
Respirazione, regolazione chimica della
Respirazione, regolazione chimica della
di Hans H. Loeschcke
Respirazione, regolazione chimica della
sommario: 1. Introduzione: a) i risultati meno recenti; b) il periodo quantitativo classico. 2. La chemosensibilità: a) i chemocettori periferici; b) chemosensibilità centrale. 3. La respirazione come meccanismo chemostatico. 4. Il controllo automatico del respiro a servizio dell'adattamento della respirazione alle necessità dei tessuti. 5. Deviazioni dal normale dell'aggiustamento del regolatore. 6. Interrelazioni tra respirazione ed escrezione renale nel mantenimento del normale equilibrio acido-base. 7. Il controllo automatico della respirazione a servizio della costanza del mezzo interno delle cellule nervose. □ Bibliografia.
1. Introduzione
Con regolazione della respirazione vengono abitualmente designati quei processi che adattano la ventilazione polmonare ai bisogni del metabolismo. Per mezzo di questa regolazione viene trasferita attraverso il polmone una sufficiente quantità di ossigeno al sangue, e quindi ai tessuti, per es. ai muscoli, e contemporaneamente viene allontanato il CO2 prodotto.
Questo adattamento respiratorio è designato come ‛regolazione chimica' della respirazione in quanto esso viene determinato da fattori chimici - specialmente dal CO2, prodotto terminale del metabolismo - oppure anche dall'azione delle sostanze chimiche che si generano nell'ipossia. A questo tipo di regolazione si potrebbero contrapporre altri processi regolatori non mediati da sostanze chimiche, per es. la regolazione ‛neurogena' della respirazione. Questa si realizza attraverso impulsi pilotanti che, partendo dall'innervazione della muscolatura, raggiungono i centri respiratori parallelamente all'innervazione stessa, o in via riflessa attraverso i recettori muscolari. Naturalmente sono esclusi da questi processi regolatori gli effetti diretti di sostanze chimiche sulla respirazione tessutale.
Per contro, negli ultimi anni è diventata sempre più chiara l'importanza della ventilazione polmonare per la regolazione dell'equilibrio acido-base nei liquidi dell'organismo (v. regolazione dell'equilibrio acido-base). In quest'ultimo caso la ventilazione polmonare non è la grandezza regolata, bensì regolatrice o, per usare un linguaggio tecnico, l'‛elemento finale di controllo del regolatore'.
Nel corso di questo articolo descriveremo l'interazione e la compatibilità di queste due funzioni, vale a dire prima l'adattamento della ventilazione polmonare alle richieste metaboliche e quindi l'impiego della ventilazione polmonare come meccanismo preminente nella regolazione dell'equilibrio acido-base.
a) I risultati meno recenti
Sul finire del secolo scorso era già noto che nel lavoro muscolare la ventilazione polmonare si adatta automaticamente all'aumentato ricambio. Si sapeva inoltre (v. Pflüger, 1868) che la respirazione poteva essere stimolata sia per aumento della pressione parziale del diossido di carbonio, sia per diminuzione di quella dell'ossigeno nell'aria inspirata. Inoltre, a opera dei lavori di LeGallois (v., 1812), Volkmann (v., 1841), Flourens (v., 1842), Markwald (v., 1890) e altri, prevaleva una concreta rappresentazione della sede dei centri respiratori nell'ambito di quella regione del midollo allungato designata come formatio reticularis. Successivamente, però, fu dimostrato che i territori interessati all'innervazione respiratoria sono ben più estesi, e inoltre che una serie di funzioni parziali prendono parte alla produzione dei movimenti respiratori fondamentali (v. Gesell, 1940; v. Pitts, 1946; v. Salmoiraghi e Burns, 1960).
Intorno al 1900 non era ancora definitivamente accertato il punto di attacco degli stimoli chimici a effetto respiratorio (CO2 e carenza di O2), né quindi la sede della ‛chemo-sensibilità', né le strutture coinvolte, designate in seguito come ‛chemocettori'. Il primo grande progresso nella soluzione di questo problema si ebbe con le ricerche di Frédéricq (v., 1901), che, mediante anastomosi dei vasi del collo, riuscì a perfondere la testa di un cane B utilizzando la circolazione di un cane A, mentre la testa di quest'ultimo era irrorata dal cane B. In queste condizioni sperimentali la chiusura delle vie aeree del cane A produceva un aumento della respirazione nel cane B e, per contro, una diminuzione nel cane A. Era cosi dimostrato che era possibile trasferire una modificazione chimica del sangue capace di produrre un aumento della respirazione, e che la sede più importante per la sensibilità a queste sollecitazioni chimiche doveva risiedere più cranialmente rispetto al punto in cui nell'esperimento di Frédéricq erano anastomizzati i vasi del collo.
Inoltre, naturalmente nei limiti delle condizioni sperimentali realizzate (narcosi), risultava dimostrato che la chemosensibilità era assente, o presente soltanto in minima parte, nel resto del corpo (per esempio muscolatura o polmoni) perché altrimenti il cane A avrebbe dovuto subire un aumento della sua attività respiratoria. Ovviamente ci si dovrebbe attendere una presa di posizione nei confronti di queste ricerche classiche da parte di alcuni autori moderni, che di recente si sono nuovamente pronunciati per una tale localizzazione.
Poiché precedentemente come possibile sede della chemosensibilità erano stati presi in considerazione prevalentemente i polmoni e i centri midollari, i risultati delle ricerche di Frédéricq furono considerati una dimostrazione indiscutibile dell'esistenza di una chemosensibilità centrale. L'ipotesi di un centro respiratorio chemosensibile è rimasta fino a oggi un dato costante in molti testi di fisiologia. Dalla nostra trattazione risulterà evidente che essa deve essere profondamente modificata.
b) Il periodo quantitativo classico
Successivamente fu ritenuto più importante determinare le relazioni quantitative esistenti tra gli stimoli respiratori, intesi inizialmente come CO2 e deficit di O2, e l'attività respiratoria. A queste ricerche contribuirono anzitutto J. S. Haldane, J. Barcroft, A. Krogh, H. Winterstein e numerosi altri. La via della ricerca quantitativa fu aperta però da Haldane (v., 1935) e dai suoi collaboratori sia con la precisazione del concetto di aria alveolare, sia con lo sviluppo dei metodi per la sua raccolta e per la sua analisi (v. Haldane e Priestley, 1905).
Dietro a queste ricerche c'era l'idea che le pressioni parziali o le concentrazioni di CO2 e O2 sciolti nel sangue fossero in equilibrio con quelle dei gas negli alveoli polmonari, sicché la loro determinazione, per es. della pressione del CO2 nell'aria alveolare, avrebbe permesso di trarre direttamente delle conclusioni sulla pressione del CO2 nel sangue arterioso. E poiché questo irrora i centri supposti chemosensibili, non era certamente una cattiva approssimazione ritenere la pressione parziale di CO2 nell'aria alveolare equivalente alla grandezza dello stimolo respiratorio. Diventava cosi possibile la determinazione sperimentale della relazione quantitativa tra stimolo (pressione di CO2) e risposta (ventilazione). Lo stesso metodo poteva naturalmente essere utilizzato per l'ossigeno.
Partendo dal fatto che il CO2 combinandosi con l'acqua dà origine ad H2CO3 che a sua volta produce ioni H+ e che in ipossia si formano nei tessuti degli acidi, per es. acido lattico, i quali ugualmente si dissociano producendo ioni H+, Winterstein (v., 1911) fece un tentativo di semplificazione proponendo una nuova ipotesi, detta ‛teoria della reazione', nella speranza di trovare un agente comune per gli effetti sulla ventilazione polmonare del CO2 e della carenza di O2. Ovviamente il denominatore comune sarebbe stato lo ione H+, considerato pertanto unico stimolo respiratorio.
