
Neocorteccia
Neocorteccia
Nell'uomo, la neocorteccia è la sede delle funzioni cognitive superiori e la sua complessità anatomica rispecchia l'importanza del compito che essa svolge nell'adattamento all'ambiente. La neocorteccia si presenta come un tessuto sottile, esteso e convoluto; la sua superficie è di 2600 cm2, lo spessore di 3÷4 mm. Essa contiene all'incirca 28×109 neuroni e, approssimativamente, un uguale numero di cellule gliali. I neuroni corticali sono collegati l'uno all'altro e agli assoni delle cellule di altre zone cerebrali da un vasto numero di terminali assonici chiamati 'sinapsi', nell'ordine di 1012. Attraverso diverse metodiche di indagine, si è visto che la corteccia è organizzata orizzontalmente in sei strati cellulari o lamine e verticalmente in gruppi di cellule collegate per mezzo di sinapsi che attraversano le lamine orizzontali. L'unità funzionale e anatomica di base della neocorteccia matura è la minicolonna, una stretta catena di neuroni che si estende verticalmente tra il II e il VI strato cellulare, perpendicolarmente alla superficie piale. Ogni minicolonna contiene circa 80÷100 neuroni, interconnessi saldamente l'uno all'altro in senso verticale; un'eccezione a questo principio è la corteccia striata dei Primati, che contiene un numero di neuroni superiore di circa 2,5 volte. La minicolonna si forma durante lo sviluppo dell'individuo attraverso la divisione iterativa di un piccolo gruppo di cellule progenitrici del neuroepitelio, tramite l'azione dell'unità ontogenetica della piastra corticale della neocorteccia in via di sviluppo.
Numerose minicolonne unite da connessioni orizzontali dense e a corto raggio formano a loro volta un'unità funzionale e anatomica chiamata 'colonna' o 'modulo corticale'. Le colonne presentano un diametro che va da 300 a 600 μm nelle specie in cui il volume della massa cerebrale varia in rapporto da 1 a 1000. Nel processo di sviluppo del cervello nella nostra specie, l'espansione corticale è stata raggiunta attraverso l'espansione dell'area della superficie corticale, con modeste variazioni di spessore. Questa espansione è determinata dall'aumento del numero delle colonne corticali, ma non della dimensione di ogni singola colonna. La proliferazione neuroepiteliale, la migrazione verticale lungo le cellule gliali e l'organizzazione radiale sono caratteristiche di tutti i Mammiferi. L'organizzazione colonnare permette una rappresentazione ricorsiva a intermittenza; in questo modo è possibile localizzare numerose caratteristiche funzionali sulla superficie bidimensionale della neocorteccia. Ciò appare con chiarezza nelle porzioni di corteccia che, in base alle afferenze talamiche e al loro ruolo funzionale, sono definite 'aree sensoriali primarie' del cervello; sembra altresì probabile che questo si verifichi in tutte quelle zone in cui sono state osservate mappe sistematiche.
La colonna corticale è una complessa unità elaborativa che collega un certo numero di informazioni in ingresso con un numero analogo di informazioni in uscita, tramite catene sovrapposte di elaborazione interna. A questo proposito, è stata osservata una segregazione parziale dei neuroni efferenti corticali con differenti bersagli estrinseci; quelli dal II al III strato laminare si proiettano verso altre zone corticali, quelli dal V al VI strato verso le strutture subcorticali. Ciò suggerisce che le operazioni di elaborazione che conducono ai diversi canali di output presentino differenze fondamentali.
Ontogenesi della neocorteccia
I neuroni corticali vengono generati nell'epitelio colonnare che riveste le zone ventricolari e subventricolari del telencefalo in via di sviluppo. Essi migrano dai rispettivi siti di generazione verso la posizione finale nella corteccia, attuando la prima divisione asimmetrica. I neuroni si differenziano ulteriormente dopo la migrazione e si dispongono in strati corticali nel sito in cui formano connessioni efferenti e ricevono connessioni estrinseche, caratteristiche dello strato. Essi sviluppano inoltre connessioni intrinseche translaminari che formano le basi reticolari delle colonne nella corteccia matura. Nella formazione della neocorteccia, l'evento iniziale è costituito dalla generazione di uno strato orizzontale denominato prepiastra, composto da un sottostrato superficiale di prime fibre nervose dirette alla corteccia, e dalle prime cellule neuronali: le cellule di Cajal-Retzius e uno strato cellulare inferiore che si trasformerà nella cosiddetta 'subpiastra'. Ondate successive di neuroni migranti formano la piastra corticale in una sequenza temporale che dall'interno si muove verso l'esterno creando i futuri strati laminari dal VI al II della corteccia matura. La piastra corticale è inserita nella prepiastra e la divide in una zona marginale estrema, il futuro I strato, e in una banda inferiore, spessa e parzialmente transitoria, la subpiastra.
I neuroni che migrano verso la neocorteccia si spostano lungo le fibre prodotte dalle cellule gliali radiali che formano barriere tra l'epitelio neuronale e la piastra corticale in via di sviluppo. Il tempo di generazione di un neurone determina la sua posizione finale laminare o verticale nella corteccia matura. La posizione spaziale originaria, nel piano x-y, di oltre il 90% dei neuroni dell'epitelio neuronale determina la posizione tangenziale finale della cellula nella neocorteccia matura. Nei 265 giorni di gestazione dell'essere umano, la neurogenesi inizia intorno al 40° giorno e termina intorno al 120°. I neuroni di proiezione e quelli intrinseci si differenziano, in maniera sequenziale, dagli strati profondi a quelli superficiali, contemporaneamente alla formazione degli strati nel processo di migrazione. Le cellule piramidali guidano gli interneuroni intrinseci nell'emissione e nella crescita di assoni e dendriti e nella formazione dei contatti sinaptici.
