature della lingua parlata.
La transizione linguaggio-scrittura-cultura, pur nella sua semplificazione estrema, ci fornisce almeno un punto di riferimento per paragonare due diversi meccanismi evolutivi nell'uomo: quello biologico, mediato dai geni, e quello culturale, mediato dal linguaggio.
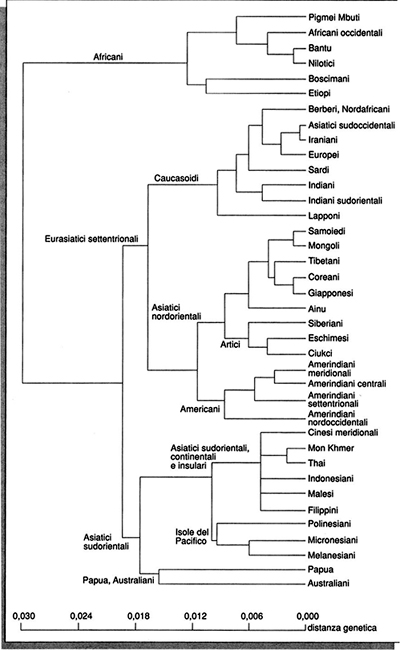
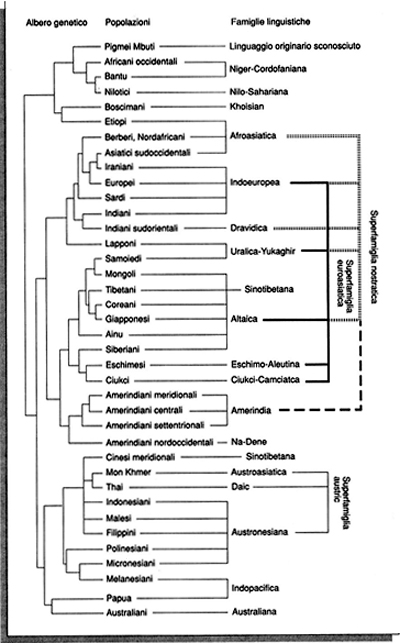
Viene naturale domandarsi: sono i geni a influenzare la cultura o è la cultura a influenzare i geni? La risposta a questa domanda è relativamente semplice: entrambi i fenomeni sono possibili, ma è quasi impossibile capire se uno dei due è più importante. Tuttavia, mentre è difficile trovare esempi di geni specifici che abbiano influenzato la cultura, abbiamo già trovato esempi di tratti culturali che hanno specificatamente influenzato la distribuzione dei geni nelle popolazioni umane. In una ricerca che ha suscitato molte discussioni (v. Cavalli-Sforza e altri, 1988) abbiamo tentato di ricostruire l'evoluzione dell'uomo integrando i dati genetici con quelli archeologici e linguistici. Un risultato che emerge chiaramente è la corrispondenza tra le grandi famiglie linguistiche in cui si suddividono le popolazioni umane e la loro evoluzione genetica. Tale corrispondenza è illustrata nelle figg. 3a e 3b. L'albero filogenetico della fig. 3a è in accordo con i dati archeologici che pongono l'origine di Homo sapiens sapiens in Africa. La corrispondenza quasi biunivoca tra la struttura dei raggruppamenti genetici e quella delle famiglie linguistiche (v. fig. 3b) ci ha indotto a ipotizzare che uno dei vantaggi adattativi di maggior rilievo che l'insediamento improvviso di Homo sapiens sapiens in tutte le parti abitate del nostro pianeta può aver prodotto sia stato proprio quello di un linguaggio completamente articolato.
La sostituzione repentina dell'uomo di Neanderthal in Europa da parte di una specie che era sulla scena della Terra da almeno 50.000 anni e la sua rapida estinzione non contraddicono l'ipotesi che il vantaggio della nostra specie sia consistito essenzialmente in un modo di comunicare più efficiente: esso avrebbe migliorato le tecniche di caccia e di approvvigionamento del cibo, favorito più stretti legami sociali, reso più semplice la diffusione di informazioni e di persone. A sostegno seppure indiretto di questa ipotesi si consideri che nella nostra società, fino a 100-150 anni fa, gli individui sordomuti avevano ben poche possibilità di riprodursi a causa della discriminazione sociale cui erano sottoposti.