Come conseguenza di questa teoria si formarono tra i più autorevoli fisiologi due correnti: mentre gli uni sostenevano la teoria della reazione, gli altri propugnavano quella della specificità di azione del CO2. Tra questi ultimi dobbiamo citare anzitutto Nielsen (v., 1936), il quale dimostrò chiaramente che un aumento nel sangue della concentrazione idrogenionica prodotto da acidi fissi, come per es. l'iniezione di acido cloridrico, o da acidi di origine metabolica, come l'acido acetico, aveva un effetto sulla respirazione di gran lunga inferiore a quello di un uguale aumento della concentrazione idrogenionica nel sangue ottenuto inspirando CO2.
Basandosi su ricerche di M. H. Jacobs, fu nuovamente Winterstein (v., 1921) a riconoscere che, a differenza degli acidi fissi, il CO2 poteva penetrare in tutti i tessuti a causa della sua elevata diffusibilità e perciò con ogni probabilità raggiungere più agevolmente i centri respiratori. Più tardi egli ipotizzò un eguale meccanismo per la diffusione nel liquor cerebrospinale (v. Winterstein e Gökhan, 1953).
L'ulteriore elaborazione della questione, allora non definitivamente risolta, se si dovesse considerare stimolo respiratorio il CO2 o l'H+ - questione questa la cui discussione aveva apportato grandi progressi all'elaborazione quantitativa dell'azione del CO2, dell'H+ e dell'O2 -, fu abbandonata da Gray (v., 1950) e sostituita con una trattazione più pragmatica del problema. Gray, infatti, studiò la relazione tra i suddetti stimoli respiratori e la ventilazione polmonare alla stregua dell'analisi di un sistema. In questo modo Gray stabili che finché queste grandezze venivano misurate nel sangue, si potevano fare previsioni quantitative per mezzo di un sistema di equazioni soltanto attribuendo a ciascuno dei tre parametri un'azione indipendente. Di conseguenza questa teoria fu chiamata da Gray ‛teoria del fattore multiplo' (multiple factor theory).
Data una serie di stimoli, si possono fare per mezzo di questa teoria ottime previsioni sulle modificazioni del respiro ottenibili, se si tengono presenti altre variabili, soprattutto l'interazione dell'O2 con gli altri due fattori. Con questo metodo risulta che con alte pressioni di ossigeno la risposta al CO2 e all'H+ è ridotta, rispetto alla respirazione a riposo, rispettivamente del 60% e del 40%. Inoltre, nella respirazione in aria l'azione dell'O2 è soltanto del 15% circa (v. Dejours e altri, 1958).
L'azione contemporanea di CO2, H+ e O2 fu definita nell'uomo da Nielsen e Smith (v., 1951) e da altri autori, e analizzata soprattutto da Lloyd e altri (v., 1958). In accordo con queste ricerche le due azioni si potenziano reciprocamente, come risulta dalla maggiore pendenza della curva di risposta al CO2 quando la tensione di ossigeno viene abbassata. Nel cane e nel gatto il quadro non è così semplice, in quanto la curva di risposta al CO2 in ipossia non soltanto diviene più ripida, ma viene spostata nella zona di basse pressioni del CO2.
Il grande vantaggio della teoria di Gray consiste nel fatto che essa si riferisce a grandezze soltanto ematiche, esattamente misurabili. Per lo stesso motivo, però, essa non può dir nulla sull'effettiva natura del meccanismo d'azione degli stimoli su qualsivoglia struttura chemosensibile, poiché prima dovrebbe essere data una risposta sperimentale al problema del trasporto dello stimolo dal sangue al recettore. Per questo è necessaria, anzitutto, la conoscenza dell'esatta localizzazione dei recettori. Infine, ancora per lo stesso motivo, la suddetta teoria non è in condizioni di escludere per gli stimoli chimici un meccanismo d'azione unico.
2. La chemosensibilità
a) I chemocettori periferici
La prima modificazione dell'ipotesi di un centro respiratorio chemosensibile si rese necessaria allorché Heymans e Heymans (v., 1927), nel corso di ricerche sulla regolazione della pressione arteriosa a opera di recettori della parete vasale nel territorio aortico, osservarono che accanto alla pressione arteriosa anche la composizione chimica del sangue esercitava effetti riflessi sulla respirazione e sul circolo.
Seguirono le classiche ricerche di C. Heymans e collaboratori, tra i quali sono da menzionare J. J. Bouckaert, L. Dautrebande e U. S. von Euler (v., per es., Heymans e Bouckaert, 1939). In queste ricerche fu dimostrato che piccolissimi organi, formati da un particolare tessuto designato dagli anatomisti come paragangliare e situati in vicinanza dell'arco aortico e della biforcazione delle carotidi, i glomera carotica e aortica, sono punti di partenza di chemoriflessi a effetto sulla respirazione e sul circolo. Come metodo in queste ricerche fu usata ancora una volta la circolazione crociata di Frédéricq in diverse modificazioni, tra l'altro quella in cui la testa di un cane B isolata era perfusa dal circolo di un cane A.
Se in queste condizioni sperimentali si faceva respirare il cane A ad elevate tensioni di CO2 o in miscele povere di O2, allora la testa B reagiva con un aumento dei movimenti respiratori. Bastava anche soltanto perfondere il glomo carotideo isolato con sangue arricchito di CO2 o povero di O2 per ottenere un incremento della respirazione. I nervi afferenti per questi riflessi sono il vago (anzitutto quel ramo designato come nervo aortico o nervo depressore di Cyon), e il nervo del seno (di Hering), ramo del nervo glossofaringeo.
Dopo sezione di questi nervi la maggior parte della sensibilità al CO2 inspirato era però ancora presente, mentre l'ipossia produceva principalmente una depressione della respirazione. Solamente in successive ricerche, eseguite su animali non in narcosi, si osservò un aumento della frequenza del respiro. Tuttavia questa risposta residua alla deficienza di ossigeno era debole e di tutt'altro tipo delle risposte all'ipossia che si osservano quando i nervi chemosensibili sono intatti. Queste lievi risposte ottenute dopo denervazione sono probabilmente da attribuirsi a effetti secondari dell'ipossia, in rapporto alla formazione di metaboliti acidi.
Con ciò rimaneva dimostrato che la sensibilità della respirazione alla carenza di ossigeno è da ricercare, prescindendo da effetti secondari, esclusivamente nei glomi chemosensibili descritti da Heymans e altri (v., per es., Gesell e altri, 1940). Ancora oggi, per contro, le opinioni sono divise per quanto riguarda il ruolo svolto dalla sensibilità al CO2 dei glomi carotidei.
Heymans, sulla base delle ricerche in circolazione crociata, pervenne alla conclusione che la partecipazione dei glomi alla sensibilità al CO2 fosse considerevole. Risultati del tutto simili sono stati ottenuti negli ultimi tempi con altra tecnica da Bouverot (v. Bouverot e altri, 1965).
