
La stima degli intervalli temporali
La stima degli intervalli temporali
La stima degli intervalli temporali si riferisce alla capacità di un animale di valutare intervalli di tempo della durata di secondi o minuti a partire da un momento arbitrario. Una prerogativa di tale stima è che l'accuratezza della misurazione è proporzionale alla durata dell'intervallo misurato. II timer utilizzato è un orologio flessibile, simile a un cronometro, che può essere fatto partire, fermato e azzerato da particolari tipi di stimoli. Non è stato ancora dimostrato per quale ragione gli animali si servano di tale cronometro, ma è probabile che esso consenta di ottimizzare le strategie di ricerca del cibo e sia implicato nell'apprendimento associativo e in altri tipi di comportamento decisionale. Le basi neuronali della stima degli intervalli temporali non sono state ancora chiarite, ma gli studi di tipo farmacologico e l'esito di diversi tipi di lesioni suggeriscono il coinvolgimento di aree cerebrali e sistemi neurotrasmettitoriali differenti nei due stadi che ne fanno parte: la misurazione e la memoria. Poiché la capacità di stimare intervalli temporali è stata osservata in molti vertebrati, è probabile che si tratti di un'abilità evoluzionistica mente molto antica.
Gli orologi biologici e la stima degli intervalli temporali
Quella del tempo è una dimensione ubiquitaria, e molti eventi biologicamente significativi possiedono caratteristiche temporali prevedibili. Non è dunque sorprendente che molti animali abbiano evoluto 'orologi' che permettono loro di sfruttare efficientemente le regolarità temporali che incontrano nel mondo.
Per la maggior parte degli etologi gli orologi biologici sono le oscillazioni di origine endogena che sono alla base delle ciclicità o ritmi comportamentali correlati a periodicità ambientali, quali l'alternarsi del giorno e della notte (ritmi circadiani), il moto delle maree (ritmi circatidali), le fasi lunari (ritmi circa lunari) e l'alternarsi delle stagioni nel corso dell'anno (ritmi circannuali). In effetti, questi sono i soli tipi di orologio che la maggior parte dei moderni testi di comportamento animale e di etologia tratta esplicitamente (v. anche il saggio di G. Tosini e M. Menaker, Orologi biologici).
Tuttavia, gli psicologi hanno operato una distinzione tra due diversi tipi di senso del tempo negli animali: il senso periodico o della fase temporale e il senso dell'intervallo temporale (Gallistel, 1990). Il senso della fase temporale si basa sulle varie oscillazioni che abbiamo appena menzionato, e si riferisce alla capacità degli animali di prevedere gli eventi che avvengono in coincidenza con determinati punti fissi della fase di un determinato oscillatore. Per esempio, le api utilizzano il senso del tempo circadiano per poter fare visita a un determinato gruppo di fiori solamente nel momento della giornata in cui essi producono nettare, e le beccacce di mare (Haematopus ostralegus) si servono probabilmente del senso del tempo circatidale per volare a cercare cibo sulla riva fangosa solamente quando c'è bassa marea.
Mentre questi orologi di tipo fasico sono utili per prevedere gli eventi che avvengono in coincidenza con determinati punti fissi di una certa fase, essi sono meno adeguati per ottenere informazioni su altri tipi di regolarità temporale che gli animali possono incontrare. Il verificarsi di un evento biologicamente significativo può essere previsto tramite uno stimolo esterno non legato a nessuna particolare ciclicità piuttosto che in base allo stato di un orologio fasico. Il tipo di senso del tempo necessario per prevedere questo tipo di evento è appunto chiamato senso dell'intervallo temporale o, anche, senso del tempo a clessidra, poiché per prevedere l'evento significativo è necessario un cronometro che, a partire da un determinato stimolo, misura l'intervallo di tempo che separa quest'ultimo dall'evento significativo. Il colibrì (Phaethornif superciliosus) in cerca di cibo offre un ottimo esempio naturale di questo tipo di senso del tempo. Un fiore che l'uccello ha appena svuotato di tutto il suo nettare avrà bisogno di una certa quantità fissa di tempo per riempirsi di nuovo. Se quindi l'uccello conosce quanto dura questo intervallo, può tornare a visitare quel fiore esattamente nel momento in cui esso è tornato a riempirsi.
Oltre a permettere agli animali di prevedere quando un certo evento si verificherà, il senso dell'intervallo temporale li rende anche capaci di prendere decisioni in base alla valutazione e alla comparazione della durata degli intervalli temporali. Per esempio, quando va a caccia in un determinato campo, uno storno (Sturnus vulgaris) potrebbe misurare l'intervallo di tempo che intercorre tra le catture delle prede. Questi intervalli potrebbero essere utilizzati al fine di ottenere una stima del tasso di cattura delle prede, ovvero la stima di un parametro che è notoriamente di notevole rilevanza per la determinazione del comportamento decisionale degli storni in cerca di cibo.
Sebbene sia teoricamente possibile ricondurre sia il senso del tempo fasico sia quello dell'intervallo temporale al funzionamento di uno stesso orologio interno, diversi studi sembrano dimostrare che non è così e che essi funzionano invece in base a meccanismi differenti. Manterremo dunque, nel seguito dell'esposizione, la distinzione tra orologi fasici e orologi per intervalli. Il termine stima degli intervalli temporali è di solito riferito esclusivamente alla stima degli intervalli di tempo dell' ampiezza di secondi o minuti, ed è su questa gamma di intervalli che la presente trattazione è focalizzata. Non saranno dunque presi in considerazione i meccanismi più specifici che si sono evoluti al fine di rendere possibile la stima degli intervalli di tempo molto più brevi, come per esempio il sistema utilizzato dai pipistrelli per misurare il ritardo nel ritorno di un'eco (v. anche il saggio di I. Nelken, nel terzo volume). Mentre gli orologi di tipo fasico sono descritti nella maggior parte dei moderni testi di etologia e di comportamento animale, gli etologi hanno prestato poca attenzione alla stima degli intervalli temporali. Storicamente, lo studio di questo fenomeno è stato appannaggio degli psicologi interessati al condizionamento operante, ovvero di coloro che studiano la risposta dei ratti (Rattus rattus) e dei piccioni (Columba livia) ai programmi di rinforzo artificiali cui questi animali sono sottoposti in laboratorio. Di conseguenza, l'apparente mancanza di collegamento tra questo campo di ricerca e i problemi incontrati dagli animali nel loro ambiente naturale ha fatto sì che lo studio della stima degli intervalli temporali non riuscisse ad attirare l'attenzione degli etologi e degli ecologi del comportamento. Diviene tuttavia sempre più evidente che tale capacità è di vitale importanza per molti aspetti del comportamento naturale degli animali. Lo scopo della presente trattazione è dunque quello di esporre le sue proprietà e di discuterle da un punto di vista etologico.
Paradigmi sperimentali per lo studio del senso dell'intervallo temporale
P.R. Killeen (Killeen et al., 1997) ha dato la seguente definizione operativa di stima del tempo: "Un organismo sta utilizzando una stima del tempo se il nostro orologio ci permette di prevedere il suo comportamento meglio di quanto non farebbe qualunque altro stimolo identificabile". Non è facile evidenziare l'utilizzo della stima degli intervalli temporali negli animali che si trovano nel loro ambiente naturale, poiché per dimostrare che il loro comportamento è basato su questo tipo di stima è necessario escludere la possibilità che essi si stiano invece servendo di qualche stimolo esterno. Per questo motivo, le prove del fatto che gli animali possiedono un orologio che permette loro di misurare gli intervalli di tempo derivano per la maggior parte dalla letteratura psicologica sul condizionamento operante, nel quale i soggetti vengono studiati in condizioni di laboratorio strettamente controllate.
