
Prime forme di vita sulla Terra: la testimonianza geologica
Prime forme di vita sulla Terra: la testimonianza geologica
L'informazione paleobiologica è conservata sotto forma di fossili morfologici e chimici, di distribuzioni isotopiche e di geochimica dei sedimenti. Le rocce sedimentarie più antiche ben conservate risalgono a 3,515 miliardi di anni fa e si trovano nell'Australia Occidentale. Intorno a 3,5 miliardi di anni fa la vita si era ben affermata tanto da permettere l'esistenza di microrganismi fotoautotrofi, forse cianobatteri, e il ciclo del carbonio produceva un frazionamento isotopico del carbonio simile a quello riscontrabile in rocce più recenti. Le stromatoliti risalenti a 3,1 miliardi di anni fa del Supergruppo di Pongola nel Sudafrica non contengono microfossili, ma presentano strutture che suggeriscono un'attività cianobatterica biocostruttiva. A partire dal Neoarcheano, 2,8 ÷ 2,5 miliardi di anni fa, testimonianze ricavate da stromatoliti, microfossili e distribuzioni isotopiche del carbonio indicano la diffusa presenza di tappeti di cianobatteri bentonici e l'esistenza di cicli biologici con produzione e consumo di metano. Poiché la produzione di metano è una caratteristica esclusiva di alcuni archeobatteri, si ha prova dell'esistenza di due dei super-regni della vita. Non si hanno invece fino a ora prove dell'esistenza di eucarioti nell'Archeano.
Introduzione
L'informazione paleobiologica è conservata sotto una varietà di forme, tra cui i fossili morfologici, i fossili chimici quali gli idrocarburi indicatori biologici, e le modalità di distribuzione degli isotopi, in particolare del carbonio e dello zolfo. Inoltre, poiché il chimismo della superficie terrestre è controllato essenzialmente dalla vita che vi si svolge, si possono dedurre informazioni di natura biologica anche dalla geochimica dei sedimenti che indicano, per esempio, la concentrazione di ossigeno nell'atmosfera. Nonostante tutte le opportunità offerte da questi diversi approcci, le interpretazioni paleobiologiche hanno gli stessi limiti delle testimonianze fossili contenute nelle rocce: quanto più antico è il periodo della storia della Terra considerato, tanto più incomplete sono le testimonianze. Le rocce più antiche che si conoscano risalgono a 4,02 miliardi di anni fa (Kroner e Nutman, 1996); sono gneiss, ovvero rocce metamorfiche di alto grado nelle quali è improbabile che si siano conservate testimonianze di vita. Le pochissime successioni dell'Adeano, più antiche di 3,8 miliardi di anni, si sono tutte profondamente alterate a causa dei numerosi eventi termici e delle deformazioni, cosicché nessuna contiene tracce univoche di vita. Tra queste successioni assai antiche, la più estesa, e anche la più studiata, è quella del Complesso dello Gneiss di nsaq in Groenlandia (Nutman et al., 1996). l dati relativi agli isotopi del carbonio in questo complesso sembrerebbero indicare che circa 3,8 miliardi di anni fa erano presenti organismi autotrofi, cioè organismi che utilizzano carbonio inorganico per la formazione delle loro cellule, anche se generalmente si considera che gli effetti dovuti alle alterazioni diagenetiche siano stati così importanti da impedire una valutazione della composizione isotopica originale (Schopf, 1983). Per un'interpretazione paleobiologica più precisa di queste rocce si attende l'applicazione di nuove tecniche; per esempio, S.I. Mojzsis e collaboratori (1996) hanno descritto l'applicazione dell'analisi a microsonda ionica a particelle di carbonio contenute in granuli minerali fosfatici provenienti da questi gneiss e hanno dimostrato che essi presentano composizioni isotopiche in accordo con l'ipotesi di un'origine da materia organica prodotta da organismi autotrofi. Essi propongono che ciò rappresenti una prova convincente dell'esistenza di vita sulla Terra 3,85 miliardi di anni fa, ma data la novità della tecnica impiegata è necessaria ancora prudenza nelle interpretazioni (Hayes, 1996). Queste rocce includono tuttavia sedimenti formati si in acqua, e da ciò risulta la presenza di acqua sulla superficie della Terra 3,8 miliardi di anni fa, o prima. Tra quelle note, le più antiche rocce sedimentarie ben conservate risalgono a 3,515 miliardi di anni fa, e la narrazione della storia paleobiologica comincia da questo periodo.
Tre aspetti principali permettono di considerare attendibili le interpretazioni delle testimonianze, peraltro assai incomplete, della vita nell' Archeano, cioè 3,8 ÷ 2,5 miliardi di anni fa: la combinazione di più testimonianze indipendenti per la costruzione di interpretazioni auto consistenti, un lavoro di estrapolazione all'indietro nel tempo dalle testimonianze del Proterozoico, che sono numerose e ben documentate, e un secolo di esperienza. Questo saggio tratta esclusivamente del primo aspetto; numerose pubblicazioni forniscono infatti informazioni dettagliate sulle testimonianze del Proterozoico e sulla storia della ricerca in questo campo (Schopf, 1983; Schopf e Klein, 1992).
