
Neoplasie
Neoplasie
di Francesco Squartini e Luigi Califano
NEOPLASIE
Oncologia umana
di Francesco Squartini
sommario: 1. Introduzione. 2. Epidemiologia e frequenza: a) i tipi più frequenti di tumori umani; b) tumori in aumento e in diminuzione; c) influenza del sesso; d) distribuzione geografica; e) classe sociale e ambiente; f) associazioni particolari. 3. Errore diagnostico: a) idoneità e attendibilità dei materiali statistici in oncologia; b) gradi di accuratezza diagnostica; c) confronto fra diagnosi cliniche e anatomopatologiche; d) errore diagnostico reale e apparente; e) effetto sulle indagini di frequenza e di distribuzione; f) variazioni in rapporto al sesso, all'età e al periodo considerati. 4. Predisposizione genetica: a) suscettibilità cromosomica al cancro; b) suscettibilità mendeliana recessiva; c) suscettibilità mendeliana dominante o familiare; d) predisposizione genetica ai cancri più comuni; e) indagini sui gemelli monocori; f) conclusioni. 5. Cause del cancro umano: a) fattori cancerogeni professionali; b) inquinamento ambientale; c) cancerogeni nella dieta; d) cancerogenesi latrogena; e) abitudini di vita e voluttuarie; f) parassiti e virus oncogeni; g) prevenzione e screening delle sostanze cancerogene. 6. Immunodepressione e malignità: a) l'ipotesi della ‛sorveglianza immunologica'; b) implicazioni cliniche; c) difetti immunologici e tumori; d) regressione spontanea dei tumori; e) azione immunosoppressiva dei cancerogeni; f) critiche sperimentali all'ipotesi e conclusioni. 7. Storia naturale della malattia neoplastica nell'uomo: a) fasi della malattia neoplastica; b) il modello del cancro mammario; c) altri comuni tipi di cancro umano; d) rappresentazione schematica della malattia neoplastica nell'uomo. 8. Precursori morfologici: a) definizione; b) analisi della definizione; c) acquisizioni sperimentali; d) variabilità e molteplicità strutturale; e) classificazione patogenetica; f) iprecursori morfologici del cancro umano; g) attuali indirizzi di ricerca e conclusioni. 9. Progressione tumorale: a) definizione; b) fonti di informazione; c) caratteri tumorali acquisibili per progressione; d) principi generali (regole) della progressione tumorale; e) esempi e considerazioni; f) meccanismi di progressione; g) notazioni critiche. 10. Diffusione metastatica: a) definizione e vie di metastatizzazione; b) fasi della metastatizzazione; c) distribuzione delle metastasi al tavolo anatomico; d) fattori che influenzano la metastatizzazione. 11. Manifestazioni cliniche, problemi di diagnosi e di prognosi: a) durata clinica; b) comportamenti particolari; c) fattori morfologici di prognosi; d) diagnosi precoce; e) sindromi paraneoplastiche; f) marcatori biologici e morfologici. 12. Stato attuale della terapia: a) chemioterapia; b) agenti chemioterapici; c) meccanismi di azione; d) protocolli terapeutici; e) limiti e danni della chemioterapia; f) terapia ormonale; g) immunoterapia; h) conclusioni. 13. Tumori dell'infanzia e dell'adolescenza: a) peculiarità e problemi; b) epidemiologia e frequenza; c) quadri di patologia; d) curve di distribuzione; e) possibili fattori causali. 14. Principali tipi di tumori maligni negli adulti: a) cancro della mammella; b) cancro del polmone; c) cancro dell'intestino; d) cancro dello stomaco; e) leucemie e linfomi. 15. Conclusioni. □ Bibliografia.
1. Introduzione
Le neoplasie o tumori, che indubbiamente assurgono a protagonisti nella patologia di questo secolo, sono ancora oggi entità biologiche e malattie poco conosciute, per le quali si può dire che manchi persino una definizione pienamente soddisfacente. Infatti, ‟poiché la natura e le cause dell'accrescimento tumorale restano ignote, le definizioni esistenti sono in realtà delle descrizioni e, come tutte le descrizioni, alcune sono migliori di altre" (v. Walter e Israel, 19672). Fra queste, la definizione fornita da Willis (v., 1948 e 19674) sembra contenere tutti i concetti essenziali insiti nello sviluppo di una neoplasia e perciò merita di essere citata: ‟Tumore è una massa abnorme di tessuto, il cui accrescimento eccede quello dei tessuti normali, è incoordinato con quello, e persiste nella stessa maniera eccessiva dopo cessazione degli stimoli che lo hanno provocato". In tal modo una neoplasia si distingue dalle proliferazioni finalistiche di natura infiammatoria e riparativa, dalle iperplasie e dalle malformazioni con eccesso di tessuto.
Qualsiasi tipo di cellula o tessuto dell'organismo uma- no, sebbene con frequenze diversissime, può dare origine allo sviluppo di un tumore. Poiché i tumori, nella maggior parte dei casi, conservano una qualche rassomiglianza morfologica con le cellule e i tessuti di origine, le classificazioni e le nomenclature in uso sono generalmente su base istogenetica. A questo proposito si deve ricordare che una neoplasia è per prima cosa e fondamentalmente un problema di morfopatologia. Infatti non esiste ancora la possibilità di diagnosticare con certezza un tumore senza o al di fuori dell'osservazione morfologica, che può essere anche solo macroscopica (sebbene sia difficile formulare un giudizio conclusivo con la sola osservazione a occhio nudo o mediante lente di ingrandimento), ma quasi sempre oggi è microscopica e cioè istologica sui tessuti o citologica sulle cellule isolate (v. Ashley, 19783).
Sul piano anatomoclinico assume tuttavia importanza fondamentale, dopo quella morfologica, la classificazione basata sul comportamento dei tumori. Infatti, i tumori di ogni tipo cellulare possono mostrare un'ampia gamma di comportamenti che riguardano ad esempio la differenziazione morfologica, la velocità di accrescimento, le modalità di diffusione, il grado di pericolosità per l'ospite. Alcuni tumori crescono lentamente, solo nel luogo di origine, non invadono i tessuti vicini e non causano pericoli al paziente, salvo il caso di particolari sedi o complicanze: questi sono i tumori innocenti o benigni. Altri tumori dello stesso tipo cellulare crescono invece rapidamente, invadono i tessuti contigui, si diffondono attraverso impianti successivi metastatici a distanza e alla fine uccidono l'ospite se non sono trattati o rimossi precocemente: questi sono i tumori maligni. Fra gli uni e gli altri esistono talora anche forme di confine, provviste appunto di un comportamento intermedio, le quali indicano quanto sia arbitraria quella distinzione netta che per necessità siamo soliti fare a scopo diagnostico e prognostico nel campo dei tumori fra l'innocenza e la malignità (v. Willis, 19674).
Nella maggior parte dei casi è fortunatamente semplice, attraverso un esame istologico, stabilire per le esigenze pratiche se un tumore è benigno o maligno. Tali criteri di diagnosi istologica possono, per i motivi detti, fallire in certi casi, ma su di essi si basa generalmente ancora oggi la condotta del medico e la terapia. Il problema anatomoclinico dei tumori nell'uomo riguarda essenzialmente le neoplasie maligne alle quali pertanto questa rassegna è in prevalenza dedicata. I tumori maligni di derivazione epiteliale, riguardanti la cute, le mucose e le strutture ghiandolari, vengono generalmente indicati col nome di carcinomi o cancri. I tumori maligni di derivazione connettivale o mesenchimale, riguardanti i tessuti di sostegno, vengono invece generalmente indicati col nome di sarcomi. Altri tumori hanno derivazioni e nomi particolari. Questo è ad esempio il caso dei tumori delle sierose (pleura, pericardio, peritoneo) e del tessuto nervoso.
Uno dei fatti che più colpiscono a proposito dei tumori maligni umani è il loro progressivo aumento di frequenza. Ciò non è vero in assoluto, poiché vi sono tumori, come ad esempio il cancro dello stomaco e del collo dell'utero, che mostrano da vari anni tendenza alla diminuzione, ma è certamente vero per le neoplasie nel loro complesso. Queste, come causa di morte, nel corso del nostro secolo sono per lo meno quadruplicate e occupano oggi il secondo posto fra tutte le cause di morte, dopo le malattie cardiovascolari (v. Lilienfeld e altri, 1972).
Alcuni motivi di questo continuo e impressionante aumento di frequenza sono facilmente intuibili. L'aumento della durata media della vita incide di certo in maniera rilevante sulla frequenza dei tumori maligni, poiché questi sono generalmente malattie dell'età media-avanzata (v. Strong, 1968). La scomparsa come causa di morte di gran numero di malattie, specie infettive, che oggi si possono curare, è di conseguenza uno dei motivi dell'aumento di frequenza dei tumori. Un'altra causa può essere trovata nelle possibilità di diagnosi delle neoplasie maligne oggi nettamente migliorate rispetto al passato prossimo e remoto. Ma purtroppo gran parte dell'incremento registrato nella morbilità e mortalità per tumori dipende certamente dalle anormali condizioni di vita, dalle abitudini dietetiche e voluttuarie, dall'inquinamento ambientale a opera di cancerogeni cui ci ha condotti nel tempo lo sviluppo della nostra civiltà (v. Hueper e Conway, 1964).
Era opinione diffusa in passato che i tumori dell'uomo fossero spontanei, cioè originati a caso. Gli studi epidemiologici, quelli riguardanti gli ambienti di lavoro, e quelli, sempre più analitici, sui fattori di rischio nei vari tipi tumorali, vanno scoprendo sotto i nostri occhi un'impressionante costellazione di cause dei tumori umani (v. Hiatt e altri, 1977). Diviene perciò sempre meglio evidente il loro meccanismo di induzione e la fondata presunzione che molti, con accorgimenti opportuni, si potrebbero evitare.
Una caratteristica che sfugge al profano circa la ma- lattia neoplastica nell'uomo è la sua lunga durata complessiva. Infatti, la fase clinica che va dalla diagnosi alla morte in casi di esito infausto è relativamente breve, di mesi o di 1-2 anni. Ma questa è preceduta in genere da una serie di tappe evolutive silenti o poco appariscenti che spesso impiegano parecchi anni prima di raggiungere la soglia della diagnosi clinica (v. Foulds, 1969-1975). Uno sguardo alla storia naturale della malattia neoplastica nell'uomo, alle sue varie fasi successive, alla progressione attraverso la quale per gradi il tumore raggiunge stadi di crescente autonomia, induce a riflessioni e approfondimenti di notevole interesse pratico. Infatti, molte misure preventive e tentativi terapeutici avrebbero probabilmente successo se applicati precocemente, nella fase preclinica delle neoplasie maligne e ciò spinge alla ricerca di nuove informazioni di base sulle prime fasi dello sviluppo tumorale nei loro aspetti morfologici e biologici (v. Antony e altri, 1976). Solo per questa strada si potrà giungere veramente a una diagnosi precoce dei tumori maligni nell'uomo, che è il presupposto di una terapia efficace.
L'epidemiologia, la frequenza, i margini di errore, la predisposizione, le cause, il controllo immunologico, le tappe evolutive, i precursori morfologici, la progressione, la diffusione metastatica, e il profilo anatomoclinico dei principali tipi di neoplasie maligne umane saranno analizzati nei capitoli seguenti per sottolineare di volta in volta lo stato delle conoscenze, i problemi aperti, le linee dominanti di ricerca e le speranze di soluzione.
2. Epidemiologia e frequenza
Parlare di frequenza dei tumori in senso generale è impossibile, poiché ciascuno dei vari tipi tumorali è fortemente condizionato dal sesso, dalla razza, dal gruppo etnico (cioè dalla costituzione genetica), dall'ambito geografico considerato, dal tipo di civiltà e quindi dalle abitudini di vita, dall'ambiente (per esempio città o campagna), dall'alimentazione, dal lavoro, ecc. Vi sono tuttavia alcuni tipi di tumori maligni molto più frequenti di altri, qualunque sia la popolazione e la latitudine considerate.
a) I tipi più frequenti di tumori umani
Nel nostro paese, considerando i due sessi insieme e i soggetti adulti, sulla base dei certificati di morte, la graduatoria per apparati, organi o sistemi vede al primo posto i tumori dell'apparato digerente (49,5%), seguiti da quelli dell'apparato genitale compresa la mammella (18,6%), respiratorio (11,2%), emopoietico (5,7%), urinario (3,8%), nervoso (2,6%), di sostegno (ossa + connettivi: 1,7%), cutaneo (1,3%), endocrino (0,6%) e di altre sedi varie (5%). I tumori degli apparati digerente, genitale, respiratorio ed emolinfopoietico sono dunque da soli responsabili dell'85% dei decessi per tumori (v. Squartini e Bolis, 1966).
Tenendo prevalentemente presenti la razza bianca, il nostro tempo e l'emisfero occidentale, i 10 tipi di tumori più frequenti come causa di morte sono, in ordine decrescente, nell'uomo: il cancro del polmone, il cancro dell'intestino, le leucemie e i linfomi, il cancro dello stomaco, il cancro della prostata, il cancro del pancreas, il cancro del rene e delle vie urinarie, le neoplasie del sistema nervoso, il cancro del fegato e delle vie biliari, i tumori del cavo orale e della faringe. Nella donna è invece al primo posto il cancro della mammella, seguito dai cancri dell'intestino e dell'utero, dalle leucemie e dai linfomi, dai tumori dell'ovaio, dai cancri dello stomaco, del polmone, del pancreas, del fegato con le vie biliari e del rene con le vie urinarie (v. Lilienfeld e altri, 1972). Ne consegue che le neoplasie maligne di gran lunga più comuni nei due sessi sono quelle dell'intestino, del polmone, degli organi emolinfopoietici, della mammella e dello stomaco, seguite a qualche distanza dalle neoplasie del pancreas, della prostata, dell'utero, dell'ovaio, del rene e vie urinarie, del sistema nervoso, del fegato e vie biliari, del cavo orale e faringe.
Questo ordine di frequenza delle neoplasie, desunto dalle statistiche di mortalità, non rispecchia esattamente quello della incidenza assoluta dei singoli oncotipi come malattie. Nelle statistiche di morbilità per tumori, infatti, meno frequenti e conosciute, per la difficoltà di collezionare i dati relativi, sono ad esempio fra le prime posizioni i tumori della cute e in posizione di rilievo spesso anche i tumori del cavo orale. Queste neoplasie, poiché facili da diagnosticare precocemente per la loro sede, si possono curare e raramente sono oggi causa di morte (v. Ashley, 19783).
b) Tumori in aumento e in diminuzione
Esaminando le frequenze dei tumori umani durante un lungo periodo di anni, si possono notare per i vari tipi tendenze alla diminuzione, alla stabilità o all'aumento che rivestono importanza nel formulare previsioni per il futuro prossimo e nell'indirizzare le ricerche sui possibili fattori causali. Mostrano attualmente tendenza alla diminuzione i tumori del cavo orale e della faringe e quelli dello stomaco nell'uomo, come pure i tumori dell'esofago, stomaco, laringe, cervice uterina, vie urinarie e tiroide nella donna. Mostrano invece tendenza all'aumento nei due sessi i tumori del polmone, del pancreas, del rene, del sistema nervoso e degli organi emolinfopoietici, e nella donna ancora i tumori dell'ovaio. Sono infine stazionari nell'uomo i tumori dell'esofago, delle vie urinarie, della tiroide e nella donna quelli della mammella, del cavo orale e della faringe. Tendenze irregolari mostrano gli altri tipi di tumori (v. Lilienfeld e altri, 1972).
c) Influenza del sesso
La marcata influenza del sesso sulla frequenza dei tumori umani non si limita a quella, già registrata, relativa ai tumori degli organi della riproduzione e sessuali secondari, ma coinvolge la maggior parte dei tipi tumorali. Con poche e rare eccezioni si può dire che gli uomini hanno in genere un'incidenza di neoplasie più elevata delle donne. Il carcinoma primitivo del fegato è nettamente più frequente nell'uomo che nella donna, con ogni probabilità per differenze di ordine endocrino. Il cancro della cute è pure più frequente nell'uomo che nella donna, fatta eccezione per il melanoma. Anche il cancro del polmone e delle vie respiratorie in genere mostra una netta predilezione per il sesso maschile, ciò che può dipendere in parte da una diversa abitudine al fumo, ma probabilmente anche da cause endocrine. Nell'uomo sono inoltre più frequenti che nella donna i cancri delle labbra, del cavo orale, della faringe, delle ghiandole salivari, dell'esofago, dello stomaco, del pancreas e delle vie urinarie. Il sesso è in grado di influenzare anche la leucemogenesi, che più frequentemente interessa gli uomini. Al contrario nella donna sono più frequenti i tumori della tiroide e, forse, i melanomi (v. Toh, 1973).
d) Distribuzione geografica
La distribuzione dei tumori nei vari organi non è uniforme, come si è già accennato, nelle diverse parti del mondo essendo fortemente influenzata da un fattore geografico che in parte rispecchia predisposizioni razziali, ma in parte è la conseguenza di profonde differenze alimentari, igieniche e ambientali. Per esemplificazione, nella tab. I si riferiscono in percentuale le frequenze di morbilità relative ai principali oncotipi ricavate da quattro fra i più accurati Registri dei Tumori attualmente esistenti e precisamente quelli di Birmingham in Inghilterra, del Quebec in Canada, di Puerto Rico nei Caraibi e di Ibadan nell'Africa tropicale (v. Doll e altri, 1970; v. Ashley, 19783).
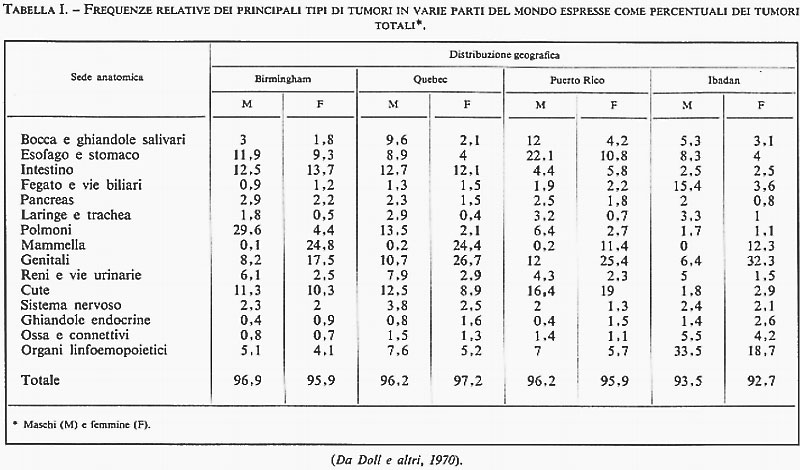
La frequenza di cancri dell'esofago e dello stomaco è particolarmente elevata a Puerto Rico, come del resto si osserva anche in altre parti del mondo (Giappone). Il cancro dell'intestino è invece molto meno frequente nei paesi poveri (Puerto Rico, Ibadan) che in quelli industrializzati (Birmingham, Quebec), ciò che sembra dipendere da una diversa composizione della dieta e delle feci. Il cancro del fegato è particolarmente frequente in Africa e lo stesso si osserva lungo una fascia estesa ai paesi costieri poveri della Cina meridionale. Sono queste geograficamente le regioni della dieta carente, squilibrata per scarsezza in genere di proteine, da cui discende una carenza di fattori epatoprotettivi, un'elevata frequenza di degenerazione grassa del fegato nei bambini (una sindrome nota col nome indigeno di Kwashiorkor), di cirrosi epatica nei giovani e di carcinoma del fegato in età più avanzata, favorito anche dalla presenza di particolari agenti cancerogeni nella dieta.
I cancri della mammella sono molto più frequenti nei paesi a più elevato tenore di vita (Birmingham, Quebec) che a Puerto Rico e Ibadan, un fatto da porsi in relazione con differenze, oltre che genetiche, nella riproduzione, nell'allattamento al seno, nell'alimentazione e nelle abitudini di vita. Il cancro dei genitali femminili è più frequente in Africa, forse per motivi igienici, così come accade per i tumori emolinfopoietici a causa della presenza in questa zona geografica del linfoma di Burkitt (o linfoma Africano), l'unica neoplasia dell'uomo per la quale sia stata dimostrata un'eziologia virale. Il cancro del polmone è invece di gran lunga più frequente nelle regioni industrializzate d'Inghilterra (v. Ashley, 19783). Molte di queste differenze geografiche, come si vede, riflettono particolari associazioni o carenze di fattori causali sui quali si ritornerà in seguito. Il cenno che ne è stato fatto è sufficiente, tuttavia, a sottolineare l'utilità degli studi di patologia geografica nella ricerca dei fattori causali in oncologia.
e) Classe sociale e ambiente
La classe sociale di appartenenza, come suggerito del resto da alcuni dati precedenti, ha pure importanza nello sviluppo dei tumori umani. Sono più frequenti oggi nelle classi agiate i tumori della mammella e dell'intestino. Sono più frequenti invece nelle classi povere i tumori dello stomaco e della cervice uterina. Come la classe sociale, anche l'ambiente di vita influenza marcatamente l'oncogenesi: nei paesi industrializzati, e per prevalenti motivi di inquinamento ambientale, il cancro del polmone è nettamente più frequente fra gli abitanti delle città che fra quelli delle campagne. Ciò ha condotto all'identificazione del ‛rischio urbano' e allo studio accurato della polluzione atmosferica come premessa per un risanamento ambientale (v. Willis, 19674).
f) Associazioni particolari
Nelle ricerche epidemiologiche più recenti, mirate all'identificazione di specifici fattori causali o predisponenti dei tumori umani, sono state identificate associazioni particolari e significative fra determinati consumi, nelle diverse aree geografiche o da parte di diverse popolazioni, e frequenza di taluni tipi di cancro. Un'associazione diretta, marcata e significativa è stata così rilevata nei paesi occidentali fra consumo di sigarette pro capite e mortalità per cancro del polmone (v. U.S. Department of..., 1964). Un'associazione analoga è stata dimostrata fra contenuto in grammi giornaliero di grassi nella dieta e mortalità per cancro della mammella (v. Fratkin, 1976). È stata inoltre messa in evidenza un'associazione geografica fra cancro del colon e della mammella nella donna, probabilmente basata sulla composizione della dieta (v. Drasar e Irving, 1973).
3. Errore diagnostico
La breve esposizione che precede sulla frequenza e sull'epidemiologia del cancro umano, non sarebbe completa e appieno valutabile nel suo significato se qui non si facesse cenno delle disponibilità di materiali, delle difficoltà di metodo, e dei margini di errore insiti nelle indagini statistiche sui tumori umani. Ciò consentirà fra l'altro di illustrare le basi anatomopatologiche della malattia neoplastica, dalle quali non si può prescindere senza rischiare largamente di sbagliare.
a) Idoneità e attendibilità dei materiali statistici in oncologia
La validità di un'indagine statistica dipende in primo luogo dall'idoneità e dall'attendibilità del materiale su cui essa si basa. La scelta di materiale idoneo è problema statistico, alla cui soluzione debbono guidare i criteri del metodo del campione, sufficientemente ampio, omogeneo e rappresentativo dell'universo che si intende studiare. Ma l'attendibilità di un materiale per uso statistico in oncologia umana dipende invece soltanto dal grado di accuratezza con cui i singoli casi che lo compongono sono stati diagnosticati, ed è perciò essenzialmente un problema di patologia. Capita molto spesso di osservare, nelle statistiche sui tumori, che il materiale giudicato idoneo per un determinato studio non sia del tutto attendibile e, viceversa, che un materiale attendibile non sia del tutto idoneo.
b) Gradi di accuratezza diagnostica
Willis (v., 19674) distingue quattro gradi di accuratezza crescente per le diagnosi di tumore, che sono: I) diagnosi a mezzo di sola visita medica; II) diagnosi a seguito di visita medica corredata da esami speciali (fisici, chimici, ematologici, radiologici, endoscopici, operatori) esclusa la biopsia; III) diagnosi a seguito di esame istologico di tessuti rimossi chirurgicamente (biopsia); IV) diagnosi a seguito di autopsia: a) senza conferma istologica o b) con conferma istologica. Soltanto i casi con diagnosi di grado IV) tipo b) possono considerarsi certi in senso assoluto. In tutti gli altri casi vi è la possibilità di un errore diagnostico, tanto più grande man mano che si sale dal grado IV) al I). Nelle diagnosi di grado I) la precisione è, ovviamente, assai scarsa, sebbene la frequenza di diagnosi esatte, come dire il livello di attendibilità, possa variare di molto in rapporto alla preparazione del medico, alla sede (superficiale o profonda) del tumore, all'ambiente urbano (con specialisti qualificati) o rurale (con prevalenza di medici generici e minore coscienza sanitaria nelle popolazioni) da cui i casi provengono.
Di qui discende la necessità di conoscere con esattezza per ogni indagine statistica in tema di tumori il tipo di materiale usato, da cui poter valutare con sufficiente rapidità e approssimazione il grado di attendibilità dei risultati. Va rilevato, tuttavia, che difficilmente esistono le condizioni per lavorare su materiale omogeneo quanto a livello di accuratezza diagnostica, e che ancor più rara in pratica è la possibilità di utilizzare materiali appartenenti ai gradi di precisione massima o assoluta.
Per conoscere, ad esempio, se la frequenza dei tumori maligni sia aumentata nel tempo e se l'aumento sia reale o fittizio, per indagare sulla distribuzione geografica dei tumori, per stabilire statisticamente se e quali rapporti esistano fra l'incidenza dei tumori nell'uomo e una qualsiasi delle variabili di interesse (tipo etnico, ambiente, alimentazione, abitudini, lavoro, condizioni sociali), ci si deve abitualmente servire di materiali compositi, cioè non omogenei. I certificati di morte rappresentano la principale sorgente di questi materiali, almeno da noi. Le diagnosi riportate sui certificati di morte sono una mescolanza dei quattro gradi di precisione diagnostica elencati sopra, con larga prevalenza dei gradi inferiori. Ne consegue che buona parte delle indagini statistiche in tema di tumori sono viziate in partenza da un errore, del quale è tuttavia possibile valutare la portata sulla base di studi che a tale problema hanno fornito un contributo di dati attraverso il confronto fra le diagnosi cliniche e anatomopatologiche per neoplasie maligne nel materiale autoptico (v. Squartini e Barola, 1965).
c) Confronto fra diagnosi cliniche e anatomopatologiche
Nel confronto fra diagnosi cliniche e autoptiche, per qualsiasi tipo di neoplasia maligna (X) si possono verificare cinque eventualità: (A) la diagnosi clinica di X è corretta, cioè verificata dall'autopsia; (B) manca una diagnosi clinica di neoplasia maligna; X è scoperto all' autopsia; (C) una neoplasia maligna è diagnosticata clinicamente, ma la sede è sbagliata (ad esempio si scambiano le metastasi nel fegato di un cancro gastrico per neoplasia primitiva del fegato) o non specificata (ad esempio neoplasia dell'addome), e X è scoperto all'autopsia; (D) la diagnosi clinica di X è errata (è l'eventualità opposta a quella che precede: ad esempio si scambia una neoplasia primitiva del fegato per metastasi di cancro gastrico e si diagnostica perciò un cancro dello stomaco inesistente), l'autopsia rivela un tumore maligno primitivo diverso da quello clinico; (E) la diagnosi clinica di X è errata, nessun tumore maligno di alcun tipo è rivelato dall'autopsia (v. Willis, 19674). A scopo indicativo nella tab. II sono riportati i dati di uno studio personale sugli errori diagnostici rilevati da 695 autopsie consecutive effettuate a Perugia su soggetti portatori di neoplasie maligne o supposti tali (v. Squartini e Barola, 1965).
d) Errore diagnostico reale e apparente
Quindi, il numero reale delle neoplasie maligne riscontrate al tavolo anatomico (X) è espresso da A + B + C (dove B + C sono gli errori diagnostici per difetto), mentre il numero totale di diagnosi cliniche di X è dato da A + D + E (dove D + E sono gli errori diagnostici per eccesso). La lettura della tab. II suggerisce alcune considerazioni interessanti. Le diagnosi cliniche esatte per il totale delle neoplasie sono pari a 70,1%. I casi falsi negativi (29,9%) sono compensati in parte dai falsi positivi (17,8%), ma rimane uno scarto negativo pari a 12,1%. Analizzando i singoli tipi tumorali si osservano notevoli variazioni di frequenza nelle diagnosi esatte, che rendono particolarmente attendibili i dati relativi a certe neoplasie (mammella, cute, utero, ovaio, leucemie e linfomi) e largamente inattendibili i dati riguardanti altre neoplasie per le quali la diagnosi esatta si rivela difficile (pancreas, prostata, fegato, colecisti, rene), sebbene un certo compenso agli errori sia fornito in ogni caso dalle diagnosi false positive. Numerosi tumori infine sono in posizione intermedia con una frequenza di diagnosi esatte oscillante fra il 65 e il 70%.

e) Effetto sulle indagini di frequenza e di distribuzione
Il compenso di alcuni errori diagnostici a opera di altri errori diagnostici introduce nel materiale clinico un fattore involontario di correzione che può rivelarsi nello stesso tempo utile e dannoso sotto il profilo statistico: è indubbiamente utile quando i casi registrati nei certificati di morte vengono usati per stabilire le frequenze dei vari tipi di tumore, mentre è invece dannoso quando si intende procedere a ulteriori analisi sulla distribuzione dei tumori, considerando l'età, il sesso, il tipo etnico, la residenza, la condizione sociale, l'attività professionale, ecc., dei soggetti portatori. Infatti, in tal caso, non è tanto grave la mancata inclusione nel gruppo considerato di soggetti portatori del tumore che si studia (B + C), quanto è grave l'inclusione al loro posto di altri soggetti portatori di tumori di altre sedi o anche privi di tumore (D + E). L'attendibilità delle indagini di questo tipo sarà quindi tanto minore quanto più grande risulterà il rapporto D + E/A. Nel materiale illustrato in tab. II tale rapporto è di 1 : 4 per la serie intera. Ciò significa che per ogni quattro casi corretti uno è sbagliato. Naturalmente il rapporto varia molto da tumore a tumore, passando da 2 : 1 per le neoplasie del fegato a 1 : 21 per quelle dell'utero.
Questi dati, largamente concordanti con altri riferiti (v. Willis, 1948; v. Münck, 1952; v. Locatelli, 1956), si riassumono in breve nelle seguenti proposizioni. Vi sono vari tipi di tumore per i quali le indagini statistiche, sia di frequenza che di distribuzione, svolte sui materiali di uso più comune (certificati di morte), comportano un errore assai piccolo, trascurabile o addirittura nullo. Vi sono altri tumori idonei solamente per indagini di frequenza, o di distribuzione, e altri ancora che condurrebbero a risultati inattendibili in ogni caso. L'errore diagnostico apparente è sempre più piccolo di quello reale, a causa delle diagnosi cliniche di tumore false positive che compensano in varia misura i casi di tumore reale clinicamente non diagnosticati. La misura dell'errore diagnostico apparente è quasi sempre di segno negativo. I casi clinici falsi positivi, che favoriscono le indagini di frequenza, ostacolano ovviamente quelle di distribuzione, fino a renderle del tutto inattendibili quando la proporzione fra casi falsi e veri superi i limiti tollerabili.
f) Variazioni in rapporto al sesso, all'età e al periodo considerati
Nelle femmine l'errore diagnostico apparente è minore che nei maschi, anche dopo esclusione dei tumori del sesso, per i quali la diagnosi è più facile. Ciò non dipende da un maggior numero di diagnosi esatte nel sesso femminile, ma al contrario da un maggior numero di false diagnosi di tumore. La frequenza di diagnosi false positive aumenta fortemente con l'età dei soggetti causando una riduzione progressiva dell'errore diagnostico apparente, nonostante il contemporaneo aumento di quello reale. L'aumento nella frequenza di diagnosi esatte verificatosi nel tempo in conseguenza del miglioramento delle condizioni diagnostiche generali è in pratica annullato dalla concomitante diminuzione, per gli stessi motivi, delle diagnosi false positive (v. Squartini e Barola, 1965).
Chi si interessa a problemi di statistica oncologica e usa le cartelle cliniche o i certificati di morte come materiali può trarre da questa breve elencazione di fatti utili spunti di meditazione. E tuttavia auspicabile che indagini dello stesso tipo vengano ripetute su altri materiali e con criteri standard. Solo in tal modo, infatti, si potrà raggiungere una somma di casi sufficientemente ampia e rappresentativa, dalla quale sia lecito trarre non solo indicazioni ma precisi indici di correzione, direttamente applicabili allo studio dei problemi.
4. Predisposizione genetica
L'esistenza di una predisposizione genetica al cancro, in certi casi, è dimostrata da numerosi esempi e non può essere oggi messa in dubbio. Il quesito da porre è se quegli esempi di suscettibilità ereditaria allo sviluppo dei tumori siano solo curiosità scientifiche, per quanto interessanti, o se non siano invece segnali intimamente connessi con l'origine del cancro in generale. E la risposta è difficile, quanto del resto lo è un'illustrazione sintetica e chiara del problema.
a) Suscettibilità cromosomica al cancro
La suscettibilità genetica al cancro può essere classificata come cromosomica, mendeliana recessiva e mendeliana dominante (v. Knudson, 1973). Numerose anomalie cromosomiche predispongono notoriamente allo sviluppo dei tumori. Nella sindrome di Down, o mongolismo, dovuta a un'anomalia cromosomica con trisomia 21 (cioè caratterizzata da un extracromosoma nella coppia cromosomica 21), o con trisomia 21-22, vi è un'incidenza di leucemia valutata 10-20 volte maggiore di quella dei soggetti normali della stessa età (v. Miller, 1970). Una maggiore incidenza di leucemia si osserva anche in altre trisomie, come nella sindrome di Klinefelter, caratterizzata da ipogonadismo (v. Sohn e Boggs, 1974), la quale predispone pure al cancro mammario (v. Harnden e altri, 1971). La trisomia 18 è stata trovata associata a tumore di Wilms, un tumore maligno embrionale del rene (v. Bove e altri, 1969). La sindrone da delezione D, caratterizzata da delezione del braccio lungo del cromosoma 13, si associa spesso a retinoblastoma (v. Orye e altri, 1974). Il cromosoma Filadelfia, un piccolo cromosoma anomalo originato per delezione o traslocazione di parte del cromosoma 21, si associa molto spesso a leucemia mieloide cronica (v. Nowell e Hungerford, 1961; v. Harnden, 1977).
L'eventualità che tutti i cancri possano essere associati con delezioni o anomalie cromosomiche sembra assai remota. Piccole delezioni potrebbero comunque essere difficili da dimostrare, per cui sarebbe anche difficile verificare sperimentalmente una simile ipotesi. Tuttavia le moderne tecniche in uso per l'identificazione di singole bande cromosomiche potranno facilitare tali studi e ci si può perciò aspettare che nuovi dati chiariscano tale argomento nel prossimo futuro (v. Knudson, 1973).
b) Suscettibilità mendeliana recessiva
Varie anomalie e sindromi congenite ereditate come carattere mendeliano autosomico recessivo sono pure associate con particolare frequenza ai tumori dell'uomo. La sindrome forse più nota in questo senso è lo Xeroderma pigmentosum, un'anomalia cutanea causante spiccata sensibilità alla luce del sole, caratterizzata da zone di atrofia e da aree isolate e confluenti di cheratosi più o meno pigmentate. Su questa base si originano spesso e presto nel corso della vita carcinomi cutanei a cellule basali, o meno frequentemente a cellule squamose, o melanomi. La malattia si osserva all'incirca in un soggetto ogni 250.000. Il meccanismo causale dei cancri in questa condizione è ormai in parte conosciuto. Si tratta di un errore congenito del metabolismo in cui un enzima necessario per la riparazione dei danni indotti dalla luce ultravioletta è difettivo (v. neoplasie: Oncologia sperimentale). L'enzima in questione è implicato nelle fasi precoci della riparazione che conduce alla escissione di dimeri di timina (v. Cleaver. 1968; v. Robbins e altri, 1974).
L'albinismo, un'anomalia ereditaria dovuta a un gene mendeliano recessivo, opposta ma in certo senso comparabile con lo Xeroderma pigmentosum, è pure causa di cancri cutanei nei portatori. Gli albini sono uno su 10.000 nella popolazione europea ma molto più numerosi presso altri gruppi etnici, come gli indiani Cuna nei Caraibi. Essi mostrano una grande riduzione della pigmentazione degli occhi, dei capelli e della cute, che è causa di particolare sensibilità alla luce del sole. Se non protetti da vestiti e lenti affumicate, tali soggetti sviluppano perciò giovanissimi una cheratosi attinica maligna e spesso finiscono per sviluppare un cancro a cellule squamose della cute. Il meccanismo della cancerogenesi in questo caso non è noto, ma una mutazione genica vi è sicuramente coinvolta (v. Heston, 1976).
Varie malattie con difetti immunitari, con o senza anomalie del timo, che si trasmettono in genere come caratteri recessivi legati al sesso, predispongono particolarmente allo sviluppo di leucemie, linfomi o altri tumori, se i soggetti non muoiono prima per malattie infiammatorie intercorrenti dovute al deficit immunitario. È questo il caso della agammaglobulinemia di Bruton, della sindrome di Di George (aplasia timica), della sindrome di Wiscott-Aldrich (depressione selettiva delle immunoglobuline M e della immunità cellulare), della sindrome di Louis-Bar o atassiateleangectasia (ipoplasia timica marcata con linfopenia e deficit di immunoglobuline A ed E; v. Smith, 19777). In questi casi, comunque, il meccanismo genetico favorente la tumorigenesi sembra da ricercare almeno in parte nella depressione del sistema immunitario causata dall'assenza o dall'ipoplasia degli organi o degli stipiti cellulari immunocompetenti. Sui rapporti fra depressione immunitaria e cancro si avrà motivo di tornare appresso.
c) Suscettibilità mendeliana dominante o familiare
Le neoplasie e le sindromi neoplastiche che si trasmettono ereditariamente come caratteri mendeliani dominanti sono anche familiari perché appunto emergenti in più membri della stessa famiglia. Queste forniscono forse la più diretta evidenza che le modificazioni genetiche possono avere importanza nell'origine del cancro. In tali casi il gene responsabile è altamente penetrante per cui il tumore diventa una manifestazione fenotipica comune. Vi sono casi di tumori a sede determinata come unica manifestazione della sindrome e casi di lesioni multiple coesistenti, a differenti sedi, neoplastiche e non neoplastiche.
Un esempio del primo tipo è dato dal retinoblastoma, un tumore retinico di tipo embrionale. Non tutti i casi di retinoblastoma, specie monolaterale, sono ereditari. Ma tutti gli individui affetti bilateralmente sono portatori del gene dominante che causa il retinoblastoma, per cui la probabilità che la loro prole sia portatrice del gene e affetta da retinoblastoma è del 50% (v. Schappert-Kimmijser e altri, 1966). Le persone portatrici del gene hanno una probabilità del 95% di sviluppare il retinoblastoma, mentre per quelle che non lo possiedono le probabilità sono meno di una su 20.000 individui (v. Knudson, 1971).
Sebbene con frequenza assai minore del retinoblastoma, numerosi altri tumori embrionali, quali il neuroblastoma e il tumore di Wilms del rene o nefroblastoma, sono stati osservati in più membri della stessa famiglia. Tuttavia non è possibile affermare che la diversità di frequenza rispetto al retinoblastoma rispecchi una reale differenza di comportamento: infatti il retinoblastoma è eccezionale non solo come esempio di neoplasia trasmissibile quale carattere ereditario dominante, ma anche come esempio di tumore maligno curabile. Il risultato è quello di un gran numero di soggetti affetti, curati, che sopravvivono e sono così in grado di produrre una prole, affetta, il cui studio ha permesso appunto di chiarire la trasmissione della malattia come carattere ereditario dominante. Le possibilità di sopravvivenza per gli altri tumori embrionali sono invece molto minori (v. Knudson, 1973).
Altri tumori a carattere familiare, per i quali si suppone o si è dimostrato un modello di trasmissione ereditaria dominante, sono il feocromocitoma (del sistema nervoso simpatico), qualche forma di melanoma, le adenomatosi endocrine multiple (tumori interessanti contemporaneamente due o più ghiandole endocrine: ipofisi, paratiroidi, tiroide, pancreas insulare, surrene; o tumori di stipiti cellulari endocrini: carcinoide bronchiale e intestinale) e la poliposi del colon, che pure rappresenta un modello fra i più noti (v. Knudson, 1973).
L'aspetto più importante della poliposi ereditaria multipla del colon è che quasi ogni soggetto che ne è portatore se non viene curato muore di carcinoma del colon prima dei 50 anni. Vi sono varie forme di poliposi intestinale, tutte caratterizzate da marcata predisposizione al carcinoma (v. McConnell, 1966). L'età media di morte nei casi sottoposti a trattamento terapeutico è sui 40 anni, cioè molto più bassa di quella per carcinoma del colon nella popolazione generale, che si aggira sui 70 anni (v. Veale, 1965). La frequenza della sindrome è fortunatamente bassa (dell'ordine di 1 caso su 6.850 individui). I polipi rappresentano lesioni iperplastiche preneoplastiche da cui i carcinomi prendono origine per progressione.
d) Predisposizione genetica ai cancri più comuni
Tutti gli esempi e i casi di cui si è fin qui parlato, di estrema importanza sul piano speculativo per una comprensione dei rapporti fra eredità e cancro, ne hanno poca in pratica, poiché la frequenza dei tumori considerati è nel complesso bassa. Particolare interesse rivestono perciò gli studi genetici volti a stabilire l'influenza dell'eredità sullo sviluppo dei cancri più comuni nella popolazione generale (v. Mulvihill e altri, 1977).
La maggior parte degli studi genetici sul cancro della mammella ha condotto alla conclusione che nelle parenti di pazienti affette da cancro mammario la frequenza di tali neoplasie è da due a tre volte superiore a quella dei controlli. Il legame fra fattore ereditario e alcuni cancri mammari emerge più chiaramente se si opera una distinzione dei cancri della mammella in premenopausali (spesso associati con lesioni mammarie preneoplastiche benigne, con tumori tiroidei e con eccessiva produzione di estrogeni ovarici) e postmenopausali (più spesso associati invece con ipertensione, obesità, diabete mellito e iperfunzione surrenalica). Infatti, nel primo tipo, le parenti mostrano un significativo eccesso di cancri della mammella in confronto ai controlli, che nel secondo tipo manca (v. Anderson, 1971 e 1972). La predisposizione genetica è inoltre più evidente nei casi di cancro bilaterale della mammella e di cancro della mammella in età giovane.
Per i cancri dell'utero, che si sviluppano in due distinte sedi con profili anatomo-clinici indipendenti, quello della cervice mostra ben pochi segni di essere correlato a fattori ereditari (v. Rotkin, 1966) mentre quello dell'endometrio è decisamente in rapporto con una predisposizione ereditaria, come può desumersi dalla sua frequente familiarità e dalla sua rimarchevole incidenza nelle parenti delle donne portatrici (v. Lynch, 1967).
Per i cancri del polmone, dell'intestino, dello stomaco, condizionati da fattori ambientali, le indagini genetiche sono meno indicative. Comunque un elevato numero di casi, più o meno rilevante e significativo, nei parenti di pazienti portatori rispetto ai controlli opportunamente selezionati, è stato più volte riferito. Analoghi risultati sono stati forniti per il cancro della prostata (v. Knudson, 1973; v. Heston, 1976).
e) Indagini sui gemelli monocori
Un tentativo classico per l'identificazione di fattori genetici nei tumori, è quello applicato allo studio delle leucemie e dei linfomi attraverso il confronto fra coppie di gemelli identici (monozigoti) e non identici (dizigoti). I primi derivano da uno stesso uovo fecondato, per cui se un fattore genetico causante leucemia è presente nei gameti, questo dovrebbe manifestarsi con la sua azione in entrambi i gemelli. I secondi derivano invece da due diverse uova fecondate, per cui il loro patrimonio cromosomico è diverso e la possibilità dell'esistenza di un eventuale fattore genetico comune per la leucemia è remota. Nei gemelli monozigoti si è trovato un tasso di concordanza per la leucemia infantile del 25%, mentre solo tre casi di concordanza risultano descritti nella letteratura per i gemelli dizigoti, nessuno dei quali ben documentato (v. MacMahon e Levy, 1964). Se pertanto un gemello monocore cade affetto da leucemia il fratello ha una probabilità su 4 di sviluppare una leucemia, che spesso appare nel giro di settimane o mesi. Nonostante che questi dati sui gemelli suggeriscano fortemente l'azione di fattori genetici nella leucemia infantile, gli studi sui parenti dei portatori di leucemie o linfomi non hanno fornito risultati altrettanto evidenti o comunque particolarmente indicativi in questo senso.
f) Conclusioni
Il problema dei rapporti fra eredità e cancro nell'uomo rimane dunque aperto (v. Mulvihill e altri, 1977). Le conoscenze attuali in questo campo sono a un punto critico, per cui non mette conto soffermarsi su modelli e ipotesi formulati per spiegare il cancro umano come una malattia da mutazione somatica (v. Knudson, 1977; v. Strong, 1977; v. Mulvihill e altri, 1977). Oggi, tenendo presente la generalità dei casi e non solo le eccezioni, si può soltanto anticipare che una certa predisposizione ereditaria ai tumori sembra esistere e avere probabilmente importanza, anche se essa non è in grado di produrre da sé l'evento (neoplasia) senza l'intervento di fattori causali.
5. Cause del cancro umano
Uno dei mutamenti di opinione più significativi della medicina moderna è quello intervenuto nel suo atteggiamento riguardo alle origini del cancro umano. Prima della seconda guerra mondiale, i cancri erano considerati malattie spontanee, inevitabilmente connessi con l'età, per i quali vi era poco da fare da parte del medico oltre a formulare una diagnosi precoce che consentisse al chirurgo e al radiologo di curarli. Oggi è comune leggere che l'80 o il 90% di tutti i cancri hanno un'origine ambientale e sono dovuti a inquinamento industriale, ad abitudini innaturali, ai contenuti e alle preparazioni della dieta, ai medicamenti, ecc. (v. Doll, 1977).
In realtà si conoscevano anche nel passato tumori dovuti a cause ambientali, ma questi erano pochi per tipo come per numero di persone interessate e perciò potenzialmente controllabili. Erano noti alcuni tipi di ‛tumori professionali', di cui il più famoso e primo a essere studiato fu il cancro della vescica, che causava la morte fra i lavoratori delle fabbriche di coloranti a base di anilina (v. Case e altri, 1954). I cancri del volto e della cute scoperta, più frequenti nelle campagne, fra i marinai, ecc., erano attribuiti alle radiazioni solari. Alcuni cancri del labbro erano posti in relazione col fumo della pipa, altri del cavo orale con l'abitudine di masticare tabacco. Oggi, invece, per quasi tutti i tipi di cancro cominciano a conoscersi, in maniera più o meno approssimata, le cause o i gruppi di cause e non vi è tanto necessità di insistere sulla natura indotta dei tumori umani quanto di fornire un panorama ordinato e schematico delle loro principali cause.
Queste si possono dividere in: a) fattori professionali, cioè collegati a un tipo di lavoro; b) inquinamento atmosferico e ambientale (acqua, suolo), che prevalentemente coinvolge gli abitanti delle città e delle zone industriali; c) uso di erbicidi, pesticidi e conservanti, che riguardano l'attività agricola e l'industria alimentare; d) dieta, importante sotto molti aspetti che vanno dagli squilibri o carenze, alla composizione chimica, a quella fisica, agli additivi alimentari, al modo di preparazione e cottura dei cibi, alle eventuali sostanze inquinanti e al condizionamento sulla flora batterica intestinale; e) medicamenti, responsabili dei tumori iatrogeni; f) fumo di tabacco e altre abitudini nocive; g) parassiti; h) virus oncogeni. In questo ambito si ritiene oggi che vadano cercate le origini della maggior parte dei tumori umani, tenendo presente che tali fattori possono agire, anzi spesso agiscono associati. Il problema, come si vede, è di primaria importanza per i suoi riflessi nel campo della prevenzione tumorale, attraverso la diminuzione e il controllo del rischio oncogeno ambientale (v. Hiatt e altri, 1977).
a) Fattori cancerogeni professionali
Alla categoria dei fattori cancerogeni professionali appartiene ormai una lunga lista di agenti: le radiazioni ionizzanti fra cui il radon (minatori di uranio, spatofluorite, ematite: cancro del polmone), i raggi X, il radium (radiologi: cancro della cute; verniciatori di quadranti luminosi: tumori delle ossa) e i raggi ultravioletti (contadini, marinai: cancro della cute); gli idrocarburi policiclici aromatici contenuti nella fuliggine, nel catrame, nel petrolio (spazzacamini: cancro dello scroto; operai dei gasometri: cancro della cute; molti altri lavoratori dell'industria: cancro dei polmone); le naftilammine, la benzidina, il 4-amminodifenile (lavoratori di industrie coloranti, chimiche, della gomma: cancro della vescica); l'asbesto (lavoratori di materiali refrattari come l'amianto: tumori della pleura); l'arsenico (lavoratori delle concerie, delle fonderie, minatori d'oro: cancro della cute e del polmone); il bisclorometiletere (lavoratori di resine a scambio ionico: cancro del polmone); il benzolo (lavoratori di colle e vernici: leucemia); il cloruro di polivinile (lavoratori della plastica: angiosarcoma del fegato); il cromo, il nichel, l'olio isopropilico (lavoratori delle raffinerie e industrie relative: cancro dei seni nasali e del polmone) (v. Hiatt e altri, 1977).
La lista tende col tempo ad allungarsi, perché molte altre sostanze sono sospettate di rischio potenziale (v. Hueper e Conway, 1964). Ma è da rilevare che molti dei sopraricordati rischi oncogeni professionali sono da tempo sotto controllo o attraverso la cessazione delle attività industriali più rischiose o dell'uso dei materiali cancerogeni, o attraverso la modificazione dei procedimenti industriali in modo da ridurre l'esposizione degli operai addetti. Fatta eccezione forse per le radiazioni ultraviolette, il numero dei lavoratori esposti è inoltre limitato rispetto alla popolazione totale, per cui si è indotti a concludere che il rischio professionale può giustificare solo un numero molto limitato del totale dei cancri (v. Doll, 1977).
b) Inquinamento ambientale
L'inquinamento ambientale - atmosferico, del suolo e delle acque - potrebbe avere maggiore importanza poiché a esso si trova esposta non intenzionalmente la popolazione generale. Cancerogeni come gli idrocarburi policiclici aromatici prodotti dalla combustione del carbone, del legno, del petrolio a uso di riscaldamento, l'arsenico, presente fra l'altro nella carta delle sigarette, i pesticidi come il DDT, i composti N-nitrosi usati come erbicidi e presenti nell'aria, nel tabacco e nel cibo, l'asbesto, comunemente presente nell'atmosfera delle città industriali, il cloruro di polivinile, entrato in ogni casa con l'uso quotidiano dei recipienti in plastica, il fluoro e il cloro usati per la fluorazione e la clorazione delle acque e che nell'acqua producono composti non del tutto innocui, le radiazioni ionizzanti, in lento ma costante aumento nell'ambiente, possono dunque essere responsabili di cancri nella popolazione generale. Difficile anche qui stabilire però in quale proporzione essi contribuiscono al totale delle neoplasie dell'uomo (v. Hiatt e altri, 1977).
Alcuni studi quantitativi sembrerebbero limitare l'importanza di simili cancerogeni ambientali per la particolare esiguità delle dosi di esposizione. È stato calcolato che gli abitanti delle grandi città inglesi, a causa della combustione per riscaldamento domestico, sono esposti a circa un centesimo della dose di benzopirene inspirato dagli operai delle industrie del gas di carbone (v. Doll e altri, 1972). Per il cloruro di polivinile, è noto che la dose giornaliera assorbita nel cibo dai recipienti di plastica è circa 50 milioni di volte minore di quella ricevuta giornalmente dai primi lavoratori della plastica che erano realmente esposti al rischio di sviluppare angiosarcomi del fegato (v. Barnes, 1976). Anche l'asbesto atmosferico delle città è circa un decimilionesimo della concentrazione ritenuta accettabile nelle industrie del settore (v. Doll, 1977).
Queste misurazioni sembrerebbero ridurre il significato delle polluzioni industriali come causa primaria o importante di cancri nell'uomo. Ma rimane difficile stabilire quale proporzione sia in definitiva da attribuire ai rischi, modesti ma diffusi e per di più combinati, di questi cancerogeni.
c) Cancerogeni nella dieta
La dieta rappresenta una sorgente alternativa di fattori cancerogeni per l'uomo e, dopo essere stata ingiustamente trascurata, va oggi riscuotendo presso i ricercatori l'interesse che apparentemente merita. La dieta può intervenire nella cancerogenesi degli organi dell'apparato digerente o anche di altri apparati con vari meccanismi. Una carenza di proteine nella dieta è probabilmente, come già detto, la causa o una concausa della elevata frequenza di cirrosi e di cancri del fegato che si verificano nelle regioni dell'Africa e dell'Asia meridionale. La dieta dei paesi industrializzati occidentali, particolarmente ricca di grassi e povera di scorie indigeribili fibrose, potrebbe avere responsabilità notevoli per il cancro dell'intestino e di altri organi.
La composizione della flora batterica intestinale dipende infatti dalla dieta e in particolare dal contenuto in grassi della dieta, che stimolano l'incremento dei batteri anaerobi quali i clostridi lecitinasi-negativi. La quantità di grasso nella dieta determina anche la quantità di steroidi biliari (colesterolo e prodotti di degradazione dei sali biliari) che raggiungono il colon. A sua volta la flora intestinale è in grado di produrre metabolicamente dagli steroidi biliari cancerogeni chimici (v. Hill, Drasar e altri, 1971) ed estrogeni (v. Hill, Goddard e altri, 1971). I primi potrebbero causare il cancro dell'intestino, i secondi quello della mammella (v. Hill, 1977). Inoltre, le feci povere di scorie indigeribili sono meno voluminose, per cui il loro transito è più lento e il ristagno aumenta così il tempo di esposizione della mucosa intestinale ai cancerogeni in esse eventualmente contenuti.
Altri rischi oncogeni della dieta possono provenire dalle sostanze chimiche impiegate in agricoltura per la produzione degli alimenti (erbicidi, pesticidi, anticrittogamici), dagli additivi chimici, dai preservanti (per es. i nitriti), dai conservanti e soprattutto dai contaminanti biologici. Aspergillus flavus, un fungo comune che si sviluppa prontamente nel grano, nel granoturco, nelle nocciole, ecc., conservati in ambienti e in condizioni climatiche inadatte, produce una micotossina, l'aflatossina B1, che è uno dei più potenti cancerogeni epatici sperimentali oggi conosciuti, attivo in dose di microgrammi. Studi epidemiologici condotti presso le popolazioni dell'Africa e dell'Asia meridionale, che consumano larghe quantità di cereali mal conservati, hanno dimostrato una correlazione fra incidenza di epatocarcinoma e contenuto di aflatossina nella dieta in quelle zone (v. Alpert e altri, 1971). La cicasina, un costituente delle noci di cicade, è un altro cancerogeno naturale dotato di potenzialità neoplastica per l'uomo. Un certo numero di piante dell'Africa, del Sudamerica e del- l'Asia producono alcaloidi che potrebbero essere cancerogeni (v. Schoental, 1968). Da ciò derivano nuovi problemi pratici e di studio che appunto oggi si sta cercando di risolvere.
Un'ulteriore possibilità di azione cancerogena della dieta dipende dal modo di cottura delle vivande. Molti tipi di sostanze organiche scaldati a temperature di combustione producono catrami cancerogeni artificiali e ciò non è certo senza significato culinario. L'abitudine di arrostire, cuocere alla griglia, friggere, tostare e affumicare i cibi potrebbe produrre infatti piccole quantità di idrocarburi cancerogeni artificiali e la loro ingestione quotidiana potrebbe essere causa non marginale dei comuni cancri del tubo digerente. I grassi portati ad alte temperature, per esempio quelli ripetutamente usati per friggere, sembrerebbero essere i più pericolosi. Il benzopirene, uno fra i più comuni cancerogeni della serie degli idrocarburi aromatici, è stato ritrovato in quantità significative nella carne e nel pesce affumicati (v. Bailey e Dungal, 1958) e nella polvere di caffè tostato (v. Hueper e Payne, 1960).
d) Cancerogenesi iatrogena
I medicamenti compongono un'altra categoria di cancerogeni per l'uomo. La lista di medicine e trattamenti pericolosi è assai lunga e comprende i raggi X usati a scopo diagnostico e terapeutico e i mezzi di contrasto radiologici come il thorotrast (tumori in ogni sede, leucemie e linfomi), gli idrocarburi cancerogeni contenuti negli unguenti e nei lassativi (cancri della cute, dello stomaco, dell'intestino), gli agenti citostatici alchilanti (melphalan, ciclofosfammide: leucemia mieloide), il cloramfenicolo (leucemia mieloide), gli estrogeni (cancri della mammella, dell'utero), l'arsenico (cancri della cute, del polmone), la clornafazina (cancro della vescica), la fenacetina (cancro della pelvi renale), gli immunosoppressivi (linfomi e altre neoplasie), il fenilbutazone (leucemie) a cui se ne devono aggiungere altri potenziali, come suggerito dalla loro azione cancerogena sugli animali, quali l'isoniazide, la griseofulvina, il tannino, il tiouracile, i glicocorticoidi, i ciclammati, gli steroidi anabolizzanti, i derivati della rauwolfia (v. Schmähl e altri, 1977).
Vi sono anche esempi di cancerogenesi iatrogena nel campo della chirurgia, quali il sarcoma da corpo estraneo e da cicatrice (v. Ott, 1970), il cancro primitivo dello stomaco operato (v. Hilbe e altri, 1968), i tumori del colon conseguenti a ureterosigmoidostomia (v. Mueller e Thornbury, 1973) e i tumori cutanei in sede di cicatrici da vaccinazione antivaiolosa (v. Schmähl e altri, 1977). Il significato delle medicine e dei trattamenti medici nell'insorgenza dei cancri in generale può essere difficilmente valutato in termini quantitativi. L'impressione è che il rischio iatrogeno non possa fornire un contributo rilevante al rischio totale, sebbene in alcuni paesi esso possa essersi reso responsabile di percentuali discrete di cancri in particolari età (infanzia) o sedi del corpo (v. Doll, 1977).
e) Abitudini di vita e voluttuarie
Importanza non secondaria come cause di cancro umano hanno anche alcune abitudini voluttuarie o di vita condizionate dalla nostra civiltà, fra cui sono essenzialmente da considerare il fumo di tabacco, l'alcool e, nel sesso femminile, le abitudini sessuali, l'allattamento al seno, il numero dei figli e l'età al primo parto. Da studi epidemiologici prospettivi e retrospettivi, chimici, sperimentali e morfologici è emerso con indubitabile chiarezza il legame esistente fra fumo di tabacco e cancro del polmone. Relazioni analoghe sono state trovate anche fra fumo di tabacco e cancri del cavo orale, della laringe, dell'esofago e della vescica. I prodotti di combustione del tabacco contengono benzopirene e altre sostanze cancerogene. I forti fumatori sono notevolmente più inclini a sviluppare cancro del polmone dei non fumatori e mostrano all'autopsia lesioni premaligne più frequenti nell'albero bronchiale (v. U.S. Department of..., 1964). L'abuso di alcool è collegato con maggiori incidenze di cancri della bocca, faringe, laringe ed esofago, sebbene il meccanismo non sia noto (v. Willis, 19674).
Il cancro della cervice uterina è più frequente nelle donne che hanno molti partners, intensa attività sessuale, numerosi figli, probabilmente a causa di infezione venerea da virus erpetico che in tal modo contraggono (v. Kessler e altri, 1974). Infine, il pratico abbandono dell'allattamento al seno, la riduzione nel numero dei figli e soprattutto la primiparità tardiva o attempata, divenute abitudini di vita nei paesi occidentali, sono in stretto rapporto con l'incremento di cancri della mammella verificatosi durante questo secolo (v. Severi e Squartini, 1962; v. MacMahon e altri, 1973).
f) Parassiti e virus oncogeni
Chiudono questo vasto panorama di cause del cancro umano le infestioni da parassiti e le infezioni da virus oncogeni. Fra le prime sono da ricordare quelle da schistosomi, frequenti in Egitto, in Giappone e in altri paesi orientali, e da Clonorchis sinensis, frequente nella Cina meridionale. In entrambi i casi il meccanismo che conduce al cancro è quello dell'infiammazione cronica. Gli schistosomi penetrano allo stato larvale attraverso la cute, seguono il sistema venoso e raggiungono così la vescica o l'intestino crasso determinandovi infezioni croniche granulomatose e poi tumori; Clonorchis sinensis, attraverso la vena porta, passa dall'intestino al fegato, ove determina granulomi seguiti nel tempo dallo sviluppo di tumori.
Il capitolo dei virus oncogeni è il più recente fra quelli dei fattori che contribuiscono alla eziologia del cancro umano e tuttavia e gia ricco di dati significativi. Il linfoma di Burkitt (v., 1968) e il carcinoma giovanile del naso-faringe, noto con il nome di linfoepitelioma, si manifestano solo in individui infetti con virus di Epstein-Barr (v. Epstein e altri, 1964), un herpesvirus oncogeno contenente DNA diffuso nel genere umano, ove causa anche la mononucleosi infettiva (v. neoplasie: Oncologia sperimentale). Il DNA virale è presente in tutte le cellule tumorali, nelle quali determina l'espressione di neoantigeni virali. La produzione virale da parte delle cellule tumorali può essere attivata in laboratorio. È difficile pensare di ottenere in laboratorio prove migliori di queste per dimostrare la natura virale dei tumori nominati. L'unico tentativo non facile che rimane da fare è quello di prevenire la malattia con un vaccino specifico (v. Epstein, 1976).
Vi sono indicazioni, come si è già detto, che un herpesvirus di tipo II possa essere causalmente associato al cancro della cervice uterina (v. Rapp, 1974), ma tali indicazioni sono ancora incomplete e necessitano di conferma (v. Doll, 1977). La patologia sperimentale suggerisce inoltre fortemente che le leucemie (v. Gross, 1951) e il cancro della mammella (v. Bittner, 1936) possano avere un'ori- gine virale. Ma fin'ora nell'uomo sono state trovate solo prove tenui o indirette della presenza di virus causali in queste malattie. I virus oncogeni in questione sono RNA virus (oncornavirus) e una prova indiretta della loro presenza è il riscontro di un enzima virale, la DNA polimerasi RNA-dipendente o trascrittasi inversa, che rappresenta appunto un marcatore biochimico specifico degli oncornavirus quando si ritrovi associato con RNA ad alto peso molecolare (v. Spiegelman e altri, 1970 ; v. Schlom e Spiegelman, 1971).
Nel caso del cancro mammario umano le tenui prove accumulate nell'ultimo decennio sulla possibile presenza e attività di un virus causale possono sintetizzarsi come segue : a) sporadica dimostrazione di particelle B, l'espressione morfologica del virus dei tumori mammari murini, nel latte di donna (v. Moore e altri, 1971); b) presenza di trascrittasi inversa associata con RNA ad alto peso molecolare nel latte di donna (v. Schlom e altri, 1972); c) omologia fra RNA del virus murino e RNA citoplasmatico del cancro mammario umano negli studi di ibridazione molecolare competitiva (v. Spiegelman e altri, 1972; v. Vaidya e altri, 1974); d) anticorpi neutralizzanti il virus dei tumori mammari murini nel siero di donna (v. Charney e Moore, 1971); e) reazione immunologica crociata fra virus dei tumori mammari murini e cancro mammario umano nei test di ipersensibilità cellulare con leucociti in vitro (v. Black e altri, 1974 e 1976). Questi dati sono tuttavia ancora in attesa di un'interpretazione unitaria e sono ritenuti al momento insufficienti per sostenere un'eziologia virale del cancro della mammella nell'uomo (v. Squartini, I virus oncogeni..., 1977). Potrà forse gettar luce in futuro su questo e analoghi problemi di oncologia umana la recente acquisizione sperimentale che oncornavirus come quello dei tumori mammari del topo possono essere trasmessi anche per via genetica come provirus germinali (v. Bentvelzen e altri, 1970).
g) Prevenzione e screening delle sostanze cancerogene
Fra i numerosi fattori causali del cancro umano ricordati si possono verificare interazioni e sinergismi di azione che portano a potenziamento degli effetti. Alcune interazioni, come quella fra asbesto o radiazioni ionizzanti e fumo di tabacco per il cancro del polmone, e ancora quella fra alcool e fumo di tabacco per il cancro dell'esofago, sono state dimostrate.
Nel concludere è giusto sottolineare il grande compito che aspetta l'organizzazione sanitaria del futuro, impegnata a tradurre questo panorama di fatti in efficaci misure di prevenzione. Un problema preliminare a questo compito è quello di sperimentare test relativamente rapidi per valutare in futuro la pericolosità delle innumerevoli sostanze che la nostra società industriale riversa ogni anno nell'ambiente. Infatti, i tradizionali test di cancerogenicità nei piccoli animali di laboratorio sono troppo lunghi, della durata di 1-3 anni, e costosi. I test di cancerogenicità in vitro sono più brevi, ma presentano qualche difficoltà applicativa (v. Heidelberger, 1973 e 1977). Perciò recentemente l'interesse si è concentrato sul fatto che i cancerogeni noti sono in genere anche mutageni, cioè sostanze in grado di produrre mutazioni nel patrimonio genetico cellulare (v. Hollaender, 1971; v. Montesano e Bartsch, 1976). L'identificazione dei cancerogeni attraverso test di mutagenesi nei Batteri e nella drosofila (v. Vogel, 1977) potrebbe perciò, per la rapidità e la semplicità dei test, risolvere il problema almeno a livello dello screening più grossolano. In una eventuale scala di approfondimento del rischio oncogeno di sostanze sconosciute, il test di cancerogenesi in vitro potrebbe poi rappresentare il secondo livello e quello su animali di laboratorio il terzo, necessario per quei composti che richiedano un'attivazione metabolica (v. Hiatt e altri, 1977). Con questo armamentario si potrebbe in futuro controllare gli effetti sull'ambiente delle sostanze chimiche nocive e prevenire così forse in larga parte il cancro umano.