I percorsi migratori e la sequenza temporale degli eventi osservati nel cervello del feto di scimmia sono stati studiati in una storica serie di esperimenti da Pasko Rakic e hanno indotto a formulare 'l'ipotesi dell'unità radiale'. La migrazione dei neuroni immaturi dall'epitelio germinale alla piastra corticale si verifica soprattutto radialmente e lungo le cellule gliali. In seguito all'ultima divisione mitotica, i neuroni immaturi della zona ventricolare e di quella subventricolare si uniscono a un insieme adiacente di fibre-guida gliali. I neuroni generati serialmente nel tempo presso lo stesso sito dell'epitelio germinale migrano in maniera sequenziale lungo gli stessi insiemi di fibre-guida gliali o lungo insiemi adiacenti e si posizionano secondo il pattern interno/esterno in una colonna radiale. I neuroni di questa colonna radiale formano un'unità ontogenetica, cioè l'elemento fondamentale di edificazione della corteccia in via di sviluppo. Perciò, l'organizzazione colonnare di base della corteccia riflette il modo in cui si è generata.
L'area superficiale e, quindi, le dimensioni di ogni neocorteccia sono determinate dal numero delle sue unità ontogenetiche, stabilito dalle divisioni simmetriche delle cellule progenitrici presenti nell'epitelio neuronale prima dell'inizio della migrazione. Ogni ulteriore divisione simmetrica fa raddoppiare il numero delle unità ontogenetiche e, di conseguenza, anche l'area della neocorteccia. Si ritiene che, nel corso dell'evoluzione, la neocorteccia si sia espansa grazie a una mutazione del sistema che regola il numero di divisioni simmetriche che si verificano nelle zone regionali del neuroepitelio. Le unità ontogenetiche moltiplicate interagiscono con altri fattori nel creare nuove aree corticali e nel promuoverne l'evoluzione attraverso la selezione naturale.
L'organizzazione modulare della neocorteccia è stata documentata attraverso lo studio di numerose aree corticali sensoriali, motorie e associative ed è stata inoltre osservata in numerose condizioni sperimentali, per esempio, nella scimmia in stato di veglia e libera di muoversi. Di seguito sono illustrate le principali evidenze sperimentali ottenute attraverso indagini condotte sulle aree primarie somatosensoriali, su quelle visive e su alcune aree associative. Le aree corticali motorie e uditive condividono i medesimi principî organizzativi, ma non saranno oggetto di questo testo.
Studi fisiologici sulle aree somatosensoriali
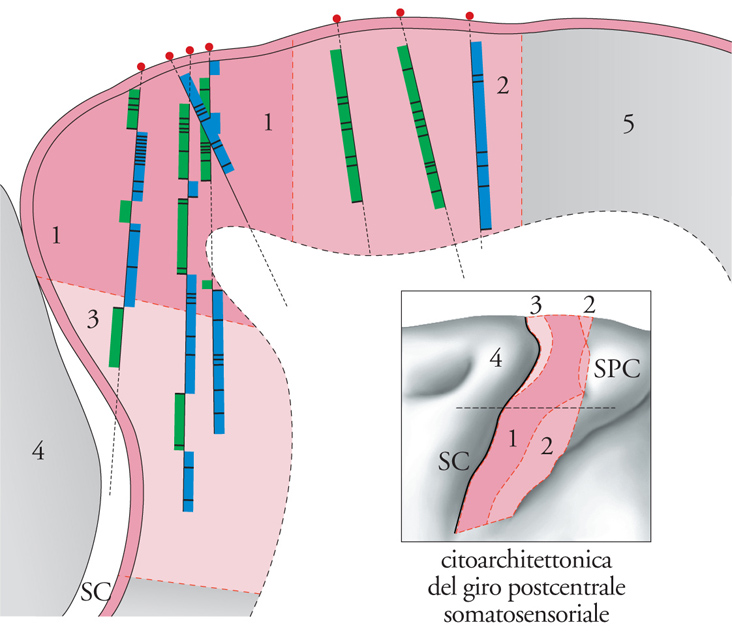
Dalle indagini elettrofisiologiche, condotte da Vernon B. Mountcastle nel 1957 e successivamente da quest'ultimo e da Thomas P.S. Powell (1959) su singoli neuroni della corteccia somatosensoriale (giro postcentrale) in gatti e scimmie anestetizzati, sono derivate le prime prove sperimentali dell'organizzazione colonnare della neocorteccia. I microelettrodi, introdotti perpendicolarmente alla superficie esterna del cervello e attraverso l'asse verticale della corteccia, incontrano in ogni strato cellulare neuroni aventi proprietà simili in termini di responsività a stimoli tattili (fig. 2). Il termine 'responsività' è impiegato in questo contesto con un significato particolare, per descrivere, cioè, la natura di determinati stimoli e il livello di adattamento a stimoli costanti. I microelettrodi incrociano zone di tessuto della dimensione di 300÷500 μm, in ognuna delle quali si incontrano neuroni con proprietà identiche. Ogni neurone riceve informazioni tattili soltanto da una specifica porzione periferica del corpo, possiede cioè un campo recettivo, ovvero un'area della cute o dei tessuti profondi che, se stimolata per esempio con una puntura di spillo o dalla pressione, determina l'attivazione elettrica (potenziale d'azione) in quel dato neurone.
L'organizzazione colonnare è di natura dinamica. In alcune aree corticali le proprietà di definizione delle colonne sono quasi totalmente stabilite da informazioni tattili provenienti dalla cute e dai tessuti; in altri casi predominano le proprietà indotte dall'elaborazione intracorticale. Il grado delle proprietà di definizione colonnare definito dal flusso afferente di entrata o dalle costruzioni intracorticali varia in rapporto alle aree corticali. Soltanto in una regione, quale la corteccia dell'area 3b, predominano le proprietà statiche stabilite dal flusso di entrata. Per esempio, le colonne direzionalmente selettive delle aree 1 e 2 sono prodotte dalla combinazione di proprietà statiche di spazio e di modalità con la selettività direzionale, un prodotto dell'elaborazione intracorticale. Un'importante scoperta è stata l'individuazione, nel giro postcentrale dei Primati, di quattro mappe somatotopiche separate della forma del corpo, distribuite nelle aree 3a, 3b, 1 e 2. Inoltre, le mappe corticali della superficie del corpo non consistono di rappresentazioni continue ma di insiemi di regioni, ognuna delle quali è deputata a un flusso afferente di entrata proveniente da una zona del corpo. Mentre la mappa disegna la sequenza generale del corpo, alcune regioni corticali adiacenti rappresentano zone del corpo molto distanti tra loro e alcune zone adiacenti del corpo sono rappresentate in zone corticali separate.