Comroe (v., 1965), basandosi su ricerche nelle quali il glomo carotideo isolato veniva perfuso e su altre ancora (v. Cropp e Comroe, 1961) nelle quali sangue arricchito in CO2 veniva iniettato nelle carotidi, ha espresso l'opinione che le oscillazioni fisiologiche della tensione del CO2 esercitino una scarsa influenza sui glomi, ma abbiano grande importanza per la respirazione, che pilotano attraverso la chemosensibilità centrale che rimane dopo esclusione dei glomi. Si è visto di recente che l'importanza dei chemocettori periferici è grande quando si verificano rapide variazioni della tensione di CO2, mentre le lente variazioni della tensione di base del CO2 sono segnalate prevalentemente attraverso i chemocettori centrali. Comunque, non è ancora definitivamente chiarito se la sensibilità dei chemocettori periferici al CO2 sia dovuta al CO2 di per sé oppure si realizzi attraverso gli ioni H+ liberati dall'H2CO3, nel senso di Winterstein. Ricerche più recenti (v. Hornbein e Roos, 1963) sono più favorevoli a quest'ultima ipotesi.
Poco dopo la scoperta dei chemocettori periferici si riuscì a registrare i potenziali d'azione nei loro nervi; la fig. 2 mostra un esperimento di von Euler e collaboratori (v., 1940). Inoltre, gli effetti sulla respirazione degli impulsi percorrenti i nervi chemosensibili possono essere studiati con la stimolazione elettrica di questi nervi. Gli impulsi originanti dai chemocettori e i riflessi da essi iniziati furono studiati esaurientemente soprattutto da Neil e collaboratori (v. Heymans e Neil, 1958) e sono tutt'ora oggetto di un'intensa indagine (v. Torrance, 1968).
Le ricerche classiche sulla morfologia dei glomi condotte da De Castro (v., 1928) sono state completate da più recenti osservazioni, fra le quali alcune al microscopio elettronico (v. Hess, 1968; v. Acker e altri, 1977).
Due tipi di cellule risultano elementi caratteristici dei glomi; le une circondano le altre e l'insieme viene considerato come una singola unità funzionale recettiva. Queste cellule sono disseminate tra vasi sanguigni discretamente ampi, con parete a struttura capillare (sinusoidi).
Il metabolismo del glomo carotideo, misurato come consumo di O2, risultò estremamente elevato e la sua irrorazione sanguigna ancora più alta, con la conseguenza che il sangue venoso refluo è quasi completamente saturo di O2 (v. Daly e altri, 1954). Ciò non esclude che nei singoli vasi e in una parte del tessuto la tensione di ossigeno possa essere bassa. Più recentemente si è visto che all'interno del glomo la tensione di O2 può scendere vicino allo zero (v. Acker e altri, 1971). Infine, poiché i glomi carotidei originano dagli archi branchiali, Schmidt e Comroe (v., 1940) suppongono che la sensibilità alla carenza di O2 di queste strutture risalga filogeneticamente alla respirazione branchiale dei Pesci.
b) Chemosensibilità centrale
Come già detto, dopo l'esclusione dei chemocettori periferici del territorio carotideo e aortico rimane una sensibilità della respirazione alla tensione del CO2, ovvero alla concentrazione degli ioni H+, del tutto sufficiente a mantenere una respirazione normale.
Il primo dubbio sulla concezione classica, e cioè che questa chemosensibilità fosse localizzata nei centri respiratori, fu avanzato da Åström (v., 1952), il quale osservò che durante la narcosi la sensibilità al CO2 è fortemente diminuita, in uno stadio in cui la risposta alla carenza di ossigeno è perfettamente conservata. Partendo da questa osservazione, in verità già nota da lungo tempo e ripetutamente confermata, Åström concluse che la chemosensibilità e il comando dell'adattamento respiratorio non potevano essere localizzati nella stessa sede, poiché l'adattamento della respirazione alla carenza di ossigeno si dimostra ancora funzionante nella narcosi, laddove invece la risposta chemosensibile centrale al CO2 risulta ridotta o del tutto abolita. Di conseguenza egli postulò una suddivisione dei centri respiratori in un ‛centro respiratorio riflesso' e in un ‛chemocentro respiratorio'.
Nello stesso anno von Euler e U. Söderberg (v., 1952) pubblicarono un lavoro nel quale descrissero potenziali d'azione derivati dall'interno del midollo allungato. Avendo osservato che in questi esperimenti la frequenza degli impulsi aumentava con la tensione del CO2 nel sangue, essi credettero di aver dato con ciò la dimostrazione dell'esistenza di chemocettori midollari. I risultati ottenuti da questi autori erano di particolare interesse, perché gli impulsi registrati non erano modulati con il ritmo respiratorio, al contrario di quanto è noto per gli impulsi delle cellule dei centri respiratori. Di conseguenza, diventava verosimile che questi impulsi fossero diretti ai centri respiratori e che questo risultato sperimentale rappresentasse una conferma del postulato di Åström.
Un nuovo approccio iniziò quando Leusen (v., 1950) dimostrò che la perfusione dei ventricoli cerebrali, dal ventricolo laterale sino alla cisterna magna, produceva un aumento della ventilazione polmonare quando effettuata con soluzioni tampone acide, e una diminuzione con quelle alcaline. Era così dimostrato che strutture chemosensibili potevano essere raggiunte con questo procedimento sperimentale. Che non si trattasse poi dei centri respiratori stessi, come supposto originariamente, fu dimostrato dalle ricerche di Loeschcke e Koepchen (v., 1958), nelle quali, dopo esclusione delle afferenze chemosensibili periferiche, la perfusione di procaina attraverso il quarto ventricolo, nel gatto, causava una considerevole diminuzione della ventilazione sino all'arresto respiratorio. Per contro, l'applicazione diretta della procaina sul pavimento della fossa romboidale, cioè relativamente vicino alla sede dei centri classici, non ebbe alcun effetto sulla ventilazione. Di conseguenza, la sede della chemosensibilità fu ricercata sulla superficie del recesso laterale del quarto ventricolo o in quella parte della base del cervello confinante con l'apertura laterale. Quest'ultima localizzazione ebbe una conferma dalle ricerche di Mitchell e altri (v., 1963), che ottennero aumenti della respirazione con l'applicazione locale di carta da filtro imbevuta di tamponi acidi sulla superficie ventro-laterale del midollo allungato in corrispondenza delle radici dei nervi cranici IX e X. La procaina applicata nella stessa sede causava depressione della ventilazione, mentre l'acetilcolina e la nicotina ne producevano un aumento. Le modificazioni della ventilazione ottenute con questo metodo comparvero spesso molto precocemente, cioè col primo atto respiratorio dopo l'applicazione della sostanza attiva, e pertanto era possibile supporre che le strutture responsabili giacessero almeno in parte molto superficialmente (secondo calcoli di Mitchell, a una profondità di circa 200 μm). Tuttavia erano necessari alcuni minuti al raggiungimento della reazione completa (tempo di semireazione 3-4 minuti) e pertanto doveva prendervi parte un processo più lento, per es. una diffusione a profondità maggiore.
La stimolazione elettrica con impulsi rettangolari del midollo allungato nell'area di Mitchell, eseguita da Loeschcke e altri (v., 1970), produsse un aumento della ventilazione. L'allestimento di una mappa degli effetti degli stimoli applicati dimostrò che un aumento simile della ventilazione poteva essere ottenuto da una seconda superficie localizzata bilateralmente sulla parte ventrale del midollo allungato in corrispondenza delle radici dell'ipoglosso. La stimolazione per mezzo di un elettrodo a infissione applicato in questa sede a profondità crescenti (v. Trouth e altri, A superficial..., 1973) mise in evidenza che la risposta diminuiva già a una profondità di 0,3 mm e scompariva del tutto a 0,8 mm, rivelando così la localizzazione superficiale di questa struttura. Trouth e altri cercarono di localizzare il massimo dell'effetto stimolante producendo uno sviluppo di gas per elettrolisi mediante l'applicazione di una corrente continua alla punta degli elettrodi, in modo tale che la sede dell'elettrolisi potesse essere riconosciuta nelle sezioni istologiche. Questa risultò essere sempre in strettissima vicinanza a un gruppo caratteristico di grandi cellule gangliari multipolari, le quali non erano state ancora descritte come una speciale unità. Era ovvia quindi la conclusione che queste cellule appartenessero al meccanismo di risposta all'applicazione dello stimolo elettrico.