Evidenze sperimentali
Stromatoliti

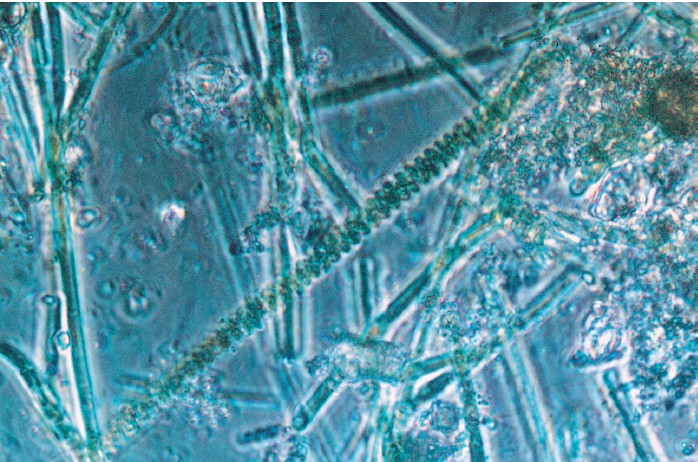
Negli ambienti marini e lacustri, tappeti di microrganismi bentonici, cioè organismi che vivono sul fondo, rimangono intrappolati, precipitano e inglobano sedimenti, generando strutture conservate sotto forma di stromatoliti (figg. 1, 2, 3). Per cercare la luce, o forse per altre ragioni, i microrganismi costruiscono strutture sedimentarie topograficamente rilevate, di dimensioni che coprono tutta la scala dal microscopico al monumentale; in vari periodi del passato essi hanno costruito scogliere grandi quanto quelle più recenti di coralli e di alghe, con un'altezza di 10 ÷ 100 m e più, e una lunghezza di centinaia di chilometri. Scogliere a stromatoliti sono molto numerose nelle successioni del Proterozoico, e molte sono note anche dall'Archeano. Il termine "stromatolite", usato in questo contesto, implica un'origine biogena, ma alcuni autori lo usano secondo una definizione morfologica che comprende anche strutture di origine abiogena. Può risultare molto difficile dimostrare in modo convincente l'origine biogena di alcune stromatoliti. J.P. Grotzinger e D.H. Rothman (1996) hanno tentato recentemente di trovare un criterio rigoroso per distinguere, tra le strutture di tipo stromatolitico, quelle biogene da quelle abiogene. Utilizzando i principi della geometria frattale per analizzare la morfologia di una forma di stromatolite del Proterozoico del Canada, essi sono giunti alla conclusione che, teoricamente, la morfogenesi delle stromatoliti può essere spiegata facendo ricorso a quattro processi abiotici: caduta di sedimento in sospensione, scivolamento gravitativo di tale sedimento (o effetti, equivalenti dal punto di vista matematico, di tensione superficiale nella precipitazione chimica), precipitazione perpendicolare alla superficie ed effetti casuali.
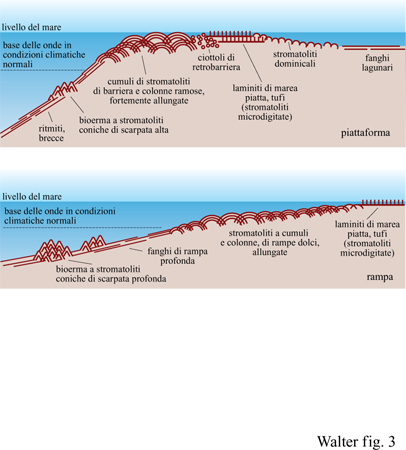
Sebbene possa essere molto difficile dimostrare l'origine biogena di strutture di tipo stromatolitico, vi sono buone ragioni per assumere che le stromatoliti siano biogene (Walter, 1996a). In primo luogo, esistono esempi di stromatoliti con microfossili disposti in modo tale da indicare che essi erano responsabili della costruzione delle lamine delle stromatoliti. In secondo luogo, la distribuzione delle facies sedimentarie carbonatiche dominate dalle stromatoliti nel Proterozoico (v. figura 3) è simile a quella di alghe e coralli del Fanerozoico, riflettendo così influenze biologiche comparabili. In terzo luogo, esistono numerose stromatoliti moderne nella cui morfogenesi si possono osservare direttamente influenze di natura biologica. Infine, è difficile trovare attualmente strutture stromatolitiche inoppugnabilmente di natura abiotica. Per esempio, recentemente, è stato dimostrato che ciò che in passato sembrava rappresentare un eccellente esempio di stromatoliti abiogene, le geyseriti di alta temperatura degli hot springs (sorgenti calde), sono in realtà abitate da biofilm microbici che sembra svolgano un ruolo critico nella morfogenesi delle geyseriti stesse (Cady e Farmer, 1996).
Microfossili
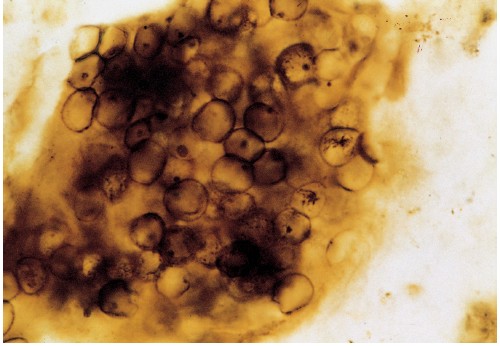
La fossilizzazione dei microrganismi (fig. 4) può avvenire per precipitazione di minerali incrostanti, caso in cui viene preservato solo il loro abito esterno, o attraverso permineralizzazione, nel qual caso il materiale cellulare è sostituito da minerali e spesso viene conservata anche materia organica. L'accuratezza nella conservazione della forma delle cellule può essere molto alta e, in rari casi, possono venire conservati persino gli organelli cellulari (Oehler, 1976; 1977), anche se quest'ultimo caso è ancora oggetto di discussione. Poiché la forma delle cellule è generalmente semplice, risulta talora problematico distinguere microfossili da arte fatti minerali (García-Ruiz, 1994). Per riconoscere i microfossili vengono utilizzati alcuni criteri quali la presenza di materia organica e l'indicazione di forme cave e di divisione cellulare. Poiché tuttavia alcuni funghi e cianobatteri sono in grado di perforare e penetrare le rocce, occorre anche dimostrare che ogni microfossile trovato non sia molto più giovane del deposito originario. In genere, questo può essere determinato in base a criteri petrografici, cioè stabilendo relazioni tra i fossili e i granuli di minerali della roccia ospite (Schopf e Walter, 1983).