Gli psicologi hanno individuato diversi paradigmi sperimentali utilizzabili per lo studio della stima degli intervalli temporali negli animali; in linea di massima essi possono essere divisi in procedure di stima e procedure di riproduzione. In un compito di stima è richiesto all'animale di prestare attenzione a un determinato intervallo di tempo e di produrre, non appena questo sia terminato, una risposta basata sulla stima della sua durata; nel secondo tipo di compito il soggetto deve riprodurre realmente la durata dell'intervallo con il proprio comportamento e in tempo reale.
Un compito di stima temporale: la bisezione
Probabilmente il metodo di stima maggiormente utilizzato è quello noto come bisezione temporale. Nel primo stadio di questa procedura, il soggetto è addestrato a compiere una semplice discriminazione temporale, per esempio tra un suono presentato per 2 s e un identico suono presentato per 8 s. In ciascuna prova, viene presentato uno dei due suoni. Una volta terminato il suono, nella gabbia dell'animale vengono inserite due leve. Se il suono presentato è per esempio quello della durata di 2 s, l'animale viene rinforzato con del cibo affinché prema la leva di sinistra, mentre se il suono è quello di 8 s esso viene rinforzato affinché prema la leva di destra (la corrispondenza tra leve e suoni è arbitraria, ma è mantenuta costante per un determinato soggetto).
Animali come ratti e piccioni sono in grado di acquisire rapidamente l'abilità di compiere questo tipo di discriminazione temporale. Quando l'animale è giunto a portare a termine il compito con sufficiente attendibilità, vengono a esso presentati suoni di durate differenti da quelle cui era stato addestrato. In queste prove, nella gabbia vengono comunque inserite le due leve, ma nessun rinforzo segue la loro pressione a prescindere da quale delle due l'animale scelga. I dati ottenuti mediante il metodo della bisezione vengono di solito rappresentati in un grafico riportando in ascissa la durata del suono presentato nelle prove sperimentali e in ordinata la probabilità di scegliere la leva associata, per esempio al suono di 8 s. La funzione che ne deriva ha una forma sigmoide, e fornisce due importanti tipi di informazioni: la pendenza della curva indica la precisione della stima temporale, e la durata del suono in corrispondenza del quale si ha il 50% di probabilità che l'animale scelga la leva associata al suono di 8 s ci dice qual è la durata che viene soggettivamente stimata come intermedia tra 2 e 8 s.
La procedura di bisezione ha molti vantaggi. Diversamente dal metodo del picco, che descriveremo qui di seguito, essa può essere utilizzata per studiare una vasta gamma di intervalli temporali diversi, compresi quelli molto brevi. Un altro vantaggio consiste nel fatto che essa fornisce una stima del tempo percepito come intermedio, un parametro che molti modelli di stima temporale sono in grado di prevedere.
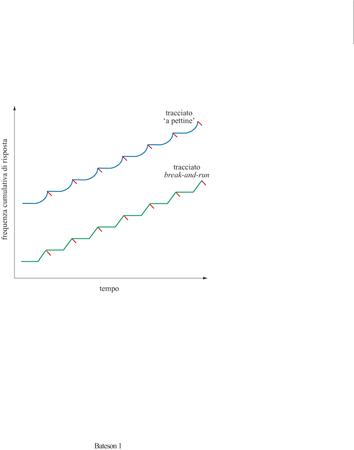
Un compito di riproduzione temporale: il metodo del picco Il metodo del picco consiste in un compito di riproduzione temporale che è una modificazione del più basilare programma di rinforzo a intervallo fisso (IF). In un programma IF, il soggetto viene rinforzato, ovvero riceve una ricompensa in cibo, alla prima risposta (beccare un tasto o premere una leva) che emette dopo che sia trascorso un intervallo di tempo di durata fissa. I programmi IF possono consistere o in prove di comportamento operante libero, in cui l'intervallo di tempo che trascorre tra le diverse ricompense alimentari costituisce l'unico stimolo discriminativo e viene azzerato ogni volta che l'animale riceve il cibo, oppure in compiti con procedura a prove discrete, a partire dai quali è stata sviluppata la procedura del picco. In una procedura a prove discrete, all'inizio dell'intervallo viene presentato un segnale, per esempio un suono o una luce, che prosegue fino a quando la risposta fornita dall' animale soddisfa i criteri stabiliti ed esso riceve il cibo. A ciò segue, prima che abbia inizio la prova successiva, un intervallo durante il quale lo stimolo non è presente. Gli animali potrebbero teoricamente rispondere con la massima frequenza nel momento in cui ricompare lo stimolo, ma in realtà essi non lo fanno e di solito mostrano un tracciato 'a pettine' (dall'inglese scallop) oppure un tracciato di risposta detto break-and-run ('darsi alla fuga'): quando compare lo stimolo si assiste a una pausa nelle risposte, che dura approssimativamente i primi due terzi dell'intervallo e che è seguita da una rapida accelerazione all'avvicinarsi del momento in cui alla risposta seguirà il rinforzo (fig. 1). La durata della pausa iniziale può essere analizzata per ottenere dati sulla stima temporale.
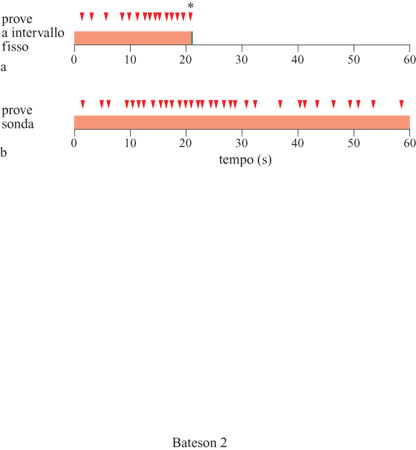
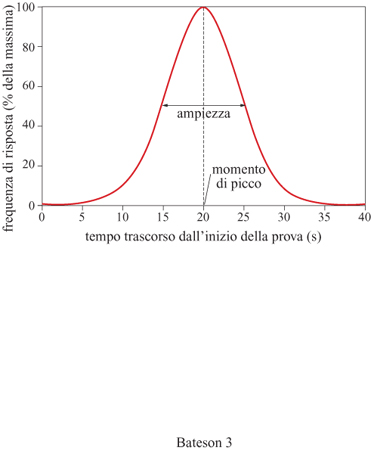
Nel metodo del picco il 50% delle prove è del tipo IF standard, ma il rimanente 50% è costituito da prove 'sonda' (probe trials) in cui lo stimolo persiste per una quantità di tempo che equivale al doppio o al triplo del normale intervallo delle prove IF. Le prove sonda sono separate dalle prove IF standard da un periodo di intervallo, e non viene fornito alcun rinforzo (fig. 2). Nella prima parte di queste prove sonda i soggetti si comportano come nelle prove IF standard (i due tipi di prove sono infatti, fino a questo punto, identici), finché, una volta trascorso l'intervallo di tempo stabilito come criterio, a un certo punto smettono di nuovo di rispondere. Quando i dati relativi a un certo numero di prove sonda vengono riportati in un grafico, riportando in ascissa il tempo trascorso dall'inizio della prova e in ordinata la frequenza media di risposta, si ottiene di solito una curva gaussiana con il suo tipico picco (fig. 3). Da essa si ricavano dati interessanti, tra i quali il momento di picco, ovvero il momento in cui si osserva la massima frequenza di risposta, la frequenza di picco, ovvero l'altezza della funzione in corrispondenza del tempo di picco, e infine l'ampiezza, misurata dall' ampiezza della curva in corrispondenza della metà della sua altezza massima. Il momento di picco viene di solito utilizzato come misura dell'accuratezza della stima temporale, mentre l'ampiezza ne indica la precisione. Una caratteristica particolare che rende il metodo del picco maggiormente vantaggioso rispetto a quello della bisezione consiste nel fatto che nel primo ciascuna prova sperimentale è in grado di fornire una stima di entrambe queste variabili, mentre nel secondo sono necessarie numerose prove sperimentali per ottenere una funzione di stima temporale da cui possano essere ricavati tali indici. Ciò rende il metodo del picco particolarmente adatto, per esempio, allo studio degli effetti transitori dei farmaci sulla stima temporale.