Schläfke e altri (v., 1970) svilupparono successivamente un metodo di perfusione locale della superficie del midollo allungato con soluzioni tampone e riscontrarono aumenti respiratori applicando tamponi acidi nei limiti della superficie di Mitchell e in quelli della superficie, descritta sopra, in corrispondenza della radice dell'ipoglosso. Di conseguenza, si rese necessario ammettere che ambedue le superfici suddette contenessero strutture chemosensibili. Shimada e altri (v., 1969) e Schläfke e altri (v., 1975) riuscirono a ottenere potenziali d'azione con elettrodi applicati medialmente alla radice dell'ipoglosso. La frequenza di questi potenziali d'azione aumentava quando la superficie del midollo allungato era perfusa con liquor artificiale più acido della norma (pH 7,0 a 37 °C) e, per contro, diminuiva fino all'arresto completo quando il liquor perfuso era più alcalino (pH 7,6 a 37 °C). Si è visto che questi potenziali d'azione possono essere registrati in tutto il territorio situato medialmente alle radici dei nervi cranici da IX a XII. Essi si estendono fino al nucleo paragigantocellulare (v. Schläfke, 1979).
Scegliendo concentrazioni diverse di bicarbonato è possibile, nel liquor artificiale usato per la perfusione, variare il pH mantenendo costante la tensione parziale di CO2, oppure la PCO2 tenendo costante il pH. Con questo metodo fu possibile dimostrare che la ventilazione era attivata al massimo quando il pH veniva abbassato mantenendo costante la tensione di CO2, e lo era meno quando il pII veniva abbassato aumentando la tensione di CO2 in presenza di una concentrazione costante di bicarbonato e, inoltre, che la ventilazione diminuiva allorché la tensione parziale di CO2 era aumentata mantenendo costante il pH (v. Loeschcke e altri, 1958). Di conseguenza, risulta inverosimile che il CO2 abbia un'azione specifica indipendente dagli ioni H+ dissociati dall'H2CO3, come era stato supposto da una parte dei più vecchi fisiologi della respirazione. Il fondamento di questa ipotesi era, infatti, che l'aumento della respirazione nella inalazione di CO2 risultasse di gran lunga maggiore dell'aumento che si manifestava quando una corrispondente modificazione del pH ematico era prodotta con iniezione di altri acidi o con acidi di origine metabolica. Come già precedentemente discusso, questa ‛specificità' è da spiegare anzitutto con il fatto che il CO2 diffonde facilmente in tutti i tessuti e nei liquidi extracellulari del cervello a contatto con le strutture chemosensibili, mentre alla diffusione degli anioni degli acidi fissi si oppone una barriera (barriera ematoencefalica, o barriera cerebrospinale), le cui basi anatomiche consistono verosimilmente nella particolare struttura dei piccoli vasi cerebrali e forse anche nel loro stretto contatto con le cellule della glia. Se quindi il CO2 non possiede alcuna azione specifica, è ovvio supporre che siano gli ioni H+, o protoni, a determinare, attraverso la loro concentrazione, l'attività del substrato chimico. In conclusione, questi risultati confermano la teoria della reazione di Winterstein per la chemosensibilità centrale. In verità, però, altri ioni sono importanti (specialmente K+ e Ca2+) il cui significato è ancora in fase di studio (v. Verstraeten, 1950; v. Berndt e altri, 1969).
Nuove ricerche sull'azione dell'acetilcolina hanno fornito importanti informazioni sulla natura dell'azione degli H+. L'acetilcolina, ben nota come mediatore dell'eccitamento a livello di determinate sinapsi, produce aumento della respirazione quando viene applicata sull'area chemosensibile del midollo allungato. Se mediante eserina si blocca l'enzima che scinde l'acetilcolina, aumenta la respirazione e l'azione dell'acetilcolina viene intensificata. L'atropina, un farmaco ben noto che blocca in parte le azioni dell'acetilcolina, impedisce l'azione sulla respirazione e inoltre produce una diminuzione dell'azione eccitante sulla respirazione che si osserva quando viene inspirato CO2 (v. Dev e Loeschcke, 1979). In base a questi risultati si conclude che l'azione degli H+ sulla respirazione ha come punto d'attacco un meccanismo di trasmissione dell'eccitamento, secondo uno schema generale che ha avuto numerose applicazioni negli studi sul sistema nervoso periferico e su quello centrale. Sarebbe stato dimostrato che l'acetilcolina intensifica la scarica delle cellule nervose della regione chemosensibile e principalmente di quegli stessi neuroni che si scaricano in risposta all'azione degli H+. Queste scariche sono aumentate dall'eserina e bloccate dall'atropina. Fukuda e Loeschcke (v., 1979) hanno dimostrato che dopo atropina la frequenza di scarica dei neuroni della zona chemosensibile non aumenta più per azione degli H+.
Infine, volendo giudicare l'impostanza del meccanismo chemosensibile sopra descritto per la regolazione della respirazione, si può partire dal fatto che la respirazione si arresta con la perfusione del midollo allungato con procaina, allorché i noti meccanismi chemosensibili del territorio carotideo e di quello aortico vengano esclusi con la sezione dei relativi nervi. In queste condizioni si ha arresto respiratorio anche quando venga inspirato CO2, malgrado la persistenza dell'eccitabilità elettrica dei centri. Ovviamente ciò significa che il descritto territorio, localizzato superficialmente nel midollo allungato, è il punto di partenza dei riflessi respiratori evocati dal CO2 e quindi del principale meccanismo di avvio della respirazione normale.
In ricerche sul cane e su capre non narcotizzate Pappenheimer e altri (v., 1965) dimostrarono che l'entità delle modificazioni respiratorie evocabili mediante perfusione degli spazi del liquor con soluzioni più o meno tamponate basta in realtà a spiegare da sola, per mezzo di questo meccanismo, la ventilazione a riposo e le sue modificazioni causate dalle inspirazioni di CO2 e dall'accumulo di acidi nell'organismo (acidosi metabolica). Inoltre, poiché la ventilazione in tutti questi casi segue in stretta correlazione l'andamento del pH del liquor cerebrospinale, si deve concludere che anche nel caso dell'acidosi la modificazione chimica del sangue abbia come conseguenza, in qualche modo, una modificazione del pH del liquor, per es. attraverso uno scambio di ioni bicarbonato del liquor con ioni cloruro del sangue. Nella fig. 9 sono riprodotte le modificazioni della concentrazione di bicarbonato nel liquor in rapporto alle variazioni della concentrazione dello stesso nel sangue, secondo ricerche di Kronenberg e Cain (v., 1968). Inoltre, dalle ricerche del gruppo di Pappenheimer risulta che la composizione del liquido extracellulare che circonda le strutture del meccanismo chemosensibile è uguale alla composizione del liquor e si modifica nello stesso senso in tutte le circostanze suddette.
Fencl e altri (v., 1969) riuscirono a eseguire ricerche simili nell'uomo con tecnica più indiretta, pervenendo alle stesse conclusioni. È evidente che la sensibilità dei chemocettori periferici all'azione del CO2 e degli ioni H+ non può essere stata di grande impoitanza in tutte queste ricerche, perché altrimenti i risultati non sarebbero stati così univoci.