Isotopi stabili
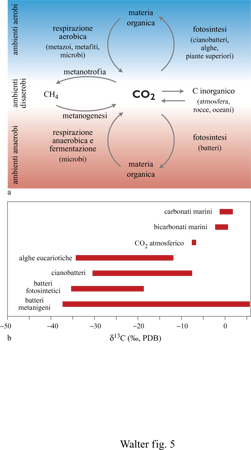
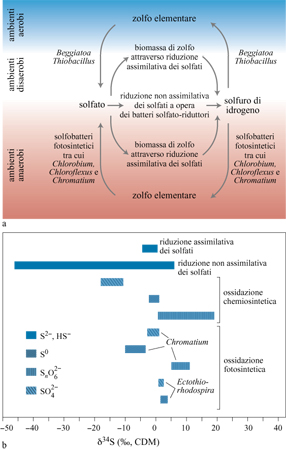
Varie reazioni metaboliche provocano il frazionamento degli isotopi del carbonio e dello zolfo (figg. 5, 6) e sono quindi registrate nella composizione della materia organica sedimentaria e in specie minerali contenenti tali elementi (Schidlowski et al., 1983). Tra i processi che possono essere individuati vi sono la fissazione autotrofica del carbonio, la produzione di metano e la riduzione catabolica del solfato, cioè la riduzione del solfato per ricavare energia. La distribuzione degli isotopi che ne risulta sopravvive, perlomeno, ai processi di metamorfismo deboli, cosicché tale approccio è largamente applicabile.
Indicatori biologici
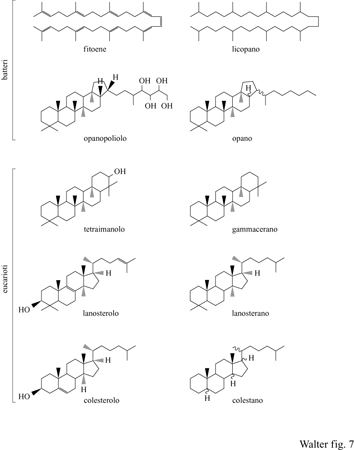
l lipidi, che caratterizzano molti organismi, quali per esempio i componenti della parete cellulare, e i pigmenti sono conservati nei sedimenti come loro derivati termicamente stabili, detti geoisomeri (fig. 7). Uno dei grandi progressi nella paleobiologia degli ultimi anni è stato lo sviluppo di tecniche analitiche che permettono di identificare con sicurezza anche piccolissime concentrazioni di questi biomarkers (indicatori biologici) nei sedimenti e di distinguerli da contaminanti di varia natura. L'utilizzo degli indicatori biologici nell'industria del petrolio è stato descritto in dettaglio da K.E. Peters e J.M. Moldowan (1993). Il loro contributo nell' avanzamento della paleobiologia del Proterozoico è stato discusso da R.E. Summons e M.R. Walter (1990). Tuttavia, tale approccio non è più applicabile qualora le rocce siano state riscaldate a temperature superiori a 200°C e quindi la materia organica sia stata deidrogenata. Questo limita l'applicazione di tali tecniche alle testimonianze dell'Archeano, in quanto si conoscono assai pochi sedimenti inalterati così antichi. Non ci sono pubblicazioni che riportino successi nell' applicazione di queste tecniche alle rocce dell'Archeano, ma recentemente sono stati ritrovati indicatori biologici in uno scisto risalente a 2,5 miliardi di anni fa nel Bacino di Hamersley nell'Australia Occidentale (R.E. Summons, comurucazione personale, 1996) e, probabilmente, tra breve il metodo verrà esteso con successo anche a rocce più antiche.
L'esplorazione dei paleoambienti
Tutte le stromatoliti e i microfossili descritti e la maggior parte dei dati sugli isotopi dell' Archeano provengono da paleoambienti marini e lacustri poco profondi. Con poche possibili eccezioni, sono tutti esempi di organismi mesofili, cioè che vivevano a temperature 'normali'. Tuttavia, di recente alcuni biologi hanno dimostrato che gli organismi più primitivi erano ipertermofili, con un optimum ecologico al di sopra di 80°C (fig. 8). Alla luce di questa scoperta si sarebbe potuto ipotizzare che i geologi avrebbero cercato tracce di termofili e ipertermofili in appropriati depositi dell'Archeano; tuttavia, ciò non è ancora avvenuto.
Nonostante i paleobiologi non abbiano rivolto quasi alcuna attenzione ai depositi di alta temperatura, sono stati ritrovati fossili in almeno tredici depositi idrotermali del Fanerozoico. Esistono molte possibilità di poter risalire al periodo di appartenenza di questi ritrovamenti: sono note infatti parecchie centinaia di casi documentati di depositi di minerali idrotermali di età archeana e proterozoica (Walter, 1996b). I depositi dell'Archeano si sono formati in una varietà di ambienti, che vanno da quello delle profondità marine a quello subaereo; per esempio, piccole stromatoliti coniche, come quelle formate attualmente da cianobatteri a 30 ÷ 59°C negli effluvi delle sorgenti termali subaeree, sono state ritrovate in tre successioni risalenti a 2,5 ÷ 2,8 miliardi di anni fa in Australia e in Canada (Grey, 1981; Hofmann et al., 1991; Hofmann e Masson, 1994).