6. Immunodepressione e malignità
L'ipotesi che la risposta immunitaria rappresenti un efficace meccanismo di difesa contro le neoplasie ha avuto una grande influenza nelle ricerche sul cancro degli ultimi 20 anni. I lavori si contano a migliaia anche se i risultati sono fin qui contraddittori e scarsi. In particolare ha preso solido sviluppo il concetto che i meccanismi immuni possano prevenire lo sviluppo di tumori incipienti e, reciprocamente, che le deficienze immunitarie possano favorire lo sviluppo tumorale. Tale concetto ha trovato la sua formulazione più articolata e completa nella teoria generale della ‛sorveglianza immunologica' da parte di Burnet (v., 1963, 1964 e 1967).
a) L'ipotesi della ‛sorveglianza immunologica'
Il primo accenno all'ipotesi si trova in una considerazione del 1957, secondo la quale ‟non è affatto impossibile che piccoli accumuli di cellule tumorali possano svilupparsi e, a causa delle loro nuove potenzialità antigeniche, provocare un'efficace reazione immunitaria con regressione del tumore e nessun segno clinico della sua esistenza" (v. Burnet, 1957). Il termine sorveglianza immunologica fu usato la prima volta nel 1963 (v. Burnet, 1963) e la prima elaborazione compiuta dell'ipotesi fu illustrata l'anno seguente (v. Burnet, 1964). Il concetto della sorveglianza immunologica può essere riassunto come segue: ‟negli esseri viventi con lunga durata della vita, come la maggior parte dei vertebrati a sangue caldo, trasformazioni genetiche ereditabili devono essere comuni nelle cellule somatiche e una proporzione di queste rappresenta un passo verso la malignità. E quindi per una necessità dell'evoluzione che dovrebbe esservi qualche meccanismo per eliminare o inattivare tali cellule mutanti potenzialmente pericolose e viene postulato che questo meccanismo sia di carattere immunologico" (v. Burnet, Immunological surveillance..., 1970).
L'ipotesi poggia dunque sui presupposti che quando dette cellule aberranti con potenzialità proliferativa compaiono nell'organismo queste possiedano nuove determinanti antigeniche e che quando una quantità sufficiente di nuovo antigene è stata prodotta questa dia inizio a una risposta immunologica dipendente dal timo, l'organo preposto all'immunità cellulare, la quale porterebbe alla eliminazione delle cellule aberranti, all'incirca nello stesso modo in cui viene rigettato un trapianto (v. Burnet, The concept of..., 1970). Nella definizione del concetto di sorveglianza immunologica sono perciò impliciti i due assiomi fondamentali e precisamente: a) che le cellule tumorali possiedano antigeni diversi; b) che tali antigeni possano essere riconosciuti come estranei provocando una risposta immune basata sugli immunociti timo-dipendenti (v. Burnet, Evaluation of..., 1970).
b) Implicazioni cliniche
Le implicazioni cliniche di un concetto così formulato sono ovviamente rilevanti. Indipendentemente dalle cause che, nelle varie circostanze, producono i tumori, questi infatti, se una sorveglianza immunologica esiste, avrebbero maggiori o minori possibilità di svilupparsi come malattie a seconda dello stato immunologico del soggetto portatore (v. Burnet, Immunological surveillance, 1970).
Le principali conseguenze cliniche prevedibili sulla base dell'ipotesi presentata sono infatti le seguenti: a) il cancro dovrebbe avere più possibilità di manifestarsi in quei periodi della vita in cui l'efficacia del sistema immunitario è bassa, vale a dire il periodo perinatale e l'età avanzata; b) le malattie genetiche che producono difetti immunologici dovrebbero essere associate con un'eccessiva frequenza di neoplasie maligne; c) gli agenti immunosoppressivi fisici (raggi X) e chimici (medicamenti) dovrebbero aumentare le probabilità di neoplasie; d) quando tessuti che comunemente soggiacciono allo sviluppo di tumori vengono esaminati per individuarvi la presenza di foci istologici di malignità in individui non selezionati (per es. nel materiale autoptico di soggetti senza segni clinici di neoplasia), tali foci dovrebbero essere trovati in numero maggiore di quanto prevedibile dalla specifica incidenza delle neoplasie cliniche a quel livello in base all'età e al sesso considerati; e) anche se la sorveglianza immunologica dovrebbe essere inefficace per tumori che hanno raggiunto la soglia clinica, ci si potrebbe in rari casi aspettare una regressione spontanea di cancri già diagnosticati; f) essendo postulato dell'ipotesi che la risposta immune alle cellule maligne è fornita dai linfociti timo-dipendenti, ci si dovrebbe aspettare che la prognosi dopo rimozione chirurgica dei tumori sia migliore per i tumori mostranti adiacenti accumuli e infiltrazione di linfociti, che non per quelli sprovvisti di tale reazione (v. Burnet, Immunological surveillance..., 1970).
c) Difetti immunologici e tumori
Un confronto fra queste previsioni e l'osservazione anatomoclinica offre svariati punti di sostegno all'ipotesi della sorveglianza immunologica contro i tumori maligni. La malattia neoplastica è caratteristicamente collegata all'età e per la maggior parte dei tipi di cancro le curve di frequenza crescono in progressione logaritmica secondo l'età. Questa concentrazione di cancri nei gruppi di età più avanzate può essere collegata con il ben noto declino della risposta immunitaria conseguente all'età. L'altro periodo di inefficienza immunologica è quello fetale e peri- natale; e forse a questo è collegato il fatto che i tumori maligni dei bambini, per quanto molto meno frequenti di quelli dei soggetti adulti e anziani, mostrano una curva rapidamente decrescente dalla nascita fino al periodo postpuberale (v. Squartini e Bolis, 1966).
L'associazione non casuale dei tumori maligni con difetti genetici del sistema immunitario è già stata qui ricordata (v. sopra, cap. 4). Le rassegne più estese in questo settore di ricerca, aperto circa un quarto di secolo fa da Bruton (v., 1952) con la scoperta dell'agammaglobulinemia nei bambini (v. immunologia e immunopatologia: Immunologia generale), sottolineano la particolare propensione dei pazienti con difetti immunologici congeniti a sviluppare neoplasie maligne (v. Bergsma e Good, 1968), più evidente, come è comprensibile, per quelle condizioni che consentono una maggiore sopravvivenza, essendo più lentamente di altre letali, come l'atassia-teleangectasia e la sindrome di Wiskott-Aldrich (v. Dent e altri, 1968; v. Kersey e altri, 1973).
L'influenza favorente dei trattamenti con agenti immunosoppressivi sullo sviluppo di tumori maligni ha pure qualche solido punto di riferimento nell' osservazione clinica. Il trapianto renale, un recente progresso della nefrologia, ha il suo rischio iatrogeno, poiché richiede per attecchire un intenso e continuo trattamento con immunosoppressivi. Nei soggetti sottoposti a trapianto, ormai molto numerosi, sono stati ripetutamente descritti tumori maligni a prevalente tipo di linfomi (v. Dent e altri, 1968; v. Kersey e altri, 1973). Le irradiazioni con raggi X, che rappresentano un potente mezzo immunosoppressivo, ma nello stesso tempo anche mutageno, usate a scopo terapeutico, aumentano di 10-15 volte il rischio di neoplasie nell'uomo (v. Doll, 1963).
d) Regressione spontanea dei tumori
Le indagini istologiche su materiali autoptici e bioptici alla ricerca di cancri occulti, microscopici, in soggetti clinicamente liberi da neoplasie, hanno ripetutamente segnalato la frequenza, del tutto incoordinata con l'aspettazione, di foci microscopici di carcinoma nella tiroide, nella prostata, nella mammella, nel collo dell'utero, o di neuroblastoma nel surrene, molti dei quali evidentemente non hanno alcuna chance di raggiungere la soglia clinica, forse per regressione spontanea o per controllo efficace a livello microscopico.
‟La regressione spontanea di un cancro già manifesto è così rara che essa è divenuta quasi il miracolo tipo per giustificare una canonizzazione moderna" (v. Burnet, Immunological surveillance..., 1970). Tali casi comunque senza dubbio si manifestano e sono ormai numerosi quelli riferiti nella letteratura medica, ben documentati e competentemente diagnosticati su base clinica e istologica (v. AA. VV., Spontaneous regression..., 1976). In un'esauriente rassegna sull'argomento Everson (v., 1964) e poi Everson e Cole (v., 1966) e Cole (v., 1976) ne hanno collezionati 176 e uno dei fatti emergenti in tale raccolta è che più della metà dei casi, ben 98, appartengono a quattro tipi tumorali: l'ipernefroma, il neuroblastoma, il melanoma e il coriocarcinoma. Si tratta di tumori di soggetti relativamente giovani per i quali, con eccezione del neuroblastoma - un tumore embrionale che mostra talora la capacità di maturare, trasformandosi così nel corso dell'evoluzione da maligno in benigno (v. Willis, 1962; v. Evans e altri, 1976) - l'ipotesi di una regressione su base immunologica appare fondata e senza alternative razionali (v. Burnet, Immunological surveillance..., 1970). A questi si può aggiungere il linfoma di Burkitt, un tumore da virus dei bambini africani, già ricordato, le cui cellule possiedono neoantigeni virali e che talora è stato visto regredire (v. David e Burkitt, 1968; v. Ziegler, 1976).
Un notevole numero di contributi anatomopatologici ha infine dimostrato il significato prognostico favorevole degli infiltrati linfatici peritumorali e delle iperplasie linfatiche o reticolari reattive nei linfonodi prossimi alle sedi tumorali (v. Black e altri, 1955; v. Fisher e altri, 1975; v. Ioachim, 1976).
e) Azione immunosoppressiva dei cancerogeni
Un altro dato, che può avere rilievo per eventuali applicazioni eziologiche del concetto esposto, è quello della immunodepressione prodotta dai fattori oncogeni sia chimici, sia fisici e virali, ben dimostrato dalle ricerche sperimentali. Così si è persino postulato che i cancerogeni agiscano come tali in quanto agenti immunosoppressivi (v. Burnet, 1967).
Ma le numerose ricerche hanno chiarito che l'azione diretta del cancerogeno sul sistema immunitario dell'ospite è un fattore, sebbene necessario, però non sufficiente per l'effetto oncogeno e che questo può essere dissociato da quello immunodepressivo del cancerogeno, diminuendo quindi l'interesse per questa associazione (v. Stutman, 1973).
f) Critiche sperimentali all'ipotesi e conclusioni
Per quanto il concetto della sorveglianza immunologica dei tumori sembri ben fondato, a esso negli anni più recenti sono state mosse varie critiche sulla base di dati sperimentali (v. Prehn, 1972 e 1974; v. Stutman, 1975). Le principali obiezioni sono le seguenti: a) non tutti i tumori sono antigenici; b) i tumori iniziali, cosiddetti in situ perché ancora non hanno invaso i tessuti vicini, non sono riconosciuti immunologicamente dall'ospite; c) nella maggior parte dei sistemi sperimentali il meccanismo di difesa immunitaria, che pure esiste, è inefficace e tardivo piuttosto che rappresentare un meccanismo di sorveglianza contro i tumori incipienti; d) i tumori sperimentali da cancerogeni chimici e virali, nei quali i principi della sorveglianza sembrano effettivi, sono abnormemente immunogenici e possono rappresentare artefatti di laboratorio i quali magari nulla hanno a che vedere con la malattia neoplastica spontanea o naturale; e) i tumori che più comunemente insorgono in soggetti con difetti immunitari congeniti o sottoposti a trattamenti con immunosoppressivi sono in genere linfomi, cioè tumori delle cellule del sistema immunitario (v. Stutman, 1977).
Tuttavia, sebbene i punti cardinali dell'ipotesi sulla sorveglianza immunitaria siano stati in tal modo posti in discussione dagli esperimenti condotti su animali (v. Stutman, 1975), l'attrazione di questa teoria è rimasta pressoché immodificata ed essa ha continuato a essere considerata più come un dogma che come un'ipotesi di lavoro, anche perché le sue implicazioni cliniche appaiono tuttora sostanzialmente corrette e rispettate. Comunque, il fatto più importante, che l'ipotesi stessa ha contribuito a controllare, dimostrare e sottolineare, è che un qualche meccanismo generale di difesa immunitaria esiste nella biologia del cancro. Indipendentemente dal suo significato nella sorveglianza contro i tumori incipienti, è auspicabile che tale meccanismo possa essere incrementato al fine di realizzare concrete possibilità di immunoterapia (v. Prehn, 1974). Lungo questa linea convergono le innumerevoli ricerche e i problemi attuali in tema di immunologia dei tumori.
7. Storia naturale della malattia neoplastica nell'uomo
Tra il momento in cui, silenziosamente, la malattia neoplastica comincia e quello in cui ci si accorge della sua presenza intercorre un periodo di tempo più o meno lungo che si designa come periodo di latenza o tempo di induzione e si definisce come ‟il periodo che passa fra la prima esposizione al cancerogeno e la comparsa del tumore" (v. Steiner, 1953).
a) Fasi della malattia neoplastica
Durante il periodo di latenza si verificano in realtà più fenomeni. Infatti, alla prima applicazione di un cancero- geno non seguono spesso per qualche tempo modificazioni apprezzabili a livello istologico, sebbene modificazioni dei caratteri biologici, chimici e ultrastrutturali delle cellule costituenti il tessuto possano realizzarsi anche in maniera istantanea o in brevissimo tempo. Dopo un certo periodo cominciano in genere a manifestarsi lesioni chiamate preneoplastiche, perché con frequenza precedono lo sviluppo di un carcinoma. Spesso tali lesioni non mostrano alcun attributo di malignità attuale, ma l'analisi del loro comportamento biologico per mezzo dei trapianti consente di dimostrarne negli animali la natura precancerosa (v. DeOme e altri, 1959), È nell'ambito di queste lesioni che si realizza di solito la trasformazione maligna (v. Foulds, The histologic analysis... II, 1956; v. Squartini e Rossi, 1962; v. Squartini e Severi, 1964). Quando la trasformazione maligna si è verificata, inizia lo sviluppo di una popolazione cellulare autonoma che però può essere apprezzata clinicamente soltanto quando, per aver superato certe dimensioni, essa diviene palpabile o comunque valutabile attraverso la semeiologia fisica, radiologica, scintigrafica o la sintomatologia soggettiva e obiettiva (v. Severi e Squartini, 1955).
Corollari di quanto detto sopra sono che la malattia neoplastica nell'uomo ha in genere una lunga durata, talora di parecchi anni, e che la sua fase clinica è come la punta di un iceberg, per la maggior parte sotto il livello clinico e perciò ben poco conosciuta.
b) Il modello del cancro mammario
Un esempio che particolarmente bene serve a illustrare le fasi della malattia neoplastica è fornito dal cancro della mammella, il più frequente nel sesso femminile e uno dei più frequenti in assoluto, particolarmente studiato in questo secolo appunto per la sua frequenza, per la sua gravità, per la sua accessibilità e anche per il fatto che dispone di uno dei modelli sperimentali meglio conosciuti (v. Squartini, 1966).
Nel modello sperimentale, come nel cancro della mammella umana, un pieno sviluppo dell'albero ghiandolare mammario è richiesto prima che compaiano alterazioni apprezzabili le quali condurranno poi alla formazione di un tumore. La mammogenesi può essere definita quindi come la prima tappa della cancerogenesi mammaria. La mammogenesi procede dalle gemme; ciascuna gemma sviluppa dei dotti, i dotti terminali sviluppano gli alveoli e quando la proliferazione alveolare è avanzata compaiono i lobuli mammari. Al termine dello sviluppo, l'albero ghiandolare della mammella, visualizzato nelle tre dimensioni, somiglia quindi molto a un albero con i suoi rami (i dotti) e le foglie o i grappoli di frutta (i lobuli e gli acini). Queste strutture sono strettamente dipendenti dagli ormoni per il loro sviluppo e per il loro mantenimento.
La seconda tappa nella cancerogenesi mammaria sperimentale e umana è rappresentata di solito da lesioni iperplastiche, preneoplastiche alveolari, ducturari o dottali. La lesione preneoplastica più nota nel modello murino è rappresentata dai noduli di iperplasia alveolare. Questi somigliano ai lobuli normali, ma differiscono da essi perché dotati di maggiore autonomia, cioè meno dipendenti dagli ormoni dei lobuli a loro simili (v. Squartini, 1959; v. DeOme e altri, 1962). Recenti studi morfologici e trapianti hanno mostrato che anche nella mammella umana vi sono lesioni, come i lobuli atipici e i lobuli postmenopausali persistenti, da interpretare come possibili lesioni preneoplastiche (v. Wellings e altri, 1975; v. Jensen e altri, 1976).
C'è oggi qualche discussione a tale riguardo circa il destino di alcune lesioni preneoplastiche, che sembrano incapaci di progredire verso il cancro, comportandosi quindi più come neoplasie abortive che come vere preneoplasie (v. Sinha e Dao, 1975; v. Dao, 1977). Ma ciò non invalida l'accordo generale circa qualche tipo di alterazione morfologica che precede, nei vari modelli tumorali, la trasformazione maligna.
La terza tappa nella cancerogenesi mammaria è rappresentata dall'accrescimento tumorale maligno che segue l'iperplasia focale. Comunque, quando la trasformazione maligna si manifesta nelle lesioni mammarie, queste sono di solito ancora nella fase preclinica. Quindi, la quarta tappa della malattia neoplastica mammaria è rappresentata dalla fase clinica del cancro che segue quella preclinica (v. Squartini, 1962).
c) Altri comuni tipi di cancro umano
Tali considerazioni, derivate da un'osservazione dei fatti morfologici, clinici e sperimentali che precedono e accompagnano lo sviluppo del cancro mammario, non sono peculiari a questo ma possono applicarsi o estendersi al cancro umano in generale. Limitando i riferimenti alle neoplasie più comuni, potranno servire come orientamento i dati seguenti.
Il cancro dell'intestino è spesso preceduto da e associato a lesioni focali iperplastiche benigne indicate col nome di polipi adenomatosi o villosi. In particolari condizioni genetiche, come si è già detto (v. sopra, cap. 4), i polipi sono numerosissimi e la trasformazione di uno o più di questi in cancro è praticamente la regola che conduce a morte i pazienti in età giovane. Ma anche il più usuale carcinoma dell'intestino è associato con polipi sporadici, dei quali talvolta è possibile cogliere la trasformazione maligna (v. Spratt e altri, 1958; v. Spratt e Watson, 1971). Una malattia oggi non rara del colon, la colite ulcerosa cronica, che decorre per lo più lentamente con ulcerazioni e proliferazioni polipoidi della mucosa su base infiammatoria irritativa, favorisce con frequenza significativa lo sviluppo di un carcinoma del colon nel giro di 10 o 20 anni (v. DeDombal, 1968; v. Devroede e altri, 1971). Ciò dà un'idea della lentezza, delle possibili concause e delle successive tappe morfologiche nel cancro intestinale.
Nello stomaco sono pure noti polipi e poliposi che con alta frequenza si trasformano in cancro, ma sono tuttavia rari. Più frequenti invece sono altre associazioni con lesioni, come la gastrite cronica, l'anemia perniciosa, o l'ulcera peptica cronica, che nel giro di molti anni possono talora esitare in cancro. I cancri delle labbra, del cavo orale, della laringe e quelli della mucosa bronchiale nei fumatori possono essere preceduti per lunghissimo tempo da lesioni ipercheratotiche benigne meglio note col nome di leucoplachie, le quali a questi livelli sono considerate lesioni precancerose.
I dermatologi hanno potuto riconoscere e classificare varie lesioni che precedono di anni lo sviluppo dei comuni cancri della cute. E i ginecologi, in associazione con i citopatologi, hanno nettamente ridotto la mortalità per cancro del collo dell'utero imparando a riconoscere una serie sequenziale di lesioni da modeste a gravi (iperplasia, displasia, carcinoma in situ o non invasivo) preliminari all'insorgenza del carcinoma invasivo (v. Johnson e altri, 1964; v. Burghardt, 1973).
d) Rappresentazione schematica della malattia neoplastica nell'uomo
Questi concetti possono essere riassunti in uno schema in cui la malattia neoplastica è rappresentata da una linea orizzontale con quattro segmenti o fasi e precisamente: la fase biologica della cancerogenesi caratterizzata da assenza di lesioni istologiche, la fase premaligna morfologica, caratterizzata da lesioni preneoplastiche, la fase preclinica del cancro caratterizzata in genere da aspetti di invasione locale e la fase clinica del cancro caratterizzata in genere da diffusione metastatica a distanza (v. tab. III; v. Severi e Squartini, 1955). Il tempo che intercorre fra (a) e (d) è il periodo di latenza ed esso sembra durare parecchi anni nell'uomo. Durante questo periodo molte misure preventive e terapeutiche probabilmente avrebbero successo e ciò stimola all'acquisizione di nuove informazioni circa le fasi tumorali precoci. Si deve imparare a conoscere quali lesioni sono veramente preneoplastiche, quando queste sono reversibili e quando divengono irreversibili. Si deve provvedere una definizione più precisa delle fasi preinvasiva e invasiva precoce del cancro preclinico. Il tempo che intercorre tra (d) ed (e) è quello che conosciamo meglio, specialmente a causa dei nostri insuccessi (v. Squartini, 1978).

Precisazioni ulteriori suggerite a questo schema sono state recensite, riassunte e analizzate in dettaglio di recente (v. Foulds, 1975). La prima fase, detta fase A o fase iniziatoria, è descritta come una zona più o meno estesa di ‛neoplasia incipiente' con ciò intendendosi una capacità di sviluppo neoplastico superiore a quella che il tessuto possedeva prima dell'applicazione del cancerogeno, senza apprezzabili alterazioni morfologiche. Sarebbe dunque un cambiamento di stato nel comportamento biologico delle cellule a ‛iniziare' la neoplasia dando origine, secondo le numerose terminologie usate, a ‛zone trasformate senza modificazioni morfologiche', o ‛campi neoplastici potenziali', o ‛tessuti normali predisposti', o semplicemente a ‛tessuti iniziali'. In realtà nella fase A si possono osservare modificazioni morfologiche di scarso rilievo indicate come A1 e A2. Le lesioni A1 sono ‛stigmate di esposizione' o puri effetti locali del danno aspecifico inflitto ai tessuti dai cancerogeni, specie se potenti. Le lesioni A2 comprendono una mescolanza di lesioni proliferative transitorie e banali che non prendono parte ulteriore nello sviluppo della neoplasia.
La seconda fase della malattia neoplastica, detta fase B, è la continuazione senza alcun limite netto della precedente ed è caratterizzata da lesioni del gruppo B, che sono le lesioni precancerose o preneoplastiche, passibili fra loro di ulteriore distinzione secondo la minore (B1) o maggiore (B2) tendenza alla malignità. Queste sono in genere lesioni focali spesso multiple le quali si sviluppano nella zona di ‛neoplasia incipiente' della fase A e da esse potrà successivamente iniziare la trasformazione maligna. Ma, indipendentemente dalla probabilità che una neoplasia maligna emerga in una lesione B, c'è una definita e talora elevata probabilità che essa emerga direttamente dai tessuti normali limitrofi inclusi nella zona di ‛neoplasia incipiente' definita (o predestinata o commissionata) dall'azione del cancerogeno nella fase A senza l'intervento di alcuna lesione del gruppo B. Vi sono ormai dati anatomoclinici sufficienti per sottolineare l'importanza di questo rilievo che, lungi dall'annullare, riqualifica il concetto di precancro.
Con la trasformazione maligna ha inizio la fase avanzata o fase C della malattia neoplastica caratterizzata dall'‛invasione' e dalla ‛metastatizzazione', i due aspetti cardinali della ‛malignità', non necessariamente associati (v. Foulds, 1975). Anche questa fase si suddivide in due tempi successivi (C1, C2), per indicare gradi progressivamente crescenti di malignità mediante sigle che possono perciò servire anche a distinguere il cancro preclinico (C1) da quello clinico (C2). Poichè l'invasione locale è il presupposto della metastatizzazione, dipenderà dalla precocità della diagnosi clinica se le metastasi compaiono in fase C1 o C2.
Tornando ai corollari di questa impostazione concettuale della malattia neoplastica, basata come si è visto su solide evidenze anatomocliniche, balzano dunque in piena luce le possibilità ancora esistenti di ricerca e di intervento nel corso di fasi meno gravi, meno conosciute e, si prevede, meglio trattabili e controllabili per interrompere il corso delle neoplasie. Tali possibilità di intervento e di ricerca sono essenzialmente collegate a una definizione, uno studio e un trattamento migliori dei precursori morfologici del cancro e a un maggiore chiarimento dei meccanismi della progressione tumorale che regolano il passaggio fra le varie fasi della malattia neoplastica. A questi argomenti sono dedicati i due capitoli seguenti.
8. Precursori morfologici
a) Definizione
Nei tre quarti di secolo trascorsi da quando il termine ‛precancerosi' venne usato forse per la prima volta a proposito di alcune malattie della pelle (v. Dubreuilh, 1898), non si è riusciti a ottenere una definizione univoca del suo significato. Infatti, questo termine è ancora oggi largamente usato per indicare, e purtroppo per confondere fra loro, due concetti assai diversi. Secondo uno di questi concetti, preneoplasia o precancro è una lesione istologica che non mostra alcuna tendenza invasiva, ma che diverrà inevitabilmente cancerosa. Secondo l'altro concetto precancro è invece una lesione istologica ritenuta capace, su base statistica, di dare origine al cancro con frequenza significativa. La prima di queste interpretazioni sembra a chi scrive troppo restrittiva, mentre la seconda appare più appropriata per una condizione patologica che, in quanto tale, è soggetta a innumeri variabili e sottratta a ogni regola precisa. La definizione che segue discende dalle premesse svolte nel capitolo precedente e tiene conto di queste considerazioni: ‟Per lesione precancerosa si intende una condizione sopraggiunta di tipo biologico o morfologico di un tessuto, caratterizzata dalla tendenza a evolvere in cancro" (v. Severi e Squartini, 1973).
b) Analisi della definizione
Per una migliore comprensione del significato è opportuno qualche breve commento alle parti che compongono la definizione. ‟Condizione sopraggiunta" sta a significare che l'inizio della lesione precancerosa si identifica con un preciso cambiamento di stato del tessuto colpito il quale, a seguito di tale cambiamento, acquista potenzialità evolutive nel senso dei tumori prima non possedute. Non è tanto la lesione apprezzabile a livello morfologico che ha importanza, quanto ciò che si trova alla sua origine. Spesso l'applicazione di un cancerogeno è seguita da alterazioni (regressive, infiammatorie) non essenziali per la cancerogenesi: infatti, con dosi opportunamente ridotte, queste possono mancare senza che l'occorrenza dei tumori ne sia modificata. La lesione precancerosa è dunque tale in quanto provocata da una modificazione cellulare non necessariamente irreversibile, ma certo persistente e capace di favorire, attraverso lo squilibrio che determina, l'instaurarsi di modificazioni cellulari successive e progressivamente più gravi. Rispetto al tempo, la condizione precancerosa si può instaurare in qualsiasi momento della vita, quindi anche prima della nascita, come probabilmente accade nel caso dei tumori congeniti o di quelli derivanti dalle malformazioni, dagli amartomi e dalle vestigia dello sviluppo embrionale (v. Squartini e Bolis, 1966; v. Severi e Squartini, 1969). La locuzione ‟di tipo biologico o morfologico" sottolinea il concetto che una lesione precancerosa non comincia nè si esaurisce nelle alterazioni della struttura istologica del tessuto interessato, ma in genere preesiste a questa come una condizione biologica prima che morfologica (v. Berenblum, 1949; v. Severi e Squartini, 1955 e 1956). All'inizio del periodo di latenza la lesione precancerosa può interessare il livello ultrastrutturale, oppure non raggiungere neanche quella soglia e svolgersi per esempio a livello biochimico (‛lesione biochimica': v. Cameron e Spector, 1961). Per quanto la fase iniziale e morfologicamente muta della malattia neoplastica non sembri avere al momento utilità pratica, questo non giustifica che essa venga trascurata.
‟Con tendenza a evolvere in cancro" esprime il concetto caratterizzante della definizione. Tendenza sembra infatti la parola più appropriata per indicare che la trasformazione di una lesione precancerosa in cancro non è inevitabile e non è nemmeno solo casuale o possibile, ma rappresenta invece un evento probabile. Per mantenere ampiezza sufficiente alla definizione si deve ammettere che la frequenza, o probabilità, della trasformazione è variabile da lesione a lesione, in rapporto a fattori che solo in parte possono essere indicati. La dipendenza dallo stimolo causale (v. DeOme e altri, 1959, 1961 e 1962) e il tempo necessario alla trasformazione (v. Mühlbock e altri, 1952) sono due di questi fattori resi noti dalla patologia sperimentale. I noduli preneoplastici di iperplasia alveolare della mammella del topo sono strutture ormonodipendenti (v. Nandi e altri, 1960; v. Nandi, 1961), per cui la rimozione o la cessazione dello stimolo che le sostiene ne può causare la scomparsa (v. Squartini, 1960 e 1966). Per quanto i noduli di iperplasia alveolare siano presenti a decine nel sistema mammario di topi suscettibili infettati con il virus di Bittner (v. Nandi, 1963), solamente uno o pochissimi cancri hanno realmente il tempo di manifestarsi (v. Olivi e altri, 1958).
Un giudizio preciso circa l'evoluzione delle lesioni precancerose è reso anche difficile dalla loro aspecificità sul piano morfologico. Per rimanere nello stesso esempio, i noduli di iperplasia alveolare della mammella del topo sono pressoché indistinguibili a livello morfologico (v. Severi e altri, 1958), istochimico (v. Harkness e altri, 1957) e biochimico (v. DeOme e altri, 1956) dalla adenosi fisiologica della gravidanza. Inoltre, lo stesso tipo di lesione si ritrova sia in topi portatori del virus di Bittner, sia in topi geneticamente uguali che ne sono privi (v. Pitelka e altri, 1964). Ma a dispetto della loro somiglianza morfologica i noduli sviluppati dai primi hanno un'alta probabilità e quelli sviluppati dai secondi una bassa probabilità di trasformarsi in cancro (v. Blair e DeOme, 1962). Sebbene gli esempi sopra riferiti provengano da studi di oncologia sperimentale, vi sono pochi dubbi che circostanze analoghe siano operative anche nel campo delle precancerosi umane.
c) Acquisizioni sperimentali
Gli studi sulla morfologia del periodo di latenza nei tumori sperimentali hanno condotto a tre constatazioni di rilievo: a) la costanza di lesioni morfologiche nei tessuti prima della malignità attuale; b) la variabilità e aspecificità di tali lesioni; c) la loro natura frequentemente solo microscopica o subclinica. Nei sistemi meglio studiati di tumori sperimentali la lesione ‛benigna' rappresenta, salvo eccezioni, un passaggio pressoché obbligato fra tessuto normale e neoplasia maligna, cosicché la sequenza consueta di eventi che si osserva è: tessuto normale → lesione benigna → neoplasia maligna. Ciò vale per la cancerogenesi cutanea (v. Berenblum, 1954), mammaria (v. De Ome, 1963), polmonare (v. Squartini e altri, 1966), delle ghiandole endocrine (v. Bielschowsky e Horning, 1958), ecc. Le lesioni premaligne hanno un volto diverso da tessuto a tessuto e talora anche in uno stesso tessuto. Per esempio, il cancro della mammella del topo può prendere origine da noduli di iperplasia alveolare, che sono lesioni microscopiche acinose (v. DeOme e altri, 1959), da placche, che sono lesioni palpabili a struttura ductulare cosi chiamate per la loro forma (v. Foulds, The histologic analysis... II, 1956), o da papillomi dei dotti (v. Orr, 1956). Inoltre, uno stesso tipo di lesione osservato nel medesimo tessuto in gruppi di ospiti diversi, può, talvolta molto frequentemente, altre volte molto raramente, sviluppare un cancro. Un esempio in questo senso è già stato fornito sopra (v. Blair e DeOme, 1962).
Se la lesione premaligna è molto piccola, il cancro che da questa si sviluppa ne cancella rapidamente le tracce, per cui l'impressione è che il cancro abbia preso origine da un tessuto normale. Se invece la lesione premaligna è palpabile, diviene possibile seguirne la morfologia e il comportamento prima, durante e dopo la trasformazione maligna. Il sistema dei tumori mammari del topo fornisce un bell'esempio di queste due diverse situazioni attraverso i noduli di iperplasia alveolare e le placche (v. Squartini e Rossi, 1962). Nelle seconde, palpabili, la trasformazione assume di solito l'aspetto di un'area focale con struttura diversa, la quale progressivamente si estende guadagnando l'intera lesione premaligna e mostrando talora gli attributi della malignità (v. Foulds, The histologic analysis. II, 1956). Nei noduli di iperplasia alveolare rimane invece difficile seguire al microscopio la trasformazione (v. Squartini e Rossi, 1960).
L'osservazione sperimentale recente che i noduli di iperplasia alveolare, mantenuti indefinitamente a mezzo di trapianto, di rado divengono maligni (v. Nandi, 1977), e che nella cancerogenesi chimica della mammella del ratto, pure in presenza di numerosi noduli di iperplasia alveolare, i tumori maligni prendono origine da porzioni di ghiandola apparentemente normali (v. Sinha e Dao, 1975; v. Dao, 1977), ha dato origine a qualche scetticismo circa il reale significato delle lesioni preneoplastiche. Ma tale scetticismo è facilmente superato se si considera il fatto che queste lesioni, ove pure non rappresentino la sede della trasformazione neoplastica, rimangono sempre segni premonitori di malignità che avrà luogo nelle loro vicinanze e se si riflette sulla possibilità che l'area del tessuto inizialmente modificato dall'azione del cancerogeno responsabile dell'accresciuto rischio di malignità sia più estesa di quella mostrante la lesione morfologica (v. Foulds, 1975).
d) Variabilità e molteplicità strutturale
Nell'uomo, in dipendenza del tessuto di origine, dei fattori causali e, forse, di altri elementi ancora sconosciuti, le alterazioni che precedono la malignità attuale possono assumere il volto dell'iperplasia, della displasia, della metaplasia, della neoplasia benigna in forma di papilloma, polipo, adenoma o simili, della disontogenia, ecc. Un'iperplasia diffusa persistente è fertile terreno per lo sviluppo della malignità. Un esempio tipico è dato dallo struma o gozzo tiroideo. Intorno ai primi decenni di questo secolo il cancro della tiroide era circa 8 volte più frequente nelle aree geografiche con gozzo endemico che nelle altre. Una certa prevalenza del cancro tiroideo nelle regioni gozzigene esiste anche oggi, sebbene non sia più evidente come in passato. Ciò è dovuto al largo impiego di sali iodati introdotti a scopo profilattico dopo il 1920 nelle aree gozzigene a deficienza di iodio, da cui si può concludere che la profilassi del gozzo è una profilassi del cancro della tiroide (v. Winder, 1952).
La displasia di un tessuto, che significa crescita non solo in eccesso ma anche incoordinata dei singoli costituenti, più spesso della semplice iperplasia rappresenta un ponte fra la normalità strutturale e la malignità. Esempi tipici in questo caso sono forniti dalla mastopatia fibrocistica, che precede talvolta il cancro mammario (v. Severi, 1952; v. Black, 1976), e dalla displasia epiteliale del collo dell'utero, che prelude al carcinoma in situ e al cancro manifesto (v. Burghardt, 1973). Un altro tipo di lesione premaligna è la metaplasia, cioè una differenziazione, per esempio epiteliale, anomala per il tessuto od organo che si considera mentre è normale altrove. Tipico è il caso della metaplasia squamosa o epidermoide degli epiteli mucosi di rivestimento (del labbro, della lingua, della laringe, ecc.). Frequente è pure il caso che la malignità si realizzi in tumori benigni quali gli adenomi o polipi adenomatosi e i papillomi o polipi villosi dell'intestino crasso, o i fibroadenomi della mammella nel caso dei sarcomi di questa ghiandola. Un terreno particolare, infine, per l'insorgenza della malignità è dato dalla disontogenia, cioè dalla condizione di alterato rapporto dei tessuti embrionali (amartomi), dalle malformazioni propriamente dette e dai teratomi (v. Willis, 1962). Come si vede, molte fra le lesioni premaligne appartengono alla categoria dei processi progressivi. Ma non è questa una regola applicabile a tutte. Ve ne sono alcune che iniziano come processi regressivi. Basterà ricordare in proposito la cirrosi epatica (v. Berman, 1962) e, per le aree geografiche in cui le è riconosciuto un significato premaligno, l'ulcera peptica dello stomaco (v. Hirafuku, 1962). Anche in queste lesioni, tuttavia, la fase regressiva mette poi in movimento un meccanismo di riparazione che porta a proliferazione dei tessuti. La sequenza irritazione → danneggiamento → riparazione costituisce infatti una delle vie alternative attraverso cui si può giungere al cancro.
e) Classificazione patogenetica
Un tentativo di classificare le lesioni premaligne su base patogenetica è stato fatto suddividendo gli stati precancerosi in: a) disontogenetici, comprendenti le anomalie riconducibili a disturbi dello sviluppo embrionale (amartomi, malformazioni, teratomi, ecc.); b) iperplasiogeni, insorgenti sulla base di un'irritazione cronica di natura infettiva, fisica o chimica che causa rigenerazione di tessuti (focolai luposi, cicatrici da ustione, ulcere da raggi, ecc.); c) disendocrini, derivanti da alterazioni nell'equilibrio ormonale (mastopia fibrocistica, iperplasia cistica dell'endometrio, ipertrofia prostatica, ecc.; v. Steiner, 1953). È improbabile che in questo schema trovino posto tutte le possibili lesioni premaligne e sarebbe perciò utile prevedere una quarta categoria per le lesioni miscellanee. Ma bisogna riconoscere a tale classificazione criteri di semplicità, logica e utilità sufficienti a mantenerla valida. Del resto, ogni altra base classificativa, dall'aspetto istologico all'evoluzione clinica, dal momento eziologico alla distribuzione topografica, risulterebbe impropria per lesioni tanto vaghe nelle cause quanto variate nella struttura e imprevedibili nel comportamento.
f) I precursori morfologici del cancro umano
La lista dei precursori morfologici del cancro umano è lunghissima. Se ne elencano molti della cute (cheratosi senile, da agenti fisici e chimici, xeroderma pigmentosum, nevo giunzionale, infiammazioni croniche, cicatrici da ustioni, ecc.), della mammella (papillomi dei dotti, carcinoma lobulare in situ, mastopatia fibrocistica, lobuli atipici e postmenopausali, fibroadenomi, ecc.), dell'endometrio (iperplasia cistica postmenopausale, iperplasia adenomatosa atipica, polipi, ecc.), della cervice uterina (displasia, metaplasia squamosa, carcinoma in situ, ecc.), dello stomaco (polipi adenomatosi, gastriti croniche, ulcera peptica, anemia perniciosa, ecc.), dell'intestino (polipi adenomatosi, adenomi villosi, colite ulcerosa cronica, ecc.), del fegato (cirrosi emocromatosica, cirrosi volgare, iperplasia dei dotti biliari, ecc.), della vescica (papillomi, cistiti croniche, bilharziosi, estrofia, ecc.), del polmone (cicatrici, metaplasie e iperplasie atipiche del rivestimento dei bronchi, tumourlets, ecc.), e praticamente di qualsiasi altra sede (v. Severi, 1962; v. Severi e Squartini, 1973; v. AA. VV., Early lesions..., 1976; v. AA. VV., 1977).
Queste lesioni sono parte integrante del processo di sviluppo del cancro (v. Gullino, 1977) e a esse dovrebbero essere dedicati studi più approfonditi volti alla ricerca di forme di controllo del cancro durante la fase premaligna, di nuovi metodi per selezionare gli individui esposti a maggior rischio, e di più accurati marcatori diagnostici che rendano possibile una migliore definizione della preneoplasia e dei suoi rapporti con la neoplasia invasiva (v. Antony e altri, 1976).
g) Attuali indirizzi di ricerca e conclusioni
Sul piano sperimentale non mancano possibilità di approfondire la conoscenza dei caratteri delle lesioni preneoplastiche attraverso studi come quelli, per esempio, sulle loro proprietà angiogeniche e sulla loro trasformazione nei trapianti. Oggi infatti è possibile trapiantare lesioni umane in topi atimici, cioè geneticamente privi del sistema immunitario, in modo da seguirne l'evoluzione nel tempo (v. Jensen e Wellings, 1976). Inoltre, le conoscenze sui fenomeni vascolari che accompagnano la trasformazione maligna consentono di esplorare un'altra via interessante. I tumori maligni per svilupparsi hanno bisogno di un'adeguata trama di vasi sanguigni e sono in grado di indurne lo sviluppo, attraverso un fattore angiogenico ancora sconosciuto, quando un loro frammento viene trapiantato (v. Folkman e Cotran, 1976). Tale proprietà è comune anche alle lesioni premaligne, perciò in grado di attecchire (v. Gimbrone e Gullino, 1976), mentre manca ai tessuti normali, che infatti non attecchiscono se sprovvisti di trama vascolare. L'identificazione del fattore angiogenico nelle lesioni premaligne potrebbe quindi consentire un'identificazione di quelle tra tali lesioni capaci di progredire verso il cancro.
A ottanta anni dal giorno in cui fu intuito forse per la prima volta, l'argomento dei precursori morfologici del cancro rimane dunque uno dei più affascinanti e aperti problemi di patologia. E più che mai aperto alla sperimentazione sugli animali e conserva intatto il suo fascino per il patologo che ancora non dispera di poter predire un giorno il futuro delle lesioni. Oggi purtroppo il ventaglio dei possibili precursori morfologici è estremamente ampio e il loro significato biologico è spesso indecifrabile. Ciò porta a conclusioni forse non troppo soddisfacenti sul piano pratico.
Infatti, poiché tanti sono i possibili precursori morfologici del cancro e tanto difficile è valutare le loro intrinseche possibilità di progressione, si deve concludere che una prevenzione del cancro per questa via sarebbe difficile. Ma il problema va visto in prospettiva. Oggi, l'osservazione di una lesione preneoplastica in un paziente dovrà condurre alla sua rimozione ove questa sia possibile, e potrà servire per aumentare la sorveglianza ed eliminare le cause che possono essere rimosse, ammesso che si sia ancora in tempo (v. Severi e Squartini, 1973). In futuro è prevedibile si disponga di una risposta o di una soluzione pratica soddisfacenti per i tre principali scopi attuali della ricerca sui precursori morfologici del cancro, che sono essenzialmente: la diagnosi precoce, l'interruzione dello sviluppo neoplastico nelle popolazioni ad alto rischio e la comprensione dell'essenza della malattia neoplastica (v. Farber, 1976).
9. Progressione tumorale
Qualche cosa è cambiato nella rigidità con cui, in tempi passati, veniva intesa la separazione fra una fase e l'altra della malattia neoplastica e soprattutto fra precancro e cancro. Negli ultimi decenni si è affermata infatti l'opinione che lo sviluppo di un tumore avvenga per ‛progressione' (v. Foulds, 1949, The histologic analysis... II, 1956, 1969 e 1975; v. Squartini, 1964), cioè attraverso la progressiva acquisizione di nuovi caratteri permanenti (v. Rous e Beard, 1935), senza dunque alcun salto drammatico fra l'innocenza e la malignità (v. Willis, 19674). In base a queste vedute diviene sempre più difficile fissare un punto definito di demarcazione fra (b) e (c) (v. tab. III). Sopra tale concetto della progressione tumorale, che direttamente incide sulla storia naturale della malattia neoplastica e sul significato stesso di lesione premaligna, come su quello di lesione maligna, è pertanto opportuno fermare l'attenzione dedicando a esso qualche riflessione.
a) Definizione
‛Progressione' è la comparsa di modificazioni qualitative stabili, ereditabili e irreversibili in uno o più caratteri di un tumore in accrescimento (v. Foulds, 1949). In senso lato questo termine è perciò usato per indicare l'intero ciclo di sviluppo di una neoplasia attraverso successive modificazioni qualitative, permanenti e irreversibili dei suoi caratteri (v. Foulds, 1951). Sotto questo profilo la progressione si differenzia dalla ‛modulazione', termine che è stato suggerito come il più appropriato per indicare l'occorrenza di quelle variazioni di portata minore, temporanee e reversibili, nella struttura e/o nel comportamento di un tumore dovute di solito a fattori ambientali (v. Foulds, The histologic analysis... I, 1956). Attraverso la progressione i tumori guadagnano un'autonomia sempre maggiore nei confronti dell'ambiente e dell'ospite. Da ciò deriva che il concetto di progressione è indissolubilmente legato a quello, opposto, di ‛dipendenza' (o responsiveness) dei tumori. ‛Dipendenza' indica infatti la capacità delle cellule di un tumore di rispondere agli stimoli estrinseci di qualsiasi natura (ormoni, agenti chemioterapici, cancerogeni chimici, ecc.), quale frequentemente si osserva nelle fasi precoci dello sviluppo di una neoplasia.
b) Fonti di informazione
Le principali fonti di informazione sulla progressione sono rappresentate, come Foulds (v., 1954) suggerisce, dallo studio, condotto con idonei mezzi clinici e patologici, della storia naturale dei tumori negli animali c nell'uomo, dall'osservazione della risposta dei tumori agli stimoli estrinseci, e dal trapianto dei tumori, che si rivela utile sia come test per lo studio della reversibilità di certi caratteri tumorali (modulazione), sia perché, prolungando nel tempo la vita del tumore, offre possibilità maggiori per il manifestarsi della progressione e per lo studio delle fasi tardive di questa.
c) Caratteri tumorali acquisibili per progressione
I ‛caratteri' elencati appresso forniscono un'esemplificazione rappresentativa di quelle che sono le più usuali modificazioni qualitative irreversibili cui i tumori vanno incontro nel corso del loro sviluppo: aumento della velocità di accrescimento, diminuzione dei segni morfologici visibili di differenziazione cellulare (per es., perdita della funzione secernente o di altre funzioni similari), indipendenza dalle influenze ormonali o da altri stimoli estrinseci, capacità di infiltrare i tessuti adiacenti, capacità di sviluppare riproduzioni metastatiche a distanza, ampliamento del raggio di trapiantabilità, acquisizione della capacità di svilupparsi in forme e ambienti diversi dagli usuali (per es., trasformazione di un tumore solido in tumore ascite), ecc. (v. Klein e Klein, Some experiments..., 1958). È attraverso la progressiva acquisizione ditali caratteri biologici e morfologici che i tumori, inizialmente dipendenti e differenziati, si trasformano, nel corso della loro evoluzione, in tumori indipendenti e altamente indifferenziati.
d) Principi generali (regole) della progressione tumorale
L'analisi effettuata da Foulds (v., 1949, 1956 e 1969) su vasti materiali ha portato all'individuazione di alcune regole o principi generali della progressione tumorale. Tali principi sono i seguenti.
1. Progressione indipendente dei tumori multipli: la progressione si manifesta indipendentemente in differenti tumori di uno stesso individuo. Questa regola si applica assai bene a numerosi esempi di tumori multipli dell'uomo e degli animali e il suo significato principale è che la progressione è un fenomeno inerente al tumore e non all'ospite.
2. Progressione indipendente dei caratteri: la progressione di differenti caratteri in uno stesso tumore avviene in maniera indipendente. Questa regola, anch'essa largamente applicabile, conduce alla più generale proposizione che la struttura e il comportamento dei tumori sono determinati da numerosi caratteri elementari i quali, entro ampi limiti, sono variabili indipendentemente e capaci di progressione indipendente, per cui possono essere assortiti e combinati in una varietà di modi. L'esistenza di legami fra certi caratteri tumorali non può essere disconosciuta. Tuttavia, ciò non diminuisce la validità del principio della progressione indipendente dei caratteri elementari, come rivela la sua indubbia utilità nell'interpretazione di quel fenomeno tanto complesso quanto oscuro che è la ‛malignità' (v. Foulds, 1954).
3. La progressione è indipendente dall'accrescimento: essa può infatti manifestarsi indifferentemente sia in cellule tumorali latenti, sia in tumori il cui sviluppo si era arrestato, o in fase di regressione, o in via di accrescimento. Da questa terza regola derivano due corollari di notevole importanza pratica: a) un tumore, quando diviene clinicamente manifesto, può essere in qualsiasi stadio di progressione; b) la progressione è indipendente dalle dimensioni e dalla durata clinica di un tumore.
4. La progressione può essere continua o discontinua: essa può cioè realizzarsi attraverso modificazioni graduali o per salti improvvisi. Ciò significa che più di un gradino dell'ipotetica scala dei caratteri di progressione può essere saltato in molti casi.
5. La progressione può svilupparsi secondo vie diverse o modelli alternativi i quali possono condurre a punti di arrivo diversi o uguali.
6. La progressione non raggiunge sempre il suo punto terminale nel corso della vita dell'ospite primario: questo principio, di frequente osservazione in patologia sperimentale, e di grande importanza pratica, non necessita di esemplificazioni.
e) Esempi e considerazioni
Tali principi sono applicabili così ai tumori indotti negli animali, come ai tumori dell'uomo. Fra i sistemi di tumori sperimentali e umani che meglio si prestano a dimostrare la generale applicabilità dei principi della progressione si possono citare: i tumori cutanei, i tumori delle vie urinarie, i tumori delle ghiandole endocrine e degli organi terminali del sistema endocrino (mammella, utero). Fra i tumori umani i seguenti esempi sono particolarmente adatti per illustrare la validità del principio della progressione indipendente dei caratteri tumorali. I cosiddetti ‛tumori misti' delle ghiandole salivari possiedono gli attributi morfologici locali della malignità, ma non metastatizzano salvo rare eccezioni (v. Thackray e Lucas, 1974). Alcuni tumori tiroidei sono definiti adenomi proprio per la mancanza di segni evidenti di invasione locale, ma metastatizzano (v. Squartini e Severi, 1964). I cancri della mammella, e persino le loro metastasi, sono sovente ancora dipendenti per l'accrescimento dagli stimoli ormonali dell'ospite (v. Currie e Illingworth, 1958; v. McGuire e altri, 1975; v. De Sombre e altri, 1976). Varie linee cellulari di cancri mammari umani coltivate in vitro mostrano di essere dipendenti dagli estrogeni, o dal progesterone, o dagli androgeni, ecc. (v. i contributi di Lippman e altri, 1976).
Il concetto della progressione tumorale e in particolare il principio della progressione indipendente dei caratteri tumorali, indicano dunque che non esiste la possibilità di una demarcazione netta fra precancro e cancro, e conducono quindi al superamento di ogni rigida contrapposizione fra queste due parole. Lo studio del comportamento biologico dei tumori associato alla conoscenza della loro struttura morfologica ha dimostrato che la dipendenza è peculiare dei periodi precoci delle neoplasie e corrisponde largamente alla fase premaligna. La progressione sopprime la dipendenza, realizzando il passaggio verso l'autonomia totale che corrisponde largamente alla fase maligna. Tuttavia la dipendenza può essere parzialmente mantenuta, anche per lunghi periodi, da tumori in progressione e la progressione non si arresta quando la malignità morfologica è raggiunta, ma si estende oltre, essendo responsabile di modificazioni successive nei caratteri dei tumori.
Lo schema della malattia neoplastica illustrato in precedenza (v. tab. III) configura pertanto una situazione teorica nella quale tutti i caratteri di malignità vengono acquisiti contemporaneamente dal tumore. In realtà, la maggior parte dei tumori esibisce una comune associazione di caratteri clinici e patologici fra loro rispondenti o armonici, ciò che consente di mantenere nella pratica quotidiana la distinzione insostituibile fra malignità e benignità. Tuttavia, una progressione ‛sfasata' dei caratteri tumorali è largamente possibile, come si è visto, e deve essere sempre tenuta presente dal patologo come dal clinico.
f) Meccanismi di progressione
Circa i meccanismi della progressione tumorale vari dati sono stati accumulati a sostegno dell'ipotesi di una selezione clonale operativa nella prima come nelle successive fasi dello sviluppo neoplastico (v. Prehn, 1976). Per clone si intende la popolazione cellulare discendente da un'unica cellula progenitrice. Si è già detto che il processo iniziale della cancerogenesi consiste in una modificazione potenziale o latente sopra una larga zona di tessuto. Ma i dati suggeriscono che questa è il risultato dell'amplificazione clonale di un'alterazione ereditabile manifestatasi dapprima in una sola cellula. Anche le tappe successive della cancerogenesi è verosimile che avvengano attraverso un analogo meccanismo di selezione clonale. Si ritiene infatti che le mutazioni spontanee siano molto frequenti nelle popolazioni di cellule tumorali. Quindi, una popolazione cellulare premaligna o maligna non mostra di solito omogeneità genetica. Ciò comporta una proliferazione competitiva di differenti doni cellulari dentro la stessa popolazione. Se uno di questi è provvisto di una maggior resistenza agli stimoli ambientali, o di una più elevata velocità di accrescimento, esso può sopravanzare gli altri. In tal modo l'indipendenza di una popolazione di cellule tumorali può progressivamente aumentare attraverso selezioni clonali successive su base mutativa (v. Klein e Klein, Some experiments..., 1958; v. Prehn, 1976).
Un esempio visivo di questo fenomeno si ha nello studio della progressione dei tumori mammari del topo gravidico-dipendenti o placche verso stadi di crescente autonomia. Le placche sono tumori con struttura tubulare organoide che crescono in gravidanza e spariscono dopo il parto. Quando vi si manifesta, la progressione di solito appare come un'area focale rotondeggiante di struttura diversa dentro la placca. Dopo il parto la placca residua regredisce, ma non così l'area di progressione focale. Per cui il corso clinico del tumore sarà irrevocabilmente modificato, dipendendo ora dal potenziale di accrescimento e dal livello di differenziazione cellulare del focolaio di progressione (v. Foulds, The histologic analysis... II, 1956; v. Squartini e Rossi, 1959; v. Squartini, 1966).
In certi casi la progressione tumorale potrebbe essere dovuta a perdita di taluni isoantigeni da parte delle cellule neoplastiche, oppure ad acquisita resistenza di queste verso l'azione di isoanticorpi specifici (v. Klein e Klein, Some experiments..., 1958). In conclusione, e senza entrare nei particolari di altri meccanismi possibili, la progressione dei tumori è il risultato di modificazioni cellulari successive le quali non seguono sempre o necessariamente la stessa strada, ma possono viaggiare lungo vie diverse che conducono allo stesso risultato finale, la neoplasia, divergendo però circa i meccanismi citologici di dettaglio (v. Klein e Klein, A system..., 1958).
g) Notazioni critiche
La progressione tumorale, un concetto come si vede solidamente fondato e costruito in 40 anni di osservazioni ed esperienze, non è tuttavia risparmiata da notazioni critiche che preludono, su basi sperimentali, a ipotesi alternative. Recentemente, per spiegare la lunga durata della malattia neoplastica, è stato proposto che le cellule trasformate siano inibite nel loro accrescimento dalle cellule normali contigue, per cui lo sviluppo tumorale sarebbe procrastinato a quando l'età o eventuali fattori promoventi riducono in maniera assoluta o relativa la popolazione cellulare limitrofa normale consentendo alle cellule trasformate, senza necessità per esse di ulteriori mutazioni, di sfuggire al controllo e così realizzare dal ‛tumore potenziale' la malattia neoplastica reale (v. Nandi, 1978).
10. Diffusione metastatica
Un tumore maligno non sarebbe la malattia drammatica e tanto spesso mortale che tutti conosciamo se non avesse la capacità di metastatizzare, cioè di diffondersi riproducendosi a distanza. La diffusione metastatica rappresenta perciò un fenomeno fondamentale nel comportamento anatomoclinico di una neoplasia e quindi la sua conoscenza risulta indispensabile alla comprensione della dinamica di questa malattia.
a) Definizione e vie di metastatizzazione
Metastasi è la ripetizione nell'ospite di un tumore, separata da questo e originata da cellule o frammenti di esso trasportati a distanza lungo le vie naturali. La metastatizzazione è conseguenza della diffusione locale diretta del tumore primitivo che avviene per infiltrazione dei tessuti lungo gli interstizi intercellulari o i vasi linfatici (permeazione linfatica) e per invasione di vene, capillari, arterie, cavità celomatiche (cavi pleurici, pericardico, peritoneale), spazi cerebrospinali e cavità epiteliali (tubo digerente, vie urinarie, vie respiratorie o lume tracheobronchiale).
L'infiltrazione e l'invasione diretta aprono quindi al tumore primitivo cinque vie naturali per la metastatizzazione e precisamente: i vasi linfatici, i vasi sanguigni, le cavità celomatiche, gli spazi cerebrospinali (più precisamente lo spazio subaracnoideo in cui circola il liquor) e le cavità epiteliali (v. Willis, 19674). Solo le prime due vie hanno però rilevanza generale, mentre le altre, e specialmente le ultime due, sono seguite solo in un limitato numero di casi particolari.
b) Fasi della metastatizzazione
Qualunque sia la via, il processo di metastatizzazione si svolge in tre fasi successive: il distacco e la penetrazione di cellule neoplastiche nelle vie naturali, il trasporto (oppure la caduta) e l'arresto di queste cellule in sedi diverse dell'organismo, l'impianto o attecchimento e lo sviluppo da esse di tumori secondari (metastasi) indipendenti da quello primitivo (v. Consolandi, 1960).
Il distacco dal tumore primario e la penetrazione di cellule neoplastiche nelle vie naturali dipende da numerosi fattori (v. Consolandi, 1960) quali la crescita progressiva del tumore (v. Willis, 1948), l'aumento della pressione tessutale (v. Young, 1959), la necrosi dei vasi tumorali, la liberazione di sostanze diffusive a opera del tumore (v. Coman, 1953), la modificazione della sostanza fondamentale del connettivo (v. Gasic e altri, 1960), la reazione stromale, la diminuzione della mutua coesione fra le cellule neoplastiche (v. Ambrose, 1958), l'attività ameboide di queste (v. Fujiwara, 1959), la struttura istologica, il grado di differenziazione, il corredo cromosomico (v. Yoshida, 1959) e la progressione tumorale (v. Wallace, 1961; v. Leighton, 1967; v. Squartini e altri, 1968).
Il trasporto a distanza dal tumore primario di cellule neoplastiche lungo le vie naturali è un evento passivo che dipende dalla via seguita e dalle leggi fisiche inerenti. Nelle cavità celomatiche o nei lumi epiteliali il trasporto si identifica con la caduta dei frustoli neoplastici. Negli spazi cerebrospinali il trasporto di cellule neoplastiche sembra avvenire per sedimentazione entro il liquor. Nel caso dei vasi linfatici rimane sempre difficile stabilire dove finisca l'infiltrazione (permeazione linfatica) e dove cominci il trasporto di cellule a distanza secondo corrente, da stazione a stazione. I vasi sanguigni, che nel trasporto di cellule o di emboli neoplastici hanno pure il significato passivo di canali di trasferimento, costituiscono perciò l'unica grande via di metastatizzazione per la quale il problema dell'arresto dei frustoli neoplastici circolanti assuma rilevanza e concretezza tali da consentire e indurre a studi più razionali (v. Wood e altri, 1961; v. Squartini e altri, 1968; v. Willis, 19732).
Per intendere i problemi che seguono è opportuno premettere alcuni richiami essenziali sul circolo sanguigno in generale ricordando che dal cuore sinistro, attraverso l'aorta, il sangue ossigenato è trasportato a ciascun organo ove giunge per mezzo di un'arteria afferente, si sfiocca nella rete di capillari destinati alla nutrizione dell'organo, da questi viene convogliato nella vena efferente che, attraverso le cave, lo scarica nel cuore destro. Il cuore destro spinge tutto il sangue, attraverso l'arteria polmonare, nel circolo capillare dei polmoni per l'ossigenazione e le vene polmonari si incaricano quindi di raccoglierlo scaricandolo nel cuore sinistro. Le reti capillari, a causa delle piccole dimensioni dei lumi capillari (intorno a 10μ), funzionano da filtri per eventuali contenuti solidi (emboli) del sangue: quelle dei singoli organi per il sangue destinato all'organo, quelle dei polmoni per il sangue refluo attraverso le vene da tutti gli organi. Il sangue refluo dal tubo digerente (stomaco, intestino, pancreas) e dalla milza ha un doppio filtro capillare interposto sul suo cammino, poiché la vena porta lo conduce al fegato e alla sua rete capillare (filtro epatico) e le vene epatiche lo conducono di qui al cuore destro e al polmone.
Per l'arresto di cellule o tessuti circolanti il problema è di sapere se questo è regolato da fattori meccanici (v. Walther, 1948; v. Consolandi, 1949), secondo le leggi della circolazione e dell'embolia, o da fattori prevalentemente biologici (v. Willis, 1952, 19674, 19733), secondo predilezioni particolari dei tumori donatori e/o idoneità differenziali degli organi e tessuti riceventi. L'importanza, o la preminenza, della teoria ‛meccanica' è stata ripetutamente illustrata e sottolineata (v. Consolandi, 1950 e 1951; v. Coman, 1953), ma sempre più peso ha assunto in tempi recenti la teoria ‛biologica' della distribuzione metastatica, in particolare dopo la dimostrazione sperimentale del passaggio diretto di emboli neoplastici attraverso il filtro capillare polmonare (v. Zeidman, 1957) o altri territori capillari (v. Korpassy, 1956; v. Korpassy e altri, 1958), dopo le prove che in condizioni sperimentali identiche tumori diversi sviluppano modelli di distribuzione metastatica diversi (v. Schmähl e Rieseberg, 1958; v. Schmähl, 1959), e specialmente dopo la sistematica documentazione della presenza di cellule neoplastiche circolanti, isolate o aggregate, nel sangue periferico (v. Engell, 1955; v. Pruitt e altri, 1958).
Tuttavia, la quantità nettamente maggiore di cellule e ammassi di cellule neoplastiche nel sangue regionale rispetto al periferico (v. Potter e altri, 1960), la documentazione della elevata efficienza filtrante del fegato per le cellule tumorali circolanti (v. Fletcher e Stewart, 1959), e gli studi attuali sulle caratteristiche angioarchitettoniche dei vari territori di irrorazione con la dimostrazione dei frequenti corto-circuiti artero-venosi d'organo (v. Bucciante, 1960; v. Semisch, 1959), hanno ricondotto anche di recente l'accento sull'importanza di alcuni fattori emodinamici nella distribuzione metastatica dei tumori (v. Wieberdink, 1957; v. Gabler e Peckholz, 1960).
Come non tutte le cellule tumorali circolanti si arrestano nei filtri capillari interposti sul loro cammino, così non tutte le cellule arrestate in un dato distretto sopravvivono (v. Engell, 1959; v. Baserga e altri, 1960) e non tutte le cellule sopravvissute producono metastasi, poiché possono rimanere a lungo latenti senza proliferare (v. Willis, 1952 e 19674), o produrre trombi neoplastici privi di ulteriore evoluzione nel senso di metastasi (v. Baserga e Saffiotti, 1955). L'impianto e lo sviluppo di tumori secondari da cellule neoplastiche circolanti nell'organismo (qualunque sia la via) e arrestatesi in un punto, rimane pertanto la fase meno conosciuta della metastatizzazione, ma é intuitivo che specialmente in questa fase grande importanza debbano avere i fattori biologici dell'idoneità del terreno e dell'organospecificità (v. Squartini e altri, 1968).
c) Distribuzione delle metastasi al tavolo anatomico
Il patologo, che al tavolo anatomico ha l'opportunità di registrare i fatti della diffusione metastatica in ciascun caso di tumore maligno, è più di ogni altro consapevole di quanto sia difficile armonizzare le singole osservazioni sopra un così complesso fenomeno biologico, e di come sia impossibile costringere i fatti osservati entro schemi troppo rigidi. Con il progresso delle conoscenze, il materiale autoptico rimane il test naturale di ogni nuova interpretazione o ipotesi a riguardo. Nella tab. IV sono riassunti i dati di un'ampia indagine personale illustranti la situazione della diffusione metastatica relativa a 730 neoplasie maligne osservate a Perugia nel corso di 717 autopsie consecutive per tumori maligni (v. Squartini e altri, 1968). Al primo posto, dopo i linfonodi, come sede di metastasi e il fegato con 187 casi, seguito dai polmoni con 112, dai reni, dai surreni e dalle ossa. Tali valori non si discostano sensibilmente, con rare e spiegabili eccezioni, da quelli rilevati in altre serie autoptiche (v. Willis, 1952, 19674 e 19733; v. Pingitore, 1972; v. Meissner e Warren, 19742) e costituiscono perciò un campione rappresentativo della diffusione metastatica dei tumori maligni al tavolo anatomico. Il campione riflette, sinteticamente, la complessità del fenomeno in esame e ne chiarisce meglio di ogni commento il grado di libertà biologica.