Un'importante variante del metodo del picco è la procedura del picco con sotto intervallo vuoto. Essa è identica al metodo del picco se non per il fatto che nel 50% delle prove sperimentali (scelte a caso) è inserito un sotto intervallo vuoto durante il quale lo stimolo non è presente. Analizzando, in questo tipo di prove, il punto in cui va a situarsi il picco una volta ricomparso lo stimolo, è possibile comprendere il modo in cui l'animale abbia trattato il sottointervallo vuoto.
Esistono tre possibilità. L'animale può ignorarlo, proseguendo con il proprio orologio come se niente fosse e mostrando così il picco nella risposta una volta trascorso il normale intervallo di criterio (cioè l'intervallo che l'animale deve imparare a discriminare) dall'inizio della prova. Oppure, l'animale può fermare il proprio orologio al sopraggiungere del sotto intervallo vuoto, memorizzare la durata dell'intervallo già trascorso, e al termine del sottointervallo far ripartire l'orologio. In questo caso, il picco risulterebbe spostato a destra di una quantità di tempo pari alla durata del sotto intervallo vuoto. La terza possibilità, infine, prevede che il soggetto azzeri il proprio orologio e lo faccia ripartire solamente al termine del sottointervallo. Se così fosse, il picco risulterebbe spostato a destra, nel grafico mostrato in figura 3, di una quantità di tempo pari alla durata dell'intervallo trascorso prima dell'inizio del sotto intervallo vuoto più la durata del sottointervallo stesso.
Esistono naturalmente molti altri metodi per analizzare la stima temporale, ma si può affermare che i paradigmi della bisezione e del picco sono quelli che hanno prodotto la maggior parte delle odierne conoscenze sulla natura della stima degli intervalli temporali.
Un metodo che va menzionato per i problemi connessi all'interpretazione dei dati che ne derivano è quello noto come rinforzo differenziale delle basse frequenze di risposta (DRL, Differential Reinforcement of Low rates of responding). In un programma di rinforzo DRL una risposta viene rinforzata se, e solo se, essa viene emessa dopo che sia trascorso un determinato intervallo fisso di tempo dall'emissione della risposta precedente. Il metodo DRL è simile a quello lF in quanto la risposta viene rinforzata solamente dopo che sia trascorso un intervallo scelto come criterio, ma ne differisce poiché le risposte anticipate rispetto al criterio penalizzano l'animale. l dati che vengono analizzati a partire da un programma DRL sono i tempi che intercorrono tra le risposte: in un soggetto la cui prestazione sia buona, questi tempi dovrebbero risultare in media leggermente più lunghi dell 'intervallo di criterio. Il problema del loro utilizzo come indici di abilità di stima temporale è che essi sono influenzati, oltre che da questa, anche dalla capacità che l'animale ha di inibire le proprie risposte. Mentre una buona prestazione in un programma DRL indica una buona capacità di stimare gli intervalli temporali, un cattivo risultato non costituisce necessariamente una prova del contrario.
Caratteristiche comportamentali dell'orologio per la stima degli intervalli temporali
l metodi appena descritti sono stati utilizzati dagli psicologi sperimentali per chiarire una serie di caratteristiche dell'orologio deputato alla stima degli intervalli temporali. Descriveremo ora alcune di queste caratteristiche e le confronteremo con quelle dell'orologio circadiano, che è l'orologio fasico maggiormente studiato. In termini generali, l'orologio per intervalli può essere paragonato a un cronometro (Church, 1978; Roberts e Church, 1978; Roberts, 1983). Esso può essere infatti azionato quando compare uno stimolo e azzerato dal rilascio di cibo o dall'intervallo tra le prove. l ratti rispondono al metodo del picco con sotto intervallo vuoto fermando il proprio orologio quando il segnale viene spento, memorizzando la durata dell'intervallo già trascorso e facendo ripartire l'orologio al riaccendersi del segnale. L'orologio per intervalli non si limita a prendere in considerazione esclusivamente gli stimoli percepibili mediante una particolare modalità sensoriale, e sia i ratti che i piccioni sono in grado di 'contare' il tempo trascorso sulla base di segnali tanto sonori quanto luminosi. È stata anche osservata una capacità definita come transJer (trasferimento) intermodale, in cui l'apprendimento di un intervallo temporale avvenuto mediante uno stimolo in una certa modalità sensoriale (per esempio, un suono) viene immediatamente trasferito a uno stimolo in una modalità differente (per esempio, un flash).
Una delle caratteristiche più importanti della stima degli intervalli temporali è la vasta gamma di durate che gli animali sono capaci di valutare (Bradshaw e Szabadi, 1997). Il metodo del picco ha prodotto funzioni di stima notevolmente accurate utilizzando sia intervalli di criterio brevi (fino a pochi secondi) sia intervalli lunghi (fino a un'ora e mezza). Al contrario, l'orologio circadiano può essere sincronizzato esclusivamente su una gamma limitata di periodi di tempo, che va dalle 19 alle 27 ore (Aschoff, 1980).
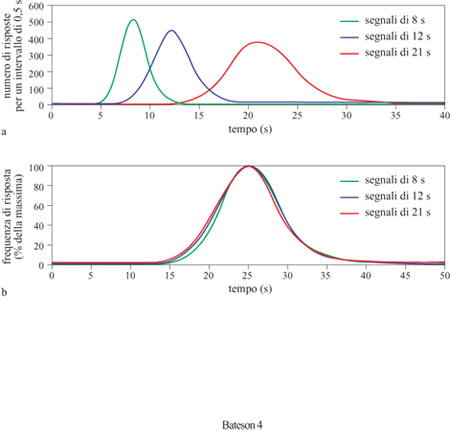
La precisione della stima degli intervalli temporali è proporzionale alla durata dell'intervallo da stimare, e questo vale per l'intera gamma di intervalli studiati, che copre 3 ÷ 4 ordini di grandezza (Gibbon et al., 1997). La larghezza della curva ottenuta mediante il metodo del picco aumenta dunque con l'aumentare dell 'intervallo di criterio; tuttavia, se la funzione viene normalizzata misurando il tempo rispetto al momento di picco e la frequenza rispetto alla frequenza massima di risposta, allora le diverse funzioni ottenute a partire da intervalli di criterio di diversa durata si sovrappongono (fig. 4). Questo fenomeno, che rappresenta una caratteristica distintiva della stima degli intervalli temporali, è stato chiamato proprietà scalare. Non è noto se anche gli orologi fasici godano o meno di questa proprietà scalare, poiché la loro precisione non è mai stata analizzata confrontando sincronizzazioni con periodi di tempo diversi. Di solito è possibile osservare che la precisione relativa della stima di tipo fasico è notevolmente maggiore di quella della stima degli intervalli temporali: la variabilità nell'inizio dell' attività giornaliera di un criceto (Mesocricetus auratus) può essere così bassa da rasentare l'1% della fase circadiana, mentre la variabilità nella stima degli intervalli temporali è usualmente di circa il 15÷35% dell'intervallo stimato. Dunque, la notevole flessibilità degli orologi per intervalli, nel senso della gamma di durate che essi possono coprire, sembra andare a scapito della loro precisione.