3. La respirazione come meccanismo chemostatico
Un sistema di regolazione può regolare, ovvero mantenere nei limiti dei valori normali, soltanto le grandezze controllabili dai recettori. In un sistema così fatto è logico attendersi che la grandezza interessata oscilli minimamente attorno al valore normale nelle immediate vicinanze dei recettori e, per contro, sia tanto meno costante quanto più lontana da questi. Altri fattori possono, allora, agire senza essere percepiti dai recettori, tanto più facilmente quanto più essi agiscono lontano dai recettori stessi. Nello stesso modo della distanza agisce anche, per es., una membrana di separazione.
Prescindendo dagli effetti collaterali del CO2 o degli ioni H+ sui recettori periferici e da quelli dell'O2 sui recettori centrali o dai meccanismi conseguenti, si può affermare, naturalmente semplificando di molto le cose, che il pH del liquido extracellulare del cervello è regolato attraverso le strutture chemosensibili centrali, mentre la tensione di ossigeno nel sangue lo è attraverso i chemocettori periferici. In un sistema di regolazione cosi fatto, la ventilazione rappresenta l'elemento finale di controllo; un aumento della ventilazione conduce all'eliminazione di CO2, vale a dire a un abbassamento della tensione del CO2 e a una diminuzione dell'acidità, laddove la tensione di O2 viene contemporaneamente innalzata, perché aumenta il rapporto tra la quantità di ossigeno apportata al polmone e quella presa dal sangue. Inoltre, poiché la ventilazione aumenta con la tensione del CO2 o con la riduzione della tensione di ossigeno, essa agisce come un ‛accoppiamento a retroazione' sia per l'innalzamento della tensione del CO2, sia per la diminuzione di quella dell'O2. In questo meccanismo l'aumento della ventilazione viene causato dagli impulsi partenti dai recettori (potenziali d'azione), i quali, raggiunti i centri respiratori, vengono trasformati in un'aumentata innervazione della muscolatura respiratoria. Lo stesso avviene quando pervengono nel sangue acidi diversi dal CO2. Infatti, il conseguente spostamento del pH ematico verso zone acide si ripercuote sul pH extracellulare del cervello. Questa modificazione del pH del sangue e del liquor viene segnalata ai centri dai recettori periferici e prima ancora da quelli centrali. Si inizia così un aumento della ventilazione di origine centrale, che allontanando il CO2 riporta vicino al valore di partenza il pH extracellulare nel cervello e di conseguenza regola contemporaneamente, anche se in modo meno efficace, il pH nel sangue.
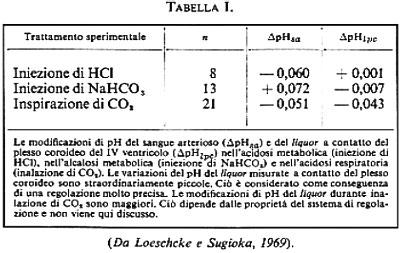
Anche se necessariamente approssimata, la prima descrizione quantitativa del processo di regolazione è dovuta a J. S. Gray (v., 1950). Essa permette di predire la ventilazione polmonare e la tensione alveolare di CO2 nello steady state di condizioni diverse. Usando le formule di Grodins e altri (v., 1954) è possibile, inoltre, calcolare approssimativamente il comportamento nel tempo della ventilazione durante il passaggio da una condizione all'altra, per es. all'inizio o alla fine di una inspirazione di CO2. In questo calcolo vengono applicate le equazioni differenziali per i processi di miscelazione nel polmone in funzione del tempo. Questi calcoli furono in seguito perfezionati da Grodins stesso e da altri autori. Negli ultimi tempi Middendorf e Loeschcke (v., 1976) hanno preso in considerazione per la prima volta il fatto che i recettori centrali misurano la concentrazione degli H+ in un liquido extracellulare che si distingue solo di poco dal liquor cerebrospinalis. Quanto sensibile sia questa regolazione del pH nel liquor risulta dalla tab. I, dove sono paragonate tra loro le modificazioni di pH del sangue e del liquor nell'acidosi metabolica acuta prodotta sperimentalmente. In queste ricerche di Loeschcke e Sugioka (v., 1969) fu misurato il pH sulla superficie del plesso coroideo, ricco di vasi sanguigni e partecipe della formazione del liquor. Il vantaggio di tale modo di procedere consiste nel fatto che le modificazioni del liquor possono essere misurate senza alcun ritardo, mentre lo svantaggio è che il liquor di recente formazione non è del tutto identico a quello che circonda il cervello, a quello dei ventricoli cerebrali o al liquido extracellulare del cervello stesso.
4. Il controllo automatico del respiro a servizio dell'adattamento della respirazione alle necessità dei tessuti
Apparentemente la respirazione con l'aiuto del meccanismo chemosensibile centrale sembra servire allo scopo di mantenere costante il pH nell'ambiente circostante le cellule nervose; il che, del resto, è senza dubbio vero. D'altra parte, in accordo con la fisiologia classica, la ventilazione polmonare dovrebbe servire a coprire le richieste d'ossigeno e ad allontanare il CO2 nelle modificazioni metaboliche, per es. nel lavoro muscolare. Quest'ultima funzione diviene quindi un caso speciale della regolazione sopra descritta. Ciò è dimostrabile come segue: la quantità di diossido di carbonio eliminata nell'unità di tempo, V•CO2, può essere calcolata dalla ventilazione alveolare, V•A, e dalla concentrazione relativa (frazione) di CO2 negli alveoli stessi, FA:
V•CO2=V•A•FA.
Sperimentalmente è dimostrato che nel lavoro muscolare, malgrado un'aumentata produzione metabolica di CO2 (V•CO2), la concentrazione relativa di diossido di carbonio (FA) non si modifica in modo sensibile.
Dall'equazione si può dedurre che ciò è possibile soltanto quando la ventilazione alveolare si modifica proporzionalmente alla produzione di CO2. Come è ovvio, ciò è esattamente quanto deve essere richiesto a un meccanismo di regolazione deputato all'adattamento della ventilazione al ricambio. Naturalmente, se questa condizione è soddisfatta, è anche sicuro che la ventilazione risulta sufficiente ad allontanare l'aumentato CO2 e ad assicurare l'apporto di ossigeno, poiché la quantità del CO2 prodotto corrisponde all'incirca alla quantità dell'ossigeno consumato. La regolazione di FA si realizza per il fatto che la frazione FA stessa determina il livello di tensione di CO2 nel sangue arterioso e quindi il pH extracellulare in corrispondenza dei recettori chemosensibili centrali, determinato anzitutto dalla tensione del CO2 arterioso. Per di più il lavoro muscolare pesante provoca la comparsa di acido lattico nel sangue e con ciò un ulteriore stimolo della ventilazione, come già precedentemente descritto. Altri fattori, per es. un aumento della temperatura corporea, agiscono nello stesso senso (v. Cunningham, 1963).
Sembrerebbe invero giustificato attendersi che in condizioni di sovraccarico funzionale del meccanismo respiratorio di regolazione, dovuto ad aumento del metabolismo, sia possibile una piccola deviazione della grandezza regolatrice FA, sufficiente a sostenere l'aumentata ventilazione. Ciò sarebbe caratteristico d'un regolatore proporzionale.
