
Shock e stress
Shock e stress
SOMMARIO: 1. Definizioni: a) stress e aggressione; b) sindrome lesionale e sindrome da reazione; c) i concetti di omeostasi e di mezzo di difesa; d) il concetto di adattamento; e) l'inibizione dell'azione. 2. Confronto fra shock e stress; generalità: a) ruolo del fattore tempo. Apprendimento e memoria; b) il concetto di percorso biologico dell'idrogeno. 3. Gli stati di shock: a) sintomatologia; b) fenomeni generali da reazione; c) integrazione dei processi metabolici e fisiopatologici; d) i fenomeni lesionali e la loro partecipazione agli stati di shock. 4. Gli stati di stress: a) l'inibizione dell'azione; b) inibizione dell'azione e asse ipotalamo-ipofiso-surrenale (IPA); c) ormoni e comportamento; d) integrazione dei fatti sperimentali nel concetto di stress; e) esame schematico delle conseguenze dello stress. 5. Il trattamento: a) trattamento degli stati di shock; b) trattamento degli stati di stress. □ Bibliografia.
1. Definizioni
a) Stress e aggressione
Come notava H. Selye (v. Selye e Heuser, 1956), il termine ‛stress' era già di uso corrente nella lingua inglese molto prima che lo stesso Selye descrivesse la ‛reazione d'allarme' nel 1936. Si parlava di stress nervoso o muscolare, quando il sistema nervoso o quello muscolare venivano sovraccaricati da un qualche stimolo, indipendentemente dallo stimolo in gioco. Il termine comprendeva sia la manifestazione sia la causa di questo sovraccarico. In realtà, però, il termine ‛stress' era più comunemente usato per indicare la causa che le relative conseguenze. Lo stesso termine era anche usato da molto tempo in fisica per descrivere l'interazione tra una forza e la resistenza contrapposta. Per esempio, pressione e tensione possono produrre uno ‛stress' nella materia inanimata. Per Selye la risposta aspecifica sarebbe l'equivalente biologico di questo stress fisico. Egli propone quindi di definire lo stress come ‟la somma di tutti i fenomeni non specifici (lesioni e reazioni comprese) provocati da una funzione (sforzo muscolare, per esempio) o da un'azione lesiva, ivi compresi i fenomeni biologici necessari al ristabilimento dello stato normale". Un ‛agente' è, secondo Selye, ‟ciò che agisce su un bersaglio biologico" (sono ‛agenti' il calore, l'elettricità, gli ormoni, le droghe, le forze meccaniche). Uno stressor, o agente stressante, è ciò che è in grado di produrre uno stress. Selye distingue quindi lo stress localizzato, che provoca la sindrome locale di adattamento (LAS), dallo stress sistemico o stimolo d'allarme, capace di produrre la sindrome generale di adattamento (GAS). Ambedue queste sindromi evolvono nel tempo. La GAS comprende: a) la fase della reazione d'allarme; b) la fase della resistenza; c) la fase dell'esaurimento. La LAS sarebbe una reazione adattativa con una fase regressiva (degenerazione, necrosi) e una fase progressiva (infiammazione, ipertrofia, iperplasia).
Va comunque notato che Selye era in primo luogo un endocrinologo e che quindi la sua GAS descrive innanzitutto una reazione endocrina aspecifica. Se chiama in causa il sistema nervoso, Selye lo fa, il più delle volte, per cercare di sapere come i neurormoni (adrenalina, noradrenalina) liberati alla periferia possano influenzare la liberazione, da parte del sistema nervoso centrale, di ormoni ipofisari (ACTH), che provocano a loro volta la secrezione di ormoni corticosurrenali; nella realtà, viceversa, l'attività del sistema nervoso centrale, nelle risposte a variazioni intervenute nell'ambiente, domina, il più delle volte, le reazioni nervose periferiche ed endocrine.
Il merito di Selye sta nell'aver mostrato l'assenza di specificità nelle reazioni organiche ad aggressioni specifiche e l'importante ruolo svolto dall'asse ipotalamo-ipofiso-surrenale in queste reazioni aspecifiche. Ma anche questo concetto è stato criticato, specialmente da Mason (v., 1971), che ha dimostrato l'esistenza di risposte endocrine differenti in corrispondenza di numerosi stress diversi.
Cercheremo in primo luogo, ripercorrendo le tappe di sviluppo delle nostre conoscenze, di precisare certe definizioni che, per l'ambiguità dei termini implicati, possono avere un significato suscettibile di trarre in inganno.
Abbiamo già visto, a proposito del termine ‛stress', come, malgrado gli sforzi di Selye per precisarne il significato, la ‛deriva' semantica sia importante. Al giorno d'oggi non si parla praticamente più di stress a proposito di un trauma circolatorio, per esempio, mentre si avrà la tendenza a riferire il termine a un'aggressione psicosociale. È questo il motivo per cui solo eccezionalmente abbiamo utilizzato il termine ‛stress', preferendo sostituirlo con i termini ‛aggressione' e ‛reazione organica all'aggressione' (ROPA, ‛reazione oscillante postaggressiva': v. Laborit, Réaction.. 1952). Lo stato di shock, come vedremo, non è altro che una forma estrema e caricaturale di aggressione e reazione all'aggressione combinate.
Per ‛aggressione' intendo l'azione su un sistema vivente di una certa quantità di energia cinetica (meccanica, termica, chimica, radiante), che tende ad accrescere l'entropia del sistema (l'entropia è la tendenza al livellamento termodinamico) ovvero, in altre parole, a turbarne la struttura. Una ‛struttura' è l'insieme delle relazioni intercorrenti tra gli elementi di un insieme (di molecole, di cellule, di tessuti, di organi, di sistemi); ogni livello di organizzazione deve essere studiato nei suoi rapporti con gli insiemi che lo contengono e con quelli che esso contiene o ‛comprende'. Al limite l'organismo non potrà essere isolato dal suo ambiente, in particolare dal suo ambiente sociale.
Esistono aggressioni ‛acute', di elevata intensità ma di breve durata, e aggressioni ‛croniche', di minore intensità ma protratte nel tempo.
Dobbiamo ora analizzare i concetti generali relativi alle aggressioni ‛fisiche' e precisare i rapporti intercorrenti fra questo tipo di aggressioni e le aggressioni ‛psicosociali'.
b) Sindrome lesionale e sindrome da reazione
Le aggressioni provocano generalmente due tipi differenti di fenomeni, strettamente collegati: a) l'azione ‛locale' produce nella zona colpita complessi disordini tessutali, limitati ma destinati ad avere, in un secondo tempo, una ripercussione generale più o meno importante; ho riunito tutti questi fenomeni locali sotto il termine ‛sindrome lesionale' (v. Laborit, Réaction..., 1952); b) l'organismo aggredito mette in atto un insieme non meno complesso di reazioni, che finiscono per influenzare l'intero equilibrio fisiobiologico: ho chiamato questi fenomeni ‛fenomeni da reazione', o, con termine cumulativo, ‛sindrome da reazione'; si tratta di una sindrome molto simile alla sindrome di adattamento generale aspecifica di Selye.
La parte assunta da ciascuna delle sindromi descritte nell'ambito del comportamento globale di un organismo soggetto a un'aggressione varia a seconda del tipo di aggressione e dello stato precedente - la storia - dell'organismo: per esempio, dipende dalla sua memoria immunitaria o nervosa. Ma quando la sindrome lesionale locale, nella zona colpita dall'agente aggressivo, non esiste, come nel caso dell'aggressione ‛psicosociale', dobbiamo chiederci quali siano i meccanismi che, malgrado ciò, portano alla reazione aspecifica. In effetti, già nel 1948 J. Trueta rilevò la comparsa di un'esclusione corticorenale nel gatto messo semplicemente al cospetto di un cane.
c) I concetti di omeostasi e di mezzo di difesa
Cannon (v., 1932) desume il suo concetto di ‛omeostasi' dalla nozione di ‟mantenimento della costanza delle condizioni di vita nell'ambiente interno", formulata da Claude Bernard (v., 1878). Ancora fino a pochi anni fa esistevano terapeuti che si ponevano come unico scopo quello di riportare i principali parametri dell'ambiente interno a valori normali. Ci si concentrava sull'ambiente interno e contemporaneamente si ignorava la cellula, in quanto si nutriva un'assoluta fiducia nella capacità delle leggi dell'osmosi di regolare gli scambi fra cellula e ambiente interno, e si ignoravano ancora le principali regolazioni metaboliche e i fenomeni di scambio che esse controllano a livello delle membrane. Questo atteggiamento derivava anche dall'interpretazione finalistica dei fatti fisiopatologici. Il termine ‛mezzo di difesa' dell'organismo, che comprendeva implicitamente la reazione organica all'aggressione (nello stesso senso in cui Selye parlava di reazione d'allarme e di fase di resistenza), fu associato alla nozione di ‛mantenimento dell'omeostasi'. Così, dal momento che l'omeostasi era necessaria alla vita, la reazione all'aggressione doveva rappresentare un mezzo di difesa dell'omeostasi. Viceversa i fatti dimostrano che questa reazione è all'origine dell'iperglicemia, dell'iperlattacidemia, dell'acidosi, dell'ipercoagulabilità, dell'iperazotemia, ecc., che sono perturbazioni profonde della stabilità dell'ambiente interno, dell'omeostasi. Questa reazione, quindi, non è omeostatica nel senso dell'omeostasi dell'ambiente interno.
d) Il concetto di adattamento
Un altro termine estremamente ambiguo è quello di ‛adattamento', che purtroppo Selye utilizzò. Sentendolo pronunciare, si è portati ad attribuirgli un significato positivo: infatti, un organismo ben adattato al suo ambiente, così come un individuo, un essere sociale, ben adattato al suo ambiente sociale, non può che attirare simpatia, rispetto o ammirazione. In realtà, i veri fenomeni di adattamento sono del tutto eccezionali. Per quel che riguarda la fisiologia, per esempio, si può citare il caso dei gruppi etnici delle Ande, che vivono a circa 4.000 m sul livello del mare; si è potuto constatare che gli individui appartenenti a queste popolazioni presentano una microcircolazione miocardica notevolmente più sviluppata di quella dei soggetti che vivono al livello del mare, il che permette ai primi di essere esposti a una debole pressione parziale di ossigeno e di realizzare, malgrado ciò, un importante lavoro meccanico. In questo caso si può definire l'adattamento come l'insieme delle trasformazioni che consentono a un organismo di conservare la stessa autonomia motoria, a lungo termine, in un ambiente differente. Viceversa, quando si passa dalla fisiologia al comportamento, sembra che si confonda, il più delle volte, l'‛adattamento' con la ‛sottomissione' (v. Laborit, 1954). Un organismo ‛reagisce' alle variazioni dell'ambiente che non si rivelano conformi al mantenimento della sua omeostasi, ed è tramite la sua ‛attività motoria' sull'ambiente e nell'ambiente che tenterà di controllarne le caratteristiche. Già Cannon (v., 1932) aveva capito perfettamente che il buon esito dell'azione di controllo sull'ambiente dipende da due tipi di comportamento, che si possono descrivere globalmente come ‛fuga' e ‛lotta': azioni che mirano a evitare o a sopprimere le variazioni ambientali pericolose per l'omeostasi. Se la fuga e la lotta si rivelano efficaci, la reazione organica che ha permesso l'attività motoria non porterà all'adattamento ma, al contrario, alla conservazione delle caratteristiche biologiche e comportamentali precedenti. Se invece sia la fuga sia la lotta risultano inefficaci, allora la reazione organica comporterà l'‛inibizione dell'azione' (v. Laborit, 1974), l'inibizione del comportamento, e questo effetto non è più un adattamento nel senso del mantenimento diretto dell'omeostasi. Le conseguenze neuroendocrine di un'inibizione che si protragga nel tempo consistono in perturbazioni profonde e pericolose delle caratteristiche abituali dell'equilibrio biologico. Malgrado ciò, questa sottomissione all'ambiente, in particolare all'ambiente socioculturale, sarà generalmente considerata alla stregua di un adattamento, mentre in realtà sarà la fonte principale di processi fisiopatologici. L'errore che tanto spesso si commette nell'uso del concetto di ‛adattamento all'ambiente' deriva da una confusione fra i diversi ‛livelli di organizzazione‛. Se si considera un individuo ‛ben adattato al suo ambiente', lo si giudica dal punto di vista del suo comportamento, e spesso i disturbi patologici che presenta cono attribuiti a microbi o a virus, alla ‛diatesi'. I disordini biologici, che per parecchio tempo possono non manifestarsi, non sono imputati alla sua sottomissione all'ambiente, cioè al suo disadattamento biologico. O, per meglio dire, saranno attribuiti allo ‛stress della vita moderna', ma non all'impossibilità, di cui spesso il soggetto non si rende conto, di controllare il proprio ambiente sociale per mantenere la propria omeostasi, né all'inibizione dell'azione gratificante neppure lo spostamento dell'azione in un altro sistema di ricompensa basterà, il più delle volte, a far scomparire le turbe biologiche, come si è po tuto constatare in un recente esperimento.
Per parecchio tempo (v. Laborit, Réaction..., 1952) ho pensato che il quadro patologico potesse essere determinato dalla messa in atto di un ‛sistema di difesa'. Insistevo sul concetto fondamentale che l'unico vero sistema di difesa è quello che permette di mantenere l'autonomia motoria, di realizzare la fuga e la lotta che rendono possibile il recupero di un ambiente in cui l'omeostasi è conservata o ritrovata. Ma a questo scopo bisogna attuare un nuovo ‛programma'. Le reazioni organiche, che in circostanze normali assicurano la costanza delle condizioni di vita nell'ambiente interno e, di conseguenza, quella dell'attività metabolica dei tessuti, devono ora assicurare ‛innanzitutto' l'autonomia motoria rispetto a un ambiente pericoloso, perturbando temporaneamente l'omeostasi. Sono quindi stato indotto a distinguere un'omeostasi ‛ristretta' all'ambiente interno e un'omeostasi ‛generalizzata' al livello dell'organizzazione superiore dell'organismo nel suo ambiente. La reazione organica all'aggressione è bensì una reazione fisiologica, purché non si confonda il suo meccanismo con quello che garantisce l'omeostasi ristretta.
A quel tempo pensavo che questa reazione fosse diventata inadatta alle circostanze della vita moderna, in cui l'‛altro', l'elemento più importante dell'ambiente, non può essere né evitato né combattuto. Pensavo che questi ‛mezzi di difesa', inutili, ma ancora funzionanti, fossero all'origine dei disordini fisiopatologici. Questa opinione è attualmente accettata e negli atti del simposio Society, stress and diseases, organizzato a Stoccolma nel 1971 da L. Levi, si legge che ‟certe situazioni psicosociali sono potenzialmente patogene, perché inducono in determinati individui reazioni adattative inadeguate [...]. L'uomo moderno, nel contesto delle società urbane tecnologiche, si trova a vivere in condizioni sociali complesse, per affrontare le quali il suo repertorio comportamentale sociale innato spesso si rivela inadatto" (v. Levi, 1971).
e) L'inibizione dell'azione
Oggi sono indotto a rivedere l'opinione espressa nel paragrafo precedente. Penso di aver dimostrato (v. Laborit, 1974 e L'inhibition..., 1979) che non è una reazione diventata inadatta che sarebbe all'origine delle complicazioni lisiopatologiche dello stress, ma un'altra reazione, corrispondente a meccanismi centrali e periferici diversi, che comanda ‛l'inibizione dell'azione' quando l'azione si dimostra impossibile. Questa reazione permette, in effetti, se messa in atto temporaneamente, di evitare la distruzione dell'aggredito da parte dell'aggressore. Permette all'aggredito di farsi dimenticare, di apparire ‛adattato' evitando il confronto, di non consumare nella lotta le sue riserve, di conservarle per il momento in cui potrà agire. Ciò che la rende pericolosa è il suo protrarsi nel tempo, quando le condizioni ambientali persistono senza cambiare. Capace di assicurare momentaneamente la sopravvivenza, questa reazione diventa una minaccia per la stessa sopravvivenza del soggetto se la soluzione che fornisce, l'inazione, l'attesa in uno stato di tensione del momento in cui l'azione diventerà Possibile non risolve il problema posto dall'ambiente. Dal punto di vista biochimico e neurofisiologico del sistema nervoso centrale, i suoi meccanismi sono diversi da quelli che assicurano la sopravvivenza controllando attivamente l'ambiente tramite la fuga e la lotta.
2. Confronto fra shock e stress; generalità
a) Ruolo del fattore tempo. Apprendimento e memoria
Abbiamo visto che una sindrome lesionale ingenera il più delle volte una reazione organica (ROPA), vale a dire un insieme di perturbazioni dell'omeostasi ristretta o sindrome da reazione. Questa preserva l'autonomia motoria dell'organismo nell'ambiente consentendogli di produrre uno sforzo supplementare: il sistema nervoso deve poter riconoscere il pericolo e dirigere la coordinazione dei muscoli scheletrici nella fuga o nella lotta. I muscoli devono fornire uno sforzo considerevole per realizzare l'uno o l'altro di questi comportamenti, e quindi devono ricevere una massa sanguigna più consistente, per il loro approvvigionamento energetico e per l'eliminazione dei prodotti del loro accresciuto metabolismo. La pompa cardiaca deve allora aumentare la sua gettata e così pure la ventilazione polmonare risultare più efficace. Dato che la massa sanguigna non si può modificare, se ne determinerà una nuova ripartizione, attraverso una vasocostrizione (diminuzione del calibro dei vasi), che riduce il letto circolatorio negli organi inutili alla fuga o alla lotta, mentre quello dei vasi degli organi impegnati nella fuga o nella lotta, che in seguito elencheremo, risulterà accresciuto. Tutta l'area splancnica (della cavità addominale) e la pelle si troveranno in uno stato di ipossia, che non potrà perdurare che per breve tempo. È necessario quindi che la fuga e la lotta risultino rapidamente efficaci e capaci di ripristinare le caratteristiche dell'ambiente. Si capisce come questa reazione sia dominata dal sistema simpatico-agrenergico.