Oleg V. Favorov e Matthew E. Diamond (1990) hanno condotto studi sperimentali con l'introduzione di numerosi elettrodi formanti un fitto reticolo. I risultati hanno permesso di stabilire che la forma della colonna corticale è quasi esagonale, generalmente circondata da altre sei colonne, doppiamente convessa nella direzione della lunghezza, con una larghezza di 300÷400 μm: tale colonna contiene circa 80 minicolonne con diametro di 50÷60 μm. La colonna corticale si presenta separata e omogenea; i campi recettivi dei neuroni delle mini colonne di ogni colonna si spostano nel sito colonnare comune senza progressione topografica.
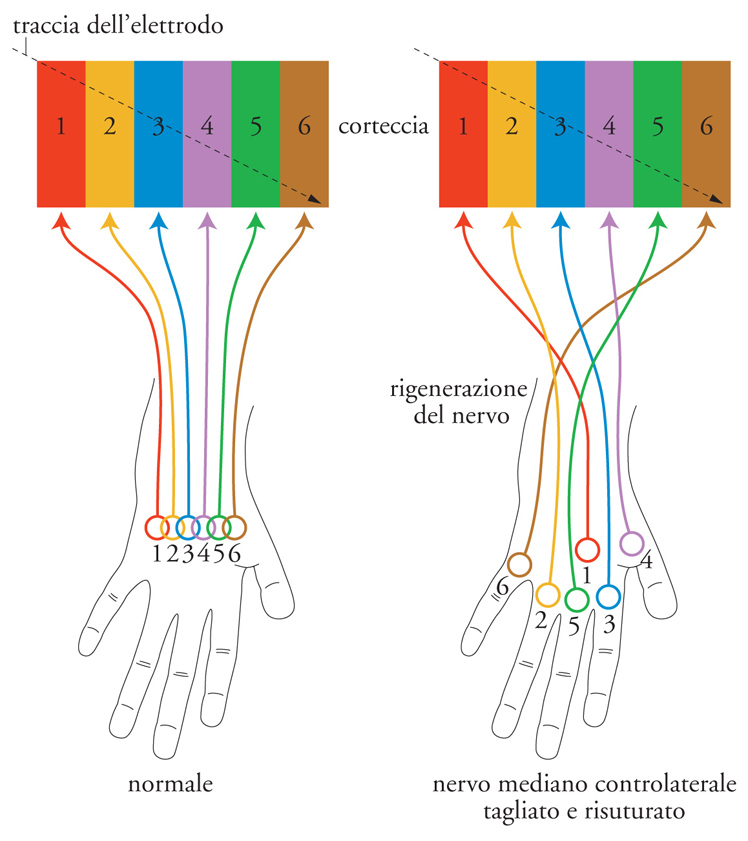
Jon H. Kaas e collaboratori (1981) hanno condotto un esperimento di rigenerazione nervosa sulle scimmie, ottenendo una prova ulteriore della composizione minicolonnare dei moduli corticali. Un esperimento iniziale di registrazione è stato effettuato con microelettrodi introdotti tangenzialmente attraverso l'area corrispondente alla mano nella corteccia postcentrale somatica. La fig. 3 a sinistra) illustra che, lungo una distanza considerevole, sono stati osservati neuroni con uno stesso tipo di proprietà modali collegati a campi recettivi che si spostano gradualmente e si sovrappongono. Successivamente è stato sezionato e risuturato il nervo mediano; si è quindi atteso il tempo necessario affinché la cute si innervasse nuovamente. L'esperimento di registrazione è stato quindi ripetuto con il risultato illustrato nella fig. 3, a destra. In luogo di una progressione regolare di campi recettivi sovrapposti, zone corticali della dimensione di 40÷60 μm seguono un percorso trasversale; al loro interno i campi recettivi sono identici ma si notano bruschi spostamenti tra zone adiacenti. Ciò è stato attribuito a una direzione impropria assunta dai fasci nervosi di reinnervazione periferica. Un fascio nervoso che innervi impropriamente una zona nuova e anormale della cute provoca la formazione di un nuovo campo recettivo sull'intera minicolonna dei neuroni verso la quale si proietta. Questo esperimento ha rivelato l'esistenza di una minicolonna con una dimensione paragonabile a quella delle unità embrionali, gli antenati delle minicolonne della corteccia adulta.
Inoltre, va detto che ognuna delle mappe nel giro postcentrale riceve l'input principale prevalentemente da uno o due degli insiemi modalmente specifici di fibre afferenti primarie. I neuroni di una porzione dell'area 3 sono modulati da fibre nervose provenienti dai muscoli. Una seconda porzione dell'area 3 e l'area 1 ricevono l'input dagli insiemi di fibre nervose ad adattamento rapido e lento provenienti dalla cute; l'area 2 riceve informazioni prevalentemente da fibre provenienti dalle articolazioni, anche se questa area contiene in aggiunta una rappresentazione completa della superficie cutanea della mano. Se una singola area riceve un input da più di un insieme di afferenti primari, i neuroni con differenti proprietà modali vengono segregati in zone diverse all'interno dell'area. Per esempio, i neuroni cutanei ad adattamento rapido e lento delle aree 3 e 1 sono segregati in colonne corticali differenti.
Infine, gli studi elettrofisiologici sono stati approfonditi da altre indagini, che hanno dimostrato come la stimolazione locale meccanica della superficie del corpo produca un aumento di attività metabolica in insiemi modulari nella corteccia controlaterale somatosensoriale di gatti e scimmie. L'attivazione coinvolge numerose regioni separate e allungate (0,5÷1,5 mm) come se numerose colonne adiacenti fossero attivate da stimoli periferici, poiché questi moduli definiti dal punto di vista metabolico sono più grandi di quelli definiti in modo elettrofisiologico. Questi insiemi collegati di moduli sono delimitati da zone con attività ridotta, dovuta all'inibizione intracorticale esercitata dal neurotrasmettitore inibitorio GABA (acido γ-amminobutirrico).