Funzioni adattative della stima degli intervalli temporali
Come già accennato, nonostante i numerosi studi condotti dagli psicologi sulla stima degli intervalli temporali, questo campo di ricerca è stato quasi completamente ignorato dagli ecologi del comportamento. Sappiamo dunque molto poco delle pressioni selettive che hanno condotto allo sviluppo di questa capacità e dell'uso che attualmente gli animali ne fanno. Sembra ovvio che possedere un orologio per misurare intervalli rappresenti un vantaggio, ma come se ne servono di fatto gli animali? il fatto che l'ottenimento di cibo azzeri facilmente il cronometro e gli intervalli delimitati da stimoli connessi al cibo siano prontamente stimati indica che la stima degli intervalli temporali è probabilmente importante per prendere decisioni riguardanti la ricerca di cibo. Ma questa capacità potrebbe essere coinvolta anche in molti altri tipi di comportamento decisionale, come la scelta del partner sessuale, varie situazioni di comportamento sociale e la navigazione.
In termini generali, l'orologio per intervalli fornisce agli animali due tipi di informazioni. Da un lato offre la possibilità di prevedere quando un determinato evento si verificherà, dall'altro permette di valutare la durata di un intervallo temporale trascorso.
Previsione dell'occorrenza di un evento significativo
La possibilità di prevedere quando un certo evento significativo si verificherà comporta potenzialmente numerosi vantaggi. Un animale che sappia quando si verificheranno gli eventi potrà organizzare il proprio tempo con maggiore efficienza e si potrà preparare adeguatamente a ciò che sta per avvenire. Nel caso dell'esempio del colibrì (P. superciliosus) precedentemente citato, un uccello che possa valutare la quantità di tempo necessaria a una particolare specie di fiore per tornare a riempirsi di nettare dopo esserne stato svuotato avrà un evidente vantaggio in termini di fitness rispetto a un conspecifico che non sia in grado di fado. Qualora tornasse a visitare il fiore troppo presto, cioè prima che questo si fosse di nuovo riempito completamente, l'uccello non ne ricaverebbe la quantità di nettare che avrebbe potuto ottenere aspettando ancora; d'altro canto, se tornasse al fiore troppo tardi, quando questo si fosse già riempito da un certo tempo, il colibrì rischierebbe di essere preceduto da un altro individuo con probabilità tanto maggiore quanto più cospicuo è il ritardo (Gill, 1988). Questi uccelli si trovano in una situazione analoga a quella del programma di rinforzo lF con comportamento operante libero, poiché l'unico stimolo discriminativo che hanno a disposizione è rappresentato dall'intervallo che intercorre tra le ricompense.
Per avere un esempio naturale che sia comparabile alla situazione prevista dal metodo del picco è possibile prendere in considerazione un problema che molti animali in cerca di cibo devono affrontare, ovvero quello di quando sia il caso di abbandonare un terreno di caccia per spostarsi in un altro. Se l'intervallo di tempo che intercorre tra le catture successive è relativamente costante, e se è possibile che la riserva di prede della zona si esaurisca improvvisamente, allora il predatore dovrebbe stimare quanto tempo è passato dall 'ultima cattura e abbandonare quella zona non appena si renda conto che è trascorsa una quantità di tempo superiore al normale intervallo tra le catture. Per esempio, per procacciarsi il cibo, i pigliamosche (Muscicapa striata) sono soliti fare incursioni partendo da una postazione fissa. In uno studio è stato osservato che se uno di questi uccelli non riesce a prendere nessun insetto per una quantità di tempo pari a 1,5 volte il normale intervallo tra le prede, esso abbandona la postazione per spostarsi altrove (Davies, 1977). Sembra dunque che i pigliamosche utilizzino la stima degli intervalli temporali per valutare la possibilità dell'esaurimento del terreno di caccia.
Questa situazione è stata simulata in laboratorio utilizzando storni (S. vulgaris) che ottenevano il cibo in base a un programma di rinforzo lF che poteva improvvisamente interrompersi (Brunner et al., 1992). Gli uccelli avevano in ogni momento la possibilità di cambiare postazione semplicemente volando da un posatoio all'altro. Analogamente ai pigliamosche, una volta trascorso l'intervallo di criterio in assenza di ricompensa, gli storni attendevano ancora per una percentuale fissa di questo intervallo prima di desistere e volare sull'altro posatoio. È interessante sottolineare che la quantità di tempo che trascorreva prima che questi uccelli decidessero di spostarsi non era di durata costante, ma era invece proporzionale alla durata dell'intervallo fisso. Se gli storni avessero atteso una quantità di tempo costante una volta stimato che l'intervallo di criterio era trascorso, con il crescere della durata di quest'ultimo essi avrebbero lasciato il posatoio troppo presto con sempre maggiore probabilità, a causa della minore precisione nella stima degli intervalli temporali più lunghi.
La capacità di stima temporale non è importante soltanto per il comportamento di ricerca del cibo, ma potrebbe avere un ruolo anche in molti altri tipi di comportamenti decisionali. La tortora dal collare (Streptopelia decaocto) offre un buon esempio di come la possibilità di prevedere un evento possa essere importante per l'esecuzione efficiente del comportamento di cooperazione. In questa specie il maschio e la femmina partecipano entrambi all'incubazione delle uova, e al primo tocca solitamente l'intervallo che ogni giorno va approssimativamente dalle 10:00 alle 17:00. Se un giorno il turno di cova del maschio inizia in ritardo, viene ritardata anche la sua fine; ciò suggerisce che l'uccello possa utilizzare un orologio per intervalli per calcolare la fine del suo turno di cova (Gibbon et al., 1984). In questo caso non è chiaro a che scopo il maschio cronometri il proprio turno anziché cogliere l'occasione per abbandonare il nido al ritorno della femmina. Non è neanche chiaro perché utilizzi un orologio per intervalli anziché il suo orologio circadiano, dato che la femmina sincronizza il proprio ritorno al nido sull'ora del giorno.
Vi sono spiccate similarità tra il metodo del picco e gli esperimenti di condizionamento classico. Ciò suggerisce che potrebbero esistere delle connessioni tra la stima degli intervalli temporali e l'apprendimento associativo. In entrambi i p aradigmi , uno stimolo arbitrario detto stimolo condizionato viene presentato anticipatamente rispetto al rinforzo, che rappresenta lo stimolo incondizionato; l'unica differenza tra un esperimento di condizionamento classico e un esperimento di stima temporale con intervallo fisso è che, in quest'ultimo, per ricevere la ricompensa il soggetto deve mettere in atto una risposta.