L'evoluzione nel tempo è contemporaneamente una caratteristica e un fattore della sindrome da shock. Si può morire d'asfissia acuta, per strangolamento o per emorragia, senza, che uno shock abbia il tempo di instaurarsi, mentre un'emorragia leggera, ma prolungata, costituisce un potente fattore di shock. Questi fatti mi hanno portato a descrivere (v. Laborit, Réaction..., 1952) una fase iniziale depressiva di ‛collasso' immediato, che non è ancora uno stato di shock, seguita da una fase di reazione, assimilabile alla fase di contro-shock di Selye, e da una fase terminale caratterizzata dall'irreversibilità e dalla morte. Ma mentre Selye considera quest'ultima come una fase di esaurimento di fallimento, da parte della reazione, nel ristabilire l'omeostasi, io, al contrario, sono portato a considerarla come la conseguenza della reazione e della sua durata.
Sono state descritte le più svariate forme di shock. A seconda dell'eziologia (del tipo di aggressione), sono stati descritti shock traumatici, emorragici, tossinfettivi, metabolici, anafilattici o respiratori. Si sono anche descritti shock da ustioni, ostetrici, medici, anestetici, operatori, da rimozione del laccio emostatico, da schiacciamento, ecc. A seconda del loro presunto meccanismo, si è parlato di shock neurogeni, vasomotori (istaminici), cardiogeni (infarti). In tutte queste forme, tuttavia, i meccanismi fondamentali restano gli stessi e costituiscono l'unità della sindrome.
Così, quando una fonte di energia (meccanica, termica, radiante, chimica) entra in rapporto con un organismo provocandogli un danno strutturale locale o diffuso, cioè vere e proprie lesioni, la reazione organica indotta, anch'essa violenta e passeggera, richiederà un trattamento distinto, ancorchè concomitante, rispetto a quello della lesione. Ma questa reazione dipenderà anche in parte, e spesso in maniera più o meno evidente, dalla ‛storia precedente' del soggetto, cioè dai suoi rapporti passati con i diversi ambienti in cui ha vissuto.
Questi precedenti storici diventano fattori della massima importanza quando si verifica ciò che si è convenuto di chiamare ‛stress psicosociale'. In questo caso mancano lesioni dovute all'azione diretta di un agente aggressore. Le lesioni compariranno spesso più tardi, dopo un periodo più o meno lungo, e saranno una conseguenza della reazione. Questo tipo di reazione, come ho già rilevato, è, secondo me, diversa dalla precedente, che consentiva la fuga o la lotta. Quest'altra reazione, viceversa, porta all'inibizione dell'azione, le cui conseguenze neuro-endocrino-metaboliche costituiscono quello che potremmo chiamare ‛il terreno' fisiopatologico. Questo è dominato dai processi di memoria, che condizionano il comportamento di un organismo nei confronti dell'ambiente, a tutti i suoi livelli di organizzazione. Entrano in gioco la memoria genetica, la memoria immunitaria e soprattutto, prevalente rispetto alle precedenti, la memoria nervosa. Tramite quest'ultima il soggetto ricorda le esperienze passate, piacevoli e spiacevoli, condizione indispensabile per cercare di rivivere le prime e di evitare le seconde. La memoria nervosa ci permette di accumulare gli ‛elementi' su cui basare un nuovo comportamento, grazie all'immaginazione, e, soprattutto, registra le azioni inefficaci o dolorose e quelle che sono state punite, o rischiano di esserlo, dall'ambiente socioculturale. È la memoria nervosa che mobilita le aree cerebrali che comandano l'inibizione dell'azione e agiscono sul sistema neuroendocrino per determinare un'‛attesa in uno stato di tensione' del momento di agire. Numerosi elementi ci permettono di affermare che questa reazione si automantiene finché l'azione gratificante non interrompe il circolo vizioso.
Riassumendo, nel passare dallo shock allo stress si va da una sindrome in cui, generalmente, la lesione primaria fa scattare una reazione aspecifica, che deve favorire la lotta o la fuga, a una sindrome in cui la lesione è la conseguenza di una reazione messa in atto dalla consapevolezza dell'inefficacia dell'azione. In entrambi i casi l'azione risulta inadeguata a controllare le caratteristiche dell'ambiente; ma nel caso dello shock la sindrome evolve rapidamente senza che vi sia stato un apprendimento precedente dell'inefficacia dell'azione. La risposta nervosa fa essenzialmente appello all'ipotalamo e al tronco cerebrale. Nello stress psicosociale il ricordo dell'inefficacia dell'azione svolge un ruolo predominante e l'intervento del sistema limbico della corteccia associativa costituisce un fattore determinante nell'insorgere delle perturbazioni fisiobiologiche.
Così, tenuto conto della difficoltà incontrata dallo stesso Selye nel dare una definizione di stress (si veda a questo proposito l'articolo di M. Maurer, 1984) e del fatto che il significato del termine ‛stress' è una questione di opinione personale, non esiterò a distinguere lo shock dallo stress. Per quel che riguarda il termine ‛stress', conformandoci alla tendenza che sembra attualmente prevalere inconsciamente, lo useremo per indicare i meccanismi e le conseguenze di un'aggressione in primo luogo psicosociale. Questo tipo di aggressione non s'accompagna a lesioni primarie, prodotte direttamente dall'agente aggressore ma, eventualmente, a lesioni secondarie, conseguenti alla reazione.
b) Il concetto di percorso biologico dell'idrogeno
Si è proposto di concepire (v. Laborit, Bases..., 1958) l'intero equilibrio biologico in funzione del percorso dell'idrogeno all'interno dell'organismo. Si può allora suddividere questo percorso in diverse tappe: a) una prima tappa in cui l'idrogeno, legato agli alimenti, è assorbito, immagazzinato (nel fegato, nel tessuto adiposo) e poi rimosso e trasportato dal sistema cardiovascolare fino alle cellule, in funzione dell'intensità dei loro bisogni energetici; b) una seconda tappa, intracellulare, in cui l'idrogeno viene prima staccato dai substrati, a opera delle deidrogenasi, e poi ionizzato (H+); ciò permette di ricavare dall'idrogeno l'energia di eccitazione dei suoi elettroni e di immagazzinarla nei composti fosforici ricchi di energia (ATP, GTP, fosfocreatina). È alla fine di questa tappa che interviene l'ossigeno, accettore di elettroni. L'ossigeno, attivato, si unisce al protone (H+) per formare, in maniera più o meno diretta, acqua (H2O). L'acqua, reagendo con il CO2 prodotto dalle decarbossilazioni, dà H2CO3, prodotto normale dei processi ossidativi intramitocondriali, che è riversato nell'ambiente extracellulare.
Se si scrive l'equazione di equilibrio:
CO2 + H2O ⇄ H2CO3 ⇄ HCO3- + H+,
si vede che la ventilazione del CO2 da parte dei polmoni sposta l'equilibrio verso sinistra e fa diminuire la concentrazione di ioni H+ ([H+]).
Essendo il pH definito come il logaritmo dell'inverso della concentrazione di ioni H+, log 1/[H+], si capisce come l'attività ventilatoria sia in grado di consentire un rapido controllo del pH. Ma acidi più dissociati, come l'acido lattico e gli acidi grassi, la cui produzione aumenta in ipossia e nel corso delle sindromi d'aggressione, possono spostare l'H2CO3 dai cationi ai quali è legato. Il CO2 sarà ventilato, ma la riserva di bicarbonato diminuirà (v. regolazione dell'equilibrio acido-base; v. respirazione, regolazione chimica della).
Così l'equilibrio biologico turbato negli stati di shock dovrà essere studiato dal punto di vista degli accidenti che possono capitare durante il percorso armonioso dell'idrogeno nell'organismo, considerando le varie tappe: quella tessutale, quella circolatoria, quella ventilatoria e, in particolare, quella escretoria, dato che i liquidi dell'ambiente interno non sono altro che un luogo di passaggio fra cellula e ambiente.
3. Gli stati di shock
Se prendiamo come modello lo shock emorragico, dato che la tecnica per provocarlo nell'animale è perfettamente standardizzata, potremo seguirne l'insorgere, sul piano dei sintomi clinici e fisiologici e su quello dei disordini biologici ed endocrino-metabolici.
a) Sintomatologia
In primo luogo dobbiamo fare una distinzione fra collasso e shock (v. Laborit, Réaction..., 1952). Il termine ‛collasso' implica un crollo della pressione arteriosa, che si incontra in certe fasi di sviluppo dello shock ma che può verificarsi anche in altre circostanze (lipotimia dei donatori di sangue, per esempio, e collasso legato ad asistolie acute, dipendenti da svariate cause). Durante lo shock la nuova ripartizione della massa sanguigna spesso manterrà, all'inizio, una pressione arteriosa quasi normale. L'assenza di correzioni vasomotorie durante il collasso è all'origine di un'ipossia cerebrale passeggera, con perdita di conoscenza, mentre il soggetto in stato di shock, generalmente, è cosciente. Il collasso è un sintomo, lo shock una sindrome che evolve nel tempo, nel cui contesto il collasso non è che un sintomo dai meccanismi polimorfi, che richiedono terapie diverse. Durante un intervento chirurgico, per esempio, se si verifica un collasso cardiovascolare, in conseguenza di un'emorragia massiccia, si farà ricorso a una trasfusione; ma se il collasso è dovuto a un'insufficienza cardiaca acuta, lo stesso trattamento provocherà, senza dubbio, un edema polmonare acuto.
Così, nella fase di scompenso degli stati di shock, il sintomo collasso, con crollo della pressione arteriosa, stasi venosa con rallentamento della circolazione di ritorno al cuore, tachicardia estrema, sarà senz'altro presente. Il malato sarà pallido, spesso cianotico, i suoi tegumenti saranno freddi a causa della vasocostrizione periferica, la sua respirazione rapida e superficiale. Benché prostrato, il malato resta, il più delle volte, lucido. La diuresi risulta raIlentata o assente.
b) Fenomeni generali da reazione
Il numero di lavori concernenti gli stati di shock, pubblicati negli ultimi cinquant'anni, è considerevole. Ma solo da trent'anni a questa parte si sono fatti dei veri progressi nella comprensione degli stati di shock e nella messa a punto di terapie adeguate, grazie ai concomitanti progressi registrati nel settore delle tecniche fisiologiche e, soprattutto, biochimiche.
La reazione ad aggressioni acute, in cui la sindrome lesionale e quella da reazione sono combinate e che conduce a uno stato di shock, ha essenzialmente come intermediari il sistema nervoso e quello endocrino. È a partire dall'intervento di questi due sistemi che le turbe cardiocircolatorie e metaboliche finiscono per coinvolgere successivamente i livelli intracellulare, cellulare, organico e sistemico. Ora, se può essere utile elencare e descrivere queste turbe, ancora più utile è comprendere come siano collegate le une alle altre e quali siano i meccanismi patologici in gioco, per sperare di controllarle all'origine.
1. I sistemi nervoso ed endocrino. - Già da tempo si supponeva che l'attivazione del sistema simpatico-adrenergico svolgesse un ruolo importante nell'insorgere dello shock. Lo stesso Cannon (1923-1934) avanzò l'ipotesi che l'iperattività del sistema simpatico-adrenergico potesse essere all'origine dello shock, ed Erlanger e Gasser (v., 1919) mostrarono che una perfusione prolungata di adrenalina (A) poteva provocarlo. Ma per dimostrare la portata e il meccanismo dell'intervento del sistema simpatico-adrenergico nell'insorgere dello shock, si sono dovuti attendere i metodi precisi di dosaggio delle catecolammine e dei loro metaboliti, metodi ideati da Weil-Malherbe e Bone (v., 1953), le cui tecniche strumentali sono state costantemente perfezionate fino a questi ultimi anni. D'altra parte, per quanto il ruolo delle catecolammine (CA) appala preponderante, quello di numerosi altri neuromodulatori periferici e centrali, scoperti in seguito, non può essere trascurato (v. neuroendocrinologia; v. neurosecrezione; v. sinapsi; v. sistema nervoso autonomo). Oltre alla noradrenalina (NA) (v. von Euler, 1948) e alla dopammina (DA), anche la Serotonina (5-HT), l'acido γ-amminobutirrico (GABA), gli oppioidi endogeni (endorfine) e numerosi altri polipeptidi intervengono, senza dubbio, nell'azione del sistema nervoso centrale durante gli stati di shock.
Lo stesso vale, evidentemente, per l'acetilcolina (ACh), ma le ricerche hanno mostrato che le variazioni di concentrazione dei neuromediatori alla periferia vanno considerate separatamente da quelle al livello dei centri. L'esistenza di una barriera ematoencefalica, impermeabile ai neuromodulatori plasmatici, fa sì che le variazioni della loro concentrazione al livello dei centri abbiano un significato autonomo. La barriera ematoencefalica manca soltanto al livello dell'ipotalamo e dell'eminenza mediana. Bisogna viceversa notare che gli ormoni peptidici possono essere trasportati nel cervello a partire dalla periferia.
Durante lo shock emorragico si registra nel cervello una significativa diminuzione di noradrenalina (NA), un aumento di dopammina (DA) e di acido dudrossifenilacetico (DOPAC), una diminuzione di acido omovanillico (HVA). Dato che quest'ultimo deriva dall'azione di un enzima, la catecolammina-O-metiltransferasi, al livello della sinapsi, mentre il DOPAC deriva dall'azione della monoamminossidasi (MAO) intramitocondriale, se ne può dedurre che la DA, durante lo shock, non viene più liberata al livello della sinapsi e si trova ossidata nel sinaptosoma. Analogamente le concentrazioni, nel cervello, della 5-HT e del suo metabolita, l'acido 5-idrossi-indolacetico (5-HIAA), aumentano. Ma il turnover della DA e della 5-HT, in seguito a ritrasfusione, due ore dopo lo shock, risulta fortemente aumentato: lo dimostra l'aumento, al di sopra del normale, delle concentrazioni di HVA e di 5-HIAA (v. Laborit e altri, Assay..., 1984). Si tratta di variazioni globali: sarebbe interessante studiare le variazioni che si determinano in certe aree specifiche durante lo shock emorragico e gli stati di shock in generale.
L'asse ipotalamo-ipofiso-surrenale. Le interrelazioni tra sistema nervoso e sistema endocrino obbligano, tuttavia, a non prendere in considerazione separatamente le loro reazioni. Al livello dei tessuti periferici, il sistema nervoso simpatico stimola la liberazione di NA in corrispondenza delle terminazioni delle sinapsi, e la liberazione di NA e di adrenalina (A) da parte della midollare del surrene; l'A proviene dalla metilazione della NA a opera della feniletanolammina-N-metiltransferasi (PNMT). D'altra parte, l'ipotalamo libera un polipeptide, il ‛fattore di liberazione delle corticotropine' (Corticotropin Releasing Factor, CRF); il CRF, che raggiunge l'ipofisi attraverso un sistema porta (vascolare), è il principale fattore stimolante la secrezione di ormone adrenocorticotropo (ACTH) da parte di tale ghiandola. L'ACTH, a sua volta, stimola la secrezione di glucocorticoidi (Gcs) da parte della corticale del surrene. Anche altri fattori endocrini e le stesse CA liberate nel plasma, agendo direttamente sul metabolismo della ghiandola, stimolano la secrezione di ACTH da parte dell'ipofisi. Inversamente, i glucocorticoidi surrenalici (Gcs) aumentano l'attività degli enzimi che presiedono alla sintesi delle CA nei surreni. L'attività della PNMT risulta fortemente accresciuta, il che favorisce la sintesi e la liberazione di A. L'attivazione degli stessi enzimi è anche sotto il controllo degli influssi colinergici nicotinici al livello della ghiandola. Infine i Gcs inibiscono la degradazione della PNMT.
Bisogna a questo punto notare che l'A possiede un effetto β-predominante, il che vuol dire che la sua azione sul metabolismo cellulare, stimolando la fosforilazione del glicogeno, alimenterà preferenzialmente le vie energetiche. La sua azione sul tono dei vasi è temporaneamente stimolante e poi rilassante: il suo effetto vasodilatatore periferico sarà dunque, in un secondo tempo, predominante (effetto β), in particolare sui vasi ricchi di β-recettori, quelli dei muscoli, del cuore, dei polmoni e del cervello, organi che necessitano di un'irrorazione sanguigna accresciuta, che permetta loro di far fronte a un comportamento di fuga o di lotta. Il suo effetto vasocostrittore si farà sentire soprattutto sui vasi della pelle e degli organi addominali (splancnici), che non sono indispensabili alla fuga e alla lotta.
La NA possiede un effetto α-predominante sui muscoli lisci dei vasi: la sua azione vasocostrittrice è generalizzata. Da ciò si può dedurre che durante gli stati di shock il rapporto A/NA nel plasma aumenterà. È la midollare del surrene che risponde in maniera predominante all'aggressione acuta (v. Fredholm e altri, 1979). L'attivazione della glicolisi da parte dell'A si accompagna a un notevole aumento del lattato serico, come vedremo; ciò spiega in parte l'iperlattacidemia da shock, dato che la NA in pratica influenza poco questo fattore. Si può altresì constatare che la stimolazione elettrica delle aree cerebrali che stimolano l'attività motoria (ippocampo ventrale, amigdala dorso-mediana) provoca un forte aumento della lattacidemia e della concentrazione di A nel plasma. La stimolazione delle aree che inibiscono l'attività motoria (ippocampo dorsale, amigdala laterale) non influenza il lattato serico né il tasso di A, ma quello di NA (v. Laborit e altri, 1977).
Bisogna qui schematizzare le azioni degli ormoni dell'asse ipofiso-surrenale. L'ACTH, per la maggior parte degli autori, aumenta il turnover delle CA cerebrali e incrementa la sintesi della DA e della NA a partire dalla (3H)tirosina. Le CA centrali hanno un effetto stimolante sul comportamento motorio, mentre gli oppioidi endogeni sembrano avere un effetto inverso. I Gcs non sembrano favorire il turnover cerebrale delle CA, nel ratto intatto, se non attraverso la liberazione primaria di ACTH, dato che la surrenalectomia, che aumenta la liberazione di ACTH a causa della scomparsa del feedback inibitore dei Gcs, aumenta ancora di più il turnover delle CA cerebrali.