Variabilità delle popolazioni umane: l'inconsistenza scientifica del concetto di razza
Quando si parla di variabilità genetica tra individui o tra popolazioni di individui occorre subito sgombrare il campo da pregiudizi. Se esaminiamo due individui a caso, non parenti tra loro, i loro corredi genetici saranno, con certezza quasi assoluta, diversi, perché diversi saranno i nucleotidi delle catene di DNA che li costituiscono: in altre parole, nel mondo biologico la diversità costituisce la regola, l'uniformità l'evento eccezionale. Di conseguenza il pregiudizio che ci porta a distinguere 'razze' diverse tra le popolazioni umane non ha alcun fondamento biologico, ma è di natura esclusivamente culturale. Il 'diverso' è classificato come tale in base a parametri che della biologia manifestano l'ignoranza più che la conoscenza: da un punto di vista genetico la persona di pelle scura è certamente diversa dalla persona di pelle chiara, ma i geni che controllano il colore della pelle (non uno solo, ma dell'ordine di qualche unità) costituiscono una percentuale irrisoria della totalità dei geni (dell'ordine di 100.000) che sono differenti in due persone con lo stesso colore della pelle. La persona di pelle scura viene 'vista' come diversa dalla persona di pelle chiara solo perché il colore della pelle è un carattere visibile, e in quanto tale viene culturalmente isolato rispetto ai caratteri la cui esistenza non è riconosciuta dai nostri sensi. Anche il colore degli occhi, la forma e le dimensioni del viso, il colore dei capelli, la statura, ecc., cioè quei caratteri umani - qualitativi e quantitativi - che possono essere 'misurati' (caratteri antropometrici), possono essere associati a differenze genetiche che hanno nella realtà un peso molto minore di quello che sembra. È il caso poi di aggiungere che l'apparenza inganna non una, ma due volte, in quanto le differenze tra caratteri antropometrici sono prevalentemente legate non all'evoluzione storica degli individui portatori di tali caratteri, ma piuttosto alle caratteristiche fisico-ambientali dei loro insediamenti. Infatti i caratteri antropometrici costituiscono per definizione l''interfaccia' tra l'uomo e l'ambiente in cui vive, e in quanto tali evolvono in funzione della geografia piuttosto che della storia di una popolazione. Pertanto il colore della pelle, la statura e molti altri caratteri non solo esprimono una minima parte del nostro corredo genetico, ma ci dicono anche poco della nostra evoluzione storica, essendo associati alle condizioni (soprattutto climatiche) dei luoghi in cui ci siamo insediati.
Le varie classificazioni delle popolazioni umane in razze, che dalla Bibbia a Erodoto e a Plinio il Vecchio, e, in tempi più moderni, da Linneo a Blumenbach, da Retzius a Boas, sono state il riflesso della nostra cultura classica più che della nostra conoscenza scientifica, si sono dimostrate un esercizio sterile per ragioni che già erano chiare a Darwin. In primo luogo le razze umane sono entità estremamente instabili nelle mani dei tassonomisti moderni: a seconda degli algoritmi di classificazione si possono distinguere da tre a più di sessanta cosiddette razze (v. Garn, 1971). Tale eterogeneità dipende in parte anche dalle diverse scuole tassonomiche, le cui tendenze a unire (lumpers) o a diversificare (splitters) le popolazioni non sono definite da criteri univoci. Nel settimo capitolo della sua opera The descent of man and selection in relation to sex, Charles Darwin scriveva: "È senz'altro indifferente indicare le razze dell'uomo in questo modo o raggrupparle in specie o sottospecie, anche se il termine sottospecie sembra più appropriato. Possiamo concludere che quando i principî dell'evoluzione saranno generalmente accettati, come ci aspettiamo avvenga presto, la diatriba tra monogenisti e poligenisti morrà di una morte silenziosa cui nessuno farà caso". Se questa profezia si è avverata solo in tempi molto recenti, grazie ai risultati che i dati archeologici e molecolari hanno fornito a sostegno della tesi di una origine unica della nostra specie in Africa, l'analisi dell'evoluzione delle popolazioni umane odierne (rappresentata nell'albero filogenetico riportato nella fig. 3a) distingue vari gruppi etnici, nella maggior parte famiglie o isolati linguistici; ma, osservando la figura da destra verso sinistra, andando cioè a ritroso nella differenziazione genetica, si incontrano raggruppamenti sempre più ampi: una loro classificazione dipende dal livello gerarchico che si considera, ed è quindi completamente arbitraria.
