
Visione
Visione
di Lamberto Maffei
Visione
sommario: 1. Introduzione. 2. Cenni sull'anatomia del sistema visivo. 3. Elettrofisiologia della visione: a) recettori retinici; b) neuroni retinici; c) organizzazione del campo recettivo delle cellule gangliari retiniche; d) corpo genicolato laterale; e) corteccia visiva; f) architettura funzionale della corteccia visiva: colonne e ipercolonne. 4. Psicofisica della visione: a) adattamento visivo; b) psicofisiologia della brillanza; c) fenomeni di interazione tra zone eccitatorie e inibitorie del sistema visivo rilevati con metodi psicofisici; d) caratteristiche temporali della visione; e) caratteristiche spaziali della visione; f) sensibilità della visione per diversi orientamenti dell'immagine; g) percezione della distanza; h) indizi monoculari; i) visione dei colori. □ Bibliografia.
1. Introduzione
Nella visione è difficile distinguere tra cosa realmente percepiamo e cosa invece è semplice inferenza o interpretazione del nostro cervello. Già Plinio, nella sua Naturalis historia, aveva affermato che l'organo della vista non è l'occhio ma la mente. Anche la vecchia teoria che supponeva che nel nostro cervello si formassero sbiadite copie della realtà o addirittura rappresentazioni punto per punto di essa, vere immagini fotografiche, non può essere più sostenuta. Alcune valutazioni delle caratteristiche degli oggetti, come, per esempio, il loro valore simbolico o estetico, dipendono dall'entrata sensoriale ma, ovviamente, anche dalla conoscenza, dalla cultura, dallo stato d'animo dell'osservatore. La visione può essere considerata quindi un processo di trasformazione e d'interpretazione di un mondo esterno, fisicamente esistente, in un nostro mondo percettivo. Sappiamo per certo che questo processo s'inizia nella retina, con l'eccitazione di milioni di recettori sensibili alla luce, e ha le sue fasi conclusive probabilmente nella corteccia del nostro cervello. Il processo di trasformazione comprende la codificazione del segnale visivo in segnali neuroelettrici, la loro trasmissione lungo le vie nervose e la loro elaborazione nelle diverse stazioni visive, con selezione di quella parte dell'informazione ritenuta più importante e attenuazione o scarto delle altre, probabilmente considerate ridondanti.
La nostra trattazione sarà, per ragioni di spazio, incompleta. Cercheremo di mettere l'accento su recenti risultati di neurofisiologia e psicofisica della visione che sembrano promettenti per sviluppi futuri.
2. Cenni sull'anatomia del sistema visivo
L'anatomia del sistema visivo dei Vertebrati può essere schematizzata come segue.

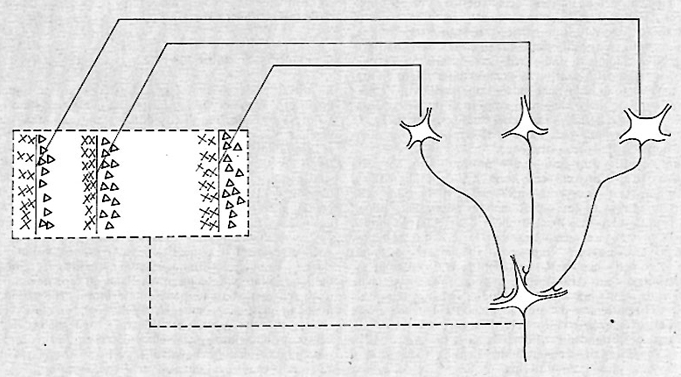
Nella retina (v. fig. 1) molti milioni di recettori trasformano il segnale luminoso in manifestazioni bioelettriche che a loro volta danno origine ai messaggi visivi che attraverso le cellule bipolari e le cellule gangliari vengono trasmessi ai centri nervosi. I fotorecettori nell'uomo, e in molti altri vertebrati, sono di due tipi: coni (6 milioni) e bastoncelli (120 milioni), cosi chiamati da M.J.S. Schultze con riferimento alla forma della loro parte più distale contenente il pigmento. I bastoncelli sono più numerosi nella periferia della retina e assenti dalla sua zona centrale, la fovea, deputata alla fine visione dei dettagli delle forme. I coni e i bastoncelli differiscono sostanzialmente anche nelle loro connessioni con le altre cellule della retina. Nella fovea, dove sono presenti solo coni, le connessioni recettori-cellule bipolari-cellule gangliari presentano scarsa o nessuna integrazione, cioè uno o pochi coni convergono su una cellula bipolare e una o poche cellule bipolari convergono su una cellula gangliare. Il grado d'integrazione aumenta andando verso la periferia della retina, più ricca di bastoncelli, ove centinaia di recettori convergono su una cellula bipolare e decine di neuroni bipolari su una cellula gangliare. È questo diverso grado di convergenza nell'organizzazione nervosa che specializza la periferia della retina, e quindi la visione bastoncellare, per la rivelazione di quantità di luce estremamente piccole (un singolo fotone).