In effetti, però, l'esperimento dimostra che il parametro FA nel lavoro muscolare per lo più non devia in modo apprezzabile dal valore a riposo; e che, ciò malgrado, la ventilazione aumenta, per l'appunto, nella misura sufficiente a mantenere costante FA. In altre parole il sistema regolatore del respiro è più efficiente di un regolatore di tipo proporzionale. Questo fatto viene indicato nel linguaggio della tecnica della regolazione come ‛spostamento dal valore prefissato'. Tale deviazione dal valore prefissato è per l'appunto sufficiente a compensare la deviazione della grandezza regolatrice necessaria per l'aumento della ventilazione. In termini fisiologici tutto ciò significa che nel lavoro muscolare i centri debbono essere raggiunti da uno stimolo aggiuntivo ben dosato, la cui intensità deve essere correlata all'intensità del lavoro muscolare (v. Dejours e altri, 1958). È noto, infatti, che nel lavoro muscolare tali eccitazioni nascono dai recettori delle articolazioni. Inoltre partono impulsi dai recettori muscolari (detti ergocettori), la cui frequenza di scarica aumenta nel lavoro muscolare. Si può pensare che questi impulsi provenienti dai recettori articolari e muscolari raggiungano i centri respiratori. Stimolazioni della respirazione vengono ottenute eccitando i monconi centrali delle radici posteriori e pertanto esiste la possibilità di un aumento della respirazione a opera d'impulsi afferenti trasportati dai nervi spinali. Un'ulteriore possibilità è quella che impulsi efferenti dal sistema piramidale o extrapiramidale, diretti ai motoneuroni delle radici anteriori e quindi all'innervazione della muscolatura, raggiungano attraverso collaterali la formatio reticularis e le cellule ivi presenti dei centri di modulazione della respirazione, producendo lo ‛spostamento dal valore prefissato'. Allo stato attuale non vi è alcun fondamento a sostegno dell'ipotesi che soltanto uno dei possibili meccanismi produca da solo questo ‛spostamento dal valore prefissato' che di per sé, però, è sicuramente dimostrato. Infine, è fuori di dubbio che l'adattamento della ventilazione polmonare all'aumentato metabolismo sia da considerare come la prestazione di un meccanismo regolatore, devoluto principalmente al mantenimento della costanza del pH del liquido extracellulare del cervello. Questo meccanismo regolatore è però adattato con particolare precisione alle speciali richieste del lavoro muscolare.
Un'ulteriore possibilità di regolazione è stata di recente discussa. Essa si basa sul fatto che il pH del sangue fluttua con la frequenza del respiro. I segnali prodotti dal pH del sangue non sono dunque uniformi ma oscillano. Le ricerche di Eldridge (v., 1972) e di altri hanno dimostrato che l'azione sul respiro prodotta da un aumento della concentrazione degli H+ dipende dalla fase dell'atto respiratorio in cui detto aumento colpisce i chemocettori periferici. L'azione è massima quando si verifica alla fine dell'inspirazione. Nel lavoro muscolare i massimi livelli della concentrazione degli H+ potrebbero essere spostati in modo da esercitare la massima azione sul respiro.
5. Deviazioni dal normale dell'aggiustamento del regolatore
Poiché il meccanismo regolatore della respirazione possiede recettori sensibili alla concentrazione degli ioni H+ e alla quantità di ossigeno disponibile, i valori assunti di volta in volta dalla ventilazione, e quindi dalla tensione di ossigeno nel sangue e dalla concentrazione degli H+ nello spazio extracellulare del cervello, devono essere in ogni momento la risultante di un compromesso tra le richieste per il mantenimento costante delle due diverse grandezze. In condizioni normali non esiste alcun pericolo per l'apporto di ossigeno e la parte svolta dalla tensione di ossigeno nel mantenimento della ventilazione è scarsa. Il ruolo svolto dalla tensione di ossigeno diventa, però, maggiore quando vi sono ostacoli alla ventilazione polmonare o in alta montagna, con pressioni di ossigeno diminuite. In queste condizioni l'iperventilazione che ne deriva dovrebbe condurre a un abbassamento della concentrazione di ioni H+ nello spazio extracellulare cerebrale. Un tale abbassamento è stato dimostrato in vario modo anche nel liquido cerebrospinale (v. Winterstein e Gökhan, 1953; v. Robin e altri, 1958; v. Severinghaus e altri, 1963). Tuttavia quest'effetto a livello della sede del recettore centrale è forse minore di quanto sia stato supposto e potrebbe essere perfino del tutto assente, dato che il cervello in ipossia produce esso stesso acido lattico.
Ulteriori effetti sul livello funzionale della ventilazione possono essere provocati dagli ormoni. Secondo Cunningham e altri (v., 1958) la noradrenalina iniettata per via endovenosa stimola la ventilazione, evidentemente nel quadro dell'azione ergotropa di questa sostanza. Azione uguale, però più persistente, hanno le iniezioni di progesterone e di estradiolo (v. Heerhaber e altri, 1948). Questi effetti hanno una loro importanza nella gravidanza. L'azione dei sopracitati ormoni steroidi basta già nelle fasi iniziali della gravidanza ad abbassare la tensione arteriosa di CO2 da 38-40 torr a circa 32 torr. Questo effetto è ulteriormente rinforzato dall'aumentata formazione di acidi nel metabolismo del feto e verosimilmente anche della madre. La tensione di CO2 nel sangue venoso dell'utero è di circa 40 torr ed è con questo sangue che avvengono gli scambi del sangue fetale. Per il feto questo sangue svolge dunque lo stesso ruolo dell'aria alveolare nel periodo post-natale. La corrispondenza delle tensioni di CO2 dimostra che il feto per quanto riguarda lo scambio di CO2 non si trova in una condizione peggiore di quella dell'adulto. Infine, per lo scambio di ossigeno l'iperventilazione gravidica non ha un ruolo così determinante, perché la saturazione del sangue arterioso della madre non può subire un forte aumento con l'iperventilazione.
6. Interrelazioni tra respirazione ed escrezione renale nel mantenimento del normale equilibrio acido-base
Con l'introduzione di cibi nell'organismo, pervengono nel sangue sostanze acide e alcaline; se ne originano anche nel metabolismo, con prevalenza, in generale, di quelle acide. Quando acidi sono introdotti improvvisamente nell'organismo, entra in azione il meccanismo chemostatico respiratorio precedentemente descritto, che protegge le cellule del cervello dalle modificazioni dell'acidità del liquido che le circonda. Poiché, però, avviene uno scambio di ioni tra questo liquido e il sangue, si potrebbe arrivare a una perdita o a un accumulo di basi tampone (bicarbonati) nel liquido extracellulare, così come l'introduzione di acidi produce inizialmente una perdita di basi tampone nel sangue e l'introduzione di alcali produce un eccesso di esse.
La restituzione a un valore normale delle basi tampone del sangue, e di conseguenza nei liquidi extracellulari, avviene attraverso il rene, che possiede tutta una serie di possibilità di escrezione dell'eccesso di acidi o di basi, la cui discussione in questa sede porterebbe troppo lontano. In questo modo il meccanismo chemostatico della respirazione assicura l'omeostasi dell'acidità del liquido extracellulare del cervello nelle modificazioni acute dell'equilibrio acido-base del sangue. La correzione dell'alterazione avviene in definitiva attraverso il rene.