È stato dimostrato statisticamente che la variazione genetica all'interno dei vari raggruppamenti, di solito definiti secondo criteri geografici o linguistici, è dello stesso ordine di grandezza di quella tra i raggruppamenti (v. Lewontin, 1972; v. Nei e Roychoudhury, 1982). Tale osservazione deriva dal fatto che in quasi tutte le popolazioni sono presenti tutti i geni che si conoscono, anche se con frequenze differenti, e che tutte le popolazioni o gruppi di popolazioni hanno rapporti di affinità diversa a seconda del gene che si considera. Ne consegue che nessun gene singolo è sufficiente a classificare le popolazioni umane in categorie sistematiche. La variabilità che esiste in tutte le popolazioni, anche in quelle di piccole dimensioni, si è 'accumulata' in tempi molto lunghi, forse fin dall'origine della specie Homo sapiens, circa 700.000 anni fa: altrimenti non potremmo spiegare la presenza della maggior parte dei polimorfismi che conosciamo in quasi tutte le popolazioni. D'altra parte, la differenziazione geografica degli uomini anatomicamente moderni (Homo sapiens sapiens) è recente, risalendo a circa 100-150.000 anni fa (il 20% del tempo di esistenza della nostra specie).
Non vi è stato pertanto tempo sufficiente per una differenziazione tra i molti gruppi etnici insediatisi in aree geografiche diverse, maggiore di quanto non sia la differenziazione all'interno di ciascun gruppo. Inoltre la nostra specie, come quella dei nostri antenati più vicini (Homo erectus), ha sviluppato un'attività migratoria molto intensa in tutte le direzioni, con fenomeni di ibridazione tra popolazioni magari separate da lungo tempo. Le mescolanze diminuiscono le differenze genetiche, introducendo gradienti continui di variabilità che rendono ancora più difficile la definizione di 'confini' genetici.
Dal punto di vista scientifico il concetto di razza è pertanto privo di fondamenti. Si può obiettare che gli stereotipi razziali hanno una coerenza che permette anche ai profani di 'classificare' le persone. Tuttavia, come già si è sottolineato, gli stereotipi più diffusi (colore della pelle, colore e forma dei capelli, tratti del viso) riflettono differenze superficiali che le analisi genetiche più fini non confermano. Con i raffinati metodi statistici di oggi, e confrontando la storia genetica di molti geni e di molte popolazioni, siamo in grado di distinguere gruppi di popolazioni diverse e di ordinarle in una gerarchia. Tali gruppi, tuttavia, non possono identificarsi con le 'razze', perché ciascun livello di gerarchia separerebbe partizioni differenti e non vi è alcuna ragione biologica per preferirne una in particolare. Inoltre piccole variazioni nei geni, o nei metodi di analisi impiegati, possono spostare certe popolazioni da un gruppo all'altro. Pertanto la speranza di realizzare una tassonomia delle popolazioni umane attendibile e duratura è vana; si tratta tuttavia di una perdita di ben poco rilievo scientifico, che viene recuperata nella ricostruzione della storia evolutiva dell'uomo moderno così come viene messa in evidenza dall'analisi genetica. Tale ricostruzione ci appare argomento di ben maggiore interesse, sia perché ipotesi evolutive sulla nostra specie possono essere verificate con l'ausilio di altri dati (archeologici, linguistici, ecc.), sia perché, sul piano metodologico, può fornire gli strumenti per verificare su problemi e dati concreti la possibile interazione tra cambiamenti genetici e cambiamenti di origine culturale. Nella maggior parte dei casi, infatti, i momenti di maggiore espansione, anche demografica, della nostra storia sono stati segnati dalle grandi innovazioni culturali e quegli stessi momenti hanno anche determinato la struttura genetica attuale delle popolazioni umane.