Vi sono tumori (come i cancri mammari) i quali, pur non essendo in posizione privilegiata rispetto al circolo sanguigno, metastatizzano con frequenza interessando un'ampia gamma di sedi, mentre ve ne sono altri i quali metastatizzano di rado e solo nelle sedi più tradizionali. Una favorevole ubicazione dei tumori primari nel circolo sanguigno conduce, ovviamente, a un aumento della loro attitudine alla metastatizzazione attraverso questa via: su 61 carcinomi del polmone con metastasi (affacciati senza interposizione di filtri al circolo generale) vi sono 63 localizzazioni metastatiche (una per tumore in media) in organi diversi da linfonodi, fegato e polmoni; su 191 carcinomi con metastasi ubicati in distretti tributari delle vene cave (cioè separati dal circolo generale per l'interposizione del filtro polmonare) il numero di metastasi osservate all'infuori dei linfonodi, fegato e polmoni è 88 (0,5 per tumore); su 188 cancri con metastasi situati in regioni dell'apparato digerente che sono tributarie della vena porta (in svantaggio anche maggiore rispetto ai precedenti, poiché gli eventuali emboli neoplastici debbono superare due filtri per raggiungere il circolo generale) le metastasi, esclusi gli organi già detti, sono 63 (0,3 per tumore).
d) Fattori che influenzano la metastatizzazione
La preminenza dei grandi filtri capillari (fegato e polmoni) quali sedi di metastasi è evidente. Anche la loro importanza come punti di arresto degli emboli o delle cellule neoplastiche circolanti appare in certo grado confermata da quanto detto sopra e dai dati seguenti: su 188 carcinomi dell'apparato digerente, tributari del circolo portale, con metastasi riscontrate al tavolo anatomico, 100 (53%) avevano metastasi epatiche, mentre su 191 carcinomi di altre sedi, tributari delle due vene cave, con metastasi riscontrate al tavolo anatomico, solo 47 (24%) avevano metastasi epatiche; reciprocamente, nei primi le metastasi polmonari erano 27 (14%), nei secondi 48 (25%).
Ma se si considerano le dimensioni degli organi colpiti, per esempio dividendo il numero di metastasi osservate in ciascun organo per il suo peso medio in grammi, la graduatoria di frequenza delle localizzazioni metastatiche quasi si capovolge e il fegato o i polmoni arretrano verso gli ultimi posti.
Una graduatoria così elaborata vedrebbe infatti al primo posto l'ipofisi con un indice di 5, seguita nell'ordine da surreni (3,8), ovaie (0,5), tiroide (0,4), meningi (0,2), reni (0,2), pancreas (0,16), fegato (0,13), polmoni (0,13), milza (0,09), utero (0,08), cuore (0,07), testicoli (0,04), cervello (0,01), ecc. Colpisce, fra le altre cose, in questa graduatoria la presenza di quattro ghiandole endocrine ai primi quattro posti.
Altro fatto evidente è la netta predilezione di alcuni tumori per particolari terreni di metastatizzazione. Delle 42 metastasi nei surreni complessivamente osservate, 19 (45%) appartengono a cancri del polmone, i quali costituiscono poco più del 10% del materiale esaminato. Su 7 metastasi ovariche osservate, 4 riguardano il cancro dello stomaco. Queste predilezioni si possono spiegare solo in parte con la situazione dei tumori primitivi rispetto al circolo sanguigno, o con peculiari vie di metastatizzazione. Del resto, la possibilità di selezionare da un tumore cellule con specificità di sede dell'impianto metastatico è stata affrontata e in certa misura dimostrata dalla patologia sperimentale (v. Stansly e Sato, 1978).
L'intensità e l'ampiezza della diffusione metastatica dipendono infine in larga misura anche dal tipo del tumore primario. I melanomi mostrano il più alto indice di metastatizzazione (24 metastasi per 4 tumori) e una diffusione pressoché ubiquitaria (14 localizzazioni), seguiti dai sarcomi e poi dai carcinomi, tra i quali i tumori della mammella e del polmone occupano, come già si è detto, i primi posti per tali caratteri.
Le considerazioni svolte richiamano l'attenzione sulla molteplicità dei fattori di interesse per la metastatizzazione dei tumori. I dati presentati sottolineano infatti equamente, in armonia con la fioritura di risultati sperimentali, l'importanza meccanica dei filtri, l'importanza quantitativa dell'irrorazione di organi e tessuti, l'importanza biologica del terreno e, infine, l'importanza del tumore primario nello stabilire se, dove e quando si realizzerà nell'ospite una ripetizione metastatica a distanza (v. Squartini e altri, 1968).
Tali fenomeni sono tuttora al vaglio della ricerca per una delucidazione dei meccanismi causali. L'impatto degli ormoni, delle medicine, degli antigeni tumorali e dei fattori immunitari sulla diffusione metastatica, come quello della composizione chimica delle sostanze fondamentali dei tessuti riceventi sull'attecchimento metastatico, sono argomenti attuali di studio, i quali allargano le prospettive di indagine e forse schiudono la via a possibili tentativi terapeutici della metastatizzazione tumorale (v. Stansly e Sato, 1978).
11. Manifestazioni cliniche, problemi di diagnosi e di prognosi
a) Durata clinica
Per alcuni tumori, come quelli cutanei o di siti facilmente accessibili ed esplorabili (per es. la cervice uterina), la fase clinica può abbracciare, in situazioni ideali, quasi l'intero arco della malattia neoplastica. Ma nella maggior parte dei casi questa è limitata ancora oggi al tardo periodo della malignità e da ciò deriva la sua durata generalmente breve, la frustrazione del medico e il prevedibile insuccesso terapeutico. I tumori maligni non trattati uccidono il 75% dei pazienti entro un periodo che oscilla fra i 14 mesi per il cancro dell'esofago e i 46 mesi per il cancro della mammella (v. Foulds, 1969). Ma vi sono moltissime eccezioni. Talora il decorso è sorprendentemente protratto (v. Shimkin, 1951); altre volte è particolarmente breve. La sopravvivenza dei tumori dopo trattamento incompleto è pure variabile e imprevedibile, come del resto lo sono i risultati del trattamento nei singoli casi. Ciò ha condotto i clinici a distinguere empiricamente fra cancri ‛buoni' e ‛cattivi', cioè ‛curabili' e ‛non curabili', attributi cui la ricerca biologica cerca di trovare una base che potrebbe risiedere anche in una diversa modalità di progressione (v. Foulds, 1969).
b) Comportamenti particolari
Alla base della fatalità dei tumori maligni in fase clinica, sta un comportamento progressivo e irreversibile che tuttavia può avere le sue eccezioni, rappresentate dai tumori occulti, dai tumori latenti, dai tumori dormienti e dai tumori che regrediscono. I ‛tumori occulti' sono quelli che si rivelano clinicamente per le loro metastasi essendo il focolaio primario piccolo o addirittura microscopico. Ciò può accadere in ogni organo, ma è comune per i carcinomi papillari della tiroide i cui pazienti spesso si rivolgono al medico per una metastasi palpabile in un linfonodo laterocervicale (v. Squartini e Severi, 1964). I ‛tumori latenti' si identificano con quei foci di carcinomi insospettati che non raramente si riscontrano all'autopsia nella prostata, o in altre sedi, in soggetti clinicamente esenti da neoplasie (v. Franks, 1956). Di questi, come dell'eccezionale regressione spontanea delle neoplasie maligne (v. AA.VV., Spontaneous regression..., 1976), si è già parlato. Il concetto di ‛tumori dormienti' si riferisce invece alle recidive locali o metastatiche a distanza di tempo eccezionale dalla rimozione del tumore primario, come 10-15 anni o addirittura più, fino a 40-50 anni. Questo stato dormiente delle cellule tumorali, del quale la ricerca biologica sta tuttora esplorando i possibili motivi, è tuttavia anch'esso evento raro, a livello di eccezione.
c) Fattori morfologici di prognosi
In genere, dopo la rimozione chirurgica, seguita o meno da terapia radiante o da chemioterapia, le recidive si manifestano nel giro dei primi 5 anni o al massimo entro 10 anni. Vi sono vari elementi anatomoclinici e istologici che guidano alla prognosi e alla terapia. Fra questi sono più importanti il livello di infiltrazione o di diffusione locale del tumore, la presenza di metastasi nei linfonodi regionali, la reazione linfatica e infiammatoria attorno alla neoplasia, lo stato del connettivo, il grado di malignità citologica e istologica, l'istotipo tumorale (v. Staquet, 1975; v. Fisher e altri, 1975). Purtroppo il livello di guarigioni da cura delle neoplasie maligne non è ancora soddisfacente e spesso accade che il tumore sia troppo avanzato per essere sottoposto a terapie chirurgiche radicali.
d) Diagnosi precoce
Da ciò discende la necessità di una diagnosi precoce che non può ottenersi con i mezzi tradizionali. Questi si basano sulla presenza del tumore come massa abnorme che provoca disturbi (da compressione, infiltrazione, ostruzione) ed è identificabile con vari metodi obiettivi, dalla palpazione, alla dimostrazione radiologica o scintigrafica. Ma a questa fase della neoplasia maligna ben raramente le terapie sono efficaci. La ricerca dei segni precoci di neoplasia già da tempo si è perciò indirizzata per altre vie rivolte alla identificazione di cellule atipiche eliminate nei liquidi organici e nelle secrezioni e di prodotti precoci del metabolismo tumorale o di antigeni tumorali, che potrebbero rappresentare utili contrassegni precoci morfologici o biologici della presenza di una neoplasia.
e) Sindromi paraneoplastiche
È noto che le cellule neoplastiche oltre a moltiplicarsi svolgono attività più squisitamente metaboliche che possono dar luogo a produzione di sostanze o secrezioni. Queste sostanze anzi sono talora responsabili di sintomi precoci e poco conosciuti della neoplasia in sviluppo, noti col nome di ‛sindromi paraneoplastiche'. Per sindromi paraneoplastiche si intendono quelle manifestazioni cliniche legate alla presenza di un tumore ma non direttamente imputabili ai fenomeni ostruttivi o invasivi della massa tumorale o delle metastasi, che regrediscono con la rimozione o la cura del tumore. Si tratta in genere di manifestazioni dolorose neurologiche, muscolari, osteoarticolari, oppure di manifestazioni endocrine e dismetaboliche, cutanee, ematologiche, cardiovascolari, dovute più spesso a tumori del polmone, a linfomi, a tumori del rene, ma presenti anche in altri tumori viscerali (v. Greenberg e altri, 1964). Benché non molto frequenti, è importante sottolineare che le sindromi paraneoplastiche sono spesso la prima e unica manifestazione della malattia, per cui su questa base un medico attento potrebbe in certi casi fondare una diagnosi precoce. Il paradosso di queste sindromi sta tuttavia nel fatto che mentre in alcuni casi possono rivelare un tumore occulto, altre volte assumono un ruolo di primo piano tanto da mascherare i fenomeni relativi alla neoplasia di cui sono emanazione, conducendo del tutto fuori strada (v. Anglesio, 1973).
f) Marcatori biologici e morfologici
Numerose indagini recenti hanno tentato di rendere più concreta e razionale la ricerca dei marcatori biologici precoci dei tumori. E stata così identificata una serie di prodotti e neoantigeni delle cellule tumorali nel siero o nelle urine di pazienti portatori, quali l'antigene carcino-embrionale (CEA = carcinoembryonic antigen) particolarmente nei soggetti portatori di cancro intestinale (v. i contributi di Gold e Freedman, 1965; v. Zamcheck e altri, 1972; v. immunologia e immunopatologia: Malattie immunoproliferative), ma anche in altri tipi di cancro (v. Chu e Nemoto, 1973); la caseina in donne con cancro mammario (v. Hendrick e Franchimont, 1974); l'idrossiprolina, particolarmente per la diagnosi precoce e il controllo successivo delle metastasi (v. Bondy e altri, 1974), ecc. L'antigene carcinoembrionale, che fu il primo marcatore biologico identificato, suscitò molte speranze, oggi purtroppo ridimensionate. Si tratta di antigeni normalmente presenti negli organi fetali (donde il nome di embrionali) che ricompaiono nelle cellule sdifferenziate del carcinoma. Sfortunatamente, le possibilità di dimostrarli nei soggetti con tumori si sono rivelate direttamente proporzionali alle dimensioni del tumore, per cui la positività è scarsa nelle lesioni piccole e iniziali. Con ciò viene meno l'interesse a questo test per una diagnosi precoce, mentre esso sembra oggi più utile per valutare la prognosi, la radicalità dell'intervento chirurgico, e la comparsa di recidive o di metastasi a questo conseguenti (v. Dhar e altri, 1972; v. Robbins, 1974).
I mezzi citologici di diagnosi precoce del cancro, iniziati con lo studio degli strisci vaginali e poi applicati a tutti i liquidi e secrezioni organiche (espettorati, urine, secrezione gastrica, ecc.), sono in uso già da molto tempo (v. Koss, 19682) per cui se ne possono valutare i risultati a distanza. Nel carcinoma della cervice uterina lo striscio vaginale consente di seguire a livello citologico la progressione tumorale, e la sua applicazione su larga scala a donne che volontariamente e periodicamente si sottopongono all'esame ha portato, attraverso diagnosi precoci e interventi limitati, a una riduzione sensibile e talora drammatica dei carcinomi cervicali invasivi nel materiale clinico e quindi della mortalità per questo carcinoma (v. Dunn, 1958; v. Boyes e altri, 1962). Vi è quindi chi prevede, per tale via, la scomparsa di certi tipi di cancro come causa di morte, ma subentrano problemi di costi sociali per ora irrisolvibili e neppure mancano note di scetticismo (v. Spriggs, 1972) circa le vite che in tal modo effettivamente possono essere salvate. Ciò probabilmente è in rapporto al fatto che la progressione dei tumori non sempre è graduale, ma può avvenire anche per salti improvvisi e i tumori a insorgenza più rapida sono spesso anche quelli a più rapida e maligna evoluzione (v. Foulds, 1975).
12. Stato attuale della terapia
La terapia di elezione in tutti i casi di tumori maligni localizzati che si possono asportare rimane l'intervento chirurgico coadiuvato dalla radioterapia, la quale rappresenta anche un'alternativa all'intervento chirurgico stesso nel caso di tumori localizzati che non si possono asportare radicalmente. Queste terapie vengono pertanto ancora oggi impiegate nella cura della maggior parte dei tumori, quali le neoplasie del tubo digerente (stomaco, intestino, esofago), dell'apparato respiratorio (laringe, bronchi, polmoni), dell'apparato genitale (mammella, utero, prostata, testicoli, ovaie), del sistema nervoso, ecc.
È tuttavia evidente dai risultati che queste terapie sono decisamente insufficienti ai bisogni. L'esperienza basata su 20 anni di trattamento del cancro mammario ha condotto a concludere che le pazienti possono essere separate in tre gruppi: il 45% di esse muoiono qualunque sia il trattamento che ricevono, il 45% sopravvivono qualunque sia il trattamento che ricevono (se cioè più o meno radicale) e solo il rimanente 10% mostrano di essere influenzate nel decorso della malattia dal trattamento che ricevono (v. Atkins, 1969; v. Handley, 1972; v. Foulds, 1975).
Nel caso di tumori sistemici, come sono ad esempio le leucemie e i linfomi, o quando i tumori localizzati si diffondono attraverso gli impianti metastatici a distanza, le terapie suddette non sono ulteriormente utilizzabili e subentra così l'esigenza di altre terapie (v. chemioterapia antineoplastica).
a) Chemioterapia
La ricerca di farmaci efficaci contro i tumori ha origini lontane, ma solo negli ultimi due decenni ha conseguito, con impostazioni più razionali, risultati apprezzabili, seppure nel complesso ancora modesti. Sono stati sintetizzati farmaci rivelatisi utili nel prolungare la sopravvivenza in certi tipi di neoplasie umane. Sono state sperimentate con successo procedure di trattamenti combinati, cioè con associazioni di più farmaci. Per tumori particolari, come il linfoma di Burkitt e il corionepitelioma, sono state talora segnalate addirittura guarigioni, probabilmente dovute però a particolari situazioni immunologiche (v. Burkitt e Wright, 1970; v. Bagshawe, 1969). La chemioterapia, divenuta così in pochi anni la principale risorsa terapeutica nelle leucemie, si pone come valida alternativa alla radioterapia nelle varie forme di linfoma, e il suo uso si va estendendo ogni giorno di più come trattamento secondario di copertura, dopo l'intervento chirurgico, o come trattamento primario, quando le neoplasie sono inoperabili e diffuse, nella maggior parte dei tumori solidi comuni, quali il cancro della mammella, dello stomaco, del polmone, ecc. (v. Stock, 1978).
Per la selezione e la prova di agenti chimici con azione antineoplastica ci si serve ampiamente, prima della sperimentazione clinica, di modelli sperimentali di tumori prevalentemente rappresentati da tumori trapiantabili nei comuni animali di laboratorio (leucemia murina, tumore ascite di Ehrlich del topo, tumore di Walker del ratto, ecc.; v. Schnitzer e Hawking, 1966). Purtroppo, per le neoplasie umane più comuni, che sono i carcinomi derivati da epiteli ghiandolari, non si dispone di modelli sperimentali adatti. Si sta perciò tentando il saggio diretto dei farmaci su tumori umani trapiantati in animali resi tolleranti mediante immunosoppressione (v. Cobb, 1972; v. Berenbaum e Sheard, 1972). I test in vitro, che al momento non sono ritenuti sostituti adeguati della sperimentazione in vivo, avranno prevedibilmente largo sviluppo in futuro (v. Hudson, 1972). Prima delle prove cliniche sui pazienti, i farmaci dimostratisi attivi e maneggevoli nei saggi sperimentali sono naturalmente sottoposti ad accurate indagini tossicologiche e farmacologiche (v. Stock, 1978).
b) Agenti chemioterapici
I farmaci antiblastici più comunemente usati comprendono gli agenti alchilanti, gli antimetaboliti, gli antibiotici, prodotti vegetali e alcuni enzimi, seguiti da una miscellanea di agenti diversi.
Gli agenti alchilanti utilizzati in terapia appartengono in prevalenza alle N-aloetilammine o mostarde azotate, alle etileneimmine, alle epossidi e agli esteri dell'acido sulfonico. Questo gruppo di farmaci comprende, oltre alla mostarda azotata, il BCNU (bis-cloroetilnitrosourea), il CCNU (doroetilcicloexilnitrosourea), il clorambucil, la ciclofosfammide, il melfalan, il merofan, il degranol e il tiotepa (v. Stock, 1978).
Gli antimetaboliti di maggiore uso comprendono il metotrexate, il 5-fluorouracile, la 6-mercaptopurina, la 6-tioguanina e la citosina-arabinoside. Gli antibiotici più attivi contro le neoplasie, prodotti da varie specie di streptomyces, comprendono l'actinomicina D, l'adriamicina, la daunomicina, la rubidomicina, la mitramicina e la streptozotocina. Ai prodotti vegetali antineoplastici appartengono gli alcaloidi della Vinca, farmacologicamente noti come vinblastina e vincristina. Fra gli enzimi con proprietà antineoplastiche sono da ricordare la L-asparaginasi e la carbossipeptidasi (v. Stock, 1978).
c) Meccanismi di azione
Gli agenti alchilanti sono nucleofili e capaci di interagire con molti costituenti cellulari (v. Ross, 1962). L'attività antineoplastica sembra comunque prevalentemente dovuta alla loro interazione col DNA nucleare (v. Roberts, 1978). Alcuni agenti alchilanti interagiscono direttamente con le macromolecole, mentre altri debbono essere prima metabolicamente attivati. I metaboliti sono spesso molto labili e perciò talora attivi solo nel distretto in cui vengono formati. Così la mostarda azo-bromica è attiva prevalentemente contro i tumori epatici (v. Bukhari e altri, 1973), il BCNU e il CCNU, mostarde azotate derivate dalla nitrosourea, sono impiegati nel trattamento dei tumori dell'encefalo (v. Carter e altri, 1972).
Gli antimetaboliti esercitano azione citotossica o citostatica. L'azione citostatica degli antifolici, di cui il metotrexate è il più noto esponente, si esplica attraverso un'inibizione dell'attività della dudrofolato-reduttasi, che impedisce la trasformazione in forma attiva dell'acido folico (v. Stock, 1978). Gli altri antimetaboliti citotossici esplicano la loro azione solo se trasformati metabolicamente in derivati attivi. La 6-mercaptopurina e la 6-tioguanidina sono i principali antagonisti delle purine. La citosina-arabinoside è il principale antagonista delle pirimidine (v. Roberts e Loehr, 1972).
Gli antibiotici con effetto antiblastico agiscono inibendo la sintesi dell'RNA o del DNA. L'actinomicina D inibisce selettivamente la sintesi dell'RNA DNA dipendente. Sembra che l'antibiotico legato all'acido nucleico impedisca in questo caso la progressione dell'RNA polimerasi lungo il DNA (v. Stock, 1978). Anche l'inibizione della sintesi dell'RNA da parte della mitramicina è dovuta al legame che questo antibiotico contrae con il DNA (v. Mihich, 1971). La daunomicina, la rubidomicina e l'adriamicina inibiscono la sintesi del DNA o quella dell'RNA secondo il sistema utilizzato (v. Bernard e altri, 1969; v. Meriwether e Bachur, 1972).
La vinblastina e la vincristina sono, come le colchicine e derivati, veleni della metafase. La loro azione antimito- tica sembra essenzialmente dovuta all'inibizione della sintesi degli acidi nucleici (v. Livingston e Carter, 1970). La L-asparaginasi è l'unica, fra le sostanze antineoplastiche, che sfrutta per l'azione una differenza qualitativa fra cellule normali e neoplastiche, e precisamente la capacità delle prime e l'incapacità delle seconde di sintetizzare l'asparagina. Perciò grandi speranze furono riposte nell'uso di questo enzima per la terapia dei tumori umani, andate purtroppo per la maggior parte deluse (v. Grundmann e Oettgen, 1970). La carbossipeptidasi 61 è un enzima batterico che scinde il folato, inibendo la sintesi del DNA (v. Chabner e altri, 1972; v. Stock, 1978).
d) Protocolli terapeutici
Le neoplasie che più favorevolmente rispondono alla chemioterapia sono quelle caratterizzate da elevati indici mitotici e da tempi di duplicazione oscillanti fra 1 e 4 giorni (v. Skipper, 1971). Questa proposizione ha naturalmente varie eccezioni in pratica ma, nonostante ciò, la sensibilità delle cellule agli agenti chemioterapici è, in linea di massima, direttamente proporzionale al loro indice mitotico (v. Stock, 1978).
Molti degli agenti chemioterapici agiscono come cito- tossici o citostatici soltanto nelle fasi cellulari dei processi metabolici di sintesi del DNA e sono perciò detti fase- specifici. Per un effetto terapeutico efficace le cellule neoplastiche dovranno pertanto essere esposte a un'adeguata concentrazione del farmaco durante la fase di sintesi (fase S), che è appunto quella suscettibile. L'efficacia del trattamento antiblastico dipende perciò non solo dalla selettività o dal tipo del farmaco, ma anche in larga parte dalle modalità di somministrazione. Così ad esempio il metotrexate somministrato a dosi elevate ogni 4 giorni prolunga la sopravvivenza di topi leucemici più che se somministrato ogni giorno a piccole dosi (v. Goldin e altri, 1956) e lo stesso accade per altri farmaci. Le alte dosi spaziate nel tempo risultano inoltre meno tossiche di quelle basse somministrate quotidianamente, poiché in apparenza le cellule dell'ospite recuperano nel periodo di sospensione dallo stato tossico più rapidamente di quelle del tumore (v. Stock, 1978).
Questi studi sperimentali sulle cinetiche cellulari hanno consentito di programmare protocolli terapeutici adeguati ai diversi tipi di neoplasia. Nei tumori umani però varie conoscenze di base fanno difetto rispetto ai modelli sperimentali e ciò rende più difficile la terapia. Per molti tumori umani, ad esempio, il tempo reale di duplicazione cellulare non è noto e il tempo di duplicazione del volume tumorale, fra l'altro assai variabile da caso a caso, non è sempre un indice attendibile dei valori cinetici di una popolazione cellulare in mitosi (v. Steel e Lamerton, 1969). A causa di queste differenze il trattamento di due tumori apparentemente simili con lo stesso farmaco può dare risultati diversissimi (v. Stock, 1978).
L'associazione di più farmaci antineoplastici con proprietà biochimiche diverse consente di ridurre gli effetti tossici e di potenziare quelli terapeutici. Per questo, sebbene la chemioterapia con un solo farmaco trovi ancora indicazioni nel trattamento di qualche forma tumorale, l'associazione di due o più farmaci è divenuta negli ultimi anni la scelta di elezione per la maggior parte dei casi. In genere si uniscono un antimetabolita con un citostatico e un ormone, ma si sono sperimentate associazioni le più varie e complesse, anche di 4 o 5 farmaci insieme, comprendenti agenti alchilanti, antimetaboliti, antibiotici, citostatici vegetali e ormoni. I risultati migliori con trattamenti combinati si ottengono nelle leucemie e nei linfomi (v. Henderson e Samaha, 1969), ma la chemioterapia combinata si è estesa ormai al trattamento di molte altre neoplasie con risultati lusinghieri. Così, nel cancro della mammella in fase avanzata la chemioterapia combinata ha fornito risultati nettamente migliori di quelli ottenibili con sola terapia ormonale (v. Griem e altri, 1973).
Talora si può ottenere sinergismo terapeutico somministrando col citostatico un secondo farmaco che ne riduca la tossicità nei confronti dell'ospite consentendo così di aumentarne la dose. Ma sono stati segnalati anche fenomeni di antagonismo fra farmaci in associazione e di autoantagonismo, con conseguente riduzione dell'efficacia terapeutica. Per esempio, vi è antagonismo tra metotrexate e L-asparaginasi, come tra citosina-arabinoside e metotrexate (v. Orindey e Nichol, 1972). Si può parlare invece di autoantagonismo nel caso di un farmaco che, agendo su più cicli metabolici, può essere meno efficace proprio a causa di questa azione multipla (v. Stock, 1978). Un esempio è fornito ancora dal metotrexate il quale uccide le cellule inibendo la sintesi del timidilato dal deossiuridilato durante la fase S del ciclo cellulare, ma nel contempo sopprime anche la sintesi ex novo delle purine impedendo alle cellule di raggiungere, o progredire attraverso, la fase 5, che è quella sensibile all'effetto principale del farmaco rappresentato appunto dalla morte cellulare per carenza timinica (v. Borsa e Whitmore, 1969; v. Stock, 1978).
e) Limiti e danni della chemioterapia
Il principale limite alla chemioterapia dei tumori è nel fatto che non si dispone ancora di farmaci ad azione selettiva sulle cellule neoplastiche, per cui si deve ricorrere a sostanze chimiche le quali esplicano un'elevata azione citotossica nei confronti delle cellule normali dell'ospite. Si impone perciò sempre l'esigenza di ridurre e di limitare al massimo gli effetti indesiderati, ciò che va a detrimento dell'efficacia terapeutica sulla neoplasia. Quando è possibile si può aggirare l'ostacolo ricorrendo alla perfusione regionale del farmaco, una metodica che però è di rado applicabile, o all'immissione diretta del farmaco nella massa tumorale, ma con scarso successo.
L'accesso dei farmaci al tumore è un altro problema non secondario della chemioterapia antiblastica. La vascolarizzazione di molte neoplasie è irregolare e scarsa (v. Willis, 19674) e ciò rende particolarmente difficile un'efficace penetrazione dei farmaci entro masse neoplastiche sovente estese e frequentemente interrotte nella loro continuità da focolai di necrosi, nei quali tuttavia sopravvivono nidi di cellule neoplastiche.
Un altro problema fondamentale, e un limite finora invalicabile per la chemioterapia dei tumori, è quello della resistenza ai farmaci. Con rare eccezioni il trattamento chemioterapico determina quasi sempre all'inizio una remissione di settimane o mesi, dopodichè però la neoplasia riprende a crescere mostrando resistenza nei confronti del farmaco. Ciò rende necessario il ricorso ad altri citostatici, per i quali il fenomeno si ripete, e così gradualmente vengono precluse le possibilità di trattamento, anche perché la rosa dei farmaci disponibili è limitata. Se si riuscisse a chiarire nei suoi diversi aspetti il fenomeno della resistenza dei tumori ai farmaci e si scoprisse il modo per impedirne la comparsa, o meglio per volgerla a favore della terapia, non vi è dubbio che si trarrebbero enormi vantaggi per il paziente (v. Stock, 1978).
Molte indagini sono state svolte in proposito e vari fattori responsabili dello sviluppo di una resistenza ai farmaci sono stati individuati per i chemioterapici oggi in maggior uso. Una sintesi bibliografica recente ne ricorda 10, che sono: il ridotto assorbimento, la ridotta attivazione del farmaco, l'aumento del catabolismo del farmaco, o dei suoi metaboliti, l'aumentata produzione di molecole bersaglio, l'aumento della concentrazione intracellulare di molecole neutralizzanti il farmaco, la ridotta sensibilità dell'enzima bersaglio, lo sviluppo di vie o siti metabolici alternativi nelle cellule neoplastiche trattate, l'aumento nella capacità riparativa delle lesioni indotte dal farmaco e, per i trattamenti ormonali, l'alterazione del recettore (v. Stock, 1978).
Il trattamento con chemioterapici, che sono particolarmente tossici per le cellule del sangue, riduce notevolmente le difese immunitarie e la competenza immunologica dei pazienti, rendendoli particolarmente recettivi alle infezioni (v. Mathé, 1971). Per ridurre i pericoli sono stati proposti sistemi di isolamento dei malati, onde proteggerli dai microrganismi ambientali. Una pratica tuttavia non seguita da tutti, perché il paziente è di per sé portatore di agenti potenzialmente patogeni (nella flora batterica saprofitica intestinale, nel cavo orale, ecc.) per cui il rischio di contrarre un'infezione letale non diminuisce con l'isolamento.
f) Terapia ormonale
Le leucemie e i tumori delle ghiandole endocrine o di organi bersaglio terminali del sistema endocrino (mammella, utero) risentono in genere favorevolmente del trattamento con ormoni, specie nelle fasi iniziali. Nella terapia delle leucemie è generalizzato l'uso dei corticosteroidi (prednisone e prednisolone) per il marcato effetto linfocitolitico e per la generica azione antineoplastica che tali ormoni dimostrano. Le pazienti con cancro mammario in età premenopausale vengono generalmente sottoposte a terapia ormonale aggiuntiva (somministrazione di ormoni antiestrogeni come il testosterone e i progestinici, o di corticosteroidi) e/o sottrattiva (ovariectomia, talora associata a surrenectomia per ridurre la concentrazione ematica degli estrogeni) con validi risultati pratici e sperimentali (v. Currie e Illingworth, 1958; v. DeS ombre e altri, 1976). A tali terapie sono sensibili i tumori le cui cellule conservano recettori estrogenici e perciò si sono sviluppate tecniche per la dimostrazione di tali recettori nei tessuti neoplastici (v. McGuire e altri, 1975). Gli estrogeni vengono usati con successo nella terapia del carcinoma prostatico, i progestinici nella cura del cancro metastatico dell'endometrio (v. Stoll, 1972).
g) Immunoterapia
La capacità dei tumori di suscitare una risposta immunitaria nell'ospite e le possibili applicazioni terapeutiche di tale fenomeno sono già state discusse o accennate nel cap. 6. Si è già detto anche che alcuni tumori (corionepitelioma, linfoma di Burkitt, neuroblastoma, melanoma, ecc.) sono particolarmente suscettibili di regressione spontanea o conseguente a un'insufficiente chemioterapia, con probabilità a seguito di un'efficace reazione immunitaria del paziente. Tuttavia, i tentativi effettuati nell'uomo di stimolare le risposte immuni dell'ospite contro i tumori hanno finora dato risultati deludenti (v. Stock, 1978). E vi sono anche problemi sperimentali ancora aperti da risolvere prima di poter procedere a un'applicazione pratica dei principi (v. Alexander, 1978). È comunque possibile che in futuro l'immunoterapia, specie in combinazione con la chemioterapia, risulti utile in certi casi.
Finora si è sperimentata, con qualche risultato, la somministrazione di immunostimolanti aspecifici (Corynebacterium parvum o bacillo di Calmette e Guérin, un ceppo attenuato di micobatterio della tubercolosi) in particolari tumori sperimentali e nella leucemia umana (v. Mathé e altri, 1969; v. Powles e altri, 1973). Sono stati inoltre provati l'attacco di un gruppo citotossico su anticorpi antitumorali specifici (v. Ghose e altri, 1972), allo scopo di guidare selettivamente il chemioterapico sul bersaglio delle cellule neoplastiche, e l'autoimmunizzazione mediante cellule tumorali autologhe irradiate in pazienti con malattia neoplastica avanzata (v. Alexander, 1978).
h) Conclusioni
In conclusione la chemioterapia, valutando la generalità dei casi e non solo le favorevoli eccezioni, ha finora fornito risultati assai modesti offrendo sostanzialmente solo un prolungamento della vita dei malati al prezzo di intossicazioni e danni biologici purtroppo rilevanti (v. Sieber e Adamson, 1975). Tale terapia è dunque ancora alla ricerca di farmaci veramente efficaci, selettivi e pertanto non nocivi, come di meccanismi che precludano lo sviluppo di una resistenza ai farmaci. La terapia ormonale concede pure solo prolungamenti o temporanee remi ssioni. L'immunoterapia è in una fase ancora più sperimentale. Il problema cruciale della terapia dei tumori resta perciò tuttora largamente non risolto e la sua quotidiana manipolazione a opera dei medici rimane a un livello empirico che chiude ulteriori possibilità di approfondimento e discussione. Ed è logico che sia cosi, poiché non si conoscono ancora i meccanismi della trasformazione tumorale.
Attualmente si possono però offrire a tale problema quei contributi che speculativamente discendono dal concetto e dai principi della progressione tumorale, elencandoli in breve qui di seguito: a) non vi è dubbio, anzitutto, che la terapia medica avrebbe raggiunto un grande risultato se riuscisse a controllare e regolare la progressione dei tumori. Tutto lascia fin qui supporre che la progressione, sebbene forse inevitabile e inarrestabile, possa essere accelerata o ritardata da stimoli vari. Il problema è di identificarli; b) il principio della progressione indipendente dei caratteri tumorali suggerisce una ristrutturazione del grossolano criterio di ‛malignità', oggi considerato dai più come entità unitaria. Ciò ha i suoi riflessi pratici, perché è difficile pensare che una singola terapia si dimostri efficace contro tutti i caratteri tumorali (dipendenza, accrescimento, infiltrazione, metastasi, ecc.) capaci di progressione indipendente; c) il blocco dell'accrescimento tumorale, quale si può ottenere con vari agenti chemioterapici, non previene né impedisce la progressione, la quale è indipendente dall'accrescimento, e non è escluso che la favorisca; d) una migliore conoscenza del livello di progressione raggiunto dal tumore, mediante il perfezionamento di test biologici e analisi morfologiche, potrebbe utilmente indirizzare le cure; e) il controllo dei tumori dipendenti, quale si può ottenere con vari mezzi, è in sé di scarsa importanza pratica, perché la dipendenza da un determinato stimolo è evento transitorio e viene prima o poi perduta per progressione. Il problema centrale è di mantenere dipendenti per il maggior tempo possibile i tumori che lo sono, non di sfruttare questo loro carattere per una temporanea cura; f) che la progressione sia fenomeno irreversibile e inevitabile in un tumore non deve scoraggiare, visto che essa può non raggiungere il suo punto d'arrivo nell'ospite primario e che sembra possibile, sperimentalmente, rallentarla prolungando la malattia anche per lungo tempo; g) sul piano sperimentale, il primo suggerimento è di dedicare gli sforzi maggiori allo studio delle possibilità di interferire competitivamente, a livello genico, con i fattori che controllano la progressione tumorale. Da queste impostazioni, in mancanza per ora di conoscenze migliori, sembra giustificato attendere sviluppi più razionali per la terapia antiblastica di domani (v. Squartini, 1964).
13. Tumori dell'infanzia e dell'adolescenza
a) Peculiarità e problemi
Il panorama dei tumori dell'infanzia, dell'età puberale e della prima adolescenza e notoriamente molto diverso da quello consueto per gli altri periodi della vita. Nei bambini e negli adolescenti prevalgono i tumori cosiddetti embrionali, i teratomi, gli amartomi, i tumori degli organi emolinfopoietici, i sarcomi, mentre sono molto rari i carcinomi, che costituiscono il tipo più comune di tumore degli adulti. Le neoplasie dei primi anni di vita sollevano dunque una serie di problemi particolari, di ordine biologico, morfologico, epidemiologico e clinico, che riguardano i loro rapporti con l'embriogenesi, la natura e la provenienza dei fattori causali, l'istogenesi, l'identificazione e la classificazione dei diversi oncotipi, la distribuzione nelle comunità infantili di alcune forme a sospetta eziologia virale e la ricerca dei veicoli di un possibile contagio, l'eventuale profilassi sulle gestanti, il controllo, la diagnosi e il decorso, le terapie efficaci, ecc. Questi problemi particolari fanno dei tumori dell'infanzia un argomento a sé nel campo della moderna oncologia, che richiede specializzazione non solamente al clinico, ma anche al patologo e al biologo.
b) Epidemiologia e frequenza
In Italia, come in altri paesi a sviluppo analogo, i tumori si avviano a diventare la principale causa di morte nell'infanzia e nell'adolescenza. Con la diminuzione di molte malattie infettive che oggi si possono curare e col miglioramento delle condizioni di vita e sanitarie generali, la frequenza relativa di decessi per tumori nell'età infantile e nell'adolescenza mostra un progressivo sensibile aumento. Se nella prima infanzia, e particolarmente nel primo anno di vita, tale frequenza appare tuttora molto piccola, ciò si deve all'elevata mortalità perinatale per cause varie ancora non sufficientemente controllabili e prevenibili, ma certo destinate a ridimensionarsi nel tempo. In realtà, il numero dei decessi per tumori è maggiore nei primi cinque anni di vita che nei quattro quinquenni successivi. Solo fra 26 e 30 anni il numero dei tumori che causano la morte torna a un livello comparabile a quello che gli compete nel primo quinquennio. Il rapporto fra i morti per tumori prima e dopo i 18 anni è di circa 1:50. Se però è riferito al numero totale dei decessi che si verificano per ciascuna età, tale rapporto sale a 1:2, fatta eccezione per i primi due anni di vita a causa dei motivi detti sopra. Dal sesto anno in poi i tumori sono la seconda causa di morte nell'infanzia e nell'adolescenza. L'insieme dei fatti ricordati dimostra che i tumori dell'infanzia e dell'adolescenza non sono come a volte si pensa un'entità trascurabile, e il problema che essi rappresentano non è sul piano quantitativo molto diverso da quello dei tumori nelle età successive della vita. Di qui la necessità di non trascurarne lo studio, sotto ogni aspetto, anche se ciò è difficile e richiede particolare competenza.
Considerando gli apparati, gli organi o i sistemi colpiti, i tumori dell'infanzia e dell'adolescenza mostrano notoriamente predilezioni molto diverse da quelle peculiari ai tumori della maturità e dell'età senile. In Italia sono nettamente al primo posto fino a 18 anni i tumori degli organi emolinfopoietici (incluse le leucemie) responsabili da soli di circa il 55% dei decessi per tumori. Seguono a distanza i tumori del sistema nervoso (16%) e quelli delle ossa e dei connettivi (6%). Poi tutti gli altri, fra cui le neoplasie degli apparati digerente, genitale e respiratorio responsabili complessivamente e nei due sessi di circa l'80% dei decessi per tumori in soggetti oltre i 18 anni di età. Poche e poco marcate sono al riguardo le differenze con i dati resi noti per altri paesi. Non vi è dubbio, quindi, che le neoplasie dei bambini e degli adolescenti siano nettamente diverse sul piano genetico e qualitativo da quelle degli adulti. Per approfondire l'indagine in tal senso occorre tuttavia trasferirsi dal terreno delle statistiche sui certificati di morte a quello della patologia, considerando quei materiali che consentono lo studio morfologico dei casi.
c) Quadri di patologia
La patologia dei tumori dell'infanzia e dell'adolescenza offre difficoltà numerose sul piano della definizione, della classificazione, a volte persino del riconoscimento delle varie forme. Incerti sono talora i limiti fra malformazioni e tumori, fra neoplasie congenite e acquisite, fra tumori benigni e maligni. Fra l'altro nei bambini sono più frequenti rispetto agli adulti i tumori benigni che causano la morte (per la sede, per l'estensione, per le complicazioni; v. Andersen, 1951). Vi sono ampie possibilità di errori diagnostici per chi non abbia familiarità con l'embriologia, con i problemi morfologici dello sviluppo così mutevoli e diversi da tessuto a tessuto, con la vastità delle modificazioni che ai livelli quantitativo e qualitativo le cellule di un tessuto possono subire in condizioni funzionali diverse (si pensi alle cellule della serie bianca del sangue) senza oltrepassare i confini della norma. Anche il concetto di progressione tumorale (v. Foulds, 1954) perde in certi casi ogni valore, potendosi assistere nei bambini alla maturazione graduale di tumori prima maligni che avevano dato metastasi (v. Fox e altri, 1959). L'impressione più viva che si ricava dal quadro generale è quella della confluenza nell'infanzia e nell'adolescenza di categorie di tumori fra loro molto diversi nell'origine, nella morfologia, nel comportamento, nelle distribuzioni e anche nelle cause. I tumori cosiddetti embrionali dei primi anni, per esempio, non hanno nulla in comune con certe neoplasie della pubertà e dell'adolescenza che preludono alla patologia tumorale degli adulti (v. Squartini e Bolis, 1966).
Articolare in una classificazione semplice e utile alla pratica categorie di tumori così diversi non è facile e per riuscirvi è necessario considerare, accanto all'istogenesi e al comportamento, criteri di cronologia e di frequenza delle neoplasie. Infatti, è giusto riservare il primo posto nella considerazione a quei tumori, frequenti nell'infanzia, che compaiono per primi: i tumori embrionali i quali, ordinati in graduatoria di frequenza, comprendono i tumori dei tessuti neurali (neuroblastomi, medulloblastomi e neuroepiteliomi, retinoblastomi, ecc.), i tumori embrionali dei visceri (nefroblastomi, epatoblastomi, orchioblastomi, ecc.) e i sarcomi embrionali. Dopo i tumori embrionali appare logico considerare i teratomi (neoformazioni derivate da più foglietti embrionali), perché a essi affini, e poi gli amartomi (sviluppo in eccesso di una componente di un tessuto) con i relativi tumori e i tumori delle vestigia embrionali, perché tutti derivati da tessuti che hanno subito un disturbo nel corso del loro sviluppo. Sistemate così le classi di neoplasie più tipiche dell'infanzia sul piano istogenetico, restano tutte le altre, che non sono diverse da quelle corrispondenti degli adulti e comprendono, in ordine di frequenza, i tumori dei tessuti emopoietici, quelli dei tessuti scheletrici, gli altri tumori mesenchimali, i tumori delle ghiandole endocrine e delle gonadi, e gli altri tumori epiteliali derivati da tessuti non endocrini. Questa classificazione (v. Willis, 1962) ha numerosi pregi e solo trascurabili difetti.
d) Curve di distribuzione
Nella tab. V e nei relativi diagrammi è illustrata la distribuzione di un ampio materiale patologico, bioptico e autoptico, da me elaborato, comprendente 579 tumori dell'infanzia e dell'adolescenza osservati a Perugia in soggetti da 0 a 16 anni e catalogati secondo il tipo istologico e l'età (v. Squartini e Bolis, 1966; v. Severi e Squartini, 1969). Nella distribuzione secondo il sesso (non riportata) c'è un'evidente predilezione dei tumori di derivazione mesenchimale per il sesso maschile e di quelli di origine epiteliale per il femminile, mentre i tumori più propriamente embrionali e assimilati sono assai equamente distribuiti fra i due sessi. La distribuzione secondo il tipo istologico e l'età è corredata da grafici esplicativi. Per analizzare le varie componenti che intervengono nella composizione della curva di distribuzione totale dei tumori, sono state tracciate separatamente anche le curve di distribuzione parziali dei tumori inclusi nelle classi morfologiche e istogenetiche considerate, riunite allo scopo in quattro gruppi secondo criteri di omogeneità e precisamente: a) le classi da 1 a 6 che comprendono tutti i tumori embrionali e i teratomi accanto a poche altre forme derivate da tessuti neurali differenziati; b) le classi 7 e 8 che includono gli amartomi, i tumori da questi derivati e quelli che prendono origine dalle vestigia dello sviluppo embrionale; c) le classi da 9 a 11 che comprendono tutti i tumori dei tessuti emopoietici, ossei e mesenchimali in genere; d) infine le classi da 12 a 14 che includono i tumori degli organi endocrini, delle gonadi, della mammella e i tumori epiteliali degli altri organi e apparati.
Esaminando le curve di distribuzione di questi quattro raggruppamenti di tumori secondo l'età si osserva che i tumori del primo e del secondo gruppo compongono tracciati regolarmente e rapidamente decrescenti con picchi di massima frequenza nel primo anno di vita, i tumori del terzo disegnano un tracciato irregolare con picchi a 1, 4, 7 e 14 anni corrispondenti a onde di ampiezza diversa e massima nell'ultimo quinquennio, mentre i tumori del quarto gruppo si allineano lungo una curva regolare ma capovolta rispetto alle prime due, con picco di massima frequenza al sedicesimo anno.
È la combinazione di questi differenti tracciati che condiziona il caratteristico andamento del grafico di distribuzione secondo l'età dei tumori dell'infanzia e dell'adolescenza nel loro complesso, espresso da una curva assai regolare ad andamento ellittico con concavità superiore, per due punti di massima frequenza quasi equivalenti in corrispondenza del primo e del quindicesimo-sedicesimo anno di vita, separati da una profonda depressione intermedia. Nella parte centrale della curva, in corrispondenza cioè del periodo di minima frequenza dei tumori, sono evidenti e riconoscibili alcuni picchi minori che deviano dall'andamento altrimenti regolare del tracciato, dovuti alle neoplasie dei tessuti emopoietici, ossei e mesenchimali in genere (v. Squartini e Bolis, 1966).
e) Possibili fattori causali
Senza volere qui entrare nel merito dei problemi eziologici dei tumori dell'infanzia e dell'adolescenza, per parlare dei quali non si dispone di dati sufficienti, è lecito tuttavia estrarre dalle curve presentate quello che con chiarezza suggeriscono. Nell'ambito delle neoplasie dell'infanzia si possono riconoscere quattro categorie principali di tumori che appaiono fra loro indipendenti sotto vari aspetti e per le quali è giustificato supporre anche una diversa eziologia.
I tumori embrionali e i teratomi, i quali si sviluppano da tessuti indifferenziati e sono concentrati nel primo o nei primi anni di vita, debbono riconoscere fattori causali che agiscono prima della nascita disturbando e deviando in senso neoplastico i normali processi di sviluppo e maturazione dei tessuti del feto e dell'embrione.
Gli amartomi e i tumori che prendono origine da questi o dalle vestigia dello sviluppo embrionale sono nel comportamento analoghi ai precedenti. Con quelli condividono la massima frequenza nei primi anni di vita, sebbene alcuni si manifestino solo più tardi. Similmente a quelli debbono riconoscere fattori causali che agiscono o prima della nascita, ma su tessuti giunti ormai al termine della loro fase di maturazione causandovi uno squilibrio di rapporti quantitativi (amartomi), o anche dopo la nascita, ma su residui di tessuti mantenutisi allo stato embrionale (tumori delle vestigia embrionali).
Per i tumori dei tessuti emopoietici, ossei e mesenchimali in genere, che si manifestano a ondate con numerosi picchi di frequenza corrispondenti all'incirca ai periodi di maggiore accrescimento corporeo postnatale, sembra giustificato pensare a fattori causali di natura particolare i quali agiscono con probabilità dopo la nascita sopra tessuti ai più alti livelli della loro attività funzionale o di sviluppo.
Infine, i tumori degli organi endocrini, delle gonadi, della mammella e gli altri tumori epiteliali in genere, praticamente assenti nell'infanzia e concentrati al massimo nelle età della pubertà e dell'adolescenza, debbono riconoscere fattori causali non diversi da quelli ai quali si attribuisce la maggiore importanza per lo sviluppo di tumori negli adulti, alcuni di natura ormonale, altri ambientale (v. Squartini e Bolis, 1966).
Quali siano i fattori causali dei tumori che si sviluppano nel corso dell'infanzia rimane comunque fino ad ora sconosciuto. La suscettibilità dei tessuti embrionali e neonatali ai cancerogeni trasmessi dalla madre o direttamente applicati (v. Shay e altri, 1950; v. Klein, 1952; v. Heston e Staffee, 1957; v. Kelly e O'Gara, 1961), come pure l'origine virale di alcune fra le neoplasie più comuni nelle prime età di vita (v. Gross, 1961 e 19702), sono fatti chiaramente dimostrati dalla patologia sperimentale. Nella patologia umana questi fatti rimangono tuttavia ancora per la maggior parte solo stimolanti argomenti di indagine e di speculazione.
14. Principali tipi di tumori maligni negli adulti
Non è possibile, nè sarebbe utile agli scopi di questa trattazione, riferire su tutti i tipi di tumori maligni umani. Nei paragrafi seguenti si descriveranno pertanto solo i principali e di questi i caratteri meglio adatti all'esemplificazione di problemi.
a) Cancro della mammella
La mammella, come organo differenziato, esiste soltanto in funzione degli stimoli ormonali che riceve. Sotto questo profilo essa rappresenta anzi il più tipico esempio nell'organismo di struttura dinamica. In rapporto alla pubertà, ai cicli mestruali, alle gravidanze, agli allattamenti, quest'organo terminale del sistema endocrino converte gli stimoli ormonali che riceve in strutture morfologiche e così aumenta l'ampiezza del suo albero ghiandolare, modifica i confini coi tessuti limitrofi, perfeziona la differenziazione istologica, adatta la struttura alle esigenze funzionali (v. Squartini e Olivi, 1962).
A causa di questo dinamismo strutturale, l'albero ghiandolare mammario è fatto di strutture transitorie e permanenti, ciò che apre nuovi problemi alla cancerogenesi mammaria, sconosciuti a livello di altri organi. In larga parte, ogni cellula epiteliale di una mammella a riposo è la depositaria potenziale di una complessa struttura ghiandolare. Reciprocamente, molte strutture ghiandolari di una mammella funzionante sono completamente riassorbite in breve tempo.
Ciò può spiegare la mancanza di informazioni precise perfino riguardo alla sede di origine del cancro mammario. La maggior parte dei carcinomi mammari infiltranti sono usualmente ritenuti di origine dottale extralobulare (v. McDivitt e altri, 1968). Ma studi sul cancro mammario sperimentale e umano suggeriscono che la più comune sede di origine possa invece essere il lobulo mammario (v. Squartini, 1959 e III Histogenesis..., 1977; v. Wellings e altri, 1975). Indagini recenti, basate sullo studio delle lesioni submacroscopiche nelle sezioni di mammelle umane intere, indicano che i focolai più precoci di carcinoma mammario sono localizzati nelle unità terminali dottali intralobulari (v. Wellings e Jensen, 1973). Ciò è importante in considerazione del controllo ormonale delle strutture lobulari e può spiegare perché il cancro della mammella è così comune nelle femmine e così raro nei maschi, i quali raramente hanno lobuli mammari a meno che non siano sottoposti per lungo tempo a trattamento con estrogeni (v. O'Grady e McDivitt, 1969).
Gli ormoni che presiedono allo sviluppo mammario sono l'estrogeno, il progesterone e la prolattina. Questa triade ormonale è in grado di produrre un pieno sviluppo dell'albero ghiandolare mammario con differenziazione lobulo-alveolare negli animali ovariectomizzati, surrenectomizzati e ipofisectomizzati (v. Cowie e Folley, 1958). Gli stessi ormoni endogeni, cioè la prolattina ipofisaria e gli steroidi ovarici e/o surrenali, sono strettamente implicati anche nello sviluppo del cancro mammario. L'ovariectomia precoce riduce fortemente o annulla addirittura il rischio di cancro della mammella (v. Feinleib, 1968). Ma vari esperimenti hanno dimostrato che livelli normali di ormoni, dunque non necessariamente livelli aumentati o esagerati, sono sufficienti per la cancerogenesi mammaria (v. Bulbrook, 1971 e 1977). Inoltre, i dati raccolti da esperimenti di trapianto nel topo suggeriscono che le cellule epiteliali mammarie trasformate dall'azione di cancerogeni chimici o virali possono trovare il supporto necessario allo sviluppo delle neoplasie nell'ambiente ormonale di ospiti endocrinologicamente normali (v. Brennan, 1975).
Sul piano clinico è ben nota l'azione protettiva delle gravidanze e degli allattamenti (v. Severi e Squartini, 1962), in particolare della prima gravidanza (v. MacMahon e altri, 1973), nei confronti del cancro mammario. Sono così stati identificati e catalogati numerosi fattori di rischio generici predisponenti al cancro mammario che includono: la nulliparità, la primiparità attempata, il mancato allattamento al seno, il menarca precoce e la menopausa tardiva (v. Tuyns, 1971; v. MacMahon e altri, 1973). Il progressivo aumento di tali fattori di rischio nel corso di questo secolo è responsabile apparentemente del forte aumento registrato nelle frequenze di cancro mammario dei paesi occidentali. Accanto ai fattori di rischio generici, ve ne sono altri più importanti, detti specifici, in quanto segnalano un alto rischio locale di cancro. Questi comprendono: la familiarità (cioè presenza di più cancri mammari nelle parenti), la displasia mammaria e i papillomi intraduttali multipli (ritenute lesioni preneoplastiche), il carcinoma lobulare in situ (una lesione considerata di sicuro significato preinvasivo) e il carcinoma della mammella controlaterale (v. Haagensen, 1971).
Sul piano morfologico, i problemi del cancro mammario riguardano ancora la classificazione, che attualmente è riferita alle sedi di origine (dai lobuli, dai dotti; ma si è già detto con quanta approssimazione), all'aspetto invasivo (cancri in situ e infiltranti), al tipo cellulare, e alle reazioni dell'ospite (proliferazione connettivale o fibrosi produttiva, infiltrati infiammatori, ecc.; v. McDivitt e altri, 1968). Le indagini ultrastrutturali tendono a ricollegare i diversi tipi di cancro con gli stipiti cellulari normali di origine che sono essenzialmente tre: le cellule secernenti dei lobuli, le cellule di rivestimento dei dotti, e le cellule mioepiteliali esterne, con funzioni di contrazione sulle precedenti per l'espulsione del secreto (v. i contributi di Murad, 1971). Altri problemi di interesse per il cancro mammario sono già stati discussi nei capitoli precedenti.
b) Cancro del polmone
Almeno tre quarti dei tumori polmonari nell'uomo prendono origine dai grossi bronchi all'ilo o in prossimità dell'ilo (v. Willis, 19674), mentre soltanto un numero molto limitato, probabilmente inferiore al 5%, ha una localizzazione periferica, sottopleurica, e un'origine bronchiolare o alveolare (v. Decker, 1955). Questa distribuzione non omogenea dei tumori lungo i rivestimenti epiteliali dell'albero respiratorio può dipendere sia da una diversa sensibilità, sia da una diversa esposizione degli epiteli bronchiale e alveolare agli agenti cancerogeni specifici. La prima di tali due ipotesi tuttavia si è dimostrata erronea all'indagine sperimentale (v. Squartini e Bolis, 1971). La seconda ipotesi è quindi molto più realistica e in accordo con l'opinione, generalmente condivisa, che la maggior parte dei tumori del polmone nell'uomo sono dovuti a cancerogeni malati. Infatti la maggior parte delle particelle presenti nell'aria malata si arrestano nell'impatto con le mucose delle vie respiratorie maggiori e in particolare dei grossi bronchi. Gli epiteli alveolari sono invece in larga misura sottratti all'azione del pulviscolo e molto più in contatto con eventuali agenti trasportati dal sangue circolante.
Uno dei fatti più evidenti, nel panorama anatomo-clinico del cancro del polmone è il suo straordinario aumento di frequenza negli ultimi decenni. Sembra logico, in base anche a quanto detto sopra, mettere tale fenomeno in rapporto con le crescenti quantità di cancerogeni malati dall'uomo per motivi voluttuari (fumo di tabacco), professionali (arsenico, cromo, nichel, asbesto) o ambientali (prodotti della combustione, fumi industriali, ecc.; v. Willis, 19674). Ma, accanto a questi, si possono considerare anche motivi o cause più strettamente inerenti la patologia dell'organo. Le cicatrici nel parenchima polmonare favoriscono notevolmente l'insorgenza di cancri del polmone. E i motivi di cicatrici polmonari sono nettamente aumentati oggi rispetto al passato: per la guarigione di processi tubercolari che un tempo non si potevano curare; per le fibrosi polmonari da polveri (silicosi, asbestosi, ecc.); per le polmoniti che, interrotte nel loro naturale decorso dall'uso frequentissimo di antibiotici, vanno talora incontro a organizzazione fibrosa degli essudati negli alveoli (la cosiddetta carnificazione polmonare) invece che a risoluzione (cioè lisi e riassorbimento degli essudati alveolari).
Il cancro del polmone può oggi essere curato mediante interventi chirurgici, terapie radianti e chemioterapie, con probabilità di successo in certi casi purché sia diagnostica- to precocemente. Per questo ha acquistato importanza in clinica la classificazione morfologica dei diversi istotipi a scopo terapeutico e prognostico, poiché ognuno esibisce comportamenti e risposte peculiari. Così il carcinoma a cellule squamose (il più frequente nei fumatori) risente il beneficio maggiore dalla terapia chirurgica mentre su di esso ha scarsissimo effetto la chemioterapia. Opposto è il comportamento del carcinoma a piccole cellule anaplastiche, il più sensibile ai trattamenti chemioterapici e radianti, mentre in posizione intermedia fra i due sono il carcinoma a grandi cellule anaplastiche e l'adenocarcinoma. Tuttavia, i carcinomi anaplastici hanno nel complesso la prognosi peggiore per il loro comportamento biologico che conduce a metastasi precoci (v. Cohen e Selawry, 1975).
Il carcinoma alveolare periferico del polmone umano è collegato da rapporti per lo meno formali con una varietà di adenomatosi polmonare papillare infettiva già da tempo descritta nella pecora, indicata col nome di Jaagsiekte (v. Duran-Reynals e altri, 1958) e ritenuta di assai probabile origine virale (v. Markson e Terleki, 1964). I tumori spontanei polmonari del topo sono anch'essi di derivazione bronchiolo-alveolare e di aspetto papillare, mostrando quindi numerosi punti di contatto da un lato con l'adenomatosi polmonare della pecora, dall'altro con il carcinoma alveolare periferico del polmone umano. L'ipotesi che i tumori spontanei del polmone nel topo siano dovuti a fattori endogeni presenti nell'ospite e trasportati al polmone dal sangue circolante, è suggerita da consistenti dati sperimentali (v. Squartini e Bolis, 1970). Ne discende che anche il carcinoma alveolare periferico del polmone umano potrebbe riconoscere cause endogene e diverse da quelle dei tumori emergenti dai grossi bronchi in prossimità dell'ilo.
c) Cancro dell'intestino
Se si pone in rapporto l'incidenza neoplastica dei vari segmenti intestinali con la loro lunghezza, cioè con l'estensione della loro superficie mucosa, ne scaturisce una considerazione a prima vista paradossale: la frequenza del cancro nei vari segmenti è inversamente proporzionale alla loro lunghezza (v. Hueper, 1942). Infatti, nell'intestino tenue, lungo 7 metri, si localizza solo il 5% di tutti i cancri intestinali, mentre il 95% è localizzato nell'intestino crasso, lungo i metro e 70 centimetri, e più del 50% di questi ha sede nella sua ultima porzione, il retto, che è lungo appena 13 centimetri (v. Squartini e Caschera, 1956). Ciò fornisce la riprova più evidente in patologia umana della distribuzione non casuale o spontanea dei tumori, la quale seguirebbe altrimenti il calcolo delle probabilità.
Analizzando più da vicino la distribuzione si osserva che nel duodeno (3,6% di tutti i cancri intestinali; 26 cm di lunghezza) la maggior parte dei tumori ha due sedi particolari e precisamente i dintorni della papilla di Vater, che conduce la bile all'intestino, e la regione parapilorica, che rappresenta il punto d'ingresso del contenuto gastrico nel lume intestinale (v. Squartini e Maltzeff, 1956). Ciò suggerisce che nel chimo gastrico e nella bile siano contenute sostanze cancerogene, un fatto anzi sul quale non dovrebbero sussistere molti dubbi se si tien conto dell'elevata frequenza di neoplasie maligne riscontrabili nel tratto oro-gastrico ed epato-biliare. Attraverso la via oro-gastrica possono giungere all'intestino i cancerogeni presenti nella dieta, di cui si è già parlato. Sembra tuttavia improbabile che questi abbiano un ruolo importante nella cancerogenesi intestinale, poiché le popolazioni con alto rischio di cancro del tratto esofago-gastrico non presentano anche un alto rischio di cancro intestinale (v. Correa e Haenszel, 1978). Attraverso la via epato-biliare, verosimilmente dunque più importante, passano le sostanze nocive condotte al fegato per la neutralizzazione ed espulse nell'intestino con la bile, fra cui possono esservi anche sostanze cancerogene (v. Cleveland e altri, 1967), così come vi sono sostanze precancerogene, rappresentate per esempio dagli acidi biliari (v. Wynder, 1975). Più a valle, infatti, i batteri intestinali, metabolizzando gli acidi biliari, il colesterolo e i grassi della dieta sono, come già detto, verosimilmente in grado di produrre da queste sostanze cancerogeni intestinali endogeni (v. Reddy e Wynder, 1973; v. Reddy e altri, 1975; v. Hill, 1975 e 1977).
Tutto ciò non sarebbe comunque sufficiente a spiegare appieno la peculiare distribuzione dei tumori intestinali se non si ponesse mente alla fisiologia del tubo digerente che di quella distribuzione è in larga parte responsabile. Infatti, all'immissione del chimo gastrico e della bile nel duodeno fa seguito un'immediata, abbondante secrezione di muco operata dalle ghiandole di Brunner che diluisce e fluidifica notevolmente il contenuto del lume facilitandone lo svuotamento, il quale è molto rapido grazie anche alla vivace peristalsi, e ricoprendo le pareti con una sottile pellicola di muco a funzione protettiva e lubrificante. Queste verosimilmente le ragioni che annullano, al di là del piloro e della papilla di Vater, il potere oncogeno del contenuto duodenale, giustificando la bassa incidenza neoplastica di tale segmento (3,6%) e quella ancora più bassa del digiunoileo (1,8%), il tratto più lungo dell'intestino, dove i pochi tumori sono localizzati in genere nell'ultima porzione, cioè in una zona in cui il contenuto intestinale, terminati i fenomeni di assorbimento, è meno fluido ed è costretto a un soggiorno più lungo per la presenza della valvola ileocecale.
A partire dal cieco e per tutta la lunghezza dell'intestino crasso, fino al retto, l'incidenza neoplastica cresce progressivamente in senso cranio-caudale e la spiegazione dei fatti ci viene ancora una volta dalla fisiologia. Le modificazioni più importanti che il contenuto intestinale subisce a livello del crasso consistono nel riassorbimento dei liquidi ancora presenti nelle feci e, di conseguenza, nella loro progressiva solidificazione. A questo punto diviene efficace anche l'azione metabolica dei Batteri sugli acidi biliari e sui grassi della dieta (v. Hill, 1975 e 1977). Questi fatti comportano un aumento di concentrazione delle sostanze cancerogene che prima erano state diluite e un aumento del tempo di soggiorno delle feci nei vari segmenti. Dopo quanto ora esposto non sembra più strano che l'incidenza tumorale nell'ambito dell'intestino crasso aumenti progressivamente nonostante che la lunghezza dei segmenti nel contempo diminuisca, potendosi ciò riassumere nella proposizione che la frequenza del cancro nei vari segmenti intestinali è direttamente proporzionale al tempo di soggiorno in essi delle feci e alla concentrazione raggiunta dalle sostanze cancerogene in queste ultime (v. Hueper, 1942; v. Squartini, 1957).
La più precoce modificazione morfologicamente dimostrabile nel cancro intestinale è la dislocazione delle mitosi negli strati più superficiali dell'epitelio, dove normalmente non si ritrovano (v. Deschner e altri, 1966). Tale fenomeno suggerisce l'avvento di alterazioni nel microambiente della mucosa e determina una sovrapproduzione di cellule che finiscono per protrudere nel lume dando origine a lesioni quali i polipi adenomatosi e villosi (v. Cole e McKalen, 1963). Le informazioni fin qui raccolte circa tali lesioni depongono per una forte e consistente associazione tra polipi e cancro intestinale, di natura causale nella maggior parte dei casi. Il polipo è un prerequisito obbligato per il cancro quando la malignità comincia nel polipo. Ma vi sono casi ben documentati nei quali il cancro non inizia in uno dei polipi, pure con esso associati. In tali casi l'associazione è indiretta nel senso di due risposte indipendenti ad una stessa causa (v. sopra, cap. 7). Questa distinzione nelle vie percorribili dal cancro intestinale ha rilevanza pratica per la strategia di controllo. Infatti, la speranza di prevenire tutti, o la maggior parte, dei cancri intestinali resecando i polipi sembra non realistica. La profilassi ideale del cancro dovrebbe invece puntare alla prevenzione dei polipi rimuovendo le cause che li producono. A questo punto, infatti, la distinzione fra associazione diretta e indiretta diverrebbe irrilevante (v. Correa e Haenszel, 1978).
Le differenze anatomiche dei vari segmenti intestinali e dei tumori che in questi si sviluppano sono causa di differenze rilevanti nelle manifestazioni cliniche e nella prognosi fra i cancri del colon destro, del colon sinistro e del retto. I primi si manifestano tardivamente con emorragie, squilibri elettrolitici da eccessiva perdita per ipersecrezione tumorale, cachessia; i secondi si manifestano invece più precocemente perché stenosanti e pertanto occludenti il lume intestinale. I cancri del retto hanno la prognosi peggiore. Numerosi fattori prognostici, rilevabili dall'esame clinico, chirurgico o morfologico, sono stati di recente identificati e introdotti nella pratica (v. Gérard, 1975).