L'alto grado di convergenza porta di conseguenza al fatto che questa regione della retina presenta elevata eccitabilità ma bassa acuità visiva, intendendo con ciò la capacità di percepire un oggetto nello spazio come distinto da un altro a esso vicino. La fovea, invece, data l'organizzazione delle sue connessioni nervose, presenta bassa sensibilità ma elevata acuità visiva; essa è anche specializzata, come vedremo, nella visione dei colori. Per elevate luminanze fotopiche, i bastoncelli contribuiscono scarsamente o non contribuiscono affatto al processo visivo, anche perché la trasmissione dei loro segnali viene inibita dall'attività dei coni, come è stato dimostrato da P. Gouras a livello delle cellule gangliari del gatto (v. Gouras, 1965).
I recettori, oltre a collegarsi con le cellule bipolari, sono anche in connessione tra loro, tramite le cellule orizzontali. A livello dello strato dei neuroni bipolari si trova un altro tipo di cellule, le amacrine, che stabiliscono contatti tra loro, con le cellule bipolari e con le cellule gangliari. Una visione schematica delle connessioni retiniche è illustrata nella fig. 1.
Recenti ricerche di microscopia elettronica hanno individuato ulteriori punti di queste connessioni. Si è visto, per esempio, che l'informazione visiva può arrivare alle cellule gangliari anche tramite la via recettore→cellula bipolare→cellula amacrina→cellula gangliare (v. Dowling e Boycott, 1967) e si è anche accertato che le cellule amacrine hanno contatti nei due sensi con le cellule bipolari. La cellula amacrina, cioè, può ricevere ma anche dare informazioni alla cellula bipolare. Ciò costituirebbe la base anatomica di un circuito di controllo dell'informazione che passa attraverso la cellula bipolare.
Da ciascuna retina dell'uomo partono circa 800.000 fibre, gli assoni delle cellule gangliari. Circa la metà di questi, e precisamente quelli provenienti dalla parte nasale delle due retine, si incrociano nel chiasma ottico e si mettono in comunicazione con un nucleo del talamo dell'altro lato, il corpo genicolato laterale. Le fibre provenienti dalla parte temporale delle due retine non si incrociano e si mettono in comunicazione con il corpo genicolato dello stesso lato. Una parte delle fibre del nervo ottico si mette in comunicazione con una formazione nervosa del mesencefalo, il corpo quadrigemello superiore (collicolo superiore). Dal corpo genicolato partono fibre che direttamente vanno alla corteccia visiva. Dal corpo quadrigemello parte invece una via indiretta che si dirige, almeno in alcuni mammiferi, principalmente all'area visiva 18, dopo aver stazionato nel pulvinar del talamo.
La corteccia visiva possiede nell'uomo un numero enorme di cellule nervose, dell'ordine di un centinaio di milioni, che operano un'ulteriore elaborazione dei segnali luminosi.
Le proiezioni retinogenicolocorticali hanno una ben precisa organizzazione topologica; ogni determinata zona della retina, cioè, proietta a una ben determinata zona del corpo genicolato e quindi della corteccia. La parte di retina che ha di gran lunga la più ampia rappresentazione corticale è la fovea.