Altri ormoni ipofisari. Accanto all'asse ipofiso-surrenale, altre secrezioni endocrine sono influenzate dalle aggressioni. Bisogna dire che la secrezione di ACTH da parte dell'ipofisi è accompagnata dalla secrezione di oppioidi endogeni che hanno lo stesso precursore peptidico, la prooppiomelanocortina (POMC). Molti autori pensano che, quando i recettori degli oppiacei, localizzati sulle terminazioni dopamminergiche, sono attivati, inibiscono la liberazione di DA e di NA centrali, mentre aumentano la DA, la NA e l'A plasmatiche (v. Dunn e Kramarcy, 1984). Sembra che anche la prolattina sia liberata durante le aggressioni, senza dubbio sotto l'azione della β-endorfina o dell'inibizione trasmessa dal sistema dopamminergico centrale inibito. L'ormone della crescita (OH) è, il più delle volte, inibito durante le aggressioni; lo stesso accade agli ormoni gonadotropi, LH e FSH, indubbiamente attraverso il medesimo meccanismo. Infine la concentrazione di vasopressina (l'ormone antidiuretico e ipertensivo) aumenta; questo fattore interviene, senza dubbio, nella diminuzione della diuresi in un individuo colpito da shock. Tuttavia alcuni autori pensano che la liberazione di vasopressina non sia dovuta all'aggressione in sé, ma all'immobilizzazione, all'ipossia, all'ischemia o a fattori emodinamici. Notiamo che la vasopressina agisce come il CRF e stimola la liberazione di ACTH.
Significato delle regolazioni. Ci si può chiedere quale sia il significato della risposta neuroendocrina all'aggressione. Per quel che riguarda le CA, l'azione sul metabolismo del glicogeno e l'azione vasomotoria che sposta la massa sanguigna verso gli organi indispensabili alla fuga e alla lotta sembrano una spiegazione sufficiente. Per quel che riguarda i glucocorticoidi, la spiegazione è meno evidente. Ricordiamo tuttavia che anche i Gcs stimolano la glicogenolisi, nonché la lipolisi; ma essi inibiscono la sintesi dei grassi, delle proteine e degli acidi nucleici, eccezion fatta per quelli che assicurano la gluconeogenesi a partire dalle proteine.
In effetti, i Gcs favoriscono la degradazione delle proteine dei tessuti in amminoacidi; questi ultimi, ripresi dal fegato, sono convertiti in glucosio (gluconeogenesi). Così i Gcs possono rifornire le riserve di glicogeno per l'alimentazione energetica del metabolismo cellulare: le CA favoriscono l'utilizzazione energetica del glicogeno.
Il sistema ipotalamo-ipofiso-surrenale è un sistema regolato in condizioni fisiologiche: la secrezione di un ormone inibisce la secrezione del o dei fattori che ne stimolano la sintesi e la liberazione. Così l'ACTH inibisce la liberazione di CRF e i Gcs inibiscono la liberazione di CRF e di ACTH. Si tratta di un sistema regolato mediante ‛retroazione negativa', che mantiene l'omeostasi del meccanismo di secrezione. Tuttavia bisogna notare che esiste anche una regolazione a ‛feedback positivo', dato che i Gcs favoriscono la sintesi e la liberazione delle CA della midollare del surrene e queste, a loro volta, favoriscono, agendo direttamente sull'ipofisi, la secrezione di ACTH. Il controllo di questi due sistemi retroattivi è possibile soltanto tramite un comando esterno al sistema, che agisce sull'ipotalamo e sulla liberazione di CRF. Questo comando è l'espressione del funzionamento del cervello, nel suo insieme, in rapporto all'ambiente. L'attività neuronica centrale catecolamminergica inibisce la liberazione di CRF, l'attività colinergica e serotoninergica centrale la stimola.
Tutto porta a credere che negli stati di shock questo comando esterno al sistema sia sopraffatto e che il feedback positivo abbia il sopravvento su quello negativo. Ciò permetterebbe di spiegare la caduta della concentrazione di NA cerebrale e la non utilizzazione della DA. Si può pensare, a quanto sembra, che le aree cerebrali che favoriscono la fuga e la lotta, ovvero l'aggressività difensiva (l'ippocampo dorsale, l'amigdala dorso-mediana, il sistema periventricolare, PVS), provochino l'intervento dell'asse ipotalamoipofiso-surrenale ma, nello stesso tempo, scatenino la reazione predominante, immediata, della midollare del surrene, reazione che permette una nuova ripartizione della massa sanguigna. Sarebbe questa reazione a retroazione positiva sull'asse che svincolerebbe il sistema dal suo comando nervoso situato più in alto. Vedremo che nei cosiddetti stati di stress entra in gioco un altro meccanismo (SIA), il cui comando centrale è diverso e che, senza dubbio, mantenendo il contatto con l'ambiente senza poterlo controllare, sembra essere all'origine delle turbe biologiche che conseguono all' ‛attesa in uno stato di tensione' del momento di agire. In questo caso è implicato il cervello anteriore.
I meccanismi di liberazione delle catecolammine. Già da parecchi anni si è giunti alla conclusione che gli influssi nervosi originati in una regione traumatizzata possono propagarsi fino ai centri cerebrali, i quali reagiscono stimolando le reazioni nervose ed endocrine che abbiamo schematizzato.
È noto che una rachianestesia che blocchi le vie nervose ascendenti ritarda o sopprime l'insorgere di uno shock traumatico dovuto, per esempio, a lesioni delle membra. Ma io ho potuto mostrare che, in stato di shock, i centri vasomotori non sono affatto ‛esauriti'. Questa constatazione induce ad ammettere l'origine periferica della sindrome. Per contro, la stimolazione del sistema adreno-simpatico periferico fa appello a quella dell'ipotalamo, tramite quella della formazione reticolare attivatrice ascendente. Nella preparazione ‛encefalo isolato' di gatto, la stimolazione elettrica del tronco cerebrale fa apparire delle CA nel circolo.
I chemo- e i barocettori esercitano anch'essi un'influenza primaria. L'ipossia stimola i chemocettori nello shock emorragico e la stimolazione elettrica delle loro fibre afferenti provoca iperpnea e ipertensione. La diminuzione della pressione di perfusione sanguigna nel glomo è un potente fattore stimolante dovuto, sembra, alla diminuzione della concentrazione di composti fosforati ad alta energia (ATP, fosfocreatina) nei chemocettori.
2. Le conseguenze periferiche della reazione all'aggressione. - Il numero di lavori dedicati alla fisiopatologia e, più recentemente, alla biologia degli stati di shock è talmente alto che non è possibile, in questa sede, citarne tutti gli autori e farne una rassegna esauriente. Consiglio, pertanto, la lettura del capitolo dedicato all'argomento in Les comportements (v. Laborit, 1973), che contiene un'abbondante bibliografia.
Turbe vasomotorie e loro conseguenze. È fuori di discussione il fatto che, in seguito a un'emorragia, la circolazione del mesentere, del sistema porta epatico e dell'intestino, della milza, del rene, del pancreas subisce notevoli turbe.
Le portate sovraepatiche e renali subiscono un calo. Dopo un'emorragia isolata, il ripristino progressivo della portata sanguigna si realizza in un'ora circa. Usando la tecnica di Wiggers o una delle sue varianti, che mantengono la pressione arteriosa sistemica intorno ai 40 mmHg (5,3 kPa), si impedisce questo ripristino. Questa caduta delle portate è dovuta alla diminuzione della pressione di perfusione prodotta dalla diminuzione della massa sanguigna, ma anche a una vasocostrizione attiva, di origine nervosa e umorale. L'osservazione al microscopio del mesentere permette di constatare questa intensa vasocostrizione. La congestione dei visceri si verifica sempre, quale che sia il tipo di shock. Esiste un accumulo di sangue nell'area venosa dell'addome e il ritorno al cuore è insufficiente. La pressione venosa centrale (PVC) subisce un calo.
Le responsabilità del sistema catecolamminergico è evidente. La perfusione di A o di NA in dosi parafisiologiche (2 ng/kg/min) provoca, in due ore, uno stato di shock irreversibile. Se si aumenta la dose di A a 8 ng/kg/min, i sintomi evolvono in due fasi: una fase ipertensiva con ipercaliemia, ipercalcemia, iponatriemia ed eccitabilità neuromuscolare aumentata. A partire dall'ottantesimo minuto, viceversa, la pressione arteriosa cala progressivamente, all'eccitabilità nervosa centrale iniziale subentra la comparsa di onde lente, l'eccitabilità neuromuscolare diminuisce. Il consumo di ossigeno si riduce fin dai primi minuti di perfusione. Si nota anche un arresto totale della circolazione capillare mesenterica e la comparsa di uno sludge (aggregazione intravascolare: vedi oltre). Il notevole aumento della differenza fra ossigeno arterioso e ossigeno venoso al livello degli organi splancnici riflette la stasi circolatoria di cui sono sede. Il quadro circolatorio è quello che si incontra durante gli stati di shock. Un ultimo gruppo di fatti permette ancora una volta di ‛incriminare' le CA: l'azione terapeutica favorevole, su tutti questi sintomi, degli agenti capaci di diminuire l'intensità dell'azione delle CA (simpaticolitici, ganglioplegici, neuroplegici). Ma questi agenti, per poter ripristinare una circolazione arterocapillare normale, necessitano, soprattutto in caso di shock emorragico o traumatico, che la loro azione sia progressivamente e parallelamente accompagnata da riempimento vascolare mediante trasfusione.
Il sistema arteriolo-capillare descritto da Chambers e Zweifach (v., 1944) è costituito da un'arteriola afferente e da una venula efferente, unite da un canale preferenziale: la metarteriola. Su quest'ultima si aprono i capillari, la cui origine è sotto il controllo di uno sfintere precapillare. Infine i capillari sono ‛shuntati' da anastomosi arterovenose, vasi corti e dotati di muscolatura, la cui apertura devia la circolazione del letto capillare. Le CA contraggono arteriole e venule ma, al contrario, aprono le anastomosi arterovenose e sottopongono a ipossia i territori tessutali su cui agiscono. Il controllo del tono delle anastomosi arterovenose sembrerebbe quindi avere una grande importanza fisiopatologica e terapeutica.
Anche il mantenimento dell'iperlattacidemia e lo sludge sono una conseguenza di una diminuzione della portata circolatoria e quindi della funzionalità degli organi (fegato e reni) che assicurano la gluconeogenesi a partire dal lattato (ciclo dei Cori). Ora, quale che sia il mezzo adottato per provocarlo (perfusione di A, shock emorragico, alcool etilico, lattato di sodio), lo sludge compare in corrispondenza di una concentrazione, sempre la stessa, di 800 mg/l di lattato serico: si tratta di un fenomeno metabolico che ritroveremo più avanti.
Anche il ruolo delle tossine microbiche intestinali, durante gli stati di shock, va inserito nel quadro delle modificazioni tessutali conseguenti alle turbe della circolazione mesenterica: Fine e collaboratori (v., 1960) lo hanno dimostrato sperimentalmente. La sterilizzazione dell'intestino tramite antibiotici produce un miglioramento nell'evoluzione degli stati di shock, pur non impedendo in tutti i casi la morte. Oltre che delle tossine microbiche, è necessario tener conto anche dell'ammoniaca, liberata dalla flora ammoniogenica intestinale: il suo tasso ematico s'innalza durante lo shock (v. Laborit e altri, 1958).
Anche le funzioni del sistema reticoloistiocitario (SRI) sembrerebbero turbate, come dimostra l'abbassamento dell'attività della colinesterasi serica (v. Laborit e Morand, 1947). Molti autori attribuiscono l'inibizione dell'SRI all'azione di tossine di origine intestinale. Vedremo che il ruolo dell'iperlattacidemia è senza dubbio predominante. In ogni modo gli animali il cui SRI viene inibito tramite un iniezione di Thorotrast (una sospensione di ossido di torio) muoiono di shock emorragico, reversibile negli animali di controllo.
Le lesioni intestinali conseguenti ai disordini vasomotori e all'anossia producono necrosi ed emorragie nella mucosa e si accompagnano a soffusioni emorragiche sul mesentere. Si manifestano con una diarrea putrida e sanguinolenta. La rimozione di una pinza posta sull'arteria mesenterica superiore provocava uno shock rapido e fatale: si è potuto prevenire l'insorgere dello shock tramite una perfusione continua di quest'arteria col sangue di un donatore.
Si deve prendere in considerazione il ruolo dei lisosomi nelle teorie che spiegano gli stati di shock postulando l'azione di tossine. Christian de Duve e collaboratori (v., 1955) hanno descritto, dopo centrifugazione differenziale, una frazione cellulare caratterizzata dal suo alto contenuto di enzimi proteolitici, e altre idrolasi, agenti a pH acido. Questi enzimi restano inattivi fintantoché la frazione con la quale si depositano, limitata da una membrana a un solo strato lipoproteico, non viene rotta. Essi proposero di chiamare queste inclusioni citoplasmatiche ‛lisosomi'. Nella fagocitosi, dopo l'invaginazione del corpo da fagocitare all'interno della membrana citoplasmatica e la costituzione di un vacuolo che lo ingloba, il fagosoma, il lisosoma entra in contatto col fagosoma e vi riversa il proprio contenuto enzimatico, costituendo il fagolisosoma, che digerisce il corpo fagocitato, i cui resti vengono espulsi insieme con gli enzimi proteolitici nello spazio extracellulare (v. cellula: Fisiologia della; v. infiammazione). La formazione del fagosoma dipenderebbe dalla glicolisi; la formazione del fagolisosoma richiederebbe la perossidazione delle membrane lisosomiali, resa possibile da una maggior produzione di H2O2 in dipendenza dalla via dello shunt dell'esosomonofosfato (via dei pentosi). I lisosomi, ‟sacchetti suicidi" (de Duve), svolgono dunque un ruolo di protezione, che può diventare pericoloso quando gli enzimi proteolitici che essi contengono vengono liberati e distruggono l'architettura protoplasmatica. È quel che può accadere in seguito a irradiazione o a intossicazione da tetracloruro di carbonio; l'azione dell'ossigeno iperbarico e gli agenti antiossidanti sono in grado di stabilizzare le membrane dei lisosomi.
Si è potuta mettere in evidenza la rottura dei lisosomi durante lo shock emorragico; in effetti nel plasma si ritrovano enzimi citoplasmatici, come le transaminasi, indici di lesioni cellulari, nonché attività enzimatiche intralisosomiali, come la fosfatasi acida e la 3-glucuronidasi (v. Laborit e altri, 1967). Ma antienzimi che si oppongono, in vitro, agli enzimi proteolitici lisosomiali non apportano alcun miglioramento all'evoluzione dello shock. Sembra quindi che la teoria tossica dello shock emorragico che attribuisce l'origine della tossicità ai batteri intestinali - che l'antibioticoterapia intestinale locale controllerebbe - non possa, da sola, spiegare le lesioni e l'evoluzione dello shock verso la morte.
Ma si è potuto mostrare che la sola iniezione intraperitoneale di A alla dose di 1 mg/kg aumenta, nel gatto, il tasso delle fosfatasi acide plasmatiche. L'aumento dell'attività ossidativa porta a un aumento della formazione endocellulare di H2O2; l'amminossidazione delle CA da parte della monoamminossidasi (MAO) produce lo stesso effetto. Sicché la reazione adrenergica all'aggressione potrebbe essere considerata responsabile della labilizzazione delle membrane lisosomiali. È possibile che ciò spieghi l'azione antinfiammatoria, riconosciuta, degli inibitori della MAO (IMAO).
Turbe metaboliche durante lo shock. Ovviamente sono dominate dall'acidosi metabolica. Le principali turbe concernono la lattacidemia e la piruvicemia. È ben noto l'interesse del rapporto lattato/piruvato e della nozione di ‛eccesso di lattato', calcolato in base alla formula di Huckabee (v., 1958):
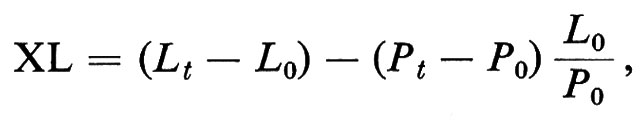
nella quale XL indica l'eccesso di lattato, Lt e Pt, rispettivamente, il tasso di lattato e quello di piruvato al tempo t, L0 e P0 i tassi di lattato e di piruvato al tempo zero. Nel nostro esperimento sullo shock emorragico (v. Laborit e altri, 1966) l'eccesso di lattato (XL) raggiunge il valore di 3,7 mM/l al momento dell'irreversibilità, un valore leggermente inferiore a quello trovato da Broder e Weil (v., 1964): 4 mM/l. Ma noi abbiamo constatato che l'evoluzione dello shock verso la morte si accompagna a una diminuzione progressiva dell'XL, il che rende dubbio il valore prognostico del rapporto, se il dosaggio è fatto dopo la comparsa del valore massimo.
Le variazioni della glicemia evolvono in due fasi: una prima fase di iperglicemia, seguita da ipoglicemia; l'ipercaliemia, invece, si accentua costantemente fino alla morte. Si è potuto mostrare che la fuoriuscita di K+ negli spazi extracellulari ha luogo soprattutto a partire dal fegato: Andersen e Shoemaker (v., 1967) imputano questo effetto all'adrenalina, ma si può anche attribuirlo all'ipossia epatosplancnica provocata dall'adrenalina.
La pCO2 presenta variazioni più complesse. Bisogna innanzitutto notare l'evoluzione divergente della pCO2 arteriosa rispetto a quella venosa: la pCO2 arteriosa si abbassa precocemente, a causa dell'iperventilazione; la pCO2 venosa, al contrario, comincia con l'innalzarsi fortemente a causa della liberazione, da parte dei tessuti, di acidi fissi, che spostano i bicarbonati. Se si preleva sangue venoso prima che passi attraverso i polmoni, si noterà una pCO2 elevata, fintantoché la riserva di bicarbonati potrà mantenerla tale; all'inizio la pCO2 evolve parallelamente alla lattacidemia.