Mark Tommerdahl e collaboratori (1993) hanno condotto studi metabolici ed elettrofisiologici collegati, dimostrando che stimoli identici eccitano sia la zona dell'aumentata attività metabolica, sia l'attività dei neuroni al suo interno. L'incremento metabolico non è uniforme all'interno del modulo, ma si manifesta in strette colonne translaminari separate da zone ristrette con attività ridotta e con una frequenza spaziale di 18÷33/mm. Questo risultato sperimentale dimostra la presenza di minicolonne attive nella corteccia sensoriale e avvalora l'ipotesi generale secondo la quale i moduli corticali sono composti da gruppi di minicolonne. Tale variazione periodica a orientamento radiale del trasporto di 2-desossiglucosio rispetto al livello di fondo è stata osservata anche nelle regioni della corteccia somatica non attivate dall'impiego di stimoli periferici. La variazione spaziale è stata simile a quella osservata in misurazioni di densità ottica in sezioni della corteccia sensoriale colorate con il violetto di cresile (colorazione di Nissl); ciò costituisce un'indicazione indiretta dell'incolonnamento verticale dei corpi cellulari neuronali in questa regione.
Studi fisiologici sulla corteccia visiva
La corteccia visiva primaria dei Mammiferi, variamente denominata 'corteccia striata', 'area 17' o 'area V1', è la struttura cerebrale dei Mammiferi più intensamente studiata, a partire dall'inizio degli anni Sessanta, in seguito alle ricerche svolte da numerosi studiosi e in particolare da David H. Hubel e Torsten N. Wiesel (1977). Nel corso dei primi studi, questi scienziati hanno suggerito un'ipotesi, in seguito elaborata ampiamente tramite numerosi esperimenti, secondo la quale l'organizzazione colonnare nell'area V1 dei Primati è definita dalle proprietà neuronali di dominanza oculare e orientamento spaziale imposte dall'input proveniente dal nucleo genicolato laterale del talamo e dalla specificità di orientamento generata dall'elaborazione intracorticale. I neuroni dell'area V1 sono sollecitati di preferenza da stimoli giunti a uno degli occhi da un punto particolare del campo visivo e sono sensibili in maniera selettiva a barre luminose brevi con angoli di orientamento particolari e limitati. Queste proprietà si presentano più o meno costanti da neurone a neurone quando i microelettrodi sono introdotti perpendicolarmente alla superficie corticale esterna. Per contro, queste proprietà cambiano da un sito all'altro quando gli elettrodi vengono introdotti tangenzialmente per un lungo tratto, quasi parallelamente alla superficie. I neuroni esaminati durante le indagini effettuate tangenzialmente presentano variazioni sistematiche di selettività oculare e orientativa. Quest'ultima sembra risentire di movimenti minimi (20 μm) degli elettrodi, e può addirittura cambiare posizione da una catena cellulare verticale all'altra. Una sequenza completa di 180° viene coperta da distanze che variano da 500 fino a 750 μm. L'insieme di queste minuscole colonne orientative viene denominato ipercolonna per l'orientamento.
La proprietà di dominanza oculare di questi stessi neuroni si modifica in modo tale da rivelare le colonne adiacenti a dominanza oculare, una per occhio. I neuroni presenti in ogni coppia di colonne passano sistematicamente dalla dominanza completa di un occhio alla dominanza completa dell'altro, attraverso passaggi graduali; questo fenomeno è stato definito ipercolonna per la dominanza oculare. Basandosi sulle misurazioni delle dimensioni tangenziali delle due classi di ipercolonne e della funzione di dispersione luminosa puntiforme, Hubel e Wiesel hanno avanzato l'ipotesi secondo la quale insiemi doppi di ipercolonne sono sufficienti a elaborare tutti gli input provenienti da una regione determinata del campo visivo, con una sovrapposizione minima nelle regioni del campo visivo servite da insiemi adiacenti di ipercolonne. Con gli esperimenti successivi, condotti da Gary G. Blasdel nel 1992, gli insiemi colonnari nell'area V1 dei Primati sono stati studiati in maniera approfondita con i metodi di imaging ottico, in particolare gli insiemi in cui le colonne a dominanza sia di orientamento sia oculare sono identificate nella stessa regione localizzata della corteccia visiva. In alcuni siti la progressione laterale del cambiamento di preferenza di orientamento ha luogo in modo lineare, mentre altrove i cambiamenti sono marcatamente non lineari. Le colonne orientative formano piastre di orientamento che misurano 0,5÷1 mm nella direzione dell'isorientamento e nelle quali si ripete una rotazione completa di 180° della preferenza di orientamento nell'arco di circa 560 μm. Queste regioni lineari occupano circa il 50% della superficie corticale e sono localizzate di preferenza sui bordi delle colonne a dominanza oculare che intersecano in senso quasi ortogonale. L'intersezione crea un modulo in cui le numerose proprietà funzionali dei neuroni dell'area V1 vengono rappresentate con ritmo reiterativo, secondo un modello originariamente denominato 'cubo di ghiaccio' da Hubel e Wiesel.
Negli esperimenti di Blasdel sono stati identificati tre tipi di cambiamenti non lineari delle preferenze orientative. Le 'singolarità' sono costituite da discontinuità puntiformi che si creano quando le preferenze di orientamento mutano, in modo continuativo, per 180° intorno a loro; questi punti tendono a localizzarsi nei centri delle colonne a dominanza oculare. Le 'fratture' sono invece costituite da discontinuità localizzate presso le quali le preferenze di orientamento variano bruscamente; questi cambiamenti localizzati e improvvisi sono stati identificati da Hubel e Wiesel nei loro esperimenti elettrofisiologici. Infine i 'punti a sella' si configurano come piccole aree a forma di chiazza al cui interno le preferenze orientative sembrano stabili. Non è stato ancora appurato in maniera definitiva il ruolo che queste regioni di cambiamento di orientazione non lineare svolgono nella funzione della corteccia visiva.