Tradizionalmente, i teorici dell'apprendimento hanno focalizzato la propria attenzione sull'associazione tra stimolo condizionato e stimolo incondizionato, ma una crescente mole di risultati sperimentali dimostra come la contiguità temporale tra i due stimoli sia altrettanto importante per determinare il verificarsi o meno dell'apprendimento e per specificare il tipo di risposte condizionate che viene messo in atto (Gallistel, 1994). Sembra che gli animali apprendano la durata degli intervalli che intercorrono tra stimolo condizionato e stimolo incondizionato e tra due stimoli incondizionati, e sembra anche che sia il rapporto tra le rappresentazioni mnestiche di queste due durate, piuttosto che una semplice associazione tra stimolo condizionato e incondizionato, a determinare il verificarsi o meno di una risposta condizionata (Gibbon et al., 1977). È anche evidente che la durata dell'intervallo tra stimolo condizionato e incondizionato è in grado di influenzare il tipo di risposta condizionata messa in atto. Questo ha certamente senso dal punto di vista funzionale, poiché il tipo di risposta condizionata maggiormente adeguata dipende appunto da quanto lo stimolo incondizionato è distante nel tempo (Timberlake et al., 1982). Per esempio, nel caso di uno stimolo condizionato che preannuncia la disponibilità di cibo, un comportamento di ricerca o di caccia potrebbe essere adeguato a un intervallo tra stimolo condizionato e incondizionato di una certa lunghezza, mentre per un intervallo più breve potrebbe essere più appropriata la messa in atto di comportamenti di manipolazione e consumazione.
Valutazione della durata degli intervalli e comportamento decisionale
La capacità di compiere una discriminazione tra intervalli temporali differenti è probabilmente importante per molti diversi tipi di comportamento decisionale. Per esempio, un animale in cerca di cibo potrebbe basare le proprie decisioni future in merito alle strategie di ricerca del cibo sulla quantità di tempo che ha trascorso cercando cibo in due zone differenti, oppure sulla quantità di tempo che ha impiegato per manipolare e ingerire due diversi tipi di preda. Facendo un altro esempio, una femmina che dovesse scegliere un partner sessuale all'interno di un lek (o arena nuziale, cioè il luogo dove si aggregano numerosi maschi allo scopo di attirare le femmine e ottenere il massimo numero di accoppiamenti) potrebbe basare la propria decisione sulla durata delle esibizioni che i diversi maschi le indirizzano. Per compiere ciascuno di questi paragoni è necessaria la capacità di valutare e comparare la durata dei diversi intervalli di tempo trascorsi.
Questo tipo di stima degli intervalli temporali potrebbe permettere agli animali di valutare molte altre variabili importanti. Per esempio, la letteratura sull'ottimizzazione del comportamento di ricerca del cibo (optimal foraging) ha evidenziato che il tasso di ingestione di cibo è spesso altamente correlato alla fitness, ed è stato dimostrato che molti animali sono in grado di prendere decisioni che rendono ottimale questo tasso (Kacelnik, 1984). Poiché il valore di questo tasso è calcolato in base al rapporto tra la quantità di cibo ottenuta e il tempo impiegato per ottenerla, per la stima del tasso di ingestione è probabilmente necessaria la stima degli intervalli temporali. Quest'ultima potrebbe anche essere utilizzata per valutare quantità di cibo diverse, poiché esiste spesso una forte correlazione tra la grandezza di una ricompensa alimentare e la quantità di tempo necessaria per manipolarla e consumarla. È dunque estremamente probabile che la stima degli intervalli temporali sia ampiamente coinvolta nel comportamento decisionale implicato nell'ottimizzazione del comportamento di ricerca del cibo. È anche possibile che essa serva a stimare la distanza coperta dagli animali nel corso dei loro spostamenti.
Infine, la stima degli intervalli temporali potrebbe avere un ruolo nell'apprendimento percettivo. Per esempio, nel corso del processo di imprinting, si forma nel pulcino domestico (Gallus gallus) una rappresentazione visiva della madre. Benché il pulcino veda la madre da molteplici prospettive e distanze diverse, esso combina queste immagini fisicamente differenti in un'unica rappresentazione, purché esse siano percepite a breve distanza l'una dall'altra. A dimostrazione di ciò, i pulcini che abbiano ricevuto un imprinting su due oggetti differenti imparano a discriminare tra i due con maggiore facilità qualora essi siano stati loro presentati, durante l'imprinting, a 5 o più minuti di distanza tra loro piuttosto che non a meno di 30 s (Chantrey, 1974). Tali risultati indicano che il senso del tempo potrebbe consentire agli animali di formare categorie.
Studi comparativi
L'approccio comparativo viene spesso utilizzato allo scopo di individuare il significato adattativo di un determinato comportamento. Nel caso della stima degli intervalli temporali, questo approccio prevede che vengano confrontate le capacità mostrate da specie differenti ma imparentate tra loro, cercando di stabilire una correlazione tra le differenze nelle capacità di stima e le differenze nell'ecologia delle diverse specie.
Il programma di rinforzo lF è stato utilizzato per studiare la stima degli intervalli temporali in diverse specie di vertebrati e almeno in un invertebrato (Richelle e Lejeune, 1980), dal ratto (Rattus rattus) allo scimpanzé (Pan troglodytes) alla cornacchia (Corvus corax), fino all'ape domestica (Apis mellifera) e molti altri ancora. È stato dimostrato che molte specie, con poche eccezioni, sono in grado di stimare gli intervalli di tempo all'interno di un programma lF. Non è chiaro se l'apparente mancanza di capacità di stima temporale in alcune altre specie sia dovuta a un reale deficit in questa capacità o a inadeguatezze nella metodologia sperimentale utilizzata per metterla in luce.
Tuttavia, i tentativi di trarre conclusioni generali da questi dati si sono rivelati notevolmente problematici. Infatti, i vari studi differiscono tra loro sotto molteplici aspetti, come il tipo di risposta osservata, la ricompensa utilizzata (per esempio, cibo o acqua), le privazioni cui sono stati precedentemente sottoposti gli animali e la gamma di intervalli temporali studiati. Ciò che sembra costituire una differenza tra le specie potrebbe dunque derivare, in realtà, da differenze metodologiche. Anche quando due o più specie diverse vengono comparate utilizzando procedure identiche, prestazioni diverse non dimostrano necessariamente capacità di stima temporale diverse, poiché potrebbero essere invece attribuibili a differenze tra le specie nelle abilità motorie nella motivazione.