Si può affermare che, dal momento in cui la pCO2 venosa comincia ad abbassarsi, lo shock raggiunge la fase di irreversibilità: a mio parere è questo il segno più affidabile dell'inizio della fase di irreversibilità ed è anche il sintomo meno sensibile alla terapia. Mentre il pH può essere controllato, una pCO2 venosa al di sotto di 20 mmHg (2,67 kPa) non si può, praticamente, normalizzare. La misura dell'‛eccesso di base' (BE) e del deficit anionico (AD) permette di precisare questi stati in terapia tramite la gasometria.
Il pH arterioso segue un andamento analogo a quello della pCO2 arteriosa, mentre il pH venoso segue una curva inversa rispetto alla pCO2 venosa e si abbassa fin dall'inizio dello shock: ciò mette in evidenza il calo dei bicarbonati. La reiniezione del sangue stravasato, associata a iniezione di sostanze tampone (Tris, bicarbonato di sodio), ristabilisce un pH normale per tutto il periodo dell'evoluzione dello shock, ma risulta incapace di impedire l'esito mortale. D'altra parte la correzione del pH non si oppone alla depressione cardiovascolare.
Il consumo di O2 (QO2) s'abbassa in un primo tempo in maniera drastica e poi più gradualmente. La ritrasfusione rialza questo consumo stabilmente, se lo shock non è ancora irreversibile, temporaneamente, se lo shock ha raggiunto la fase di irreversibilità.
L'attività colinesterasica serica diminuisce (v. Laborit e Morand, 1947) e le transaminasi seriche (SGOT e SGPT) aumentano, esprimendo così la sofferenza dei tessuti; lo stesso accade per le fosfatasi acide. Si nota un aumento rilevante degli acidi grassi liberi (AGL), che partecipano all'acidosi metabolica. Infine il catabolismo proteico si accompagna a un aumento dell'urea e dell'ammoniaca plasmatiche.
Riassumendo, la lattacidemia e la piruvicemia permettono di avere informazioni sull'attività della glicolisi e dei processi ossidativi, dato che la reazione fra acido piruvico e acido lattico è una ‛piattaforma girevole' posta fra processi aerobi e processi anaerobi. Il pH segnala la gravità dell'acidosi, mentre le pCO2 arteriosa e venosa mostrano come l'organismo possa compensarla. Glicemia e caliemia forniscono informazioni complementari. Abbiamo anche studiato le variazioni di 2,3-difosfoglicerato globulare (2,3-DPG) (v. Laborit e altri, 1972), intermediario della glicolisi, che permette la dissociazione, al livello dei tessuti, dell'ossiemoglobina e il rilascio di ossigeno ai tessuti. Il tasso di 2,3-DPG si alza all'inizio dello shock emorragico a causa di un aumento dell'attività glicolitica, ma in fase di irreversibilità si abbassa, per il blocco dell'attività glicolitica. In questo caso l'iperossigenazione del sangue non può essere completamente efficace: agli stessi risultati è giunto Esina (v., 1981).
Fisiopatogenesi metabolica degli stati di shock. L'A e le CA, in generale, insieme con l'anossia, sono i fattori essenziali dell'iperlattacidemia. Piccolissime dosi di A, di appena 0,9 ng/kg/min, innalzano la lattacidemia da 180 mg/l a 380 mg/l.
Lo stesso vale per quel che riguarda l'iperglicemia e l'ipercaliemia dell'acidosi e l'abbassamento della pCO2, ancorché l'iperventilazione iniziale, stimolata per via riflessa, ma mantenuta dall'acidosi, svolga un ruolo precoce nella caduta della pCO2 arteriosa.
Le CA sono anche responsabili del mantenimento dell'acidosi, dato che la diminuzione della portata epatorenale che esse provocano diminuisce le possibilità di riciclaggio dei lattati nella gluconeogenesi. Senza una perfusione di CA, il lattato iniettato sparisce rapidamente dal circolo. Un α-litico sotto perfusione di A riporta alla norma il Q02, mentre un β-litico ne rivela ancora la diminuzione. Il primo fattore di stabilità dell'iperlattacidemia, accanto all'effetto β-mimetico sulla formazione del 3′,5′-AMP ciclico e il conseguente aumento della glicolisi, è l'ipossia di numerosi territori tessutali in vasocostrizione. L'apertura degli shunts arterovenosi favorisce questa ipossia. Soltanto gli α-litici, sopprimendo la vasocostrizione, sono capaci di ridurre la differenza arterovenosa, grazie all'aumento della portata dei tessuti. Essi riducono anche la lattacidemia, assicurando una migliore perfusione epatorenale e favorendo così la gluconeogenesi. Infine l'abbassamento della pCO2 dovuto alla dissociazione dei bicarbonati spiega anche l'impossibilità di mantenere il funzionamento dei cicli ossidativi da parte della carbossilazione del piruvato.
Riassumendo, sembra dunque che tutte le turbe metaboliche registrate durante lo shock abbiano all'origine, direttamente o indirettamente, l'iperattività adreno-simpatica in risposta all ‛aggressione.
c) Integrazione dei processi metabolici e fisiopatologici.
Bisogna prestare attenzione a un fatto: a partire da una concentrazione di lattato nel plasma variabile da 800 mg a 1.000 mg/l, compare un certo numero di lesioni anatomiche e funzionali. Ora, queste lesioni appaiono su elementi cellulari o su tessuti, la cui attività ossidativa è ridotta, il cui equipaggiamento mitocondriale è relativamente limitato e in cui il ruolo della glicolisi è importante.
1. L'aggregazione intravascolare della microcircolazione e le coagulopatie di consumo. - Il fenomeno fu descritto sotto il nome di ‛rulli' da R. Fåhraeus nel 1921. Nel 1945 M. H. Knisely e collaboratori descrissero il fenomeno dell'aggregazione intravascolare o ‛sludge', che essi considerano sempre patologico. Per Fåhraeus il fenomeno faciliterebbe il trasporto sanguigno diminuendo la viscosità del sangue, ammettendo però che nei vasi capillari più piccoli il raggrumarsi (sludge) aumenti lo scorrimento.
Numerosi fattori sono capaci di provocare lo sludge: plasmatici (iperglobulinemia, introduzione di macromolecole nel sangue), emodinamici (diversi shock, ipotensione, vasocostrizione adrenalinica), eritrocitari (lesioni globulari, sangue conservato), parietali (vasocostrizione arteriosa, contatto con adrenalina, eccitazione simpatica). Ora, si può constatare sperimentalmente che la maggior parte di questi fattori provoca una iperlattacidemia. La perfusione di lattato di sodio provoca anch'essa lo sludge se la concentrazione raggiunge gli 800 mg/l. Ricordiamo che la glicolisi porta alla formazione del piruvato che, in aerobiosi, sarà utilizzato come acetil-CoA per iniziare il ciclo degli acidi tricarbossilici. Se l'attività di questo viene perturbata (ipossia, turba di origine tossica), l'acido piruvico si orienterà verso la formazione di acido lattico nella reazione promossa dalla latticodeidrogenasi (LDH) o suoi isoenzimi:
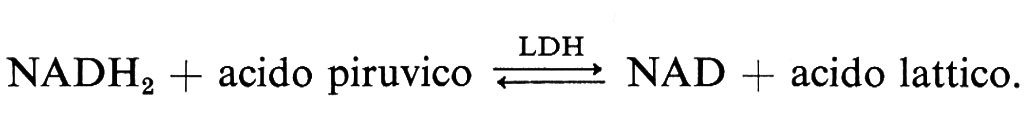
Si tratta di una reazione d'equilibrio che può quindi invertirsi in seguito all'aumento del prodotto della reazione: l'acido lattico. In questo caso NADH2 non può più essere ossidato e la glicolisi si blocca. Ora, sugli elementi debolmente ossidativi, la glicolisi è la fonte principale di ATP, che permette la conservazione della struttura e della funzione. Il blocco della glicolisi produce quindi depolarizzazione, invasione di sodio, edema.
La cellula endoteliale dei vasi sembra appartenere a questo tipo, così come il globulo rosso, la cui formazione di 2,3-DPG risulta bloccata con una concentrazione plasmatica di lattato di 800 mg/l (v. Laborit e altri, 1972). È in corrispondenza di questa concentrazione che compare lo sludge. L'acido piruvico non ne è capace e può anche opporsi momentaneamente alla sua comparsa. Schummer (v., 1966) ha ottenuto gli stessi risultati con il lattato, per una concentrazione nel plasma di 720 mg/l.
Abbiamo potuto mettere in evidenza lesioni endoteliali aortiche identiche, prodotte tanto da un'iniezione di A quanto da una perfusione di lattato di sodio, tale da ottenere un'iperlattacidemia al tasso indicato, e anche con un'iperventilazione sufficientemente intensa e prolungata da provocare un'acidosi metabolica di pari valore. Si tratta di un rigonfiamento edematoso della cellula, la quale si separa dalle cellule adiacenti; ciò produce la comparsa di pori intercellulari dove il plasma e le piastrine entrano in contatto con il collageno della membrana basale. L'adesione delle piastrine è la prima tappa della trombosi.
Abbiamo anche potuto mostrare che l'aggregazione piastrinica in vitro si ottiene per una stessa concentrazione plasmatica di acido lattico, e senza che la si possa imputare a un abbassamento del pH. L'acido piruvico alle stesse dosi non la provoca e anzi si oppone a quella prodotta dall'acido lattico, indubbiamente ossidando NADH2 e ripristinando la glicolisi.
Nella ‛degenerazione viscosa' delle piastrine, ultima tappa della trombosi, sembra probabile il ruolo dei lisosomi. L'iperlattacidemia, bloccando la glicolisi, favorirà la penetrazione del glucosio-6-fosfato nella via dei pentosi e la formazione di H2O2 che perossida le membrane dei lisosomi piastrinici. Lo sludge, immobilizzando i molteplici fattori della coagulazione, può condurre a sindromi emorragiche per lungo tempo confuse con la fibrinolisi. Ma le sindromi emorragiche non sono che l'espressione di una coagulopatia intravascolare disseminata (v. Hardaway e Drake, 1963). Ora, per garantire una microcircolazione corretta, che permetta un'alimentazione normale dei tessuti, bisogna ottenere la lisi dei microtrombi. Hardaway e Drake hanno anche proposto il trattamento degli stati di shock irreversibili con la fibrinolisina.
2. Inibizione del sistema reticolo-istiocitario. - Abbiamo studiato questo fenomeno usando la tecnica di Halpern e collaboratori (v., 1954), basata sulla cinetica della scomparsa dal sangue di particelle di carbone iniettate per via venosa, la cui curva di scomparsa rappresenta l'attività fagocitaria dell'SRI (v. sistema reticoloendoteliale). Per la solita concentrazione di 800 mg/l, l'iperlattacidemia è capace di diminuire considerevolmente l'indice fagocitario.
3. Le strutture ossidative. - L'iperlattacidemia da shock è il fattore fondamentale della caduta della pCO2 arteriosa e poi, raggiunto lo stadio di irreversibilità, di quella della pCO2 venosa. Ora, la carbossilazione dell'acido piruvico a ossalacetico è necessaria alla rigenerazione degli intermediari del ciclo degli acidi tricarbossilici e al suo funzionamento. Perché questa carbossilazione resti possibile, è necessano che la pCO2 sia mantenuta al di sopra di 1 5 mmHg (2 kPa). Perché i cicli ossidativi siano efficaci, sia su sezioni di tessuti nell'apparecchio di Warburg, sia in vivo, è quindi necessaria una certa pressione parziale di CO2 . Altrimenti la glicolisi sarà favorita e l'acido lattico formato in maggior quantità non sarà utilizzato nel ciclo degli acidi tricarbossilici, e neppure il piruvico. L'acidosi metabolica crescente non può, in questo caso, che abbassare ulteriormente la pCO2 Abbiamo potuto constatare che una grave ipocapnia da iperventilazione, capace di scatenare un'acidosi metabolica, abbassa il consumo di O2 , mentre un'ipercapnia moderata, incapace di stimolare una risposta catecolamminergica d'origine centrale, favorisce una caduta della lattacidemia. Si può così capire l'effetto favorevole del ‛carbogeno' (miscela di O2 al 95% e CO2 al 5%), messo in evidenza da L. J. Henderson (1908-1917), e si capisce anche come una sezione di tessuto nell'apparecchio di Warburg possa consumare O2 solo in un'atmosfera comprendente anche il 5% di CO2. Inoltre, dato che una delle fonti più attive di CO2 è l'attività del ciclo degli acidi tricarbossilici, la depressione del suo funzionamento instaura un circolo vizioso, che deve essere uno degli elementi caratteristici dell'irreversibilità degli stati di shock.
d) I fenomeni lesionali e la loro partecicazione agli stati di shock.
I fenomeni locali che interessano la zona colpita dall'aggressione assumono maggiore o minore importanza a seconda dei tipi di shock.
1. Conseguenze generali delle lesioni locali. - Si è dovuto attendere il lavoro di Moon (v., 1942) per capire che la riduzione del volume del sangue durante lo shock traumatico non era dovuta a un edema generalizzato, ma le perdite idriche locali erano sufficienti a spiegarne la genesi. Si sa che le cellule endoteliali dei vasi lasciano passare nei tessuti sottostanti molecole voluminose, grazie a un fenomeno di trasporto vescicolare endoplasmatico analogo alla pinocitosi: per invaginazione della membrana superficiale. Questo processo fa ricorso alla glicolisi e può essere bloccato dall'iperlattacidemia. Ne risultano un gonfiore edematoso della cellula endoteliale, che protrude nel lume vascolare, e la comparsa di falle intercellulari che favoriscono l'aggregazione globulare e piastrinica, che è all'origine dello sludge. Tale fenomeno è bensì di carattere generale, ma si farà sentire particolarmente nelle lesioni focalizzate e nell'area splancnica.
Questi fenomeni si accompagnano a turbe ‛elettrolitiche'. Nella zona traumatizzata si accumula Na+, mentre, in tutti i tipi di shock, aumenta il K+ plasmatico: da ciò conseguono ipernatriemia e ipercaliemia. Dato che il mantenimento del sodio extracellulare e del potassio intracellulare esige un metabolismo cellulare attivo, necessario alla conservazione del potenziale di membrana, se ne può dedurre che ogni tipo di sofferenza cellulare, tossica o anossica, sarà all'origine di queste turbe elettrolitiche.
2. Ruolo del tessuto connettivo. - Gli scambi fra i liquidi circolanti - sangue e linfa - e le cellule parenchimatose avvengono attraverso una formazione complessa: il tessuto connettivo. Il connettivo è costituito da cellule - i fibroblasti e i mastociti - e da una sostanza fondamentale nella quale si distinguono le fibre collagene e le fibre elastiche. La sostanza fondamentale è costituita essenzialmente da mucopolisaccaridi, da eteropolisaccaridi neutri e da glicoproteine, ma contiene anche elementi provenienti dal sangue: acqua, enzimi, sali minerali, albumina, globuline, vitamine, ormoni.
I mucopolisaccaridi derivano dall'attività metabolica del fibroblasto e il loro compito sembra essere soprattutto quello di mantenere un ambiente ‛fortemente carico negativamente' intorno alle cellule parenchimatose. Il mastocito contiene eparina, istamina e serotonina. Ogni aggressione, e in particolare ogni sostanza carica positivamente, provoca una ‛esplosione' del mastocito, che riversa i suoi granuli nella sostanza fondamentale, dove parteciperanno ai fenomeni vasomotori locali.
Infine, numerosi polipeptidi (callidina, bradichinina) sono liberati nel tessuto connettivo a partire da α2-globuline plasmatiche, loro precursori, sotto l'azione di un enzima fibrinolitico, la callicreina. Queste chinine sono ipotensive e algogene. Le catecolammine sarebbero capaci, in vivo, di attivare le proteasi, nonché di provocare la degranulazione dei mastociti. Anche i leucociti polinucleati invadono la regione traumatizzata, si alterano e liberano il contenuto dei loro lisosomi, che aumenta la permeabilità capillare e provoca la rottura dei mastociti. Il ruolo delle prostaglandine ha attirato più recentemente l'attenzione.
L'insieme di queste turbe può essere riassunto dicendo che, localmente, il tessuto connettivo si trova invaso da sostanze cariche positivamente (diverse ammine, proteine del sangue ecc.), che devono essere bloccate dall'eparina e dai mucopolisaccaridi prima di essere catabolizzate dal fibroblasto. Ogni fattore ‛pro-ossidante' favorisce la fagocitosi, la labilizzazione delle membrane cellulari ed endocellulari (granuli, lisosomi) e quindi i fenomeni detti ‛infiammatori'.
La variabilità della partecipazione dei fenomeni locali alla sindrome generale dipenderà dal tipo di shock. La perdita locale di liquidi e di elettroliti assume una grande importanza attraverso la diminuzione della massa sanguigna circolante nello shock da laccio emostatico, da ustioni estese e da schiacciamento (crush injury). Questi fattori possono anche intervenire sui fenomeni generali tramite la messa in circolo di diversi prodotti d'attrito tessutale nello shock traumatico.