Le terapie chirurgiche hanno fatto progressi. Tuttavia le prospettive di guarigione o di sopravvivenza globali dei soggetti con cancro intestinale curato sono ancora oggi assai poco brillanti. Infatti, su 100 casi di carcinoma dell'in- testino crasso, al momento della diagnosi 5 sono inoperabili, 95 sono giudicati operabili. Di questi ultimi, al momento dell'intervento si appura che in realtà 15 sono inoperabili e solo 80 sono giudicati operabili. Tuttavia, per 10 di questi 80 nel corso della resezione si rileva che l'intervento sarà solo palliativo, perché non tutto il tumore è asportabile. Dei 70 che rimangono e vengono curati con intervento chirurgico radicale, 3 muoiono in conseguenza dell'intervento, 35 muoiono entro cinque anni per recidiva o metastasi e solo 32 su 100 dunque sopravvivono per più di cinque anni con buone probabilità di essere guariti (v. Robbins, 1974).
d) Cancro dello stomaco
Sebbene l'incidenza del carcinoma gastrico sia in diminuzione, esso rimane il tipo più frequente di cancro in Giappone, nell'Unione Sovietica, nell'America Latina e anche in alcuni paesi europei, nonché una delle più comuni cause di morte per tumori ovunque. Il carcinoma gastrico è invece eccezionale negli animali domestici e di laboratorio, praticamente assente in quelli selvatici e ciò indica chiaramente che le cause sono da ricercare nelle innaturali condizioni di vita dell'uomo, specie riguardo alle sue abitudini alimentari. I cancerogeni possono derivare dagli additivi, dai modi di cottura dei cibi, dalle sostanze inquinanti il suolo e l'atmosfera che possono venire raccolte dalle acque e di cui si è già parlato (v. sopra, cap. 5). Il cancro gastrico per motivi genetici e/o ambientali ha una distribuzione geografica molto varia. La sua frequenza è influenzata anche dallo stato sociale dei soggetti, in quanto nelle classi sociali più povere la sua incidenza è circa doppia rispetto a quella delle classi agiate (v. Willis, 19674), ciò che non è forse estraneo al fatto già rilevato della sua tendenza negli ultimi decenni a diminuire di frequenza.
Questo tipo di tumore fornisce utili esempi negativi e positivi circa il possibile significato concausale di lesioni gastriche concomitanti. L'ulcera gastrica peptica è frequente, talora è associata al cancro dello stomaco e lo precede, per cui si potrebbe supporre che l'ulcera favorisca l'insorgenza del cancro rappresentandone un precursore morfologico e una concausa importante. Ma tale ipotesi non resiste alla critica ove si consideri che l'ulcera peptica duodenale è ancora più frequente di quella gastrica, mentre il cancro duodenale è raro. Pertanto, fatta eccezione per situazioni particolari, all'ulcera dello stomaco si riconosce oggi solo il significato di un fattore di localizzazione del cancro realizzato da altre cause (v. Willis, 19674). Diverso è il caso per la gastrite cronica atrofica associata ad anemia perniciosa, la quale è considerata un precursore morfologico e un fattore causale o concausale importante, capace di evolvere nel 10% dei casi in cancro dello stomaco. In tale malattia è frequente anche il riscontro di carcinomi in situ. Circa il meccanismo patogenetico che la legherebbe al cancro si attribuisce importanza alle modificazioni cellulari instaurantisi a seguito della carenza di vitamina B12, non assorbita per l'atrofia della mucosa gastrica, carenza che direttamente influisce sulla sintesi del DNA e sulla replicazione cellulare (v. Robbins, 1974).
Il cancro gastrico non trattato o incurabile porta a morte nel giro di un anno dall'inizio dei sintomi. Solo il 10% dei pazienti supera l'anno e appena il 2% i 5 anni. Poiché l'intervento chirurgico si rivela difficilmente radicale, anche il carcinoma gastrico curabile diviene presto un problema di terapia medica con scarse prospettive per ora di efficaci risultati. Gli agenti chemioterapici possono produrre regressioni in una certa percentuale di casi prolungando la sopravvivenza. Le risposte favorevoli sono più frequenti nei maschi, nell'età di mezzo e negli istotipi maggiormente differenziati, che sono i carcinomi a struttura ghiandolare o adenocarcinomi (v. Moertel, 1975).
e) Leucemie e linfomi
Sono questi i tumori del sistema emo-linfo-reticolare che comprende i linfonodi, il timo, la milza e i tessuti linfatici sparsi nell'organismo (come per es. le tonsille), produttori di linfociti e plasmacellule, e il midollo osseo che produce le cellule staminali progenitrici di tutto il sistema, i globuli rossi o eritrociti (serie eritroide) e i globuli bianchi o leucociti (serie mieloide). Si parla di leucemia quando le cellule neoplastiche circolano nel sangue, come fanno le corrispondenti cellule normali, un fenomeno oggi considerato alla stregua di una metastasi circolante (v. Willis, 19674), mentre col termine di linfomi in senso lato si intendono comunemente i tumori solidi del sistema. Per molti anni tali neoplasie sono state considerate, osservate e classificate dal solo punto di vista morfologico, senza cioè alcun riferimento alla funzione dei rispettivi stipiti cellulari. Oggi, dopo i rilevanti progressi dell'immunologia negli ultimi vent'anni, si cominciano giustamente a riconsiderare molte di queste neoplasie come tumori del sistema immunitario. (v. sangue: Leucemie e Organi emopoietici ; v. sistema reticolo endoteliale).
Limitando i ricordi funzionali a un solo tipo cellulare, i linfociti, va ricordato che oggi, in base alle osservazioni sui difetti immunologici congeniti nell'uomo e agli esperimenti di ablazione negli animali di parti dei sistemi immunitari murini (timo) e aviari (borsa di Fabrizio), si ritiene che esistano due sistemi linfocitari funzionanti: il sistema a cellule T (timo dipendente) responsabile della immunità cellulare e il sistema a cellule B (borsa equivalente) responsabile della immunità umorale. Vi sono tuttavia anche cellule dette U (undefined) o N (null), perché non appartenenti né all'uno né all'altro dei sistemi nominati. Tutte queste cellule deriverebbero da un progenitore comune, la cellula germinativa o staminale prodotta nel midollo e circolante col sangue. A seconda che questa cellula si diriga nel timo o nelle strutture linfatiche del tubo digerente (ritenute l'equivalente nell'uomo della borsa di Fabrizio degli uccelli), la sua discendenza sarebbe condizionata o commissionata per la funzione T o B. Dagli organi linfatici centrali (timo e stazioni linfatiche del digerente) le cellule così commissionate per la funzione si dirigerebbero attraverso il circolo sanguigno a popolare gli organi linfatici periferici o effettori (linfonodi, milza, ecc.; v. Smith, 19777; v. immunologia e immunopatologia).
Le cellule B e T sono distribuite in maniera caratteristica negli organi linfatici effettori periferici: le cellule T occupano le aree paracorticali dei linfonodi e le regioni perivascolari della milza, le cellule B invece sono concentrate nei centri germinativi dei linfonodi e della milza (i cosiddetti follicoli linfatici a struttura rotondeggiante concentrica), nelle mucose del tratto gastroenterico e disseminate nel midollo. Le plasmacellule, che sono cellule mature e funzionanti del sistema B, appaiono tipicamente localizzate nell'area midollare dei linfonodi, nelle aree perivascolan della milza e del midollo osseo. Le cellule T rappresentano normalmente il 70% dei linfociti del sangue circolante mentre il rimanente 30% è costituito da cellule B e N. I linfociti B e T, sotto l'influenza di mitogeni vegetali e antigeni ai quali siano stati precedentemente esposti, si trasformano in vitro da piccoli linfociti in cellule voluminose capaci di moltiplicarsi e metabolicamente attive, dette immunoblasti, ciò che ha meritato al fenomeno il nome di blastizzazione.
La fitoemoagglutinina è un mitogeno specifico per i linfociti T, come la fitolacca lo è per i linfociti B, i quali blastizzano comunque sotto l'azione di svariati antigeni. I centri germinativi dei linfonodi e della milza sono le sedi della blastizzazione dei linfociti B in vivo dopo stimolazione antigenica. I linfociti B possono essere anche identificati selettivamente in laboratorio per la presenza di immunoglobuline di superficie e di recettori superficiali per il complemento. I linfociti T sono identificabili per la formazione di rosette con eritrociti di montone dietro stimolazione da parte di antigeni timo-dipendenti. Anche la morfologia secondo alcuni servirebbe a differenziare i linfociti T dai B in quanto i primi avrebbero una superficie relativamente liscia, i secondi una superficie filamentosa o pelosa per numerosi microvilli che al microscopio elettronico tridimensionale appaiono come ciuffi di peli (v. Smith, 19777).
Questi progressi dell'immunologia stanno rivoluzionando gli schemi patologici tradizionali di classificazione dei linfomi (v. Collins e Lukes, 1971; v. Lukes e Collins, A functional approach..., 1974). Una classificazione immunologica dei linfomi è già stata proposta e sta entrando sperimentalmente nell'uso, almeno come ipotesi di lavoro (v. Lukes e Collins, Immunologic..., 1974). La sindrome di Sézary, la micosi fungoide, che sono tipi particolari di linfomi cutanei, la leucemia linfatica acuta dei bambini e, forse, il linfogranuloma maligno o malattia di Hodgkin sono oggi considerate neoplasie a linfociti T. La leucemia linfatica cronica, il linfoma linfocitico differenziato, i linfomi del centro germinativo, di aspetto spesso nodulare come le strutture da cui traggono origine, i linfomi caratterizzati da abnorme produzione di immunoglobuline e con essi il plasmocitoma, il mieloma, la macroglobulinemia di Waldeström e il linfoma di Burkitt o africano sono invece ritenute oggi neoplasie a linfociti B. Accanto a tali due gruppi principali si considerano poi ancora i sarcomi immunoblastici (a blasti T o B) e i sarcomi istiocitari, cioè composti da cellule reticolari macrofagiche indifferenziate corrispondenti alla cellula midollare originaria dei sistemi (v. Lukes e Collins, 1975; v. Smith, 19777).
Sulla base dei nuovi criteri, numerosi fattori prognostici e di valutazione della risposta alle terapie sono stati vagliati e saggiati (v. Rozencweig e altri, 1975; v. Bergsagel, 1975; v. Mathé e altri, 1975; v. Silver, 1975; v. Eckhardt, 1975). Per il futuro si attende, da un'impostazione concettuale più razionale, un approccio migliore ai problemi della prognosi e della terapia.
15. Conclusioni
‟Il cancro è sempre stato un ‛problema'. In principio, per quanto personalmente possa ricordare, era un ‛problema intellettuale' che molti pensavano fosse oltre i poteri della mente umana di risolvere. Oggi è divenuto un ‛problema biologico', che, come a volte si dice, i biologi potrebbero risolvere nei ritagli di tempo, se ne avessero. Il pendolo è probabilmente sfuggito troppo oltre. Il cancro è ancora una malattia che uccide la gente e i limitati contributi delle ricerche di laboratorio alla alleviazione delle sofferenze umane bandiscono ogni facile ottimismo" (v. Foulds, 1969).
Di fronte alla impressionante mole di ricerche e di bibliografia sul cancro accumulatesi durante questo secolo, sta ancora la scarsezza dei risultati utili in pratica. Di fronte alla fioritura di ipotesi per interpretare il problema biologico del cancro, sta l'attuale povertà di valide e accettate conclusioni. Ciò forse dipende anche dal fatto che questa malattia clinica e questo affascinante problema di laboratorio trascendono i limiti e il linguaggio della patologia. Le basi patologiche rimangono ancora oggi fondamentali per una impostazione razionale del problema, ma l'accento va forse posto sulla patologia neoplastica intesa come processo epigenetico (v. Foulds, 1969), o come accidente evoluzionistico (v. Burnet, Immunological surveillance, 1970) e non meramente come sviluppo di tumefazioni palpabili.
È difficile immaginare che soluzioni pratiche di portata rivoluzionaria possano essere raggiunte fino a quando il problema dell'accrescimento neoplastico non avrà trovato un'interpretazione biologica soddisfacente nel suo insieme. Ed è perciò da ritenersi un progresso la formulazione di concetti nuovi che trascendono l'aspetto anatomo-clinico attingendo al vasto patrimonio di conoscenze della moderna biologia. Da questo orientamento di pensiero discende l'intuizione che il cancro è ‟un processo inevitabile quanto il progresso evoluzionistico e della stessa natura generale" (v. Burnet, 1967), come l'affermazione che ‟nessuna teoria sul cancro è accettabile se non considera le neoplasie come una delle possibili conseguenze della organizzazione biologica" (v. Foulds, 1969).
Nel corso dell'evoluzione si è reso necessario per gli organismi più complessi di preservare l'uniformità del tipo cellulare imparando a riconoscere dalle proprie le strutture estranee e a eliminarle attraverso il fenomeno immunologico del rigetto (v. Thomas, 1959). Per tali scopi si è sviluppato negli organismi superiori il sistema dell'immunità cellulare imperniato sul timo, sugli organi linfatici periferici, sui linfociti circolanti, dotato della capacità di rigettare i tessuti eterogenei. Questo sistema sembra rivelarsi oggi come un meccanismo primario nella difesa naturale contro le neoplasie attraverso la ‛sorveglianza immunologica'. Ma è probabile che la stessa modificazione flessibile del controllo genetico, necessaria per lo sviluppo del sistema immunitario e per la comparsa in ogni specie degli antigeni specifici di istocompatibilità, sia stata anche responsabile della comparsa del cancro come malattia nei Vertebrati (v. Burnet, Immunological surveillance..., 1970).
L'insieme delle informazioni derivate dallo studio delle metastasi dei tumori nell'uomo e del trapianto dei tumori negli animali, indica che il comportamento neoplastico dipende da una modificazione cellulare stabile, probabilmente irreversibile, replicabile e verosimilmente presente in tutte le cellule neoplastiche. La prova decisiva della donazione (trapianto di una sola cellula), necessaria per sostanziare questa opinione, è tecnicamente difficile e di rado ha successo. Ma la donazione di un teratocarcinoma trapiantabile fornisce evidenze convincenti di un meccanismo stabile e replicabile entro le singole cellule, responsabile di tutti i dettagli della struttura neoplastica e del comportamento, il quale dunque possiede e trasporta con sé la ‛memoria morfogenetica' o il ‛programma di sviluppo' di un archetipo tumorale estremamente complesso, analogamente a quanto avviene per la ‛differenziazione biologica' lungo tutta la scala degli esseri viventi (v. Foulds, 1969). Inoltre, dalla sperimentazione con virus dei tumori mammari murini, è emersa la dimostrazione che informazioni genetiche complete per la struttura morfologica e il comportamento biologico dei tumori sono possedute e trasmesse stabilmente dagli acidi nucleici virali (v. Squartini e altri, 1963).
Lo studio anatomo-clinico delle neoplasie e la ricerca sperimentale hanno prodotto un'inverosimile quantità di conoscenze empiriche o di fatti senza fornire nel contempo un adeguato linguaggio per la loro comprensione, o concetti efficaci per la loro sintesi (v. Foulds, 1969). Tali concetti, e il linguaggio biologico necessario alla decodificazione del problema, cominciano oggi ad affiorare, offrendosi per l'approfondimento e la meditazione. Al termine di una rassegna, necessariamente sommaria, dei risultati accumulati in quasi mezzo secolo conforta percepire, nella pensosa rielaborazione concettuale dei fatti, quasi il presagio di una soluzione.
bibliografia
AA. VV., Early lesions and the development of epithelial cancer, in ‟Cancer research", 1976, XXXVI, pp. 2475-2706.
AA. VV., Spontaneous regression of cancer, in ‟National Cancer Institute Monographs", 1976, XXXXIV, pp. 1-150.
AA. VV., Preneoplasia, in ‟The American journal of pathology", 1977, LXXXIX, pp. 401-482.
Alexander, P., Immunità e tumori, in Biologia dei tumori (a cura di E. J. Ambrose e F. J. C. Roe), Napoli 1978, pp. 282-314.
Alpert, M. E., Hutt, M. S. R., Wogan, G. N., Davidson, C. S., The association between aflatoxin content of food and hepatoma frequency in Uganda, in ‟Cancer", 1971, XXVIII, pp. 253-260.
Ambrose, E. J., Invasiveness and surface properties of cancer cells, in ‟Nature", 1958, CLXXXII, pp. 1419-1421.
Andersen, D. H., Tumors of infancy and childhood. I. A survey of those seen in the Pathology Laboratory of the Babies Hospital during the years 1935-1950, in ‟Cancer", 1951, IV, pp. 890-906.
Anderson, D. E., Some characteristics of familial breast cancer, in ‟Cancer", 1971, XXVIII, pp. 1500-1504.
Anderson, D. E., A genetic study of human breast cancer, in ‟Journal of the National Cancer Institute", 1972, XLVIII, pp. 1029-1034.
Anglesio, E., Le sindromi paraneoplastiche, in Trattato di oncologia clinica (a cura di P. Bucalossi e U. Veronesi), Milano 1973, pp. 232-243.
Antony, P. P., Bontwell, R. K., Farber, E., Friedell, G. H., Gullino, P. M., Lipkin, M., Netttesheim, P., Price, J., Sporn, M. B., Early lesions and the development of epithelial cancer (Symposium).
Summary statement, in ‟Cancer research", 1976, XXVI, p. 2706.
Ashley, D. J. B., Evans' histological appearances of tumours, Edinburgh-London-New York 19783.
Atkins, H. J. B., Breast cancer symposium. Discussion, in ‟British journal of surgery", 1969, LVI, pp. 782-796.
Bagshawe, K. D., Choriocarcinoma, London 1969.
Bailey, E., Dungal, N., Polycyclic hydrocarbons in Icelandic smoked food, in ‟British journal of cancer", 1958, XII, pp. 348-350.
Barnes, A. W., Vinyl chloride and the production of PVC, in ‟Proceedings of the Royal Society of Medicine", 1976, LXIX, pp. 277-281.
Baserga, R., Putong, P. B., Tyler, S., Wartman, W. B., The dose-response relationship between the number of embolic tumor cells and the incidence of blood-borne metastates, in ‟British journal of cancer", 1960, XIV, pp. 173-185.
Baserga, R., Saffiotti, U., Experimental studies on histogenesis of blood-borne metastates, in ‟Archives of pathology", 1955, LIX, pp. 26-34.
Bentvelzen, P., Daams, J. H., Hageman, P., Calafat, J., Genetic transmission of viruses that incite mammary tumor in mice, in ‟Proceedings of the National Academy of Sciences", 1970, LXVII, pp. 377-384.
Berenbaum, M. C., Sheard, C. E., Testing anti-cancer drugs, in ‟Lancet", 1972, I, pp. 1116-1117.
Berenblum, I., The biological mechanism of carcinogenesis, in Scripta varia. Semaine d'étude sur le problème biologique du cancer, Città del Vaticano 1949, pp. 1-13.
Berenblum, I., Carcinogenesis and tumor pathogenesis, in ‟Advances in cancer research", 1954, II, pp. 129-175.
Bergsagel, D. E., Plasma cell myeloma. Prognostic factors and criteria of response to therapy, in Cancer therapy. Prognostic factors and criteria of response (a cura di M. J. Staquet), New York 1975, pp. 73-87.
Bergsma, D., Good, R. A. (a cura di), Immunological deficiency diseases of man, New York 1968.
Berman, C., The precancerous lesions associated with primary carcinoma of the liver, in The morphological precursors of cancer (a cura di L. Severi), Perugia 1962, pp. 229-249.
Bernard, J., Paul, R., Boiron, M., Jacquillat, C., Maral, R., Rubidomycin, Berlin-Heidelberg-New York 1969.
Bielschowsky, F., Horning, E. S., Aspects of endocrine carcinogenesis, in ‟British medical bulletin", 1958, XIV, pp. 106-115.
Bittner, J. J., Some possible effects of nursing on the mammary gland tumor incidence in mice, in ‟Science", 1936, LXXXIV, pp. 162-163.
Black, M. M., Structural, antigenic and biological characteristics of precancerous mastopathy, in ‟Cancer research", 1976, XXVI, pp. 2596-2604.
Black, M. M., Moore, D. H., Shore, B., Zachrau, R. E., Leis, H. P., Effect of murine milk samples and human breast tissues on human leucocyte migration indices, in ‟Cancer research", 1974, XXXIV, pp. 1054-1060.
Black, M. M., Opler, S. R., Speer, F. D., Survival in breast cancer cases in relation to the structure of primary tumor and regional lymphnodes, in ‟Surgery, ginecology and obstetrics", 1955, C, pp. 543-551.
Black, M. M., Zachrau, R. E., Shore, B., Leis, H. P., Biological considerations of tumor-specific and virus-associated antigens of human breast cancer, in ‟Cancer research", 1976, XXXVI, pp. 769-774.
Blair, P. B., DeOme, K. B., Mammary tumour development in transplanted hyperplastic alveolar nodules of the mouse, in ‟Proceedings of the Experimental Society of Biology and Medicine", 1962, CVIII, pp. 289-291.
Bondy, P. K., Powles, T. J., Leese, C. L., Hydroxyproline excretion as an indicator of progress in treating breast cancer, in Abstracts of the 9th meeting on mammary cancer in experimental animals and man (a cura di F. Squartini), Pisa 1974, p. 29.
Borsa, J., Whitmore, G. F., Cell killing studies on the mode of action of methotrexate on L-cells in vitro, in ‟Cancer research", 1969, XXIX, pp. 737-744.
Bove, K. E., Hoffler, H., McAdams, A. J., Nodular renal blastema. Definition and possible significance, in ‟Cancer", 1969, XXIV, pp. 323-332.
Boyes, D. A., Fidler, H. K., Lock, D. R., Significance of in situ carcinoma of the uterine cervix, in ‟British medical journal", 1962, I, pp. 203-205.
Brennan, M. J., Endocrinology in cancer of the breast. Status and prospects, in ‟American journal of clinical pathology", 1975, LXIV, pp. 797-809.
Bruton, O. C., Agammaglobulinemia, in ‟Pediatrics", 1952, IX, pp. 722-728.
Bucciante, L., Sui fondamenti angioarchitetturali e strutturali della regolazione del circolo sanguigno, in ‟Archivio De Vecchi", 1960, XXXII, pp. 117-239.
Bukhari, A., Connors, T. A., Gilsenan, A. M., Ross, W. C. J., Tisdale, M. J., Warwick, G. P., Wilman, D. E. V., Cytotoxic agents designed to be selective for liver cancer, in ‟Journal of the National Cancer Institute", 1973, L, pp. 243-247.
Bulbrook, R. D., The role of endocrine factors in the aetiology of mammary cancer, in Corso superiore sul trattamento dei tumori della mammella (a cura di P. Bucalossi e A. Ratti), Milano 1971, pp. 41-44.
Bulbrook, R. D., VIII. Endocrinology, in Breast cancer research, UICC technical report series (a cura di M. J. Brennan), vol. XXVII, Genève 1977, pp. 89-111.
Burghardt, E., Early histological diagnosis of cervical cancer, Stuttgart 1973.
Burkitt, D., The African lymphoma. Epidemiological and therapeutic aspects, in Proceedings of the international conference on leukemia-lymphoma (a cura di C. Zarafonetis), Philadelphia 1968, pp. 321-330.
Burkitt, D. P., Wright, D. H., Burkitt's lymphoma, Edinburgh-London 1970.
Burnet, F. M., Cancer: a biological approach, in ‟British medical journal", 1957, I, pp. 779-786, 841-847.
Burnet, F. M., The evolution of bodily defence, in ‟Medical journal of Australia", 1963, II, pp. 817-821.
Burnet, F. M., Immunological factors in the process of carcinogenesis, in ‟British medical bulletin", 1964, XX, pp. 154-158.
Burnet, F. M., Immunological aspects of malignant disease, in ‟Lancet", 1967, I, pp. 1171-1174.
Burnet, F. M., Evaluation of the evidence for immune surveillance: impressions and comments, in Immune surveillance (a cura di R. T. Smith e M. Landy), New York 1970, pp. 512-540.
Burnet, F. M., Immunological surveillance, Oxford 1970.
Burnet, F. M., Immunological surveillance: an evolutionary approach, in Immunity and tolerance in oncogenesis (a cura di L. Severi), Perugia 1970, pp. XLV-LXI.
Burnet, F. M., The concept of immunological surveillance, in ‟Progress of experimental tumour research", 1970, XIII, pp. 1-37.
Cameron, G. R., Spector, W. G., The chemistry of the injured cell, Springfield 1961.
Carter, S. K., Schabel, F. M., Broder, L. E., Johnston, T. P., 1,3-bis (2-chloroethyl)-1-nitrosourea (BCNU) and other nitrosoureas in cancer treatment: a review, in ‟Advances in cancer research", 1972, XVI, pp. 273-332.
Case, R. A. M., Hosker, M. E., McDonald, D. B., Pearson, J. T., Tumours of the unirary bladder in workmen engaged in the manufacture and use of certain dyestuff intermediates in the British chemical industry. Part I. The role of aniline, benzidine, alpha-naphthylamine and betanaphthylamine, in ‟British journal of industrial medicine", 1954, XI, pp. 75-104.
Chabner, B. A., Chello, P. L., Bertino, J. R., Antitumor activity of a folate-cleaving enzyme, carboxypeptidase G1, in ‟Cancer research", 1972, XXX, pp. 2114-2119.
Charney, J., Morre, D. H., Neutralization of murine mammary tumor virus by sera of women with breast cancer, in ‟Nature", 1971, CCXXIX, pp. 627-628.
Chu, T. M., Nemoto, T., Evaluation of carcinoembryonic antigen in human mammary carcinoma, in ‟Journal of the national Cancer Institute", 1973, LI, pp. 1119-1122.
Cleaver, J. E., Defective repair replication of DNA in xeroderma pigmentosum, in ‟Nature", 1968, CCXVIII, pp. 652-656.
Cleveland, J. C., Litvak, S. F., Cole, J. W., Identification of the route of action of the carcinogen 3 : 2′, dimethyl-4-aminobiphenyl in the induction of intestinal neoplasia, in ‟Cancer research", 1967, XXVII, pp. 708-714.
Cobb, L. M., Metastatic spread of human tumour implanted into thymectomized, athymocyte serum treated hamsters, in ‟British journal of cancer", 1972, XXVI, pp. 183-189.
Cohen, M. H., Selawry, O. S., Bronchogenic carcinoma. Prognostic factors and criteria of response, in Cancer therapy. Prognostic factors and criteria of response (a cura di M. J. Staquet), New York 1975, pp. 185-197.
Cole, J. W., McKalen, A., Studies on the morphogenesis of adenomatous polyps in the human colon, in ‟Cancer", 1963, XVI, pp. 998-1002.
Cole, W. H., Opening adress: spontaneous regression of cancer and the importance of finding its cause, in ‟National Cancer Institute Monographs", 1976, XXXIV, pp. 5-9.
Collins, R. D., Lukes, R. J., Studies on possible derivation of some malignant lymphomas from follicular center cells (Abstract), in ‟American journal of pathology", 1971, LXII, p. 63 a.
Coman, D. R., Mechanisms responsible for the origin and distribution of blood-borne tumor metastases. A review, in ‟Cancer research", 1953, XIII, pp. 397-404.
Consolandi, G., Per una nuova classificazione dei tipi di cancrometastasi. Critica della classificazione di H. E. Walther. Le emometastasi tipo ‛cuore', in ‟Lavori dell'Istituto di Anatomia e Istologia Patologica, Università degli Studi, Perugia", 1949, VIII, pp. 169-253.
Consolandi, G., Le cancro-metastasi. I. Frequenza e distribuzione delle metastasi cancerigne in rapporto alla nuova classificazione dei tipi di cancrometastatizzazione. (Studio statistico sul materiale autoptico di Perugia e rassegna della letteratura moderna), in ‟Lavori dell'Istituto di Anatomia e Istologia Patologica, Università degli Studi, Perugia", 1950, IX, pp. 181-261.
Consolandi, G., Le cancro-metastasi. II. Verso una sistematica definitiva dei tipi di metastatizzazione cancerigna, in ‟Lavori dell'Istituto di Anatomia e Istologia Patologica, Università degli Studi, Perugia", 1951, X, pp. 41-71.
Consolandi, G., Concetti attuali sulla diffusione metastatica dei tumori, in ‟Lavori dell'Istituto di Anatomia e Istologia Patologica, Università degli Studi, Perugia", 1960, XX, pp. 213-236.
Correa, P., Haenszel, W., The epidemiology of large bowel cancer, in ‟Advances in cancer research", 1978, XXVI, pp. 1-141.
Cowie, A. T., Folley, S. J., Endocrine aspects of mammary growth and function, particularly in relation to pituitary hormones, in Endocrine aspects of breast cancer (a cura di A. R. Currie e C. F. W. Illingworth), Edinburgh-London 1958, pp. 266-275.
Currie, A. R., Illingworth, C. F. W. (a cura di), Endocrine aspects of breast cancer, Edinburgh-London 1958.
Dao, T. L., IX. Carcinogenesis, in Breast cancer research, UICC technical report series (a cura di M. J. Brennan), vol. XXVIII, Genève 1977, pp. 112-125.
David, J., Burkitt, D., Burkitt's lymphomas. Remission following seemingly non-specific therapy, in ‟British medical journal", 1968, IV, pp. 288-291.
Decker, H. R., Alveolar cell carcinoma of the lung (pulmonary adenomatosis): a study of 155 cases, in ‟Journal of thoracic surgery", 1955, XXX, pp. 230-258.
DeDombal, F., Ulcerative colitis: definition, historical background, etiology, diagnosis, natural history and local complications, in ‟Post-graduate medical journal", 1968, VL, pp. 648-659.
Dent, P. B., Peterson, R. D. A., Good, R. A., The relationship between immunologic function and oncogenesis, in Immunologic deficiency disease of man (a cura di D. Bergsma e R. A. Good), New York 1968, pp. 443-450.
DeOme, K. B., The role of the mammary tumor virus in mouse mammary noduligenesis and tumorigenesis, in Viruses, nucleic acids and cancer, Baltimore 1963, pp. 498-507.
DeOme, K. B., Bern, H. A., Berg, W. E., Pissott, L. E., Radiophosphorus uptake by normal, hyperplastic and tumorous mammary tissues of mice, in ‟Proceedings of the Society of Experimental Biology and Medicine", 1956, XCII, pp. 55-58.
DeOme, K. B., Blair, P. B., Faulkin, L. J., Some characteristics of the preneoplastic alveolar nodules in the C3H/Crgl mice, in ‟Acta Unionis internationalis contra cancrum", 1961, XVII, pp. 973-982.
DeOme, K. B., Faulkin, L. J., Bern, H. A., Blair, P. B., Development of mammary tumors from hyperplastic alveolar nodules transplanted into gland-free mammary fat pads of female C3H mice, in ‟Cancer reseearch", 1959, XIX, pp. 515-520.
DeOme, K. B., Nandi, S., Bern, H. A., Blair, P. B., Pitelka, D. R., The preneoplastic hyperplastic alveolar nodule as the morphologic precursor of mammary cancer in mice, in The morphological precursors of cancer (a cura di L. Severi), Perugia 1962, pp. 349-368.
Deschner, E. E., Lipkin, M., Solomon, C., Study of human rectal epithelial cells in vitro. II. H3-thymidine incorporation into polips and adjacent mucosa, in ‟Journal of the National Cancer Institute", 1966, XXVI, pp. 849-857.
DeSombre, E. R., Johnson, E. S., White, W. F., Progression of rat mammary tumors effected by a gonadoliberin analog, in ‟Cancer research", 1976, XXXVI, pp. 3830-3833.
Devroede, G. J., Taylor, W. F., Saner, W. G., Jackman, R. J., Sticklen, G. B., Cancer risk and life expectancy of children with ulcerative colitis, in ‟New England journal of medicine", 1971, CCLXXXV, pp. 17-21.
Dhar, P., Moore, T., Zamcheck, N., Kupchick, H. Z., Carcinoembryonic antigen (CEA) in colonic cancer, in ‟Journal of the American Medical Association", 1972, CCXXI, pp. 31-35.
Doll, R., Interpretation of epidemiologic data, in ‟Cancer research", 1963, XXIII, pp. 1613-1623.
Doll, R., Introduction, in Origins of human cancer (a cura di H. H. Hiatt, J. D. Watson e J. A. Winsten), Cold Spring Harbor 1977, pp. 1-12.
Doll, R., Muir, C. S., Waterhouse, J. A. H., Cancer incidence in five continents, vol. II, Genève 1970.
Doll, R., Vassey, M. P., Beasley, P. W. R., Buckley, A. P., Fear, E. C., Fisher, P. E. W., Gammon, E. J., Gunn, W., Hughes, G. O., Lee, K., Norman-Smith, B., The mortality of gas workers: final report of a prospective study, in ‟British journal of industrial medicine", 1972, XXIX, pp. 394-406.
Drasar, B. S., Irving, D., Environmental factors and cancer of the colon and the breast, in ‟British journal of cancer", 1973, XXVII, pp. 167-172.
Dubreuilh, W., Des hypercheratoses circonscrites, in Official transactions of the third international congress of dermatology held in London, August 1896 (a cura di J. J. Pringle), London 1898, pp. 125-176.
Dunn, J. E., Preliminary findings of the Meenphis-Shelby County uterine cancer study and their interpretation, in ‟American journal of public health", 1958, XXXXVIII, pp. 861-868.
Duran-Reynals, F., Jungher, E., Cuba-Caparo, A., Rafferty, K. A., Helmboldt, C. F., The pulmonary adenomatosis complex in sheep, in ‟Annals of the New York Academy of Sciences", 1958, LXX, pp. 726-742.
Eckhardt, S., Lymphosarcoma and reticulum cell sarcoma. Prognostic factors and criteria of response, in Cancer therapy: prognostic factors and criteria of response (a cura di M. J. Staquet), New York 1975, pp. 131-138.
Engell, H. C., Cancer cells in the circulating blood, in ‟Acta chirurgica scandinavica" (Suppl.), 1955, CCI, pp. 1-70.
Engell, H. C., Cancer cells in the blood. A five to nine year follow up study, in ‟Annals of surgery", 1959, CIL, pp. 457-461.
Epstein, M. A., Epstein-Barr virus: is it time to develop a vaccine program?, in ‟Journal of the National Cancer Institute", 1976, LVI, pp. 697-700.
Epstein, M. A., Achong, B. C., Barr, J. M., Virus particles in cultured lymphoblasts from Burkitt's lymphoma, in ‟Lancet", 1964, I, pp. 702-703.
Evans, A. E., Gerson, J., Schnaufer, L., Spontaneous regression of neuroblastoma, in ‟National Cancer Institute Monographs", 1976, XXXXIV, pp. 49-54.
Everson, T. C., Spontaneous regression of cancer, in ‟Annals of the New York Academy of Sciences", 1964, XCIV, pp. 721-735.
Everson, T. C., Cole, W. H., Spontaneous regression of cancer, Philadelphia-London 1966.
Farber, E., Early lesions and the development of epithelial cancer (Symposium). Introductory remarks, in ‟Cancer research", 1976, XXXVI, p. 2481.
Feinleib, M., Breast cancer and artificial menopause: a cohort study, in ‟Journal of the National Cancer Institute", 1968, XXXXI, pp. 315-329.
Fisher, E. R., Gregorio, R. M., Fisher, B., The pathology of invasive breast cancer. A syllabus derived from findings of the national surgical adjuvant breast project (Protocol n. 4), in ‟Cancer", 1975, XXXVI, pp. 1-85.
Fletcher, W. S., Stewart, J. W., Tumour cells in the blood with special reference to pre- and post-hepatic blood, in ‟British journal of cancer", 1959, XIII, pp. 33-37.
Folkman, J., Cotran, R., Relation of vascular proliferation to tumor growth, in ‟International review of experimental pathology", 1976, XVI, pp. 207-248.
Foulds, L., Mammary tumours in hybrid mice. Growth and progression of spontaneous tumours, in ‟British journal of cancer", 1949, III, pp. 345-375.
Foulds, L., Experimental study of the course and regulation of tumour growth, in ‟Annals of the Royal College of Surgeons", 1951, IX, pp. 93-101.
Foulds, L., The experimental study of tumour progression. A review, in ‟Cancer research", 1954, XIV, pp. 327-339.
Foulds, L., The histologic analysis of mammary tumors of mice. I. Scope of investigation and general principles of analysis, in ‟Journal of the National Cancer Institute", 1956, XVII, pp. 701-711.
Foulds, L., The histologic analysis of mammary tumors of mice. II. The histology of responsiveness and progression. The origins of tumors, in ‟Journal of the National Cancer Institute", 1956, XVII, pp. 713-753.
Foulds, L., Neoplastic development, vol. I, London-New York-San Francisco, 1969; vol. II, London-New York-San Francisco 1975.
Fox, F., Davidson, J., Thomas, L. B., Maturation of sympathicoblastoma into ganglioneuroma. Report of 2 patients with 20 and 46 year survivals respectively, in ‟Cancer", 1959, XII, pp. 108-116.
Franks, L. A., Latency and progression in tumours. The natural history of prostatic cancer, in ‟Lancet", 1956, II, pp. 1037-1039.
Fratkin, L. B., Epidemiology of breast cancer. British Columbia. Canada and the world (Abstract), Vancouver 1976, pp. 1-14.
Fujiwara, J., The motility of gastric cancer cells, in ‟Gann" (Suppl.), 1959, L, pp. 188-189.
Gabler, G., Peckholz, I., Die Metastasierung des bronchial Karzinoms. (Morphologische Untersuchungen zur Frage der Organselektivität der hämatogenen Absiedlungen), in ‟Beiträge zur pathologische Anatomie", 1960, CXXII, pp. 452-494.
Gasic, G., Loebel, F., Badinez, O., Cementing substances in metastasizing and non metastasizing transplantable tumours in mice, in ‟Nature", 1960, CLXXXV, pp. 864-865.
Gérard, A., Carcinoma of the colon and rectum. Prognostic factors and criteria of response, in Cancer therapy. Prognostic factors and criteria of response (a cura di M. J. Staquet), New York 1975, pp. 199-227.
Ghose, T., Norvell, S. T., Guclu, A., Cameron, D., Bodurtha, A., Macdonald, A. S., Immunochemotherapy of cancer with chlorambucilcarrying antibody, in ‟British medical journal", 1972, III, pp. 495-499.
Gimbrone, M. A., Gullino, P. M., Angiogenic capacity of preneoplastic lesions of the murine mammary gland as a marker of neoplastic transformation, in ‟Cancer research", 1976, XXXVI, pp. 2611-2620.
Gold, T., Freedman, S. O., Demonstration of tumor-specific antigens in human colonic carcinomata by immunological tolerance and absorption techniques, in ‟Journal of experimental medicine", 1965, CXXI, pp. 439-462.
Gold, T., Freedman, S. O., Specific carcinoembryonic antigens of the human digestive system, in ‟Journal of experimental medicine", 1965, CXXII, pp. 467-481.
Goldin, A., Venditti, J. M., Humphreys, S. R., Mantel, N., Modification of treatment schedules in the management of advanced mouse leukemia with amethopterin, in ‟Journal of the National Cancer Institute", 1956, XVII, pp. 203-212.
Greenberg, E., Divertie, M. B., Woolner, L. B., A review of unusual systemic manifestations associated with carcinoma, in ‟American journal of medicine", 1964, XXXVI, pp. 106-120.
Griem, M. L., Jensen, E. V., Ultmann, J. E., Wissler, R. W., Breast cancer: a challenging problem, London-Berlin 1973.
Grindey, G. B., Nichol, C. A., Interaction of drugs inhibiting different steps in the synthesis of DNA, in ‟Cancer research", 1972, XXXII, pp. 527, 531.
Gross, L., ‛Spontaneous' leukemia developing in C3H mice following inoculation in infancy with AK leukemia extracts or AK embryos, in ‟Proceedings of the Society of Experimental Biology and Medicine", 1951, LXXVI, pp. 27-32.
Gross, L., Viral etiology of mouse leukemia, in ‟Advances in cancer research", 1961, VI, pp. 149-180.
Gross, L., Oncogenic viruses, Oxford-Braunschweig 19702.
Grundmann, E., Oettgen, H. F., Experimental and clinical effects of asparaginase, Berlin-Heidelberg-New York 1970.
Gullino, P. M., Considerations on the preneoplastic lesions of the mammary gland, in ‟The American journal of pathology", 1977, LXXXIX, pp. 413-430.
Haagensen, C. D., Predisposition to carcinoma of the breast, in Corso superiore sul trattamento dei tumori della mammella (a cura di P. Bucalossi e A. Ratti), Milano 1971, pp. 25-39.
Handley, R. S., Observations and thoughts on cancer of the breast, in ‟Proceedings of the Royal Society of Medicine", 1972, LXV, pp. 437-444.
Harkness, M. N., Bern, H. A., Alfert, M., Goldstein, N. O., Cytochemical studies of hyperplastic alveolar nodules in the mammary gland of the C3H/HeCRGL mouse, in ‟Journal of the National Cancer Institute", 1957, XIX, pp. 1023-1029.
Harnden, D. G., Cytogenetics of human neoplasia, in Genetics of human cancer (a cura di J. J. Mulvihill, R. W. Miller e J. F. Fraumeni), New York 1977, pp. 87-104.
Harnden, D. G., Mclean, N., Langlands, A. O., Carcinoma of the breast and Klinefelter's syndrome, in ‟Journal of medical genetics", 1971, VIII, pp. 460-461.
Heidelberger, C., Chemical oncogenesis in culture, in ‟Advances in cancer research", 1973, XVIII, pp. 317-366.
Heidelberger, C., Oncogenic transformation of rodent cell lines by chemical carcinogenes, in Origins of human cancer (a cura di H. H. Hiatt, J. D. Watson e J. A. Winsten), Cold Spring Harbor 1977, pp. 1513-1520.
Henderson, E. S., Samaha, R. J., Evidence that drugs in multiple combinations have materially advanced the treatment of human malignancies, in ‟Cancer research", 1969, XXIX, pp. 2272-2280.
Hendrick, J. C., Franchimont, P., Radio-immunoassay of casein in the serum of normal subjects and of patients with various malignancies, in ‟European journal of cancer", 1974, X, pp. 725-730.
Heston, W. E., Genetic aspects of human cancer, in ‟Advances in cancer research", 1976, XXIII, pp. 1-21.
Heston, W. E., Staffee, C. H., Development of tumours in fetal and adult lung transplants, in ‟Journal of the National Cancer Institute", 1957, XVIII, pp. 779-788.
Hiatt, H. H., Watson, J. D., Winsten, J. A. (a cura di), Origins of human cancer, Cold Spring Harbor 1977.
Hilbe, G., Salzer, G. M., Hussl, H., Kutschera, H., Die Carcinom-Fefährdung der Resektionsmagens, in ‟Lagenbecks Archiv für klinische Chirurgie", 1968, CCCXXIII, pp. 142-150.
Hill, M. J., Metabolic epidemiology of dietary factors in large bowel cancer, in ‟Cancer research", 1975, XXXV, pp. 3398-3402.
Hill, M. J., The role of unsaturated bile acids in the etiology of large bowel cancer, in Origins of human cancer (a cura di H. H. Hiatt, J. D. Watson e J. A. Winsten), Cold Spring Harbor 1977, pp. 1627-1640.
Hill, M. J., Drasar, B. S., Aries, V. C., Growther, J. S., Hawksworth, G. M., Williams, R. E. O., Bacteria and etiology of cancer of large bowel, in ‟Lancet", 1971, I, pp. 95-100.
Hill, M. J., Goddard, P., Williams, R. E. O., Gut bacteria and aetiology of cancer of the breast, in ‟Lancet", 1971, II, pp. 472-473.
Hirafuku, I., On the histopathological changes of the gastric mucosa with reference to the early stages of stomach cancer, in the morphological precursors of cancer (a cura di L. Severi), Perugia 1962, pp. 601-604.
Hollaender, A., Chemical mutagens: principles and methods for their detection, New York 1971.
Hudson, C. N., Testing anti-cancer drugs, in ‟Lancet", 1972, I, p. 1116.
Hueper, W. C., Occupational tumors and allied diseases, Springfield 1942.
Hueper, W. C., Conway, W. D., Chemical carcinogenesis and cancers, Springfield 1964.
Hueper, W. C., Payne, W. W., Carcinogenic studies on soot of coffee-roasting plants, in ‟Archives of pathology", 1960, LXIX, pp. 716-727.
Ioachim, H. L., The stromal reaction of tumors: an expression of immunesurveillance, in ‟Journal of the National Cancer Institute", 1976, LVII, pp. 465-475.
Jensen, H. M., Rice, J. R., Wellings, S. R., Preneoplastic lesions in the human breast, in ‟Science", 1976, CXCI, pp. 295-297.
Jensen, H. M., Wellings, S. R., Preneoplastic lesions of the human mammary gland transplanted into the nude athymic mouse, in ‟Cancer research", 1976, XXXVI, pp. 2605-2610.
Johnson, L. D., Easterday, C. L., Gore, H., Hertig, A. T., The histogenesis of carcinoma in situ of the uterine cervix, in ‟Cancer", 1964, XVII, pp. 213-229.
Kelly, M. G., O'Gara, R. W., Induction of tumours in newborn mice with dibenza, hanthracene and 3-methylcholanthrene, in ‟Journal of the National Cancer Institute", 1961, XXVI, pp. 651-679.
Kersey, J. H., Spector, B.D., Good, R. A., Primary immunodeficiency and cancer, in ‟Advances in cancer research", 1973, XVIII, pp. 211-230.
Kessler, I. I., Kulcar, Z., Zimolo, A., Grgurevic, M., Strnad, M., Goodwin, B.J., Cervical cancer in Yugoslavia. II. Epidemiologic factors of possible etiologic significance, in ‟Journal of the National Cancer Institute", 1974, LIII, pp. 51-60.
Klein, E., Klein, G., A system for the detection of single gene mutations in mouse tumours, in Proceedings of the II international symposium on mammary cancer (a cura di L. Severi), Perugia 1958, pp. 709-712.
Klein, G., Klein, E., Some experiments on the mechanism of progression in mouse mammary carcinoma, in Proceedings of the II international symposium on mammary cancer (a cura di L. Severi), Perugia 1958, pp. 713-717.
Klein, M., Lung adenomas in offspring following injection of pregant mice with urethan, in ‟Cancer research", 1952, XII, p. 275.
Knudson, A. G., Mutation and cancer. Statistical study of retinoblastoma, in ‟Proceedings of the National Academy of Sciences", 1971, LXVIII, pp. 820-823.
Knudson, A. G., Mutation and human cancer, in ‟Advances in cancer research", 1973, XVII, pp. 317-352.
Knudson, A. G., Genetic predisposition to cancer, in Origins of human cancer (a cura di H. H. Hiatt, J. D. Watson e J. A. Winsten), Cold Spring Harbor 1977, pp. 45-52.
Korpassy, B., Concezioni nuove sul meccanismo dello sviluppo di metastasi neoplastiche, in ‟Lo sperimentale", 1956, CVI, pp. 344-350.
Korpassy, B., Szönyi, F., Baradnay, G., Experimentelle Untersuchungen zur Frage der Seltenheit von Geschwulstmetastasen in der Skeletmuskulatur, in ‟Zeitschrift für Krebsforschung", 1958, LXII, pp. 370-374.
Koss, L. G., Diagnostic cytology and its histopathologic bases, Philadelphia 19682.
Leighton, J., The spread of cancer: pathogenesis, experimental methods, interpretation, New York-London 1967.
Lilienfeld, A. M., Levin, M. L., Kessler, I. I., Cancer in the United States, Cambridge, Mass., 1972.
Lippman, M., Bolan, G., Huff, K., The effects of estrogens and antiestrogens on hormone-responsive human breast cancer in long-term tissue culture, in ‟Cancer research", 1976, XXXVI, pp. 4595-4601.
Lippman, M., Bolan, G., Huff, K., The effects of glucocorticoids and progesterone on hormone-responsive human breast cancer in long-term tissue culture, in ‟Cancer research", 1976, XXXVI, pp. 4602-4608.
Lippman, M., Bolan, G., Huff, K., The effects of androgens and antiandrogens on hormone-responsive human breast cancer in long-term tissue culture, in ‟Cancer research", 1976, XXXVI, pp. 4610-4618.
Livingston, R. B., Carter, S. K., Single agents in cancer chemotherapy, New York 1970.
Locatelli, L., Alcuni aspetti del problema del cancro del polmone: concordanze e discordanze diagnostiche, in ‟Rivista di anatomia patologica e di oncologia", 1956, XI, pp. 1023-1060.
Lukes, R. J., Collins, R. D., A functional approach to the classification of malignant lymphoma, in Diagnosis and therapy of malignant lymphoma (a cura di K. Musshoff), Berlin-Heidelberg-New York 1974, pp. 18-30.
Lukes, R. J., Collins, R. D., Immunologic characterization of malignant lymphoma, in ‟Cancer", 1974, XXXIV, pp. 1488-1503.
Lukes, R. J., Collins, R. D., New approaches to the classification of the lymphoomata, in ‟British journal of cancer", 1975, XXXI, suppl. 11, pp. 1-27.
Lynch, H. T., Hereditary factors in carcinoma, Berlin-Heidelberg-New York 1967.
McConnell, R. B., The genetics of gastrointestinal disorders, London-New York 1966, pp. 156-169.
McDivitt, R. W., Stewart, F. W., Berg, J. W., Tumors of the breast, in AFIP Atlas of tumor pathology (a cura di H. I. Firminger), second series, fasc. 3, Washington 1968, pp. 1-156.
McGuire, W. L., Carbone, P. P., Vollmer, E. P., Estrogen receptors in human breast cancer, New York 1975.
MacMahon, B., Cole, P., Brown, J., Etiology of human breast cancer, in ‟Journal of the National Cancer Institute", 1973, L, pp. 21-42.
MacMahon, B., Levy, M. A., Prenatal origin of childhood leukemia, in ‟New England journal of medicine", 1964, CCLXX, pp. 1082-1085.
Markson, L. M., Terleki, S., The experimental transmission of ovine pulmonary adenomatosis, in ‟Pathologic veterinary", 1964, I, pp. 269-288.
Mathé, G., Aseptic environments and cancer treatment, Berlin-Heidelberg-New York 1971.
Mathé, G., Amiel, J. L., Scharzenberg, L., Schneider, M., Cattan, A., Schlumberger, J. R., Hayat, M., De Vassal, F., Active immunotherapy for acute lymphoblastic leukaemia, in ‟Lancet", 1969, I, pp. 697-699.
Math, G., Pouillart, P., Schwarzenberg, L., Hayat, M., De Vassal, F., Lafleur, M., Acute leukemias. Prognostic factors and criteria of response, in Cancer therapy. Prognostic factors and criteria of response (a cura di M. J. Staquet), New York 1975, pp. 89-106.
Meissner, W. A., Warren, S., Tumori, in Patologia (a cura di W. A. D. Anderson), vol. I, Roma 19742, pp. 542-575.
Meriwether, W. D., Bachur, N. R., Inhibition of DNA and RNA metabolism by daunorubicin and adriamycin in L 1210 mouse leukemia, in ‟Cancer research", 1972, XXXII, pp. 1137-1142.
Mihich, E., Drugs and cell regulation, New York-London 1971.
Miller, R. W., Neoplasia and Down's syndrome, in ‟Annals of the New York Academy of Sciences", 1970, CLXXI, pp. 637-644.
Moertel, C. G., Carcinoma of the stomach. Prognostic factors and criteria of response (a cura di M. J. Staquet), New York 1975, pp. 229-236.
Montesano, R., Bartsch, H., Mutagenic and carcinogenic N-nitroso compounds: possible environmental hazards, in ‟Mutation research", 1976, XXXII, pp. 179-185.
Moore, D. H., Charney, J., Kramarski, B., Lasfargues, E. Y., Sarkar, N. H., Brennan, M. J., Burrows, J. H., Sirsat, S. M., Paymaster, J. C., Vaidya, A. B., Search for a human breast cancer virus, in ‟Nature", 1971, CCXXIX, pp. 611-615.
Mueller, C. F., Thornbury, J. R., Adenocarcinoma of the colon complicating ureterosigmoidostomy. A case report and review of the literature, in ‟Journal of urology", 1973, CIX, pp. 225-227.
Mühlbock, O., Ebbenhorst Tengbergen, W. van, Rijssel, T. G. van, Studies on the development of mammary tumors in dilutebrown DBAb mice without the agent, in ‟Journal of the National Cancer Institute", 1952, XIII, pp. 505-531.
Mulvihill, J. J., Miller, R. W., Fraumeni, J. F. (a cura di), Genetics of human cancer, New York 1977.
Münck, W., Autopsy finding and clinical diagnosis. A comparative study of 1000 cases, in ‟Acta medica scandinavica", 1952, CXLII, suppl. 266, pp. 775-781.
Murad, T. M., A proposed histochemical and electron microscopic classification of human breast cancer according to cell origin, in ‟Cancer", 1971, XXVII, pp. 288-299.
Murad, T. M., Cytologic differentiation of carcinoma of the breast by electron microscopy, in ‟Acta cytologica", 1971, XV, pp. 400-409.
Nandi, S., Effect of hormones on the maintenance of hyperplastic alveolar nodules in the mammary glands of various strains of mice, in ‟Journal of the National Cancer Institute", 1961, XXVII, pp. 187-201.
Nandi, S., New method for detection of mouse mammary tumor virus. I. Influence of foster nursing on incidence of hyperplastic mammary nodules in BALB/cCrgl mice, in ‟Journal of the National Cancer Institute", 1963, XXXI, pp. 57-73.
Nandi, S., IV. Precancer, in Breast cancer research, UICC technical report series (a cura di M. J. Brennan), vol. XXVII, Genève 1977, pp. 59-66.
Nandi, S., Models for transformation of breast cells in animals and man, in Abstracts of the 11th meeting on mammary cancer in experimental animals and man (a cura di M. J. Brennan e M. A. Rich), Detroit 1978, p. 25.
Nandi, S., Bern, H. A., DeOme, K. B., Hormonal induction and maintenance of precancerous hyperplastic alveolar nodules in the mammary glands of hypophysectomized female C3H/HeCrgl mice, in ‟Acta Unionis internationalis contra cancrum", 1960, XVI, pp. 211-224.
Nowell, P. C., Hungerford, D.A., Chromosome studies in human leukemia. II. Chronic granulocytic leukemia, in ‟Journal of the National Cancer Institute", 1961, XXVII, pp. 1013-1035.
O'Grady, W. P., McDivitt, R. W., Breast cancer in a man treated with diethylstilbestrol, in ‟Archives of pathology", 1969, LXXXVIII, pp. 162-166.
Olivi, M., Caschera, F., Barbieri, G., Mammary cancer development in the RIII/Dm/Se substrain of mice studied by the histological and ‛whole mount' methods, in Proceedings of the II international symposium on mammary cancer (a cura di L. Severi), Perugia 1958, pp. 617-626.
Orr, J. W., The chemical induction of mammary and ovarian tumours, in ‟Acta Unionis internationalis contra cancrum", 1956, XII, pp. 682-689.
Orye, E., Delbeke, M. J., Vandenabeele, B., Retinoblastoma and arm deletion of chromosome 13. Attempts to define the deleted segment, in ‟Clinical genetics", 1974, V, pp. 457-464.
Ott, G., Fremdkörpersarkome, in Experimentelle Medizin, Pathologie und Klinik (a cura di F. Lenthardt, R. Schoen, H. Schwiegk, A. Studer e H. U. Zollinger), Berlin-Heidelberger-New York 1970.
Peller, S., Cancer in childhood and youth, Bristol 1960.
Pingitore, R., La diffusione metastatica di 783 tumori maligni al tavolo anatomico, in ‟Tumori", 1972, LVIII, pp. 319-326.
Pitelka, D. R., Bern, H. A., Nandi, S., DeOme, K. B., On the significance of virus-like particles in mammary tissues of C3Hf mice, in ‟Journal of the National Cancer Institute", 1964, XXXIII, pp. 867-885.
Potter, J. F., Longenbaugh, G., Chu, E., Dillon, I., Romsdahl, M., Malmgren, R. A., The relationship of tumor type and resectability to the incidence of cancer cells in blood, in ‟Surgery, gynecology and obstetrics", 1960, CX, pp. 734-738.
Powles, R. L., Growther, D., Bateman, C. J. T., Beard, M. E. J., McElwain, T. J., Russel, J., Lister, T. A., Whitehouse, J. M. A., Wrighley, P. F. M., Pike, M., Alexander, P., Hamilton Fairley, G., Immunotherapy for acute myelogenous leukaemia, in ‟British journal of cancer", 1973, XXVIII, pp. 365-376.
Prehn, R. T., Role of tumor antigens in the biology of neoplasia, in Cellular antigens (a cura di A. Nowotny), New York 1972, pp. 308-320.
Prehn, R. T., Immunological surveillance: pro and con, in Clinical immunobiology (a cura di F. H. Bach e R. A. Good), vol. II, New York 1974, pp. 191-210.
Prehn, R. T., Tumor progression and homeostasis, in ‟Advances in cancer research", 1976, XXIII, pp. 203-236.
Pruitt, J. C., Hilberg, A. W., Kaiser, R. F., Malignant cells in peripheral blood, in ‟New England journal of medicine", 1958, CCLIX, pp. 1161-1164.
Rapp, F., Herpesvirures and cancer, in ‟Advances in cancer research", 1974, XIX, pp. 265-302.
Reddy, B. S., Mastromarino, A., Wynder, E. L., Further leads on metabolic epidemiology of large bowel cancer, in ‟Cancer research", 1975, XXXV, pp. 3403-3406.
Reddy, B. S., Wynder, E. L., Large bowel carcinogenesis. Fecal constituents of populations with diverse incidence rates of colon cancer, in ‟Journal of the National Cancer Institute", 1973, L, pp. 1437-1442.
Robbins, J. H., Kraemer, K. H., Lutzner, M. A., Festoff, B. W., Coohn, H. G., Xeroderma pigmentosum. An inherited disease with sun sensitivity, multiple cutaneous neoplasms and abnormal DNA repair, in ‟Annals of internal medicine", 1974, LXXX, pp. 221-248.
Robbins, S. L., Pathologic basis of disease, Philadelphia-London-Toronto 1974.
Roberts, D. W., Loehr, E. V., Depression of thymidylate synthetase activity in response to cystosine arabinoside, in ‟Cancer research", 1972, XXXII, pp. 1160-1169.
Roberts, J. J., Modificazioni degli acidi nucleici e tumori, in Biologia dei tumori (a cura di E. J. Ambrose e J. C. Roe), Napoli 1978, pp. 212-239.
Ross, W. C. J., Biological alkylating agents, London 1962.
Rotkin, I. D., Further studies in cervical cancer inheritance, in ‟Cancer", 1966, XIX, pp. 1251-1268.
Rous, P., Beard, J. W., The progression to carcinoma of virus-induced rabbit papillomas (Shope), in ‟Journal of experimental medicine", 1935, LXII, pp. 523-548.
Rozencweig, M., Kenis, Y., Staquet, M., Hodgkin's disease. Prognostic factors and clinical evaluation, in Cancer therapy. Prognostic factors and criteria of response (a cura di M. J. Staquet), New York 1975, pp. 49-72.
Schappert-Kimmijser, J., Hemmas, G. D., Nijland, R., The heredity of retinoblastoma, in ‟Ophtalmologica", 1966, CLI, pp. 197-213.
Schlom, J., Spiegelman, S., Simultaneous detection of the reverse transcriptase and high molecular weight RNA unique to the oncogenic RNA viruses, in ‟Science", 1971, CLXXIV, pp. 840-843.
Schlom, J., Spiegelman, S., Moore, D. H., Reverse transcriptase and high molecular weight RNA in particles from mouse and human milk, in ‟Joural of the National Cancer Institute", 1972, XXXXVIII, pp. 1197-1203.
Schmähl, D., Experimental studies in rats on the mechanism of the blood-borne metastases process, in ‟Acta Unionis internationalis contra cancrum", 1959, XV, pp. 898-906.
Schmähl, D., Rieseberg, T., Experimentelle Untersuchungen an Ratten über die Metastasierung von Tumoren, in ‟Zeitschrift für Krebsforschung", 1958, LXII, pp. 456-480.
Schmähl, D., Thomas, C., Auer, R., Iatrogenic carcinogenesis, Berlin-Heidelberg-New York 1977.
Schnitzer, R. J., Hawking, F., Experimental chemotherapy, New York-London 1966.
Schoental, R., Toxicology and carcinogenic action of pyrrolizidine alkaloids, in ‟Cancer research", 1968, XXVIII, pp. 2237-2246.
Semisch, R., Neue Ansichten über die peripheren Lungenzirkulation und ihre Folgerungen bezüglich der Metastasierung Fatt und Thromboembolie, in ‟Archiv für klinische Chirurgie", 1959, CCLXXXXII, pp. 294-301.
Severi, L., La mastopatia fibrocistica con particolare riguardo al cancro della mammella, in ‟Lavori dell'Istituto di Anatomia e Istologia Patologica, Università degli Studi, Perugia", 1952, XII, pp. 145-187.
Severi, L. (a cura di), The morphological precursors of cancer, Perugia 1962.
Severi, L., Olivi, M., Biancifiori, C., Istopatologia del carcinoma mammario negli ‛inbred strains' del topo, in ‟Atti della Società Italina di Cancerologia", 1958, I, pp. 85-158.
Severi, L., Squartini, F., I quattro tempi della cancerogenesi mammaria del topo, in ‟Lavori dell'Istituto di Anatomia e Istologia Patologica, Università degli Studi, Perugia", 1955, XV, pp. 101-117.
Severi, L., Squartini, F., Die biomatrischen Probleme des Krebses, in ‟Zeitschrift für Krebsforschung", 1956, LXI, pp. 180-194.
Severi, L., Squartini, F., Problems arising in the control of breast cancer, in ‟Acta Unionis internationalis contra cancrum", 1962, XVIII, pp. 755-759.
Severi, L., Squartini, F., Epidemiological aspects of tumours among children in Italy with some pathological data for Umbria, in ‟Lavori dell'Istituto di Anatomia e Istologia Patologica, Università degli Studi, Perugia", 1969, XXIX, pp. 147-163.
Severi, L., Squartini, F., I precursori morfologici del cancro, in Trattato di oncologia clinica (a cura di P. Bucalossi e U. Veronesi), Milano 1973, pp. 104-126.
Shay, H., Weinhouse, S., Gruenstein, M., Marx, H. E., Friedman, B., Development of malignant lymphoma in some of the young rats suckled by mothers receiving methylcholanthrene by stomach tube only during the lactation period, in ‟Cancer", 1950, III, pp. 891-895.
Shimkin, M. B., Duration of life in untreated cancer, in ‟Cancer", 1951, IV, pp. 1-8.
Sieber, S. M., Adamson, R. H., Toxicity of antineoplastic agents in man. Chromosomal aberrations, antifertility effects, congenital malformations, and carcinogenic potential, in ‟Advances in cancer research", 1975, XXII, pp. 57-155.
Silver, R. T., Chronic lymphocytic leukemia. Proposed guidelines for protocol studies, in Cancer therapy. Prognostic factors and criteria of response (a cura di M. J. Staquet), New York 1975, pp. 115-129.
Sinha, D., Dao, T. L., Site of origin of mammary tumors induced by 7, 12-dimethylbenz(a) anthracene in the rat, in ‟Journal of the National Cancer Institute", 1975, LIV, pp. 1007-1009.
Skipper, H. E., Cancer chemotherapy is many things. G. H. A. Cloves memorial lecture, in ‟Cancer research", 1971, XXXI, pp. 1173-1180.
Smith, S. B., Thymus, in Pathology (a cura di W. A. D. Anderson e J. M. Kissane), vol. II, Saint Louis 19777, pp. 1575-1600.
Sohn, K. Y., Boggs, D. R., Klinefelter's syndrome, LSD usage and acute lymphoblastic leukemia, in ‟Clinical genetics", 1974, VI, pp. 20-22.
Spiegelman, S., Axel, R., Schlom, J., Virus related RNA in human and mouse mammary tumours, in ‟Journal of the National Cancer Institute", 1972, XXXXVIII, pp. 1205-1211.
Spiegelman, S., Burny, A., Das, M. R., Keydar, J., Schlom, J., Travniceck, M., Watson, K., Characterization of the products of RNA-directed DNA polymerase in oncogenic RNA viruses, in ‟Nature", 1970, CCXXVII, pp. 563-567.
Spratt, J. S., Ackerman, L. V., Moyer, C. A., Relationship of polyps of the colon to colonic cancer, in ‟Annals of surgery", 1958, CXXXXVIII, pp. 682-698.
Spratt, J. S., Watson, F. R., The rationale of pratice for polypoid lesions of the colon, in ‟Cancer", 1971, XXVIII, pp. 153-159.
Spriggs, A. I., Population screening by the cervical smear, in ‟Nature", 1972, CCXXXVIII, pp. 135-137.
Squartini, F., Nuovi orientamenti in tema di cancerogenesi intestinale umana, in ‟Annali della Facoltà di Medicina e Chirurgia della Università degli Studi di Perugia", 1957, XXXXVIII, pp. 67-74.
Squartini, F., Mammogenesis and breast carcinogenesis in virgin female mice of BALB/cf substrain with the milk agent, in ‟Journal of the national Cancer Institute", 1959, XXIII, pp. 1227-1238.
Squartini, F., Mammary tumors in mice as a model in studies on carcinogenesis, in Recent contributions to cancer research in Italy (a cura di P. Bucalossi e U. Veronesi), Milano 1960, pp. 289-335.
Squartini, F., Responsiveness and progression of mammary tumors in high-cancer-strain mice, in ‟Journal of the National Cancer Institute", 1962, XXVIII, pp. 911-926.