Problemi sociali ed etici: il progetto Genoma umano
I tre miliardi di nucleotidi che compongono il DNA dell'uomo, distribuiti in tutti i cromosomi delle sue cellule, si articolano in un numero di geni conosciuto solo per ordine di grandezza - 10⁵ -, l'insieme dei quali prende il nome di 'genoma umano'. Il progetto Genoma umano è un'iniziativa di ricerca internazionale cui partecipano gli Stati Uniti, il Giappone e alcuni Stati della Comunità Europea: essa ha lo scopo di 'mappare' il genoma umano, cioè di localizzare sui cromosomi tutti i geni, e di ricostruirne la sequenza di DNA. Il costo di un'impresa del genere è stato stimato in circa un dollaro per nucleotide, distribuito nell'arco di 15 anni con una spesa annua, quindi, di circa 200 milioni di dollari. La possibilità di programmare una ricerca così impegnativa è maturata nella comunità scientifica verso il 1985, quando cioè i genetisti medici hanno incominciato a sfruttare le risorse della biologia molecolare per isolare alcuni geni causa di patologie. Apparve allora evidente che uno studio sistematico di tutti i geni umani, della loro localizzazione precisa sui cromosomi, della struttura molecolare dei loro prodotti avrebbe permesso non solo una capacità diagnostica di estrema raffinatezza e tempestività, ma anche, più generalmente, un avanzamento profondo delle conoscenze biologiche, e avrebbe consentito di dare alcune risposte a problemi medici ritenuti insolubili, quale la terapia di patologie genetiche mediante la sostituzione dei geni responsabili (terapia genica). Negli Stati Uniti il progetto incominciò a diventare operativo nel 1988, quando il Congresso americano raccomandò al National Institute of Health e al Department of Energy di ridistribuire una parte consistente dei loro fondi per finanziarlo.
Negli stessi Stati Uniti, ma forse ancor più negli Stati europei, il progetto ha però destato preoccupazioni, sia per problemi di strategia scientifica (il timore per esempio di far convergere su di esso la fetta più consistente dei finanziamenti a scapito di altri progetti), sia per le implicazioni sociali, etiche e legali di un progetto giudicato, per certi aspetti, tecnologicamente troppo avanzato rispetto al contesto sociale e politico della comunità cui è destinato. A ciò si aggiunga la complicazione della variabilità genomica: il genoma di un individuo è diverso da quello di un altro e, poiché a tutt'oggi (1992) non avrebbe senso identificare la sequenza intera del DNA di ognuno di noi, si tratta di capire entro quali limiti lo studio di un numero limitato di genomi sia generalizzabile. L'esperienza insegna, tuttavia, che auspicare una moratoria sulle conoscenze che comunque saranno accumulate attraverso il perseguimento anche parziale di questo progetto sarebbe un atteggiamento inutile se non addirittura dannoso: sembra più costruttivo prendere coscienza dei problemi e anticiparli responsabilmente piuttosto che doverli affrontare, impreparati, in un secondo momento.
Le ipotesi su cui fondare tale presa di coscienza si possono così indicare: 1) il progetto Genoma umano si realizzerà entro qualche decennio in quanto i progressi scientifici associati alle leggi di mercato in campo medico e farmaceutico porteranno quasi certamente alla 'mappatura' dei geni umani sui cromosomi e successivamente all'identificazione della loro sequenza di DNA; 2) la conoscenza dei propri geni diventerà un'acquisizione normale della nostra vita quotidiana e l'informazione genetica diventerà sempre più accessibile; 3) i servizi di genetica medica dovranno sempre più integrarsi agli altri servizi sanitari di diagnosi e cura, e l'accesso a essi sarà in qualche modo garantito. Quest'ultima ipotesi può essere meno realistica delle due precedenti, perché è direttamente collegata alle scelte sull'aborto selettivo e sulla manipolazione dell'embrione umano a fini di diagnosi e, in prospettiva, di terapia. Se, per esempio, la legislazione di uno Stato non permettesse in alcuna forma le ricerche che richiedono l'uso di embrioni umani, di feti o tessuti fetali, le applicazioni cliniche del progetto Genoma umano non potrebbero essere che limitate, così come i finanziamenti per sperimentarle e distribuirle sotto forma di servizi pubblici.