3. Ruolo del dolore. - Un problema più complesso e che meriterebbe una lunga trattazione è quello del ruolo del dolore nell'insorgere degli stati di shock. Ci limiteremo all'essenziale (v. somatoestesia; v. neoplasie: Dolore e terapia del dolore, suppl.). Alcuni considerano il dolore come un'aggressione di per sé, mentre esso non sembra altro che un epifenomeno cosciente di un'aggressione locale propagata per via neuroendocrina ai centri encefalici. La partecipazione affettiva, fondamentale nel fenomeno del dolore, rende la sua responsabilità difficile da apprezzare. Sembra che si debba distinguere la consapevolezza del dolore dalle reazioni generali a questa sensazione. Il dolore prende generalmente origine a partire dai fenomeni lesionali locali che abbiamo appena elencato; istamina, serotonina, chinine, prostaglandine locali partecipano alla sua genesi. Sappiamo che l'anestesia locale o quella spinale, pur non proteggendo dallo shock, sopprimono il dolore. Ma questo effetto è la conseguenza della diffusione nervosa ai centri encefalici degli stimoli aggressivi o dolorosi? I potenti morfino-mimetici che oggi possediamo sopprimono il dolore, permettendo l'intervento chirurgico, ma non sopprimono la reazione organica alle aggressioni e le turbe vasomotorie e metaboliche che le accompagnano, come fanno i neurolettici. La reazione neuroendocrina all'aggressione deriva dalla diffusione al di fuori della via lemniscale di stimoli provenienti dalla periferia. Questa diffusione ai centri encefalici si presenterà con la complessità di un fenomeno al quale parteciperanno la coscienza, l'attenzione, la memoria e quindi l'affettività, a seconda del passato personale e dunque della personalità dell'individuo. Ma durante l'ultimo decennio il notevole lavoro intrapreso, concernente gli oppioidi endogeni che indubbiamente partecipano a certi tipi di analgesie fisiologiche conseguenti ad aggressioni, è giunto a complicare ulteriormente la questione. In effetti, certi autori hanno trattato con successo le ipotensioni da shock emorragico o endotossinico somministrando nalossone, antagonista degli oppiacei (v. Holaday e Faden, 1978).
Riassumendo, se accettiamo la distinzione fatta all'inizio tra shock e stress, non è possibile affermare che il dolore, fenomeno cosciente, aggiunge ai meccanismi dello shock quelli dello stress? In questo caso è la consapevolezza del dolore che sarebbe utile sopprimere, senza sperare di agire, con ciò, sui meccanismi dello shock.
4. Gli stati di stress
La confusione che spesso viene fatta fra shock, stress, adattamento, aggressioni traumatiche o psicosociali, ci ha indotto, agli inizi di questo articolo, a un tentativo di chiarire i vari concetti. Un fatto è certo, ed è che i numerosi lavori recenti di provenienza anglosassone riguardano non lo shock, ma lo stress; infatti è il termine ‛stress' che compare generalmente nei titoli. Ora, i metodi sperimentali usati per provocarlo non portano mai a uno stato di shock. Che si tratti di elettroshock plantari inevitabili (ECPi), di contenzione di durata variabile, di nuoto forzato, di isolamento, di combattimento, ecc., l'esperienza si realizza su un animale vigile e non sotto anestesia generale. Questi lavori esplorano sempre l'asse ipotalamo-ipofiso-surrenale (IPA) e il sistema simpatico-adrenergico (SA), come quelli che riguardano gli stati di shock. Invece, le turbe dell'equilibrio omeostatico periferico, dell'equilibrio acido-base, le turbe metaboliche, i fenomeni vasomotori e arteriolo-capillari, per esempio, non sono mai presi in considerazione. Ma sono a questi ultimi che gli studi sullo shock si sono principalmente interessati; le attività biochimiche e neurofisiologiche centrali, viceversa, sono state, in questo caso, poco studiate. È probabile che lo sviluppo delle nostre conoscenze riguardanti la biochimica cerebrale, registrato in questi ultimi decenni, abbia permesso di vedere in questa biochimica e nelle sue variazioni il fattore essenziale delle variazioni dei nostri comportamenti. Si è cercato quindi di utilizzare test sperimentali implicanti un ‛comportamento' attivo o passivo nei confronti dell'agente aggressore, comportamento che non esiste negli stati di shock, anche se alla loro origine c'è stato un tentativo di fuga o di lotta. Così il cervello permetterà certe risposte nei confronti dell'agente ‛stressante': in primo luogo cercherà di controllarlo, di neutralizzarlo, o attraverso la fuga (evitamento attivo) o attraverso la lotta (combattimento, aggressività difensiva). Il cervello registrerà poi l'esperienza, cioè il successo o l'insuccesso di questo controllo, e questo apprendimento avrà delle conseguenze importanti per il suo comportamento successivo, che sarà dunque legato a un processo di memoria.
Ora, è ovvio che se il controllo dell'evento da parte dell'animale risulta efficace, ne conseguono poche turbe biologiche o fisiologiche, centrali o periferiche (v. Weiss, 1968). Soltanto quando questo controllo diventa impossibile appaiono le turbe.
a) L'inibizione dell'azione
Come abbiamo più volte ripetuto, il sistema nervoso serve esclusivamente a che l'organismo possa controllare le caratteristiche del suo ambiente. È tramite questo controllo che l'organismo manterrà la ‟costanza delle condizioni di vita nel suo ambiente interno" (v. Bernard, 1878), la sua omeostasi (v. Cannon, 1932), il suo piacere (v. Freud, 1953).
Mi limiterò a ricordare le conclusioni tratte (v. Laborit, 1973 e L'inhibition..., 1979) a proposito dei principali comportamenti: a) il primo comportamento rinnova l'azione gratificante; esso ha bisogno di un'esperienza precedente del risultato di questa azione, di un processo di memoria, e fa appello alle formazioni catecolamminergiche (dopammina e noradrenalina) cerebrali, mettendo in gioco, in particolare, il Medial Forbrain Bundle (MFB) (v. Olds e Milner, 1954); b) il secondo risponde in maniera innata agli stimoli nocicettivi, e fa appello al sistema periventricolare (PVS) - la cui mediazione è colinergica - che comanda la fuga e, se la fuga risulta impossibile, la lotta, l'aggressività difensiva. Se questi comportamenti sono efficaci e permettono di evitare la punizione, saranno rinforzati, come i precedenti (v. de Molina e Hunsperger, 1962); c) infine, se nessuno dei precedenti comportamenti è efficace, si giunge all'inibizione dell'azione, preferibile alla lotta sfiancante o alla distruzione da parte dell'aggressore. Questo comportamento fa appello a quello che abbiamo chiamato sistema inibitore dell'azione (SIA), di cui fanno parte certe aree corticali, l'area settale mediana, il nucleo caudato, l'ippocampo dorsale, e in particolare l'amigdala laterale, che convergono verso il nucleo ventro-mediano dell'ipotalamo, origine della via inibitrice finale comune. Questo sistema è colinergico e serotoninergico (v. Laborit, 1974).
Questo sistema inibitore dell'azione comanda la liberazione del CRF (fattore di liberazione della corticotropina), che fa intervenire l'asse ipofiso-corticosurrenale. Esso comanda anche la liberazione di noradrenalina da parte delle terminazioni simpatiche. Addurrò argomenti che autorizzano a pensare che i glucocorticoidi, la cui liberazione è stimolata dal funzionamento di questo sistema, stimolano a loro volta questo stesso funzionamento, facendo così entrare il soggetto in un circolo vizioso da cui non può uscire se non attraverso l'azione gratificante (v. Laborit, 1976).
L'inibizione promuove quell'effetto che si chiama ‛angoscia'. Le situazioni comportamentali che provocano l'inibizione possono riassumersi in alcuni grandi meccanismi che si possono considerare all'origine dello stress: a) conflitto interneuronico tra pulsione connessa a un bisogno fondamentale (bere, mangiare o copulare) e un fattore acquisito, derivato dall'apprendimento socioculturale, in altri termini fra la pulsione e il divieto socioculturale. Il fattore acquisito può essere anche una ricompensa rinforzata e poi, secondariamente, soppressa; b) impossibilità di agire per mancanza di informazioni circa le conseguenze dell'azione, in presenza di un evento non ancora inventariato dall'esperienza precedente, o, al contrario, per eccesso di informazioni, senza possibilità di classificarle in un registro comportamentale appropriato, nè di controllarle efficacemente (coping degli anglosassoni). L'angoscia di morte appartiene a queste due categorie; c) infine, l'angoscia propriamente umana, derivante dalla capacità dell'uomo di immaginare, a partire dalla sua esperienza passata, un evento doloroso, anche se è possibile che non si verifichi.
L'angoscia può risolversi nell'azione, anche inefficace: è il caso dell'aggressività esplosiva, nella tossicomania e in certi comportamenti nevrotici. Ma soprattutto, dato che i glucocorticoidi distruggono il timo e bloccano il sistema immunitario, favorisce gli accidenti patologici nel loro insieme, infettivi, tumorali o quelli che si è convenuto di chiamare ‛psicosomatici'. Si è potuto dimostrare che i glucocorticoidi, opponendosi alla sintesi proteica, sopprimono il sonno paradosso. Essi favoriscono la ritenzione d'acqua e dei sali, aumentando la massa sanguigna, mentre la noradrenalina eleva il tono vascolare: da ciò derivano le cosiddette ipertensioni essenziali e le loro complicazioni. Ma l'ansioso spera ancora in una soluzione della sua angoscia, mentre il depresso non si aspetta più niente. I due comportamenti spesso si alternano nel tempo o si combinano fra loro.
1. Sistema inibitore dell'azione e acetilcolina. - Altrove ho passato in rassegna i numerosi lavori sul ruolo del sistema colinergico in questa inibizione (v. Laborit, L'inhibition..., 1979); in questa sede citerò solo i più recenti. Anisman e altri (v., 1979) mostrano che il trattamento con uno stimolante catecolamminergico, la L-dopa, o un bloccante colinergico, la scopolamina, contrasta il deficit di reazione (inibizione) prodotto dall'apprendimento precedente di uno shock plantare inevitabile.
Russel e Macri (v., 1979) mostrano le strette relazioni esistenti tra il tasso cerebrale di ACh e l'attività motoria. L'emicolinium-3′ per via intracerebroventricolare (i.c.v.), che riduce la concentrazione di ACh, provoca un'iperattività. In seguito a lesioni settali che riducono il tasso di ACh, si nota un'iperattività iniziale che scompare col tempo: ciò suggerisce il ristabilimento di un equilibrio fra i neuromediatori.
Mason e Fibiger (v., 1979) constatano che i farmaci anticolinergici (scopolamina, atropina) provocano una stimolazione locomotoria nel ratto, e Stephens e Herberg (v., 1979) mostrano che scopolamina e benzatropina (i.p.) aumentano l'autostimolazione. Dato che la scopolamina non inibisce la ricaptazione della dopammina (DA) alle sinapsi, come la benzatropina, l'attività stimolante di questi farmaci anticolinergici non può essere attribuita che all'inibizione di un sistema inibitore colinergico della ricompensa.
Schmidt e altri (v., 1980) fanno la curiosa constatazione che i ratti del ceppo F-344 sono particolarmente portati a realizzare un evitamento attivo di uno shock elettrico plantare (ESP), mentre quelli del ceppo Z-M si inibiscono e riducono la loro attività motoria. Ora, un funzionamento colinergico accresciuto si osserva nell'ippocampo dorsale degli Z-M e non in quello degli F-344, dopo ESP. Gli autori ne deducono che un meccanismo colinergico controlla la risposta comportamentale repressiva allo stress. Analogamente Zolman e altri (v., 1978) constatano che la scopolamina aumenta, nei pulcini, l'attività motoria e la resistenza all'estinzione di un condizionamento attivo, teso a ottenere il cibo. Gli animali, in questo caso, difficilmente apprendono un evitamento passivo: questi risultati sono strettamente analoghi a quelli ottenuti con i ratti e indicano il ruolo di un sistema colinergico nell'inibizione. È questa anche la conclusione del lavoro di Kelsey (v., 1983).
Risch (v., 1982) mostra nell'uomo una supersensibilità del sistema colinergico centrale nella depressione, che Caroll (v., 1982) e Doerr e Berger (v., 1983) considerano responsabile della reazione del sistema IPA al test al desametasone, corticoide di sintesi che, nell'individuo normale, inibisce la secrezione di ACTH e di cortisone.
2. Sistema inibitore dell'azione e serotonina (5-HT). - Schenberg e Graeff (v., 1978) riferiscono che in ratti addestrati ad arrestare una stimolazione nocicettiva provocata da elettrodi collocati nella sostanza grigia mesencefalica, due antagonisti della 5-HT (metisergide e ciproeptadina) facilitano la risposta e diminuiscono la latenza della reazione. Tachiki e altri (v., 1978) mostrano che deboli dosi di 5-idrossitriptofano (5-HTP), precursore della serotonina, diminuiscono l'attività locomotoria e che lo stesso effetto mostrano dosi inattive di tetrabenazina, combinate con dosi inattive di 5-HT. La paraclorofenilalanina (pCPA), inibitrice della sintesi di serotonina, si oppone all'azione sedativa della tetrabenazina, che sarebbe quindi dovuta a un eccesso di 5-HT funzionale. Graeff e Silveira Filho (v., 1978) mostrano che la stimolazione dei nuclei del rafe serotoninergico nei ratti addestrati ad azionare una leva per ottenere acqua inibisce il loro comportamento.
La pCPA ripristina questo comportamento, mentre 5-HTP + benzerazide si oppongono all'azione della pCPA. Così la via serotoninergica mesencefalica, che prende origine nei nuclei del rafe mediano e si proietta sull'area settale e sull'ippocampo, sembra implicata nell'inibizione del comportamento. Ad analoghe conclusioni giunge Redgrave (v., 1978), che in un esperimento ha prodotto nell'animale un comportamento di autostimolazione consistente nell'azionamento di una leva collegata con un elettrodo inserito nell'MFB e nel tegmento mesencefalico ventrale (VMT): Redgrave constata che la 5-HT fa diminuire il numero degli azionamenti della leva e fa aumentare la loro durata, mentre la DA e la noradrenalina (NA) producono l'effetto opposto. Quindi il sistema della ricompensa parrebbe stimolato dalle catecolammine (CA) e inibito da un sistema serotoninergico. Plaznik e altri (v., 1980) provocano delle lesioni dei nuclei del rafe dorsale e mediano e constatano che, con la diminuzione massiccia della concentrazione di 5-HT, facilitano l'evitamento attivo e l'attività globale degli animali. Inversamente, Thornton e Goudie (v., 1978), usando pCPA e producendo lesioni dei nuclei del rafe mediano, impediscono l'acquisizione di una risposta di evitamento passivo. Anche Yamamoto e Ueki (v., 1978) riscontrano un'iperattività motoria negli animali con lesioni del rafe mediano; ma, dato che questa iperattività è soppressa dall'alfametiltirosina (α-MT), che blocca la sintesi delle CA, essi pensano che essa dipenda dall'attivazione di un sistema catecolamminergico secondario all'inibizione della funzione serotoninergica. Le lesioni del rafe sembrano, invece, responsabili del comportamento muricida; Waldbillig (v., 1979) sostiene la stessa opinione. Il sistema serotoninergico-inibitore, che prende origine nel rafe dorsale, si proietta sull'amigdala, le cui lesioni provocano anch'esse il comportamento muricida.
Montanero e altri (v., 1979) pensano che l'effetto antiamnesico di due sostanze antiserotoninergiche nei confronti di un evitamento passivo sia in relazione con un effetto antipunizione di un trattamento antiserotoninergico. Per finire, Brown e altri (v., 1982) mostrano l'esistenza di un meccanismo serotoninergico come mediatore di un comportamento ‛senza speranza' (helplessness).
3. Comportamento d'inibizione, memoria e nuclei ventromediani dell'ipotalamo (VMI). - Il ‛rinforzo', la ripetizione dell'azione gratificante, fa ricorso a un processo di memorizzazione che pertanto va collegato all'attività dell'MFB e delle vie catecolamminergiche.
Viceversa, la risposta a uno stimolo nocicettivo, la ‛fuga' e la ‛lotta', che, secondo l'opinione corrente, fa intervenire il PVS, è una risposta immediata che sarà rinforzata solo nel caso che porti all'evitamento della punizione.
Infine, l'‛inibizione dell'azione' implica anch'essa un processo di memoria, il ‛ricordo' dell'inefficacia dell'azione, come abbiamo potuto mostrare (v. Kunz e altri, 1974) sottoponendo gli animali, cui applicavamo elettroshock plantari inevitabili, a elettroshock (ES) convulsivanti alla fine di ogni seduta di apprendimento. L'ES convulsivante impedisce il passaggio dalla memoria a breve termine a quella a lungo termine (v., Dawson e McGaugh, 1969). Da un giorno all'altro l'animale non si ricorda dell'inefficacia della sua azione e così evita le conseguenze fisiopatologiche dell'inibizione dell'azione. Le vie inibitrici colinergiche messe in gioco fanno capo al nucleo ventromediano dell'ipotalamo, che si trova così all'origine della via finale comune verso la quale confluiscono quelle provenienti, in particolare, dall'area settale dell'ippocampo dorsale e dell'amigdala laterale. Bruce e altri (v., 1978) mostrano che le lesioni del nucleo ventromediano dell'ipotalamo (VMI) sopprimono l'evitamento passivo: essi attribuiscono questo fatto all'incapacità di inibire un comportamento precedentemente rinforzato e a un mutamento nelle tendenze a rispondere a stimoli avversi. Beagley e Beagley (v., 1978), dal canto loro, osservano che ratti che abbiano subito lesioni settali presentano un evitamento attivo pur essendo stati precedentemente addestrati all'inibizione tramite shock inevitabili: così le lesioni settali, secondo i due autori, ehminano la risposta inibitrice provocata dall'apprendimento dell'inefficacia dell'azione. King e Grossman (v., 1978) mostrano che le lesioni del VMI facilitano l'acquisizione di un comportamento di evitamento attivo, e Hoffman e Trowill (v., 1979) mostrano che le sinapsi colinergiche del VMI costituiscono una parte del sistema periventricolare, che controlla la risposta inibitrice del comportamento in seguito a punizione.
b) Inibizione dell'azione e asse ipotalamo-ipofiso-surrenale (IPA)
Numerosi lavori hanno mostrato: a) il ruolo del VMI nell'intervento dell'asse ipotalamo-ipofiso-surrenale (IPA); b) il ruolo scatenante dell'ACh e della 5-HT in questo meccanismo, nonché il ruolo inibitore delle catecolammine. Qualche anno fa ho pubblicato una rassegna generale sull'argomento (v. Laborit, 1976).