Squartini, F., Progressione e chemioterapia dei tumori, in ‟Lavori dell'Istituto di Anatomia e Istologia Patologica, Università degli Studi, Perugia", 1964, XXIV, pp. 83-93.
Squartini, F., Tumours arising from vertical transmission, in Handbuch der experimentellen pharmakologie, vol. XV, t. 13 (a cura di O. Eichler), Berlin-Heidelberg-New York 1966, pp. 1-73.
Squartini, F., III. Histogenesis, in Breast cancer research, UICC technical series (a cura di M. J. Brennan), vol. XXVII, Genève 1977, pp. 51-58.
Squartini, F., I virus oncogeni nel carcinoma mammario, in I tumori della mammella (a cura di U. Veronesi, A. Perussia, H. Emanuelli e M. De Lana), Milano 1977, pp. 15-25.
Squartini, F., The pathophysiology of breast cancer: speech of the Convener, in Proceedings of the XIth meeting on mammary cancer in experimental animals and man (a cura di M. J. Brennan e M. A. Rich), Detroit 1978.
Squartini, F., Barola, E., Confronto fra diagnosi cliniche e anatomo-patologiche per neoplasie maligne nel materiale autoptico del settorato di Perugia (1921-1964), in ‟Lavori dell'Istituto di Anatomia e Istologia Patologica, Università degli studi, Perugia", 1965, XXV, pp. 5-27.
Squartini, F., Bolis, G. B., Tumori dell'infanzia e dell'adolescenza: analisi statistica e classificazione di 579 casi, in ‟Lavori dell'Istituto di Anatomia e Istologia Patologica, Università degli Studi, Perugia", 1966, XXVI, pp. 59-84.
Squartini, F., Bolis, G. B., Detection of a lung tumour agent in BALB(c)/Cb/Se irradiated mice, in ‟Lavori dell'Istituto di Anatomia e Istologia Patologica, Università degli Studi, Perugia", 1970, XXX, pp. 125-128.
Squartini, F., Bolis, G. B., Sensibilità al metilcolantrene degli epiteli dell'albero respiratorio in topi BALB/c/Cb/Se, in ‟Lavori dell'Istituto di Anatomia e Istologia Patologica, Università degli Studi, Perugia", 1971, XXXI, pp. 35-56.
Squartini, F., Bolis, G. B., Barola, E., Metastasi in 717 autopsie consecutive per tumori maligni, in ‟Lavori dell'Istituto di Anatomia e Istologia Patologica, Università degli Studi, Perugia", 1968, XXVIII, pp. 109-118.
Squartini, F., Caschera, F., Basi patogenetiche del cancro intestinale (da uno studio sulla distribuzione di 1183 tumori dell'apparato digerente), in ‟Lavori dell'Istituto di Anatomia e Istologia Patologica, Università degli Studi, Perugia", 1956, XVI, pp. 375-381.
Squartini, F., Maltzeff, N., Nuovi orientamenti in tema di cancerogenesi duodenale umana, in ‟Lavori dell'Istituto di Anatomia e Istologia Patologica, Università degli Studi, Perugia", 1956, XVI, pp. 323-347.
Squartini, F., Olivi, M., Cancerogenesi mammaria da metilcolantrene: parte prima, in ‟Lavori dell'Istituto di Anatomia e Istologia Patologica, Università degli Studi, Perugia", 1962, XXII, pp. 5-72.
Squartini, F., Olivi, M., Severi, L., A reconsideration of lung tumours observed in the course of experiments on mammary carcinogenesis, in Lung tumours in animals (a cura di L. Severi), Perugia 1966, pp. 273-283.
Squartini, F., Rossi, G., Analisi morfologica della ‛responsiveness' e della progressione nei tumori mammari del ‛substrain' RIII/Dm/Se, in ‟Lavori dell'Istituto di Anatomia e Istologia Patologica, Università degli Studi, Perugia", 1959, XIX, pp. 165-214.
Squartini, F., Rossi, G., Studio dei tumori mammari nel BALB/cf/Cb/Se substrain: accrescimento, ‛responsiveness' e progressione, origine istologica, morfologia, in ‟Lavori dell'Istituto di Anatomia e Istologia Patologica, Università degli Studi, Perugia", 1960, XX, pp. 133-165.
Squartini, F., Rossi, G., Responsiveness and progression of the morphological precursors of breast cancer in inbred mice. A review, in The morphological precursors of cancer (a cura di L. Severi), Perugia 1962, pp. 319-327.
Squartini, F., Rossi, G., Paoletti, I., Characters of mammary tumours in BALB/c female mice foster-nursed by C3H and RIII mothers, in ‟Nature", 1963, CLXXXVII, pp. 505-506.
Squartini, F., Severi, L., Strain differences in the mammary tumour-inducing virus as detected by the characters and behaviour of neoplasms, in A Ciba Foundation symposium on tumour viruses of murine origin (a cura di G. E. W. Wolstenholma e M. O'Connor), London 1962, pp. 82-103.
Squartini, F., Severi, L., Problema biologico dei tumori della tiroide, in Corso superiore sulla terapia dei tumori della testa e del collo, Milano 1964, pp. 305-322.
Stansly, P. G., Sato, H., Cancer metastasis. Approaches to the mechanism, prevention and treatment, Gann monograph on cancer research n. 20, Baltimore 1978.
Staquet, M. J. (a cura di), Cancer therapy. Prognostic factors and criteria of response, New York 1975.
Steel, G. G., Lamerton, L. F., Cell population kinetics and chemotherapy, in ‟National Cancer Institute monographs", 1969, XXX, pp. 29-50.
Steiner, P. E., Human significance of experimental carcinogenesis, in ‟Archives of pathology", 1953, LV, pp. 227-244.
Stock, J. A., Chemioterapia dei tumori, in Biologia dei tumori (a cura di E. J. Ambrose e J. C. Roe), Napoli 1978, pp. 315-354.
Stoll, B. A., Endocrine therapy in malignant disease, London 1972.
Strong, L. C., Biological aspects of cancer and aging, Oxford-Braunschweig 1968.
Strong, L. C., Theories and pathogenesis: mutation and cancer, in Genetics of human cancer (a cura di J. J. Mulvihill, R. W. Miller e J. F. Fraumeni), New York 1977, pp. 401-415.
Stutman, O., Immunological aspects of resistance to the oncogenic effects of 3-methylcholanthrene in mice, in ‟Israel journal of medical sciences", 1973, IX, pp. 217-224.
Stutman, O., Immunodepression and malignancy, in ‟Advances in cancer research", 1975, XXII, pp. 261-422.
Stutman, O., Immunological surveillance, in Origins of human cancer (a cura di H. H. Hiatt, J. D. Watson, J. A. Winsten), Cold Spring Harbor 1977, pp. 729-750.
Thackray, A. C., Lucas, R. B., Tumors of the major salivary glands, in AFIP Atlas of tumor pathology (a cura di H. I. Firminger), second series, fasc. 10, Washington 1974, pp. 1-144.
Thomas, L., Discussion, in Cellular and humoral aspects of the hypersensitive states (a cura di H. S. Lawrence), New York 1959, pp. 529-532.
Toh, Y. C., Physiological and biochemical review of sex differences and carcinogenesis with particular reference to the liver, in ‟Advances in cancer research", 1973, XVIII, pp. 155-209.
Tuyns, A. J., L'épidémiologie du cancer du sein, in Corso superiore sul trattamento dei tumori della mammella (a cura di P. Bucalossi e A. Ratti), Milano 1971, pp. 11-23.
U.S. Department of Health, Education and Welfare, Smoking and health. Report of the Advisory Committee to the Surgeon General of the Public Health Service, Washington 1964.
Vaidya, A. B., Dion, A. S., Moore, D. H., Homology of mouse mammary tumor viral RNA with RNA from human breast cancer, in ‟Nature", 1974, CCIL, pp. 565-567.
Veale, A. M. O., Intestinal polyposis, London-New York 1965.
Vogel, E., Identification of carcinogens by mutagen testing in Drosophila. The relative reliability for the kinds of genetic damage measured, in Origins of human cancer (a cura di H. H. Hiatt, J. D. Watson e J. A. Winsten), Cold Spring Harbor 1977, pp. 1483-1497.
Wallace, A. C., Metastasis as an aspect of cell behavior, in ‟Canadian cancer conferences", 1961, IV, pp. 139-165.
Walter, J. B., Israel, S. M., General pathology, London-Toronto 19672.
Walther, H. F., Krebsmetastasen, Basel 1948.
Wellings, S. R., Jensen, H. M., On the origin and progression of ductal carcinoma of the human breast, in ‟Journal of the National Cancer Institute", 1973, L, pp. 1111-1118.
Wellings, S. R., Jensen, H. M., Marcum, R. G., An atlas of subgross pathology of the human breast with special reference to possible precancerous lesions, in ‟Journal of the National Cancer Institute", 1975, LV, pp. 231-273.
Wieberdink, J., Foetal haemic metastasis. An explanation of the ‛Papper-type' of metastasis in adrenal neuroblastoma, in ‟British journal of cancer", 1957, XI, pp. 378-383.
Willis, R. A., Pathology of tumours, London 1948, 19674.
Willis, R. A., The spread of tumours in the human body, London 1952, 19733.
Willis, R. A., The pathology of the tumours of children, Edinburgh-London 1962.
Winder, E. L., Some practical aspects of cancer prevention, in ‟New England journal of medicine", 1952, CCXXXXVI, pp. 573-582.
Winder, E. L., The epidemiology of large bowel cancer, in ‟Cancer research", 1975, XXXV, pp. 3388-3394.
Wood, S., Holyoke, D., Yardley, J. H., Mechanisms of metastasis production by blood-borne cancer cells, in ‟Canadian cancer conferences", 1961, IV, pp. 167-223.
Yoshida, T. H., Karyotype of tumor cells and their ability to invade organs, in ‟Gann" (Suppl.), 1959, L, pp. 210-211.
Young, J. S., The invasive growth of malignant tumours. An experimental interpretation based on elastic-jelly models, in ‟Journal of pathology and bacteriology", 1959, LXXVII, pp. 321-339.
Zamcheck, N., Moore, T. L., Dhar, P., Kupchik, H., Immunologic diagnosis and prognosis of human digestive tract cancer. Carcinoembryonic antigens, in ‟New England journal of medicine", 1972, CCLXXXVI, pp. 83-86.
Zeidman, I., Metastasis. A review of recent advances, in ‟Cancer research", 1957, XVII, pp. 157-162.
Ziegler, J. L., Spontaneous remission in Burkitt's lymphoma, in ‟National Cancer Institute monographs", 1976, XXXXIV, pp. 61-62.
Oncologia sperimentale
di Luigi Califano
sommario: 1. Introduzione: a) cenni storici sullo sviluppo delle moderne ricerche di oncologia; b) i tumori spontanei degli animali; c) i ceppi inbred; d) la coltura di cellule in vitro; e) il fenomeno della trasformazione. 2. Cancerogenesi chimica: a) idrocarburi; b) azocomposti; c) uretano; d) mostarde, etilenimmine, epossidi; e) aflatossina e cicasina; f) agenti chimici endogeni; g) etionina; h) metalli e non metalli; i) cenni sui meccanismi d'azione dei carcinogeni chimici; l) cocancerogenesi; m) il fumo del tabacco. 3. Cancerogenesi fisica: a) radiazioni ultraviolette; b) raggi X, isotopi radioattivi, radiazioni ionizzanti; c) cancerogenesi da solidi. 4. Cancerogenesi virale: a) caratteristiche generali dei virus oncogeni; b) virus polioma e SV40; c) tumori da virus erpetici: il linfoma di Burkitt e l'adenocarcinoma di Lucké del rene di rana; d) il sarcoma di Rous; e) il complesso sarcoma-leucemia del topo; f) il fattore latte. 5. Cenni di biologia molecolare della cancerogenesi. 6. Metabolismo energetico della cellula neoplastica. 7. Cenni di immunologia oncologica. 8. Ormoni e cancro. 9. Conclusioni. □ Bibliografia.
1. Introduzione
a) Cenni storici sullo sviluppo delle moderne ricerche di oncologia
Il problema del cancro, uno dei più travagliati della medicina per le difficoltà diagnostiche di molte delle sue forme e per la limitata efficacia dei sussidi terapeutici, da pochi anni è divenuto anche uno dei più complessi della biologia.
Si può a ragione dire ‛da pochi anni', perché in effetti solo molto recentemente si è avuta tale concentrazione di sforzi nelle ricerche da dare l'impressione che la cancerologia sia disciplina nuova o nuovissima; se si paragona ciò che si conosce oggi con quanto si sapeva nei primi trenta anni di questo secolo, si ha l'impressione di due mondi culturali diversi.
Ciò non significa che non fosse viva, già negli ultimi decenni del secolo scorso, l'ansia di conoscere da una parte la causa o le cause del cancro, dall'altra il meccanismo o i meccanismi con cui un tessuto diventa canceroso. I progressi nelle conoscenze erano, però, pochi e in parte incerti, onde la diffusa sfiducia di conseguire solide acquisizioni, e ciò in contrasto con quanto era avvenuto o avveniva per le malattie infettive, delle quali venivano progressiva- mente isolati i vari agenti patogeni, con scoperte seguenti a scoperte in quella che ancora oggi si indica con la romantica espressione di ‛epoca d'oro della batteriologia'.
Sembrava, in sostanza, che il mistero del cancro fosse insolubile in quanto legato al mistero stesso della vita, e quando affiorò qualche scoperta, che solo dopo molti anni fu riconosciuta di essenziale importanza, essa fu riguardata, anche da eminenti patologi, con scetticismo e considerata piuttosto come curiosità biologica, priva di significato generale.
Tanto scetticismo proveniva principalmente dal fatto che i tumori colpiscono l'uomo a caso, come a caso insorgono in uno o in un altro organo, e che mai era stato possibile dimostrare la loro contagiosità anche tra persone viventi nello stesso ambiente e con abitudini di vita praticamente identiche, sebbene fossero note le cosiddette ‛famiglie a cancro', cioè famiglie con notevole incidenza di tumori maligni.
L'epoca che potrebbe dirsi scientifica dell'oncologia ebbe inizio nel 1908, quando V. Ellermann e O. Bang (v., 1908) scoprirono che una forma di leucemia dei polli era prodotta da un virus filtrabile, e si consolidò pochi anni dopo, quando P. Rous (v., 1911) scoprì che un sarcoma del pollo era anch'esso prodotto da un virus filtrabile. In realtà, Giuseppe Sanarelli nel 1898 aveva già scoperto che il mixoma del coniglio era prodotto da un virus filtrabile (v. Sanarelli, 1898), ma né la scoperta di Ellermann e Bang, né quella di Rous e meno ancora quella di Sanarelli suscitarono molto interesse tra gli studiosi, per la già accennata considerazione che nell'uomo la leucemia e il sarcoma non mostrano caratteri di contagiosità tali da far sospettare una eziologia da agenti infettivi quali sono i Virus; per quanto riguarda il mixoma del coniglio, inoltre, sorsero dubbi sulla sua natura neoplastica, dubbi che tuttora permangono.
Successivamente fu dimostrata l'eziologia virale di altri tumori di Mammiferi, ma la situazione essenzialmente non mutò fino a quando K. Yamagiwa e K. Ichikawa (v., 1914) scoprirono che la pennellatura con catrame della cute del padiglione dell'orecchio del coniglio, ripetuta per molti mesi, causava un tumore epiteliale in tutto simile ad alcune forme di tumori umani. Da tale scoperta prese l'avvio l'affannosa ricerca della sostanza o delle sostanze contenute nel catrame capaci di determinare formazione di tumori, e con ciò ebbe inizio l'era degli studi sulla cancerogenesi chimica che tanta luce doveva portare nell'oscuro problema.
In quegli anni inizio, in altra direzione, l'opera di O. Warburg (v., 1926) che portò alla scoperta di quel peculiare metabolismo della cellula cancerosa, la glicolisi aerobica, che tuttora è considerato la sua più importante diversità metabolica rispetto alla cellula normale.
Successivamente, dopo la scoperta che da attività cancerogena sono caratterizzati non solo molti idrocarburi, ma anche sostanze che con questi non hanno alcuna relazione strutturale, l'elenco dei carcinogeni chimici si è enormemente accresciuto e sono stati isolati o sintetizzati sia idrocarburi naturali o artificiali a enorme attività oncogena, sia sostanze da questi differenti in grado di produrre tumori di vari organi. Oggi si dispone pertanto di composti capaci di indurre tumori sperimentali in animali di differenti specie e in organi diversi, si e cioè in grado di cancerizzare un tessuto a volontà.
Con tali scoperte, naturalmente, sono sorti nuovi e più intricati problemi, primo tra tutti quello di comprendere come sostanze chimicamente tanto dissimili tra loro producano praticamente sempre lo stesso effetto, e se i vari carcinogeni chimici, o almeno le varie classi di questi, agiscano infine agli stessi livelli del metabolismo cellulare ovvero in siti differenti.
Alla quasi indifferenza per le prime scoperte di virus oncogeni, è succeduto negli ultimi anni un gran fervore di ricerche su di essi, non inferiore a quello che suscitò la cancerogenesi chimica. Le scoperte, da parte di Shope, del fibroma (v. Shope, 1932) e poi del papilloma del coniglio (v. Shope, 1934) segnarono certamente passi importanti nello sviluppo dell'oncogenesi virale. Questa, al momento attuale, costituisce il campo di maggiore interesse, sia per sperimentazione sia per considerazioni generali, soprattutto da quando è stato dimostrato che vari altri tumori di animali, appartenenti a generi e specie diverse, sono prodotti da virus, identificati alla microscopia elettronica e in colture di cellule. La scoperta del virus polioma (v. Gross, 1953; v. Stewart, 1953) ha segnato una svolta decisiva in questo settore di studi, perché esso è risultato capace di determinare in uno stesso ospite tumori a differente struttura istologica, mentre gli altri virus studiati in precedenza determinano in genere un solo tipo di tumore o tumori di tipi istologici molto affini. La possibilità di ottenere allo stato di grande purificazione e in quantità cospicue il virus polioma ha consentito di eseguire indagini altrimenti impossibili. Molto ha contribuito all'attuale interesse per i virus oncogeni anche la scoperta effettuata da J. J. Bittner, nel 1936 (v. Bittner, Some possible..., e The receptibility..., 1936), di un virus presente nel latte di alcuni ceppi di topi, che si trasmette dalla madre alla prole, scoperta che ha suscitato la speranza di poter individuare un analogo fattore per il cancro della mammella della donna. Molto ha contribuito e contribuisce alle ricerche sui virus oncogeni la scoperta del virus del sarcoma-leucemia del topo, di quello dell'adenocarcinoma del rene della rana e, per quanto più strettamente riguarda la patologia umana, lo studio del tumore di Burkitt dei bambini dell'Uganda; ancor più recentemente, inoltre, sono stati scoperti i virus di tumori di scimmie, di Felini e di vari altri animali.
Tutto ciò ha portato a quella che può dirsi la riscoperta del virus di Rous, di cui sono stati isolati vari ceppi con proprietà alquanto differenti tra loro, alcuni dei quali sono anche trasmissibili in Mammiferi. Solo, dunque, dopo cmquant'anni è stata giustamente valorizzata una grande scoperta, così che al Rous venne conferito il premio Nobel per la medicina nel 1966, quando egli aveva ormai ottantasette anni.
Naturalmente, le ricerche di virologia oncologica sono state rese possibili dagli enormi sviluppi delle tecniche di identificazione e di isolamento dei Virus in generale, e in particolare dai metodi elaborati per lo studio del fago e dalla grande mole di notizie raccolte abbastanza rapida- mente su questo agente.
Uno dei mezzi tecnici più utili per tali ricerche è risultato quello delle colture in vitro, largamente impiegate in biologia e particolarmente in virologia quale mezzo principale per la coltura dei Virus; sono stati in tal modo ottenuti risultati interessanti anche dal punto di vista pratico, in quanto si è giunti a selezionare ceppi di virus a scarsissimo o nullo potere patogeno, ma a capacità immunizzante corrispondente a quella dei ceppi virulenti. In seguito a questi studi si sono potuti preparare il vaccino Sabin contro il virus della poliomielite, e successivamente il vaccino antimorbilloso. Dell'impiego delle colture in vitro negli studi di oncologia sarà detto successivamente; qui basti accennare che esse hanno reso possibile realizzare in vitro la cosiddetta trasformazione, cioè l'induzione alla cancerizzazione di una singola cellula che, in dipendenza di tale evento, diventa distinguibile dàlle cellule normali circostanti. È stato anche possibile, per mezzo delle colture in vitro, isolare e distinguere per uno stesso virus tipi diversi, alcuni dotati e altri privi di attività oncogena; un tipico esempio è rappresentato dagli adenovirus dell'uomo.
La penetrazione di un virus in una cellula della quale determina la trasformazione significa introduzione di materiale genico virale nel genoma della cellula ospite. Si arriva così dal livello cellulare e subcellulare a quello più strettamente molecolare, cioè all'interazione tra acido nucleico cellulare e acido nucleico virale o tra prodotti della loro attività. Con ciò il problema dell'oncogenesi è diventato, nella sua essenza, problema di biologia molecolare e non pochi progressi di questa relativamente nuova disciplina traggono origine e sviluppo dall'oncologia sperimentale.
La constatazione della relativa frequenza di neoplasie in persone ripetutamente esposte all'azione di radiazioni di vario tipo e la produzione sperimentale di tumori a mezzo di radiazioni ha portato a riconoscere anche una cancerogenesi fisica. Fisica è anche la cancerogenesi da solidi, il cui recente riconoscimento è dovuto all'osservazione di quei tumori che si sviluppano per introduzione negli animali di lamine di sostanze plastiche o di alcuni metalli.
Si suole, pertanto, classificare gli agenti oncogeni in chimici, fisici e biologici (Virus): classificazione arbitraria, come tutte le classificazioni, ma tuttora usata, anche se appare chiaro che il meccanismo dell'oncogenesi deve essere indagato, come si è detto, a quei livelli del metabolismo sui quali direttamente o indirettamente si esercita l'influenza degli agenti carcinogenetici, quale che ne sia l'origine e la natura.
Lo studio dell'eziologia e della patogenesi delle malattie dell'uomo è tanto più agevole quanto più fedele ne è la riproduzione in animali da laboratorio. Secondo uno dei postulati di R. Koch, perché un germe possa sicuramente ritenersi responsabile di una malattia infettiva la sua inoculazione in animali da esperimento deve determinare la comparsa di espressioni cliniche e anatomopatologiche proprie della malattia in studio.
Con la produzione della malattia sperimentale è altamente facilitata la scoperta di mezzi terapeutici quali i chemioterapici e gli antibiotici.
Se la malattia umana non è riproducibile negli animali o se in questi non si riscontra una malattia spontanea a essa paragonabile, i progressi nelle conoscenze sono molto più lenti, perché la base di sperimentazione è enormemente più limitata.
Da ciò l'origine di una delle più importanti discipline medico-biologiche, cioè della medicina sperimentale, e la conseguente creazione di istituti di libera ricerca nel mondo, quali l'Institut Pasteur di Parigi, il Preussisches Institut (poi Koch's Institut) für Infektionskrankheiten di Berlino, il Rockefeller Institute for Medical Research di New York, e tanti altri.
b) I tumori spontanei degli animali
Sino agli inizi del XIX secolo si riteneva, in generale, che il cancro fosse una malattia esclusiva dell'uomo non riscontrabile negli animali. Tale convinzione doveva poi cadere, quando si incominciò a osservare l'incidenza di tumori benigni e maligni in molte specie animali. Talvolta furono anche compiuti tentativi di trapianto di tumori spontanei in animali della stessa specie, con risultati in alcuni casi positivi, ma invalidati dall'interruzione della trasmissione. I primi consistenti tentativi di trapiantare tumori spontanei da animale ad animale della stessa specie furono compiuti in vari laboratori (v. Loeb, 1901; v. Borrel, 1903; v. Jensen, 1903), ma il maggiore impulso fu dato da P. Ehrlich e H. Apolant nel 1905 con il trapianto di sarcomi e specialmente di adenocarcinomi mammari del topo.
Tumori di quest'ultimo tipo sono tuttora impiegati in molti laboratori, specialmente nella forma ascitica; l'introduzione endoperitoneale di cellule isolate è seguita dalla comparsa di ascite, e in tale liquido le cellule continuano a moltiplicarsi rimanendo libere le une dalle altre, sospese nel liquido. In tal modo è possibile eseguire ricerche confrontabili, almeno entro certi limiti, con quelle condotte su cellule cancerose coltivate in vitro.
Nei trapianti iniziali di tumori la percentuale di attecchimento variava molto, per cui non si ottenevano risultati costanti. Ehrlich vide che il trapianto frequente faceva diminuire lo scarto e credette che tale fatto fosse dovuto a un fenomeno di ‛virulentazione' del ceppo di tumore, in analogia a quanto era stato osservato per alcuni microbi che, trasferiti frequentemente da animale ad animale, acquistano maggiore virulenza, cioè incremento della capacità di uccidere gli animali da esperimento: in altre parole, a seguito dei ripetuti passaggi si otteneva una progressiva diminuzione, fino a un dato livello stazionario, del numero dei microbi necessario per l'effetto letale. E ora ben noto che il fenomeno dell'esaltazione della virulenza dei Batteri dipende dalla selezione che i poteri immunitari dell'organismo operano nei loro confronti, per cui muoiono i germi meno virulenti e sopravvivono quelli dotati di virulenza più elevata, onde alla fine solo questi si moltiplicano. In modo analogo si può spiegare la virulentazione delle cellule neoplastiche, cioè come un fenomeno in rapporto non all'acquisizione di nuove proprietà, bensì all'eliminazione delle cellule immunosensibili.
Nonostante tali acquisizioni, il trapianto di tumori da animale ad animale, specie per alcuni tipi di neoplasie e per alcune specie, dava sempre risultati non soddisfacenti proprio per l'incostanza dei risultati. Un passo decisivo fu compiuto con la selezione di ceppi di topi geneticamente puri.
c) I ceppi inbred
I ceppi inbred si selezionano incrociando fratelli e sorelle e talvolta genitori e figli. I primi risultati furono ottenuti da C. C. Little dell'Università di Harward selezionando il ceppo DBA (Dilute Brown), ma solo nel 1919, dopo varie traversie, Little a Cold Spring Harbor (v. Little, 1947) riuscì a ottenere allevamenti consistenti di tale ceppo, che tuttora è largamente impiegato (v. Gross, 1970). Un ceppo famoso è il C3H, ottenuto da L. C. Strong (v., 1935), che presenta alta incidenza spontanea di tumore della mammella. In parallelo fu isolato un ceppo detto X (CBA di Strong), sempre inbred, a bassissima frequenza di tumore della mammella; e così fu resa possibile la scoperta di Bittner (v., Some possible..., e The receptibility..., 1936) del fattore del latte, cui si è precedentemente accennato.
Un ceppo di topi molto in uso è lo Swiss, allevato originariamente in un laboratorio svizzero e poi in America e in altri paesi.
Si conosce anche un ceppo inbred con frequenza del cinquanta per cento di tumori delle ossa. I ricercatori dispongono oggi di ceppi nei quali la frequenza spontanea di un dato tipo di tumore è assai elevata e di ceppi, utilizzabili come controllo, nei quali tale frequenza è al contrario bassa o nulla. Sono stati progressivamente isolati ceppi, inbred ad alta e rispettivamente bassa incidenza di tumori di vari organi e di leucemie, che costituiscono praticamente il materiale sperimentale di ogni laboratorio di oncologia.
Al fine di disciplinare e standardizzare la nomenclatura dei ceppi inbred di topi fu costituito nel 1952 un comitato il quale periodicamente integra la lista dei ceppi già noti con quelli di nuovo isolamento.
Anche per altre specie animali sono disponibili ceppi inbred, ma i caratteri sono meno selezionati che per il topo: ciononostante, essi costituiscono un materiale per le prove sperimentali di gran lunga più adatto che i ceppi non selezionati.
Si conoscono ora oltre cinquanta tipi di tumori trapiantabili, così che si dispone di tumori praticamente di tutti gli organi compresi quelli del sistema nervoso e di ghiandole endocrine; essi sono disponibili presso il National Institute of Health di Bethesda.
d) La coltura di cellule in vitro
Altro ausilio tecnico di essenziale importanza per la cancerologia, come si è già accennato, è offerto dalla possibilità di coltivare le cellule, malgrado alcune limitazioni dipendenti dalle diversità di condizioni di vita rispetto alle cellule dell'organismo. Lo studio delle colture cellulari presenta inestimabili vantaggi tra i quali importantissimo quello di consentire osservazioni in assenza di fenomeni immunitari, che in vivo interferiscono spesso validamente con l'attecchimento dei trapianti di tumori o di cellule trasformate in vitro.
Le colture in vitro hanno messo in evidenza fenomeni del più alto interesse: uno di questi consiste nel fatto che mentre le cellule normali generalmente si sviluppano in vitro solo per alcune generazioni (a eccezione di alcune linee di recente osservazione), quelle dei tumori si sviluppano invece indefinitamente.
Con adatti accorgimenti tecnici, e in particolare con la frequente sostituzione del liquido di sviluppo, si riesce a mantenere in coltura alcune cellule normali per vari mesi. A tali colture si dà il nome convenzionale di ceppi (cell strains); una buona percentuale di questi, però, dopo un tempo più o meno lungo muore. Talvolta, invece, le colture diventano stabili, acquisiscono cioè la proprietà della moltiplicazione illimitata, e in tal caso a queste cellule si dà il nome di linee cellulari (cell lines). Sono al presente note varie linee cellulari normali e tumorali. Alcune linee derivano da fibroblasti di embrioni di topo di alcuni ceppi inbred, altre da fibroblasti di embrioni di hamster, una da cellule renali di scimmia verde africana. Finora non si è riusciti a ottenere linee cellulari umane, mentre si dispone di linee di tumori umani tra cui famosa, perché largamente impiegata, la linea Hela proveniente da cellule di un tumore uterino. Un'altra linea di tumore umano usata nell'oncologia virale è quella indicata con la sigla Kb, proveniente da un carcinoma del nasofaringe, che permette la crescita di adenovirus isolati dall'uomo.
e) Il fenomeno della trasformazione
Un'altra importante proprietà cellulare messa in evidenza con gli studi delle colture in vitro è quella della perdita dell'inibizione da contatto, per azione sia di carcinogeni chimici sia di virus.
Per l'allestimento delle colture, si opera il distacco delle cellule dal tessuto di provenienza per mezzo della digestione con tripsina delle strutture interstiziali. Le cellule, libere le une dalle altre, si muovono nel liquido di coltura fino a quando non entrano in contatto tra loro; allora cessano i movimenti e si affiancano. Il contatto reciproco delle cellule determina anche la cessazione dell'attività mitotica. In tal modo per la scomparsa della motilità e della capacità di moltiplicarsi si forma un tappeto monostratificato di cellule contigue e la coltura entra in fase stazionaria. A tale fenomeno di cessazione dei movimenti e blocco della moltiplicazione si dà il nome, proposto da M. Abercrombie (v., 1961 e 1962) che scopri il fenomeno, di ‛inibizione da contatto'. Le cellule di tumore, invece di crescere in tappeto, restano libere fra loro nei terreni liquidi, mentre si sovrappongono in più strati formando dei cumuli ben identificabili in colture in agar molle, costituendo quelli che sono anche chiamati foci di trasformazione.
La morfologia delle cellule trasformate è molto diversa da quella delle cellule di colture normali: i fibroblasti normali, ad esempio, hanno forma fusata, ma quando sono trasformati diventano rotondeggianti e molto spesso presentano variazioni del numero dei cromosomi. Altra importante caratteristica è che le cellule trasformate crescono con maggiore velocità, così che la loro densità, cioè il numero di cellule per un dato volume o una data superficie, aumenta nettamente.
Molti ritengono che l'inibizione da contatto sia l'espressione di un meccanismo di regolazione dello sviluppo, e che la perdita ditale capacità potrebbe, almeno in parte, spiegare la crescita illimitata o, come si dice, sfrenata delle cellule neoplastiche: queste, cioè, sarebbero caratterizzate dalla perdita di uno dei controlli della moltiplicazione.
Alcuni fatti indicano che nella trasformazione si verificano alterazioni di una certa entità a livello della membrana cellulare. Si è visto che nelle cellule trasformate da virus di Rous diminuisce notevolmente la concentrazione dell'adenosinmonofosfato ciclico, c-AMP, potente regolatore di molti processi metabolici, mediatore della risposta cellulare a vari ormoni.
Tale diminuzione del c-AMP pare sia di essenziale importanza nel processo di trasformazione, come risulta dall'importante constatazione che la sua aggiunta a cellule infettate con virus di Rous ne blocca la trasformazione in cellule neoplastiche. Ricerche sull'agglutinabilità delle cellule condotte con l'impiego di una sostanza di natura proteica, la concanavalina A, e di due glicoproteine, tutte di origine vegetale e in grado di legarsi a tre differenti carboidrati, hanno consentito di dimostrare anzitutto che nel processo di trasformazione le alterazioni della membrana cellulare svolgono un ruolo di primaria importanza; inoltre, che sulla superficie di fibroblasti trasformati in vitro esistono almeno tre siti differenti, corrispondenti ai tre carboidrati legabili dalle tre diverse agglutinine, ai quali per analogia con altre situazioni biologiche si dà il nome di recettori. Non è escluso che oltre ai tre recettori individuati, ne esistano altri con diversi carboidrati. Ciò dimostra l'esistenza di una sostanziale differenza tra superfici, cioè membrane delle cellule agglutinabili, in particolare di quelle trasformate, e membrane delle cellule normali. Tale differenza consisterebbe nella posizione dei recettori, che nella cellula normale sarebbero situati più all'interno della membrana, quindi non esposti e non combinabili con agglutinine, mentre nella cellula trasformata sarebbero più all'esterno, quindi esposti.
Questa spiegazione è convalidata dal fatto che cellule normali sottoposte a digestione con tripsina divengono agglutinabili sia dalla concanavalina sia dall'agglutinina da germi di grano. Si ha ragione di credere che i recettori per l'agglutinina da semi di soia sarebbero situati più profondamente degli altri nella membrana cellulare, in quanto solo alcune cellule trasformate sono da essa agglutinabili, e per rendere agglutinabili quelle normali il trattamento con tripsina deve essere protratto.
Il meccanismo con il quale le cellule neoplastiche divengono agglutinabili dalle tre sostanze non è chiaro, ma il fenomeno verosimilmente indica che nel passaggio da cellula normale a trasformata si ha perdita di qualche costituente chimico del tutto periferico nella cellula.
Si è anche prospettata l'idea che nelle cellule trasformate i recettori che legano le agglutinine siano più ad- densati in alcuni tratti della membrana, quindi più facilmente combinabili con le agglutinine vegetali (fenomeno del caping).
Connessi probabilmente con questo sono altri due fenomeni indicati come adhesivity e stickiness: parole che, se pure traducibili in italiano ambedue con ‛adesività', esprimono tuttavia due concetti differenti e pertanto vengono comunemente impiegate in inglese. Secondo D. R. Coman (v., 1961), il primo di questi due termini indica la forza che si oppone alla separazione meccanica di due cellule unite tra loro, il secondo la tendenza di una cellula ad aderire a un substrato, ad esempio alla superficie di un vetro. Secondo lo stesso Coman, mentre le cellule normali sono dotate di grande adhesivity ma di scarsa stickiness, quelle neoplastiche mostrano caratteri opposti. In altri termini, le cellule normali tendono ad aderire fortemente tra loro, quelle neoplastiche molto meno ; ma mentre le normali si attaccano debolmente alle superfici, quelle neoplastiche, invece, vi si fissano più fortemente. Probabilmente questi due fenomeni sono solo aspetti particolari dell'inibizione da contatto, cioè delle alterazioni alle quali lo strato periferico della cellula va incontro nel passaggio a cellula trasformata, per cui le cellule normali aderiscono tra loro e formano tappeto, mentre le neoplastiche non aderiscono tra loro e restano libere o si pluristratificano. Sarebbe interessante conoscere quale ruolo i vari siti sensibili alle diverse agglutinine svolgano nei due fenomeni di adhesivity e di stickiness.
La diminuzione della adhesivity viene ritenuta da Kojima e Sakai (v., 1964) fattore di rilievo per la formazione delle metastasi, perché, essendo deboli le forze intercellulari, alcune cellule facilmente si staccano dalle altre, passano nel sangue o nella linfa e possono colonizzare a distanza.
2. Cancerogenesi chimica
Frequentemente in medicina l'osservazione clinica ha determinato l'inizio di ricerche biologiche che, a loro volta, hanno giovato alla soluzione del problema pratico; così pure in cancerologia partendo da osservazioni cliniche si è giunti allo studio e alle scoperte della cancerogenesi chimica. Tali ricerche hanno portato all'identificazione di composti ad attività carcinogenetica e, conseguentemente, hanno prospettato il problema delle modalità secondo le quali queste sostanze determinano la trasformazione di una cellula normale in cellula neoplastica.
Nel 1775-1778 P. Pott, famoso medico inglese noto anche per la prima descrizione della tubercolosi dei corpi vertebrali, richiamò l'attenzione sulla frequenza di tumori dello scroto negli spazzacamini (v. Pott, 1778).
Un secolo più tardi, R. V. Volkmann (v., 1875) descrisse un secondo tipo di cancro professionale, cioè l'epitelioma cutaneo nei lavoratori del catrame o della paraffina. Tuttavia, i primi tentativi di riprodurre sperimentalmente tumori in animali da laboratorio mediante spennellatura della cute con catrame di carbone non furono coronati da successo: infatti A. N. Hanau (v., 1889) non ottenne alcun risultato, né vero risultato positivo ottenne H. Bayon (v., 1912).
Il problema fu quindi abbandonato per alcuni anni, per quanto fosse sempre evidente uno stretto rapporto tra catrame e cancro, finché Yamagiwa e Ichikawa (v., 1914), dopo aver spennellato pazientemente per mesi con catrame la cute del padiglione di orecchi di conigli, ottennero lo sviluppo di veri cancri cutanei a larga capacità infiltrativa nel derma. Fu così dimostrato sperimentalmente quanto l'osservazione clinica aveva notato, cioè che il cancro dei lavoratori del catrame è dovuto all'azione di tale sostanza, ed ebbe quindi inizio la cosiddetta era della cancerogenesi chimica.
Ma l'interesse per i tumori da catrame si destò in Europa solo alcuni anni dopo la prima guerra mondiale, e dopo che la dimostrazione fornita da H. Tsutsui (v., 1918) della possibilità di provocare sperimentalmente cancri cutanei da catrame sulla cute di topi stimolò e facilitò lo sviluppo delle ricerche in proposito. Infatti, qualche anno dopo, R. D. Passey (v., 1922) con l'applicazione di estratti eterei di fuliggine sulla cute dei topi ottenne lo sviluppo di cancri cutanei, riproducendo così sperimentalmente il cancro dello scroto degli spazzacamini, secondo il citato nesso di dipendenza prospettato dal Pott.
Il catrame è una complessa miscela di sostanze organiche e inorganiche: si imponeva, quindi, la necessità di isolare da esso la sostanza o le sostanze dotate di potere oncogeno. Eliminata la possibilità che questo fosse dovuto a costituenti inorganici del catrame e in particolare all'arsenico, nel quale vari ricercatori avevano inizialmente creduto di individuare l'elemento responsabile della cancerizzazione per la diffusa opinione circa il presunto potere oncogeno di tale elemento (cancro da arsenico), fu presto accertato che l'attività oncogena è riferibile a idrocarburi aromatici e precisamente a quelli che distillano tra 300 e 400 °C: dunque, idrocarburi a elevato peso molecolare. Un importante progresso fu realizzato da E. L. Kennaway (v., 1924 e 1925), che ottenne composti cancerogeni molto potenti trattando l'acetilene e l'isoprene in atmosfera di idrogeno ad alta temperatura (idrocarburi artificiali). Il riconoscimento degli idrocarburi nei distillati, e nei tessuti con essi trattati, fu inoltre facilitato dalla scoperta che le sostanze biologicamente attive sono contenute nelle frazioni del distillato il cui spettro di assorbimento presenta tre massimi, cioè a 400, 418 e 440 mm (v. Hieger, 1930).
Carcinogeni furono anche ottenuti da sostanze presenti nell'organismo, come il colesterolo, nonché da estratti di pelle, di capelli ecc. (v. Hieger, 1946, 1947, 1949 e 1959). Anche da oli minerali furono ottenuti carcinogeni (v. Leitch, 1922).
a) Idrocarburi
L'1-2-benzantracene fu il primo composto puro, individuato nel 1933 da I. Hieger (v.), il cui spettro di fluorescenza si avvicinava a quello della frazione attiva, anche se non si identificava con esso. Peraltro né tale composto né l'antracene da cui esso deriva avevano attività cancerogena.
Tuttavia l'identificazione dell'1,2-benzantracene servì da guida per scoprire altri tipi molecolari a esso analoghi e rese così possibile trovare il primo potente carcinogeno, cioè l'1,2:5,6-dibenzantracene (v. Kennaway e Hieger, 1930).
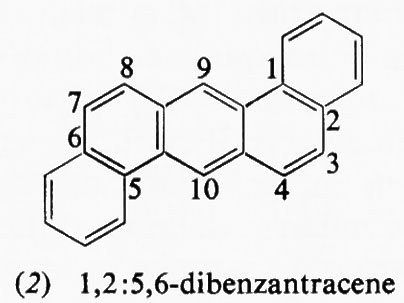
Da allora sono stati isolati oltre trecento idrocarburi cancerogeni a potenza variabile. Alcuni, che determinano l'insorgenza di tumore solo in qualche specie animale e solo in qualcuno degli animali trattati, in genere dopo un lungo periodo di latenza, vengono indicati come carcinogeni deboli o debolissimi. Al contrario, si conoscono idrocarburi che in varie specie animali (topo, ratto, coniglio, criceto, piccione ecc.) e in vari tessuti inducono la formazione di tumori che ripetono la struttura dei tessuti dai quali originano (epiteliomi negli epiteli, sarcomi nel connettivo sottocutaneo, rabdomiomi nel tessuto muscolare, osteosarcomi nelle ossa ecc.). Non è accertata la possibilità della produzione di tumori del tubo digerente per mezzo di idrocarburi anche molto potenti, come dubbia è quella di indurre con tali sostanze leucemie e tumori della mammella.
Di due altri idrocarburi aromatici occorre far cenno, per il particolare significato che hanno avuto in questi studi (v. Hieger, 1961). Uno è il 3,4-benzopirene

la cui importanza nella storia della cancerologia deriva dal fatto che il suo spettro di fluorescenza è il più intenso tra tutti, onde è servito da guida per il riconoscimento e per l'isolamento delle molecole biologicamente più attive in una miscela di idrocarburi.
L'altro è il 3-metilcolantrene, uno tra i più potenti carcinogeni che si conoscano.
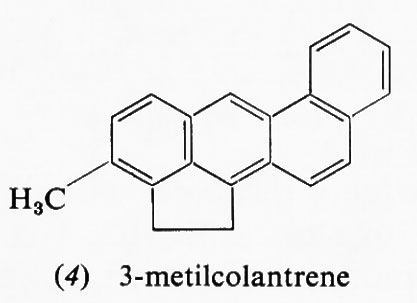
È interessante il fatto che il metilcolantrene fu preparato per la prima volta da H. Wieland ed E. Dane (v., 1933) dall'acido desossicolico e poi da W. Rossner (v., 1937) dal colesterolo, cioè da due componenti naturali degli organismi. Con ciò si poneva il quesito, peraltro non ancora risolto, se sia possibile che nell'organismo dall'acido desossicolico e in particolare dal colesterolo abbiano origine molecole tipo metilcolantrene.
Tra gli idrocarburi debolissimi e quelli ad altissima attività si inseriscono, in graduale successione, gli altri. Nella sperimentazione si impiegano in genere o quelli debolissimi o, all'opposto, quelli altamente attivi. I primi vengono specialmente usati nelle ricerche con i cosiddetti fattori cocancerogeni, cioè fattori di per sé innocui o a debolissima attività oncogena, ma capaci di potenziare fortemente l'attività dei carcinogeni deboli.
L'alchilazione in determinati punti dello scheletro di alcuni idrocarburi aromatici induce la comparsa di attività cancerogena o il suo potenziamento. L'alchile più attivo è il metile, poi l'influenza decresce con l'aumentare dei carboni fino ad annullarsi in corrispondenza dei derivati butilici. L'introduzione di un secondo metile incrementa l'attività cancerogena, ma solo per alcuni idrocarburi, e forse anche per un terzo metile si determina un lieve rafforzamento rispetto al dimetilderivato; ulteriori metilazioni non fanno aumentare l'attività. Un fatto importante, risultato dalle numerose ricerche eseguite sullo scheletro fondamentale dell'1,2-benzantracene, è che esistono in esso punti di massima sensibilità all'alchilazione situati a livello dei carboni in posizione 5, 9 e 10; molto meno sensibili sono quelli in posizione 6, 7, 8, 3, 4, e del tutto insensibili quelli in posizione 1′, 2′, 3′ e 4′. Il punto di massima sensibilità del crisene è, invece, il carbonio in posizione 2, un poco meno sensibili sono quelli in 1 e in 6.
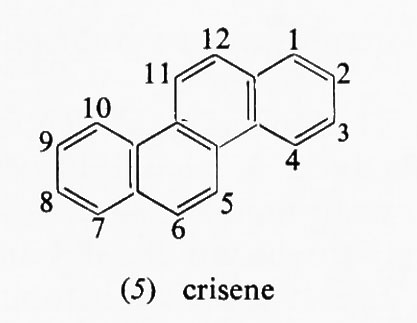
Si può anche, per metilazione in alcuni punti, far perdere all'idrocarburo ogni attività cancerogena: tale è il caso del 3,4-benzopirene, molecola che per metilazione dei carboni 2′ e 3′ diviene inattiva. L'introduzione nelle molecole di idrocarburi di altri gruppi altamente reattivi come OH, COOH, CH2Cl ecc. porta alla loro inattivazione come cancerogeni; ma anche questa non è regola generale, perché in alcuni, come il 10-metil-1,2-benzantracene, alcune sostituzioni non alterano praticamente il potere oncogeno.
L'idrogenazione inattiva i carcinogeni; l'introduzione di un idrogeno in uno dei doppi legami di uno degli anelli aromatici induce abbassamento del potere oncogeno, che va completamente perduto se l'idrogenazione della molecola è completa.
L'attività cancerogena degli idrocarburi aromatici è massima nei composti tetra- e pentaciclici nei quali si sia determinata o la metilazione in alcuni punti di sensibilità o la coniugazione con un altro anello benzenico in posizione laterale (anellazione), con l'effetto che H. Druckrey (v., 1950) chiamò auxocancerogeno. Secondo lo stesso ricercatore, gli idrocarburi a tre o anche a quattro anelli benzenici, a combinazione laterale, non sono cancerogeni ma hanno in sé la possibilità di diventarlo: sono cioè, come egli si esprime, cancerofori. Tutte quelle azioni che deformano la molecola o agiscono sui doppi legami degli anelli determinano attenuazione o perdita dell'attività cancerogena (v. Butenandt e Dannenberg, 1956); uguale effetto causa la sostituzione con gruppi atomici molto reattivi o che hanno proprietà acide.
Nel 1940 O. Schmidt (v.) richiamò l'attenzione sulla relazione esistente tra potere carcinogenetico degli idrocarburi e densità di elettroni mobili - gli elettroni π - in alcuni punti della molecola, cioè nella cosiddetta regione K e nella regione L. La regione L è chimicamente più reattiva, perché più facilmente vi avvengono sostituzioni, e la regione K è quella dei doppi legami più altamente reattivi degli idrocarburi tipo fenantrene. La densità elettronica della regione K deve, secondo A. e B. Pullman (v., 1954) e altri studiosi di chimica-fisica, superare un certo valore perché la molecola dell'idrocarburo sia cancerogena.
Un altro cancro professionale - quello della vescica dei lavoratori di anilina e derivati, messo in evidenza da osservazioni cliniche di L. Rehn (v., 1895) - ha da tempo posto il problema dell'importanza dell'anilina come cancerogeno. La semplice molecola dell'anilina non ha tale potere, che è invece posseduto dalla β-naftilammina, dalla β-antrammina e dal 2-amminofluorene.

Quest'ultimo composto, specialmente attivo come acetilderivato (9), è di notevole importanza: esso induce in varie specie animali tumori di organi vari a struttura istologica diversa. Molto probabilmente è carcinogeno non il 2-acetilamminofluorene, bensì una sostanza originata dalla sua degradazione metabolica: F. Bielschowsky e W. H. Hall (v., 1951) dimostrarono infatti che ponendo due ratti in parabiosi il tumore si sviluppa solo nell'animale cui è stato somministrato con l'alimento il composto, i cui prodotti di degradazione vengono rapidamente fissati nei tessuti, senza diffondere nell'animale a lui unito.
Anche per quanto riguarda la β-naftilammina è verosimile che il corpo attivo sia un prodotto del suo metabolismo, probabilmente identificabile nel 2-ammino-1-naftolo che viene eliminato con le urine e pertanto permane in vescica: l'applicazione di tale sostanza sulla mucosa vescicale induce infatti formazione di tumore. Il 2-amminofluorene e la β-naftilammina si citano come esempi della cosiddetta cancerizzazione a distanza, espressione con la quale si suole indicare lo sviluppo di cancro in sede diversa da quella dell'applicazione del carcinogeno. Ciò significa che solo dal metabolismo del composto ha luogo la formazione della molecola attiva che si fissa e causa sviluppo di tumore: questo può avvenire nell'organo stesso della captazione e della successiva degradazione metabolica; oppure il composto viene fissato in un organo, ad esempio nel fegato, ove si svolge la sua degradazione metabolica seguita da rapida eliminazione dei metaboliti e quindi dall'induzione del tumore negli organi di eliminazione (vescica, vie biliari ecc.).
Anche per il 2-acetilamminofluorene molte ricerche riguardano il raggruppamento atomico attivo. La molecola è abbastanza resistente a trattamenti chimici, onde sostituzioni varie non ne alterano essenzialmente l'attività; questa è ridotta nell'N-dimetilamminofluorene e rafforzata, invece, per introduzione di un secondo gruppo acetilamminico o di un atomo di fluoro nel carbonio in posizione 7.
b) Azocomposti
Altra serie di composti a cancerizzazione a distanza è quella degli azocomposti. Le indagini in proposito traggono origine dal cosiddetto fenomeno di Fischer: questi nel 1906 osservò che l'introduzione di rosso scarlatto o di Sudan III nel sottocutaneo del padiglione dell'orecchio dei conigli determinava l'insorgenza di proliferazioni epiteliali tendenti anche all'invasione e alla corneificazione, con caratteri cioè di tipo epiteliomatoso. A tale fenomeno, in generale, non fu data importanza e lo si ritenne espressione eccito-proliferativa senza rapporto con epiteliomi, anche perché le proliferazioni facilmente regredivano. In seguito T. Sasaki e T. Yoshida (v., 1935) scoprirono che l'o-amminoazotoluene, che precedenti ricerche di E. Hayward (v., 1909) avevano dimostrato essere la sostanza attiva del rosso scarlatto producente il fenomeno di Fischer, determinava nei ratti ai quali era stato somministrato con gli alimenti lo sviluppo di tumori epatici. Questa scoperta aprì un vasto campo di studi, tuttora in pieno sviluppo, e alimentò la speranza di scoprire la sostanza o le sostanze in grado di causare tumori del fegato nell'uomo.
Un fatto molto importante, messo in luce dalle ricerche di H. Druckrey e K. Küpfmüller (v., 1948), è che l'attività cancerogena risulta, in una certa maniera, dipendente non dal tempo nel quale viene somministrato un composto, ma dalla soglia critica della sua somministrazione, che può essere raggiunta in tempi variabili. Tra le altre dimostrazioni appare particolarmente valida quella della Stop- Versuche (ricerca con stop): se un ratto è nutrito per un certo tempo con dosi basse di cancerogeno del quale si sospende poi la somministrazione, il tumore non si sviluppa; se però dopo un certo tempo si riprende a somministrare il composto con la dieta, l'epatoma compare quando è stata raggiunta la quantità corrispondente a somministrazioni giornaliere continuative. Il fenomeno è analogo a quello che si riscontra nel caso delle radiazioni, per le quali ha importanza la sommazione delle dosi, anche se tale sommazione avviene in periodi molto lunghi.
La dose critica varia molto in rapporto al ceppo di ratti e alle condizioni sperimentali, oscillando da 350 a 1.200 mg. Per uno stesso ceppo genetico, in condizioni determinate, particolarmente per quanto riguarda l'alimentazione, tale dose critica è abbastanza costante. Sembra inoltre che l'arricchimento della dieta con vitamina B2 ritardi notevolmente lo sviluppo del tumore.
La molecola madre è costituita dall'azobenzene
che non è cancerogeno. Il suo derivato dimetilato, 2,3′-azotoluene, possiede una certa attività. I composti più attivi originano dal 4-amminoazobenzene, molecola ancora a debole attività carcinogena.


Tale attività diviene elevata nel 4-dimetilamminoazobenzene e specialmente nel 4-metiletilamminoazobenzene. I composti (12) e (13) vengono trasformati facilmente l'uno nell'altro nei processi metabolici cellulari.
Il 4-dimetilamminoazobenzene era un tempo usato nelle industrie dei grassi come colorante del burro (giallo burro), ma in seguito alla scoperta della sua attività cancerogena ne è stato, naturalmente, vietato l'impiego.
La sostituzione del ponte di azoto con un gruppo etilenico porta alla produzione di stilbeni, alcuni dei quali, come il 4-dimetilamminostilbene, sono cancerogeni;

al contrario, il 4-amminostilbene inibisce la crescita tumorale.
Già nel 1937 A. Haddow e A. M. Robinson (v.) avevano visto che l'inoculazione di alcuni idrocarburi inibiva la crescita di tumori negli animali; successivamente, nel 1947, Haddow e Kon (v.) trovarono che il 4-amminostilbene era molto più attivo di essi come antineoplastico.
La constatazione che alcuni composti di una stessa serie agiscono da deboli e alcuni da potenti carcinogeni, che altri risultano del tutto indifferenti e altri ancora esercitano addirittura attività biologica opposta, ha posto già da tempo numerosi e complessi problemi teorici e sperimentali.
c) Uretano
Un carcinogeno chimico di particolare interesse è l'uretano (etilcarbammato).
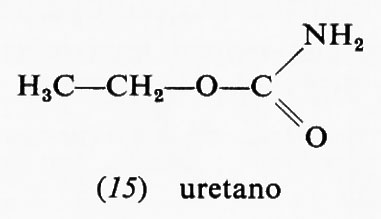
La scoperta dell'attività carcinogenetica di tale sostanza avvenne, come talora accade, fortuitamente. Nel 1943 A. Nettleship e altri (v.) dovendo realizzare, per le loro ricerche sugli effetti delle radiazioni nel topo, un tipo di anestesia di più lunga durata di quella che si otteneva con il pentobarbitale, impiegarono una preparazione a base di etiluretano, usata in veterinaria: notarono allora il sorprendente fatto che 26 su 29 giovani femmine di topo presentarono, dopo un certo tempo, tumori del polmone causati, come fu poi possibile accertare con i controlli, non dall'irradiazione, ma dal narcotico.
Tale scoperta diede origine a numerose ricerche sull'azione dell'uretano a livello di varie funzioni cellulari e fu possibile mettere in evidenza come gli effetti maggiori del composto si esplicassero sul nucleo, e in particolare sul DNA, con conseguenti alterazioni dei cromosomi.
L'uretano è cancerogeno, oltre che per il topo, anche per il ratto; per altre specie, invece, come cavia e pollo, è del tutto inattivo. Esso risulta cancerogeno anche per applicazione locale, cioè per pennellazioni della cute, e tale effetto è potenziabile con cocarcinogeni come l'olio di croton. L'uretano è un carcinogeno multipotenziale perché, somministrato per via alimentare con l'acqua da bere a topi neonati, causa non solo tumori del polmone, ma anche della mammella e del fegato, e inoltre emangiomi e linfomi del timo. Ciò dipende dal fatto che il composto, quale che sia la via d'introduzione, si ripartisce, press'a poco alla stessa concentrazione, tra tutti i tessuti dai quali, però, scompare in poche ore. Non si conoscono ancora esattamente le varie tappe metaboliche dell'uretano: si sa comunque che esso è metabolizzato dal fegato ed escreto come urea e che il 90% del carbonio del gruppo carbossilico è eliminato sotto forma di CO2. Tale ultimo punto, dimostrato dal reperto nell'aria espirata della massima parte della radioattività del carbonio marcato del residuo carbammico, potrebbe spiegare l'elevata incidenza di tumori polmonari in animali sottoposti all'azione oncogena dell'uretano. Desta tuttora sorpresa il fatto che una sostanza a modesto peso molecolare, quale è l'uretano, solubile in acqua e rapidamente eliminabile, eserciti notevole potere oncogeno a sede multipla. Occorre ricordare che O. Warburg (v., 1921) ritenne che l'attività cancerogena dell'uretano fosse ascrivibile al suo potere narcotico, in definitiva, quindi, a limitazione della respirazione cellulare: egli tentò di spiegare la narcosi da uretano come dovuta a competizione, sulla superficie cellulare, tra molecole del narcotico e molecole di sostanze nutritive, quali glucosio e amminoacidi, e interpretò tale fenomeno come conseguenza di una inibizione delle ossidazioni cellulari, dimostrando che l'azione narcotica aumenta con l'allungarsi della catena carboniosa negli omologhi superiori dell'etiluretano. Tuttavia tali composti non pare che esercitino attività oncogena, come invece ci si sarebbe aspettato qualora l'attività oncogena fosse stata realmente correlata con quella narcotica.
L'uretano, oltre a possedere attività cancerogena, è anche in grado di esercitare potere antiproliferativo, come fu messo in evidenza da A. Haddow e W. A. Sexton (v., 1946) prima su tumori solidi e poi sulle leucemie, nelle quali l'azione è particolarmente rilevante: per un certo tempo, quindi, si diffuse l'impiego del composto come farmaco antineoplastico specialmente in alcune forme di leucemia e nel mieloma, e ancora oggi alcuni ricercatori paragonano i risultati che si conseguono con antileucemici vari a quelli che, per controllo e riferimento, si ottengono con l'uretano. Si include comunemente il farmaco nella vasta serie dei veleni antimitotici: la sua somministrazione induce infatti caratteristiche alterazioni nucleari (v. chemioterapia antineoplastica).
Anche per alcuni composti della classe degli idrocarburi cancerogeni era da tempo conosciuta l'azione antineoplastica: si tratta, tuttavia, di molecole vicine strutturalmente, ma diverse in uno o più raggruppamenti atomici rispetto a quelle dotate di potere oncogeno. Per quanto riguarda l'uretano, invece, sembrerebbe che la stessa semplice molecola possegga entrambe le capacità, cancerogena e anticancerogena, e il fenomeno viene da alcuni studiosi messo in relazione a momenti di sensibilità diversa delle fasi del ciclo cellulare.
Sembra però più probabile che in conseguenza delle attività metaboliche specifiche dei diversi tipi cellulari si formino dalla stessa molecola prodotti diversi, taluni capaci di esercitare potere oncogeno nei riguardi di alcuni tipi di cellule, altri invece dotati di potere antiproliferativo e antineoplastico. Si potrebbe cosi ammettere che a contatto con le cellule neoplastiche, caratterizzate da un particolare metabolismo parzialmente anaerobico, l'uretano dia luogo alla formazione di molecole ad attività antiproliferativa, contrariamente a quanto si verifica nel caso delle cellule normali, nei cui confronti il composto agisce da carcinogeno.
d) Mostarde, etilenimmine, epossidi
Per altri gruppi di sostanze, come le mostarde, le etilenimmine e gli epossidi, si osservano fatti analoghi: attività carcinogenetica esplicata da alcuni tipi molecolari, attività anticarcinogenetica svolta da altri a questi correlati. C'è troppo riscontro, anche per composti a struttura molecolare estremamente diversa, tra le due azioni biologiche, quella favorente e quella inibente la crescita neoplastica, per poter sospettare che si tratti di evenienze casuali disgiunte. Più logico appare, pertanto, ammettere l'esistenza di uno stretto nesso, e sulla guida della struttura chimica delle varie sostanze cercare il punto bersaglio dell'una o dell'altra azione.
Le mostarde (iprite e derivati) erano da tempo note come sostanze ad azione analoga o almeno confrontabile con quella delle radiazioni. Il corpo base, noto come gas mostarda o iprite, è il bis(2-cloroetil)solfuro.
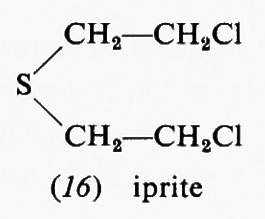
Questa sostanza, somministrata per inalazione, provoca nei topi la comparsa di tumori polmonari e introdotta per via sottocutanea determina in topi e ratti sviluppo di sarcomi (v. Heston, 1950). Molto più attive risultano le mostarde azotate, nelle quali lo zolfo è sostituito da un azoto mono- o dimetilato, come la metilbis(2-cloroetil)ammina e la tris(2-cloroetil)ammina.
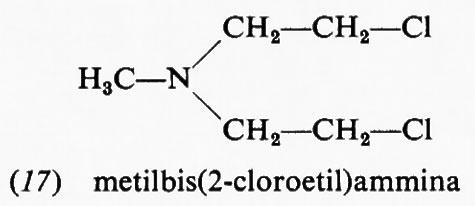

L'attività di questi composti è in rapporto con il loro potere fortemente alchilante, che si esplica su vari aggruppamenti atomici e in primo luogo sui gruppi amminici, dunque su amminoacidi e ammine biogene; ma essi sono in grado di alchilare anche gruppi carbossilici e gruppi fosforati, così che la loro azione risulta particolarmente intensa sulle proteine e sugli acidi nucleici. In tal modo, secondo molti autori, si spiegano le alterazioni cromosomiche che si riscontrano a seguito di applicazioni di mostarde azotate in cellule normali o in cellule neoplastiche. L'azione biologica di tali composti è molto complessa e vari sono i sistemi enzimatici a essi sensibili. Le mostarde azotate sono potenti mutageni sia per organismi relativamente semplici come Batteri e Funghi (Neurospora), sia per Drosophila, e fu proprio la scoperta della loro attività mutagena che destò l'interesse per quella carcinogenica.
Le mostarde azotate sono altresì in grado di esercitare una notevole azione citostatica e inibente lo sviluppo di alcune forme di tumori, come linfosarcomi e linfogranulomi (morbo di Hodgkin), e per vario tempo furono impiegate per la terapia di tali affezioni.
Da potenti alchilanti agiscono tutte le etilenimmine, e in particolare le N-aciletilenimmine,
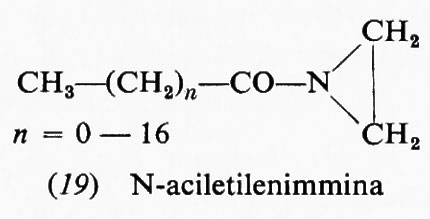
che, inoculate sotto cute, determinano nei ratti formazione di sarcomi.
L'etilenimmina svolge anche attività mutagena e alcuni suoi derivati (metilolammide) sono dotati di potere antiproliferativo. La funzione cancerogena è dipendente dalla struttura molecolare; si sa a tale proposito che le etilenimmine monofunzionali (acil-etilenimmine), cioè con un solo gruppo
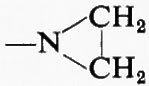
sono carcinogene, mentre quelle bifunzionali, cioè con due gruppi
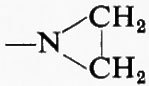
sono sempre prive di potere oncogeno e agiscono solo da inibenti lo sviluppo neoplastico.
Azione alchilante esercitano gli epossidi, composti nei quali due atomi di carbonio sono uniti da un ponte di ossigeno:
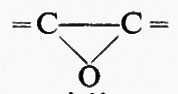
Gli epossidi come tali sono inattivi, ma alcuni loro derivati esercitano notevole attività oncogena, che è massima nel 4-vinil-cicloesano-1,2:7,8-diepossido; altri derivati sono invece caratterizzati da proprietà antiproliferativa.
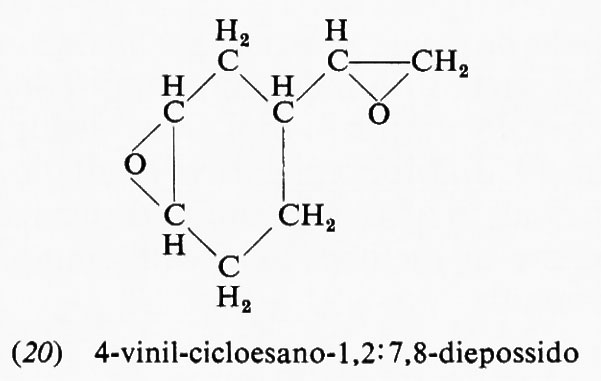
e) Aflatossina e cicasina
Recentemente due potenti carcinogeni sono stati scoperti in sostanze alimentari: l'aflatossina, dimostratasi poi sostanza non unitaria, e la cicasina.
La storia dell'aflatossina è interessante per la singolare coincidenza delle varie osservazioni. Nel 1961 si osservò in Inghilterra una larghissima mortalità, dovuta principalmente a gravi lesioni epatiche, negli allevamenti di tacchini, anatre e polli alimentati con farina di arachide.