Quando si stabilì con certezza che le diverse parti della retina proiettano alle diverse parti della corteccia e del collicolo superiore in maniera ordinata, producendo vere e proprie mappe del mondo visivo, alcuni fisiologi e psicologi pensarono che probabilmente la sola funzione delle vie visive era di ricostruire un'immagine in termini elettrici sulla superficie del cervello. Tuttavia questa ipotesi, anche solo su base strettamente logica, non può essere vera basta ricordare, per esempio, che la mappa del cervello è bidimensionale, mentre lo spazio percettivo è tridimensionale e comprende altre dimensioni, come quella temporale e cromatica.
La corteccia visiva è situata nella porzione occipitale del cervello ed è classicamente divisa in un'area cosiddetta ‛striata', o area 17, dove la via genicolocorticale proietta direttamente, e in due aree associative, aree 18 e 19, dove l'informazione arriva o direttamente dalla corteccia striata o dalla via corpo quadrigemello superiore-pulvinar, come è suggerito dagli esperimenti sul gatto. Vi sono altre aree della corteccia cerebrale che ricevono informazioni correlate con la visione, come la corteccia del lobo inferotemporale, alla cui fisiologia accenneremo in seguito.
3. Elettrofisiologia della visione
a) Recettori retinici
I fotorecettori della retina dei Vertebrati sono, come abbiamo già detto, i coni e i bastoncelli. Oltreché forme e collegamenti nervosi diversi, essi posseggono anche pigmenti diversi. Il più studiato è il pigmento dei bastoncelli, la rodopsina. Questa è formata da un gruppo cromoforo, derivato dalla vitamina A, unito a una proteina, l'opsina, e ha uno spettro di assorbimento con massimo a circa 500 nm.
Uno dei primi eventi causati dalla luce è una isomerizzazione che porta l'aldeide della vitamina A, il retinale, dalla forma cis a quella trans. Si postula che questo sia l'evento critico che causa una variazione elettrica nella membrana del recettore. Le successive reazioni chimiche che avvengono nel pigmento portano al distacco del gruppo cromoforo dall'opsina. Nel recettore, poi, dai prodotti della scissione si riforma il pigmento iniziale, la rodopsina.
Per ragioni di spazio non sono qui trattati i dettagli della fotoscissione della rodopsina.
I pigmenti dei coni presentano tre spettri di assorbimento diversi e saranno trattati successivamente. La sensibilità dei fotorecettori è molto alta: un singolo ‛quanto' di luce è in grado di eccitare un bastoncello adattato allo scuro. Questa scoperta, ormai accertata con tecniche diverse, si deve a S. Hecht, S. Shlaer e M. H. Pirenne (1942), che, sulla base di esperimenti psicofisici, stabilirono per primi la sorprendente sensibilità della retina adattata allo scuro.
L'assorbimento di un singolo quanto di luce da parte di un bastoncello può eccitare solo una molecola di rodopsina, ma è molto improbabile che questo evento di per sé sia capace di generare i segnali bioelettrici del messaggio visivo. È verosimile che un processo di amplificazione segua l'azione della luce sul pigmento visivo e determini i segnali elettrici: la luce potrebbe semplicemente innescare un processo autocatalitico a livello delle reazioni fotochimiche, per esempio attivando qualche enzima.
La scissione del pigmento dei fotorecettori provoca in essi, con modalità ancora in gran parte oscure, l'insorgenza di un segnale elettrico atto a trasmettere l'informazione visiva alle altre cellule della retina.
Gli studi di elettrofisiologia hanno dimostrato che l'eccitamento di un recettore è usualmente legato a una depolarizzazione della membrana cellulare; la risposta dei recettori visivi dei Vertebrati alla luce è invece rappresentata da una iperpolarizzazione della membrana dei recettori stessi. Questo tipo di risposta elettrica dei recettori visivi è abbastanza raro: fra tutti i tipi di recettori solamente quelli olfattivi presentano un comportamento simile e si può affermare che, in genere, la risposta a uno stimolo da parte di un recettore sensoriale, compresi i recettori visivi degli Invertebrati, è la depolarizzazione della membrana (v. recettori). È chiaro che un fenomeno di iperpolarizzazione è particolarmente efficace per la trasmissione dell'informazione, se diminuisce un'attività tonica eccitatrice preesistente. Questo è proprio quello che si verifica nei recettori visivi dei Vertebrati: R. D. Penn e W. A. Hagins (1969) hanno infatti dimostrato che nel ratto, anche in condizioni di oscurità, esiste una corrente diretta dal segmento esterno a quello interno dei recettori, e che questa corrente viene diminuita dall'illuminazione.