1. Ruolo del VMI nell'intervento dell'asse IPA. - Makara e altri (v., 1980) hanno constatato che, in seguito a sezione completa o antero-laterale intorno al VMI, le aggressioni non innalzano più la cortisolemia, mentre Filaretov e Vassilevskaya (v., 1978) notano che l'immobilizzazione si accompagna a una stimolazione dei neuroni del VMI, all'inibizione di quelli dell'ipotalamo anteriore e laterale e all'innalzamento della cortisolemia. D'altra parte Briaud e altri (v., 1979) mostrano che il fattore di liberazione delle corticotropine (CRF) si trova localizzato nella frazione mitocondriale dei sinaptosomi dei neuroni del VMI. Analogamente Nishizawa e Bray (v., 1978) constatano che, mentre l'aumento degli acidi grassi liberi (AGL) e del glicerolo, in risposta all'iniezione di NA, a causa della sua azione periferica, è lo stesso sia negli animali di controllo sia in quelli che abbiano subito una lesione del VMI, le diverse aggressioni (nuoto forzato, freddo, ecc.) si accompagnano invece a un aumento molto meno evidente e degli AGL e del glicerolo negli animali con lesioni del VMI: ciò dimostra che questa regione modula l'attività del sistema nervoso simpatico.
Infine Shimazu e altri (v., 1978) inducono, per stimolazione elettrica del VMI, un aumento rapido e forte del contenuto epatico in fosforilasi attiva del glicogeno, senza influenzare la sintetasi. L'opposto accade se si stimolano i nuclei laterali dell ‛ipotalamo (IL). La concentrazione di insulina diminuisce nel primo caso, non varia nel secondo. D'altra parte Van Bogaert e de Schepper (v., 1978) provocano una vasocostrizione pre- e post-capillare negli arti posteriori del cane stimolandone i nuclei del VMI e sopprimono queste risposte con un α-litico, il che mostra la loro dipendenza noradrenergica. Il VMI è dunque all'origine della via simpatica splancnica che controlla la glicolisi. La via vagale, antagonista, che controlla la glicogenesi, dipende dall'IL.
2. Ruolo dell'A Ch e della 5-HT nell'intervento dell'asse IPA e ruolo delle catecolammine (CA) nella sua inibizione. Il VMI sembra quindi essere il luogo d'incontro di differenti meccanismi soggiacenti, che intervengono nell'inibizione dell'azione, e il luogo d'origine di una via finale comune, che garantisce questa inibizione. D'altra parte questa inibizione, come abbiamo visto, è assicurata da meccanismi centrali colinergici e serotoninergici. Se ne può concludere che i due mediatori centrali, l'ACh e la 5-HT, stimolano l'intervento dell'asse IPA, in quanto quest'ultimo dipende dalla stimolazione del VMI. Per quel che riguarda questo argomento, cioè il comando nervoso centrale e il ruolo dei neuromediatori nell'intervento dell'asse IPA, si rinvia il lettore all'abbondante bibliografia già menzionata (v. Laborit, 1976 e L'inhibition..., 1979), nonché alle rassegne di Dunn e Kramarcy (v., 1984), di Sourkes (v., 1983) e di Axelrod e Reisine (v., 1984).
Buckingham e Hodges (v., 1979) inducono la liberazione di CRF applicando dell'ACh, della nicotina o del betanecolo sull'ipotalamo del ratto; Risch e altri (v., 1980) constatano che, nell'uomo stesso, i mutamenti d'umore e del comportamento provocati dalla fisostigmina (inibitore della colinesterasi) sono accompagnati da un aumento della concentrazione serica di cortisolo e di β-endorfina. Lo stesso accade con la 5-HT, mentre, come era già stato segnalato da diversi autori, questa liberazione di CRF è bloccata dalla NA e dal GABA, ma non dall'isoprenalina. Analogamente Sithichoke e altri (v., 1978) constatano che la risposta dell'asse IPA alle aggressioni viene ridotta da un'alimentazione carente di colina, effetto parzialmente corretto da un'aggiunta di coline all'acqua da bere. Sithichoke e Marotta (v. Sithichoke e altri, 1978) precisano che i recettori nicotinici centrali eccitano l'IPA nell'aggressione, ma restano inattivi in uno stato di non aggressione. Davis e Davis (v., 1980), per quel che riguarda l'uomo, prospettano anche la possibilità dell'intervento di un meccanismo colinergico centrale nella risposta dell'asse IPA allo stress.
Da parte loro Szafarczyk e altri (v., 1979) constatano che l'ACTH e il corticosterone plasmatico, nel ratto, diminuiscono in seguito a lesioni soprachiasmatiche e iniezione di pCPA, inibitore della sintesi di 5-HT, mentre l'iniezione di 5-HTP dopo somministrazione di pCPA stimola secrezione di ACTH. L'attività locomotoria diurna aumenta in seguito a somministrazione di pCPA. Nello stesso senso Modlinger e altri (v., 1980) mostrano che la somministrazione di triptofano stimola la liberazione di ACTH e di cortisolo. Le stesse constatazioni sono state fatte da Meyer e altri (v., 1978) e da Fuller e Snody (v., 1980).
Spindel e Negro-Vilar (v., 1983) mostrano che la 5-HT agisce direttamente anche sulle cellule in coltura di ipofisi anteriore, provocando la liberazione di ACTH e potenziando quella provocata dall'arginina-vasopressina (AVP).
Quanto al ruolo dell'inibizione della liberazione di CRF da parte dei neuroni noradrenergici, si rinvia il lettore ai numerosi lavori sull'argomento, fra cui quello di Weiner e Ganong (v., 1978). Quest'azione, a quanto pare, mette in gioco un α-recettore. Anche le lesioni del sistema noradrenergico, afferente all'ipotalamo, potenziano la liberazione di β-endorfina (v. Millan e altri, 1982). Sembra che anche i neuroni noradrenergici centrali controllino la liberazione di NA periferica. La somministrazione per via intraventricolare cerebrale di un α-bloccante, la dibenamina, provoca un'ipertensione arteriosa considerevole (v. Laborit e altri, 1974), che l'animale sia surrenalectomizzato o meno. Lo stesso effetto provoca un'iniezione di cortisone nell'animale surrenalectomizzato (v. Laborit e altri, 1975).
c) Ormoni e comportamento
Un certo numero di lavori ci mostrano, nel ratto, l'azione scatenante del corticosterone nel comportamento di sottomissione, vale a dire nell'inibizione. Leshner e Potitch (v., 1979) constatano che, aumentando il tasso di corticosterone plasmatico, si incrementa il comportamento di sottomissione. Essi continuano il loro esperimento mostrando che deboli dosi di corticosterone favoriscono la sottomissione secondaria durante un secondo test (v. Leshner e altri, 1980). La combinazione di sconfitta e corticosterone facilita notevolmente il comportamento secondario di sottomissione. Ely e Henry (v., 1978) osservano che i maschi dominanti sono più attivi e rispondono alle interazioni sociali utilizzando soprattutto il sistema adrenomidollare, mentre i maschi subordinati sono meno attivi e rispondono tramite l'asse ipotalamo-ipofiso-surrenale. Questo comportamento neuroendocrino è invertito quando è invertita la funzione sociale dell'animale. I maschi subordinati presentano una risposta del corticosterone plasmatico all'ACTH e allo stress da immobilizzazione più forte che non i maschi dominanti. Valeri e altri (v., 1978) mettono in evidenza, nel topo, le relazioni esistenti tra la captazione di (3H)idrocortisone nell'ippocampo e nel setto e il rango gerarchico dell'animale, nonché tra i recettori specifici per il corticosterone e l'asse ipofiso-surrenale. Dantzer e altri (v., 1980) constatano che la frustrazione dovuta alla soppressione di una ricompensa ottenuta attraverso un comportamento operante si accompagna, nel maiale, a un aumento dei corticosteroidi plasmatici.
Katz e Carroll (v., 1978) provocano, tramite iniezione di desametasone, una forte riduzione dell'attività motoria diurna. Micco e altri (v., 1979) ottengono, con un'iniezione di corticosterone, che si lega selettivamente ai neuroni della commissura anteriore dell'ippocampo, una repressione cronica e moderata dell'attività spontanea. Analogamente, secondo Mobley e Sulser (v., 1980) i corticoidi possono alterare la sensibilità del sistema dell'adenilatociclasi accoppiata ai recettori della NA nel cervello, sistema che, come sappiamo, facilita l'attività spontanea e di evitamento.
Ancora di recente, aggiungendo le loro osservazioni a quelle di numerosi autori precedenti, Diebold e altri (v., 1978) hanno notato l'aumento della cortisolemia nei pazienti depressi, e Hall e altri (v., 1979) hanno constatato che i pazienti che ricevevano 40 mg di prednisone al giorno presentavano, fin dai primi cinque giorni, fragilità emotiva, ansia, distrazione, eloquio contratto, insonnia, depressione, perplessità, mutismo, turbe dello schema corporeo, deliri e apatia. Träskman e altri (v., 1980) trovano un aumento altamente significativo del cortisolo e della 5-HIAA nell'LCR dei depressi e dei suicidi.
L'asse ipofiso-surrenale non è l'unico a essere turbato dall'inibizione dell'azione. Ruzsas e altri (v., 1979) mettono in evidenza l'aumento del rapporto fra 131I della tiroide e del siero (T/S), l'aumento del tasso ematico e pituitario di TSH, quello del tasso ematico di tiroxina e quello di TRH nell'ipotalamo, in seguito a somministrazione di pCPA e a sezione delle vie serotoninergiche del tronco cerebrale. Il sistema serotoninergico sarebbe quindi un inibitore di questo insieme endocrino. Ora, si conosce l'importanza, altrove discussa, che si è recentemente attribuita alla TRH nel trattamento delle depressioni. Di Renzo e altri (v., 1979) confermano questa azione inibitrice della 5-HT sulla liberazione di TRH-TSH.
Un altro fatto noto è che la pCPA favorisce il comportamento sessuale del ratto maschio. Larsson e altri (v., 1978) hanno appurato sperimentalmente, su basi morfologiche e comportamentali, che le vie ascendenti serotoninergiche, che innervano l'ipotalamo e l'area preottica, esercitano un'azione inibitrice sul controllo, da parte del testosterone, del comportamento sessuale del ratto maschio. Se i glucocorticoidi favoriscono l'inibizione comportamentale e l'estinzione di un comportamento operante, l'ACTH, viceversa, aumenta l'attività locomotoria e ritarda l'estinzione (v. de Wied, 1967; v. Bohus e Lissak, 1968; v. Bohus, 1968; v. McEwen e Weiss, 1970).
d) Integrazione dei fatti sperimentali nel concetto di stress
A. La risposta a stimoli nocicettivi attiva il sistema reticolare e l'ipotalamo posteriore. La fuga e la lotta sono la conseguenza comportamentale dell'attivazione del PVS, cui si accompagna una liberazione adrenomidollare di CA, soprattutto di A. Il PVS, sistema colinergico, provocherà anche una liberazione di CRF e, di conseguenza, di ACTH, che promuoverà l'attività locomotoria di fuga e di lotta. La liberazione secondaria di glucocorticoidi ristabilirà un'attività media, attraverso il ripristino dell'equilibrio tra sistema attivatore e sistema inibitore dell'azione. Se l'azione si è dimostrata efficace, si nota un ritorno all'eccitabilità neuronica di partenza; in queste condizioni non si può parlare di ‛stress': si tratta di una risposta fisiologica all'ambiente.
B. Se l'azione risulta inefficace, interverrà il sistema inibitore dell'azione e la liberazione di glucocorticoidi manterrà lo stato di allarme ‛in tensione'. Le vie nervose utilizzate in questo caso a partire dal sistema reticolare saranno vie talamiche specifiche, che raccorderanno i nuclei intralaminari del talamo, la cui eccitabilità dipende dai glucocorticoidi (v. Koryani ed Endroczi, 1970). Lo stato di ‛attesa in tensione' del momento di agire è all'origine della liberazione periferica di NA e sarà mantenuto fintantoché non si presenterà la possibilità di realizzare un'azione efficace: si tratta, a mio avviso, di ciò che si è convenuto di chiamare uno ‛stato di stress'. Questo stato implica necessariamente il ‛ricordo' dell'inefficacia dell'azione, visto che la risposta neuroendocrina può riapparire se si rimette l'animale nell'ambiente dove è stato aggredito, senza però fargli subire una nuova aggressione (v. Cassens e altri, 1980).
e) Esame schematico delle conseguenze dello stress
Contrariamente a quanto abbiamo visto a proposito degli stati di shock, negli stati di stress le lesioni sono secondarie al persistere della reazione all'ambiente. Favorevole in un primo tempo, questa reazione diventa nociva per la sua persistenza, che dipende dall'impossibilità di controllare le caratteristiche dell'ambiente attraverso l'azione.
Abbiamo già indicato che le conseguenze nocive di questa reazione provengono soprattutto dal fatto che i glucocorticoidi bloccano le difese immunitarie e la liberazione di NA provoca una vasocostrizione generalizzata. A partire dalle mie prime osservazioni (v. Laborit, 1974 e L'inhibition..., 1979), i lavori concernenti l'inibizione dell'azione come fattore che provoca effetti di patologia generale si sono moltiplicati: citiamo l'importante lavoro di Riley (v., 1981). Numerosi lavori sono stati anche dedicati ai rapporti fra il sistema nervoso centrale e il sistema immunitario (v. Spector e Korneva, 1981; v. Stein e altri, 1981; v. Gross e altri, 1982; v. Fauman, 1982). Certi lavori recenti mostrano anche che gli ESP inevitabili (ESPi) aumentano, nel ratto, il numero di metastasi polmonari del carcinoma di Walker-256, iniettato dieci giorni prima (v. Simon e altri, 1980), o la crescita del mastocitoma P 815 nel topo (v. Sklar e Anisman, 1979), mentre gli ESP evitabili non producono questi effetti, contrariamente alla stimolazione elettrica del mesencefalo, che invece li produce. Analogamente, solo il 27% dei ratti sottoposti a un ESPi rigettano il carcinoma di Walker-256, mentre ben il 63% dei ratti sottoposti a uno shock evitabile lo rigettano (v. Visintainer e altri, 1982).
Ora, gli ESPi aumentano il tasso di corticosterone plasmatico e diminuiscono il numero di linfociti T circolanti, per involuzione timica (v. Laudenslager e altri, 1983; v. Keller e altri, 1981). La distruzione del VMI e dell'eminenza mediana (ME) sopprime l'ipertensione in seguito a iniezione i.v.c. di angiotensina Il (v. Johnson e altri, 1981); Johnson e altri mostrano che le vie nervose che controllano la risposta cardiovascolare e renale discendono dal VMI e dall'ME. Certo è che il SIA, che comanda la liberazione di NA da parte delle terminazioni nervose del simpatico e la liberazione di glucocorticoidi da parte delle surrenali (il primo effetto provoca la diminuzione della capacità del letto vascolare, il secondo la ritenzione d'acqua e di sali), non può che favorire l'ipertensione che ho chiamato ‛nevrotica', con tutte le sue conseguenze (emorragie cerebrali, infarto, aterosclerosi, ecc.).
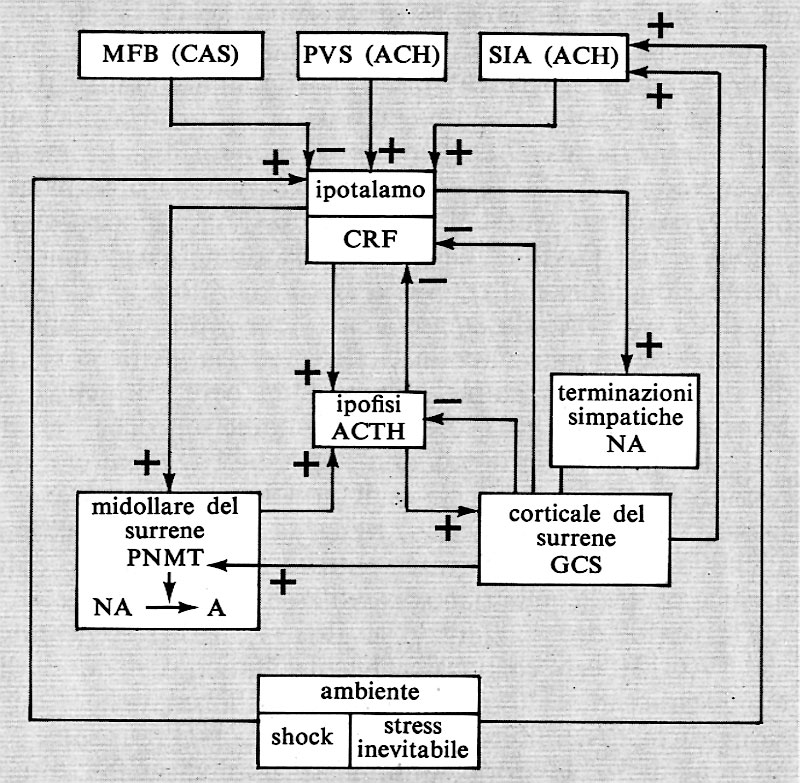
È probabile che la metilazione dei fosfolipidi delle membrane, il loro attacco da parte della fosfolipasi A2 e i prodotti del metabolismo dell'acido arachidonico (leucotrieni, prostaglandine, tromboxano) intervengano anch'essi negli eventi patologici dovuti allo stress.
Così l'inibizione dell'azione e le sue conseguenze parrebbero inglobare tutte le forme patologiche somatiche, infettive, tumorali e degenerative. Tsuda e altri (v., 1983) mostrano che le lesioni e le ulcere gastriche sono molto più gravi nei ratti che subiscono un ESPi che in quelli che possono evitare la punizione attraverso un comportamento operante. Per quel che riguarda le malattie mentali, non possono non dipendere anch'esse dai fattori descritti (v. Laborit, L'inhibition..., 1979 e 1981). Si comprende anche come ciò che si è convenuto di chiamare ‛stress' sia un problema sempre più attuale, dovuto alla vita moderna, ma la sua soluzione, forse, va ricercata più a livello economico, sociale e politico, che non medico (v. Laborit, L'inhibition..., 1979 e 1983). V. anche psicosomativa.