Una delle soluzioni migliori a questo tipo di problema è quella di comparare soltanto specie strettamente imparentate, che si presume siano molto simili tra loro in tutto eccetto che nella capacità di stima temporale. Purtroppo gli studi rigorosi di questo tipo sono stati sorprendentemente pochi. Un'eccezione è costituita da un lavoro comparativo sulle capacità di stima temporale di colombi e cornacchie (Powell, 1974). Questi uccelli furono addestrati con un programma DRL, il cui intervallo di criterio veniva progressivamente allungato allo scopo di determinare fino a quale punto i soggetti avrebbero continuato a rispondere adeguatamente. l colombi si rivelarono incapaci di rispondere a intervalli che andassero oltre i 10 o 30 s, mentre le cornacchie furono in grado di stimare intervalli temporali fino a 120 s. Questo studio non è soggetto alla critica che viene di solito mossa alla procedura DRL, ovvero che i risultati potrebbero essere attribuibili a differenze tra le specie nella capacità di inibire la risposta piuttosto che alla loro capacità di stimare gli intervalli, perché entrambe le specie mostravano una buona prestazione su intervalli di tempo fino a 120 s quando la fine dell'intervallo era segnalata da un cambiamento di colore del tasto e veniva in tal modo a mancare la necessità di effettuare una stima temporale. Sembra dunque che le cornacchie siano capaci di stimare intervalli di tempo più lunghi rispetto ai colombi. Benché la conclusione di questo studio sembri valida, è tuttavia difficile, in mancanza di dati su altre specie, fare ipotesi sul possibile significato adattativo di queste differenze.
Modelli teorici e meccanismi funzionali
Una delle ragioni per cui la stima degli intervalli temporali non è stata studiata in modo più approfondito va forse rintracciata nel fatto che non esiste un organo palesemente deputato alla misurazione del tempo. Tuttavia sono stati elaborati molti modelli teorici al fine di spiegare come questa misurazione possa essere effettuata.
Descriveremo e confronteremo tre diversi modelli, da ciascuno dei quali derivano risultati che presentano un buon accordo con i dati ottenuti mediante il metodo del picco che abbiamo già discusso (Bradshaw e Szabadi, 1997). In ciascun modello è possibile identificare tre diversi stadi del processo di elaborazione dell'informazione temporale. In primo luogo, la misurazione del tempo richiede un orologio. In secondo luogo, ciascun modello implica una memoria in cui l'intervallo temporale possa essere rappresentato. Infine, in ciascun modello è presente uno stadio decisionale, nel quale la misurazione effettuata dall'orologio e la rappresentazione mnestica vengono utilizzate per la determinazione del comportamento.
La teoria scalare
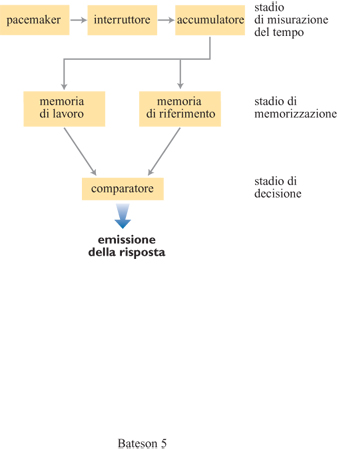
La teoria scalare della stima temporale (Gibbon e Allan, 1984) rappresenta un modello quantitativo di elaborazione dell'informazione che contiene moduli corrispondenti esplicitamente ai processi di misurazione (orologio), memorizzazione e decisione (fig. 5). L'orologio consiste in questo caso in una sorta di pacemaker che emette impulsi in modo continuo. Quando viene attivato l'interruttore, questi impulsi vengono convogliati in un accumulatore: in tal modo, la durata di un lasso di tempo appena trascorso costituisce una funzione lineare del numero di impulsi giunti all'accumulatore. Questo numero di impulsi passa poi alla memoria di lavoro (o a breve termine), e infine alla memoria di riferimento (o a lungo termine), nella quale si trova immagazzinata la distribuzione degli intervalli temporali precedentemente sperimentati dall'individuo. Dalla memoria di riferimento viene richiamata la durata di un intervallo estraendo un campione dalla distribuzione che vi è immagazzinata. In un programma IF la decisione di cominciare a rispondere, o smettere di farlo, viene presa confrontando continuamente il campione preso dalla memoria di riferimento con la durata dell'intervallo di tempo appena trascorso che si trova nella memoria di lavoro. Quando il rapporto tra il tempo trascorso e il valore in memoria è sufficientemente prossimo a uno, l'animale emette la risposta.
Tra i modelli di stima temporale esistenti, la teoria scalare costituisce quello più avanzato dal punto di vista formale; uno dei suoi grandi punti di forza sta nel fatto che se ne possono ricavare predizioni quantitative esplicite in merito ai risultati di molte diverse procedure di stima.
La teoria comportamentale
Come suggerito dal suo stesso nome, la teoria comportamentale della stima temporale (Bradshaw e Szabadi, 1997) costituisce un modello basato sul comportamento osservabile. Come la teoria scalare, anche quella comportamentale prevede la presenza di un pacemaker che emette impulsi. Secondo questo modello, ogni impulso è associato a uno stato comportamentale specifico, come il muoversi all'interno della gabbia, il premere la leva o il mangiare. In tal modo, all'emissione di ciascun impulso da parte del pacemaker, l'animale passa allo stato comportamentale a esso associato. La velocità del pacemaker è controllata principalmente dalla frequenza di rinforzo. Ci si può dunque attendere che un animale sottoposto a un programma IF completi la sequenza di stati comportamentali con maggiore rapidità se il criterio è di 10 s piuttosto che di 30 s, poiché nel primo caso la frequenza con cui l'animale riceve il rinforzo è maggiore. La ricompensa che viene offerta rinforza la sequenza comportamentale che l'ha preceduta, e tale sequenza viene immagazzinata in memoria: la rappresentazione di un intervallo di tempo corrisponde dunque a una sequenza di stati comportamentali. Quando l'animale è posto nuovamente di fronte a stimoli simili a quelli già sperimentati, esso dà inizio alla sequenza comportamentale che in tale situazione è stata maggiormente rinforzata. Il maggiore pregio della teoria comportamentale è costituito dalla sua 'parsimonia': tale teoria offre un modello di stima temporale in cui non è necessario fare ricorso a costrutti di tipo cognitivo quali interruttori, accumulatori e comparatori.
Il modello dell'oscillatore multiplo
L'orologio descritto dal modello dell'oscillatore multiplo (Church e Broadbent, 1990) è composto da diversi oscillatori, i cui differenti periodi coprono l'intera gamma di intervalli da stimare. Questi oscillatori sono completamente accoppiati e vengono azzerati all'inizio dell'intervallo. Il tempo trascorso viene determinato riferendosi alla metà della fase di ciascun oscillatore, e ottenendo in tal modo un vettore le cui componenti sono pari a +1 o −1. La memoria di lavoro è rappresentata in questo caso da una matrice autoassociativa di questo vettore, e la memoria di riferimento è rappresentata da un'altra matrice avente le stesse dimensioni di quella della memoria di lavoro. Il modello prevede che, quando viene ricevuto un rinforzo, la memoria di riferimento viene aggiornata mediante il calcolo della media ponderata dei valori presenti nella memoria di riferimento esistente e dei contenuti attuali della memoria di lavoro. Per decidere quando emettere la risposta in un programma IF, viene effettuata una comparazione o valutazione della similarità tra il vettore che rappresenta il tempo trascorso e la matrice della memoria di riferimento.
Il principale vantaggio del modello dell'oscillatore multiplo risiede nel fatto che esso permette di fare predizioni molto particolareggiate di dati sulla stima temporale di cui gli altri modelli non danno alcuna spiegazione. Quale dei tre modelli appena delineati offra la migliore descrizione dei meccanismi che sono alla base della stima temporale, posto che vi sia una descrizione migliore delle altre, è motivo di intenso dibattito. Ciascuno di questi modelli ha i suoi punti di forza e di debolezza, che riflettono tanto la struttura teorica nella quale il modello è stato sviluppato quanto i diversi tipi di dati che con ognuno di essi si è voluto spiegare. In futuro, i modelli di stima temporale dovranno probabilmente tenere in considerazione non solo i risultati comportamentali ma anche la crescente quantità di dati di tipo neurofarmacologico concernenti l'orologio interno.