7. Il controllo automatico della respirazione a servizio della costanza del mezzo interno delle cellule nervose
La costanza del pH nel liquido extracellulare del cervello è di così grande importanza per l'organismo da avere sviluppato durante l'evoluzione la formazione di un meccanismo di regolazione estremamente preciso e complesso. Al perché di questa importanza può essere data la seguente risposta: si tratta, in effetti, di un meccanismo parziale, che assieme ad altri processi regolatori (per es. la temperatura, la concentrazione di potassio e di sodio, la pressione osmotica) coopera per realizzare un ambiente costante per le cellule nervose. Se si considerano questi meccanismi come elementi del circuito di un calcolatore, si comprende come dalla costanza delle condizioni dipenda la riproducibilità, vale a dire la stessa attendibilità, di questo strumento d'importanza vitale. Deviazioni di piccola entità vengono prese in considerazione soltanto nell'interesse di funzioni vitali, come per es. l'apporto di ossigeno o la riproduzione.
bibliografia
Acker, H., Fidone, S., Pallot, D., Eyzaguirre, C., Lübbers, D. W., Torrance, R. W. (a cura di), Chemoreception in the carotid body, Berlin-New York 1977.
Acker, H., Lübbers, D. W., Purves, M. J., Local oxygen tension field in the glomus caroticum of the cat and its change at changing arterial PO2 in ‟Pflügers Archiv", 1971, CCCXXIX, pp. 136-155.
Åström, A., On the action of combined carbon dioxide excess and oxygen deficiency in the regulation of breathing, in ‟Acta physiologica scandinavica", 1952, XXVII, suppl. 98, pp. 7-61.
Berkenbosch, A., Adan, A. J. M., Influence of CSF calcium concentration on the ventilatory response to CO2 and O2, in ‟Pflügers Archiv", 1974, CCCXLVIII, pp. 33-50.
Berndt, J., Fenner, A., Berger, K., Influence of calcium and magnesium on the respiratory response to changes in CSF pH, in ‟Respiration physiology", 1969, VII, pp. 216-229.
Bouverot, P., Flandrois, R., Puccinelli, R., Dejours, P., Étude du rôle des chémorécepteurs artériels dans la régulation de la respiration pulmonaire chez le chien éveillé, in ‟Archives internationales de pharmacodynamie et de thérapie", 1965, CLVII, pp. 253-271.
Comroe, J. H., Physiology of respiration, Chicago 1965.
Cropp, J. H., Comroe, H. H., Role of mixed venous blood pCO2 in respiratory control, in ‟Journal of applied physiology", 1961, XVI, pp. 1029-1033.
Cunningham, D. J. C., Some quantitative aspects of the regulation of human respiration in exercise, in ‟British medical bulletin", 1963, XIX, pp. 25-30.
Cunningham, D. J. C., Hey, E. N., Lloyd, B. B., The effect of intravenous infusion of noradrenaline on the respiratory response to carbon dioxide in man, in ‟Quarterly journal of experimental physiology", 1958, XLIII, pp. 394-399.
Cunningham, D. J. C., O'Riordan, J. L. H., The effect of a rise in the temperature of the body on the respiratory response to carbon dioxide at rest, in ‟Quarterly journal of experimental physiology", 1957, XLII, pp. 329-345.
Daly, B., Lambertsen, C. J., Schweitzer, A., Observations on the volume of blood flow and oxygen utilization of the carotid body in the cat, in ‟Journal of physiology", 1954, CXXV, pp. 67-89.
De Castro, F., Sur la structure et l'innervation du sinus carotidien de l'homme et des Mammifères. Nouveaux faits sur l'innervation et la fonction du glomus caroticum. Études anatomiques et physiologiques, in ‟Trabajos del Instituto Cajal de Investigaciones Biológicas", 1928, XXV, pp. 331-380.
Dejours, P., Labrousse, Y., Raynaud, J., Girard, T., Teiclac, A., Stimulus oxygène et la ventilation au repos et au cours de l'exercise musculaire, à basse altitude (50 m) chez l'homme, in ‟Revue française d'études cliniques et biologiques", 1958, III, pp. 105-129.
Dempsey, J. A., Forster, H. V., Do Pico, G. A., Ventilatory acclimatization to moderate hypoxemia in man, in ‟Journal of clinical investigation", 1974, LIII, pp. 1091-1100.
Dev, N. B., Loeschcke, H. H., A cholinergic mechanism involved in the respiratory chemosensitivity of the medulla oblongata in the cat, in ‟Pflügers Archiv", 1979, CCCLXXIX, pp. 29-36.
Eldridge, F. L., The importance of timing on the respiratory effects of intermittent carotid sinus nerve stimulation, in ‟Journal of physiology", 1972, CCXXII, pp. 297-318.
Euler, C. von, Söderberg, U., Medullary chemosensitive receptors, in ‟Journal of physiology", 1952, CXVIII, pp. 545-554.
Euler, U. S. von, Liljestrand, G., Zottermann, Y., The excitation mechanism of the chemoreceptors of the carotid body, in ‟Skandinavisches Archiv für Physiologie", 1940, LXXXIII, pp. 132-152.
Fencl, V., Vale, J. R., Broch, A., Respiration and cerebral blood flow in metabolic acidosis and alkalosis in humans, in ‟Journal of applied physiology", 1969, XXVII, pp. 67-76.
Flourens, P., Recherches expérimentales sur les fonctions du système nerveux dans les animaux vertébrés, Paris 1842.
Frédéricq, L., Sur la cause de l'apnée, in ‟Archives de biologie", 1901, XVII, pp. 561-563.
Fukuda, Y., Loeschcke, H. H., A cholinergic mechanism involved in the neuronal excitation by H+ in the respiratory chemosensitive structures of the ventral medulla oblongata of cats in vitro, in ‟Pflügers Archiv", 1979, CCCLXXIX, pp. 125-135.
Gesell, R., A neurophysiological interpretation of the respiratory act, in ‟Ergebnisse der Physiologie", 1940, XLIII, pp. 477-539.
Gesell, R., Lapides, J., Levin, N., The interaction of central and peripheral chemical control of breathing, in ‟American journal of physiology", 1940, CXXX, pp. 155-169.
Gray, J. S., Pulmonary ventilation and its physiological regulation, Springfield, Ill., 1950.
Grodins, F. S., Gray, J. S., Schroeder, K. R., Novins, A. L., Jones, R. W., Respiratory responses to CO2 inhalation. A theoretical study of a nonlinear biological regulator, in ‟Journal of applied physiology", 1954, VII, pp. 283-308.
Haldane, J. S., Respiration, Oxford 1935.
Haldane, J. S., Priestley, J. G., The regulation of the lung ventilation, in ‟Journal of physiology", 1905, XXXII, pp. 225-266.
Heerhaber, J., Loeschcke, H. H., Westphal, U., Eine Wirkung des Progesterons auf die Atmung, in ‟Pflügers Archiv", 1948, CCL, pp. 42-55.
Hess, A., Electron microscopic observations of normal and experimental cat carotid bodies in arterial chemoreceptors, in Arterial chemoceptors. Proceedings of the Wates Foundation Symposium (a cura di R. W. Torrance), Oxford-Edinburgh 1968, pp. 51-56.
Heymans, C., Bouckaert, J. J., Les chémorécepteurs du sinus carotidien, in ‟Ergebnisse der Physiologie", 1939, XLI, pp. 28-55.
Heymans, C., Neil, E., Reflexogenic areas of the cardiovascular system, London 1958.
Heymans, J. F., Heymans, C., Sur les modifications directes et sur la régulation réflexe de l'activité du centre respiratoire de la tête isolée du chien, in ‟Archives internationales de pharmacodynamie et de thérapie", 1927, XXXIII, pp. 273-370.
Hornbein, T. F., Roos, A., Specificity of H ion concentration as a carotid chemoreceptor stimulus, in ‟Journal of applied physiology", 1963, XVIII, pp. 580-584.