La comprensione dell'architettura funzionale dell'area V1 dei Primati è stata resa più ardua dalla scoperta che lungo le colonne a dominanza oculare sono presenti, a intermittenza, zone localizzate di attività metabolica al di sopra della media. Queste zone, definibili come punti o macule (detti 'blob'), sono state rivelate mediante 2-desossiglucosio o citocromoossidasi. Sono zone localizzate con diametro di circa 150 μm più evidenti nel II e nel III strato, ordinate lungo il centro delle colonne a dominanza oculare a intervalli regolari di 500÷550 μm. Le file parallele sono distanti tra loro circa 350 μm. Le regioni maculate ricevono un input diretto dagli strati intercalati del nucleo genicolato laterale; le regioni che si alternano a esse ricevono solo un input corticale interno. I neuroni presenti nelle macule sono sensibili a particolari colori e non sono specifici per l'orientamento; i neuroni delle regioni intermedie si comportano in modo opposto. Le regioni a macula e quelle intermedie si differenziano per tipo di connettività interna ed esterna e per la diversa distribuzione di alcuni marcatori molecolari.
Studi fisiologici sulle aree associative
Una caratteristica sorprendente delle aree corticali associative è l'evidenza che i parametri di definizione delle colonne sono costituiti da proprietà neuronali che si sono formate all'interno dei sistemi di elaborazione intracorticale. Si tratta di proprietà complesse e quelle collegate all'input sensoriale che definiscono i moduli delle aree sensoriali primarie 3b o V1 raramente appaiono immodificate nelle aree associative. La plasticità delle sinapsi corticali sembra suggerire che tali proprietà possano essere modificate dall'esperienza, ovvero dall'apprendimento, e le evidenze preliminari sembrano confermare questa ipotesi. L'organizzazione modulare delle cortecce associative dipende in parte dal fatto che le connessioni tra queste aree sono distribuite in modo discontinuo nella dimensione corticale trasversale, sia all'origine sia al bersaglio. Esse terminano in colonne larghe 200÷500 μm, con particolare densità soprattutto negli strati sopragranulari, separate da zone di uguale larghezza in cui i terminali con quella origine sono rari. Le cellule provenienti dall'area di origine sono distribuite in modo simile in zone discontinue del II e del III strato.
Si considerano brevemente tre aree associative per le quali l'organizzazione modulare è stata documentata da esperimenti elettrofisiologici condotti su scimmie in stato di veglia: l'area parietale posteriore, la temporale mediale e la temporale inferiore. Nella prima, detta 'area 7a' (attualmente divisa in varie sottoaree) i neuroni con proprietà similari sono ordinati in moduli verticali che si estendono attraverso gli strati cellulari, collegati ad altri insiemi nei quali i neuroni sono caratterizzati da proprietà differenti. Queste aree corticali non riflettono singoli input sensoriali, né sono connesse in modo incondizionato a effettori periferici. Le proprietà neuronali che le caratterizzano sono state scoperte grazie al coordinamento di esperimenti elettrofisiologici e di indagini comportamentali condotte su scimmie addestrate a eseguire azioni scelte in base ad alterazioni comportamentali prodotte da lesioni dell'area e dei suoi tessuti connettivi. Le proprietà degli insiemi modulari della corteccia parietale posteriore differiscono notevolmente ma hanno un elemento in comune: sono tutte collegate alle azioni svolte dall'animale nello spazio immediatamente circostante, o alla sua percezione degli oggetti presenti e degli eventi accaduti in quello spazio e all'attenzione rivolta a essi.
Insiemi neuronali differenti si attivano in diverse situazioni: quando lo sguardo e l'attenzione si fissano su un oggetto che suscita interesse; quando l'oggetto viene seguito lentamente; quando il braccio si estende verso l'oggetto; quando l'oggetto viene manipolato; quando si produce una stimolazione visiva. L'ultima classe di neuroni ha prodotto proprietà visive non individuabili in alcuna delle aree corrispondenti gerarchicamente inferiori; essi forniscono un'immagine dinamica dello spazio circostante, del movimento dell'animale attraverso quello spazio o del movimento degli oggetti attraverso di esso. Quando perpendicolarmente alla superficie del lobo parietale inferiore vengono introdotti microelettrodi, esiste un'elevata probabilità di incontrare neuroni appartenenti a una sola di queste classi, mentre il punto di registrazione si sposta attraverso gli strati cellulari. Penetrando con gli elettrodi ortogonalmente alla dimensione verticale della corteccia, si incontrano masse di neuroni in successione, prima di una classe e poi di un'altra. Attualmente, non è stato ancora chiarito in quale modo questi vari insiemi modulari siano collegati nell'area 7a.
La corteccia temporale mediale (MT) è una piccola area con un'organizzazione cellulare interna molto particolare, posta nella sponda posteriore del solco temporale superiore della scimmia; è stata scoperta da Semir Zeki ed è specializzata per l'elaborazione delle informazioni sul movimento di oggetti visibili e sulla loro direzione di movimento. Zeki l'ha denominata V5, area visiva prestriata di quinto ordine; essa riceve input sia dall'area V1 sia dalla V2. L'omologa della V5 delle scimmie è stata identificata da Josef Zihl e collaboratori (1983), nel cervello umano con la PET (Positron emission tomography). Una paziente con lesioni bilaterali e quasi simmetriche che includevano quest'area era stata oggetto di indagini minuziose; la donna non percepiva o non riconosceva gli oggetti in movimento, ma unicamente se erano fermi. La sua lesione era selettiva in modo specifico per la visione del movimento; gli aspetti della visione e del movimento percepibili attraverso gli altri sensi apparivano ragionevolente integri.
I neuroni dell'area MT delle scimmie con assi similari di preferenza per la direzione di movimento sono ordinati in colonne verticali e queste colonne sono ordinate a loro volta in piastre in cui una rotazione completa di 180° dell'asse di movimento è rappresentata in 400÷500 μm di corteccia. Le colonne dell'asse di movimento sono intersecate da un secondo insieme in cui le due opposte direzioni di movimento lungo ogni asse sono rappresentate in due colonne adiacenti. Quindi le colonne dell'area MT sono specificate dall'intersezione di due insiemi parametrici, ognuno dei quali viene prodotto dall'elaborazione intracorticale.