5. Il trattamento
Sarò estremamente succinto per quel che riguarda il trattamento degli stati di shock e di stress, a causa dell'estensione dei problemi e della varietà delle risposte terapeutiche proposte. Non fornirò che le linee essenziali delle metodiche terapeutiche, in relazione allo studio fisiopatologico presentato.
a) Trattamento degli stati di shock
Non si ripeterà mai abbastanza che, dato l'intreccio fra sindrome lesionale e sindrome da reazione, le due sindromi vanno trattate congiuntamente. In quest'ottica il trattamento della lesione ha la precedenza, e ciò è evidente: a seconda della natura della lesione, si farà ricorso alla trasfusione, all'arresto delle emorragie, al trattamento chirurgico delle ferite, all'immobilizzazione delle fratture, ai vari antibiotici negli shock tossinfettivi, agli antitossici nelle intossicazioni, ecc. Si ripristinerà una ventilazione efficace e un corretto funzionamento cardiovascolare e si provvederà al trattamento del dolore.
Il trattamento della sindrome da reazione, altrettanto importante, rientra più propriamente nel quadro del nostro studio fisiopatologico.
1. Trattamento dell'ipovolemia. - Le perdite di liquidi, dovute a emorragia, a perdite di plasma in caso di ustioni, a perdite di elettroliti in caso di diarrea, di vomito, di disidratazione, a perdite locali a livello dei tessuti (traumi, crush injury, ecc.) debbono essere compensate iniettando sangue o soluzioni sostitutive. Le quantità da iniettare saranno controllate attraverso le variazioni della pressione arteriosa, la frequenza cardiaca, la portata urinaria, la irrorazione cutanea e sottoungueale, l'auscultazione e la radioscopia dei polmoni. Il controllo della pressione venosa centrale fornirà utili informazioni. La parte del sangue e dei cristalloidi nella rianimazione liquida è ben codificata e le decisioni da prendere spettano al rianimatore.
2. Trattamento metabolico. - Insisto sul fatto che l'apporto di liquidi deve costituire il supporto di un apporto metabolico. Il ruolo del glucosio nel ripristino di un'attività metabolica sia glicolitica sia mitocondriale è ormai un fatto assodato (v. Laborit, Bases..., 1958). Il glucosio sarà somministrato in soluzioni ipertoniche (dal 10 al 30%), poiché la sua utilizzazione metabolica rischia, in soluzione isotonica, di lasciare una massa idrica troppo rilevante (v. Laborit, Intérêt majeur..., 1958; v. Stremple e altri, 1976): associato all'insulina e al potassio in una soluzione ‛ripolarizzante' (v. Laborit e Huguenard, 1956) si è dimostrato efficace in tutte le sindromi di insufficienza cardiovascolare, grazie alla sua azione sulla dinamica cardiaca e alla sua azione vasodilatatrice nel dominio splancnico. La somministrazione di un sale di calcio agisce spesso in maniera favorevole sulla contrattilità cardiaca.
Il diidrossiacetone, che consente una ripresa della glicolisi, ha dato buoni risultati sperimentali, pur non essendo stato adottato nella clinica, se non per quel che riguarda la conservazione del sangue e del tasso di 2,3-DPG (v. Laborit e Lamothe, 1972). Abbiamo anche mostrato che un amminoacido precursore delle catecolammine, la L-tirosina, possiede proprietà centrali, la cui utilizzazione nel trattamento dello shock emorragico è estremamente benefica, mentre, di per sé, non esercita alcuna azione sul sistema vasomotore o endocrino (v. Laborit, 1972).
3. Trattamento farmacologico. - Malgrado sia ben dimostrato il ruolo scatenante delle catecolammine nell'insorgere dello shock e nel suo evolvere verso l'irreversibilità, certi terapeuti, obnubilati da un sintomo, il crollo della pressione arteriosa, ancora somministrano dei simpaticomimetici. Di fronte all'evidente nocività della NA e dell'A, alcuni si sono limitati alla dopammina, senza conseguire risultati sempre convincenti. Pur essendo utilizzabili in certi casi di collasso, attualmente si ammette in generale che i simpaticomimetici non sono indicati nel trattamento degli stati di shock. La distinzione tra collasso e shock assume in questo caso tutta la sua pregnanza.
I simpaticolitici e i vasodilatatori, al contrario, hanno conquistato un posto di rilievo nella profilassi e nel trattamento dello shock. La simpaticolisi si può ottenere attraverso neuroplegici (fenotiazine, aloperidolo), che presentano il vantaggio di esercitare un'azione centrale e periferica, o, meno spesso, attraverso ganglioplegici e simpaticolitici (fentolammina) ad azione periferica specifica. Il miglioramento della irrorazione dei tessuti deve, evidentemente, accompagnare il riempimento vascolare (v. Laborit, Réaction..., 1952), senza il quale l'ipotensione associata allo shock rischierebbe di compromettere la circolazione coronarica e cerebrale.
L'utilizzazione dei glucocorticoidi, di cui alcuni hanno descritto gli effetti favorevoli, è ancora in discussione. Resta da parlare del nalossone, antagonista degli oppiacei: Holaday e Faden (v., 1978) e numerosi autori dopo di loro hanno mostrato che esso può opporsi all'ipotensione secondaria, all'emorragia e all'iniezione di endotossina, senza dubbio opponendosi all'effetto ipotensivo delle endorfine.
Sapendo che la pro-oppiomelanocortina liberata al livello dei centri fornisce contemporaneamente l'ACTH e le endorfine, l'azione del fredback negativo dei corticoidi potrebbe spiegare la loro azione terapeutica favorevole, secondo alcuni. Notiamo, di sfuggita, che queste terapie si oppongono anche a una reazione detta ‛di difesa', supposta favorevole.
4. Trattamento dello squilibrio acido-base. - È evidente che, essendo l'acidosi la conseguenza dell'ipossia dei tessuti, è soprattutto attraverso il ripristino di una normale pressione di perfusione dei tessuti e di una normale attività metabolica che questi squilibri vanno trattati. Tuttavia è utile correggere rapidamente il pH, il cui abbassamento si oppone ai summenzionati scopi terapeutici (bicarbonato di sodio o tris(idrossimetil)amminometano: Tris) e correggere, se possibile, la ‛caduta' della pressione parziale di CO2 (carbogeno).
Infine, nei casi gravissimi, un'ipotermia generale moderata (da 30 a 35 °C), sotto neuroplegia (ibernazione artificiale); permette ancora di equilibrare l'intensità del metabolismo cellulare, riducendolo; ciò è necessario, date le scarse possibilità di approvvigionamento, dovute alle funzioni ventilatorie e cardiovascolari difettose. L'ipotermia offre anche la possibilità di una dilazione, necessaria perché si faccia sentire l'azione di tutte le terapie farmacologiche (v. Laborit, Clinical..., 1979).
b) Trattamento degli stati di stress
Lo sviluppo della neuropsicofarmacologia rende attualmente impossibile una qualsiasi sistematizzazione. Se si ammette che l'inibizione dell'azione è all'origine dello sviluppo di quasi tutti i processi morbosi, non genetici e non traumatici, si comprende come si faccia ricorso ad agenti farmacologici estremamente vari. Per trattare le conseguenze dello stress - ansia, depressione con le sue conseguenze, insonnia, turbe somatiche varie - si fa ricorso ad ansiolitici (benzodiazepine) e ad antidepressivi, il cui scopo è di facilitare i meccanismi catecolamminergici centrali (inibitori della monoamminossidasi, IMAO, o composti triciclici, imipramina e analoghi).
Vorrei limitarmi a segnalare, in conclusione, che la farmacologia finora si è preoccupata principalmente di ripristinare l'attività favorendo i sistemi centrali che ne sono responsabili. Tuttavia si può ugualmente pensare di inibire il sistema inibitore (SIA). Certe molecole recentemente sintetizzate sembrano capaci di raggiungere questo scopo (v. Laborit, 1981).
L'interdisciplinarità sembra ancora più necessaria nella profilassi e nel trattamento degli stati di stress che non nella profilassi e nel trattamento degli stati di shock, dato che psicologia, antropologia, sociologia, economia e politica, vale a dire l'insieme delle scienze umane, partecipano alla loro comprensione.
BIBLIOGRAFIA
Andersen, D., Shoemaker, W.L., Effect of hemorrhage on hepatic potassium movements, in ‟Proceedings of the Society for Experimental Biology and Medicine", 1967, CXXIV, pp. 840-844.
Anisman, H., Remington, G., Sklar, L., Effect of inescapable shock on subsequent escape performance: catecholaminergic and cholinergic mediation of response initiation and maintenance, in ‟Psychopharmacoloby", 1979, LXI, pp. 107-124.
Axelrod, J., Reisine, T.D., Stress hormones: their interaction and regulation, in ‟Science", 1984, CCXXIV, pp. 452-459.
Beagley, G.H., Beagley, W.K., Alleviation of leamed hellesness following septal lesions in rats, in ‟Physiological psychology", 1978, VI, pp. 241-244.
Bernard, Cl., Leçons sur les phénomènes de la vie communs aux animaux et aux végétaux, Paris 1878.
Bohus, B., Pituitary ACTH release and avoidance behaviour of rats with cortisol implants in mesencephalic reticular formation and median eminence, in ‟Neuroendocrinology", 1968, VI, pp. 355-365.
Bohus, B., Lissak, K., Adrenocortical hormones and avoidance behavior of rats, in ‟International journal of neuropharmacology", 1968, VII, pp. 301-306.
Briaud, B., Enjalbert, A., Mialhe, C., Kordon, C., Subcellar distribution of corticotropin-releasing factor in the medio-basal hypothalamus of the rat, in ‟Neuroendocrinology", 1979, XXVIII, pp. 371-376.
Broder, G., Weil, M. H., Excess lactate: an index of reversibility of shock in human patients, in ‟Science", 1964, CXLIII, pp. 1457-1459.
Brown, L., Rosellini, R.A., Samuels, O.B., Riley, E.P., Evidence for a serotonergic mechanism of the learned helplessness phenomenon, in ‟Pharmacology, biochemistry and behavior", 1982, XVII, pp. 877-884.
Bruce, B.M., Carrington, C.D., Grossman, S.P., Passive avoidance behavior in lean and obese rats with ventromedial hypothalamic lesion, in ‟Physiology and behavior", 1978, XX, pp. 57-66.
Buckingham, J.C., Hodges, J.R., Hypothalamic receptors influencing the secretion of corticotropin releasing hormone in the rat, in ‟Journal of physiology", 1979, CCXC, pp. 421-432.
Cannon, W.B., The wisdom of the body, New York 1932 (tr. it.: La saggezza del corpo, Milano 1956).
Caroll, B.J., The dexamethasone suppression test for melancholia, in ‟British journal of psychiatry", 1982, XXXVIII, pp. 15-22.
Cassens, G., Roffman, M., Kuruc, A., Orsulak, P. J., Schildkraut, J. J., Alterations in brain norepinephrine metabolism induced by environmental stimuli previously paired with inescapable shock, in ‟Science", 1980, CCIX, pp. 1138-1139.
Chambers, R., Zweifach, B.W., Topography and function of the mesenteric capillary circulation, in ‟American journal of anatomy", 1944, LXXV, pp. 173-205.
Dantzer, R., Arnone, A.H., Mormede, P., Effects of frustration on behavior and plasma corticosteroid level in pigs, in ‟Physiology and behavior", 1980, XXIV, pp. 1-4.
Davis, B.M., Davis, K.L., Cholinergic mechanism and anterior pituitary hormones secretion, in ‟Biological psychiatry", 1980, XV, pp. 303-310.
Dawson, R.G., McGaugh, J.L., Electroconvsulsive shock effects on a reactivated memory trace; further examination, in ‟Science", 1969, CLXVI, pp. 525-527.
Diebold, K., Vecsei, P., Jackenkroll, R., Marquetand, D., Reindell, A., Investigation of the plasma cortisol in psychiatric diagnostic, in ‟Archiv für Psychiatrie und Nervenkrankheiten", 1978, CCXXVI, pp. 29-36.
Di Renzo, G.F., Quattruone, A., Schettini, G., Preziosi, P., Effect of selective lesioning of serotonin-containing neurons on the TSH-ihibiting action of d-fenfluramine in male rats, in ‟Life sciences", 1979, XXIV, pp. 489-494.
Doerr, P., Berger, M., Physostigmine-induced escape from dexamethasone suppression in normal adults, in ‟Biological psychiatry", 1983, XVIII, pp. 261-268.
Dunn, A. J., Kramarcy, N.R., Neurochemical responses in stress: relationships between the hypothalamic-pituitary-adrenal and catecholamine systems, in Handbook of psychopharmacology, vol. XVIII (a cura di L.L. Iversen, S.D. Iversen, S.H. Snyder), New York 1984, pp. 455-515.
Duve, C. de, Pressman, B.C., Gianetto, R., Wattiaux, R., Appelmans, F., Tissue fractionation studies. VI. Intracellular distribution patterns of enzymes in rat liver tissue, in ‟Biochemical journal", 1955, LX, pp. 604-(17.
Ely, D.K., Henry, J. P., Neuroendocrine response, patterns in dominant and subordinate mice, in ‟Hormones and behavior", 1978, X, pp. 156-169.
Erlanger, J., Gasser, H., Studies in secondary shock. III. Circulatory failure due to adrenaline, in ‟American journal of physiology", 1919, IL, pp. 345-353.
Esina, N. I., Changes in 2,3-diphosphoglyceric acid concentration in erythrocytes during the postresuscitation period in patients with a history of massive blood loss and trauma, in ‟Patologiceskaja fiziologija i eksperimental'naja terapija", 1981, pp. 27-31.
Euler, U.S. von, Identification of sympathomimetic ergone of adrenergic nerve of cattle. (Simpathyn N with laevo-noradrenaline), in ‟Acta physiologica scandinavica", 1948, XVI, pp. 63-74.
Fahreus, R., The suspension-stability of the blood, in ‟Acta medica scandinavica", 1921, LV, pp. 1-228.
Fauman, M.A., The central nervous system and the immune system, in ‟Biological psychiatry", 1982, XVII, pp. 1459-1481.
Filaretov, A.A., Vassilevskaya, L.V., Electrical activity of hypothalamus during activation of pituitary-adrenocortical system, in ‟Brain research", 1978, CLVI, pp. 67-74.
Fine, J., Frank, E.O., Ravin, H.A., Ruttenburg, S.H., Schweinburg, F.B., The bacterial factor on traumatic shock, in The biochemical response to injury, vol. I (a cura di H.B. Stoner e C.J. Threlfall), Oxford 1960, pp. 377-396.
Fredholm, B.B., Farnebo, L.O., Hamberger, B., Plasma catecholamines cyclic AMP and metabolic substrates in hemorrhagic shock of the rat. The effect of adrenal demedullation and 6-OH-dopamine treatment, in ‟Acta physiologica scandinavica", 1979, CV, pp. 490-495.
Freud, S., The standard edition of the complete work of Sigmund Freud, London 1953.
Fuller, R.W., Snody, H.L., Effect of serotonin-releasing drugs on serum corticosterone concentration in rats, in ‟Neuroendocrinology", 1980, XXXI, pp. 96-100.
Graeff, E.G., Silveira Filho, N.G., Behavioral inhibition induced by electrical stimulation of the median raphe nucleus of the rat, in ‟Physiology and behavior", 1978, XXI, pp. 477-484.
Gross, R.J., Brooks, W.H., Roszman, T.L., Markesbery, W.R., Hypothalamic immune interactions. Effects of hypophysectomy on neuroimmunomodulation, in ‟Journal of neurological science", 1982, LIII, pp. 557-566.
Hall, R. C. W., Popkin, M. K., Stickney, S. K., Gardner, E.R., Presentation of the steroid psychoses, in ‟Journal of nervous and mental disease", 1979, CLXVII, pp. 229-236.
Halpern, B.N., Stiffel, C., Biozzi, G., Benacerraf, B., Détermination des constantes de l'activité granulopexique du système réticulo-endothélial chez le lapin, in ‟Comptes rendus de la Société de Biologie", 1954, CXLVIII, pp. 240-243.
Hardway, R.M., Drake, D.E., Prevention for ‛irreversible' hemorrhagic shock with fibrinolysis, in ‟Annals of surgery", 1963, CLVII, pp. 39-46.
Hoffman, D.L., Trowill, J.A., Modulation of intracranial reward punishment interaction by scopolamine, in ‟Physiology and behavior", 1979, XXIII, pp. 223-228.
Holaday, J.W., Faden, A.I., Naloxone reversal of endotoxine hypotension suggest role of endorphines in shock, in ‟Nature", 1978, CCLXXV, pp. 450-451.
Huckabee, W.E., Relatinships of pyruvate and lactate during anaerobic metabolism. I. Effect of infusion of pyruvate or glucose and of hyperventilation, in ‟Journal of clinical investigation", 1958, XXXVII, pp. 244-254.
Johnson, A.K., Buggy, J., Fink, G.D., Brody, M. J., Prevention of renal hypertension and of the central pressor effect of angiotensin by ventromedial hypothalamic ablation, in ‟Brain research", 1981, CCV, pp. 255-264.
Katz, R.J., Carroll, B.J., Endocrine control of psychomotor activity in the rat: effects of chronic dexamethasone upon general activity, in ‟Physiology and behavior", 1978, XX, pp. 25-30.
Keller, S.E., Weiss, J. M., Schleiffer, S.J., Miller, N.E., stein, M., Suppression of immunity by stress: effect of a graded series of stressors on lymphocyte stimulation in the rat, in ‟Science", 1981, CCXIII, pp. 1397-1400.
Kelsey, J.E., The role of norepinephrine and acetylcholine in mediating escape deficits produced by inescapable shocks, in ‟Behavioral and neural biology", 1983, XXXVII, pp. 325-331.