Nello stesso anno E. M. Wood e C. P. Larson (v., 1961) richiamarono l'attenzione sul fatto che nel Nord-Ovest degli Stati Uniti si osservavano, con una certa frequenza, trote con epatomi, e il fenomeno fu messo in relazione con la contaminazione di Aspergillus flavus della farina di arachide con la quale gli animali venivano nutriti. Dalle colture di tale fungo furono inizialmente isolate quattro afiatossine che, inoculate in trote normali, produssero epatomi ed epatocarcinomi identici a quelli naturali. Le originali afiatossine furono indicate con le sigle B1, B2, G1 e G2. In seguito sono state identificate altre due tossine, indicate come B2a e G2a. Dal latte di ratti e di mucche nutrite con farina di arachide contenente afiatossine sono stati isolati due prodotti, M1 e M2, che pare siano prodotti idrossilati di B1 e B2.
Contemporaneamente Lancaster e altri (v., 1961) dimostrarono l'alta incidenza di epatomi in ratti nutriti con farina di arachide contenente aflatossine. Finora è stata dimostrata l'attività carcinogenetica delle aflatossine in ratti, anatre e trote, mentre topi di varie linee genetiche non sono sensibili a queste sostanze.
Per quanto riguarda l'uomo, è stato dimostrato che alcuni alimenti, particolarmente in Asia e in Africa, contengono aflatossine in quantità variabili. È stata prospettata, perciò, l'ipotesi che la frequenza di epatomi in alcuni paesi, come l'Uganda, si possa attribuire alle afiatossine prodotte dai microrganismi del genere Aspergillus flavus contaminanti gli alimenti.
Inquietante è anche il problema della contaminazione del riso con muffe. In alcuni campioni di riso di origine giapponese sono state isolate circa duecento specie di muffe tra le quali cinquanta di Penicillium. Nell'avaria del riso conosciuta come riso ingiallito è stato isolato il Penicillium islandicum, nelle cui colture sono stati identificati la luteoschirina e un peptide contenente cloro.
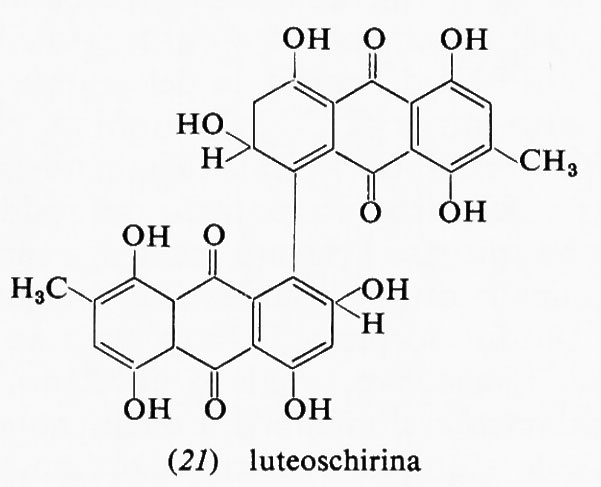
Entrambe le sostanze sono fortemente tossiche per il tessuto epatico, in quanto determinano infiltrazione grassa, necrosi centrolobulare, atrofia e cirrosi.
Era noto sin dall'antichità che la farina di semi di Cycadaceae - piante antichissime e per la maggior parte fossi- li, con pochi generi superstiti in varie parti del mondo - risultava tossica se ingerita cruda, mentre la cottura la rendeva innocua. In Australia il bestiame che si nutriva con foglie, frutti e parti del tronco di tali piante presentava gravi manifestazioni morbose, per cui in alcune zone esse vengono ora estirpate. Si osservò anche che nel Guam, un'isola delle Marianne, si riscontravano numerosi casi di sclerosi laterale amiotrofica (v. Whiting, 1963), da mettere forse in relazione col largo consumo di farina di Cycadaceae.
Nutrendo varie specie animali con tale farina si riscontrò non già insorgenza di fenomeni neurologici, ma, nei ratti, sviluppo di tumori di vari organi, in particolare del fegato e del rene (v. Laquer e altri, 1963), e, nella cavia, di carcinomi del fegato. Fu isolata la sostanza attiva, cui fu dato il nome di cicasina.
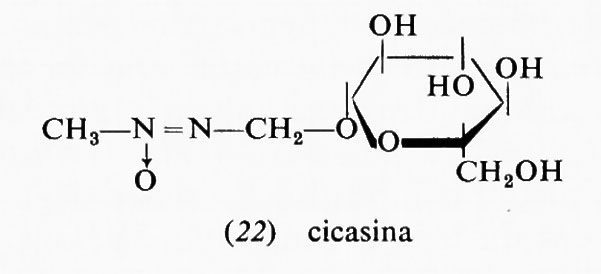
La somministrazione di tale sostanza determina nei ratti un'elevata incidenza di tumori del rene, e un'incidenza inferiore di tumori dell'intestino e del fegato. Fu poi scoperto che la sostanza introdotta per via parenterale determina effetti tossici ma non formazione di tumore, e che viene eliminata come tale; inoltre la cicasina in ratti germ free non esercita azione oncogena, ma solo effetti tossici. Tutto ciò portava a concludere che il carcinogeno viene originato dalla metabolizzazione della cicasina da parte di batteri intestinali.
La sostanza ad azione carcinogenica venne identificata nell'aglicone della cicasina, cioè nel metilazossimetanolo (MAM), che si forma per azione di glucosidasi batteriche (v. Kobayashi e Matsumoto, 1964). La sostanza determina comparsa di tumori in diversi organi, come l'originaria farina di Cycadaceae, e manifesta tale proprietà anche se viene inoculata in ratti germ-free. Il MAM, che nel 1965 fu sintetizzato da H. Matsumoto e altri (v., 1965), è chimicamente simile alla dimetilnitrosammina, per cui J. A. Miller (v., 1964) suppose che i due composti siano convertiti in uno stesso effettivo cancerogeno.
Si conoscono varie altre sostanze di origine vegetale dotate di attività carcinogenetica più o meno spiccata, come la griseofulvina estratta dal fungo Penicillium griseofulvum e il safrolo contenuto nell'olio di sassafrasso (corteccia della radice dell'albero americano Sassafras officinale). Ratti nutriti con segale cornuta, sclerozio del fungo Claviceps purpurea, parassita della spiga di segale, presentano numerosi neurofibromi, che regrediscono se tale alimentazione viene sospesa, ricompaiono se ripristinata.
La tiourea, contenuta nelle piante del genere Brassica, produce adenomi e anche carcinomi della tiroide. Gli alcaloidi della pirrolizidina, ricavabili soprattutto da Senecio, sono sostanze ad azione epatotossica e carcinogenetica per il fegato. Varie altre sostanze di origine vegetale sono state identificate come carcinogene, e la lista evidentemente è destinata ad allungarsi nei prossimi anni. L'interesse che si rivolge ai vegetali quali fonti di carcinogeni deriva, naturalmente, dalla possibilità che alcuni di essi causino tumori epatici nell'uomo: ciò potrà essere assodato soprattutto in base a ricerche epidemiologiche, del tipo di quelle, già citate, compiute sulle aflatossine nelle trote e in alcuni volatili.
f) Agenti chimici endogeni
Per quanto riguarda la cancerogenesi da agenti chimici endogeni il problema è di appurare se nell'organismo si possano formare sostanze carcinogene, nel corso di reazioni metaboliche dipendenti dalla presenza di specifici sistemi enzimatici. Nonostante le innumerevoli ricerche eseguite, iniziate poco dopo la scoperta dei primi idrocarburi cancerogeni e tuttora in corso, non si può dare una risposta certa. Come si è già detto, H. Wieland ed E. Dane (v., 1933), riuscendo a ottenere metilcolantrene dall'acido desossicolico, dimostrarono la possibilità della formazione per via chimica di idrocarburi cancerogeni da steroidi costituenti normali dei tessuti e dei liquidi dell'organismo; tuttavia, la prova che una tale reazione possa realmente svolgersi nell'organismo non fu fornita. Come fanno notare R. Butenandt e H. Dannenberg (v., 1956), mentre gli steroidi sono composti idroaromatici saturi, il metilcolantrene e gli altri cancerogeni di questo tipo sono idrocarburi formati esclusivamente da anelli aromatici: per tale ragione, l'attenzione dei ricercatori è stata rivolta in particolare alle molecole ditale tipo strutturale, come gli ormoni femminili estrone, equilina ed equilenina, per appurare se dalla loro deidrogenazione possano originare idrocarburi cancerogeni. Non si è riusciti a dimostrare tale evenienza, anche perché non si sono trovati nei tessuti enzimi catalizzanti tali reazioni, e quindi l'ipotesi che da ormoni naturali possano derivare carcinogeni non è stata comprovata. Né è stata dimostrata la possibilità che germi della normale flora intestinale siano capaci di trasformare steroidi normali in carcinogeni.
Di un certo interesse è la possibilità, dimostrata per primo da L. Shabad (v., 1937), di provocare sviluppo di neoplasie in animali da esperimento mediante inoculazione di estratti di tessuto. L'estratto di Shabad proveniva dal fegato di un canceroso, ma in seguito fu osservato da altri, come ad esempio da P. E. Steiner (v., 1942 e 1943), che anche gli estratti di fegato di individui normali sono in grado di svolgere identica azione. Questa non è attribuibile a idrocarburi cancerogeni originati nei processi metabolici che si svolgono nel fegato, perché risultati simili si ottengono perfino con estratti di fegato di bambini nati morti e di fegato di maiale.
Il riconoscimento che la sostanza attiva era contenuta nella frazione insaponificabile degli estratti lipidici indusse I. Hieger (v., 1946 e 1947) a ritenere che si trattasse o di colesterolo o di suoi derivati e non di idrocarburi aromatici, anche perché nel corso di queste ricerche non era stato possibile mettere in evidenza sostanze i cui spettri di assorbimento corrispondessero a quelli caratteristici degli idrocarburi aromatici.
Lo stesso Hieger dimostrò come il colesterolo possa determinare sviluppo di tumori: egli infatti iniettando tale sostanza, anche altamente purificata, nel tessuto sottocutaneo di topi ottenne insorgenza di sarcomi in 70 su 1.424 animali (v. Hieger, 1959).
Risultati talvolta simili, talvolta opposti, furono ottenuti in vari altri laboratori: per motivi ancora non spiegabili, secondo i dati riferiti dai vari autori è possibile osservare l'insorgenza di tumori entro limiti notevolmente ampi, dallo 0 al 15%, e addirittura per uno stesso ceppo inbred di topi, come ad esempio il C57, dallo 0 all'11%. Il problema, perciò, permane attuale, soprattutto in considerazione del ben noto reperto anatomopatologico dell'accumulo di colesterolo nel connettivo circostante alcuni tumori, in particolare quelli della mammella. Di recente è stata presa in esame la possibilità che il colesterolo agisca non come carcinogeno chimico ma come corpo allo stato solido, in rapporto alle facce dei suoi cristalli (v. sotto, cap. 3, È c).
Alcuni dei derivati del colesterolo, quali il 6-idrossi-Δ4- colestene-3-one, il Δ4-colestene-3,6-dione, l'ossido di colesterile e il 6-idroperossido-Δ4-colestene-3-one, svolgono azione oncogena (v. Fieser e altri, 1955; v. Bischoff e altri, 1955; v. Bischoff, 1957). Hieger, invece, è riuscito a ottenere sviluppo di tumori solo con l'ultimo di tali composti. È interessante notare che le percentuali di positività circa l'attività oncogena sono notevolmente elevate, in particolare proprio per l'ultima sostanza, con la quale si registra fino al 60% di risultati positivi. Anche nel caso di questi derivati del colesterolo le conoscenze sul meccanismo d'azione sono incomplete, e si impongono quindi ulteriori ricerche.
g) Etionina
Una sostanza strettamente vicina a un costituente normale delle cellule è l'etionina, un omologo dell'amminoacido naturale metionina.

L'etionina, considerata per molto tempo un amminoacido artificiale, cioè non esistente in natura, fu identificata nel 1957 da F. Schlenk (v., 1957) tra i costituenti di un lievito e successivamente, nel 1961, venne individuata da I. F. Fisher e M. F. Mallette in Escherichia coli e in altri batteri. A tale ultimo reperto potrebbe essere attribuito un particolare significato in considerazione della ricchezza in Escherichia coli della flora intestinale: infatti nella stasi intestinale, o per sopravvento di altri germi, Escherichia coli va incontro a lisi e libera etionina, il cui assorbimento potrebbe determinare effetti tossici.
L'etionina certamente non fa parte degli amminoacidi che entrano nella costituzione delle proteine di organismi pluricellulari; è anzi un antimetabolita di un amminoacido per questi indispensabile, la metionina. Somministrata ad animali provoca gravi lesioni soprattutto a livello epatico, consistenti in infiltrazione grassa e necrosi degli acini, e inoltre necrosi dei tubuli renali, emorragie dei surreni, alterazioni metaboliche del miocardio e delle cellule della mucosa intestinale, ecc. Si può quindi affermare che esiste una patologia sperimentale da etionina, che può essere limitata o annullata dalla somministrazione di metionina: tra i due amminoacidi, infatti, si determinano, anche nei tessuti neoplastici, fenomeni di scambio competitivo.
L'azione cancerogena dell'etionina fu messa in evidenza da H. Popper e altri (v., 1953) che, somministrandola per via alimentare, osservarono nei ratti lo sviluppo di noduli epatici con qualche modesto carattere di malignità. Successivamente, riducendo la percentuale dell'etionina dallo 0,50% allo 0,25%, E. Farber (v., 1959) ottenne, in rapporto alla più lunga sopravvivenza degli animali dipendente dalla riduzione degli effetti tossici, sviluppo di veri cancri del fegato. L'ipotesi oggi più accreditata è che l'etionina costituisca una molecola donatrice di alchili per gli acidi nucleici.
h) Metalli e non metalli
L'interesse suscitato da vari metalli e non metalli come agenti carcinogenetici è negli ultimi anni notevolmente diminuito a causa dell'incostanza dei risultati sperimentali e della sempre maggiore importanza che ha assunto la cancerogenesi da composti organici e, più recentemente, quella da virus.
Tra i metalli è stata attribuita attività cancerogena principalmente al cromo, al cobalto, al berillio e al nichel; di questi il più importante sembra essere il cromo, in relazione al non raro riscontro di tumori del polmone in operai addetti alla manipolazione di minerali contenenti il metallo. Da tempo era nota la frequenza di più o meno gravi infiammazioni, a volte a carattere necrotizzante, della mucosa nasale in operai addetti all'estrazione del cromo e alla fabbricazione di colori da questo derivati; nel 1936 W. Alwens e altri (v.) segnalarono numerosi casi di tumore polmonare in lavoratori nelle industrie di cromo, e successivamente la casistica è notevolmente aumentata così che al presente il cancro polmonare da cromo viene considerato da molti autori come un cancro professionale. Anche se molte volte i due tipi di alterazioni sono concomitanti, non è stato sicuramente accertato alcun rapporto tra lesioni nasali e tumori del polmone. Per quanto riguarda la presunta azione oncogena, pare che i cromati siano molto più attivi dei dicromati. Sperimentalmente, però, nonostante qualche risultato parzialmente positivo (v. Schinz e Uehlinger, 1942), non si è riusciti a produrre tumori negli animali; tale fatto sosterrebbe l'ipotesi secondo la quale il cromo non rappresenta la causa del cancro polmonare e i casi osservati sono verosimilmente cancri da fumo.
Ancora meno precise sono le conoscenze sulla possibile attività oncogena del berillio e del nichel, mentre può essere del tutto esclusa quella dell'alluminio, ipotesi che aveva inizialmente suscitato notevole allarme per il diffuso impiego di tale metallo nella fabbricazione di recipienti da cucina.
Per quanto manchino ancora sicure dimostrazioni, maggiore importanza come agente cancerogeno sembra avere l'arsenico. Era da tempo nota la relativa frequenza di cancro del polmone nei minatori dei giacimenti arseniferi dello Schneeberg e di Joachimsthal, come erano noti sin dal 1820 i tumori cutanei di alcuni abitanti in prossimità dei giacimenti d'arsenico di Reichenstein. Mentre fu accertato che i cancri polmonari delle due prime località erano imputabili non già all'azione dell'arsenico, bensì a quella di emanazioni radioattive (v. Rajewsky e altri, 1943), fu altresì dimostrato che l'acqua che bevevano gli abitanti di Reichenstein proveniva da una falda idrica situata al di sotto dei giacimenti e conteneva elevate quantità di arsenico: si pensò quindi che a tale elemento fosse presumibilmente imputabile l'elevata incidenza di tumori. A dimostrazione indiretta di tale interpretazione veniva fatta notare la scomparsa dell'incidenza neoplastica verificatasi nel 1928, a seguito della costruzione di un nuovo acquedotto. È certo che negli operai esposti all'azione cronica dell'arsenico sono frequenti alcune alterazioni cutanee localizzate preferibilmente sul tronco e sulle dita, in particolare ipercheratosi e papillomi, che sono considerate di tipo precanceroso e che in alcuni casi si trasformano in veri cancri cutanei. L'avvelenamento cr0nico da arsenico determina inoltre con una certa frequenza cirrosi epatica, sulla quale si osserva talvolta l'impianto di un tumore epatico, fatto che pone molti e gravi problemi patogenetici, allo stato attuale non solubili. Di fronte a queste e a varie altre osservazioni cliniche sta però la scarsa rilevanza delle ricerche sperimentali, perchè non è possibile produrre negli animali veri cancri da arsenico, a eccezione di rari casi. D'altra parte, è anche vero che ricercatori di alto livello, come A. Carrel (v., Le principe..., 1925) e A. Fischer (v., Die Erzeugung..., e Dauerzüchtung..., 1927), comunicarono di essere riusciti a trasformare in vitro cellule normali in cellule cancerose in presenza di tracce di arsenico, ma tali ricerche non hanno avuto seguito.
Pertanto, contrariamente a quanto si è verificato in altri casi, il reperto clinico non è sostenuto da inconfutabili prove sperimentali, e allo stato delle conoscenze non si può certo dubitare del primo nè trascurare le altre.
i) Cenni sui meccanismi d'azione dei carcinogeni chimici
Si è accennato che con particolari idrocarburi aromatici si possono ottenere tumori in varie specie animali; è relativamente facile provocare con queste sostanze carcinomi nel topo e sarcomi nel topo e nel ratto. Non sono ancora note le ragioni della differente sensibilità all'azione dei cancerogeni di due specie tanto filogeneticamente vicine; si consideri in proposito che per effetto dell'applicazione di idrocarburi, che è in grado di provocare lo sviluppo di carcinomi cutanei anche nel coniglio, si determina la comparsa di cancri soltanto nella cute del topo e non in quella del ratto, ma insorgenza di sarcomi sottocutanei in entrambe le specie.
Il fatto che sostanze a struttura chimica nota inducano crescita neoplastica fece sorgere l'idea, subito dopo l'isolamento dei primi idrocarburi cancerogeni, che si potesse studiare il meccanismo della carcinogenesi a livello morfologico. Tuttavia, le più accurate indagini istologiche non riuscirono a recare validi contributi alla soluzione del problema; le ragioni di tale insuccesso possono oggi essere comprese se si considera che i fenomeni che determinano la cancerizzazione si svolgono a livello molecolare, cioè a grandezze enormemente inferiori a quelle accessibili morfologicamente. È stato comunque possibile documentare alcuni eventi terminali espressi a livello di determinate strutture, che sono valsi soltanto a precisare e a definire quanto già si conosceva sull'istogenesi di noduli tumorali iniziali dell'uomo o di tumori spontanei o sperimentali di animali. D'altra parte, era facilmente supponibile che nella zona trattata con idrocarburi si sviluppassero fatti infiammatori o anche necrotici, con stimolo alla proliferazione vasale ed eventuale incapsulazione del veicolo lipoideo. Generalmente le sostanze cancerogene, quale che sia la loro struttura molecolare, non sono dotate della sola attività oncogena, ma possono agire in modo assai vario, svolgendo il ruolo di eccitatori, di mediatori chimici dell'infiammazione, di agenti necrotizzanti per alcuni tipi cellulari ed eccito-proliferativi per altri, e così via. Sulla cute gli idrocarburi determinano varie reazioni aspecifiche di tale tipo. La proliferazione da essi indotta mostra, in alcuni casi, qualche carattere di specificità, con conseguente formazione di cellule atipiche e quindi comparsa di caratteri di anaplasia cellulare.
Di particolare interesse nello studio dell'istogenesi dei tumori cutanei da idrocarburi appare il reperto, piuttosto frequente anche se non costante, della formazione di papillomi, che si possono comparare con alcune lesioni precancerose dell'uomo. E tuttavia difficile stabilire anche in questo caso se a tali manifestazioni debba essere attribuito un significato generale ed essenziale o non rappresentino piuttosto soltanto un fenomeno collaterale.
Il frequente riscontro, in fase precoce di formazione dei tumori, della paralisi delle mitosi lascia supporre che la stessa sostanza cancerogena o un prodotto del suo metabolismo esercitino azione antimitotica, alla stessa guisa di vari altri antimitotici.
La vera cancerizzazione è un fenomeno che si avvera ex novo: in accordo con P. Rondoni (v., 1946), si può dire che, in un determinato momento, tra le varie reazioni aspecifiche si manifesta una violenta attivazione delle mitosi e si forma l'ammasso di cellule neoplastiche. Si ritiene che non sia una cellula sola a subire la trasformazione neoplastica, ma che il processo interessi contemporaneamente un gruppo di cellule e che in generale lo sviluppo neoplastico sia pluricentrico, cioè non limitato a un solo centro di proliferazione: successivamente i vari centri continuano a proliferare, si fondono tra loro e costituiscono la massa tumorale il cui accrescimento è agevolmente osservabile. Anche in patologia umana, d'altronde, erano ben noti i fenomeni di accrescimento pluricentrico di tumori in fase iniziale.
La crescita ex novo del tessuto neoplastico si accorda, forse, con quanto si osserva nella trasformazione maligna delle cellule coltivate in vitro nelle quali la trasformazione dei caratteri che segna il passaggio dalla cellula normale alla cellula cancerosa è fenomeno critico, se è lecito paragonare quanto avviene in vivo con quanto accade nelle colture di cellule.
l) Cocancerogenesi
Strettamente connesso con la cancerogenesi chimica è il fenomeno della cocancerogenesi. Nel 1938 M. J. Shear (v.) scoprì che l'attività cancerogena del benzopirene è incrementata dall'olio di creosoto: a tale sostanza egli diede il nome di cocancerogeno per precisarne proprio l'azione cooperativa con il cancerogeno. Il termine ha avuto fortuna, non solo perché furono scoperte varie sostanze dotate di attività cocancerogena, ma soprattutto perché le ricerche sull'argomento misero in luce alcuni fenomeni la cui interpretazione è servita di base alla concezione della teoria dei due stadi dell'oncogenesi. Tale teoria fu espressa da I. Berenblum (v., 1941), il quale individuò nell'olio di croton, che si estrae dai semi di crotontiglio (Croton tilium), un potente cocarcinogeno.
Pennellando la cute di topo con quantità troppo esigua di benzopirene o con solo olio di croton non si osserva comparsa di tumore, ma il trattamento combinato con le stesse dosi di idrocarburo e con il cocancerogeno è in grado di determinare l'effetto oncogeno: l'olio di croton, cioè, potenzia l'azione del benzopirene. Il potenziamento è svelato dalla percentuale di animali nei quali insorge la neoplasia, dalla notevole abbreviazione del tempo di latenza, cioè dell'intervallo di tempo che intercorre tra l'applicazione del cancerogeno e l'insorgenza del cancro, e infine dal fatto che, mentre con determinate dosi di benzopirene si provoca la comparsa di un solo centro di sviluppo, con la contemporanea applicazione di entrambe le sostanze si induce un'insorgenza pluricentrica. Di conseguenza, il fenomeno può anche essere espresso quantitativamente (v. Berenblum, 1941).
L'olio di croton, che è un agente irritante cutaneo per molte specie, si comporta come cocancerogeno soltanto per la cute del topo; la sua azione cocancerogena, inoltre, si svolge nettamente nei confronti di alcuni idrocarburi aromatici, soprattutto del benzopirene, ma è nulla o assai scarsa nei confronti dell'1,2-benzantracene e dell'1,2:5,6-dibenzantracene. Esso agisce come cocancerogeno anche nei confronti dell'uretano che, pur essendo un carcinogeno generale capace di provocare tumori in vari organi, riesce a indurre comparsa di carcinomi cutanei con grande difficoltà e soltanto in particolari ceppi inbred di topi; questi carcinomi però si sviluppano facilmente nelle zone di cute pretrattate con uretano sulle quali si applichi il cocancerogeno.
Oltre all'olio di croton, sono state individuate numerose altre sostanze ad azione cocancerogena, tra le quali di notevole importanza alcuni detergenti: la dimostrazione della loro attività è stata fornita da ingegnosi esperimenti, condotti con trapianti di zone cutanee pretrattate con vari carcinogeni. La constatazione che l'azione del cocancerogeno si esercita anche varie settimane dopo la cessazione della pennellatura della cute con il carcinogeno induce a pensare che questo abbia prodotto nella cellula modificazioni funzionali irreversibili.
Sui fenomeni ora accennati e su vari altri, che non è possibile riportare in questa sede, è fondata la teoria dei due stadi, concepita da P. Rous e I. G. Kidd (v., 1941) e da I. Berenblum (v., 1941) e conosciuta con il nome di ‛carcinogenesi a due stadi' o anche di ‛carcinogenesi a molti stadi'. Fondamentalmente, secondo tale teoria, il carcinogeno durante un primo stadio determina la trasformazione delle cellule normali in cellule maligne. A questo punto possono verificarsi due condizioni: o il carcinogeno oltre a causare la trasformazione maligna è anche in grado di determinare fenomeni proliferativi a tendenza iperplastica, e allora il tumore si sviluppa in quanto le cellule hanno subito quella che attualmente si designa come trasformazione cellulare e sono avviate alla moltiplicazione; o il carcinogeno non possiede, o possiede solo in debole grado, attività eccitoproliferativa, e allora le cellule cancerizzate restano quiescenti per un tempo anche assai lungo. In quest'ultimo caso, tuttavia, è ancora possibile il rapido completamento del ciclo e lo sviluppo del tumore in seguito all'intervento di uno stimolo specifico, rappresentato dal cocarcinogeno. Alla cancerizzazione pura Berenblum diede il nome di ‛iniziazione' e alla stimolazione proliferativa quello di ‛promozione', distinguendo corrispondentemente gli agenti in ‛inizianti' e ‛promoventi'. Rous indicò l'iniziazione con il termine di ‛potenzialità neoplastica'. In Italia F. Pentimalli, in base alle sue ricerche sull'importanza dei fenomeni rigenerativi nello sviluppo del sarcoma del pollo da virus di Rous, indicò quest'ultimo come ‛fattore potenziale' e la rigenerazione, cioe la moltiplicazione cellulare, con il termine di ‛fattore realizzante'.
Più recentemente Berenblum (v., 1969), in un lucido inquadramento dei fattori che in senso positivo o negativo agiscono sullo sviluppo neoplastico, ha messo in evidenza che una certa confusione è generata dal frequente uso come sinonimi dei due termini ‛azione cocarcinogena' e ‛azione promovente'.
Il termine ‛cocancerogenesi' si usa in senso generale per indicare ogni tipo di aumento dell'induzione di tumore realizzata, generalmente, per mezzo della concorrente applicazione di carcinogeno e di cocancerogeno, sebbene in alcuni casi quest'ultimo possa esplicare la sua azione prima o dopo rispetto al primo.
Il termine azione ‛promovente', invece, è più limitativo, vale cioè a designare la condizione in cui il cocancerogeno viene applicato dopo completamento dell'azione iniziante propria del carcinogeno. Si conoscono molti cocarcinogeni che non esercitano azione promovente.
Per tali considerazioni, secondo Berenblum si debbono distinguere vari tipi di cocancerogenesi, a seconda dell'azione che esplicano le diverse sostanze: 1) azione ‛additiva', che si esplica nei confronti di un carcinogeno in grado di provocare da solo la comparsa del tumore; 2) azione sinergica, in senso farmacologico, che si verifica quando gli effetti combinati eccedono la sommazione degli effetti separati; 3) azione carcinogenica incompleta, consistente in attività nei riguardi di una sola fase della carcinogenesi, cioè solo per l'iniziazione o solo per la promozione; 4) azione preparativa, che si svolge sull'organo bersaglio rendendolo più sensibile all'azione carcinogenica; 5) azione permissiva, consistente in facilitazione della funzione carcinogenetica, che si esplica, per esempio, aumentando la solubilità o la velocità di assorbimento del carcinogeno da parte della cellula, o affrettandone il metabolismo o la velocità di escrezione; 6) azione sui Virus, che si svolge secondo meccanismi diversi: a) favorendo la liberazione delle particelle virali dal sito nel quale sono localizzate; b) deprimendo la reazione immunitaria dell'ospite; c) attivando un virus incompleto; d) rendendo le cellule bersaglio più sensibili al virus; e) deprimendo l'azione dell'interferon; 7) azione condizionante, consistente nell'intervento di alcuni fattori richiesti per la crescita di un tumore ormono-dipendente o di un tumore il cui sviluppo è frenato dalla reazione immunitaria dell'ospite.
L'azione cocancerogena studiata più a fondo è quella dell'olio di croton, che agisce come carcinogeno incompleto (tipo 3 dell'elenco), cioè come promovente che sollecita focolai di cellule, sulle quali si è già esplicata l'azione iniziatrice.
Si tratta naturalmente di schematismi di grande utilità per una materia così complessa come la cocancerogenesi, in quanto facilitano i processi di distinzione e specificazione, pur presentando a volte il rischio di indurre a inquadramenti semplicistici e non rigorosi.
m) Il fumo del tabacco
Connessa con la cancerogenesi chimica è la questione dell'importanza del fumo di tabacco quale agente eziologico del cancro polmonare. È fuori dubbio che negli ultimi anni la frequenza di tale tipo di tumore è assai aumentata: mentre in passato il cancro polmonare era raro, attualmente esso rappresenta un'alta percentuale dei tumori maligni, con differenze notevoli tra le singole popolazioni e in rapporto alle varie condizioni di vita. In passato l'importanza del tabacco quale causa di cancro aveva attratto principalmente l'attenzione per la frequenza di tumori localizzati nella cavità orale nei fumatori, in particolare di quelli delle labbra nei fumatori di pipa.
Il rapporto fumo di tabacco/cancro del polmone si è desunto fondamentalmente da dati statistici molto elaborati, che hanno dimostrato una netta prevalenza dei tumori polmonari tra i fumatori. Il problema ha assunto significato sociale e molte commissioni nei vari paesi, in particolare in Inghilterra, Stati Uniti e Paesi scandinavi, hanno svolto estesissime indagini con risultati che, in linea di massima, concordano nel porre in guardia l'umanità verso la pericolosità del fumo di tabacco, nell'indicare la durata media del tempo di induzione e la zona di pericolosità, cioè il numero di sigarette fumate per giorno, nel riconoscere l'indispensabilità di filtri idonei, ecc.
Le ricerche sperimentali non hanno dato, però, risultati significativi, nonostante le molte modalità cui si è fatto ricorso. La grande difficoltà di tali indagini consiste nell'impossibilità di riprodurre condizioni corrispondenti a quelle reali, soprattutto per quanto riguarda la meccanica del fumare. Inoltre, bisogna tener conto del fatto che il cancro polmonare si manifesta nei fumatori dopo molti anni dall'inizio dell'abitudine di fumare: tale lungo periodo di preparazione non è attuabile negli animali da esperimento per la relativa brevità della loro vita, e d'altra parte non si può accettare senz'altro l'idea che un evento morboso, che nella vita dell'uomo evolve durante un lungo periodo di tempo, negli animali a breve vita debba invece compiersi in un tempo a questa proporzionato.
Se è vero d'altronde che una relazione abbastanza stretta tra fumo e tumore polmonare si desume statisticamente per la diversa incidenza della malattia nei fumatori rispetto ai non fumatori, è altresì vero che anche per questi ultimi la frequenza di tali tumori è andata progressivamente aumentando, anche se in misura minore che nei primi. Tutto ciò induce a ritenere che l'eziologia dei tumori polmonari sia molteplice, dovuta cioè a più fattori, dei quali alcuni personali come il fumo, altri ambientali e tra questi in primo luogo il grave inquinamento dell'ambiente nell'epoca attuale.
Per la produzione sperimentale di tumori da fumo si sono usati metodi diretti e indiretti. I primi consistono nell'abituare alcuni tipi di scimmie a fumare (v. Jarvick, 1967): tuttavia gli animali fumano, e sembra volentieri, un numero molto limitato di sigarette ma rifiutano ulteriori inviti, e pertanto non sono stati conseguiti risultati di qualche importanza. E stato anche tentato di realizzare nei cani il fumo forzato attraverso tracheotomia, ma l'insorgenza di fenomeni morbosi collaterali, come emboli polmonari ed enfisema, non consente esperimenti di lunga durata.
I metodi indiretti sono basati invece sull'inalazione passiva, che si attua mantenendo gli animali da esperimento in ambiente saturo di fumo di tabacco. Nonostante alcuni studiosi, in particolare E. L. Wynder e D. Hoffmann (v., 1967), abbiano potuto osservare l'insorgenza di cancri ghiandolari del polmone in topi di ceppi sensibili ai tumori polmonari mantenuti in tali condizioni sperimentali, secondo altri autori (v. Di Paolo e Moore, 1959; v. Di Paolo e Levin, 1965) queste non riprodurrebbero in realtà gli effetti determinati dal fumo di tabacco nell'uomo.
È comunque dimostrato che con il catrame o con il condensato di tabacco, cioè con il materiale che si accumula, per esempio, nei cannelli di pipa, si producono tumori epiteliali negli animali da esperimento (v. Wynder e altri, 1953; v. Wynder e Hoffmann, 1967). È da tener presente che l'attività carcinogenetica dei condensati freschi, cioè di meno di 24 ore, è maggiore di quella dei condensati di più tempo, in rapporto alla presenza nei primi di sostanze volatili. Alcuni condensati sono attivi sugli epiteli, ove provocano l'insorgenza di carcinomi, e sul sottocutaneo, ove danno luogo a sviluppo di sarcomi; altri invece esplicano attività oncogena solo sui tessuti epiteliali. Frazionando con vari metodi il condensato di fumo di tabacco è stato possibile compiere importanti osservazioni, tra le quali di particolare interesse appaiono quelle di Wynder e Hoffmann che hanno identificato fattori inizianti e fattori promoventi la crescita neoplastica, cioè cocancerogeni: gli iniziatori sono risultati idrocarburi aromatici localizzati in una singola frazione, mentre i promoventi sono stati localizzati nella frazione acida, in quella neutra e in quella fenolica. La cancerogenesi da condensato di fumo avverrebbe cosi a opera dei due fattori, secondo la concezione di Berenblum dei cocarcinogeni.
Il problema dell'eziologia del cancro del polmone, notevolmente complesso per le molteplicità dei fattori in causa, è in ogni modo ancora lontano dalla sua soluzione.
3. Cancerogenesi fisica
Anche in questo caso la patologia sperimentale ha affrontato e per alcuni aspetti risolto il problema posto dall'osservazione clinica. Già da lungo tempo era nota la frequenza di tumori cutanei sulle parti scoperte di individui esposti per molti anni all'azione diretta delle radiazioni solari, in prevalenza quindi marinai e contadini. Più recentemente, in rapporto al rapido ed esteso sviluppo della diagnostica e terapia radiologica, è divenuto progressivamente più frequente e allarmante il riscontro di lesioni cutanee, evolventi talvolta in cancri, in medici e tecnici radiologi.
Da tali osservazioni sorse anzitutto la necessità di adeguate protezioni del personale addetto all'uso di apparecchi di radiologia e degli stessi ammalati che per scopi diversi venivano irradiati, protezioni da molti anni obbligatorie per legge.
In campo sperimentale, è stato possibile riprodurre agevolmente sia i tumori da irradiazioni solari, sia quelli da raggi Rüntgen, e la mole delle ricerche è veramente notevole.
a) Radiazioni ultraviolette
Nel 1928 G. M. Findlay (v.) vide che l'irradiazione con raggi ultravioletti della cute di ratti spennellata con catrame determinava più rapida comparsa di cancri cutanei rispetto agli animali di controllo sottoposti alla sola azione del catrame. Due anni dopo lo stesso autore dimostrò che la sola irradiazione è in grado di provocare l'insorgenza di cancri cutanei.
Seguirono naturalmente numerose altre ricerche, e al presente non v'è dubbio che i raggi ultravioletti esplichino azione cancerogena sia sull'epitelio, ove danno luogo alla comparsa di epiteliomi, sia sul derma e sull'ipoderma, ove provocano lo sviluppo di sarcomi; anzi, in condizioni sperimentali quest'ultimo tipo di tumori si osserva molto più frequentemente che non gli epiteliomi, contrariamente a quanto si verifica spontaneamente nell'uomo.
È stato precisato che dello spettro ultravioletto solo le radiazioni a lunghezza d'onda comprese tra 260 e 300 nm sono dotate di attività carcinogenetica, anzi per luci ultraviolette monocromatiche si è visto che tale attività è strettamente limitata a 297 nm. Si supponeva, e sembrava logico ritenere, che l'effetto cancerogeno fosse correlato con quello eritematoso, che cioè fosse dipendente da questo; si è potuto invece stabilire che non esiste alcuna relazione tra i due effetti, perché le radiazioni che inducono eritema hanno una lunghezza d'onda di 253,7 nm e come tali non sono cancerizzanti.
La prova diretta che i raggi solari sono efficaci nel produrre tumori fu fornita da A. H. Roffo (v., 1934) che esponendo topi e ratti a intensa irradiazione solare ottenne sviluppo di carcinomi.
b) Raggi X, isotopi radioattivi, radiazioni ionizzanti
L'attività oncogena dei raggi X fu dimostrata sperimentalmente da P. Marie e altri (v., 1910) che riuscirono con tale mezzo a indurre sviluppo di tumori nel topo, e venne poi ampiamente studiata da B. Bloch e N. Dreyfus (v., 1921).
I risultati delle numerose ricerche sull'argomento, condotte secondo varie modalità, hanno consentito di stabilire senza alcun dubbio che i raggi X, così come gli isotopi radioattivi, esplicano attività oncogena, e hanno quindi imposto la necessità di adottare adeguati mezzi di protezione per tutti coloro, chimici, fisici, biologi, mineralogisti, che per ragioni di lavoro sono esposti a fonti di radiazioni.
È ormai perfettamente chiarita la causa della notevole frequenza di tumori del polmone negli operai delle miniere di Schneeberg e di Joachimsthal: come si è già detto, misurazioni di radioattività dell'aria eseguite in tali ambienti hanno consentito di dimostrare che il contenuto in emanazione è assai alto e certamente di molto superiore alla concentrazione ritenuta tollerabile. Anche in una larga percentuale di animali da esperimento mantenuti per lungo tempo in miniera si osserva la comparsa di cancro del polmone. Fra le varie sostanze radioattive presenti nell'aria delle miniere il radon sembra svolgere il ruolo principale nell'eziologia della malattia (v. Hueper, 1954 e 1955; v. Hueper e altri, 1952). Così pure è stato possibile accertare che la frequenza di sarcomi delle ossa nelle operaie di una fabbrica di orologi nel New Jersey addette alla verniciatura dei quadranti con colori fluorescenti dipendeva dall'ingestione di sostanze radioattive. Le operaie, infatti, per l'abitudine di affilare la punta del pennellino con le labbra, ingerivano continuamente tracce di vernice contenente derivati del tono, il cui accumulo nelle ossa determinava entro un periodo relativamente breve, di pochi anni, l'insorgenza del tumore (v. Martland, 1929).
L'importanza delle radiazioni ionizzanti quale causa di tumori e di malattie correlate è risultata tragicamente evidente dopo la constatazione dell'impressionante aumento dei casi di leucemia tra i soggetti esposti agli effetti della esplosione delle bombe atomiche a Hiroshima e Nagasaki nel 1 945, nell'area compresa in un raggio di due chilometri dalle città l'incidenza della malattia, nella forma di leucemia mieloide, fu calcolata essere da sei a sette volte maggiore in tali popolazioni rispetto a quelle non comprese nelle aree contaminate. Si sono osservati due periodi di massima incidenza della leucemia mieloide nelle suddette popolazioni il primo compreso tra 4 e 8 anni dopo l'esplosione, con circa il 50% di casi acuti, il secondo tra 13 e 14 anni dopo l'esplosione, con l'84% di casi acuti, e il fenomeno non ha ancora ricevuto una soddisfacente spiegazione.
Già da tempo erano stati segnalati numerosi casi di leucemia tra i radiologi (v. Jagic e altri, 1911): è stato calcolato che questi specialisti presentano una probabilità di contrarre la malattia circa dieci volte maggiore rispetto agli altri medici.
Accurate statistiche hanno inoltre dimostrato l'aumento di frequenza della leucemia in bambini le cui madri siano state sottoposte a panirradiazione, durante la gravidanza.
Dal punto di vista sperimentale in alcuni topi adulti del ceppo LAF1 esposti a esplosioni atomiche fu possibile osservare lo sviluppo di tumori in vari organi, in particolare linfomi timici e leucemie e nelle femmine anche tumori ovarici e ipofisari. Più dimostrative furono invece le ricerche di H. S. Kaplan e M. B. Brown (v., 1952) e di L. Gross e altri (v., 1959), che ottennero alte percentuali di casi di leucemia in topi di ceppi inbred nei quali, in condizioni normali, l'incidenza della malattia è molto bassa: in particolare il Kaplan e il Brown dimostrarono la possibilità di osservare dopo panirradiazione X l'incidenza di leucemia nel 90% dei topi del ceppo C 57 Black - nei quali, spontaneamente, la malattia si manifesta sporadicamente o non si manifesta affatto - con frequenza massima negli animali di 2-4 settimane di vita e inversamente proporzionale all'età. Da rilevare che la leucemia consegue, come già detto, all'irradiazione di tutto il corpo, mentre l'irradiazione del solo timo ottenuta proteggendo con adeguata schermatura il resto del corpo è priva di effetto leucemogeno; inoltre, la timectomia preventiva riduce a circa un decimo i casi di leucemia da irradiazione o da sostanze chimiche, e questa inibizione dello sviluppo della malattia è a sua volta annullata dall'impianto di timo. Tali rilievi inducono a ritenere che il fatto che la leucemia sperimentale da irradiazione sia di tipo linfatico, sia del tutto indipendente da una azione diretta dei raggi sugli organi linfatici: Kaplan e Brown ritengono possibile la liberazione dai tessuti irradiati di un fattore leucemogeno, e la sua inattivazione da parte degli organi non irradiati nel caso di irradiazione parziale. L'inoculazione di cellule di midollo osseo esercita un certo potere inibente sullo sviluppo della leucemia, se praticata per via endovenosa subito dopo l'irradiazione. Inoltre, è stato osservato che un analogo potere inibente esercita sul topo l'inoculazione di poltiglia di milza della stessa specie e di milza di pecora, mentre un effetto inibente più modesto consegue all'inoculazione di estratti splenici acellulari. Si è pertanto supposta l'esistenza di un fattore antileucemogeno, che è stato indicato con la sigla RLP (Radiation Leukaemia Protecting factor), attivo nei confronti della leucemia da raggi ma privo di effetti nei confronti di quella da sostanze chimiche (v. Berenblum e altri, 1965).
Di notevole interesse è il rilievo che l'irradiazione di topi appartenenti a ceppi con alta incidenza di leucemia non determina aumento di tale incidenza: sembrerebbe cosi dimostrato che in tali ceppi la potenzialità leucemica è già espressa ai valori massimi e pertanto non incrementabile ulteriormente dalle radiazioni, mentre nei ceppi a bassa incidenza di leucemia questa potenzialità è pressoché inespressa e suscettibile di essere esaltata fino alla completa espressione dal trattamento irradiante.
L. Gross (v., 1958) in topi del ceppo C3H e M. Liebermann e H. S. Kaplan (v., 1959) in topi del ceppo C57 Black poterono fornire l'importantissima dimostrazione che dagli organi degli animali nei quali si è sviluppata la leucemia da irradiazione è possibile isolare un virus che può essere trasmesso in serie in animali normali neonati: l'irradiazione, cioè, non sarebbe causa diretta di leucemia, ma agirebbe attivando un virus leucemogeno latente, che sarebbe il vero effettore della malattia. Tale virus risulterebbe identico al virus A della leucemia spontanea di Gross sia per l'aspetto delle particelle osservate al microscopio elettronico, sia dal punto di vista immunologico, differenziandosene soltanto per particolarità minori.
Contrariamente a quanto accade per il topo, nel ratto la panirradiazione non induce lo sviluppo di leucemia bensì l'elevata incidenza di tumori solidi in vari organi, nei quali però non si è finora riusciti a dimostrare particelle virali. Ciò non autorizza tuttavia ad ammettere senz'altro una refrattarietà dei ratti alla leucemia da virus, perché è ben noto che questi animali sono sensibilissimi all'azione leucemogena del virus A; piuttosto si può pensare che nel ratto non esista un virus potenzialmente leucemogeno, e che la panirradiazione attivi invece virus in grado di indurre lo sviluppo di tumori solidi (v. Gross, 1970).
Secondo Gross, nel topo il virus attivabile da radiazioni è trasmesso dai genitori alla prole, cioè in via verticale, e solo così se ne spiegherebbe la presenza in ceppi come il C3H a bassissima incidenza leucemica. Gross non esclude la possibilità, finora in realtà non dimostrata, di trasmissione del virus latente con il latte.
Non si conosce ancora il meccanismo con il quale i vari tipi di irradiazione determinano tumori solidi o leucemie, ma certamente esso è molto complesso e va inquadrato nell'ambito delle conoscenze radiobiologiche che hanno avuto negli ultimi anni uno sviluppo veramente notevole (v. radiologia medica; v. radiobiologia). Occorre inoltre ricordare che le stesse radiazioni che esplicano attività cancerogena possono, in determinate condizioni, determinare regressione almeno temporanea di tumori e di leucemie, onde il loro ben noto impiego terapeutico.
Sarà in seguito fatto cenno all'azione delle radiazioni su qualche fenomeno cellulare elementare.
c) Cancerogenesi da solidi
La scoperta dello sviluppo di tumori in corrispondenza di superfici solide si deve a F. C. Turner (v., 1941) che osservò in ratti l'insorgenza di sarcomi in prossimità di dischi di bachelite introdotti nel tessuto sottocutaneo.
Al caso si deve la scoperta di B. S. Oppenheimer e altri (v., 1948) dello sviluppo di sarcomi in ratti nei quali, allo scopo di produrre ipertensione sperimentale, i reni erano stati avvolti con un foglio di cellofan. Successivamente, si riuscì a produrre sarcomi mediante impianto sottocutaneo di fogli di cellofan. Tuttavia il concetto di carcinogenesi da superfici solide fu delineato con precisione da H. Nothdurft (v., 1955), il quale vide che mentre la polvere di cellofan risultava poco attiva nell'indurre la formazione di sarcomi, i dischi di tale materiale mostravano di possedere attività oncogena in grado direttamente proporzionale all'estensione della loro superficie, cioè erano fortemente attivi quelli interi e meno attivi quelli forati. Tali prove consentivano di escludere un'azione da sostanze chimiche, e il processo venne indicato come cancerogenesi da superficie di corpi allo stato solido. Il rapido sviluppo delle ricerche mise in evidenza la possibilità di produrre, sia pure in grado diverso, sarcomi con l'impiego di fogli di molti tipi di plastiche, come il dracon, il teflon, l'ivalon, il polietilene, il polistirene e vari altri, mentre le polveri o trucioli di tali plastiche risultavano inattivi.
Un'ulteriore sorpresa provocarono le ricerche di G. Hecht (v., 1952), il quale osservò sviluppo di sarcomi nel tessuto sottocutaneo di ratti in seguito all'impianto di foglioline d'oro e successivamente di lamine d'avorio. Oppenheimer e altri (v., 1956) riprodussero i tumori con l'impianto di lamine d'acciaio, di tantalio o di vitallio (lega a base di cromo, cobalto, nichel e molibdeno). Successivamente si dimostrò che anche l'asbesto induce sviluppo di sarcomi e che, mentre polveri di quarzo o di vetro comune non svolgono attività oncogena, questa è svolta invece da cristalli o lamine di quarzo o da lastrine di vetro.
Ricerche istogenetiche hanno consentito di risolvere parzialmente il problema, facendo luce sulle modalità di sviluppo delle masse neoplastiche.
Attorno alle lamine di materiale solido si forma una capsula connettivale la quale per vari mesi resta quiescente, cioè senza segni di attiva proliferazione cellulare o di infiammazione, fino a quando in corrispondenza di una piccola zona inizia la moltiplicazione cellulare di elementi atipici e quindi lo sviluppo del tumore. Nel periodo di quiescenza la capsula è costituita da fibre collagene e da numerosi fibroblasti immaturi, con scarsa partecipazione vasale. Le polveri, invece, determinano reazioni infiammatorie più o meno vivaci, con partecipazione di macrofagi, neoformazione vasale e comparsa di cellule giganti. Di particolare interesse appaiono due osservazioni: asportando la lastrina di metallo o di plastica, con una porzione della capsula che l'include, si verifica egualmente sviluppo di sarcoma sui residui della capsula (v. Oppenheimer e altri, 1958); il trapianto nel tessuto sottocutaneo di ratti normali della sola capsula connettivale, priva cioè della lamina, dopo alcuni mesi dalla sua formazione, è seguito dallo sviluppo di sarcoma (v. Nothdurft, 1960). Molto persuasiva è l'interpretazione di L. S. Salyamon (v., 1961), secondo il quale la carcinogenesi da solidi è il risultato dell'inibizione della infiammazione che si determina a livello delle superfici dei solidi, mentre le polveri, come si è detto, costituiscono lo stimolo di una cospicua reazione infiammatoria, frequentemente sfociante anche nella formazione di granulomi.
A F. Bischoff e G. Bryson (v., 1964) si deve un'analisi dei termini di confronto tra formazione di sarcomi da solidi e trasformazione in cellule sarcomatose di fibroblasti normali coltivati in vitro. Nel 1941 C. P. Gey (v.) comunicò di aver osservato trasformazione neoplastica in colture di cellule, suscitando un enorme interesse perché per la prima volta veniva dimostrata una cancerizzazione spontanea, cioè senza intervento di carcinogeno. Successivamente W. R. Earle (v., 1943) confermò tale osservazione, escludendo in modo assoluto la possibilità di contaminazione delle colture con tracce di carcinogeni chimici o di virus. Attualmente non si hanno più dubbi sulla possibilità di cancerogenesi spontanea di cellule coltivate in vitro.
La cancerogenesi in vitro e quella da solidi presentano il carattere comune dell'adesione dei fibroblasti a superfici inerti e l'interposizione di uno strato di proteine tra cellule e superfici, che limita gli scambi gassosi. Secondo H. Goldblatt e G. Cameron (v., 1953) l'anaerobiosi, in armonia con la teoria di Warburg, è condizione favorente la trasformazione spontanea delle cellule coltivate in vitro in cellule neoplastiche.
Il meccanismo della cancerogenesi da solidi è tuttora ignoto. L'ipotesi che dalle sostanze plastiche si formino radicali liberi, cioè si verifichi trasferimento di elettroni instabili dalla superficie del polimero alla cellula, così come accade per le radiazioni, è probabilmente da escludersi, perché ricerche condotte da H. Druckrey e D. Schmäll (v., 1954) mediante l'impiego di polietilene, che contiene radicali liberi, e di cellofan, che ne è esente, non mettono in evidenza significative differenze nella capacità delle due sostanze di indurre formazione di sarcomi.
A. Caputo (v., 1973) prende in considerazione la possibilità che per formazione di legami a idrogeno tra polimeri e membrane dei lisosomi si determini un danno di tali strutture che darebbe luogo alla liberazione di DNA-asi e conseguentemente ad alterazioni delle catene polinucleotidiche del DNA.
È certo che il problema dell'attività carcinogenetica delle sostanze plastiche è di notevole interesse medico non solo per l'impiego attualmente assai diffuso di plastiche sostitutive, ma anche per l'inquinamento ambientale da parte di componenti a base di plastiche poliviniliche. La dimostrazione fornita da P. L. Viola e altri (v., 1971) che l'inalazione di aria con un contenuto del 30% di cloruro di vinile induce tumori della cute, del polmone e delle ossa è quindi di grande importanza teorica e pratica.
4. Cancerogenesi virale
a) Caratteristiche generali dei virus oncogeni
Gli studi sull'oncogenesi virale hanno dimostrato che sono in grado di produrre la formazione di tumori sia virus a DNA sia virus a RNA.
I virus a DNA dotati di potere oncogeno sono il virus del polioma e il virus SV40, il virus del papilloma, gli adenovirus, i poxvirus, gli herpesvirus; il loro DNA ha una struttura a doppia elica, e nei virus del polioma, SV40 e del papilloma presenta una forma ad anello in quanto le estremità di ogni catena sono legate tra loro da legami covalenti di natura ancora non precisata (v. acidi nucleici). Questi tre virus sono inoltre caratterizzati dal piccolo diametro delle particelle e dal più basso peso molecolare del DNA (rispettivamente per i primi due 450 Å e 3•106 dalton, per il terzo 550 Å e 5•106 dalton), e dal fatto che il loro esiguo genoma può codificare al massimo dieci proteme ciò consente di distinguere facilmente nella cellula trasformata i prodotti genici di origine virale e quelli di origine cellulare, e di precisare il numero dei geni che intervengono nel processo di trasformazione cellulare. Adenovirus, poxvirus ed herpesvirus hanno diametro delle particelle maggiore, peso molecolare del DNA più elevato (per ognuno dei tre: 21-23•106 dalton, 160•106 dalton e circa 100•106 dalton) e morfologia più complessa; i poxvirus causano tumori benigni cutanei in alcune scimmie e talvolta nell'uomo e un fibroma nel coniglio, gli herpesvirus sono ritenuti gli agenti eziologici di alcuni tumori degli animali e dell'uomo, quale l'adenocarcinoma di Lucké del rene di rana e il linfoma maligno di Burkitt.
I virus a RNA sono causa di tumori in molte specie animali: appartengono a tale gruppo proprio il virus della eritroleucemia dei polli, il primo virus di cui fu scoperto il potere oncogeno (v. Ellermann e Bang, 1908), quello del sarcoma dei polli (v. Rous, 1911), e altri come il virus di Bittner del carcinoma mammario del topo, quelli della leucemia-sarcoma del topo, quelli del sarcoma-leucemia del gatto, identificati molto più tardi e oggetto di ricerche ancor oggi molto estese. Sono virus di dimensioni piuttosto grandi (diametro delle particelle fino a 1.000 Å e oltre), il cui genoma è costituito da un solo filamento di RNA caratterizzato da un peso molecolare relativamente poco elevato.
Per quanto riguarda le caratteristiche generali dell'oncogenesi virale, i problemi più interessanti sono quelli relativi ai meccanismi con i quali i virus inducono la trasformazione cellulare. Come abbiamo già detto, questa consiste essenzialmente nel fatto che le cellule si sottraggono ai meccanismi di controllo della moltiplicazione per l'intervento di fattori in grado di agire sulla loro superficie o sulle loro strutture interne. I virus sono risultati in grado di indurre alterazioni sia della membrana cellulare (come nel caso del virus del sarcoma di Rous, già citato), sia dei processi intracellulari determinando alterazioni di alcune attività enzimatiche e della sintesi del DNA e conseguente comparsa di aberrazioni cromosomiche. Un altro aspetto importante della trasformazione cellulare indotta dai virus oncogeni a eccezione dei virus a RNA della leucemia murina, dell'adenocarcinoma mammario del topo e della eritroleucemia dei polli, è che le cellule trasformate contengono l'acido nucleico virale, DNA o RNA, in forma funzionalmente incompleta: le ragioni di tale differente comportamento degli acidi nucleici virali nelle cellule trasformate non sono note. Il problema della conoscenza del meccanismo della trasformazione è strettamente connesso al riconoscimento dell'alterazione metabolica che ne è responsabile, e quindi del prodotto o dei prodotti genici che determinano il fenomeno, e all'identificazione di quelle variazioni metaboliche che ne rappresentano invece una conseguenza. Di grande importanza sono, a tale riguardo, gli studi con i mutanti temperatura-sensibili. Cellule infettate con uno di tali mutanti crescono in vitro come cellule normali, e non sono da queste differenziabili, se tenute a temperatura non permissiva, cioè 41 °C; sebbene producano virus, e quindi siano infettate, tali cellule non sono dunque trasformate. Se le stesse cellule sono invece mantenute a 36 °C, cioè a temperatura permissiva, si determina la trasformazione, che retrocede tuttavia se la temperatura è nuovamente portata a 41 °C. Appare chiaro che in tali mutanti si è determinata una particolare alterazione responsabile della codificazione di una o più proteine denaturabili dalla temperatura, la cui identificazione e il cui studio sono essenziali per la comprensione del processo di trasformazione che esse sono in grado di indurre e di mantenere.
Un altro vantaggio offerto dallo studio dei mutanti temperatura-sensibili è rappresentato dalla possibilità di ottenere sincronizzazione della trasformazione: infatti, infettando le cellule a temperatura più bassa e lasciando trascorrere il tempo necessario perché tutte siano infettate, è possibile ottenere in poche ore la trasformazione di tutti gli elementi con l'elevazione della temperatura a 36 °C.
Esaminiamo ora alcuni dei tumori sperimentali da virus, tra quelli più studiati e diffusamente impiegati nelle ricerche.
b) Virus polioma e SV40
La storia della scoperta di questo virus è incerta. Nel suo libro Oncogenie viruses, L. Gross (v., 1970) riferisce che nel 1951, in alcuni dei topi neonati di ceppo C3H ai quali aveva inoculato un filtrato di tessuti di topi di ceppo AK affetti da leucemia spontanea, osservò, anziché l'atteso sviluppo della leucemia, la comparsa di tumori del collo in ambo i lati. Egli pubblicò questa sua osservazione soltanto due anni dopo, cioè proprio quando S. E. Stewart (v., 1953) comunicava di aver rilevato lo stesso fenomeno. È difficile dire a chi tocchi la priorità della scoperta, nonostante che Gross la rivendichi a sé con molte argomentazioni.
Gross indicò il virus come parotid tumor virus, perché i tumori del collo comparsi in seguito all'inoculazione di filtrato erano adenocarcinomi della parotide. Qualche anno dopo Stewart e altri (v., 1957) riuscirono a isolare un virus capace di produrre tumori anche di altri organi, e proposero quindi di chiamarlo polyoma virus (virus di tumori multipli), nome che ha incontrato fortuna ed è divenuto di uso corrente.
È questo uno dei virus più profondamente studiati non solo nelle ricerche di oncogenesi, ma anche in quelle di virologia generale e di biologia molecolare.
La sua inoculazione determina comparsa di alterazioni dei tubuli renali di aspetto adenocarcinomatoso e, in una certa percentuale di casi, di sarcomi del rene talvolta bilaterali, sviluppo di carcinomi della mammella, di tumori delle ossa, di carcinomi dei surreni e di altri tipi di neoplasmi. Complessivamente, secondo le osservazioni di Stewart e collaboratori, il virus del polioma è in grado di provocare ben 23 tipi di tumori a struttura istologica differente, sebbene con frequenze molto diverse.
In generale non esiste alcuna relazione tra la leucemia e i vari tumori solidi indotti dal virus, nel senso che solo eccezionalmente la sua inoculazione è seguita dalla contemporanea comparsa dei due tipi di processi morbosi. Si è pertanto supposto che questi siano in realtà provocati da due virus diversi occasionalmente presenti in uno stesso animale o in un particolare ceppo inbred di topi, e che tra essi si stabilisca un fenomeno di interferenza tale che la penetrazione di uno dei due in una cellula impedisce l'azione dell'altro. Questa ipotesi sembra confermata da varie prove abbastanza convincenti, quali la possibilità di annullare con il riscaldamento a 56 °C per 30 minuti l'attività leucemogena, ma non quella cancerogena, dell'estratto di colture cellulari infettate e l'inattivazione della sola attività cancerogena ma non di quella leucemogena del virus, a opera del siero di topo immunizzato mediante trattamento con liquido di colture cellulari in cui si è moltiplicato il virus polioma.
Il virus polioma viene facilmente coltivato in colture soprattutto di cellule embrionali di topo; è di notevole interesse il fatto che la coltivazione in serie determina progressivo aumento non solo della produzione, ma anche dell'attività del virus, nel senso che la capacità di indurre tumori a strutture istologiche diverse è maggiore per il virus coltivato su cellule in vitro che per quello che si estrae dalla massa tumorale.
Il virus polioma è attivo su molti ceppi, inbred e non inbred, di topi; in criceti neonati provoca lo sviluppo di vari tumori, come nel topo, e talvolta sarcomi del muscolo cardiaco, che nell'uomo e negli animali da esperimento è raramente sede di neoplasmi. Questa particolare sensibilità di topi e criceti neonati e di varie altre specie (ratti, furetti, conigli, cavie, un tipo di ratto selvatico) ha determinato un larghissimo impiego del virus nelle ricerche di oncologia sperimentale. La varietà di specie sensibili al virus e la capacità di questo di indurre tumori di molteplici tipi istologici, se per certi aspetti rappresentano un vantaggio nella soluzione di determinati problemi tecnici, di fatto danno tuttavia luogo a una situazione sperimentale multiforme e complessa.
A tale proposito va anche considerato che il virus polioma è presente in un'altissima percentuale di topi normali di ceppi inbred diversi dall'originario AK, cioè in quelli C3H, C57 Brown ecc., senza alcuna relazione con la frequenza in questi ceppi di incidenza spontanea di leucemia o di carcinoma della mammella.
Furono inizialmente identificati diversi ceppi del virus polioma, dei quali uno caratterizzato da maggiore attività oncogena nei confronti di ratti e criceti rispetto al topo, uno da una particolare affinità per il timo, altri forniti di elevata attività citopatogena e bassa attività carcinogenetica. La vera fase scientifica sui mutanti del virus polioma iniziò, però, quando M. Fried (v., 1965) dimostrò che al trattamento con acido nitroso resistono solo alcune particelle virali : se con queste si infettano colture di cellule embrionali di pollo mantenute a temperature di 31 °C (temperatura permissiva) e di 38 °C (temperatura non permissiva), si osserva che il 25% delle placche di cellule trasformate che ne derivano risultano sensibili alla temperatura, cioè possono svilupparsi solo a temperatura permissiva. I mutanti del virus in grado di indurre la formazione di tali placche vennero indicati con la sigla ts (temperature-sensitive). Da questo primo esperimento numerosissime ricerche si sono susseguite, sia con altri mutageni come l'idrossilammina, sia con varie linee cellulari come la 3T3 (v. Di Mayorca e altri, 1969), tanto che finora sono stati isolati circa 200 mutanti ts. Analoghe osservazioni sono state in seguito compiute sul virus SV4O e su quello di Rous. Si ritiene oggi che la temperatura permissiva consenta la sintesi di alcuni prodotti genici, impossibile alla temperatura non permissiva: si dispone così di un espediente tecnico di grandissima importanza per la virologia oncologica, perché consente di distinguere alcuni fatti della trasformazione cellulare come dipendenti o indipendenti dall'attività virale, soprattutto di precisare quanta parte del genoma virale è indispensabile per il mantenimento nella progenie cellulare e quindi anche nel tumore in vivo del carattere di trasformazione.
Nel 1959 G. A. Di Mayorca e altri (v.) fornirono l'im- portante dimostrazione che il DNA del virus polioma, isolato con metodi chimici e inoculato in colture di cellule di embrione di topo, determina effetto citopatogeno e, do- po due passaggi, produzione di particelle virali che, a loro volta inoculate nel topo, provocano la comparsa di tumori. La prova decisiva che la sostanza isolata biologicamente attiva fosse DNA fu fornita dalla sua inattivazione specifica, cioè dalla perdita della sua attività infettante, dopo trattamento con desossiribonucleasi. L'estrazione del DNA infettivo è facilitata dal riscaldamento delle cellule a temperature comprese tra 75 e 100 °C, con un meccanismo e per ragioni tuttora non conosciuti; trattasi comunque di un DNA dotato di una notevole termoresistenza, che può essere mantenuto per 30 minuti a 100 °C senza che perda la sua attività. Successivamente, in altri laboratori fu realizzata l'estrazione del DNA anche con altri solventi, e si dimostrò inoltre la capacità infettante di DNA estratti da virus del papilloma di Shope (v. Ito, 1960) e SV40 (v. Boiron e altri, 1962).
Numerose sono state le ricerche di microscopia elettronica per l'identificazione delle particelle virali del polioma e la dimostrazione delle loro dimensioni, condotte iniziai- mente su liquido di colture cellulari, poi su sezioni ultrasottili: nel corso di tali studi fu possibile anche dimostrare la presenza nel nucleo delle cellule infettate di cospicui addensamenti di particelle virali a struttura cristallina. La grandezza dei virus può anche essere determinata, oltre che con la microscopia elettronica, mediante l'ultrafiltrazione attraverso membrane di collodio a porosità nota e per mezzo dell'ultracentrifugazione.
Nel nucleo sono state messe in evidenza anche formazioni filamentose di diametro corrispondente a quello delle particelle di tipo cristallino, la cui comparsa precede quella delle particelle: basandosi su tali osservazioni W. Bernard e altri (v., 1959) hanno concluso che in realtà i filamenti sono costituiti dalle particelle stesse. Quando queste divengono così numerose da riempire i nuclei, passano nel citoplasma: qui non è possibile osservare filamenti, ma si rinvengono soltanto particelle piccole e grandi, queste ultime facenti parte di corpi inclusi.
Nelle cellule dei tumori, di qualsivoglia struttura istologica, le particelle virali si riscontrano in numero molto piccolo o non si riscontrano affatto: ciò corrisponde al fatto, noto già dagli studi iniziali, come si è accennato, che dai tumori si estraggono solo piccole quantità di virus, contrariamente a quanto si verifica per le cellule coltivate in vitro e infettate con virus polioma dalle quali se ne ottengono grandi quantità.