È noto che negli ambienti idrotermali attuali vi sono comunità biologiche diversificate, anche se sono state studiate solo le parti superficiali di questi sistemi. La 'terza dimensione', il sistema sotterraneo in tutta la sua profondità, rimane inesplorata (Walter, 1996b). Questi sistemi offrono particolari opportunità per i paleobiologi, visto che loro esempi abbondano nelle testimonianze geologiche e sono molto più accessibili delle sorgenti calde (vents) attuali. Occorre ipotizzare la presenza di vita microbica in questi ambienti, così come è stata ritrovata in altri ambienti sotterranei (Pederson, 1993; Stetter et al., 1993; Parkes et al., 1994). Nei sistemi idrotermali le superfici sotterranee abitabili sono molto estese e associate a cospicui depositi minerali. È inevitabile, ma non noto con sicurezza, che i depositi in cui si sono formati minerali a temperature inferiori a 120°C siano stati influenzati da microrganismi o li racchiudano.
Le possibilità di rinvenire tracce della presenza di organismi ipertermofili nelle rocce più antiche inalterate presenti sulla Terra sono notevoli. Per esempio, depositi minerali di solfuri risalenti a 3,26 miliardi di anni fa del tipo black smoker nell' Australia Occidentale mostrano tessiture solfuriche eccezionalmente ben conservate e indistinguibili, apparentemente, da quelle degli attuali black smokers dei fondali marini; essi si sono formati ad almeno 1000 m di profondità. La geochimica di questi depositi indica temperature dei fluidi di almeno 300°C (Vearncombe et al., 1995), ma gli organismi devono essere vissuti a temperature assai più basse. In questi depositi sono presenti probabili microfossili filamentosi (S. Veamcombe, comunicazione personale, 1996).
Paleobiologia dell'Archeano

Si conoscono stromatoliti dell' Archeano provenienti da poco meno di trenta località nel mondo. Microfossili probabili o accertati sono noti solo da sei località (Walter, 1994; Altermann e Schopf, 1995). Informazioni sugli isotopi provengono da numerose località. Una discussione dettagliata sulle evidenze paleobiologiche dell'Archeano è presentata in Schopf (1983) e Schopf e Klein (1992). Paleoarcheano (3,8 ÷ 3,2 miliardi di anni fa) Le stromatoliti del Gruppo di Warrawoona in Australia Occidentale risalgono a 3,5 miliardi di anni fa (fig. 9) e sono le più antiche conosciute (Lowe, 1980; 1983; Walter et al., 1980; Buick et al., 1981; Groves et al., 1981; Walter, 1983; 1994; Hickman, 1990). Le stromatoliti del Gruppo del Fig Tree in Sudafrica (Byerly et al., 1986) sono forse leggermente più giovani. L'origine biogena di tutte queste stromatoliti è stata contestata da D.R. Lowe (1994) mentre quella di alcune forme del Gruppo di Warrawoona è stata riasserita da Walter (1994) e da R. Buick e collaboratori (1995a).
Le stromatoliti del Warrawoona sono stratiformi, pseudocolonnari e domiformi. Poiché si trovano in selci, si suppone che siano state formate in ambienti lagunari iperalini in una regione vulcanica attiva. La loro origine biogena è stata discussa a lungo, ma la maggior parte degli autori concorda sulla presenza di almeno 'probabili' stromatoliti, intendendo che tale attribuzione ha il medesimo grado di affidabilità delle stromatoliti di simile forma semplice del Proterozoico. Accenni di filamenti microbici sono stati trovati in una stromatolite domiforme.
Le stromatoliti del Fig Tree sono le più ipotetiche. Il loro aspetto è convincente, ma esse incrostano rocce di origine vulcanica in un ambiente dove è lecito attendersi la formazione di geyserite, assai difficile da distinguere dalle stromatoliti. Visto che di recente è stato scoperto che almeno alcune geyseriti sono in parte il prodotto di biofilm microbici ipertermofili (Cady e Farmer, 1996), i ritrovamenti del Fig Tree ben ripagherebbero uno studio più dettagliato.
I microfossili del primo Archeano sono estremamente rari e, come le stromatoliti, tutti sono stati o possono essere contestati (Schopf e Walter, 1983; Schopf, 1992; 1993). Molti microfossili segnalati per questa era, attualmente, non sono più considerati biogeni. Le forme filamentose della Formazione di Towers del Gruppo di Warrawoona sono sicuramente biogene, ma i tentativi di trovare ulteriori esempi nella stessa località sono falliti; sono così sorti dubbi sulla provenienza degli esemplari originali (Schopf, 1992). Nelle selci del Basalto di Apex del Gruppo di Warrawoona sono stati rinvenuti numerosi microfossili filamentosi e alcuni sferoidali (Schopf, 1993). Poiché la descrizione delle caratteristiche della zona di ritrovamento è piuttosto sommaria, occorre una certa cautela nel prendere in considerazione anche questa segnalazione. Questi filamenti sono stati attribuiti a numerosi taxa differenti e consistono di catene di cellule con una larghezza di 0,5 ÷ 19,5 μm. Non presentano guaine né si ritrovano in lamine stromatolitiche accertate. Secondo J.W. Schopf (1993), i più grandi sono probabilmente cianobatteri, perché hanno dimensioni maggiori della maggior parte delle forme dei batteri e, presumibilmente, degli archeobatteri. Tuttavia, la morfologia offre scarsi indizi per trovare somiglianze, soprattutto in assenza di ulteriori evidenze a supporto. Il cherogene ritrovato nella selce di Apex e nella Formazione di Towers (Strauss e Moore, 1992) è povero in 13C. Per indicare quantitativamente l'abbondanza relativa degli isotopi del carbonio si utilizza la scala δ, che esprime l'abbondanza degli isotopi stabili 12C e 13C rispetto a quelli di uno standard detto PDB. Per questo cherogene, δ¹³C va da -18 a -37, inaccordo con un'origine da metabolismo autotrofo, anche se non necessariamente da fotosintesi con produzione di ossigeno da parte dei cianobatteri.