Le implicazioni sociali e politiche di questo progetto sono esemplari per le loro ramificazioni negli ambiti più diversi. Partendo dai problemi di politica e deontologia scientifica ci si può chiedere: 1) entro quali limiti sia giustificato destinarvi una parte non indifferente delle risorse finanziarie destinate alla ricerca biomedica (nel caso degli Stati Uniti, il 2-3% del bilancio del National Institute of Health viene usato per finanziare proposte di ricerca di questo progetto); 2) come possa essere assicurata la segretezza dei dati genetici raccolti (l'anonimato è spesso insufficiente, in quanto in questo genere di studi sono preferite le famiglie di grandi dimensioni, che nelle piccole comunità risultano facilmente riconoscibili); 3) in che misura sia lecito scambiare informazioni e materiale biologico tra gruppi di ricerca (in particolare: che ruolo assume chi fornisce il materiale biologico senza partecipare all'analisi del medesimo? nel caso in cui l'informazione genetica rivesta anche un interesse commerciale, chi ne è il proprietario? con quali strumenti se ne regola l'accesso?); 4) considerato il necessario impegno di elevate risorse finanziarie, con quali strumenti si possa garantire un coordinamento del progetto a livello internazionale al fine di evitare ridondanze e duplicazioni di sforzi.La ricerca sul genoma umano non può non suscitare anche problemi di politica sociale. In primo luogo è la nozione stessa di responsabilità che può cambiare i suoi confini: la determinazione genetica parziale o totale di un carattere, soprattutto se legato al comportamento (per esempio l'alcolismo), può rendere incerti i confini tra il patologico e l'illecito, con uno spettro di sfumature intermedie al quale gli interventi di protezione sociale e gli strumenti legali devono potersi adeguare.
Il problema centrale di una conoscenza sempre più estesa dei nostri geni sarà tuttavia quello della discriminazione genetica. L'origine di questo pericolo sta nell'intervallo di tempo sicuramente lungo tra la previsione o la diagnosi di una patologia genetica (o comunque associata a geni) e la possibilità pratica di adottare una terapia. Se ogni malattia diagnosticata in base alla conoscenza del genoma potesse essere immediatamente curata, il problema non sarebbe diverso da quello generale di una politica sanitaria in grado di valutare priorità, costi, benefici, efficacia, ecc. La specificità di una malattia causata o prevista dal nostro assetto genico, ma non curabile, può essere discriminante: a) per l'individuo sano cui venga prognosticata la malattia, nel caso in cui tale informazione sia accessibile a terzi, per esempio al datore di lavoro o alla compagnia assicuratrice; b) per l'individuo affetto dalla malattia, nel caso in cui l'accesso ai servizi sanitari e sociali venga differenziato a seconda delle possibilità terapeutiche; c) a causa della scarsa disponibilità dei servizi di consulenza genetica nell'ambito dei sistemi sanitari nazionali per l'alto costo di formazione del personale qualificato.
Un atteggiamento che storicamente ha sempre condotto alla discriminazione genetica nella sua forma deteriore, cioè al razzismo, è quello di chi ritiene che sia interesse della società indirizzare i suoi membri a conservare i geni ritenuti 'benefici' (eugenica positiva) o a eliminare i geni ritenuti 'dannosi' (eugenica negativa). Tale atteggiamento, che si esprime in vincoli del comportamento riproduttivo o nell'adozione dell'aborto selettivo, ha giustificazioni ben diverse se è il risultato di una scelta responsabile da parte di una coppia di coniugi o se invece riflette la tutela di un interesse pubblico, di un bene ritenuto comune da parte dello Stato. In questo secondo caso (purtroppo un peccato originale della comunità scientifica, che ha trovato fertile terreno di crescita nell'ideologia politica) si commette un errore grossolano: quello di supporre di poter cambiare in modo significativo la frequenza dei geni dannosi. In realtà, anche immaginando di poter sterilizzare tutti gli individui affetti da malattie genetiche, si può calcolare che il numero di generazioni necessarie a farle scomparire completamente sarebbe comunque assai alto, dell'ordine di secoli, se non di millenni. A prescindere da ogni considerazione etica, è molto difficile giustificare seriamente un provvedimento di politica sociale quando si prevede il suo effetto proiettato in tempi così lontani. La giustificazione addotta in sede dottrinale (lo Stato avrebbe il dovere di intervenire per proteggere quegli interessi che i singoli individui non sono in grado di proteggere) è molto discutibile in materia di scelta riproduttiva: si dovrebbe accettare che lo Stato possa, meglio degli stessi genitori, decidere quale genere di figli essi debbano avere. Una delega tanto rischiosa da mettere in pericolo i più elementari diritti di libertà. (V. anche Etologia; Evoluzione culturale; Razzismo).
Bibliografia
Ammerman, A. J., Cavalli-Sforza, L. L., Neolithic transition and the genetics of populations in Europe, Princeton, N.J., 1984 (tr. it.: La transizione neolitica e la genetica di popolazioni in Europa, Milano 1986).