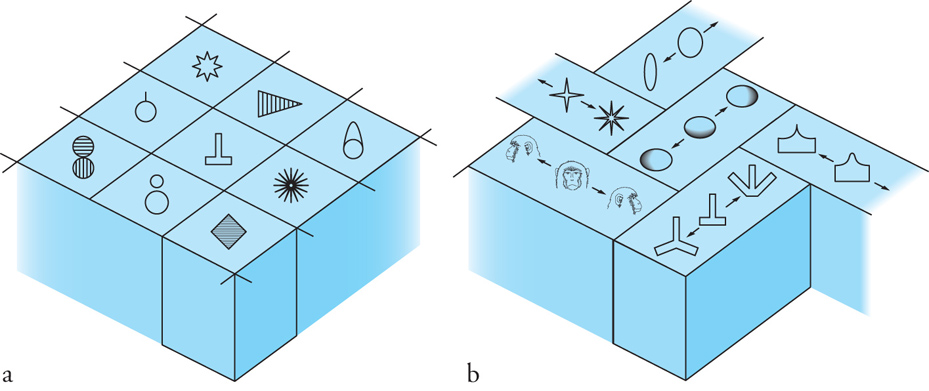
La corteccia infero-temporale è fondamentale per la percezione visiva degli oggetti; infatti la sua rimozione nella scimmia danneggia gravemente le capacità di apprendere a distinguere o a riconoscere un oggetto attraverso la vista. Molti neuroni di quest'area rispondono selettivamente alle forme degli oggetti e alcune cellule vengono attivate soprattutto da forme particolari quali, per esempio, le sagome della mano o del muso della scimmia o quella del volto umano. In questo contesto è di particolare interesse il lavoro sperimentale condotto da Keiji Tanaka e collaboratori (1993). Tanaka è riuscito a ridurre i contorni di stimoli complessi, come per esempio i volti, nelle sagome essenziali all'attivazione neuronale e ha confermato che la maggior parte delle cellule presenti nella zona anteriore della corteccia temporale inferiore è attivata da sagome moderatamente complesse. I neuroni con selettività simili o strettamente correlate sono raggruppati in colonne che si estendono attraverso gli strati cellulari, mentre i neuroni con selettività diverse sono ordinati in un mosaico simile a quello illustrato nella fig. 4.
Basi anatomiche dell'organizzazione colonnare
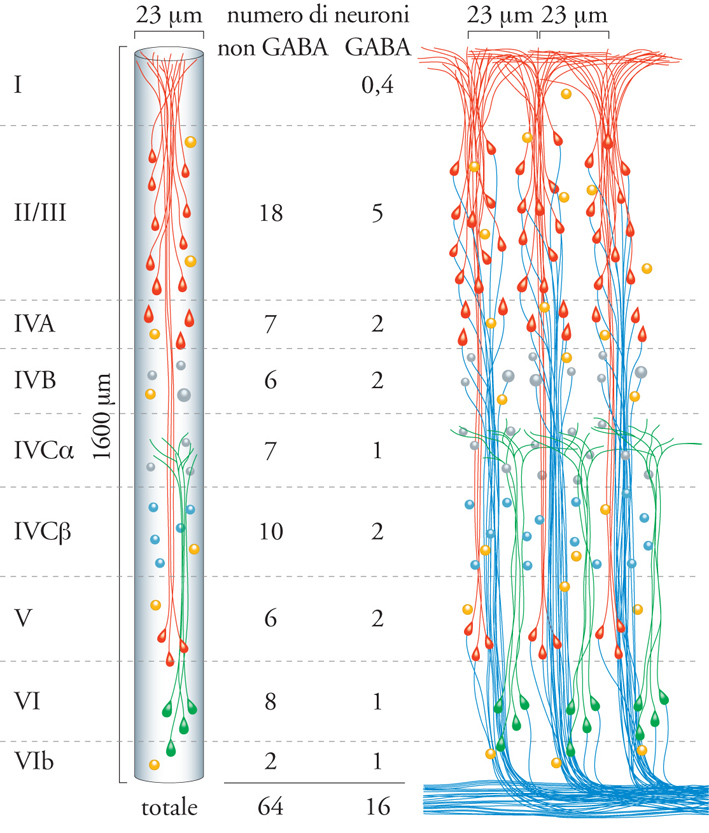
Quando venne avanzata per la prima volta (Mountcastle 1957), l'ipotesi dell'organizzazione colonnare fu accolta con diffidenza dalla maggior parte degli studiosi di neuroanatomia. Negli anni Quaranta del XX sec., era dominante l'idea classica di un'organizzazione laminare della corteccia e venivano avanzate con frequenza ipotesi circa la specificità funzionale di ogni strato cellulare; si riteneva, per esempio, che gli strati sopragranulari fossero specializzati per le funzioni psichiche. La specificità colonnare era di natura dinamica ed era sostenuta dall'inibizione pericolonnare, la quale non poteva essere rivelata dai metodi anatomici allora in uso. L'introduzione di nuovi metodi nella ricerca neuroanatomica ha permesso di dimostrare l'esistenza di un'organizzazione colonnare della neocorteccia, di cui si descriveranno gli aspetti essenziali (fig. 5).
Incolonnamento per mezzo di proiezioni afferenti
Attualmente siamo in possesso di descrizioni dei due ordinamenti anatomici che convalidano l'ipotesi colonnare (Jones 1983). Numerose indagini anatomiche e fisiologiche, collegate tra loro, hanno dimostrato che il sistema sensoriale afferente, chiamato lemnisco mediale, è organizzato in moduli a partire dal livello di entrata della radice dorsale del midollo spinale fino a quello del giro postcentrale. I fasci di assoni provenienti dalle cellule dei moduli talamici si estendono verso le zone colonnari di terminazione nel IV strato e nel III della corteccia del giro postcentrale, formando insiemi separati da zone in cui i terminali sono molto meno densi. Le zone apicali della rappresentazione postcentrale di mani, piedi e volto sono collegate all'altro emisfero mediante un grosso fascio di fibre chiamato 'corpo calloso'. All'interno delle regioni della mappa somatotopica collegate tra loro, gli afferenti callosali terminano in zone con dimensioni di 0,5÷1 mm. Queste zone e quelle terminali dei sistemi ipsilaterale cortico-corticale si sovrappongono per una distanza non ancora definita agli insiemi dei terminali degli afferenti talamici degli strati IV e IIIb, ma si estendono anche agli strati più superficiali; in molte altre aree corticali le zone terminali di questi due sistemi sono interconnesse. Esiste una convergenza perché le cellule piramidali dello strato IIIb che emettono fibre callosali ricevano un input sinaptico dagli afferenti talamici.