È importante notare che fino a oggi è stata effettuata l'analisi isotopica solo del cherogene contenuto globalmente in campioni di roccia e che i risultati ottenuti non possono essere attribuiti direttamente ad alcun microfossile. La medesima considerazione vale per tutte le analisi effettuate sulle rocce dell'Archeano e del Proterozoico. Alla luce di quanto noto sui meccanismi di formazione delle stromatoliti attuali (Walter, 1994), le stromatoliti della Formazione di T owers sembrano essere state costruite da organismi filamentosi fotoautotrofi ma, ancora una volta, non necessariamente tali organismi devono essere stati cianobatteri. Sono stati trovati microfossili filamentosi anche nel Gruppo di Onverwacht in Sudafrica (Walsh e Lowe, 1985; Schopf, 1992), risalenti a 3,5 miliardi di anni fa; alcuni di questi si rinvengono in selce laminata che potrebbe essere stromatolitica. Come per il Gruppo di Warrawoona, il cherogene associato è povero in 13C, in accordo con un'origine autotrofa.
Nell'Australia Occidentale sono stati rinvenuti depositi sedimentari di solfuri metallici risalenti a 3,26 miliardi di anni fa, eccezionalmente ben conservati, che si formarono in sorgenti termali del tipo black smoker (Vearncombe et al., 1995). ln questi depositi sono presenti probabili microfossili filamentosi (S. Vearncombe, comunicazione personale 1996).
l minerali sedimentari solfatici dell' Archeano antico sono leggermente arricchiti in 34S se confrontati con i solfuri associati, e ciò risulta in accordo con un'origine da ossidazione fotosintetica di zolfo ridotto (Schidlowski et al., 1983). La deplezione in 34S nei solfuri, caratteristica della riduzione batterica dei solfati, non è osservabile fino al Paleoproterozoico, cioè a partire da 2,5 miliardi di anni fa.

Considerando tutte queste evidenze assieme alla geochimica dei sedimenti, le implicazioni biologiche sono le seguenti: l) la vita esisteva già almeno 3,5 miliardi di anni fa; 2) in ambienti lagunari poco profondi, dove i microrganismi dovevano combattere l'ipersalinità, l'essiccazione e la forte intensità di luce, si formarono tappeti di microbi bentonici; 3) i microbi erano sia a cocco che filamentosi; 4) alcuni microbi potrebbero aver utilizzato la luce come sorgente di energia, sebbene quest'ipotesi sia ancora controversa; rapporti isotopici del carbonio suggeriscono che essi fossero autotrofi. In figura (fig. 12) è riportata una ricostruzione dell'ambiente di quel tempo.
Mesoarcheano (3,2 ÷ 2,8 miliardi di anni fa)
Le stromatoliti del Gruppo di lnsuzi in Sudafrica, risalenti a circa 3,1 miliardi di anni fa, sono gli unici esempi di benthos nell'intervallo tra 3,2 e 2,8 miliardi di anni fa (Walter, 1983; Beukes e Lowe, 1989). Esse si sono formate in piane tidali e nei canali tidali adiacenti (Beukes e Lowe, 1989).
In queste stromatoliti sono evidenti due tipi di strutture chiaramente biogene. Stromatoliti bulbo se ampie e alte fino a 30 cm hanno un'organizzazione interna colonnare a strati. Le lamine sono increspate e costituite da lenti di carbonato semisferoidali giustapposte, alcune delle quali hanno una tessitura radiale che indica la possibile preesistenza di sottili microrganismi filamentosi disposti in modo radiale. Varie specie degli attuali cianobatteri formano 'ciuffi' paragonabili a tali strutture; tuttavia, anche alcuni depositi minerali inorganici hanno una tessitura simile. Altre stromatoliti nodulari e bulbose, grandi fino a 15 cm, possiedono al loro interno strutture finemente laminate in cui le colonne raggiungono 1 ÷ 5 mm di larghezza. Le lamine entro le colonne sono da convesse a coniche. Esistono anche colonne isolate di dimensioni maggiori con lamine a cono (Beukes e Lowe, 1989). Tali strutture possono essere paragonate in dettaglio con le stromatoliti attuali costruite dal cianobatterio oscillatoraceo Phormidium tenue. Di conseguenza, si può ipotizzare che queste stromatoliti dell' Archeano siano state costruite da sottili microrganismi filamentosi a fototassia positiva, aventi cioè la capacità di dirigersi verso la luce (Walter, 1983).
Con il Mesoarcheano, per la prima volta si hanno strutture che, in virtù delle loro caratteristiche sedimentologiche, della loro complessità macro- e microstrutturale e della somiglianza con le stromatoliti attuali, possono essere interpretate in modo convincente come vere stromatoliti. Non si conoscono microfossili di questa età. l dati sugli isotopi del carbonio sono in accordo con la presenza di organismi autotrofi.
Neoarcheano (2,8 ÷ 2,5 miliardi di anni fa)
A partire dal Neoarcheano le testimonianze geologiche divengono assai più numerose. Possono essere considerate come parte di un continuum della testimonianza fossile del Proterozoico, dove "letteralmente migliaia di esempi di microfossili sono stati ritrovati in centinaia di unità geologiche" (Altermann e Schopf, 1995), sono stati segnalati circa 1000 esempi di stromatoliti (Bertrand-Sarfati e Walter, 1981), esiste una solida documentazione sugli isotopi del carbonio e dello zolfo (Schopf e Klein, 1992), e inizia a delinearsi una documentazione di idrocarburi biomarkers (Summons e Walter, 1990). Nonostante ciò, nel Neoarcheano vi sono solo circa venti ritrovamenti documentati di stromatoliti e tre di microfossili (Walter, 1983; Altermann e Schopf, 1995).