Nei Vertebrati e negli Invertebrati esiste tra la cornea e il fondo dell'occhio una differenza di potenziale che si modifica quando l'occhio viene illuminato: il complesso di variazioni elettriche registrate prende il nome di elettroretinogramma (ERG). Esso può essere registrato facilmente anche nell'uomo ed è di notevole aiuto per stabilire la funzionalità della retina, specie in alcune forme di patologia dell'occhio. Si distinguono in esso quattro onde: a, b, c e d.
L'onda a rivela l'attività dei recettori; l'onda b sembra che si origini nello strato nucleare interno della retina e si è ritenuto per molto tempo che fosse indice dell'attività delle cellule bipolari. Recentemente, però, registrazioni intracellulari dalla retina di Necturus (v. Miller e Dowling, 1970) indurrebbero ad attribuire l'onda b dell'elettroretinogramma all' attività delle cellule gliali, cellule di supporto e di nutrizione interposte tra le altre cellule della retina. Anche se l'origine dell'onda b è incerta, è di fondamentale importanza il fatto che le variazioni della sua ampiezza sono nella maggior parte dei casi un indice fedele dell'andamento del processo visivo a livello retinico. L'onda c dell'elettroretinogramma è un'onda che si origina nell'epitelio pigmentato, membrana nutritizia che ricopre i recettori della retina: essa non dà nessuna informazione sul processo visivo. L'onda d segnala la fine dello stimolo luminoso e, quindi, un cambiamento di attività di tutte le sorgenti sopra ricordate. Per una descrizione dettagliata delle componenti dell'elettroretinogramma si rimanda al lavoro di K. T. Brown (v., 1968).
b) Neuroni retinici
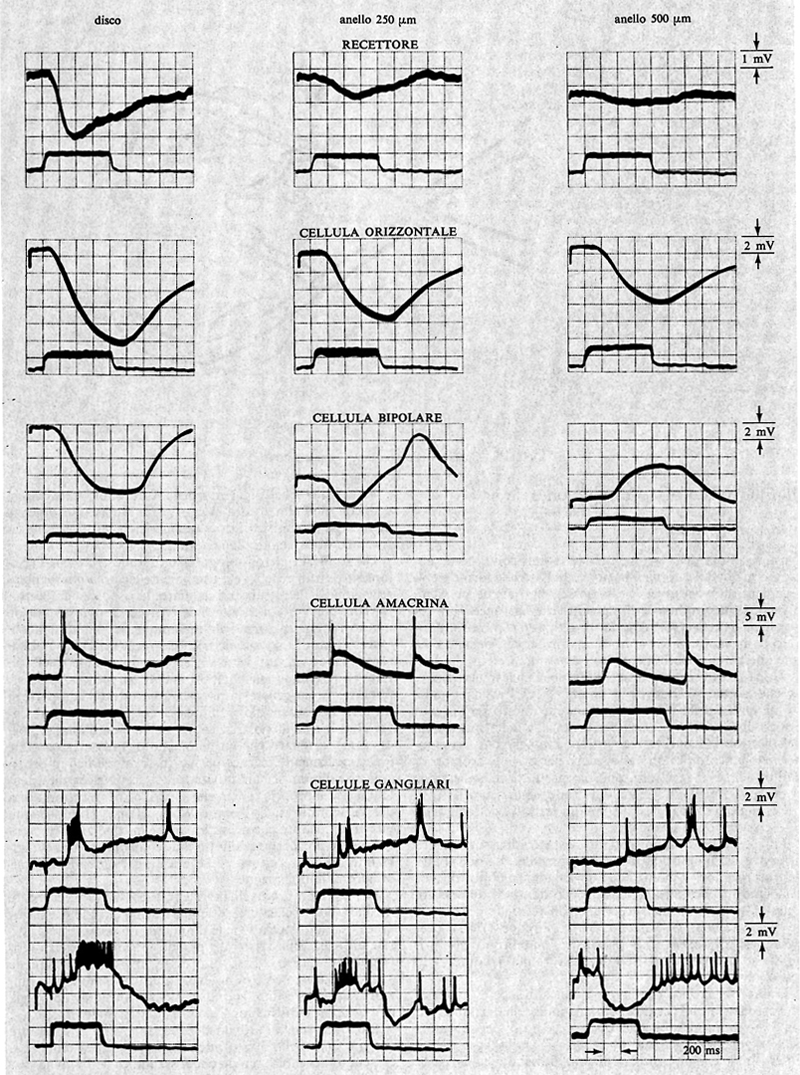
L'informazione visiva è codificata nei recettori così come nei primi strati della retina in variazioni graduate di potenziali; con un meccanismo analogico quindi, per prendere a prestito una parola del linguaggio della teoria delle comunicazioni. È solo a livello delle cellule gangliari che questa informazione viene codificata in impulsi nervosi d'ampiezza e forma costante (‛tutto - o - nulla'), cioè in maniera digitale. Esempi di registrazioni intracellulari da ogni tipo di cellula retinica sono riportati nella fig. 4. L'analisi ha confermato il quadro precedentemente descritto, ma ha messo in evidenza anche rari impulsi nervosi nelle cellule amacrine.