King, B.M., Grossman, S.P., Impaired and enhanced shuttle box avoidance behavior following ventro-medial hypothalamic lesions in rats, in ‟Physiology and behavior", 1978, XX, pp. 51-56.
Knisely, M. H., Stratman-Thomas, W.K., Elliott, T.S., Block, E.H., Knowlesi malaria in monkeys. I. Microscope pathological circulatory physiology of rhesus monkeys during acute plasmodium knowolesi malaria, in ‟Journal of the National Malaria Society", 1945, IV, pp. 285-300.
Koryani, L., Endroczi, E., Influence of pituitary-adreno-cortical hormones on hypothalamo-cortical and brainstem limbic circuits, in Progress in brain research, vol. XXXII, Pituitary, adrenal and the brain (a cura di D. de Wied e J.A.W.N. Weijnen), Amsterdam 1970, pp. 120-130.
Kunz, E., Valette, N., Laborit, H., Rôle de l'apprentissage dans le mécanisme d'inhibition comportemental et de l'hypertension artérielle consécutive à l'application de stimulus aversifs sans possibilité de fuite ou de lutte, in ‟Agressologie", 1974, XV, pp. 381-385.
Laborit, H., La vasoplégie progerssive dans la conduite de la perfusion au cours des déséquilibres vaso-moteurs post-agressifs, in ‟Presse médicale", 1952, LX, p. 693.
Laborit, H., Réaction organique à l'agression et choc, Paris 1952, 19542.
Laborit, H., Résistance et soumission en physiobiologie, Paris 1954.
Laborit, H., Bases physiobiologiques et principes généraux de réanimation, Paris 1958.
Laborit, H., Intérêt majeur du glucose hypertonique en réanimation, in ‟Presse médicale", 1958, LXVI, pp. 444-445.
Laborit, H., Treating of agression and reduction of fatigue syndromes with L-tyrosine, U.S. Patent Office, n. 3.651.231 del 21-3-1972.
Laborit, H., Les comportements, Paris 1973.
Laborit, H., Bases neurophysiologiques et biologiques des comportements d'évitements actifs et passifs. Conséquences somatiques. Le niveau comportemental. Problèmes sémantiques, in ‟Annales médico-psychologiques", 1974, I, pp. 573-603.
Laborit, H., On the mechanism of activation of the hypothalamopituitary-adrenal reaction to changes in the environment (the ‛alarm reaction'), in ‟Resuscitation", 1976, V, pp. 19-30.
Laborit, H., La dihydroxyacétone dans le traitement des états de choc et la conservation du sang, in ‟Médecine et hygiène", 1978, XXXVI, pp. 1072-1085.
Laborit, H., Clinical hypothermia, in Body temperature (a cura di P. Lomax ed E. Schönbaum), New York-Basel 1979.
Laborit, H., L'inhibition de l'action. Biologie, physiologie, psychologie, sociologie, Paris-Montréal 1979.
Laborit, H., Physiopathologie de l'inhibition et proposition d'une réponse thérapeutique originale, in ‟Agressologie", 1981, XXII, pp. 45-55.
Laborit, H., La colombe assassinée, Paris 1983.
Laborit, H., Baron, C., Ferran, C., Henriet, I., Assay of cerebral catecholamines, serotonin and their metabolites in experimental hemorrhagic shock in rats, in ‟Research communications in chemical pathology and pharmacology", 1984, XLIII, pp. 425-433.
Laborit, H., Baron, C., Ferran, C., Henriet, I., Action of certain pharmacological associations on the level of cerebral biogenic amines and some metabolites in the curative treatment of hemorrhagic shock in rats, in ‟Research communications in chemical pathology and pharmacology", 1984, XLIII, pp. 435-447.
Laborit, H., Baron, C., Lamothe, C., Laborit, G., Variations du 2,3-diphosphoglycérate (2,3-DPG) globulaire au cours du choc hémorragique et sous l'action d'agents activateurs de la glycolyse, in ‟Agressologie", 1972, XIII, pp. 233-241.
Laborit, H., Baron, C., Laurent, J., Faits expérimentaux neurophysiologiques et biologiques montrant l'existence d'activités fonctionnelles séparées pour l'hippocampe dorsal et ventral, in ‟Agressologie", 1977, XVII, pp. 27-32.
Laborit, H., Baron, C., Weber, B., Essai de mise en évidence expérimentale du rôle des lysosomes dans l'évolution du choc hémorragique irréversible, in ‟Agressologie", 1967, VIII, pp. 23-33.
Laborit, H., Huguenard, P., Échanges ioniques, tonus vasculaire et métabolisme cellulaire. Bilan d'une étude expérimentale et clinique, in ‟Presse médicale", 1956, LXIV, pp. 605-608.
Laborit, H., Kunz, E., Baron, C., Mise en évidence d'une commande centrale distincte des systèmes adrénergiques et noradrénergiques périphériques. Action d'un α-bloquant (dibénamine) par voie intraventriculaire cérébrale sur la pression artérielle du lapin surrénalectomisé ou non, in ‟Agressologie", 1974, XV, pp. 327-332.
Laborit, H., Kunz, E., Thuret, F., Baron, C., Action de l'hydrocortisone sur le taux de norépinéphrine plasmatique che zle lapin surrénalectomisé, in ‟Agressologie", 1975, XVI, pp. 351-354.
Laborit, H., Lamothe, C., Sur l'utilisation de la dihydroxyacétone comme nouveau substrat dans les milieux de conservation du sang, in ‟Agressologie", 1972, XIII, pp. 243-246.
Laborit, H., Morand, P., Choc traumatique et cholinestérase, in ‟Mémoires de l'Académie de Chirurgie. Paris", 1947, LXXIV, pp. 261-263.
Laborit, H., Obrenovitch, L., Leguen, P., Jouany, J. M., Action des sels potassiques et magnésiens de l'acide DL-aspartique sur l'hyperammoniémie du choc hémorragique expérimental, in ‟Comptes rendus de la Société de Biologie", 1958, CLII, pp. 942-944.
Laborit, H., Weber, B., Baron, C., Étude expérimentale des perturbations métaboliques du choc hémorragique irréversible et de leur correction thérapeutique. Rôle de la carboxylation du pyruvate, in ‟Agressologie", 1966, VII, pp. 395-425.
Larsson, K., Fuxe, K., Everitt, B. J., Holmgren, M., Sodersten, P., Sexual behavior in male rats after intracerebral injection of 5,7-dihydroxytryptamine, in ‟Brain research", 1978, CXLI, pp. 293-303.
Laudenslager, M. L., Ryan, S. M., Drugan, R.C., Hyson, R.L., Maier, S.F., Coping and immunosuppression: inescapable but not escapable shock suppresses lymphocyute proliferation, in ‟Science", 1983, CCXXI, pp. 568-570.
Leshner, A.L., Kom, S.J., Mixon, J.F., Rosenthal, C., Besser, A. K., Effects of corticosterone on submissiveness in mice: some temporal and theoretical considerations, in ‟Physiology and behavior", 1980, XXIV, pp. 283-288.
Leshner, A.L., Potitch, J.A., Hormonal control of submissiveness in mice: irrelevance of the androgens and relevance of the pituitary-adrenal hormones, in ‟Physiology and behavior", 1979, XXII, pp. 531-534.
Levi, L. (a cura di), Society, stress and diseases, vol. I, The psychosocial environment and psychosomatic diseases, London-New York 1971.
McEwen, B.S., Weiss, J. M., The uptake and action of corticosterone: regional subcellular studies on rat brain, in Progres in brain research, vol. XXXII, Pituitary, adrenal and the brain (a cura di D. de Wied e J.A.W.N. Weijnen), Amsterdam 1970, pp. 57-70.
Makara, G.B., Stark, E., Palkovits, M., Re-evaluation of the pituitary-adrenal response to ether in rats with various cuts around the medial basal hypothalamus, in ‟Neuroendocrinology", 1980, XXX, pp. 38-44.
Mason, J.W., A re-evaluation of the concept of ‛non-specificity' in stress theory, in ‟Journal of psychiatric research", 1971, VIII, pp. 323-333.
Mason, S.T., Fibiger, H.C., Interaction between noradrenegic and cholinergic systems in the rat brain: behavioral function in locomotor activity, in ‟Neuroscience", 1979, IV, pp. 517-526.
Maurer, M., Le ‛stress', une définition impossible: Selye au centre de la controverse, in ‟Médecine et hygiène", 1984, XLII, pp. 2240-2249.
Meyer, J.S., Buckholtz, N.S., Boggan, W.O., Serotonergic stimulatinoof pituitary-adrenal activity in the mouse, in ‟Neuroendocrinology", 1978, XXVI, pp. 312-324.
Micco, D.J., McEwen, B.S., Shein, W., Modulation of behavioral inhibition in appetitive extinction following manipultaion of adrenal steroids in rats: implications for involvement of the hippocampus, in ‟Journal of comparative and physiological psychology", 1979, XCIII, pp. 323-329.
Millan, M.J., Millan, M. H., Herz, A., Lesions of ventral noradrenergic (norepinephrine) bundle affect brain and pituitary pools of endorphines and the response of these in rats, in ‟Neuroendocrinology", 1982, XXXV, pp. 449-456.
Mobley, P.L., Sulser, F., Adrenal corticoids regulate sensitivity of noradrenaline receptor-coupled adenylate cyclase in brain, in ‟Nature", 1980, CCLXXXVI, pp. 608-609.
Modlinger, R.S., Schonmuller, J.M., Aroba, S.P., ACTH release by triptophan in man, in ‟Clinica de endocrinología y metabolismo", 1980, L, pp. 360-363.
Molina, A.F. de, Hunsperger, R.Q., Organisation of subcortical system governing defence and fight reactions in the cats, in ‟Journal of physiology", 1962, CLX, pp. 200-213.
Montanero, N., Dall'Olio, R., Gandolfi, O., Bromolysergide and methyusergide protection against ECS-induced retrograde amnesia, in ‟Neuropsychobiology", 1979, V, pp. 174-180.
Moon, V.H., Shock, its dynamics, occurrence and management, Philadelphia 1942.
Nishizawa, N., Bray, G.A., Ventromedial hypothalamic lesions and the mobilisation of fatty acids, in ‟Journal of clinical investigation", 1978, XLI, pp. 714-721.
Olds, J., Milner, P., Positive re-enforcement produced by electrical stimulation of septal area and other regions of rat brain, in ‟Journal of comparative and physiological psychology", 1954, XLVII, pp. 419-427.
Plaznik, A., Kostowski, W., Bidzinski, A., Hauptmann, M., Effects of lesions of the midbrain raphe nuclei on avoidance learning in rats, in ‟Physiology and behavior", 1980, XXIV, pp. 257-262.
Redgrave, P., Modulation of intracranial self-stimulation behaviour by local perfusions of dopamine, noradrenaline and serotonin within the caudate nucleus and nucleus accumbens, in ‟Brain research", 1978, CLV, pp. 277-296.
Riley, V., Psychoneuroendocrine influences on immunocompetence and neoplasia, in ‟Science", 1981, CCXII, pp. 1100-1109.
Risch, S.C., β-endorphin hypersecretion in depression: possible cholinergic mechanisms, in ‟Biological psychiatry", 1982, XVII, pp. 1071-1079.
Risch, S.C., Cohen, R.M., Jancowsky, D.S., Kalin, N. H., Murphy, D.L., Mood and behavioral effects of physostigmine on humans are accompanied by elevation in plasma β-endorphin and cortisol, in ‟Science", 1980, CCIX, pp. 1545-1546.
Russel, R.W., Macri, J., Central cholinergic involvement in behavioral hyper-reactivity, in ‟Pharmacology, biochemistry and behavior", 1979, X, pp. 43-48.
Ruzsas, C.R., Jozsa, R., Mess, B., Inhibitory role of brain stem serotoninergic neuron system on thyroid function in rat, in ‟Endocrinologia experimentalis", 1979, XIII, pp. 9-18.
Schenberg, L.C., Graeff, E.G., Role of the periaqueducal gray substance in the antianxiety action of benzodiazepines, in ‟Pharmacology, biochemistry and behavior", 1978, IX, pp. 287-296.
Schmidt, D.E., Cooper, D.O., Barrett, R.J., Strain specific alterations in hippocampal cholinergic function following acute footshock, in ‟Pharmacology, biochemistry and behavior", 1980, XII, pp. 277-280.
Schummer, W., Lactic acid as a factor in the production of irreversibility in olighaemic shock, in ‟Nature", 1966, CCXII, pp. 1210-121.
Selye, H., A syndrome produced by diverse noxious agents, in ‟Nature", 1936, CXXXVIII, p. 32.
Selye, H., Heuser, G., Fifth annual report on stress, New York 19566.
Shimazu, T., Matsushita, H., Ishikawa, K., Hypothalamic control of liver glycogen metabolism in adult and aged rats, in ‟Brain research", 1978, II, pp. 343-352.
Simon, R.H., Lovett, E.I.III, Tomaszek, D., Lundy, J., Electrical stimulation of the midbrain mediates metastatic tumor growth, in ‟Science", 1980, CCIX, pp. 1132-1133.
Sithichoke, N., Malasanos, L.J., Marotta, S.F., Cholinergic infuences on hypothalamic-pituitary-adrenocortical activity of stressed rats on approach utilizing choline deficients diets, in ‟Acta endocrinologica", 1978, LXXXIX, pp. 737-743.
Sklar, L.S., Anisman, H., Stress and coping factors influence tumor growth, in ‟Science", 1979, CCV, pp. 513-515.
Sourkes, T.L., Pathways of stress in the CNS, in Progress in neuropsychopharmacological and biological psychiatry, vol. VII, n. 2, Oxford-New York 1983, pp. 389-411.
Spector, N.H., Korneva, E.A., Neurophysiology, immunophysiology and neuroimmunomodulation, in Psychoneuroimmunology, behavioral medicine (a cura di R. Ader), vol. XXI, New York 1981, pp. 449-472.
Spindel, E., Negro-Vilar, A., Serotonin and ACTh release: direct effects at the anterior pituitary level and potentiation of arginine vasopressin-induced ACTH release, in ‟Endocrinology", 1983, CXII, pp. 1217-1223.
Stein, M., Schleifer, S.J., Keller, S.E., Hypothalamic influences on immunoresponses, in Psychoneuroimmunology, behavioral medicine (a cura di R. Ader), vol. XXI, New York 1981, pp. 429-447.
Stephens, D.N., Herberg, L.J., Dopamine-acetuylcholine ‛balance' in nucleus and corpus striatum and its effects on hypothalamic self-stimulation, in ‟European journal of pharmacology", 1979, LIV, pp. 331-344.
Stremple, J.F., Thomas, H., Sakach, V., Trelka, D., Myocardial utilization of hypertonic glucose during hemorrhagic shock, in ‟Surgery", 1976, LXXX, pp. 4-43.
Szafarczyk, A., Ixart, G., Malaval, F., Nougier Soule, J., Assenmacher, I., Effects of lesions of the suprachiasmatic nuclei and of p-chlorophenylalaline on the circadian rhythms of ACTh and corticosterone in the plasma and a locomotor activity of rats, in ‟Journal of endocrinology", 1979, LXXXIII, pp. 1-16.
Tachiki, K.H., Tateishi, T.T., Nishiwaki, K.K., Nakamura, E., Nagayama, H., Takahashi, R., Animal model of depression. III. Mechanisms of action of tetrabenazine, in ‟Biological psychiatry", 1978, XIII, pp. 429-444.
Thornton, E.W., Goudie, A.J., Evidence for the role of serotonin in the inhibition of specific motor responses, in ‟Psychopharmacology", 1978, LX, pp. 73-80.
Träskman, L., Tybring, G., Asberg, M., Bertilsson, L., Lantto, O., Schralling, D., Cortisol in the CSF of depressed and suicidal patients, in ‟Archives of general psychiatry", 1980, LXXIV, pp. 722-725.
Tsuda, A., Tanaka, M., Nishikawa, T., Hirai, H., Effects of coping behavior on gastric lesions in rats as a function of the complexity of coping tasks, in ‟Physiology and behavior", 1983, XXX, pp. 805-808.
Valeri, P., Angelucci, L., Palmery, M., Specific (3H) corticosterone uptake in the hippocampus and septum varies with social settings in mice: hormone receptor autoregulation may be involved, in ‟Neuroscience letters", 1978, IX, pp. 249-254.
Van Bogaert, A., Schepper, J. de, Arterial and venous reponses to hypothalamic stimulation in the dog, in ‟Archives internationales de physiologie et de biochimie", 1978, LXXXVI, pp. 1091-1098.
Visintainer, M.A., Volpicelli, J.R., Seligman, M.E.P., Tumor rejection in rats after inescapable or escapable shock, in ‟Science", 1982, CCXVI, pp. 437-439.
Waldbillig, J.R., The role of the dorsl and median raphe in the inhibition of muricide, in ‟Brain research", 1979, CLX, pp. 341-346.
Weil-Malherbe, H., Bone, A.D., The adrenergic amines of human blood, in ‟Lancet", 1953, I, p. 974.
Weiner, R.I., Ganong, W.F., Role of brain catecholamines and histamine in regulation of anterior or pituitary secretion, in ‟Physiological reviews", 1978, LVIII, pp. 905-976.
Weiss, J.M., Effects of coping responses on stress, in ‟Journal of comparative and physiological psychology", 1968, LXV, pp. 251-260.
Wied, D. de, Opposite effects of ACTH an glucocorticoids on extinction of conditioned avoidance behaviour of rats, in Proceedings of the 2nd International congrss on hormonal steroids. Milan May 1966. Excerpta medica. Congress series n. 132, Amsterdam 1967, pp. 945-951.
Yamamoto, T., Ueki, S., Effects of drugs on hyperactivity and agression induced by raphe lesions in rats, in ‟Pharmacology, biochemistry and behavior", 1978, IX, pp. 821-826.
Zolman, J.F., Mattingly, B.A., Sahley, C.L., Cholinergic involvement in inhibitory behavior of the young domestic chick, in ‟Behavioral biology", 1978, XXIII, pp. 415-432.