Bankowsky, Z., Capron, A. M. (a cura di), Genetics, ethics and human values: human genome mapping, genetic screening and gene therapy, Geneva 1991.
Bodmer, W. F., Cavalli-sforza, L. L., Genetics, evolution and man, San Francisco 1976 (tr. it.: Genetica, evoluzione, uomo, 3 voll., Milano 1977).
Cavalli-Sforza, L. L., Menozzi, P., Piazza, A., The history and geography of human genes, Princeton, N.J., 1994.
Cavalli-Sforza, L. L., Piazza, A., Menozzi, P., Mountain, J., Reconstruction of human evolution: bringing together genetic, archaeological, and linguistic data, in "Proceedings of the National Academy of Sciences USA", 1988, LXXXV, pp. 6002-6006.
Cook-Degan, R. M., The human genome project: formation of federal policies in United States 1986-1990, Washington 1990.
Curtoni, E. S., Dallapiccola, B., De Marchi, M., Mattiuz, P., Momigliano, P., Piazza, A., Manuale di genetica, Torino 1991.
Darwin, C., On the origin of species by means of natural selection, London 1859, 1872⁶ (tr. it.: L'origine delle specie, Torino 1959).
Darwin, C., The descent of man and selection in relation to sex, 2 voll., London 1871 (tr. it.: L'origine dell'uomo, Roma 1983).
Diamond, J. M., The earliest horsemen, in "Nature", 1991, CCCL, pp. 275-276.
Dolgopolsky, A. B., The Indo-European homeland and lexical contacts of Proto-Indo-European with other languages, in "Mediterranean language review", 1988, III, pp. 7-31.
Dunn, L. C., A short history of genetics, New York 1965 (tr. it.: Breve storia della genetica, Milano 1978).
Garn, S. M., Human races, Springfield, Ill., 1971.
Gimbutas, M., The civilization of the goddess: the world of Old Europe, San Francisco 1991.
Jordan, E., The human genome project: where did it come from, where it is going, in "American journal of human genetics", 1992, LI, pp. 1-6.
Lerner, I. M., Heredity, evolution and society, San Francisco 1968 (tr. it.: Eredità, evoluzione, società, Milano 1972).
Lewontin, R. C., The apportionment of human diversity, in "Evolutionary biology", 1972, VI, pp. 381-398.
Lewontin, R. C., The genetic basis of evolutionary change, New York 1974.
Mallory, J. P., In search of the Indo-Europeans: language, archaeology and myth, London 1989.
Mange, A. P., Mange, E. J., Genetics: human aspects, Philadelphia 1980.
Mayr, E., The growth of biological thought, Cambridge, Mass., 1982 (tr. it.: Storia del pensiero biologico, Torino 1990).
Menozzi, P., Piazza, A., Cavalli-Sforza, L. L., Synthetic maps of human gene frequencies in Europe, in "Science", 1978, CCI, pp. 786-792.
Muller-Hill, B., Murderous science, New York 1988.
Nei, M., Roychoudhury, A. K., Genetic relationships and evolution of human races, in "Evolutionary biology", 1982, XIV, pp. 1-59.
Piazza, A., Evoluzione biologica e cultura, in "Scientia", 1988, CXXIII, pp. 237-248.
Polsinelli, M., Buiatti, M., Ottaviano, E., Ritossa, F., Genetica, Firenze 1989².
Rendine, S., Piazza, A., Cavalli-Sforza, L. L., Simulation and separation by principal components of multiple demic expansions, in "American naturalist", 1986, CXXVIII, pp. 681-709.
Renfrew, C., Archaeology and language: the puzzle of Indo-European origins, London 1987 (tr. it.: Archeologia e linguaggio, Roma-Bari 1989).
Ruhlen, M., A guide to the world's languages, Stanford, Cal., 1987.
Thompson, J. S., Thompson, M. W., Genetics in medicine, Philadelphia 1966 (tr. it.: La genetica in medicina, Torino 1982).
Vogel, F., Motulsky, A. G., Human genetics, New York 1982 (tr. it.: Genetica umana, Milano 1988).
Weatherall, D. J., The new genetics and clinical practice, Oxford 1982 (tr. it.: Genetica e pratica clinica, Bologna 1990).