Dati neurofarmacologici
Rispetto a quanto ci è noto sull'orologio circadiano, sappiamo piuttosto poco dei meccanismi neuronali che sono alla base della stima degli intervalli temporali dell'ampiezza di secondi o minuti. Tuttavia, i risultati di recenti ricerche di tipo neurofarmacologico sembrano indicare che i diversi stadi dall'elaborazione dell'informazione temporale che abbiamo descritto implicano l'attività di aree cerebrali differenti e sono modulati da sistemi neurotrasmettitoriali differenti (Meck, 1996).
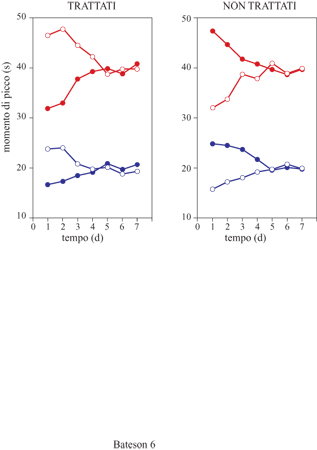
È stato dimostrato che la velocità dell'orologio è modulata dal livello funzionale di dopamina presente nei gangli della base. Per esempio, se un ratto viene addestrato a riprodurre un certo intervallo di tempo mediante la procedura del picco, e in seguito si esamina la sua prestazione dopo avergli somministrato sostanze che agiscono sul sistema dopaminergico, si osserva che gli agonisti della dopamina quali la metanfetamina e la cocaina producono un immediato spostamento del picco verso sinistra (cioè verso tempi minori) proporzionale alla durata dell'intervallo da stimare (v. figura 3). Questo risultato è compatibile con l'ipotesi che gli agonisti della dopamina siano in grado di accelerare l'orologio, poiché il ratto situa la fine dell'intervallo di tempo in anticipo rispetto al vero intervallo di criterio. Al contrario, gli antagonisti della dopamina come l'aloperidolo producono uno spostamento proporzionale del picco verso destra (tempi maggiori), il che è compatibile con un rallentamento dell'orologio. Se i ratti vengono addestrati per molti giorni consecutivi sotto l'effetto di queste sostanze, si osserva che le loro funzioni di stima tornano gradualmente a normalizzarsi, e questo dato sembra indicare che l'intervallo di criterio viene nuovamente appreso in base a un orologio interno funzionante a una velocità differente. A conferma di questa ipotesi, si è trovato che se a questo punto viene improvvisamente interrotta la somministrazione della sostanza, la funzione di stima temporale si risposta immediatamente nella direzione opposta allo spostamento osservato inizialmente (fig. 6). L'insieme degli effetti di queste sostanze è noto come effetto sull'orologio. W.H. Meck ha suggerito che le cellule che fanno da pacemaker per la stima degli intervalli temporali possano essere costituite dai neuroni della pars compacta della substantia nigra, poiché le lesioni di quest'area cerebrale producono una perdita totale della capacità di controllare le risposte in base al tempo, mentre la somministrazione sistemica di L-DOPA (un precursore della dopamina) è in grado di ristabilire questa capacità (Meck, 1996).
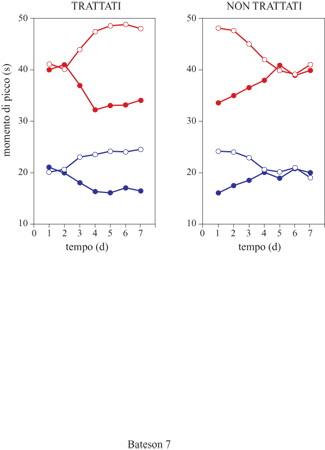
La memoria temporale e i meccanismi attenzionali sembrano essere correlati con la funzione dell' acetilcolina nella corteccia frontale. Nei ratti le sostanze che agiscono sul sistema colinergico producono distorsioni sistematiche della memoria temporale. Per esempio, se un ratto viene addestrato a riprodurre un intervallo di tempo mediante la procedura del picco, e la sua prestazione viene esaminata dopo avergli somministrato una dose di fisostigmina, un inibitore dell'acetilcolinesterasi, si osserva uno spostamento graduale e proporzionale del picco verso sinistra. Tale spostamento è compatibile con l'ipotesi che la fisostigmina faccia sì che nella memoria di riferimento vengano immagazzinati intervalli più brevi, che gradualmente sostituiscono la rappresentazione temporale più verosimile. Al contrario, gli antagonisti colinergici come l' atropina producono uno spostamento graduale e proporzionale del picco verso destra, compatibile con l'ipotesi di un immagazzinamento degli intervalli più lunghi. In entrambi i casi, quando la somministrazione della sostanza viene interrotta le funzioni tornano gradualmente a normalizzarsi nel giro di alcune sessioni; ciò suggerisce che gli intervalli vengano nuovamente appresi con parametri mnestici normali. Il pattern degli effetti di queste sostanze è stato denominato effetto sulla memoria (fig. 7). È stato osservato che, oltre alle manipolazioni di tipo farmacologico, anche le lesioni della corteccia frontale producono spostamenti di picco paragonabili a quelli tipici dell' effetto sulla memoria.
Meck (1996) ha suggerito che l'orologio interno contenuto nei gangli della base possa essere connesso ai meccanismi mnestici e attenzionali della corteccia frontale mediante circuiti striato- fronto-striatali. Alcuni dati a favore di questa ipotesi stanno emergendo da studi di risonanza magnetica funzionale condotti su esseri umani durante l'esecuzione di compiti di stima temporale.
Considerazioni ontogenetiche
Sappiamo poco circa lo sviluppo ontogenetico della capacità di stimare intervalli temporali. Non sappiamo se la capacità di regolazione temporale sia presente sin dalla nascita o dalla schiusa, o se compaia invece in qualche stadio ontogenetico successivo. Non sappiamo nemmeno se per il suo normale sviluppo sia necessaria un'esperienza ambientale specifica.
Esistono alcuni problemi di natura tecnica che hanno probabilmente ostacolato lo studio della stima degli intervalli temporali in animali giovani. In primo luogo, il fatto che gli esperimenti di stima temporale prevedano un periodo di addestramento dei soggetti che dura molti giorni costituisce un problema per lo studio di specie che maturano rapidamente. Inoltre, le tecniche tradizionalmente utilizzate per lo studio della stima temporale si basano sul condizionamento operante, che può essere di difficile realizzazione in soggetti giovani il cui sistema motorio non sia ancora pienamente sviluppato. Una soluzione a quest'ultimo problema è rappresentata dallo studio di specie a prole precoce. Una ricerca condotta su pulcini domestici (G. gallus) addestrati con programmi di rinforzo lF di 2 o 3 min ha riscontrato un buon tracciato 'a pettine' in soggetti esaminati tra 3 e 10 giorni dopo la schiusa, dimostrando che almeno in questa specie la capacità di stimare intervalli temporali è presente a partire da uno stadio di sviluppo molto precoce (Marley e Morse, 1966). Se riflettiamo sul fatto che questo tipo di stima è intimamente connesso al condizionamento classico, quest'ultimo risultato non dovrebbe sorprenderci poiché è stato ampiamente dimostrato che è possibile produrre un condizionamento classico persino nello stadio fetale dello sviluppo.