Le cellule gangliari dei Mammiferi presentano una scarica spontanea, anche nella più completa oscurità. Ciò corrisponderebbe al fatto che, anche nella più completa oscurità, non si ha una sensazione di nero, ma piuttosto di grigio. Hering chiamò questa sensazione Eigengrau, e già nel secolo scorso ipotizzò un'attività della retina in assenza del segnale luminoso. Si è discusso molto sull'origine e sulla funzione di questa scarica detta ‛scarica all'oscuro' o dark discharge. Oggi l'interpretazione più soddisfacente è quella secondo cui la scarica all'oscuro rappresenta la frequenza portante di un sistema a modulazione di frequenza. Lo stimolo luminoso, infatti, non fa che diminuire o aumentare la frequenza di questa scarica all'oscuro. Il parametro essenziale della scarica, modulato dallo stimolo luminoso, è verosimilmente rappresentato dalla probabilità che un impulso nervoso ha di avvenire o meno in un certo tempo.
La distribuzione temporale degli eventi nervosi (impulsi nervosi) in ogni singola cellula gangliare all'oscuro o in condizioni stazionarie di illuminazione è una distribuzione di tipo casuale e precisamente una distribuzione di tipo gamma. Recenti ricerche, basate su un modello matematico della retina, hanno messo in evidenza che una distribuzione di tipo casuale della scarica è ottimale per la trasmissione dell'informazione. Un'altra proprietà, che contribuisce a far sì che il sistema visivo abbia un'alta capacità di trasmissione dell'informazione, è data dal fatto che gli eventi nervosi delle diverse cellule gangliari sono indipendenti fra di loro. È noto dalla teoria delle comunicazioni che ogni legame fra i canali di un sistema aumenta la ridondanza del sistema stesso e diminuisce quindi la capacità di trasmissione dell'informazione.
c) Organizzazione del campo recettivo delle cellule gangliari retiniche
Le prime ricerche sulle cellule gangliari della retina dei Vertebrati sono dovute a H. K. Hartline (v., 1938) e a R. Granit (v., 1947). Dai loro esperimenti risulta che la retina si serve di due tipi di cellule gangliari per trasmettere l'informazione visiva: cellule che aumentano la loro scarica in risposta a uno stimolo luminoso, dette cellule on, e cellule che la diminuiscono, dette cellule off.