Le zone focali delle terminazioni di questi sistemi, cioè le callosali, sono ordinate in strisce mediolaterali che incrociano in modo quasi ortogonale le rappresentazioni generali anteriori e posteriori di ogni parte del corpo attraverso le aree 3b, 1 e 2. Le intersezioni specificano un modulo secondo il luogo, la modalità e la connessione interemisferica, anche se il significato di quest'ultima per ciò che concerne la funzione è ancora oscuro. La specificazione di un modulo per mezzo dell'intersezione delle strisce sembra essere una proprietà corticale generale poiché è individuabile sia nella corteccia uditiva sia in quella visiva, nonché nella corteccia postcentrale somatica. I risultati di questi studi anatomici e le relative numerose osservazioni di natura fisiologica permettono di formulare un'importante conclusione di carattere generale: la reale unità operativa di questo sistema distribuito non è rappresentata dal singolo neurone con il proprio assone, ma da gruppi di cellule con connessioni anatomiche e proprietà funzionali similari.
Incolonnamento per mezzo della connettività intrinseca
Gli interneuroni (cioè piccoli neuroni modulatori che connettono fra loro più cellule nervose vicine) locali del IV strato e dello stato IIIb, le cellule spinose non piramidali o stellate, sono uno dei principali bersagli postsinaptici di afferenti provenienti dal nucleo talamico ventro-posteriore. Gli assoni di questi interneuroni si estendono verticalmente in piccoli fasci delle dimensioni di 100÷200 μm che attraversano tutti gli strati cellulari e terminano sopra i dendriti e le spine dendritiche delle cellule piramidali e sopra gli interneuroni inibitori; essi proiettano la zona locale dei terminali talamici in una colonna translaminare.
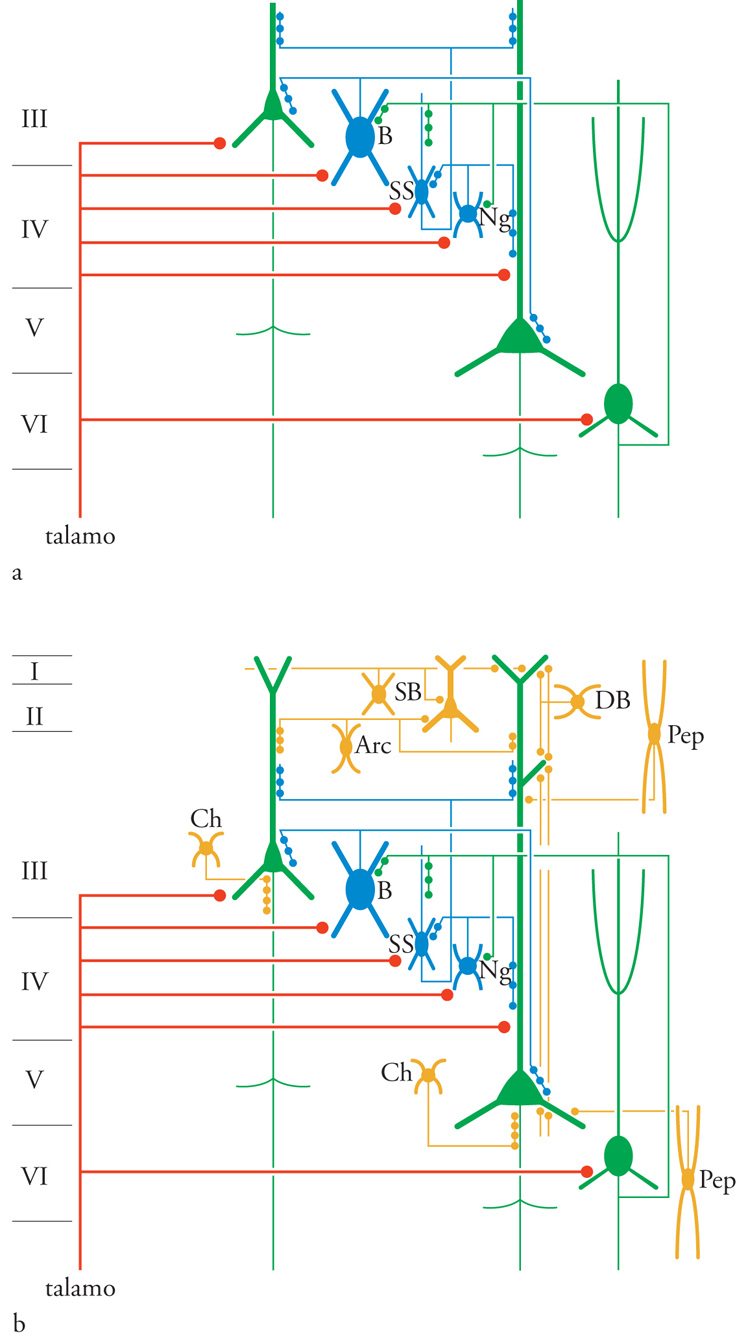
Come regola generale, tutte le altre azioni sinaptiche eccitatorie intrinseche alle colonne vengono imposte tramite le abbondanti ramificazioni collaterali ricorrenti degli assoni delle cellule piramidali. Questi circuiti di rientro creano un sistema eccitatorio bidirezionale, poiché le cellule piramidali degli strati sopragranulari innervano in questo modo le cellule piramidali degli strati infragranulari e viceversa. I collaterali di ogni assone staminale evitano la propria cellula d'origine ma si allungano diffusamente all'interno della zona ristretta costituita dalla popolazione colonnare locale delle cellule piramidali, fatta salva un'eccezione. Anche i collaterali ricorrenti terminano sopra gli interneuroni inibitori fornendo un meccanismo di inibizione pericolonnare. La fig. 6A illustra i vari tipi di cellule piramidali e non piramidali che ricevono afferenti talamocorticali e la fig. 6B cellule non piramidali intrinseche che non ricevono questo tipo di afferenti.