Kronenberg, R. S., Cain, S. M., Effects of acetazolamide and hypoxia on cerebrospinal fluid bicarbonate, in ‟Journal of applied physiology", 1968, XXIV, pp. 17-20.
LeGallois, C. J. J., Expériences sur le principe de la vie, notamment sur celui des mouvements du coeur, et sur le siège de ce principe, Paris 1812.
Leusen, I., Influence du pH du liquide céphalo-rachidien sur la respiration, in ‟Experientia", 1950, VI, p. 272.
Lloyd, B. B., The chemical stimulus to breathing, in ‟British medical bulletin", 1963, XIX, pp. 10-14.
Lloyd, B. B., Jukes, M. G. M., Cunningham, D. J. C., The relation between alveolar oxygen pressure and the respiratory response to carbon dioxide in man, in ‟Quarterly journal of experimental physiology", 1958, XLIII, pp. 214-227.
Loeschcke, H. H., Koepchen, H. P., Beeinflussung von Atmung und Vasomotorik durch Einbringen von Novocain in die Liquorräume, in ‟Pflügers Archiv", 1958, CCLXVI, pp. 611-627.
Loeschcke, H. H., Koepchen, H. P., Gertz, K. H., Über den Einfluss von Wasserstoffionenkonzentration und CO2 Druck im Liquor cerebrospinalis auf die Atmung, in ‟Pflügers Archiv", 1958, CCLXVI, pp. 569-585.
Loeschcke, H. H., Lattre, J. de, Schläfke, M. E., Trouth, C. O., Effects on respiration and circulation of electrically stimulating the ventral surface of the medulla oblongata, in ‟Respiration physiology", 1970, X, pp. 184-197.
Loeschcke, H. H., Sugioka, K., pH of cerebrospinal fluid in the cisterna magna and on the surface of the choroid plexus of the 4th ventricle and its effect on ventilation in experimental disturbances of acid-base balance. Transients and steady states, in ‟Pflügers Archiv", 1969, CCCXII, pp. 161-188.
Markwald, M., Die Bedeutung des Mittelhirns für die Athmung, in ‟Zeitschrift für Biologie", 1890, XXVI, pp. 259-289.
Middendorf, T., Loeschcke, H. H., Mathematische Simulation des Respirationssystems, in ‟Journal of mathematical biology", 1976, III, pp. 149-177.
Mitchell, R. A., Loeschcke, H. H., Massion, W. H., Severinghaus, J. W., Respiratory responses mediated through superficial chemosensitive areas on the medulla, in ‟Journal of applied physiology", 1963, XVIII, pp. 523-533.
Mitchell, R. A., Loeschcke, H. H., Severinghaus, J. W., Richardson, B. W., Massion, W. H., Regions of respiratory chemosensitivity on the surface of the medulla, in ‟Annals of the New York Academy of Sciences", 1963, CIX, pp. 661-681.
Nielsen, M., Untersuchungen über die Atemregulation beim Menschen, in ‟Skandinavisches Archiv für Physiologie", 1936, LXXIV, suppl. 10, pp. 83-208.
Nielsen, M., Smith, H., Studies on the regulation of respiration in acute hypoxia, in ‟Acta physiologica scandinavica", 1951, XXIV, pp. 293-305.
Pappenheimer, J. R., Fencl, V., Heisey, S. R., Held, D., Role of cerebral fluids in control of respiration as studied in anesthetized goats, in ‟American journal of physiology", 1965, CCVIII, pp. 436-450.
Pflüger, F., Über die Ursache der Atembewegungen, sowie der Dyspnoe und Apnoe, in ‟Pflügers Archiv", 1868, I, pp. 61-106.
Pitts, R. F., Orgnisation of the respiratory center, in ‟Physiological reviews", 1946, XXVI, pp. 609-630.
Robin, E. D., Whaley, R. D., Crump, C. H., Bickelmann, A. G., Travis, D. M., Acid-base relations between fluid and arterial blood with special reference to control of ventilation, in ‟Journal of applied physiology", 1958, XIII, pp. 385-392.
Salmoiraghi, G. G., Burns, B. D., Localization and patterns of discharge of respiratory neurones in brain-stem of cat, in ‟Journal of neurophysiology", 1960, XXIII, pp. 2-13.
Schläfke, M. E., Origin and afferent modification of respiratory drive from ventral medullary areas, in Nervous control mechanism in regular, periodic and irregular breathing. Physiological and clinical aspects in the adult and perinatal states (a cura di C. von Euler), Stockholm 1979.
Schläfke, M. E., Pokorski, M., See, W. R., Prill, R. K., Loeschcke, H. H., Chemosensitive neurons on the ventral medullary surface, in ‟Bulletin of physiopathological respiration", 1975, XI, pp. 277-284.
Schläfke, M. E., See, W. R., Loeschcke, H. H., Ventilatory response to alterations of H+ concentration in small areas of the ventral medullary surface, in ‟Respiration physiology", 1970, X, pp. 198-212.
Schmidt, C. F., Comroe, J. H., Functions of carotic and aortic bodies, in ‟Physiological reviews", 1940, XX, pp. 115-157.
Severinghaus, J. W., Mitchell, R. A., Richardson, B. W., Singer, M. M., Respiratory control at high altitude suggesting active transport regulation of CSF pH, in ‟Journal of applied physiology", 1963, XVIII, pp. 1155-1166.
Shimada, K., Trouth, C. O., Loeschcke, H. H., Von der Wasserstoffionenkonzentration des Liquor abhängige Aktivität von Neuronen im Gebiet der chemosensiblen Zonen der Medulla oblongata, in ‟Pflügers Archiv", 1969, CCCXII, p. R55.
Torrance, R. W., Prolegomena in Arterial chemoceptors. Proceedings of the Wates Foundation Symposium (a cura di R. W. Torrance), Oxford-Edinburgh 1968, pp. 1-40.
Trouth, C. O., Loeschcke, H. H. Berndt, J., A superficial substrate on the ventral surface of the medulla oblongata influencing respiration, in ‟Pflügers Archiv", 1973, CCCXXXIX, pp. 135-152.
Trouth, C. O., Loeschcke, H. H., Berndt, J., Histological structures in the chemosensitive regions of the ventral surface of the cat's medulla oblongata, in ‟Pflügers Archiv", 1973, CCCXXXIX, pp. 171-183.
Verstraeten, J. M., Influences de la concentration en ions potassium, calcium et magnésium du liquide céphalo-rachidien sur la respiration, in ‟Revue belge de pathologie et de médecine expérimentale", 1950, XX, pp. 1-21.
Volkmann, A. W., Über die Bewegungen des Athmens und Schluckens mit besonderer Berücksichtigung neurologischer Streitfragen, in ‟Archiv für Anatomie, Physiologie und wissenschaftliche Medizin", 1841, pp. 342-360.
Winterstein, H., Die Regulierung der Atmung durch das Blut, in ‟Pflügers Archiv", 1911, CXXXVIII, pp. 167-184.
Winterstein, H., Die Reaktionstheorie der Atmungsregulation, in ‟Pflügers Archiv", 1921, CXCI, pp. 184-185.
Winterstein, H., Atmungsregulation und Reaktionsregulation, in ‟Naturwissenschaften", 1923, XI, pp. 625-630.
Winterstein, H., Gökhan, N., Ammoniumchlorid-Acidose und Reaktionstheorie der Atmungsregulation, in ‟Archives internationales de pharmacodynamie et de thérapie", 1953, XCIII, pp. 212-232.