La maggior parte dei pochi microfossili noti del Neoarcheano sono stati interpretati come cianobatteri; sono stati però individuati anche 'batteri' filamentosi. Tra gli esemplari del Gruppo di Fortescue in Australia e del Gruppo di Campbell in Sudafrica la presenza di microfossili interpretati come cianobatteri è avvalorata dalla presenza di questi ultimi in stromatoliti. Pochi esemplari sono ben conservati o particolarmente caratterizzati e le loro dimensioni relativamente grandi costituiscono l'argomento più saldo a favore di affinità con i cianobatteri (Altermann e Schopf, 1995).
La maggior parte delle stromatoliti sinora descritte provenienti da rocce neoarcheane sembrano essere di origine marina, ma due o tre sono probabilmente lacustri. Quelle di origine marina si trovano nelle cosiddette zone a pietre verdi, soggette ad attività tettonica, a notevoli fenomeni vulcanici e, quindi, a intense deformazioni. Probabilmente, esse formavano "scogliere strette ed effimere che fiancheggiavano vulcani subsidenti" (Grotzinger, 1989). Le stromatoliti lacustri si trovano in depositi di piattaforma (Walter, 1994); quelle tipo Conophyton dell'Australia Occidentale descritte da K. Grey (1981) e le forme tipo Thyssagetes del Canada descritte da Hofmann e collaboratori (1991), sono particolarmente significative perché queste caratteristiche forme coniche possono essere interpretate in analogia a forme ancora esistenti (Walter et al., 1976). Esse suggeriscono, in particolare, costruzioni da parte di microrganismi filamentosi sottili, a fototassia positiva, che potrebbero essere stati microaerofili, cioè organismi che vivono in ambienti con poco ossigeno libero; le forme attuali sono costituite da cianobatteri.

Le stromatoliti lacustri, fluviali e forse marine del Gruppo di Fortescue e del Ventersdorp dell'Australia Occidentale e del Sudafrica sono conservate in maniera eccezionale (fig. 13). Molte contengono tracce dei filamenti dei microorganismi che li hanno costruiti. In base alle tessiture presenti vi si possono riconoscere differenti comunità microbiche (Walter, 1983). Una sembra rappresentare un cianobatterio oscillatorio simile a Lyngbya, a filamenti grossolani, fototattico e fototropico; un'altra mostra caratteri di fototassia positiva, forse da parte di un cianobatterio oscillatorio a filamenti sottili. Il complesso dei dati isotopici del carbonio nel Neoarcheano è stato interpretato come indizio dell' esistenza di processi di fotoautotrofia coinvolgente la produzione di ossigeno, di metano genesi e di metilotrofia, cioè, rispettivamente, di produzione e consumo biologico di metano (Hayes, 1983; Buick, 1992). Il cherogene di parecchie unità litologiche di questa età è assai povero in 13C, avendo un δ¹³C pari a -50, a differenza del tipo comune che va da -25 a -35. Un frazionamento estremo degli isotopi del carbonio avviene nella produzione di metano da parte dei metano geni; ma non si conoscono altri processi simili che, in condizioni naturali, lo producano nei paleoambienti in cui le rocce si sono formate. Per essere incorporato nella materia organica sedimentaria, il metano dovrebbe essere stato quindi utilizzato da altri batteri come fonte di carbonio. Ciò rappresenterebbe una fase ossidativa che potrebbe essere stata associata alla riduzione di ossigeno. Se l'ossigeno fosse stato coinvolto, questo rappresenterebbe una prova indiretta dell' esistenza antecedente di fotosintesi con produzione di ossigeno, visto che nessun altro processo noto è in grado di produrre quantità significative di ossigeno libero. Si può anche ipotizzare che negli ambienti dove è stata registrata la forma isotopicamente leggera di cherogene, l'acqua fosse l'unico possibile donatore di elettroni disponibile in quantità sufficiente e, visto che i cianobatteri sono gli unici eubatteri che utilizzano questa via metabolica, ci deve essere stata una fotoautotrofia che generava ossigeno. Tale interpretazione risulta coerente con i ritrovamenti di stromatoliti e microfossili, e con i dati geochimici che confermano l'esistenza di grandi quantità di ossigeno libero nell'atmosfera del Paleoproterozoico, cioè a partire da 2,5 miliardi di anni fa.
Conclusioni
I contributi della geologia alla ricerca della vita sono limitati dalla scarsa disponibilità di successioni di rocce poco alterate rispetto alla loro deposizione iniziale. Malgrado che la rarità di depositi relativamente inalterati aumenti proporzionalmente all'età delle rocce, anche rari reperti possono fornire numerose informazioni, offrendoci una 'fotografia' delle forme di vita ad esse contemporanee. I depositi di 3,46 miliardi di anni del Gruppo di Warrawoona in Australia Occidentale ne sono un esempio. Per quanto ci è dato di sapere, già in quei tempi la vita era ben affermata sulla Terra, tanto che vi erano microrganismi fotoautotrofi, forse cianobatteri, ed era attivo un ciclo del carbonio tale da produrre frazionamenti isotopici del carbonio simili a quelli che si ritrovano in rocce più recenti. Da ciò possiamo estrapolare la presenza non solo di produttori primari diversificati, ma anche di eterotrofi efficienti nel riciclaggio del carbonio organico. È plausibile supporre che nei depositi minerali idrotermali dell'Archeano ci si possano attendere testimonianze paleobiologiche significative di ipertermofili, ma questi depositi sono ancora studiati in modo insufficiente.