Altri collaterali ricorrenti degli assoni staminali delle cellule piramidali del II e del V strato di ognuno dei campi citoarchitettonici della corteccia postcentrale si proiettano orizzontalmente per lunghe distanze all'interno della propria area. Essi generano centri locali di contatti terminali simili a colonne che si incontrano a intervalli orizzontali di circa 800 μm. Queste proiezioni costituiscono vie di unione tra colonne aventi proprietà modali similari, oppure permettono a quelle con proprietà differenti di convergere trasversalmente; gli aspetti funzionali di queste proiezioni sono ancora sconosciuti. Anche le fibre nervose afferenti talamocorticali specifiche terminano direttamente sopra i dendriti delle cellule piramidali del III, V e VI strato e forniscono vie alternative alle connessioni monosinaptiche attraverso la corteccia somatosensoriale. I potenziali postsinaptici inibitori evocati nelle cellule piramidali dalle scariche talamocorticali indicano che anche gli afferenti talamocorticali terminano direttamente sopra gli interneuroni inibitori locali.
Le cellule GABA-ergiche inibitorie del tipo a doppio bouquet sono presenti in tutti gli strati, ma più densamente nel II e nel III. Esse proiettano i propri assoni nei fasci verticali della corteccia e terminano sopra le cellule piramidali e gli interneuroni inibitori. Queste cellule impongono una potente corrente inibitoria a direzione verticale e possono anche esercitare una disinibizione a direzione verticale delle cellule piramidali sopra le quali si proiettano gli altri interneuroni inibitori. La stretta distribuzione verticale degli assoni a doppio bouquet è così specifica e limitata che crea uno stretto cilindro verticale di inibizione/disinibizione che attraversa verticalmente la corteccia. Inoltre, molte altre classi di interneuroni inibitori formano anelli locali di controllo sul corpo delle cellule piramidali e sopra i segmenti iniziali degli assoni. Le proiezioni orizzontali degli assoni mielinizzati delle cellule a canestro grandi si estendono per 1÷2 mm e forniscono un altro percorso all'inibizione pericolonnare. Le cellule nervose a forma gliale si proiettano sopra gli interneuroni eccitatori locali di tipo spinoso degli strati IV e IIIb formando un saldo anello inibitorio presso il sito dell'entrata corticale. Le cellule che usano peptidi come neurotrasmettitori terminano sopra i corpi e i dendriti delle cellule piramidali ma le conoscenze riguardanti la loro funzione sono scarse.
Tra tutti i sistemi corticali, i circuiti del I strato sono i meno noti; non si conoscono ancora le modalità attraverso le quali essi incidono sulla formazione e l'operatività delle colonne e rimane tuttora un mistero il ruolo da essi svolto in una funzione corticale più globale.
Bibliografia
Blasdel 1992: Blasdel, Gary G., Differential imaging of ocular dominance and orientation selectivity in monkey striate cortex, "Journal of neuroscience", 12, 1992, pp. 3115-3138.
Blasdel 1992: Blasdel, Gary G., Orientation selectivity, preference and continuity in monkey striate cortex, "Journal of neuroscience", 12, 1992, pp. 3139-3161.
Favorov, Diamond 1990: Favorov, Oleg V. - Diamond, Matthew E., Demonstration of discrete place-defined columns - segregates - in the cat SI, "Journal of comparative neurology", 298, 1990, pp. 97-112.
Horton 1984: Horton, Jonathan C., Cytochrome oxidase patches: a new cytoarchitectonic feature of monkey visual cortex, "Philosophical transactions of the Royal Society of London. Series B", 304, 1984, pp. 199-253.
Hubel, Wiesel 1977: Hubel, David H. - Wiesel, Torsten N., Functional architecture of macaque monkey cortex, "Proceedings of the Royal Society of London. Series B", 198, 1997, pp. 1-59.
Jones 1983: Jones, Edward, G., The columnar basis of cortical circuitry, in: The clinical neurosciences, edited by Roger N. Rosenberg, New York, Churchill Livingstone, 1983, V, pp. 199-236.
Kaas 1981: Kaas, Jon H. e altri, Organization of somatosensory cortex of primates, in: Organization of the cerebral cortex of primates, edited by Francis O. Schmitt e altri, Cambridge (Mass.)-London, MIT Press, 1981.
Keller 1993: Keller, Asaf, Intrinsic synaptic organization of the motor cortex, "Cerebral cortex", 3, pp. 430-441.
Mountcastle 1957: Mountcastle, Vernon B., Modality and topographic properties of single neurons of cat's somatic sensory cortex, "Journal of neurophysiology", 20, 1957, pp. 408-434.
Powell, Mountcastle 1959: Powell, Thomas P.S. - Mountcastle, Vernon B., Some aspects of the functional organization of the cortex of the postcentral gyrus of the monkey: a correlation of findings obtained in a single unit analysis with cytoarchitecture, "Bulletin of the Johns Hopkins Hospital", 105, 1959, pp. 133-162.
Rakic 1990: Rakic, Pasko, Critical cellular events in cortical evolution: radial unit hypothesis, in: The neocortex: ontogeny and phylogeny, edited by Barbara L. Finley, Giorgio Innocenti and Henning Scheich, New York, Plenum Press, 1991, pp. 21-32.
Sur 1984: Sur, Mriganka - Wall, John T. - Kaas, Jon H., Modular distribution of neurons with slowly adapting and rapidly adapting responses in area 3b of somatosensory cortex in monkeys, "Journal of neurophysiology", 51, 1984, pp. 724-744.
Tanaka 1993: Tanaka, Keiji e altri, Serial processing of visual object features in the posterior and anterior parts of inferotemporal cortex, in: Brain mechanisms of perception and memory: from neuron to behavior, edited by Taketoshi Ono e altri, New York, Oxford University Press, 1993.
Tommerdahl 1993: Tommerdhal, Mark e altri, Minicolumnar activation patterns in cat and monkey SI cortex, "Cerebral cortex", 3, 1993, pp. 399-411.
Zeki 1991: Zeki, Semir e altri, A direct demonstration of functional specialization in human visual cortex, "Journal of neurophysiology", 11, 1991, pp. 641-649.
Zihl 1983: Zihl, Josef - von Cramon, D. Yves - Mai, Norbert, Selective disturbance of movement vision after bilateral brain damage, "Brain", 106, 1983, pp. 313-340.