Nelle cellule infettate con virus polioma sono presenti due antigeni, che mancano nelle corrispondenti cellule normali, uno dei quali compare nel nucleo prima che inizi la sintesi del DNA virale e prima della comparsa dell'antigene capsidico, e può essere dimostrato per mezzo dell'immunofluorescenza o della fissazione del complemento: tale antigene, inizialmente designato con la sigla ICFA (Inducing Complement Fixation Antigen), è stato poi denominato T per indicarne la presenza oltre che nelle cellule infettate anche in quelle trasformate e in quelle dei tumori. Anticorpi antiantigene T sono dimostrabili nel siero di criceti adulti portatori di tumori trapiantabili da virus polioma o nel siero di topi neonati infettati con tale virus. Antigeni T sono anche presenti in cellule infettate con SV40 o con adenovirus, ma poiché i corrispondenti anticorpi non danno reazioni crociate si ritiene che questi antigeni siano prodotti dal genoma virale e quindi specifici di ciascun virus.
Un'altra classe di antigeni dimostrata nei tumori indotti da virus polioma, SV40 e adenovirus, indicata con la sigla TSTA (Tumor Specific Transplantation Antigens), favorisce la resistenza al trapianto di cellule tumorali in animali adulti precedentemente infettati con tali virus. In queste condizioni sperimentali, infatti, in un certo numero di animali nei quali si è sviluppato il tumore a seguito dell'inoculazione di virus si determina il rigetto, così come avviene nei trapianti di organi o di tessuti tra animali della stessa specie, fenomeno essenzialmente di natura immunitaria dovuto all'attività che i linfociti dell'ospite esplicano nei confronti del tessuto trapiantato. È probabile che questi antigeni siano localizzati sulla membrana cellulare.
I quesiti relativi ai due tipi di antigeni sono numerosi e complessi, e investono il problema più generale dell'immunologia dei tumori: a tale proposito, occorre tener presente che è stata dimostrata un'antigenicità anche nei tumori indotti da sostanze dotate di attività carcinogenetica, nei quali una delle più tipiche caratteristiche è la rarità delle reazioni crociate immunologiche tra i vari tipi di tumori.
Il virus SV4O, molto simile al virus polioma per quanto riguarda sia le caratteristiche fisiche e chimiche, sia quelle biologiche quali la capacità di indurre la trasformazione cellulare e di formare antigeni, si trova largamente diffuso nelle colture di rene di scimmia impiegate per la coltivazione di virus patogeni per l'uomo e in particolare di quello della poliomielite. La scoperta del virus SV4O si deve a B. H. Sweet e M. R. Hilleman (v., 1960), i quali videro che il liquido di coltura di cellule renali di rhesus o di cynomolgus produceva in quelle di cercopiteco un intenso effetto citopatico, consistente essenzialmente nella formazione di vacuoli. Il virus responsabile di tale alterazione fu quindi chiamato agente vacuolizzante e successivamente indicato con la sigla SV4O proposta da R. N. Hull e altri (v., 1956) e facente seguito alla serie di sigle progressive SV1, SV2 ecc. con le quali erano stati designati vari virus isolati in colture di cellule renali di scimmia. L'attività oncogena del virus fu dimostrata in criceti neonati da B. E. Eddy e altri (v., 1961) e confermata in vari altri laboratori; fu tra l'altro messa in evidenza la sua capacità di indurre tumori del cervello in un roditore (Mastomys natalensis), e soprattutto quella di trasformare cellule umane coltivate in vitro.
Naturalmente, il fatto che il virus SV4O fosse riscontrato in preparazioni di vaccino Sabin destò un certo allarme, sebbene l'infezione di volontari con tale virus non abbia dato luogo a fenomeni morbosi. Sono stati fatti innumerevoli tentativi per evitare la contaminazione di vaccini con SV40, e si sono adoperati in proposito anticorpi specifici o sostanze chimiche come il beta-propiolattone, che è risultato in grado di inattivare il virus SV40 senza distruggere quello poliomielitico.
c) Tumori da virus erpetici: il linfoma di Burkitt e l'adenocarcinoma di Lucké del rene di rana
Come accennato, certi virus erpetici sono considerati da alcuni studiosi, sulla base di dimostrazioni molteplici e di rilievi epidemiologici di notevole importanza, agenti eziologici di tumori umani e di tumori degli animali. Ciò nonostante, esistono tuttora non pochi dubbi principalmente per il fatto che, specie nell'uomo, i virus erpetici sono molto diffusi così come diffusa è la presenza di anticorpi antierpetici. È ben noto, inoltre, che herpesvirus persistono in molti individui anche per tutta la vita, provocando occasionalmente eruzioni erpetiche nonostante la presenza di anticorpi specifici nel sangue circolante. La struttura morfologica, la composizione chimica e le varie fasi dei virus nella cellula ospite sono state ampiamente indagate negli ultimi anni, ma le conoscenze acquisite sulla loro oncogenicità sono tuttora relativamente scarse, nonostante gli sforzi compiuti.
I due tumori per i quali l'eziologia erpetica è altamente probabile sono il tumore di Burkitt dell'uomo e l'adenocarcinoma di Luckè della rana; meno certe sono le conoscenze sull'eventuale significato eziologico dei virus erpetici nei riguardi del carcinoma a cellule squamose del collo dell'utero della donna e per alcuni altri tumori di animali. Poiché per i primi due le ricerche sono state estesissime, specie negli ultimi anni, e da esse sono derivati fatti e idee di ordine generale, si riporta in sintesi la sequenza delle scoperte.
D. Burkitt (v., 1958), chirurgo inglese nell'ospedale di Kampala in Uganda, individuò una particolare sindrome, occorrente con una certa frequenza in bambini sino ai 14 anni di età viventi in determinate zone, nelle quali rappresenta circa il 50% dei tumori dell'infanzia. Si tratta di un linfoma la cui localizzazione più frequente è nei mascellari, ma che può manifestarsi in vari organi e in modo particolare nelle ovaie; tuttavia, contrariamente a quanto si osserva in altri linfomi, non determina aumento di volume della milza nè dei linfonodi, ma solo occasionalmente rare adenomegalie a sede addominale, e non si accompagna a comparsa di cellule abnormi nel sangue periferico nemmeno negli stadi terminali.
Burkitt e collaboratori visitarono cinquanta ospedali dell'Africa centrale e poterono osservare numerosi casi della malattia, diversamente diagnosticati. Per l'Uganda fu accertata l'esistenza, prima dell'osservazione di Burkitt, di almeno duecento casi del linfoma. Fu così individuata la cintura del linfoma (lymphoma belt), una vasta zona compresa all'incirca fra i 15 gradi di latitudine nord e i 20 gradi di latitudine sud. Inoltre, l'incidenza della malattia apparve limitata ad altitudini inferiori ai 1.600 metri, e ciò fece concepire l'idea che l'agente eziologico fosse trasmesso da un insetto vettore incapace di vivere al di sopra ditale altezza: esso però non è stato identificato nemmeno nel corso delle indagini più recenti. Va sottolineato, e in seguito ne sarà dimostrata la ragione, che vari casi di linfoma di Burkitt sono stati riscontrati nella Nuova Guinea, alcuni in Canada, alcuni negli Stati Uniti, in Colombia e in regioni del Sudafrica, un caso tra gli eschimesi.
Per un certo tempo sembrò che tali reperti potessero infirmare il concetto di lymphoma belt, che tuttavia successivamente ricevette piena conferma dalla constatazione che la frequenza del tumore nei territori della cintura è enormemente più alta, tanto da assumere aspetto epidemiologico; solo nella Nuova Guinea è possibile rilevare un'incidenza relativamente più elevata del linfoma, che invece si manifesta negli altri paesi in casi isolati.
Vari virus furono di volta in volta osservati e descritti, alcuni di tipo erpetico, altri di tipo reovirus (v. Bell e altri, 1964), ma la conclusione fu che essi erano solo virus occasionali, di frequente osservazione nelle cellule neoplastiche, e conosciuti con il nome di opportunist passengers. Si era, così, nella condizione di non riuscire a individuare il virus agente eziologico di una malattia che pure, per molte considerazioni, si doveva ritenere di natura virale.
M. A. Epstein e Y. M. Barr (v., 1965) considerarono la possibilità che il virus nelle cellule del tumore fosse mascherato dagli anticorpi specifici formatisi nel corso della malattia, e la conseguente estrema difficoltà di dimostrarlo con la microscopia elettronica; essi ritennero quindi che solo in colture di cellule avulse da ogni possibile contatto con anticorpi si sarebbe riusciti a porlo in evidenza. La coltivazione in vitro di materiale bioptico di linfoma di Burkitt, compiuta in Uganda dagli stessi autori nel 1964 e da R. J. V. Pulvertaft (v., 1967) in Nigeria, dette luogo allo sviluppo di linee continue di linfoblasti, eliminando così ogni possibile dubbio sulla natura neoplastica della malattia in quanto è noto che i linfociti normali non danno linee continue. Nel corso di tali ricerche, dopo alcuni passaggi, sia in sezioni ultrasottili degli elementi linfoblastici sia su pellets di loro estratti furono osservate particelle virali di tipo erpetico che vennero indicate con la sigla EB, Epstein-Barr (v. Epstein e Barr, 1964; v. Epstein e altri, 1964 e 1966). Successivamente, S. Toshima e altri (v., 1967) poterono osservare tali particelle anche all'esame diretto delle cellule del linfoma.
Osservazioni oltremodo interessanti sono scaturite nel corso di tali studi: Stewart (v., 1969) ha dimostrato che in criceti neonati e timectomizzati l'inoculazione combinata di virus EB e di dimetilsulfossido (DMSO) determina l'insorgenza di un'encefalite trasmissibile in serie; questa malattia in altre specie - scimmie, topi, cavie, conigli e gattini neonati - è invece provocata dal solo virus anche in assenza di DMSO. La spiegazione di questi dati sperimentali non appare semplice: si potrebbe infatti ammettere sia la contaminazione con un virus neurotropo, sia un vario potere patogeno dell'EB che sarebbe in grado di provocare nell'uomo il linfoma e negli animali l'encefalite. Di estrema importanza è senza dubbio la capacità mostrata dall'EB di indurre in vitro crescita illimitata di linfociti del sangue periferico, sicura espressione di trasformazione neoplastica.
Una svolta importante negli studi sul linfoma di Burkitt è stata segnata dalle ricerche immunologiche: con tecniche di immunofluorescenza si è dimostrato che i sieri di soggetti ammalati e i sieri di conigli nei quali sia stato inoculato l'EB reagiscono solo con le cellule contenenti il virus, la cui presenza è facilmente controllabile al microscopio elettronico; si è inoltre osservato che reagiscono con tali cellule anche i sieri di un'elevata percentuale di individui normali.
Tutto ciò induce ad ammettere che il virus EB sia largamente diffuso e determini negli organismi la formazione di anticorpi che ne impediscono l'attività patogena, e che nelle zone ove la sua diffusione è meno estesa, più elevata è la frequenza di soggetti non immuni e quindi suscettibili ad ammalarsi.
L'immunologia ha inoltre convalidato il ruolo eziologico del virus EB per il linfoma con la dimostrazione della comparsa nelle cellule infettate con esso, analogamente a quanto è possibile osservare in cellule trasformate da altri virus, di un neoantigene al quale sarebbe dovuta la reattività immunologica nelle prove di immunofluorescenza.
Per quanto riguarda la possibile identificazione dell'EB con altri virus, occorre anzitutto precisare che esso presenta una notevole affinità con il virus dell'herpes simplex, ma se ne differenzia nettamente perché non è in grado come questo di indurre effetto citopatogeno e per l'assenza di corrispondenza immunologica.
Suggestiva appare l'ipotesi della probabile esistenza di rapporti tra EB e virus della mononucleosi infettiva dell'uomo: già da tempo infatti è stata segnalata una certa frequenza di tale malattia in individui a contatto con ammalati di linfoma, e a opera di numerosi ricercatori e soprattutto di G. Henle e altri (v., 1968) è stato dimostrato che in ammalati di mononucleosi gli anticorpi antivirus EB - presenti come abbiamo già detto negli individui normali - sono assenti nel siero nelle fasi iniziali e fanno la loro comparsa più tardi durante il decorso della malattia.
Lo studio delle colture cellulari ha consentito inoltre osservazioni di estremo interesse. Anzitutto, è stato dimostrato che al pari delle cellule del linfoma i linfociti del sangue periferico degli ammalati di mononucleosi danno linee continue: tale comportamento, caratteristico delle cellule neoplastiche, è in netto contrasto con la natura assolutamente benigna della mononucleosi che anzi regredisce spontaneamente, ma occorre ricordare che, con una certa frequenza, regrediscono anche linfomi di Burkitt. Si è poi accertato che sia nelle cellule del linfoma coltivate in vitro, o nei linfociti normali infettati con EB, sia nelle cellule della mononucleosi è presente un'anomalia cromosomica consistente nell'accorciamento del braccio lungo del cromosoma 10.
La scoperta che anticorpi antivirus EB sono largamente presenti nelle popolazioni ha portato necessariamente ad ammettere che tale virus sia molto diffuso nel mondo e che la valida immunizzazione che precocemente si determina nei suoi confronti ne renda impossibile l'identificazione anche al microscopio elettronico e la coltivazione con i tradizionali metodi virologici: la sua attività patogena può quindi esplicarsi solo nei rari casi nei quali l'individuo non è venuto a contatto con esso, con una certa analogia a quanto avviene per la poliomielite. Lo studio di questi fenomeni ha prospettato una serie di problemi quanto mai complessi, tra i quali soprattutto la ricerca e il chiarimento dei motivi per cui in assenza di immunizzazione in alcuni soggetti si sviluppa il linfoma e in altri la mononucleosi.
Nel 1934 B. Lucké (v.) pubblicò uno studio accurato di un adenocarcinoma renale che aveva osservato in molte delle rane catturate nell'area del lago Camplain nel Vermont; il tumore fu riscontrato anche in rane viventi in zone lacustri del Wisconsin e del Quebec, e in altre zone. La frequenza con la quale era possibile osservare tale neoplasia negli Anfibi fece per la prima volta prospettare l'ipotesi della trasmissione di un tumore maligno per contagio.
Lo studio di tale neoplasia permise di accertare una serie di fatti inaspettati e singolari. Anzitutto fu dimostrato che il tumore può essere trapiantato a rane dello stesso ceppo, ma non a quelle della stessa specie ma di ceppo diverso: così, il trapianto non attecchisce se un adenocarcinoma renale di una rana del Vermont, ad esempio, viene trapiantato in una del Wisconsin, pur appartenendo ambedue gli anfibi alla stessa specie, Rana pipiens. Tuttavia, malgrado questa strettissima specificità nell'ambito della stessa specie, la trasmissibilità del tumore fu operata con successo nel tritone (Triturus viridiscens o Diemictylus viridiscens), cioè addirittura in un animale appartenente ad altro genere: nel nuovo ospite il trapianto si sviluppa come adenocarcinoma del rene, ma talvolta anche come condrosarcoma, vale a dire come un tumore di struttura istologica completamente differente. Fu inoltre dimostrato che il successivo trapianto in Rana pipiens di questi due tipi di tumori sviluppatisi nel tritone dà luogo a sua volta a sviluppo di entrambi i tipi istologici: di questi, però, il condrosarcoma regredisce, mentre l'adenocarcinoma si sviluppa ulteriormente perdendo la peculiare originaria specificità di ceppo così da poter essere trapiantato in rane di altre zone.
Sin dalle prime indagini è apparso verosimile che la neoplasia sia causata da un virus, in quanto non solo lo stesso Lucké aveva dimostrato la possibilità di riprodurla mediante inoculazione di estratti di tumore essiccato e conservato a bassa temperatura o anche tenuto in glicerina al 50%, ma si osservò anche che l'introduzione di frammenti di tumore in parti diverse del corpo della rana dà luogo, per la diffusione dalla sede di impianto di un virus che si fissa elettivamente nel tessuto renale, a sviluppo di adenocarcinoma del rene anziché di un tumore locale: questo fenomeno è particolarmente evidente se si opera l'innesto nella camera anteriore dell'occhio, ove l'attecchimento è notevole e rapido, ma seguito dalla regressione della massa che inizialmente si sviluppa e dalla comparsa di un adenocarcinoma del rene.
Singolare è anche il fatto che la crescita del tumore è vivace durante i mesi estivi, si attenua o cessa del tutto durante quelli invernali e riprende in primavera. Dai tumori invernali il virus si estrae facilmente in grande quantità, da quelli estivi se ne estrae pochissimo o non se ne estrae affatto: questo fenomeno dell'esistenza di un rapporto inverso tra crescita tumorale e produzione di virus pare dipendere esclusivamente dalla temperatura, perché M. Mizell e altri (v., 1969) hanno osservato che è possibile estrarre virus solo da tumori di rane tenute artificialmente a bassa temperatura e non da quelli di animali mantenuti a temperatura elevata. Si tratta con ogni probabilità di un fenomeno analogo a quello noto per alcuni fagi, per il polioma, l'SV40, il virus di Rous, i cui mutanti temperatura- sensibili esplicano a bassa temperatura - temperatura permissiva - alcune attività che non sono invece in grado di esplicare a temperatura elevata, cioè non permissiva, come si è già accennato.
Connessa con la produzione di virus nelle rane ibernanti è la diffusione dell'infezione: in primavera parte del virus prodotto viene eliminato con le orme, diffuso nell'acqua, assunto dai girini, con conseguente sviluppo di tumore, mentre ciò non può avvenire durante l'estate.
La prima prova diretta dell'esistenza di un virus quale agente eziologico dell'adenocarcinorna renale fu fornita da W. R. Duryll (v., 1956), il quale dimostrò che un estratto di tumore filtrato su filtro da batteriologia e inoculato nelle rane dà luogo a sviluppo dell'adenocarcinoma.
Con le prime osservazioni di microscopia elettronica si poté dimostrare su sezioni ultrasottili di tumore l'esistenza di particelle virali endonucleari e di particelle virali citoplasmatiche, le prime di tipo erpetico, le seconde di tipo poliedrico, ma non si riuscì a stabilire se le une o le altre o entrambe fossero i reali agenti eziologici.
Dalle colture in vitro di cellule del tumore e di cellule normali di Anfibi e di specie diverse, compresi i Mammiferi, infettate con materiale proveniente dal tumore della rana, sono stati isolati numerosi ceppi di virus, che sono stati indicati con una sigla seguita da un numero: quelli isolati dalla Rana pipiens con sigla FV (Frog Virus) seguita da numeri progressivi, FV1, FV2, FV3, ecc. (l'FV3 è stato isolato da un tumore renale, FV1, FV2 da rane normali); altri ceppi con la sigla L (L1, L2, L3, ecc.), altri ancora con quella LT (LT1, LT2, LT3, ecc.).
Alcuni di tali ceppi sono stati anche isolati da tumori fatti sviluppare in ospiti intermedi, ad esempio nel tritone, allo stato larvale e a quello adulto.
La maggior parte dei ceppi isolati è di tipo virus cito- plasmatici, e si riscontrano in cellule parenchimali, ad esempio del fegato, sia di animali normali sia di animali portatori di tumore. Nella compagine del tumore i virus citoplasmatici sono anche frequenti, ma sono localizzati solo in cellule stromali anziché in quelle neoplastiche. Attualmente, però, si ritiene che i virus citoplasmatici non abbiano significato eziologico e sono quindi considerati come opportunist passenger viruses. Con maggiore probabilità, invece, possono essere identificate come agenti eziologici della neoplasia le forme virali di tipo erpetico osservabili al microscopio elettronico nelle sezioni del tumore.
Dei ceppi isolati l'FV4 è il solo di natura erpetica ; poi- ché la sua curva di crescita in colture cellulari è bifasica, si ammette il concorso di un helper, cui è stata data la sigla CAV (Cell Associated Virus), in analogia a ciò che si verifica per virus di neoplasie di altre specie e in particolare del sarcoma di Rous.
Tuttavia, né con i virus citoplasmatici né con FV4 è possibile indurre sviluppo di tumore in animali adulti o in embrioni, girini, larve ecc., o provocare la trasforma- zione neoplastica in cellule coltivate in vitro. Quindi, per lo stesso virus di tipo erpetico, che è maggiormente indi- ziato quale agente del tumore di Lucké, è mancata la prova eziologica decisiva della produzione della neoplasia.
È probabile che, in base alle conoscenze del virus EB del tumore di Burkitt, anche la questione dell'eziologia del tumore di Lucké venga chiarita: infatti è certo che tra i due tipi di tumore, per quanto riguarda la situazione virologica, le affinità o analogie sono molto strette.
d) Il sarcoma di Rous
Nel 1911 Peyton Rous, nell'Istituto Rockefeller di New York, descrisse un sarcoma dei polli provocato da un virus, che fu indicato con il n. 1 quando, di lì a poco, seguirono le scoperte di numerose altre neoplasie di questo tipo: tra queste, con il n. 5 fu designato il tumore osservato nel 1915 a Napoli da F. Pentimalli (v., Quinto tumore..., 1916), mentre con il n. 7 venne indicato un osteocondrosarcoma che lo stesso Rous in collaborazione con J. B. Murphy e W. H. Tyler individuò nel 1912 (v. Rous e altri, 1912). Circa 50 sono complessivamente i sarcomi provocati da virus scoperti finora nei polli e in altri volatili. Attualmente il sarcoma di Rous è considerato uno dei più importanti materiali di ricerca in oncologia sperimentale.
Dal punto di vista più strettamente patologico il tumore di Rous è, tra i tumori degli animali, uno di quelli che più si avvicinano ai tumori dell'uomo sia per l'aspetto morfologico sia per l'evoluzione maligna. Le cellule sono fortemente anaplastiche, la crescita è di tipo infiltrativo con distruzione del tessuto invaso, con grande frequenza compaiono metastasi soprattutto nel polmone e nelle ovaie e gli animali vanno rapidamente incontro a cachessia.
L'iniziale ipotesi che il trapianto del tumore fosse possibile solo in alcune razze di polli, comunque non in altri volatili, fu in seguito smentita: come sarà chiarito più avanti, furono studiate le ragioni della differente recettività delle varie razze al sarcoma, e fu possibile isolare ceppi di polli sensibili o insensibili all'azione del virus. Si è comunque accertato che in generale gli animali giovani sono molto più suscettibili a tale azione di quelli adulti, e che nei più vecchi, specialmente nei galli, il trapianto del sarcoma non attecchisce o tutt'al più è seguito dallo sviluppo di un fibroma che abitualmente non contiene virus.
Nel 1914 in Giappone A. Fujinami e K. Inamoto (v.) avevano osservato un tumore del pollo trapiantabile in animali della stessa specie, e quattro anni dopo ne aveva- no dimostrano la filtrabilità dell'agente eziologico. Successivamente Fujinami e K. Suzue (v., 1928) riuscirono a trapiantare il tumore in anatre, dimostrando così per la prima volta la possibilità di eterotrapianti che, innestati nel pollo, riproducevano il tipico tumore. Queste ricerche, che hanno avuto in seguito larghissima estensione, hanno dimostrato la possibilità di effettuare il trapianto in molte altre specie animali, in particolare in Mammiferi, consentendo così la scoperta di alcuni fatti di fondamentale importanza.
Quando Rous, nel 1911, descrisse il sarcoma indicato con il suo nome, fornì pure la prima dimostrazione dell'eziologia virale della neoplasia, poiché riuscì a riprodurre un sarcoma identico come struttura a quello originario inoculando nei muscoli di un pollo un filtrato per candela di tale tumore. Inoltre, lo stesso Rous in collaborazione con Murphy (v. Rous e Murphy, 1912 e 1914) dimostrò pure che frammenti del tumore essiccati o conservati in glicerina al 50% inoculati in polli danno luogo a sviluppo della neoplasia.
Innumerevoli seguirono poi le ricerche tendenti a definire i caratteri del virus responsabile del sarcoma; soltanto nel 1947 fu possibile ad A. Claude e altri (v., 1947) osservare al microscopio elettronico su sezioni ultrasottili di tumore o di pellets da centrifugazione di estratto di tumore la presenza di particelle virali, e il reperto fu successivamente confermato con varia frequenza da numerosi altri ricercatori.
Sin dalle prime ricerche apparve chiaro che le cellule stimolate in modo aspecifico alla proliferazione risultano più recettive al virus: fu infatti osservato che l'inoculazione di filtrato acellulare di tumore, che specialmente se ottenuto da neoplasia a lento accrescimento può non dar luogo a crescita neoplastica, è prontamente seguita dalla comparsa di sarcoma se si aggiunge al materiale inoculato polvere di diatomee, e Pentimalli dimostrò inoltre che un focolaio rigenerativo, determinato meccanicamente in un tessuto, diviene sede di sarcoma in seguito all'introduzione endovenosa del filtrato acellulare (v. Pentimalli, Lesioni dei tessuti..., 1916). Tali fenomeni possono ora essere interpretati sulla base della differente sensibilità delle cellule ai carcinogeni nelle varie fasi del ciclo cellulare. E stato osservato che il virus del sarcoma di Rous può provocare manifestazioni diverse, neoplastiche o emorragiche, a seconda dell'età dell'organismo infettato: infatti, F. Duran-Reynals (v., 1940) dimostrò che la sua inoculazione determina generalmente nei pulcini appena nati una malattia emorragica caratterizzata dalla presenza di sangue fluido o coagulato nel peritoneo, da emorragie in vari organi, da splenomegalia; in quelli di qualche settimana di vita comparsa di emorragie e noduli neoplastici; in animali adulti formazioni di tumori nel cui contesto sono talvolta osservabili emorragie. È interessante rilevare che nel liquido emorragico degli animali infettati si rinvengono notevoli quantità di virus, prodotti, come è facilmente intuibile, nelle cellule dell'organismo ospite. È pensabile che le cellule dell'animale giovane, o almeno alcune tra esse, vadano incontro per azione del virus esclusivamente a infezione progressiva, e che solo quelle dell'animale adulto presentino quella particolare forma di infezione abortiva che è caratterizzata da trasformazione e contemporanea produzione di virus.
In vari modi si è tentato di isolare il virus di Rous; W. R. Bryan (v. Bryan e altri, 1954) è riuscito a ottenere preparazioni che conservano per anni l'attività se mantenute sotto vuoto e a bassa temperatura: si tratta, però, di colture eterogenee di virus, alcune pure, altre contenenti virus della leucosi del pollo. Attualmente, i vari ceppi di virus isolati dal pool di Bryan e da altri ricercatori, cioè il CarrZilber particolarmente adatto per la trasmissione in Mammiferi, i due Bryan standard strain e high titer strain, il Mill Hill, l'Harris, il Praga, il ceppo Schmidt-Ruppin patogeno anche per i Mammiferi, non mostrano proprietà biologiche univoche.
Gross, in base a una serie di dati raccolti da P. J. Simons e R. M. Dougherty (v., 1966), ritiene che tutti questi ceppi provengano dal sarcoma n. 1 di Rous, e che i numerosi passaggi in polli di razze differenti abbiano dato luogo a fenomeni di selezione e in qualche caso abbiano favorito la contaminazione con virus occulti esistenti allo stato latente in quegli animali.
Un nuovo orientamento venne impresso agli studi sul virus di Rous, e in genere sui virus oncogeni a RNA, dalla dimostrazione fornita da H. Hanafusa e altri (v., 1963 e 1964) che il ceppo high titer strain è in grado di indurre formazione di tumori anche a rapida crescita, ma non di dar luogo a progenie di virus. Questo fatto fu messo in relazione con la mancanza di un fattore necessario alla formazione di particelle virali complete, il quale venne designato con il termine helper o con la sigla RAV (Rous Associated Virus), e fu poi identificato come virus della leucosi aviaria; la presenza contemporanea dei due virus nelle cellule sensibili induce formazione di virioni completi, la cui parte interna cioè il nucleoprotide contenente il genoma è codificata dal virus di Rous e quella esterna cioè l'envelope dal RAV. Poiché i due virus possono coesistere in uno stesso ospite ma anche apparire isolati, si ammette l'esistenza di virus di Rous completi capaci di codificare tutte le componenti delle particelle virali, e di virus difettivi che abbisognano dell'helper per la formazione delle particelle. I virus di Rous possono quindi venire distinti in completi o difettivi, ed essere indicati con una sigla nella quale le lettere comprese tra parentesi denotano se l'envelope è codificato dallo stesso virus o da quello delle leucosi: RSV.A (RSVA) ceppo autosufficiente, RSV.O (ALV.A) ceppo difettivo completabile con ALV.A (Avian Leucosis Virus).
La paziente e laboriosa selezione di razze di polli caratterizzate da differente sensibilità ai vari ceppi ha consentito poi di classificare il virus di Rous nei quattro sottogruppi A, B, C, D; la linea di polli che si dimostrò sensibile a tutti questi sottogruppi fu indicata con la sigla Ch/O. Tuttavia, soltanto alcuni degli animali di tale linea apparvero sensibili al RSV.O, così che fu poi possibile selezionare due ceppi di polli Ch/O, rispettivamente sensibili e resistenti al RSV.O. Corrispondentemente le cellule dei Ch/O sensibili sono sensibili in vitro ai cinque virus, A, B, C, D, O, mentre quelle dei Ch/O resistenti sono sensibili ai primi quattro virus e insensibili al quinto sottogruppo.
Gli studi di H. Rubin (v., 1960 e 1961) dimostrarono che i sottogruppi dei virus del sarcoma e di quelli della leucosi interferiscono tra loro: ad esempio, una coltura di cellule Ch/O sensibili se è infettata con virus A della leucosi non fissa il virus A del sarcoma, ma fissa invece gli altri virus, B o C o D; la situazione inversa si determina se la coltura è infettata con un sottogruppo di virus di sarcoma. Queste ricerche hanno offerto la possibilità non solo di identificare determinati sottogruppi di virus e di approfondire gli studi di genetica del virus di Rous, ma anche di compiere importanti osservazioni di ordine immunologico: è stato infatti scoperto che in alcuni polli sono presenti antigeni capaci di reagire con anticorpi specifici antiantigeni dei virus della leucosi e del sarcoma. Tali polli sono indicati come COFAL positivi (COFAL, Complement Fixing Avian Leucosis), mentre vengono detti COFAL negativi quelli privi di antigeni. Si ammette che nelle cellule dei polli COFAL positivi deve necessariamente essere presente un antigene o un sito antigenico identico a quello posseduto dal virus della leucosi o dal virus del sarcoma, e quindi un gene che ne codifichi la sintesi: ciò fa supporre che nel genoma di tali cellule esista un frammento del genoma del virus del sarcoma o della leucosi, stabilmente integrato. Recentemente R. A. Weiss e altri (v., 1971) hanno dimostrato che con vari agenti chimici e fisici è possibile indurre la produzione di virus strutturalmente simili a quelli della leucemia-sarcoma del pollo da parte di cellule di pollo normale, cioè infetto. Si è osservato che il fattore COFAL è trasmissibile come un virus della leucosi aviaria che può essere ripreso da un particolare sottoceppo, RSV.B (RSV.O), ed è costituito da RNA a 65S. Come si può facilmente comprendere, queste ricerche, tuttora in pieno svolgimento, sono estremamente interessanti anche da un punto di vista biologico generale.
Di grande interesse è la possibilità di trasmettere il tumore non solo ad altri uccelli - anatre, come abbiamo già ricordato, e fagiani (v. Andrewes, 1932) - ma addirittura in mammiferi: L. A. Zilber e I. N. Kriukova (v., 1957) trapiantarono con successo il ceppo Carr-Zilber del sarcoma di Rous in ratti, nei quali osservarono poi la formazione di cisti in parte simili a quelle descritte da F. Duran-Reynals (v., 1940) nei pulcini neonati. Presto si vide che agli stessi risultati si poteva giungere anche impiegando i ceppi Praga e Schmidt-Ruppin, con i quali ceppi anzi fu pure possibile il trapianto in topi. Successivamente, si operarono trapianti in ratti a coda cotonosa, in criceti siriani, in cavie, in cani e in alcune specie di scimmie.
I tumori si inducono sia per innesto di tessuto sia, ma con minore percentuale di attecchimento, mediante inoculazione di supernatante di estratto tessutale.
Lo studio degli eterotrapianti ha messo in evidenza che mentre dai tumori facilmente inducibili con trapianto da pollo a mammifero e con quello in serie da mammifero a mammifero non si ottiene produzione di virus o se ne estrae una quantità minima, l'impianto del tumore del mammifero in polli determina invece lo sviluppo di virus. Se si coltivano contemporaneamente cellule di tumore di ratto con cellule di pollo di ceppo sensibile e si determina la loro fusione a mezzo di virus Sendai, inattivato con raggi ultravioletti, si forma un heterokaryon in grado di produrre virus.
Recentemente D. Simkovic (v., 1972) ha schematizzato la situazione dei rapporti tra virus del sarcoma aviario e cellule di tumori dei Mammiferi prodotti da virus di Rous indicando le seguenti possibilità: a) produzione di virus da parte delle cellule del tumore del mammifero; b) produzione di virus soltanto in presenza di cellule sensibili di polli combinate in heterokaryon a mezzo di virus Sendai; c) nessuna produzione di virus con qualunque mezzo; d) persistenza o moltiplicazione per vario tempo del virus nelle cellule, nelle quali provoca alterazioni morfologiche e sintesi di antigene virale ma non trasformazione maligna.
Questi fatti sono di estremo interesse, ma la loro ancora incompleta conoscenza non consente, al momento attuale, di trarre conclusioni generali.
I rapporti a volte molto stretti che esistono tra il virus del sarcoma e quello della leucosi dei polli inducono oggi ad ammettere l'esistenza di un complesso indicato come sarcoma-leucemia del pollo, anche se in realtà, come si è detto, si tratta di due virus diversi dei quali è possibile la separazione. La forma per la quale più evidenti appaiono tali connessioni è la mieloblastosi aviaria, i cui virus conosciuti, 1 e 2, sono frequentemente associati al virus di Rous, tanto che correntemente si designano con una sola sigla: AMV
Un problema che ha sempre appassionato gli studiosi è quello dei rapporti tra carcinogeni chimici e agenti virali. L'affermazione di A. Carrel (v. Carrel, Un sarcome..., 1925) che da tumori indotti da idrocarburi aromatici era possibile estrarre un virus riproducente il tumore in animali normali, inizialmente contrastata soprattutto da P. R. Peacock (v., 1933), è stata poi ampiamente confermata dalle ricerche di C. Oberling e M. Guérin (v., 1950): sembra così dimostrato che gli agenti chimici attivano nel pollo un virus oncogeno preesistente in stato latente o potenziale. Pur non essendo possibile al momento attuale attribuire a questi dati sperimentali un valore generale, occorre comunque tener presente che il virus di Rous e quelli delle leucosi sono largamente diffusi tra i polli e non necessariamente in rapporto a presenza di tumore o di leucosi; d'altra parte, pare assodato che tutti i tumori spontanei che sono stati trovati nel pollo siano di natura virale, e si è potuto osservare come il complesso sarcoma-leucosi sia in alcuni allevamenti una malattia largamente diffusa.
Con la dimostrazione che il virus di Rous è in grado di determinare la trasformazione di fibroblasti in coltura, H. Temin e H. Rubin (v., 1958) realizzarono un sistema semplice ed essenziale per studiare il meccanismo con il quale il virus provoca la formazione di tumore. Nel corso di tali ricerche fu possibile osservare che le cellule trasformate dal virus continuano a moltiplicarsi indipendentemente dalla densità cellulare, la quale invece rappresenta un fattore limitante nelle colture di fibroblasti normali.
e) Il complesso sarcoma-leucemia del topo
Nel 1951 L. Gross (v.) scoprì che una forma di leucemia del topo è determinata da un virus; in seguito furono isolati numerosi altri virus, alcuni identici, altri per qualche aspetto diversi da quello di Gross, per un totale di oltre duecento ceppi, la cui classificazione tuttavia è ancora incerta.
Sicuramente sette ceppi hanno caratteristiche proprie, e vengono indicati con la sigla MuLV (Murine Leukemia Virus), o più comunemente con quella MLV (Mouse Leukemia Virus), preceduta dal nome del ricercatore: Gross, Moloney, Rauscher, Friend, Kaplan, Graffi.
L'inoculazione in topi, ratti o criceti di un virus leucemogeno determina sviluppo di linfosarcomi negli organi emolinfopoietici, più frequentemente nel timo, spesso nella milza, nel fegato, nei gangli linfatici. Nel contesto del quadro morboso così provocato, il reperto di cellule immature nel sangue periferico non è affatto costante, e limitato agli stadi terminali, onde appare evidente che il termine leucemia è in realtà usato in senso largo.
Si conoscono anche forme di leucemia mieloide da virus, caratterizzate da presenza nel sangue di cellule immature della serie mieloide, ed è noto che il virus di Friend e quello di Rauscher determinano una eritroleucemia.
Nel 1964 J. J. Harvey (v.) inoculando in topi neonati BALB/C plasma di ratto portatore di leucemia da virus di Moloney conservato a lungo a bassa temperatura, osservò insorgenza di sarcoma in prossimità della zona di inoculazione oltre allo sviluppo di linfosarcomi nella milza. Due anni più tardi J. B. Moloney (v., 1966) inoculando ancora in topi neonati del ceppo BALB/C lo stesso virus da plasma conservato e centrifugato, ottenne soltanto sviluppo di rabdomiosarcomi nel sito di inoculazione, senza comparsa di linfosarcomi o di leucemie. Apparve quindi evidente il ruolo eziopatogenetico di un virus differente da quelli precedentemente isolati, cui fu dato il nome di virus del sarcorna murino e del quale furono poi individuati tre ceppi, indicati ognuno con la sigla MSV o MuSV (Mouse o Murine Sarcoma Virus) preceduta dall'iniziale del cognome del ricercatore che l'ha isolato. In considerazione dei rapporti assai stretti tra virus leucemogeni e virus del sarcoma, si usa comunemente la sola espressione di complesso sarcoma-leucemia murina, così come nel caso dei polli si parla di complesso sarcoma-leucemia aviaria.
La morfologia delle particelle virali del complesso sarcoma-leucemia murina è molto simile a quella del complesso aviario; il loro RNA comprende due tipi di molecole con il primo dei quali è identificabile il materiale genetico, costituito da un unico filamento. Gli studi sulla costituzione di queste particelle hanno consentito di individuare un componente polipeptidico che viene indicato come antigene gs (gruppo specifico), poichè è risultato comune a tutti i componenti del complesso sarcoma-leucemia murino; esso è presente anche in virus di sarcoma-leucemia di criceti e di gatti, così che si considera la possibilità di un suo eventuale impiego per la ricerca di virus oncogeni a RNA dell'uomo. I virus della leucemia e del sarcoma del topo si comportano in modo del tutto simile ai corrispettivi virus aviari: infatti, i leucemogeni infettano i fibroblasti in coltura senza trasformarli e passano nel mezzo dopo essersi moltiplicati, mentre quelli del sarcoma sono in grado di indurre la trasformazione dei fibroblasti. Il virus del sarcoma è difettivo, può cioè riprodursi soltanto se le cellule vengono infettate anche con virus leucemico: si forma in tal caso un fenotipo misto o pseudo-tipo, denominato MSV (MLV) per indicare che il materiale genico è di virus sarcomatoso e l'envelope è di virus leucemico con funzione di helper. In genere, i due virus coesistono in uno stesso materiale, così come accade per quelli del complesso aviario nell'high titer virus di Bryan, e conseguentemente dalle cellule trasformate con la miscela derivano particelle virali complete. I fibroblasti trasformati crescono meno rigogliosamente di quelli normali, forse per un accumulo di metaboliti tossici inibenti la moltiplicazione, e pertanto non si sviluppano quelle generazioni successive alla moltiplicazione di una cellula trasformata indicate come foci clonali; si formano invece, in prossimità di cellule che producono gli pseudo-tipi del virus del sarcoma, foci proliferativi che sono stati denominati da W. P. Rowe e altri (v., 1970) ‛foci a placca'.
L'iniziale ipotesi dell'intervento di due fattori nella trasformazione, ossia di un two-hits phenomenon consistente nella contemporanea infezione di una stessa cellula da parte di MSV (MLV) e di MLV, fu poi dimostrata errata quando in varie linee cellulari, quali ad esempio la BALB/C, si osservò crescita di tipo clonale indotta dal virus di uno stock molto diluito, in situazione quindi caratterizzata da assai scarse probabilità di infezione contemporanea di una stessa cellula con i due virus. Come è stato già detto, l'infezione di una cellula con MSV (MLV) è seguita dalla trasformazione e da crescita di tipo clonale, ma non dalla produzione di virus: la possibilità del recupero del genoma dell'MSV con la superinfezione di virus MLV e della conseguente produzione di particelle virali complete ma sempre fenotipicamente miste - cioè MSV (MLV) - è un'ulteriore conferma che la trasformazione è un single-hit phenomenon. È da notare che, finora, non si conosce un ceppo di virus sarcomatoso del topo indipendente da quello della leucemia, mentre per il virus di Rous sono stati dimostrati ceppi autonomi, cioè senza helper.
La constatazione che il genoma del virus del sarcoma si trasmette da una cellula alla progenie sempre come virus difettivo indusse H. Temin (v., 1964) a introdurre il concetto di provirus, cui sarà accennato in seguito.
Complessi virali analoghi a quelli aviari e murini sono stati dimostrati anche per i criceti, per i gatti e, forse, per i bovini.
f) Il fattore latte
La scoperta del fattore latte segnò un'importante svolta nelle conoscenze sulla cancerogenesi. Era da tempo nota la relativa frequenza di cancri spontanei della mammella in alcuni allevamenti di topi: dal 1935 L. C. Strong (v.) era riuscito a isolare ceppi inbred, denominati CBH e A, caratterizzati da un'elevatissima incidenza di cancro mammario, pari nel primo al novanta per cento di tutte le femmine, nell'altro al novanta per cento delle femmine che avevano partorito e al cinque per cento di quelle vergini. Successivamente (v. Strong, 1936-1942) furono isolati altri ceppi di topi, CBA, BALB/C, C57 Black, con incidenza di cancro mammario assai bassa o addirittura nulla, e ceppi simili furono isolati anche in altri laboratori. Si ritenne inizialmente che la diversa frequenza della neoplasia, apparentemente ereditaria, fosse in rapporto a fattori genici; tuttavia, l'osservazione che la prole da padre appartenente a un ceppo con alta incidenza e da madre appartenente a un ceppo con bassa incidenza non era affetta da tumore, contrastava fortemente con l'ipotesi dell'eredità cromosomica. Si ammise allora la possibilità di una trasmissione extracromosomica, termine ampio comprendente l'intervento di fattori sia citoplasmatici, sia esterni; e in seguito all'osservazione di J. J. Bittner (v. Some possibile..., e The receptibility..., 1936) che delle topine appartenenti a un ceppo con alta incidenza di tumore mammario, allattate, anzichè dalla propria madre, da una femmina di ceppo a bassa frequenza, soltanto tre su nove presentarono cancro mammario, si pensò che tra questi fattori extracromosomici almeno uno dovesse essere identificato tra i componenti del latte. Seguirono numerose altre ricerche, condotte dallo stesso Bittner, da H. B. Andervont e da un folto gruppo di sperimentatori in vari laboratori su un gran numero di animali.
In particolare i lavori dello Andervont (v., 1941) contribuirono in maniera risolutiva a spiegare il motivo per cui una sia pur esigua percentuale della prole sottratta all'allattamento materno si ammalava, prima o poi, di cancro mammario: egli infatti, in luogo di isolare la madre dai figli entro le prime 24 ore dal parto secondo gli schemi seguiti dai vari ricercatori, procedeva al taglio cesareo delle femmine giunte al termine della gestazione, così da avere l'assoluta certezza che i topini neonati non potessero ingerire la benchè minima quantità di latte materno. In tali condizioni, la incidenza della neoplasia nelle topine si riduceva a zero o al massimo eguagliava quella del ceppo della nutrice. All'agente trasmesso con il latte responsabile dello sviluppo di carcinoma mammario venne dato il nome di fattore latte o fattore di Bittner.
La constatazione che l'incidenza del tumore è molto più elevata nelle topine che hanno partorito che in quelle vergini e di gran lunga maggiore nelle femmine che nei maschi, poneva naturalmente il problema dell'influenza ormonica sullo sviluppo del cancro mammario. Era da tempo noto l'effetto della castrazione sullo sviluppo di tumori della mammella del topo, in riflesso a quanto noto in patologia umana sulla diversa frequenza del cancro mammario nella donna in relazione alle diverse situazioni ormoniche (gravidanza, allattamento, menopausa ecc.); interessante per la oncologia sperimentale era stata inoltre la dimostrazione di A. Lacassagne (v., 1932 e 1939) della possibilità di induzione del cancro mammario in topi maschi trattati a lungo con benzoato di estrone, trattamento che fu poi da altri dimostrato efficace solo in animali appartenenti a ceppi caratterizzati da elevata incidenza di tumore mammario spontaneo nelle femmine. Si vide inoltre che nei maschi di tali ceppi già la castrazione è da sola sufficiente a indurre lo sviluppo di carcinoma mammario, e si pensò che i fattori ormonali agissero aumentando la sensibilità delle cellule della ghiandola mammaria al fattore latte. Riassumendo i suoi studi sull'argomento, Bittner concluse che lo sviluppo di carcinomi mammari nei topi appartenenti a ceppi con alta incidenza della malattia è con ogni probabilità in relazione a tre fattori: a) un fattore latte, trasmesso attraverso l'allattamento; b) un fattore genetico ereditario; c) un fattore ormonale causale.
Secondo la maggior parte degli autori, la natura virale del fattore di Bittner sembra sicuramente dimostrata dalle sue caratteristiche di filtrabilità, di lunga conservabilità in glicerina al 50%, di inattivabilità al calore (61 °C per 30 minuti) e di ultracentrifugabilità, e soprattutto dalle fotografie al microscopio elettronico di sezioni ultrasottili di tessuto del tumore che dimostrano la presenza di particelle virali localizzate nel citoplasma. Il fattore latte, pertanto, viene frequentemente indicato come virus del carcinoma mammario del topo (mouse mammary carcinoma virus) o come MTA.
Si è tentato di trasferire in patologia umana i risultati di questi studi per identificare un possibile fattore responsabile dello sviluppo del carcinoma mammario nella donna: la ricerca di particelle di tipo virale nelle sezioni ultrasottili di questi tumori ha spesso avuto esito negativo, tuttavia occorre ricordare che anche in tumori sperimentali di sicura eziologia virale la microscopia elettronica sovente non consente il reperto del virus.
5. Cenni di biologia molecolare della cancerogenesi
La trasformazione neoplastica della cellula rappresenta l'evento terminale di complesse fasi delle quali alcune sono soltanto deviazioni di vario grado dalla norma, altre invece costituiscono aspetti peculiari dell'abnorme processo biologico. Lo studio di tali alterazioni, che si trasmettono poi nelle successive discendenze cellulari, riguarda essenzialmente i processi metabolici e le caratteristiche strutturali e dinamiche delle molecole che sono alla base del meccanismo della cancerizzazione: il problema entra così nell'ambito della biologia molecolare, cioè della disciplina che studia in generale struttura e funzione delle macromolecole biologiche, in primo luogo degli acidi nucleici cui è essenzialmente devoluta la trasmissione dei caratteri ereditari (v. acidi nucleici; v. biologia molecolare).
La moltiplicazione cellulare è uno dei più complessi fenomeni della vita cellulare, del quale è stato possibile operare uno schematismo fondato su particolari espressioni morfologiche (v. cellula: Fisiologia della cellula); per comodità di studio, essa viene distinta in fasi e le varie attività che vi si svolgono sono designate con il termine cinetica cellulare. A. Howard e S. Pelc (v., 1953) indicarono le varie fasi del ciclo cellulare come G1-S-G2-M, con una ulteriore suddivisione delle G1 e G2 nelle due sottofasi G1 e G2 precoce e G1 e G2 tardiva, ognuna corrispondente a eventi biochimici diversi e caratterizzata da una particolare sensibilità ad agenti esterni, tra i quali carcinogeni chimici o fisici o biologici, e chemioterapici.
La fase M rappresenta la mitosi, cioè l'evento conclusivo e più appariscente dell'intero ciclo, mentre nella fase 5 ha luogo quel complesso fenomeno di programmazione biochimica che è la duplicazione del DNA. Uno dei meccanismi preparatori di questa sintesi, ritenuto indispensabile da molti ricercatori, è quello cosiddetto ‛a ping pong', consistente in una successione alternata di sintesi di RNA e di proteine. Si usa dire che una cellula che si moltiplica è in ciclo, mentre quella che non si moltiplica - potendo permanere in tale situazione stabilmente o temporaneamente - è fuori ciclo. Stabilmente fuori ciclo sono gli elementi cellulari maggiormente differenziati, come ad esempio le cellule nervose, cioè quegli elementi che G. Bizzozero indicò come perenni; temporaneamente fuori ciclo sono quelle cellule labili di Bizzozero, delle mucose o della cute, che dopo essersi divise entrano in stato temporaneo di quiete, e gli elementi stabili come le cellule epatiche, le quali si moltiplicano solo in alcune particolari condizioni. Le cellule fuori ciclo si trovano nella cosiddetta fase Q, di quiete, e in tale stato sono relativamente insensibili ad agenti interni ed esterni tra cui mutageni e carcinogeni di vario tipo, ai quali sono invece sensibilissime le cellule in ciclo: è nota la frequenza notevolmente più elevata di sviluppo di tumori in tessuti a elementi labili in confronto di quelli a elementi stabili. Una differente sensibilità agli agenti carcinogeni caratterizza anche le varie fasi del ciclo cellulare: la maggiore sensibilità sembrerebbe peculiare della fase G1 tardiva, secondo quanto dimostrato dalle ricerche di G. P. Warwick (v., 1971) sulla facilità di trasformazione delle cellule epatiche in rigenerazione durante questa fase.
Le cellule che compongono un tumore sono in parte in ciclo, in parte fuori ciclo, con ritmi che determinano la rapidità di crescita della massa neoplastica : è probabile che, anche in condizioni così lontane dai normali fenomeni di crescita armonizzata, esista una certa regolazione della proliferazione cellulare. Per spiegare le modalità di una tale regolazione, si ammette che l'inizio della sintesi del DNA corrisponda a un segnale che dà l'avvio al meccanismo ‛a ping pong', e analogamente che un segnale ne determini l'arresto: non si conosce ancora, tuttavia, la natura di questi due segnali.
La sintesi del DNA, che come è noto è di tipo semiconservativo, per cui il filamento di DNA risulta costituito da una catena polinucleotidica originaria e da una complementare neosintetizzata (v. acidi nucleici; v. biologia; v. cellula: Fisiologia della cellula), può essere spontanea, stimolata e indotta. La sintesi spontanea è quella che si svolge nella cellula in condizioni normali. La sintesi stimolata è, invece, quella che avviene sotto l'influenza di un agente che inattiva il fattore o i fattori normalmente deputati a inibire la moltiplicazione cellulare, la cui esistenza e per varie ragioni ammessa e che si ritiene siano localizzati verosimilmente a livello della membrana cellulare. L'inibizione esercitata da tali fattori è con ogni probabilità dovuta a blocco della traduzione dell'RNA; la disinibizione attiva l'RNA rendendolo così disponibile a funzionare da template, e avvia in tal modo la catena di eventi che sfociano nella sintesi di DNA e infine nella mitosi. La sintesi stimolata costituirebbe il meccanismo con il quale la cellula fuori ciclo può rientrarvi e quindi riprodursi.
Più complesso, e per alcuni lati ancora molto oscuro, è il meccanismo della terza forma di sintesi, quella indotta, connessa con l'esistenza di un fattore o di un complesso di induzione, identificabile con l'alterazione intervenuta nella continuità della molecola dell'acido nucleico, a opera per esempio di un carcinogeno o di un cocarcinogeno. Come si vede, si tratterebbe di un processo che presenta qualche analogia con il crossing over, nel quale pure intervengono fenomeni di riparazione (v. biologia; v. genetica: Citogenetica).
Comunque, la sintesi indotta si distinguerebbe da quella spontanea fondamentalmente per due motivi: per il segnale di inizio, che avvia la sintesi spontanea e non quella indotta; e per il sito di inizio stesso della sintesi, localizzato, in quella spontanea, a livello di un ben determinato tratto della lunga catena dell'acido nucleico denominato punto di iniziazione, e in un punto qualunque corrispondente al tratto ove si è verificato il danno in quella indotta.
Secondo A. Caputo (v., 1973), la comparsa del segnale è in rapporto alla provenienza dell'energia, identificabile nel caso della sintesi spontanea nelle comuni reazioni esoergoniche che si svolgono in prossimità della membrana cellulare, in quello della sintesi indotta nella rottura dei legami chimici a livello dei tratti danneggiati della molecola. Ciò determinerebbe una diversa disponibilità di energia in dipendenza sia del punto ove è avvenuta la rottura, sia della struttura chimica del carcinogeno.
La sintesi del DNA inizia con lo svolgimento della doppia elica, conseguente alla rottura di legami che tengono unite le due eliche, ognuna delle quali è così disponibile a funzionare da template; la successiva formazione delle due nuove catene polinucleotidiche, resa possibile dalla sintesi di nucleotidi, dà luogo alla costituzione di un nuovo filamento di DNA. Rottura dei legami e sintesi dei nucleotidi avvengono a opera di enzimi specifici, endonucleasi e polimerasi in grado rispettivamente di scindere il legame 3-idrossil-5-fosforil e di operare la formazione di legami fosfo-diesterici tra i desossiribonucleotidi trifosfati.
La rottura di una regione del DNA può avvenire a opera di vari agenti, ma sempre tramite l'intervento di endonucleasi: si possono così formare frammenti di DNA di varia lunghezza, costituiti comunque da almeno cinque mononucleotidi. La riparazione si effettua per l'intervento di polimerasi che promuovono l'incorporazione di mononucleotidi sul template della catena polinucleotidica omologa a quella degenerata, o di ligasi che determinano coniugazione di corte catene di polinucleotidi, sintetizzati da polimerasi, con la molecola di DNA mediante legami fosfodiesterici.
Di notevole interesse è il problema se gli enzimi agenti nella funzione di repair siano preesistenti nella cellula allo stato attivato, ovvero presenti in essa ma inattivi, o vengano invece formati ex novo nel momento della trasformazione cellulare. Per quanto riguarda la trasformazione indotta dai carcinogeni chimici, le cognizioni sono scarse e non univoche; al contrario, per quella determinata dai virus oncogeni moltissimi dati sperimentali sono stati accumulati negli ultimi anni e, come si è accennato a proposito del polioma, si è sicuramente dimostrato che alcuni enzimi sono di nuova sintesi, alcuni altri presenti ma inattivi nella cellula vengono attivati. Inoltre, almeno per quanto riguarda il polioma, gli enzimi di nuova sintesi sono di origine cellulare; in altri casi, invece, come per il virus vaccinico, alcuni di questi enzimi sono certamente codificati dal virus.
Appare evidente che l'infezione da polioma induce nella cellula sintesi di nuove molecole enzimatiche, delle quali alcune operano la rottura delle catene polinucleotidiche cellulari, altre promuovono la sintesi di acido nucleico virale e cellulare con preponderanza dell'uno o dell'altro a seconda rispettivamente del tipo produttivo o abortivo dell'infezione. Un virus è quindi in grado di determinare notevoli variazioni biochimiche nella cellula ospite, sia per effetto di enzimi, in grado di operare la sintesi polinucleotidica, in esso presenti o contenuti come informazione nel suo patrimonio genetico così che la cellula stessa è indotta a produrli, sia in alcuni casi mediante la capacità di promuovere la formazione di eso- ed endonucleasi che staccano polinucleotidi dalla catena del DNA cellulare e operano l'inserimento successivo di porzioni del genoma virale. In conseguenza di questo complesso di interazioni si determina quella deviazione più o meno completa dei processi biosintetici cellulari secondo i modelli strutturali del virus che rappresenta in sostanza l'intima essenza dell'infezione virale.
L'esistenza di virus oncogeni a RNA e di altri a DNA era stata spiegata da Temin (v., 1964), già da vari anni, con l'ipotesi che i primi fossero in grado di indurre sintesi di DNA virale. Tale spiegazione non era in realtà apparsa convincente, soprattutto perché in contrasto con uno dei dogmi della biologia molecolare, secondo il quale è possibile lo stampo dell'RNA sul DNA e non viceversa. Tuttavia, nel 1970 5. Mizutani e Temin (v., 1970) nel virus di Rous, e D. Baltimore (v., 1970) nello stesso virus di Rous e in quello di Rauscher, scoprirono un enzima, la transcriptasi inversa, che determina la trascrizione dell'RNA virale in DNA; la dimostrazione che il prodotto dell'attività di tale enzima è insensibile alla ribonucleasi, ma idrolizzabile dalla desossiribonucleasi, costituisce la prova migliore dell'effettiva sintesi di DNA a partire da RNA come template (stampo).
L'enzima, ora noto anche con il nome di polimerasi RNA-dipendente, è stato purificato e attentamente studiato. È stato così possibile precisarne alcune caratteristiche chimiche e fisicochimiche e accertare che la sua attività si svolge in due fasi successive: dapprima si forma una sola elica di DNA sullo stampo dell'unico filamento di RNA, quindi si determina la formazione di catene ibride RNA/DNA; la catena di DNA funziona poi da template e, a opera di una DNA-polimerasi DNA-dipendente, si forma una doppia elica di DNA (v. acidi nucleici; v. biologia).
Si scoprì inoltre che alcuni polimeri sintetici, ribo- e desossiribonucleotidi, artificialmente uniti possono funzionare da template: S. Spiegelmann e altri (v., 1970) e Mizutani e altri (v., 1970) hanno dimostrato che poliossicitosina/poliriboguanina agiscono da template per la transcriptasi inversa, con un'efficienza molto maggiore degli acidi nucleici naturali.
Le ricerche di Temin, di Baltimore, di Spiegelmann e di Mizutani hanno realizzato una delle più importanti scoperte biologiche degli ultimi anni. Si è, di conseguenza, aperto un vasto campo di indagini intese a dimostrare la presenza di polimerasi RNA-dipendente in tumori umani quale prova, sia pure indiretta, della loro eziologia virale.
Finora, vari ricercatori hanno dimostrato nell'uomo la presenza di entrambe le attività polimerasiche, RNA- e DNA-dipendenti, in cellule leucemiche, ma non in leucociti normali, e la loro scomparsa quando, in seguito a terapia, il quadro ematologico si normalizza. Sembrerebbe pertanto di poter concludere che le leucemie dell'uomo, così come quelle di varie specie animali, sono provocate da virus. Tuttavia non va dimenticato che le due attività polimerasiche sono presenti anche in virus non oncogeni, onde la loro dimostrazione, se costituisce una generica indicazione della presenza di un virus, non è comunque sufficiente a fornire la prova sicura dell'eziologia virale di un tumore o di una leucemia. Le ricerche condotte in numerosi laboratori di oncologia sperimentale hanno consentito di accertare che nei virioni di virus oncogeni a RNA sono presenti, oltre a quella polimerasica, un'attività ribonucleasica che agisce sul substrato costituito dall'ibrido RNA/DNA e idrolizza l'RNA, e attività esonucleasiche ed endonucleasiche. L'interazione di questi diversi enzimi, di importanza essenziale per la replicazione dei virus, è responsabile della formazione di una copia a DNA del genoma virale la quale poi, integratasi in quello cellulare, è in grado sia di dirigere la sintesi di nuove particelle virali, sia di causare la trasformazione maligna della cellula. I dati sperimentali sembrano confermare l'ipotesi che tra acido nucleico della cellula ospite e acido nucleico virale si formino ibridi, in modo analogo a quanto avviene nel caso dei fagi, generalmente in seguito all'unione solo di quei tratti dei due filamenti nei quali vi è complementarità delle basi.
Le numerose ricerche sulle caratteristiche di queste corrispondenze complementari tendono attualmente a dimostrare affinità tra acido nucleico di cellule umane e quello di virus a DNA e a RNA maggiormente indiziati come probabili agenti eziologici di tumori.
La formazione di molecole ibride di acido nucleico virale e cellulare può spiegare le modalità della trasmissione, in vivo e in vitro, della trasformazione operata da virus oncogeni nelle successive generazioni cellulari, cioè della trasmissione del virus o di parte di esso in senso verticale. R. J. Huebner e G. J. Todaro (v., 1969) hanno formulato la cosiddetta teoria sull'‛oncogene', quella parte cioè del genoma virale che, stabilmente integrato in quello delle cellule, viene trasmesso di generazione in generazione. Il dato sperimentale più importante sul quale si fonda tale teoria è la dimostrazione che alcune linee cellulari, come la 3T3 o la 3T12, dopo molte generazioni e dopo che le cellule sono divenute aneuploidi, producono in vitro virus del tipo della leucemia del topo (MLV): ciò si può spiegare ammettendo che il virus in fase intracellulare si trovi in forma inespressa e che particolari condizioni, come il frequente trasferimento su nuovo terreno o l'alta densità di cellule, ne favoriscano l'espressione, cioè la formazione di particelle virali complete.
Si è inoltre osservata produzione di virus, specialmente della leucemia, in topi sottoposti all'azione di radiazioni o di carcinogeni chimici, che quindi non determinerebbero la diretta trasformazione neoplastica delle cellule, ma attiverebbero l'oncogene inducendo nella cellula un profondo disordine e la conseguente perdita della normale capacità di regolazione.
A modifica della teoria ora esposta il Temin (v., 1971) ha introdotto la teoria del ‛protovirus', secondo la quale non esisterebbe l'oncogene nelle cellule germinali, ma si determinerebbe nel corso dello sviluppo; attraverso il passaggio RNA → DNA l'informazione verrebbe poi inserita nel genoma cellulare con possibilità di indurre formazione di particelle complete a RNA. Tale processo, però, dovrebbe comportare la comparsa di antigeni virali durante lo sviluppo tardivo e non, come in realtà accade, già durante la vita embrionale; la precocità di comparsa di antigeni virali e la conseguente tolleranza immunitaria svelata dall'assenza negli animali di anticorpi specifici contro tali antigeni, costituiscono invece un valido sostegno della teoria dell'oncogene.
È opportuno ricordare il significato di alcuni termini usati in virologia per definire situazioni tra loro distinte: come provirus si indica qualsiasi informazione allo stato potenziale; come ‛virogene' si designa un provirus naturale, costituito dall'insieme di geni che codificano l'intera particella virale; infine, come abbiamo visto, il termine di ‛oncogene' si riferisce a quel particolare provirus responsabile della trasformazione cellulare, rappresentato da un gene o da un gruppo di geni nei quali è contenuta la relativa informazione.
Conseguentemente all'integrazione di parte del genoma virale con quello della cellula, in determinate condizioni si verifica il fenomeno della trasformazione cellulare, che può o meno essere accompagnata dalla replica di particelle virali; in alcuni casi, invece, la suddetta integrazione è seguita dalla sola replica di particelle virali, mentre manca il processo di trasformazione.
È molto probabile che diversi fattori siano in grado di determinare una rottura della o delle catene di polinucleotidi dell'acido nucleico cellulare; esiste allora la possibilità che a tale rottura segua, a mezzo delle polimerasi e ligasi, la riparazione del danno (fenomeno del repair) con ristabilimento della normale struttura o di parte di essa o, nel caso che l'agente lesivo sia un virus, la sostituzione con frammenti di catene polinucleotidiche virali. Se il repair avviene secondo il programma normale, non si determinano variazioni geniche; in caso contrario, si stabiliscono mutilazioni del codice genetico cellulare o, come ad esempio quando avviene l'integrazione con parte di quello virale, ha luogo il fenomeno della trasformazione.
Il momento del repair è quindi, secondo molti studiosi, quello che decide il destino della cellula per quanto riguarda la sua possibile cancerizzazione.
L'azione dei cancerogeni si esplicherebbe in un primo tempo determinando il danno, secondariamente impedendo la riparazione o determinando saldature con polinucleotidi fuori programma; questa seconda modalità d'azione sarebbe quella prevalentemente esercitata dai cocancerogeni.
Il fatto che solo alcune delle cellule sottoposte all'azione di carcinogeni diventano cancerose esprimerebbe la maggior frequenza della riparazione secondo programma rispetto a quella anomala fuori programma.
6. Metabolismo energetico della cellula neoplastica
Nel 1910 O. Warburg (v., 1930, p. 35) poté dimostrare che la moltiplicazione cellulare implica un'attivazione della respirazione: egli infatti osservò che nelle uova di riccio di mare, dopo la fecondazione, aumenta il consumo di ossigeno rapportato all'unità di peso. In seguito a questa osservazione, si suppose che anche le cellule neoplastiche, data l'elevata moltiplicazione cellulare propria di un gran numero di tumori, fossero caratterizzate da un aumento degli scambi respiratori.
Tale ipotesi fu però smentita dallo stesso Warburg, che proseguendo nei suoi studi vide che con una discreta frequenza la maggiore richiesta di energia è soddisfatta dalla fermentazione del glucosio, con conseguente produzione di acido lattico: a tale fenomeno egli diede il nome di glicolisi, distinguendone una forma anaerobica che si svolge in ambiente privo di ossigeno e una aerobica che si svolge in presenza di ossigeno. Nel processo di glicolisi anaerobica si determina un accumulo di acido lattico, perché, conseguentemente alla mancanza di ossigeno, si arresta l'ossidazione dell'acido piruvico: in condizioni di anaerobiosi, pertanto, ogni tessuto normale glicolizza. La glicolisi aerobica, invece, ha luogo quando la cellula non è capace di ossidare tutto l'acido piruvico che produce, una parte del quale si trasforma quindi in acido lattico. La determinazione dei due tipi di glicolisi, che si esegue facendo metabolizzare il materiale biologico in ambiente di azoto e di ossigeno rispettivamente per la glicolisi anaerobica e per l'aerobica, consente di valutare la quantità di acido lattico prodotta per ora e per milligrammo di tessuto secco: questi valori vengono espressi rispettivamente con le sigle
N2 O2
Q e Q
CO2 CO2
ove Q è la quantità di acido lattico prodotto desunta dal volume di CO2 sviluppato in presenza di bicarbonato.