Considerazioni filogenetiche
Quando si è evoluta la capacità di stimare intervalli temporali, e qual è la sua storia filogenetica? Considerata la sua amplissima diffusione dal punto di vista tassonomico, sembra molto probabile che si tratti di un'abilità primitiva dei vertebrati, e dunque di qualcosa di evoluzionisticamente molto antico. Per determinare con certezza se tale abilità abbia preceduto o meno l'origine dei vertebrati, è indispensabile che vengano raccolti dati sulla stima temporale negli invertebrati. Se è vero, come abbiamo visto, che sembra esistere una stretta connessione tra la capacità di stimare intervalli temporali e l'apprendimento associativo, allora è molto probabile che tale capacità sarà documentata anche negli invertebrati, poiché molte specie appartenenti a questo gruppo sono capaci di questa forma di apprendimento.
La comprensione dei meccanismi che consentono la stima temporale potrebbe chiarire se gli orologi delle diverse specie siano o meno omologhi. A tale proposito, gli studi di tipo farmacologico stanno cominciando a offrire qualche supporto all'ipotesi di un orologio per intervalli comune a tutti i vertebrati. È stato recentemente dimostrato che le sostanze che agiscono sul sistema dopaminergico, come la metanfetamina, e che, come abbiamo visto, nel ratto (R. rattus) sono in grado di incrementare la velocità dell'orologio, influenzano questa variabile anche nel colombo (Kraemer et al., 1997) e nello storno europeo (s. vulgaris). Questi dati sembrano confermare l'ipotesi che un orologio per la stima degli intervalli temporali simile a quello di cui sono dotati gli uccelli e i mammiferi attuali fosse presente anche nei comuni pro genitori di queste due classi di animali. Analoghi studi farmaco logici condotti sui pesci potranno forse dimostrare che anche la capacità di stima degli intervalli temporali osservata in questa classe è attribuibile a questo stesso tipo di orologio.
Bibliografia citata
ASCHOFF, J. (1980) Ranges of entrainrnent: a comparative analysis of circadian rhythm studies. In Proceedings of the XIIIth Conference of the International Society of Chronobiology, Pavia, a c. di Halberg F., Scheving L.E., Powell, E.W., Milano, Il Ponte, pp. 105-112.
BRADSHAW, C.M., SZABADI, E., a c. di (1997) Time and behaviour: psychological and neurobehavioral analyses. In Advances in psychology, voI. 120., Amsterdam-New York, Elsevier.
BRUNNER, D., KACELNIK, A., GIBBON, J. (1992) Optimal foraging and timing processes in the starling, Sturnus vulgaris: effect of inter-capture interval. Anim. Behav., 44, 597-613.
CHANTREY, D.F. (1974) Stimulus preexposure and discrimination leaming by domestic chicks: effect of varying interstimulus time. J. Comp. Physiol. Psychol., 87, 517-525.
CHURCH, R.M. (1978) The internaI clock. In Cognitive processes in animaI behavior, a c. di Hulse S.H., Fowler H., Honig W.K., Hillsdale, L. Erlbaum Associate, pp. 277-310.
CHURCH, R.M., BROADBENT, H. (1990) Alternative representations of time, number and rate. Cognition, 37, 55-81.
DAVIES, N.B. (1977) Prey selection and the search strategy of the spotted flycatcher (Muscicapa striata). Anim. Behav., 25, 1016-1033.
GALLISTEL, C.R. (1990) The organisation of learning. Cambridge, Mass., MIT Press.
GALLISTEL, C.R. (1994) Space and Time. In AnimaI Iearning and cognition, a c. di Mackintosh N.J., San Diego, Academic Press., pp. 221-253.
GIBBON, J., ALLAN, L.G., a c. di (1984) Timing and time perception. New York, New York Academy of Sciences.
GIBBON, J., BALDOCK, M.D., LOCURTO, C., GOLD, L., TERRACE, H.S. (1977) Trial and intertrial durations and autoshaping. J. Exp. Psych.: Anim. Behav. Proc., 3, 264-284.
GIBBON, J., MALAPANI, C., DALE, C.L., GALLISTEL, C.R. (1997) Toward a neurobiology of temporal cognition: advances and challenges. Curr. Opin. Neurobiol., 7, 170-184.
GIBBON, J., MORRELL, M., SILVER, R. (1984) Two kinds of timing in circadian incubation rhythm of ring doves. Am. J. Physiol., 247, RI083-RI087.
GILL, F.B. (1988) Trapline foraging by hermit hummingbirds: competition for an undefended renewable resource. Ecology, 69, 1933-1942.
KACELNIK, A. (1984) CentraI pIace foraging in starlings (Sturnus vulgaris). I. Patch residence time. J. Anim. Ecol., 53,283-299.
KILLEEN, P.R., FETTERMAN, J.G., BIZO, L.A. (1997) Time's causes. In Time and behaviour: psychological and neurobehavioral anaIyses, a c. di Bradshaw C.M., Szabady E. In Advances in psychology, voI. 120., Amsterdam-New York, Elsevier.
KRAEMER, P.J., RANDALL, C.K., DOSE, J.M., BROWN, R.W. (1997) Impact of d-amphetamine on temporal estimation in pigeons tested with a production procedure. Pharm. Biochem. Behav., 58, 323-327.
MARLEY, E., MORSE, W.H. (1966) Operant conditioning in the newly hatched chicken. J. Exp. Anal. Behav., 9, 95-103.
MECK, W.H. (1996) Neuropharmacology of timing and time perception. Brain Res. Cogn. Brain Res., 3, 227-242.
POWELL, R.W. (1974) Comparison of differential reinforcement of low rates (DRL) performance in pigeons (Columba livia) and crows (Corvus brachyrhynchos). J. Comp. Physiol. Psychol., 86, 736-746.
RICHELLE, M., LEJEUNE, H. (1980) Time in animaI behavior. Oxford-New York, Pergamon.
ROBERTS, S. (1983) Properties and function of an internaI clock. In AnimaI cognition and behavior, a c. di Mellgren R.L., Amsterdam, North-Holland Publishing Company.
ROBERTS, S., CHURCH, R.M. (1978) Control of an internaI clock. J. Exp. Psych.: Anim. Behav. Proc., 4, 318-337.
TIMBERLAKE, W., WAHL, G., KING, D. (1982) Stimulus and response contingencies in the misbehavior of rats. J. Exp. Psych.: Anim. Behav. Proc., 8, 62-85.
Bibliografia generale
BRADSHAW, C.M., SZABADI, E., a c. di, Time and behaviour: psychological and neurobehavioral anaIyses. In Advances in psychology, voI. 120, Amsterdam-New York, Elsevier, 1997.
GALLISTEL, C.R. The organisation of Iearning. Cambridge, Mass., MIT Press, 1990.
GIBBON, J., ALLAN, L.G., a c. di, Timing and time perception. New York, New York Academy of Sciences, 1984.
GIBBON, J., MALAPANI, C., DALE, C.L., GALLISTEL, C.R. (1997) Toward a neurobiology of temporal cognition: advances and challenges. Curr. Opin. Neurobiol., 7, 170-184.
RICHELLE, M., LEJEUNE, H. Time in animaI behavior. OxfordNew York, Pergamon, 1980.