Senza lo sviluppo di nuove tecniche di ricerca, che appaiono ancora lontane, non ci possiamo aspettare di penetrare la barriera della storia innalzata dagli stadi finali dell'accrezione della Terra, nel periodo detto del tardo e violento bombardamento, continuato fino a 3,8 miliardi di anni fa. Tutte le rocce più antiche devono essere state alterate così pesantemente che risulta impossibile ipotizzare una conservazione di informazioni biologiche. Comunque, recenti studi isotopici suggeriscono la presenza di vita 3,8 miliardi di anni fa (v. anche il saggio di C.F. Chyba e G.D. McDonald, Gli ambienti planetari e l'origine della vita). Finalmente, possiamo aspettarci di poter colmare in maniera sommaria lo iato temporale compreso tra i 3,8 e i 3,5 miliardi di anni fa. Per esempio, recentemente è stata segnalata al di sotto del Gruppo di Warrawoona nell'Australia Occidentale una successione ben conservata di rocce sedimentarie vulcaniche datata 3,515 miliardi di anni fa (Buick et al., 1995b). Sebbene, nonostante una ricerca preliminare, ancora non si conoscano stromatoliti o microfossili provenienti da questa successione, i primi risultati di studi isotopici sul carbonio indicano che il ciclo di questo elemento era funzionante come in tempi più recenti (R. Buick, comunicazione personale, 1996).
Le stromatoliti antiche 3, l miliardi di anni del Supergruppo di Pongola in Sudafrica non sono prive di microfossili, ma hanno delle forme particolari che suggeriscono costruzioni operate da cianobatteri. Dal Neoarcheano, 2,8 ÷ 2,5 miliardi di anni fa, la presenza di stromatoliti e microfossili e i rapporti isotopici del carbonio indicano la diffusa presenza di tappeti di cianobatteri bentonici e suggeriscono che erano attive la produzione e il consumo biologico di metano. Dato che la produzione biologica di metano è una caratteristica esclusiva di alcuni degli archeobatteri, abbiamo prove dell'esistenza di due dei super-regni della vita. Fino a oggi, invece, non si conosce nessuna prova dell'esistenza di eucarioti durante l'Archeano.
Bibliografia citata
ALTERMANN, W., SCHOPF, J.W. (1995) Microfossils from the Neoarchean Campbell group, Griqualand West Sequence of the Transvaal Supergroup, and their paleoenvironmental and evolutionary implications. Precambrian Research, 75, 65-90.
BERTRAND-SARFATI, J., WALTER, M.R. (1981) Stromatolite biostratigraphy. Precambrian Research, 15, 353-371.
BEUKES, N.l, LOWE, D.R. (1989) Environmental control on diverse stromatolite morphologies in the 3000 Myr Pongola Supergroup, South Africa. Sedimentology, 36, 383-397.
BUlCK, R. (1992) The antiquity of oxygenic photosynthesis: evidence from stromatolites in sulphate-deficient Archean lakes. Science, 255, 74-77.
BUlCK, R., DUNLOP, J.S.R., GROVES D.I. (1981) Stromatolite recognition in ancient rocks, and appraisal of irregularly laminated structures in an Early Archean chert-barite unit from North Pole, Western Australia. Alcheringa, 5, 161-181. BUlCK, R., GROVE, D.I., DUNLOP, J.S.R. (1995a) Commento Geology, Feb. 1995, 191.
BUlCK, R., THORNETT, J.R., McNAUGHTON, N.J., SMITH, J.B., BARLEY, M.E., SAVAGE, M. (1995b) Record of emergent continental crust ∼ 3.5 billion years ago in the Pilbara Craton of Australia. Nature, 375, 574-577.
BYERLY, G.R., LOWE, D.R., WALSH, M.M. (1986) Stromatolites from the 3,300-3,500 Myr Swaziland Supergroup, Barbeton Mountain Land, South Africa. Nature, 319, 489-491.
CADY, S.L., FARMER, J.D. (1996) Fossilization processes in silicieous thermal springs: trends in preservation along thermal gradients. In Evolution of hydrothermal ecosystems on Earth (and Mars?), Ciba Found. Symp. 202. Chichester, Wiley, pp. 150-173.
GARCÍA-RUlZ, J.M. (1994) Inorganic self-organisation in Precambrian cherts. Origins of life and Evolution of the Biosphere, 24, 451-467.
GREY, K. (1981) Small conical stromatolites from the Archean near Kanowna, Western Australia. Western Australia Geological Survey Annual Report Jor 1980, 90-94.
GROTZINGER, J.P. (1989) Facies and evolution ofthe Precambrian carbonate depositaI systems: emergence of the modern platform archetype. Society of Economic Paleontologists and Mineralogists Special Publication, 44, 79-106.
GROTZINGER, J.P., ROTHMAN, D.H. (1996) An abiotic model for stromatolite morphogenesis. Nature, 383, 423-425. GROVES, D.I., DUNLOP, J.S.R., BUlCK, R. (1981) An early habitat of life. Scientific American, 245, 56-65.
HAYES, J.M. (1983) Geochemical evidence bearing on the origin of aerobiosis. In Earth 's earliest biosphere; its origin and evolution, a c. di Schopf J.W., Princeton, Princeton University Press, pp. 291-301.
HA YES, J.M. (1996) The earliest memories of life on Earth. Nature, 384, 21-22.
HICKMAN, A.H., a c. di (1990) Pilbara and the Hamersley Basin. In Excursion guidebook, third international archean symposium, a c. di Ho S.E., Glover J.E., Myers J.S., Muhling J.R., University of Western Australia, pp. 1-57.
HOFMANN, H.I., SAGE, R.P., BERDUSCO, E.N. (1991) Archean stromatolites in Michipicoten Group siderite ore at Wawa, Ontario. Economic Geology, 86, 1023-1030.
HOFMANN, H.J., MASSON, M. (1994) Archean stromatolites from Abitibi greenstone belt, Quebec, Canada. Geological Society of America Bulletin, 106, 424-429.
KRONER, A, NUTMAN, AP., a C. di (1996) The Oldest Rocks on Earth. Precambrian Research, 78, 1-196.
LOWE, D.R. (1980) Stromatolites 3,400-Myr old from the Archean of Western Australia. Nature, 284, 441-443.