Warburg (v., 1925, 1926, 1943 e 1955) poté dimostrare che molti tessuti normali, specialmente quelli in vivace proliferazione, e molti tumori in rapido accrescimento svolgono un'intensa attività glicolitica in condizioni di anaerobiosi, mentre in aerobiosi la funzione glicolitica è frequente in tessuti neoplastici e soppressa a favore dell'ossidazione in quelli normali. La conferma della scoperta di Warburg provenne soprattutto dalla dimostrazione della possibilità per le cellule neoplastiche di sopravvivere in ambiente privo di ossigeno ma in presenza di glucosio (v. Warburg, 1930).
Sin dalle prime ricerche Warburg ritenne che le cellule neoplastiche avessero la normale respirazione danneggiata e fossero pertanto obbligate a operare la fermentazione del glucosio per integrare il fabbisogno energetico deficitario. Egli inoltre interpretò in vario modo la glicolisi aerobica osservata nei tessuti non neoplastici: quella retinica quale espressione del danneggiamento subito dal tessuto durante la preparazione, quella dei tessuti embrionali come conseguenza di maggior fabbisogno energetico nel corso della moltiplicazione cellulare. Grazie ai miglioramenti tecnici razionalmente usati e perfezionati fino agli ultimi anni della sua vita, egli vide che i valori della glicolisi aerobica della retina erano notevolmente inferiori a quelli inizialmente trovati e che il metabolismo respiratorio e glicolitico delle cellule embrionali è diverso da quello delle cellule neoplastiche.
All'iniziale entusiasmo per queste ricerche subentrò un periodo di critiche demolitrici, specialmente a opera di S. Weinhouse (v., 1951 e 1955) e collaboratori (v. Weinhouse e altri, 1950 e 1951) in aspra polemica con Warburg e i suoi allievi; tra questi ultimi, D. Burk (v., 1939) riuscì a operare un confronto tra respirazione e fermentazione di cellule di fegato normale, di fegato rigenerante e di fegato cancerizzato con o-amminoazotoluene, e concluse che nelle cellule tumorali i due processi sono peculiari.
Fu rilevato che negli epatomi a lenta crescita (a deviazione minima) di Morris, nel ratto, non si notava né riduzione della respirazione cellulare né comparsa di glicolisi aerobica: questa è, forse, la critica meno valevole all'ipotesi di Warburg, in quanto tali epatomi, appunto perché a deviazione minima, sono poco diversi dal tessuto epatico normale, certamente non comparabili a neoplasie in attivo sviluppo.
Allo stato attuale delle conoscenze, anche se appare scarsamente sostenibile con valide dimostrazioni il concetto di Warburg di un danneggiamento respiratorio delle cellule neoplastiche, è tuttavia certo che queste sono caratterizzate da un'elevata glicolisi aerobica e che con grande frequenza mostrano una netta riduzione dei mitocondri, cioè degli organuli sede della catena delle ossidazioni cellulari (v. cellula: Fisiologia della cellula).
L'esistenza di glicolisi aerobica nella retina costituisce tuttora un problema non risolto.
Gli studi sul metabolismo delle cellule neoplastiche hanno messo in evidenza fenomeni di un certo rilievo: tra questi il cosiddetto effetto Crabtree (v. Crabtree, 1959), cioè l'abbassamento dei valori della respirazione per aggiunta di glucosio al mezzo di sospensione di cellule neoplastiche, che non si determina per quelle normali, e la riduzione dell'effetto Pasteur, cioè della fermentazione in presenza di ossigeno, il cui meccanismo è in realtà tuttora poco chiaro onde è impossibile trarne un sicuro significato fisiopatologico.
Negli epatomi di Morris sono state osservate alterazioni di alcuni enzimi in rapporto al grado di malignità. Sono state poste in evidenza alterazioni della produzione di enzimi a funzione limitante a livello di punti chiave della demolizione del glucosio, cioè dell'esochinasi, della fosfofruttochinasi e della piruvatochinasi, con comparsa di isoenzimi di tipo fetale; la sintesi ditali enzimi è repressa durante la vita cellulare normale, ma viene derepressa nella trasformazione neoplastica.
7. Cenni di immunologia oncologica
I brillanti successi ottenuti con l'impiego di sieri e vaccini nella terapia e nella profilassi delle malattie infettive fecero nascere la speranza che sussidi di tale tipo potessero essere utilizzati anche per la prevenzione e la cura dei tumori. Ciò naturalmente implicava l'esistenza e l'individuazione di costituenti antigenici specifici della cellula neoplastica: tale presupposto teorico, che dette luogo a primi approcci sperimentali i cui risultati debbono oggi essere valutati con la massima cautela, ottenne in realtà una valida conferma soltanto quando si osservò il rigetto di isotrapianti neoplastici da parte di animali ospiti singenici. Si raggiunse pertanto la certezza dell'esistenza di antigeni neoplastici specifici, e fu così possibile superare la difficoltà delle prime ricerche sperimentali: stabilire cioè se la risposta immunitaria in un animale in seguito al trapianto di un tumore sia suscitata da antigeni specifici del tumore ovvero da isoantigeni presenti nella massa di tessuto trapiantato.
La comparsa di antigeni specifici tumorali rappresenta un evento importante della cancerogenesi, ma non indi- spensabile ai fini della trasformazione neoplastica: non mancano, difatti, esempi di tumori dotati di scarsa antigenicità e anche di tumori nei quali si è verificata una delezione dei normali costituenti antigenici senza sostituzione con antigeni di nuova sintesi. Comunque, la maggior parte dei tumori sperimentali possiede antigeni specifici, ma non è ancora possibile comprendere l'importanza dell'antigenicità della cellula neoplastica nei riguardi dell'organismo ospite e della sua capacità di risposta immunitaria in relazione sia alla genesi, sia all'ulteriore controllo dello sviluppo del tumore.
Gli antigeni individuati nei tumori e nelle leucemie sperimentali sono distinti in base alla loro associazione con virus oncogeni a RNA o a DNA, alla loro solubilità, alla loro localizzazione. In particolare, per quanto riguarda quest'ultimo carattere è importante la distinzione in antigeni superficiali, ovviamente più esposti e a più immediato contatto con le cellule immunitarie, e antigeni intracellulari in grado di dar luogo a una risposta immunitaria in seguito a fenomeni di citolisi.
Si ritiene che gli anticorpi diretti contro tali ultimi antigeni difficilmente possano esercitare un proficuo sistema di controllo della neoplasia, anche se la loro eventuale di- mostrazione potrebbe essere utilizzabile in clinica a scopo diagnostico.
Occorre tuttavia ricordare che alcuni di quegli antigeni tumorali che definiamo specifici compaiono in realtà nelle cellule anche nelle malattie non neoplastiche e talora, sia pure transitoriamente, anche in condizioni fisiologiche in alcune fasi del ciclo cellulare: prescindendo da più complesse considerazioni sulla genesi dei nuovi costituenti antigenici, si comprende la necessità di un'attenta valutazione ai fini diagnostici della dimostrazione di un antigene, soprattutto in rapporto alla presenza di altri segni probativi per l'esistenza di una neoplasia. La dimostrazione, ad esempio, di anticorpi anti EBV (Epstein Barr Virus) è comunque di notevole interesse diagnostico, ma assume significato diverso nel caso della mononucleosi infettiva e in quello del linfoma di Burkitt o del carcinoma nasofaringeo.
Vi è una differenza di ordine generale tra i tumori indotti nell'animale con cancerogeni chimici e quelli indotti da virus oncogeni. I primi hanno infatti una propria caratteristica individualità antigenica, tipica di ogni singolo tumore: se nello stesso animale si inducono, con lo stesso agente chimico, diversi tumori in varie zone dell'organismo, ogni tumore sarà fornito di almeno un costituente antigenico individuale, e conseguentemente la reattività crociata sarà assente o notevolmente rara. Al contrario, i tumori da virus posseggono antigeni tra di loro correlati o identici sierologicamente, dei quali è difficile definire la natura e l'origine, anche se numerose osservazioni sperimentali sono riuscite a metterne in evidenza in alcuni casi la provenienza dal genoma virale, in altri quella dalla cellula ospite.
Le acquisizioni suddette sono di estremo interesse per la comprensione di alcuni fenomeni generali della cancerogenesi. Tralasciando altre considerazioni di ordine più strettamente immunologico, basterà qui ricordare che nel caso di tumori indotti da un carcinogeno chimico la diversità degli antigeni dimostrabili indica chiaramente da un lato che il carcinogeno di per sè non può essere l'antigene, dall'altro il disordine genico della cellula trasformata: i vari antigeni singolarmente distinti di uno stesso tumore, infatti, rappresentano con molta probabilità prodotti di geni alterati o anche di geni normali che, repressi nelle cellule differenziate, sono resi liberi dalla trasformazione maligna.
Altri interessanti aspetti del problema dell'immunologia dei tumori riguardano i fenomeni di resistenza e di tolleranza: la dimostrazione, ad esempio, della possibilità di ottenere immunizzazione attiva con vari metodi contro tumori indotti da agenti chimici o da virus, e la constatazione dell'esistenza di condizioni di tolleranza immunologica fino alla completa soppressione della risposta immune, aprono interessanti campi di indagine anche nei confronti di possibili applicazioni pratiche.
Si è accennato alla possibilità che alcuni antigeni di tumori indotti da carcinogeni chimici rappresentino l'espressione fenotipica di geni normali repressi nelle cellule differenziate. In alcuni casi tali antigeni appartengono al gruppo di quelli embrionali, presenti nella cellula durante il periodo della vita embrionale, e la loro biosintesi, come si è detto, è in rapporto a fenomeni di derepressione che hanno luogo nella trasformazione maligna. Sono oggi ben conosciuti, e svelabili nell'uomo con indubbio valore diagnostico, antigeni CEA (Carcino-Embryonic Antigen) dei tumori dello stomaco e dell'intestino, e una α-fetoproteina dei tumori del fegato (v. immunologia e immunopatologia: Malattie immunoproliferative).
Tuttavia, i numerosi studi condotti nel tentativo di dimostrare che i tumori umani posseggono antigeni specifici similmente a quanto si osserva nelle neoplasie sperimentali degli animali, non sono stati finora coronati da successo.
8. Ormoni e cancro
La ben nota cognizione, derivata da numerose osservazioni cliniche, dell'esistenza di tumori ormonodipendenti, in modo particolare il cancro della mammella, ha indotto gli studiosi a ricercare sperimentalmente i rapporti tra alcune neoplasie e determinate attività endocrine. Interessanti dati sperimentali si sono andati così accumulando fino a costituire prove certe e incontrovertibili della stretta relazione esistente tra complesse interazioni endocrine e sviluppo di tumori. Basterà qui ricordare che M. S. Biskind e G. S. Biskind (v., 1944) nel ratto, H. A. Bali e J. Furth (v., 1949) e W. V. Gardaer (v., 1955) nel topo osservarono lo sviluppo di tumori in ovaie escisse e successivamente innestate nella milza dello stesso animale: per spiegare ciò si è pensato che gli ormoni ovarici che continuano a formarsi negli innesti intrasplenici vengano direttamente immessi nel fegato ove sono inattivati, così da non essere più in grado di esercitare la normale azione di blocco a livello ipofisario; conseguentemente si determina un'iperproduzione di gonadotropine ipofisarie che agiscono sul tessuto ovarico in sede ectopica, determinandone l'iperplasia e la successiva cancerizzazione. Probabilmente con analogo meccanismo, cioè provocando una prolungata iperproduzione di tireotropina ipofisaria, agiscono i vari metodi impiegati per produrre sperimentalmente carcinomi della tiroide; più recentemente, D. Sinha e altri (v., 1965) sono riusciti a indurre un cancro della tiroide mediante somministrazione di tireotropina.
Tra tutti i tumori ormonodipendenti, il cancro mammario è senza dubbio quello più studiato e per il quale sono stati raccolti in maggior numero interessanti rilievi clinici e sperimentali. A. E. C. Lathrop e L. Loeb (v., 1916) dimostrarono, per es., che la castrazione di topine giovani induce una notevole diminuzione della frequenza del cancro mammario spontaneo in ceppi di topi caratterizzati dalla elevata frequenza di tale tumore. Successivamente si osservò che l'inoculazione di ormone ovarico, in dosi elevate e protratta nel tempo, è in grado di determinare un considerevole aumento dell'incidenza del cancro nelle topine nonché la sua comparsa anche in alcuni maschi nei quali la neoplasia spontanea è praticamente inesistente. Già si è fatto cenno come fosse apparso chiaro che anche l'azione del fattore latte è strettamente dipendente, oltre che da condizioni genetiche, dalla presenza degli ormoni ovarici.
Un più accurato studio del problema fu reso possibile a partire dagli anni trenta, quando si riuscì a ottenere in forma pura gli estrogeni. Si ritenne inizialmente che la molecola dell'estradiolo, di gran lunga il più attivo tra gli ormoni ovarici e dal punto di vista chimico assai simile agli idrocarburi, a seguito di particolari trasformazioni operasse come agente carcinogeno: tuttavia non si è mai riusciti a dimostrare la presenza nei tessuti di enzimi capaci di in- durre tale conversione molecolare. Si è inoltre osservato che l'equilenina, il più insaturo degli estrogeni e quindi teoricamente il più probabilmente trasformabile in idrocarburi cancerogeni, è assai meno attiva nella cancerizzazione, mentre sostanze sintetiche chimicamente molto lontane dagli ormoni ovarici, come lo stilbestrolo, agiscono come gli estrogeni steroidei. Per tali ragioni l'ipotesi della formazione di idrocarburi aromatici in seno ai tessuti a opera di enzimi è apparsa sempre meno probabile.
Molti autori hanno supposto che gli estrogeni agirebbero non inducendo direttamente la trasformazione della cellula della ghiandola mammaria, bensì sensibilizzandola all'azione di quei fattori che sono i veri responsabili della cancerizzazione. È stato infatti dimostrato che gli estrogeni sono in grado di determinare iperplasia e trasformazione cistica della mammella, condizioni nelle quali per intervento di vari fattori locali la comparsa di un cancro è assai probabile.
Gli studi dei rapporti tra neoplasie mammarie e ormoni hanno consentito di distinguere determinate categorie sperimentali, corrispondenti a vari stadi di sviluppo del tumore. La prima categoria e il primo stadio, della dipendenza, esprimono una condizione in cui la proliferazione neoplastica avviene solo in presenza di un alto livello di ormone: tale è il caso di un tumore che si sviluppa in una topina vergine non appena diventi gravida, quindi regredisce e scompare, per ricomparire poi a una nuova gravidanza, e così successivamente. La seconda categoria, della responsività, corrisponde a uno stadio di proliferazione accelerata in seguito a somministrazione di alte dosi di ormone, ma che, cessata questa, prosegue, sia pure a ritmo più lento. La terza categoria, dell'indipendenza, corrisponde all'assenza di sensibilità alla somministrazione di ormone. In termini rigorosamente corretti, quando si parla di condizione si deve intendere il carattere permanente del tumore nei riguardi della dipendenza ormonale, mentre l'espressione stadio indica un mutevole e successivo svolgersi di eventi di una neoplasia che può iniziare come ormonodipendente, per diventare poi ormonoindipendente, passando per lo stato di responsività. I tumori della mammella della donna possono appartenere a una delle tre categorie.
La recente acquisizione sperimentale della possibilità di studiare i rapporti tra estrogeni e cancro mammario oltre che nei topi anche nei ratti, nei quali a differenza dei primi non si conosce un virus in qualche modo correlato allo sviluppo della neoplasia, ha consentito di semplificare notevolmente il problema; attualmente si conoscono almeno cinque ceppi di ratti nei quali è possibile indurre con estrogeni formazione di tumori della mammella. Interessanti osservazioni sono state anche condotte sui possibili rapporti tra carcinogeni chimici ed estrogeni. È stato dimostrato che il 3-metilcolantrene e il 7,1 2-dimetilbenzantracene, ad esempio, somministrati per via alimentare, in parte passano nelle feci senza subire modificazioni, in parte vengono assorbiti e metabolizzati o depositati nei tessuti, particolarmente nel tessuto adiposo. La persistenza di tali sostanze nei vari organi, fegato, polmone, reni, tubo gastroenterico, è limitata dalla presenza in essi di un'idrolasi per il benzopirene, la cui attività è stimolata proprio da alcuni idrocarburi cancerogeni; la mammella è invece priva di questo enzima, così che l'accumulo nel suo tessuto adiposo di carcinogeni assunti con l'alimentazione e non metabolizzati può rappresentare la ragione per cui è indotto lo sviluppo del cancro mammario (v. Dao, 1964). La possibilità di provocare in tal modo la neoplasia non è modificata in femmine di ratto castrate immediatamente prima della somministrazione del carcinogeno, nelle quali è quindi ancora circolante una sufficiente quantità di estrogeni; tuttavia, se la somministrazione del carcinogeno segue di trenta o più giorni la castrazione, il tumore non si sviluppa. Questi rilievi inducono a ritenere che il ruolo degli estrogeni nell'induzione del cancro mammario nel ratto è essenziale. D'altro canto, è stato sperimentalmente dimostrato che la somministrazione di 17β-estradiolo sopprime l'inibizione della sintesi dell'RNA, e probabilmente delle proteine, determinata come effetto iniziale da un idrocarburo cancerogeno a livello delle cellule della ghiandola mammaria, le quali possono così proliferare.
La stretta dipendenza dello sviluppo del cancro mammario da idrocarburi da complesse situazioni endocrine è documentata da numerosi dati: la neoplasia cresce più rapidamente durante la gravidanza, regredisce dopo il parto, riprende a svilupparsi nel corso di una successiva gravidanza; la sua crescita è notevolmente influenzata dall'attività dell'ipofisi, delle ovaie e dei surreni, e la rimozione ditali ghiandole ne determina una rapida regressione; un tumore in regressione in un animale ovariectomizzato riprende a crescere in seguito alla somministrazione di ormoni; un tumore regredito per effetto di ovariectomia e ipofisectomia riprende a svilupparsi se nell'animale si innesta un tumore mammotrofico, cioè secernente prolattina, ormone della crescita e forse ACTH.
L'analisi dei meccanismi con i quali i vari ormoni influenzano lo sviluppo del cancro mammario nel topo ha compiuto negli ultimi anni notevoli progressi.
Si deve a E. V. Jensen e H. I. Jacobson (v., 1962) l'importante osservazione che alcuni organi, come utero e vagina, hanno la capacità di concentrare e trattenere il 17β- estradiolo marcato; in seguito è stato dimostrato che tale caratteristica è comune a tutti gli organi cosiddetti bersaglio, tra cui principalmente la ghiandola mammaria. Dopo che F. Bresciani e G. A. Puca (v., 1965) e Bresciani e altri (v., 1967, 1969 e 1973) hanno identificato le proteine che legano il 17β-estradiolo, alle quali pertanto compete la funzione di recettori, i cancri della mammella sia del topo sia della donna possono essere distinti in due categorie: quelli con recettori e quelli senza recettori per l'ormone. È stato dimostrato che l'estrogeno reagisce con i recettori proteici del citoplasma; qui un enzima proteolitico, attivabile dal calcio, scinde il frammento del recettore al quale è fissato l'ormone, questo quindi passa nel nucleo e si lega a una proteina basica della cromatina. È probabile che in tal modo sia indotta un'attivazione di geni, e quindi sintesi di RNA e conseguentemente di proteine, cioè il primo evento della moltiplicazione cellulare.
Nella donna è stato possibile dimostrare l'esistenza di una stretta relazione tra cancri privi di recettori e insensibilità alla terapia ormonica, mentre notevolmente variabile è apparsa la risposta a tale trattamento dei cancri provvisti di recettori.
9. Conclusioni
I progressi realizzati nel corso degli ultimi anni dall'oncologia sperimentale, esposti sinteticamente in questo articolo, hanno consentito importanti acquisizioni su molti processi biologici abnormi, anche se la conoscenza dei meccanismi che sono alla base del loro svolgersi è ancora parziale.
È stato sicuramente accertato che agenti di natura diversa, chimica, fisica, biologica, sono in grado di determinare l'insorgenza del cancro in animali da esperimento. Alcuni di tali agenti sono responsabili dell'insorgenza di determinati tipi di cancro anche nell'uomo: tale è il caso degli idrocarburi cancerogeni, all'azione di alcuni dei quali è da ascrivere il cancro dei lavoratori del catrame; dei composti di anilina, che provocano l'insorgenza di tumori della vescica; delle radiazioni ultraviolette, causa di epiteliomi cutanei in persone esposte a lungo alle radiazioni solari; dei raggi Röntgen e delle radiazioni di corpi radioattivi, inducenti tumori del tutto comparabili a quelli che con essi si producono sperimentalmente negli animali.
In tal modo, l'oncologia sperimentale ha chiarito, attraverso un'enorme massa di ricerche, l'eziologia di alcuni tumori dell'uomo, quelli cosiddetti professionali; essa è stata favorita in questa ricerca dalla possibilità di identificare in determinati fattori ambientali presumibili agenti carcinogenetici, grazie all'evidenza di una relazione causa- effetto non riscontrabile nella maggior parte dei tumori che colpiscono apparentemente a caso l'individuo o l'organo.
Diversa è la situazione per quanto concerne i tumori da virus. Come si è già detto, numerosi virus sono stati definitivamente riconosciuti quali agenti eziologici di vari tumori degli animali: il virus di Rous del sarcoma del pollo, i virus della leucemia del pollo, alcuni dei quali correlati con il virus di Rous, i virus del complesso sarcoma-leucemia del topo, il fattore di Bittner del cancro mammario del topo, il virus polioma del tumore della parotide e di vari altri organi nel topo, e così molti altri virus di tumori o leucemie spontanee di altre specie animali. La dimostrazione della contagiosità per tutti questi virus è mancata o è ancora dubbia, con la sola eccezione del virus di Lucké dell'adenocarcinoma del rene di rana.
Soprattutto le ricerche sui virus oncogeni hanno contribuito in notevole misura allo studio del meccanismo o dei meccanismi attivi nel trasformare una cellula normale in cellula tumorale. Infatti, poiché si conosce con sufficiente approssimazione la capacità di virus oncogeni molto piccoli di codificare un numero limitato di proteine, è possibile distinguere nella cellula trasformata quanto è da riferire ad attività propria della cellula e quanto, invece, a quella del virus infettante.
Diversa è, invece, la situazione riguardo all'ansiosa domanda se l'eziologia di alcuni o di molti o di tutti i tumori umani sia di natura virale. Le numerose ricerche finora eseguite sia per scoprire al microscopio elettronico forme virali, sia per isolare in coltura virus provenienti da tessuti umani, non hanno finora condotto a risultati probativi; si spiega pertanto lo scarso credito che vari studiosi concedono all'ipotesi dell'eziologia virale del cancro dell'uomo, in considerazione anche dell'insorgenza imprevedibile e del tutto casuale della malattia per la quale non è dimostrabile la contagiosità propria dei processi morbosi sostenuti da un agente infettivo.
Peraltro, contro tale scetticismo fa riscontro la convinzione di molti ricercatori, il cui numero è sensibilmente aumentato negli ultimi anni, dell'origine virale del cancro dell'uomo, probabilmente di tutte le forme di cancro.
L'eziologia virale di due tumori dell'uomo, la verruca e il condiloma acuminato, è stata sicuramente dimostrata dalla possibilità della trasmissione diretta delle malattie in volontari - in genere gli stessi ricercatori. È vero che verruca e condiloma non sono tumori maligni e possono regredire spontaneamente, tuttavia non va dimenticato che alcuni papillomi di animali, come quello di Shope, spontaneamente o in risposta ad alcuni stimoli si trasformano in cancro; che per azione di idrocarburi cancerogeni si sviluppano papillomi, i quali successivamente si trasformano in carcinomi; infine che, con relativa frequenza, papillomi di alcuni organi dell'uomo, come quello della vescica, si trasformano in carcinomi.
Si tratta pur sempre, in ogni caso, di ragionamenti analogici, contrastati dalla mancanza di dimostrazioni dirette.
Tuttavia, la recente acquisizione che il tumore di Burkitt è dovuto al virus di Epstein e Barr, il quale è anche l'agente eziologico della mononucleosi infettiva, ha richiamato l'attenzione degli studiosi su alcuni importanti aspetti del problema: infatti, non si riesce a porre sicuramente in evidenza in sezioni ultrasottili alla microscopia elettronica il virus di Epstein-Barr né a coltivarlo, e la sua esistenza è ammessa in base a criteri immunitari. Questi sono, è vero, criteri indiretti, ma di tale enorme specificità da non lasciar dubbi sulla relazione tra il virus che costituisce l'antigene e gli anticorpi presenti nel sangue degli uomini a eccezione di quelli affetti da morbo di Burkitt e da mononucleosi infettiva nelle loro fasi iniziali. Non sembrerebbe, dunque, indispensabile per sostenere l'origine virale di un tumore dimostrare la presenza di particelle virali nell'interno delle sue cellule né isolare il virus in coltura.
D'altra parte, le ricerche sui tumori sperimentali da virus hanno messo in evidenza che solo di alcuni di essi è possibile la dimostrazione al microscopio elettronico o l'isolamento nelle colture di cellule; invece altri virus, il cui genoma è intimamente legato a quello della cellula ospite, non si riproducono più come tali, ma integrati nella cellula ne operano la trasformazione maligna, cosicché l'acido nucleico virale viene trasmesso alle cellule figlie e si perpetua il carattere della malignità.
Ancora a sostegno della natura virale dei tumori dell'uomo sta la constatazione che cellule umane normali sono trasformabili in vitro in cellule neoplastiche a opera di virus di origine umana - Adenovirus in particolare - o di virus di origine animale. D'altra parte, sembra difficile pensare che mentre tante specie animali sono spontaneamente affette da tumori di natura virale, l'uomo ne sarebbe del tutto esente. A tale proposito occorre anche considerare che l'esistenza di alcuni virus in grado di indurre neoplasie, quale ad esempio quello del polioma, fu documentata grazie a particolari tecniche di ricerca in feti o in animali appena nati, alle quali fecero poi seguito scoperte di fondamentale importanza che hanno determinato un nuovo corso nelle ricerche di oncologia sperimentale.
Naturalmente, le considerazioni esposte, e varie altre che si omettono, hanno valore relativo, ma allo stato attuale è impossibile non tenerne debito conto.
Il problema, già di per sé tanto complesso, si complica ulteriormente se si tenta di esaminare la relazione tra agenti chimici e agenti fisici da una parte e virus dall'altra. Esistono realmente tanti agenti cancerogeni con peculiari modalità di azione quanti sono quelli che si conoscono, o piuttosto, come si è già accennato, tutti hanno a bersaglio lo stesso o gli stessi punti del metabolismo cellulare? La possibilità di indurre lo sviluppo di tumori con agenti chimici o fisici è completamente indipendente dalla presenza di virus oncogeni, ovvero, come molti credono in base a convincenti ricerche, soltanto questi debbono essere considerati i veri effettori della cancerizzazione mentre gli altri agenti cancerogeni determinerebbero l'attivazione di un virus oncogeno per cosi dire latente?
È questo non solo uno dei maggiori problemi dell'attuale oncologia sperimentale, per motivi di ordine strettamente teorico e speculativo, ma anche di grande interesse umano, perché la conoscenza del tipo di molecole, ad esempio virali, interagenti con quelle specifiche della cellula ospite, è presumibile che possa rendere più razionale ed efficace la terapia dei tumori.
bibliografia
Abercrombie, M., Behaviour of normal and malignant connective tissue cells in vitro, in ‟Canadian cancer conference", 1961, IV, pp. 101-118.
Abercrombie, M., Contact-dependent behavior of normal cells and the possible significance of surface changes in virus-induced transformation, in ‟Cold Spring Harbor symposia on quantitative biology", 1962, XXVII, pp. 427-431.
Alwens, W., Banke, E. E., Jonas, W., Auffallende Häufigkeit von Bronchialkrebs bei Arbeitern der chemischen Industrie, in ‟Münchener medizinische Wochenschrift", 1936, LXXXVII, pp. 485-487.
Andervont, H. B., Spontaneous tumors in a sublime of strain C3H mice, in ‟Journal of the National Cancer Institute", 1941, I, pp. 737-744.
Andervont, H. B., Effect of ingestion of strain C3H milk in the production of mammary tumors in strain C3H mice of different ages, in ‟Journal of the National Cancer Institute", 1941, II, pp. 13-16.
Andervont, H. B., Note on the transfer of the strain C3H milk influence through successive generations of strain C mice, in ‟Journal of the national Cancer Institute", 1941, II, pp. 307-308.
Andrewes, C. H., The transmission of fowl-tumors to pheasants, in ‟Journal of pathology and bacteriology", 1932, XXXV, pp. 407-413.
Bali, H. A., Furth, J., Morphological and biological characteristics of X-ray induced transplantable ovarian tumors, in ‟Cancer research", 1949, IX, pp. 449-472.
Baltimore, D., Viral RNA-dependent DNA polymerase, in ‟Nature", 1970, CCXXVI, pp. 1209-1211.
Bayon, H., Epithelial proliferation induced by the injection of gasworks tar, in ‟Lancet", 1912, II, p. 1579.
Bell, T. M., Massie, A., Ross, M. G. R., Williams, M. C., Isolation of a reovirus from a case of Burkitt's lymphoma, in ‟British medical journal", 1964, I, pp. 1212-1213.
Berenblum, I., The cocarcinogenic action of croton resin, in ‟Cancer research", 1941, I, pp. 44-48.
Berenblum, I., A re-evaluation of the concept of cocarcinogenesis, in ‟Progress in experimental tumor research", 1969, XI, pp. 21-30.
Berenblum, I., Cividalli, G., Trainin, N., Hodes, M. E., Some properties of RLP. A factor from sheep capable of inhibiting radiation leukemogenesis in mice, in ‟Blood", 1965, XXVI, pp. 8-19.
Bernard, W., Febure, H. L., Cramer, R., Mise en evidence au microscope électronique d'un virus dans des cellules infectées in vitro par l'agent du polyome, in ‟Comptes rendus hebdomadaire des séances de l'Académie des Sciences", 1959, CCIL, pp. 483-485.
Bielschowsky, F., Hall, W. H., Carcinogenesis in parabiotic rats. Tumors of liver and seminal vesicles induced by acetylaminofluorene in normal males joined to castrated males or females, in ‟British journal of cancer", 1951, V, pp. 106-114.
Bischoff, F., Cocarcinogenic activity of cholesterol oxidation products and sesame oil, in ‟Journal of the national Cancer Institute", 1957, XIX, pp. 977-985.
Bischoff, F., Bryson, G., Carcinogenesis through solid state surfaces, in ‟Progress in experimental tumor research", 1964, V, pp. 85-133.
Bischoff, F., Lopez, G., Rupp, I. I., Gray, C. L., Carcinogenic activity of cholesterol degradation products, in ‟Federation proceedings", 1955, XIV, pp. 183-184.
Biskind, M. S., Biskind, G. S., Development of tumors in the rat ovary after transplantation into the spleen, in ‟Proceedings of the Society for Experimental Biology and Medicine", 1944, LV, pp. 176-179.
Bittner, J. J., Some possible effects of nursing on the mammary gland tumor incidence in mice, in ‟Science", 1936, LXXXIV, p. 162.
Bittner, J. J., The receptibility to cancer of the mammary glands of mice, in ‟Proceedings of the Society for Experimental Biology and medicine", 1936, XXXV, pp. 42-48.
Bloch, B., Dreyfus, N., Die experimentelle Erzeugung von Röntgen-Carcinomen bei Kaninchen nebst allgemeinen Bemerkungen über die Genese der experimentellen Carcinome, in ‟Schweizerische medizinische Wochenschrift", 1921, LI, pp. 1033-1037.
Boiron, M., Paoletti, C., Thomas, M., Rebiere, J. P., Bernard, J., Acide désoxyribonucléique infectieux extrait de cultures de cellules de rein de Singe babouin infectées par le virus SV40, in ‟Comptes rendus hebdomadaires des séances de l'Académie des Sciences; D: Sciences Naturelles", 1962, CCLIV, pp. 2097-2099.
Borrel, A., Épithélioses infectieuses et épithéliomas, in ‟Annales de l'Institut Pasteur de Lille", 1903, XVII, pp. 81-122.
Bresciani, F., Nola, E., Sica, V., Puca, G. A., Early stages in estrogen control of gene expression and its derangement in cancer, in ‟Federation proceedings", 1973, XXXII, pp. 2126-2132.
Bresciani, F., Puca, G. A., Fissazione del 17 β-estradiolo in ghiandola mammaria e in tumori mammari del topo C3H/HeJ, in ‟Atti della Società Italiana di Patologia", 1965, pp. 661-664.
Bresciani, F., Puca, G. A., Maffettone, G. A., Riduzione dei siti leganti l'estradiolo nella frazione citoplasmatica solubile dei tumori mammari del topo C3H, in ‟Atti della Società Italiana di Patologia", 1967, pp. 711-715.
Bresciani, F., Puca, G. A., Nola, E., Salvatore, M., Ardovino, I., Meccanismo dell'azione estrogena e trasformazione neoplastica, in ‟Atti della Società Italiana di Patologia", 1969, pp. 204-224.
Bryan, W. R., Moloney, J. B., Calnan, D., Stable standard preparations of the Rous sarcoma virus preserved by freezing and storage at low temperature, in ‟Journal of the National Cancer Institute", 1954, XV, pp. 315-329.
Burk, D., A colloquial consideration of the Pasteur and neo-Pasteur effects, in ‟Cold Spring Harbor symposia on quantitative biology", 1939, VII, pp. 420-459.
Burkitt, D., A sarcoma involving the jaws in African children, in ‟British journal of surgery", 1958, XLVI, pp. 218-223.
Butenandt, R., Dannenberg, H., Die Biochemie der Geschwülste, in Handbuch der allgemeinen Pathologie, vol. VI, Geschwülste, Berlin-Göttingen-New York 1956, pp. 107-241.
Caputo, A., I presupposti molecolari della trasformazione neoplastica, Roma 1973.
Carrel, A., Le principe filtrant des sarcomes de la poule produits par l'arsenic, in ‟Comptes rendus des séances de la Société de Biologie", 1925, XCIII, pp. 1083-1085.
Carrel, A., Un sarcome fusicellulaire produit par l'indol et transmissible par un agent filtrant, in ‟Comptes rendus des séances de la Société de Biologie", 1925, XCIII, pp. 1278-1280.
Claude, A., Parter, K. R., Pickels, E. G., Electron microscopy study of chicken tumor cells, in ‟Cancer research", 1947, VII, pp. 421-430.
Coman, D. R., Adhesiveness and stickiness; two independent properties of the cell surface, in ‟Cancer research", 1961, XXI, pp. 1436-1438.
Chrabtree, H. G., Observations on the carbohydrate metabolism of tumors, in ‟Biochemical journal", 1959, XXIII, pp. 535-545.
Dao, T. L., Carcinogenesis of mammary gland in rat, in ‟Progress in experimental tumor research", 1964, V, pp. 157-216.
Di Mayorca, G., Callender, G., Marin, G., Giordano, R., Temperature-sensitivite mutants of polyoma virus, in ‟Virology", 1969, XXXVIII, pp. 126-133.
Di Mayorca, G. A., Eddy, B. E., Stewart, S. E., Hunter, W. S., Friend, C., Bendich, A., Isolation of infectious deoxyribonucleic acid from SE polyoma-infected tissue culture, in ‟Proceedings of the National Academy of Sciences", 1959, XLV, pp. 1805-1808.
Di Paolo, J. A., Levin, M. L., Tumor incidence in mice after oral painting with cigarette smoke condensate, in ‟Journal of the National Cancer Institute", 1965, XXXIV, pp. 595-600.
Di Paolo, J. A., Moore, G. E., Effect on mice of oral painting of cigarette smoke condensate, in ‟Journal of the National Cancer Institute", 1959, XXIII, pp. 529-534.
Druckrey, H., Die Pharmakologie krebserregender Substanzen, in ‟Zeitschrift für Krebsforschung und klinische Onkologie", 1950, LVII, pp. 70-85.
Druckrey, H., Küpfmüller, K., Quantitative Analyse der Krebsentstehung, in ‟Zeitschrift für Naturforschung; Teil C: Biochemie, Biophysik, Biologie, Virologie", 1948, IIIb, pp. 254-266.
Druckrey, H., Schmäll, D., Cancerogene Wirkung von Polyäthylenfolien an Ratten, in ‟Zeitschrift fur Naturforschung; Teil C: Biochemie, Biophysik, Biologie, Virologie", 1954, IXb, pp. 529-530.
Duran-Reynals, F., A hemorragic disease occurring in chicks inoculated with the Rous and Fujinami viruses, in ‟Yale journal of biology and medicine", 1940, XIII, pp. 77-98.
Duryll, W. R., Seminar on transmission studies on renal carcinoma of the frog, in ‟Journal of the Franklin Institute", 1956, CCLXI, pp. 377-379.
Earle, W. R., Production of malignancy in vitro, in ‟Journal of the National Cancer Institute", 1943, IV, pp. 131-248.
Eddy, B. E., Borman, G. S., Berkeley, W. H., Young, R. D., Tumors induced in hamsters by injection of rhesus monkey kidney cell extracts, in ‟Proceedings for the Society of Experimental Biology and Medicine", 1961, CVII, pp. 191-197.
Ehrlich, P., Apolant, H., Beobachtungen über maligne Mäusetumoren, in ‟Klinische Wochenschrift (Berlin)", 1905, LXII, pp. 871-874.
Ellermann, V., Bang, O., Experimentelle Leukämie bei Hühner, in ‟Zentralblatt für Bakteriologie, Parasitenkunde, Infektionskrankheoiten und Hygiene", Abteilung I. Originale, 1908, XLVI, pp. 595-609.
Epstein, M. A., Achong, B. G., Barr, Y. M., Virus particles in cultured lymphoblasts from Burkitt's lymphoma, in ‟Lancet", 1964, I, pp. 702-703.
Epstein, M. A., Achong, B. G., Barr, Y. M., Zajac, B., Henle, G., Henle, W., Morphological and virological investigations on cultured Burkitt's tumor lymphoblasts (strain Raji), in ‟Journal of the National Cancer Institute", 1966, XXXVII, pp. 547-559.
Epstein, M. A., Barr, Y. M., Cultivation in vitro of human lymphoblasts from Burkitt's malignant lymphoma, in ‟Lancet", 1964, I, pp. 252-253.
Epstein, M. A., Barr, Y. M., Characteristic and mode of growth of a tissue cultur strain (EBI) of human lymphoblasts from Burkitt's lymphoma, in ‟Journal of the National Cancer Institute", 1965, XXXIV, pp. 231-240.
Farber, E., Studies on the chemical pathology of lesions produced by ethionine, in ‟Archives of pathology", 1959, LXVII, pp. 1-8.
Fieser, L. F., Greene, T. W., Bischoff, F., Lopez, G., Rupp, I. I., A carcinogenic oxidation product of cholesterol, in ‟Journal of the American Chemical Society", 1955, LXXVII, pp. 3928-3929.
Findlay, G. M., Ultraviolet light and skin cancer, in ‟Lancet", 1928, CCXV, pp. 1070-1073.
Fischer, A., Dauerzüchtung reiner, stamme von Carcinomzellen in vitro, in ‟Zeitschrift für Krebsforschung", 1927, XXV, pp. 89-96.
Fischer, A., Die Erzeugung bösartiger Geschwülste, in ‟Zentralblatt für Bakteriologie, Parassitenkunde, Infektionskrankheiten und Hygiene", 1927, CIV, pp. 31-36.
Fisher, I. F., Mallette, M. F., The occurrence of ethionine in bacteria, in ‟Journal of general physiology", 1961, XLV, pp. 1-13.
Fried, M., Cell-transforming ability of a temperature-sensitive mutant of polyoma virus, in ‟Proceedings of the National Academy of Sciences", 1965, LIII, pp. 486-491.
Fujinami, A., Inamoto, K., Über Geschwülste bei japanischen Haushühnern, insbesondere über einem transplantablen Tumor, in ‟Zeitschrift für Krebsforschung", 1914, XIV, pp. 94-119.
Fujinami, A., Suzue, K., Contribution to the pathology of tumor growth. Experiments on the growth of chicken sarcoma in the case of heterotransplantation, in ‟Transaction of the Japanese Pathology Society", 1928, XVIII, pp. 616-622.
Gardner, W. V., Development and growth of tumors in ovaries transplanted into the spleen, in ‟Cancer research", 1955, XV, pp. 109-117.
Gey, C. P., Cytological and cultural obsdervations on transplantable rat sarcomata produced by the inoculation of altered normal cells maintained in continous culture, in ‟Cancer research", 1941, I, p. 737.
Goldblatt, H., Cameron, G., Induced malignancy in cells from rat myocardium subjected to intermittent anaerobiosis during long propagation in vitro, in ‟Journal of experimental medicine", 1953, XCVII, pp. 525-552.
Gross, L., Spontaneous leukemia developing in C3H mice following inoculation in infancy, with AK leukemic extracts, or AK-embryos, in ‟Proceedings of the Society for Experimental Biology and Medicine", 1951, LXXVI, pp. 27-32.
Gross, L., A filterable agent, recovered from AK-leukemic extracts, causing salivary gland carcinomas in C3H mice, in ‟Proceedings of the Society for Experimental Biology and Medicine", 1953, LXXXIII, pp. 414-421.
Gross, L., Attempt to recover filterable agent from X-ray-induced leukemia, in ‟Acta haematologica", 1958, XIX, pp. 353-361.
Gross, L., Oncogenic viruses, Oxford-London 1970.
Gross, L., Roswit, B., Wada, E. R., Dreyfuss, Y., Moore, L. A., Studies on radiation-induced leukemia in mice, in ‟Cancer research", 1959, XIX, pp. 316-320.
Haddow, A., Kon, G. A. R., Chemistry of carcinogenic compounds, in ‟British medical bulletin", 1947, VII, pp. 314-326.
Haddow, A., Robinson, A. M., The influence of various polycyclic hydrocarbons on the growth rate of transplantable tumours, in ‟Proceedings of the Royal Society of London", 1937, CXII, pp. 442-476.
Haddow, A., Sexton, W. A., Influence of carbamic esters (urethanes) on experimental animal tumors, in ‟Nature", 1946, CLVII, pp. 500-502.
Hanafusa, H., Hanafusa, T., Rubin, H., The defectiveness of Rous sarcoma virus, in ‟Proceedings of the National Academy of Sciences", 1963, XLIX, pp. 572-580.
Hanafusa, H., Hanafusa, T., Rubin, H., Analysis of the defectiveness of Rous sarcoma virus. II. Specification of RSV antigenicity by helper virus, in ‟Proceedings of the National Academy of Sciences", 1964, LI, pp. 41-48.
Hanau, A. N., Erfolgreiche experimentelle Übertragung von Carcinom, in ‟Fortschritte der Medizin", 1889, VII, pp. 321-339.
Harvey, J. J., An identified virus which causes the rapid production of tumors in mice, in ‟Nature", 1964, CCIV, pp. 1104-1105.
Hayward, E., Weitere klinische Erfahrungen über die Anwendung der Scharlachfarbstoffe und deren Komponenten zur beschleunigten Epithelialisierung granulierender Flächen, in ‟Münchener medizinische Wochenschrift", 1909, LVI, pp. 1836-1838.
Hecht, G., Zur Wirkung von Dimethylaminoazobenzol, in ‟Archive für experimentelle Pathologie und Pharmacologie", 1952, CCXV, pp. 610-618.
Henle, G., Henle, W., Diehl, V., Relation of Burkitt's tumor-associated herpes-type virus to infectious mononucleosis, in ‟Proceedings of the National Academy of Sciences", 1968, LIX, pp. 94-101.
Heston, W. E., Carcinogenic action of the mustards, in ‟Journal of the national Cancer Institute", 1950, XI, pp. 415-423.
Hieger, I., The spectra of cancer-producing tars and oil and of related substances, in ‟Biochemical journal", 1930, XXIV, pp. 505-511.
Hieger, I., The isolation of a cancer-producing hydrocarbon from coal tar. Part I. Concentration of the active substance, in ‟Journal of the Chemical Society", 1933, pp. 395-405.
Hieger, I., Carcinogenic substance in human tissues, in ‟Cancer research", 1946, VI, pp. 657-667.
Hieger, I., Carcinogenic activity of preparations rich in cholesterol, in ‟Nature", 1947, CLX, pp. 270-271.
Hieger, I., Carcinogenic activity of lipoid substances, in ‟British journal of cancer", 1959, III, pp. 123-139.
Hieger, I., Carcinogenesis by cholesterol, in ‟British journal of cancer", 1959, XIII, pp. 439-451.
Hieger, I., Carcinogenesis, London-New York 1961.
Howard, A., Pelc, S., Synthesis of DNA in normal and irradiated cells and its relation to chromosome breakage, in ‟Heredity", 1953, VI, suppl., pp. 261-273.
Huebner, R. J., Todaro, G. J., Oncogenes of RNA tumor viruses as determinants of cancer, in ‟Proceedings of the National Academy of Sciences", 1969, LXIV, pp. 1087-1094.
Hueper, W. C., Recent developments in environmental cancer, in ‟Archives of pathology", 1954, LVIII, pp. 360-399, 475-523, 645-682.
Hueper, W. C., Experimental studies in metal cancerigenesis. IV, in ‟Journal of the National Cancer Institute", 1955, XVI, pp. 55-73.
Hueper, W. C., Zueffle, J. H., Link, A. M., Johnson, M. G., Experimental studies in metal cancerigenesis. II. Experimental uranium cancers in rats, in ‟Journal of the National Cancer Institute", 1952, XIII, pp. 291-301.
Hull, R. N., Minner, J. R., Smith, J. W., New viral agents recovered from tissue cultures of monkey kidney cells. I. Origin and properties of cytophatic agents SV1, SV2, SV4, SV5, SV6, SV11, SV12, and SV15, in ‟American journal of hygiene", 1956, LXIII, pp. 204-215.
Ito, Y., A tumor-producing factor extracted by phenol from papillomatous tissue (Shope) of Cottontail rabbits, in ‟Virology", 1960, XII, pp. 596-601.
Jagic, N., Schwartz, G., Schwartz, V., Siebenrock, L., Blutbefunde bei Röntgenologen, in ‟Klinische Wochenschrift", 1911, XLVIII, pp. 1220-1222.
Jarvick, M. E., Tobacco smoking in monkeys, in ‟Annals of the New York Academy of Sciences", 1967, CXLII, pp. 280-294.
Jensen, C.O., Experimentelle Untersuchungen über krebs bei Mäusen, in ‟Zentralblatt für Bakteriologie, Parsitenkunde, Infektionskrankheiten und Hygiene", Abteilung I. Originale, 1903, XXXIV, pp. 28-34.
Jensen, E. V., Jacobson, H. I., Basic guides to the mechanism of estrogen action, in ‟Recent progress in hormone research", 1962, XVIII, pp. 387-408.
Kaplan, H. S., Brown, M. B., Protection against radiation-induced lymphoma development by shielding and partial-body irradiation of mice, in ‟Cancer research", 1952, XII, pp. 441-444.
Kennaway, E. L., The formation of a cancer-producing substance from isoprene (2 methylbutadiene), in ‟Journal of pathology and bacteriology", 1924, XXVII, pp. 233-238.
Kennaway, E. L., Experiments on cancer-producing substances, in ‟British medical journal", 1925, II, pp. 1-4.
Kennaway, E. L., Hieger, I., Carcinogenic substances and their fluorescence spectra, in ‟British medical journal", 1930, I, pp. 1044-1046.
Kobayashi, A., Matsumoto, H., Methylazoxymethanol, the aglycone of cycasin, in ‟Federation Proceedings", 1964, XXIII, pp. 1354-1356.
Kojima, K., Sakai, I., On the role of stickiness of tumor cells in the formation of metastases, in ‟Cancer research", 1964, XXIV, pp. 1887-1891.
Lacassagne, A., Apparition de cancers de la mammelle chez la souris mâle, soumise à des injections de folliculine, in ‟Comptes rendus hebdomadaires des séances de l'Académie des Sciences", 1932, CXCV, pp. 630-632.
Lacassagne, A., Relationship of hormones and mammary adenocarcinoma in the mouse, in ‟American journal of cancer", 1939, XXXVII, pp. 414-424.
Lancaster, M. C., Jenkins, F. P., Philip, J. M., Toxicity with certain samples of groundnuts, in ‟Nature", 1961, CXCII, pp. 1095-1097.
Laquer, G. L., Mickelsen, O., Whiting, M. G., Kurland, L. T., Carcinogenic properties of nuts from Cycas cirunalis L., indigenous to Guam, in ‟Journal of the National Cancer Institute", 1963, XXXI, pp. 919-951.
Lathrop, A. E.C., Loeb, L., Further investigations on the origin of tumors in mice. On the part played by hormones in the spontaneous development of tumors, in ‟Journal of cancer research", 1916, I, pp. 1-20.
Leitch, A., Paraffin cancer and its experimental production, in ‟British medical journal", 1922, II, pp. 1104-1106.
Liebermann, M., Kaplan, H. S., Leukemogenic activity of filtrates from radiation-induced lymphoid tumors of mice, in ‟Science", 1959, CXXX, pp. 387-388.
Little, C. C., The genetics of cancer in mice, in ‟Biological reviews", 1947, XXII, pp. 315-343.
Loeb, L., On transplantation of tumors, in ‟Journal of medical research", 1901, I, pp. 28-38.
Lucké, B., A neoplastic disease of the kidney of the frog, Rana pipiens, in ‟American journal of cancer", 1934, XX, pp. 352-379.
Marie, P., Clunet, J., Raulot-Lapointe, G., Contribution à l'étude des tumeurs malignes sur ulcères de Roentgen, in ‟Bulletin de l'Association Française pour l'Étude du Cancer", 1910, III, pp. 404-426.
Martland, H., Occupational poisoning in manufacture of luminous watch dials. General review of hazard caused by ingestion of luminous paint with special reference to the New Jersey case, in ‟Journal of the American Medical Association", 1929, XCII, pp. 466-552.
Matsumoto, H., Nagahama, T., Larson, H. O., Studies on methylazoxymethanol the aglycone of cycasin. A synthesis of methylazoxymethyl acetate, in ‟Biochemical journal", 1965, XCV, pp. 13P-14P.
Miller, J. A., Comments on chemistry of cycades, in ‟Federation proceedings", 1964, XXIII, pp. 1361-1362.
Mizell, M., Stackpole, C. W., Isaac, J. J., Herpestype virus latency in the Lucké tumor, in Biology of amphibian tumors, Heidelberg 1969, pp. 337 ss.
Mizutani, S., Boettiger, T., Temin, H., A DNA-dependent DNA polymerase and a DNA endonuclease in virions of Rous sarcoma virus, in ‟Nature", 1970, CCXXVIII, pp. 424-427.
Mizutani, S., Temin, H., RNA-dependent DNA polymerase in virions of Rous sarcoma virus, in ‟Nature", 1970, CCXXVI, pp. 1211-1213.
Moloney, J. B., A virus-induced rhabdomyosarcoma of mice, in Conference on murine leukemia. National Cancer Institute Monograph N. 22, U.S. Public Health Service, Bethesda 1966, pp. 139-142.
Nettleship, A., Hensaw, P. S., Meyer, H. L., Induction of pulmonary tumors in mice with ethyl carbamate (urethan), in ‟Journal of the National Cancer Institute", 1943, IV, pp. 309-317.
Nothdurft, H., Die experimentelle Erzeugung von Sarkomen bei Ratten und Mäusen durhc Implantation von Rundscheiben und Gold, Silber, Platin oder Elfenbein, in ‟Naturwissenschaften", 1955, XLII, pp. 75-76.
Nothdurft, H., Tumorerzeugung durch Fremdkörperimplantation, in ‟Abhandlungen der deutschen kademie der Wissenschaften zu Berlin", 1960, III, pp. 80-89.
Oberling, C., Guérin, M., Sarcome de la poule par méthylcholanthrène devenu filtrable, in ‟Bulletin du cancer", 1950, XXXVII, pp. 5-14.
Oppenheimer, B. S., Oppenheimer, E. T., Danishefsky, I., Stout, A. P., Carcinogenic effect of metals in rodents, in ‟Cancer research", 1956, XVI, pp. 439-441.
Oppenheimer, B. S., Oppenheimer, E. T., Danishefsky, I., Stout, A. P., Eirich, F. R., Further studies of polymers as carcinogenic agents in animals, in ‟Cancer research", 1955, XV, pp. 330-340.
Oppenheimer, B. S., Oppenheimer, E. T., Stout, A. P., Sarcomas induced in rats by implanting cellophane, in ‟Proceedings of the Society for Experimental Biology and Medicine", 1948, LXVII, pp. 33-34.
Oppenheimer, B. S., Oppenheimer, E. T., Stout, A. P., Willhite, M., Danishefsky, I., The latent period in carcinogenesis by plastics in rats and its relation to the precancerous stage, in ‟Cancer", 1958, XI, pp. 204-213.
Passey, R. D., Experimental soot cancer, in ‟British medical journal", 1922, I, pp. 1112-1113.
Peacock, P. R., Producton of tumors in fowl by carcinogenic agents: (1) Tar; (2) 1:2:5:6 Dibenzanthracene-lard, in ‟Journal of pathology and bacteriology", 1933, XXXVI, pp. 141-152.
Pentimalli, F., Lesioni dei tessuti come fattori di sviluppo dei tumori sperimentali, in ‟Lo Sperimentale (Archivio di biologia normale e patologica)", 1916, LXX, pp. 337-344.
Pentimalli, F., Quinto tumore trapiantabile, essiccabile e filtrabile dei polli, in ‟Rendiconti della R. Accademia dei Lincei", 1916, XXV, serie V, pp. 382-385.
Popper, H., Huerga, J. de la, Yesineck, C., Hepatic tumors due to prolonged ethionine feeding, in ‟Science", 1953, CXVIII, pp. 80-82.
Pott, P., The chirurgical works, Dublin 1778.
Pullman, A., Pullman, B., Sur les transformations métaboliques des hydrocarbures cancérogènes, in ‟Bulletin de la Société de Chimie Biologique", 1954, XXI, pp. 1097-1104.
Pulvertaft, R. J. V., Cytology of Burkitt's tumour (African lymphoma), in ‟Lancet", 1967, I, pp. 238-240.
Rajewsky, B., schraub, A., Kahlan, G., Experimentelle Geschwülsterzeugung durch Einatmung von Radiumemanation, in ‟Naturwissenschaften", 1943, XXXI, pp. 170-171.
Rehn, L., Blasengeschwülste bei Fuchsin-Arbeitern, in ‟Archiv für klinische Chirurgie vereinigt mit deutsche Zeitschrift für Chirurgie", 1895, L, pp. 588-600.
Roffo, A. H., Krebs und Sarkom durch Ultraviolett-und Sonnenstrahlen, in ‟Zeitschrift für Krebsforschung", 1934, XLI, pp. 448-467.
Rondoni, P., Il cancro. Istituzioni di patologia generale dei tumori, Milano 1946.
Rossner, W., Über neue Derivate des Methylcholanthrens und über einige vom Cholesterin abgeleitete heterocyclische Verbindungen, in ‟Hoppe-Seyler's Zeitschrfit für physiologische Chemie", 1937, CCIL, pp. 267-274.
Rous, P., A sarcoma of the fowl transmissible by an agent separable from tumor cells, in ‟Journal of experimental medicine", 1911, XIII, pp. 397-411.
Rous, P., Kidd, I. G., Conditional neoplasms and sub-threshold neoplastic states, in ‟Journal of experimental medicine", 1941, LXXIII, pp. 365-389.
Rous, P., Murphy, J. B., The nature of the filterable agent causing a sarcoma of the fowl, in ‟Journal of the American Medical Association", 1912, LVIII, p. 1938.
Rous, P., Murphy, J. B., On the causation by filterable agents of three distinct chicken tumors, in ‟Journal of experimental medicine", 1914, XIX, pp. 52-69.
Rous, P., Murphy, J. B., Tyler, W. H., The relation between a chicken sarcoma's behaviour and the growth's filterable cause, in ‟Journal of the American Medical Association", 1912, LVIII, pp. 1840-1841.
Rowe, W. P., Pugh, W. E., Hartley, J. W., Plaque assay techniques for murine leukemia viruses, in ‟Virology", 1970, XLII, pp. 1136-1139.
Rubin, H., A Virus in chick embryos which induces resistance in vitro to infection with Rous sarcoma virus, in ‟Proceedings of the National Academy of Sciences", 1960, XLVI, pp. 1105-1119.
Rubin, H., The nature of virus-induced cellular resistance to Rous sarcoma virus, in ‟Virology", 1961, XIII, pp. 200-206.
Salyamon, L. S., The role of inflammation in the mechanism of carcinogenic, cocarcinogenic and certain anti-carcinogenic effects, in ‟Problems of oncology", 1961, VII, pp. 44-50.
Sanarelli, G., Das myxomatogene Virus. Beitrag zum Studium des Krankheitserreger ausserhalb des Sichtbaren, in ‟Zentralblatt für Bakteriologie, Parassitenkunde, Infectionskrankheiten und Hygiene", Abteilung I. Originale, 1898, XXIII, pp. 865-873.
Sasaki, T., Yoshida, T., Experimentelle Erzeugung des Leber-carcinoms durch Füttertung mit O-Amidoazotoluol, in ‟Virchows Archiv für pathologische Anatomie und Physiologie und für klinische Medizin", 1935, CCXCV, pp. 175-200.
Schinz, H. R., Uehlinger, E., Der Metallkrebs. Ein neues Prinzip der Krebserzeugung, in ‟Zeitschrift für Krebsforschung", 1942, LII, pp. 425-437.
Schlenk, F., Observations on the metabolism of alkylmercaptans in yeast, in ‟Archives of biochemistry and biophysics", 1957, LXIX, pp. 67-73.
Schmidt, O., Das Kastenmodell in Theorie und Praxis der aromatischen Verbindungen, in ‟Berichte der deutschen chemischen Gesellschaft", A., 1940, LXXIII, pp. 97-116.
Shabad, L., Production expérimentale de tumeurs malignes par une extrait benzénique du foie d'un cancéreux. A propos de la question des substances cancérigènes endogènes, in ‟Comptes rendus des séances de la Société de Biologie", 1937, CXXIV, pp. 213-216.
Shear, M. J., Carcinogenic activity of some anthracene derivatives, in ‟Journal of biological chemistry", 1938, CXXIII, p. 108.
Shope, R. E., A filterable virus causing tumor-like condition in rabbits and its relationship to virus myxomatosum, in ‟Journal of experimental medicine", 1934, LVIII, pp. 607-624.
Simkovic, D., Characteristics of tumors induced in Mammals, expecially Rodents, by viruses of the avian leukosis sarcoma groups, in ‟Advances in virus research", 1972, XVII, pp. 95-127.
Simons, P. J., Dougherty, R. M., Antigenic characteristics of three variants of Rous sarcoma virus, in ‟Journal of the National Cancer Institute", 1966, XXXVI, pp. 389-404.
Sinha, D., Pascal, R., Furth, J., Transplantable thyroid carcinoma induced by thyrotropin: its similarity to human Hurtle cell tumors, in ‟Archives of pathology", 1965, LXXIX, pp. 192-198.
Spiegelmann, S., Burny, A., Das, M. R., Kaydar, J., Schlom, J., Travnicek, M., Watson, K., DNA-directed DNA polymerase activity in oncogenic RNA viruses, in ‟Nature", 1970, CCXXVII, pp. 10-29.
Steiner, P. E., Induction of tumors with extracts from human livers and human cancers, in ‟Cancer research", 1942, II, pp. 425-435.
Steiner, P. E., Incidence of carcinogenic factor in livers of cancer, non cancer, cirrhotic and negro patients, in ‟Cancer research", 1943, III, pp. 385-395.
Stewart, S. E., Leukemia in mice produced by a filterable agent present in AKR leukemic tissues with notes on a sarcoma produced by the same agent, in ‟Anatomical record", 1953, CXVII, p. 532.
Stewart, S. E., Studies on the herpes-type recovered from the Burkitt's tumor and other human lymphomas, in ‟Advances in virus research", 1969, XV, pp. 291-305.
Stewart, S. E., Eddy, B. E., Gochenour, A. M., Borgese, N. G., Grubbs, G. E., The induction of neoplsms with a substance released from mouse tumor by tissue culture, in ‟Virology", 1957, III, pp. 380-400.
Strong, L.C., The establishment of the C3H inbred strain of mice for the study of spontaneous carcinoma of the mammary gland, in ‟Genetics", 1935, XX, pp. 586-591.
Strong, L. C., The establishment of the ‛A' strain of inbred mice, in ‟Journal of heredity", 1936, XXVII, pp. 21-24.
Strong, L. C., The origin of some inbred mice, in ‟Cancer research", 1942, II, pp. 531-539.
Sweet, B. H., Hilleman, M. R., Detection of a ‛non-detectable' simian virus (vacuolating agent) present in rhesus and cynomolgus monkey kidney cell culture material, in Second International Conference on Live Poliovirus Vaccines, Pan American Health Organization and World Health Organization, Washington, June 1960, pp. 79-85.
Temin, H., Nature of the provirus of the Rous sarcoma, in ‟National Cancer Institute monographs", 1964, XVII, pp. 557-570.
Temin, H., The protovirus hypothesis: speculations on the significance of RNA-directed DNA synthesis for normal development and carcinogenesis, in ‟Journal of the National Cancer Institute", 1971, XLVI, p. III.
Temin, H., Rubin, H., Characteristics of an assay for Rous sarcoma virus and Rous sarcoma cells in tissue culture, in ‟Virology", 1958, VI, pp. 669-688.
Toshima, S., Takagi, N., Minowada, J., Moore, G. E., Sandbery, A. A., Electron microscope and cytogenetic studies of cells derived from Burkitt's lymphoma, in ‟Cancer research", 1967, XXVII, pp. 753-771.
Tsutsui, H., Über das künstlich erzeugte Cancroid bei der Maus, in ‟Gann: Japanese journal of cancer research", 1918, XII, pp. 17-21.
Turner, F. C., Sarcomas at sites of subcutaneously implanted bakelite disks in rats, in ‟Journal of the National Cancer Institute", 1941, II, pp. 81-83.
Viola, P. L., Bigotti, A., Caputo, A., Oncogenic response of rat skin, lungs and bones to vinyl chloride, in ‟Cancer research", 1971, XXXI, pp. 516-522.
Volkmann, R. V., Über Teer-Paraffin und Russkbres (Schornsteinfegerkrebs), in Beiträge zur Chirurgie, Leipzig 1875.
Warburg, O., Physikalische Chemie der Zellatmung, in ‟Biochemische Zeitschrift", 1921, CXIX, pp. 134-166.
Warburg, O., Über Milchsäurebildung beim Wachstum, in ‟Biochemische Zeitschrift", 1925, CLX, pp. 307-311.
Warburg, O., Über den Stoffwechsel der Tumoren, Berlin 1926.
Warburg, O., The metabolism of tumors, London 1930.
Warburg, O., Stoffwechsel der Carcinomzelle, Verhandlungen der Deutschen Akademie Wissenschaften, Berlin 3, 1943.
Warburg, O., Über die Enstehung der Krebszellen, in ‟Naturwissenschaften", 1955, XLII, pp. 401-406.
Warwick, G. P., Effect of the cell cycle on the carcinogens, in ‟Federation proceedings", 1971, XXX, pp. 1760-1765.
Weinhouse, S., Studies on the fate of isotopically labeled metabolites in the oxidative metabolism of tumors, in ‟Cancer research", 1951, XI, pp. 585-591.
Weinhouse, S., Oxidative metabolism of neoplastic tissues, in ‟Advances in cancer research", 1955, III, pp. 269-325.
Weinhouse, S., Millington, R. H., Wenner, C. E., Occurrence of the citric acid cycle in tumors, in ‟Journal of the American Chemical Society", 1950, LXXII, pp. 4332-4333.
Weinhouse, S., Millington, R. H., Wenner, C. E., Metabolism of neoplastic tissue. I. The oxidation of carbohydrate and fatty acids in transplanted tumors, in ‟Cancer research", 1951, XI, pp. 845-850.
Weiss, R. A., Früs, R. R., Katz, E., Vogt, P. K., Induction of avian tumor viruses in normal cells by physical and chemical carcinogens, in ‟Virology", 1971, XLVI, pp. 920-938.
Whiting, M. C., Toxicity of cycads, in ‟Economic botany", 1963, XVII, pp. 270-302.
Wieland, H., Dane, E., Über die Konstitution der Gallensäuren. III. Über die Haftstelle der Seitenkette, in ‟Hoppe-Seyler's Zeitschrift für physiologische Chemie", 1933, CCXIX, pp. 240-244.
Wood, E. M., Larson, C. P., Hepatic carcinomas in rainbow trout, in ‟Archives of pathology", 1961, LXXI, pp. 471-479.
Wynder, E. L., Graham, E. A., Croninger, A. B., Experimental production of carcinoma with cigarette tar, in ‟Cancer research", 1953, XIII, pp. 855-864.
Wynder, E. L., Hoffmann, D., Tobacco and tobacco smoke, New York-London 1967.
Yamagiwa, K., Ichikawa, K., Pathogenese der Epithelgeschwülste. Hervorrufung von Carcinomen am Kaninchenohr durch Teerpinselung, in ‟Gann: Japanese journal of cancer research", 1914, VIII, p. 132.
Zilber, L. A., Kriukova, I. N., Haemorrhagic disease of rats caused by Rous sarcoma virus, in ‟Voprosy virusologii", 1957, II, pp. 239-243.