LOWE, D.R. (1983) Restricted shallow-water sedimentation of EarIy Archean stromatolitic and evaporitic strata of the Strelley Pool Chert, PiI bara Block, Western Australia. Precambrian Research, 19, 239-283.
LOWE, D.R. (1994) Abiological origin of described stromatolites older than 3.2Ga. Geology, 22, 387-390.
MOJZSIS, S.J., ARRHENIUS, G., McKEEGAN, K.D., HARRISON, T .M., NUTMAN, AP., FRIEND, C.R.L. (1996) Evidence of life on Earth before 3,800 million years ago. Nature, 384, 55-59.
NUTMAN, A.P., McGREGOR, V.R., FRIEND, C.R.L., BENNET, V.C., KINNY, P.D. (1996) The Itsaq Gneiss Complex of southern West Greenland; the world's most extensive record of early crustal evolution (3900-3600 Ma). Precambrian Research, 78, 1-39.
OEHLER, D.Z. (1976) Transmission electron microscopy of organic microfossils from the late precambrian bitter springs formation of Australia: techniques and survey of preserved ultrastructure. Journal of Paleontology, 50, 90-106.
OEHLER, D.Z. (1977) Pyrenoid-like structures in Late Precambrian algae from the Bitter Springs Formation of Australia. Journal of Paleontology, 51, 885-901.
PARKES, R.I., CRAGG, B.A., BALE, S.l GETLIFF, J.M., GOODMAN, K., RocHELLE, P.A., FRY, lC., WEIGHTMAN, A.l, HARVEY, S.M. (1994) Deep bacterial biosphere in Pacific Ocean sediment. Nature, 371, 410-413.
PEDERSON, K. (1993) The deep subterranean biosphere. EarthScience Reviews, 34, 243-260.
PETERS, K.E., MOLDOWAN, J.M. (1993) The biomarker guide; interpreting molecular fossils in petroleum and ancient sediments. Englewood Cliffs, Prentice Hall.
SCHIDLOWSKI, M., HAYES, J.M., KAPLAN, I.R. (1983) Isotopic inferences of ancient biochemistries: Carbon, Sulfur, Nitrogen, and Hydrogen. In Earth's earliest biosphere; its origin and evolution, a c. di Schopf J.W., Princeton, Princeton University Press, pp. 149-187.
SCHOPF, J.W. (1983) Earth's earliest biosphere: its origin and evolution. Princeton, Princeton University Press.
SCHOPF, J.W. (1992) Paleobiology of the Archean. In The proterozoic biosphere: a multidisciplinary study, a c. di Schopf J.W., Klein C., Oxford Univo Press, pp. 25-39.
SCHOPF, J.W. (1993) Microfossilis of the early Archean Apex chert: new evidence of the antiquity of life. Science, 260, 640-646.
SCHOPF, J.W., KLEIN, C., a c. di (1992) The Proterozoic Biosphere: A Multidisciplinary Study. Cambridge, Cambridge University Press.
SCHOPF, J.W., W ALTER, M.R. (1983) Archean microfossils: new evidence of ancient microbes. In Earth 's Earliest Biosphere: Its origin and Evolution, a c. di Schopf J.W., Princeton, Princeton University Press, pp. 214-339.
STETTER, K.O., HUBER, R., BLOCHL, E., KURR, M., EDEN, R.D., FIELDER, M., CASH, H., VANCE, I. (1993) Hyperthermophilic archea are thriving in deep North Sea and Alaskan oil reservoirs. Nature, 365, 743-745.
STRAUSS, H., MOORE, T.B. (1992) Abundances and isotopic compositions of carbon and sulfur species in whole-rock and kerogen samples. In The proterozoic biosphere: a multidisciplinary study, a c. di Schopf J.W., Klein C., Oxford Univo Press, pp. 709-798.
SUMMONS, R.E., WALTER, M.R. (1990) Molecular fossils and microfossils of prokaryotes and protists from Proterozoic sediments. American Journal of Science, 290-A, 212-244.
VEARNCOMBE, S., BARLEY, M.E., GROVES, D.I., McNAUGHTON, N.l, MIKUCKI, E.I., VEARCOMBE, J. R. (1995) 3.26 Ga blacksmoker-type mineralization in the Strelley Belt, PiI bara Craton, Western Australia. Journal of the Geological Society of London, 152, 587-590.
WALSH, M.M., LOWE, D.R. (1985) Filamentous microfossilis from the 3500 Myr-old Onverwacht Group, Barbeton Mountain Land, South Africa. Nature, 314, 530-532.
WALTER, M.R. (1983) Archean stromatolites: evidence of the Earth's earliest benthos. In Earth 's earliest biosphere: its origin and evolution, a c. di Schopf J.W., Princeton, Princeton University Press, pp. 187-213.
WALTER, M.R. (1994) Stromatolites: the major source of information on the evolution of the early benthos. In Early life in Earth, Nobel Symposium No. 84, a c. di Bengston S., New York, Columbia University Press, pp. 270-286.
WALTER, M.R. (1996a) Old fossils could be fractal frauds. Nature, 383, 385-386.
WALTER, M.R. (1996b) Ancient Hydrothermal Ecosystems on Earth: a New Paleobiological Frontier. In Evolution of hydrothermal ecosystem on Earth (and Mars?), Ciba Found. Symp. 202. Chichester, Wiley, pp. 112-127.
WALTER, M.R., BAULD, J., BROCK, T.D. (1976) Microbiology and morphogenesis of colurunar stromatolites (Conophyton, Vacerrilla) from hot springs in Yellowstone National Park. In Stromatolites, Developments in Sedimentology, 20, a c. di Walter M.R. Amsterdam, Elsevier, pp. 273-310.
WALTER, M.R., BUlCK, R., DUNLOP, J.R.S. (1980) Stromatolites 3,400-3,500 Myr old from the North Pole area, Western Australia. Nature, 284, 443-445.