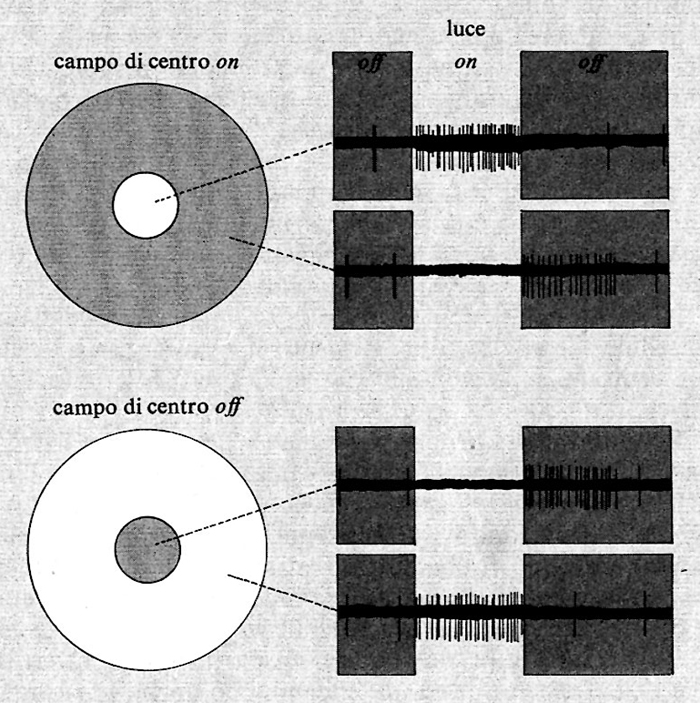
Una conoscenza più approfondita dell'elettrofisiologia delle cellule gangliari è derivata dai successivi esperimenti di S. W. Kuffler (v., 1953). Questi, nel 1952-1953, analizzò la risposta delle cellule gangliari del gatto a piccoli stimoli luminosi puntiformi ben localizzati sulla retina. Egli vide che ogni cellula gangliare risponde solo alla stimolazione luminosa di un'area ben localizzata del campo visivo, che può variare da alcuni minuti di arco ad alcuni gradi, definita da Hartline ‟campo recettivo" della cellula. Kuffler si accorse che la cellula non rispondeva uniformemente allo stimolo luminoso su tutto il campo recettivo. Questo poteva essere diviso in due zone concentriche, a risposta praticamente opposta. Kuffler distinse due classi di cellule, le cellule a centro on e le cellule a centro off. In entrambe le classi la parte centrale del campo recettivo presenta una sensibilità allo stimolo luminoso maggiore che non la parte periferica. Nelle cellule on l'illuminazione della parte centrale del campo recettivo provoca una risposta eccitatoria, e cioè un aumento della scarica della cellula, mentre l'illuminazione della parte circostante del campo recettivo provoca una risposta inibitoria. Le cellule off sono invece organizzate all'opposto, con una parte centrale del campo recettivo la cui illuminazione provoca una risposta inibitoria, mentre l'illuminazione della parte periferica provoca una risposta eccitatoria. Su entrambi i tipi di cellule una diminuzione del livello di illuminazione dà risposte opposte a quelle descritte. Esempi delle risposte delle cellule gangliari del gatto sono riportati nella fig. 5.
Se il campo recettivo di una cellula gangliare, per esempio di tipo a centro on, è illuminato uniformemente sia nella parte centrale eccitatoria sia nella parte periferica inibitoria - la risposta della cellula è nella maggioranza dei casi ancora di tipo eccitatorio; la frequenza di scarica è però molto minore di quella che si ha quando viene stimolata solo la parte centrale del campo recettivo. Ciò dimostra che le attività evocate dalla stimolazione delle parti eccitatoria e inibitoria del campo recettivo influenzano in senso opposto la cellula gangliare. Ricerche quantitative hanno messo in evidenza che questa interreazione è in prima approssimazione lineare: si ha cioè una vera e propria somma algebrica. Di conseguenza, una cellula gangliare a centro on risponderà al massimo delle sue possibilità quando la parte centrale del campo recettivo è illuminata e la parte periferica oscurata. Recenti ricerche hanno dimostrato che anche le cellule bipolari presentano un'organizzazione in zone antagoniste del loro campo recettivo, simile a quella che abbiamo descritto per le cellule gangliari. Sulla base di questi risultati fu prospettata l'ipotesi che le cellule on servissero principalmente per vedere oggetti luminosi in campo oscuro, e le cellule off per vedere oggetti scuri in campo luminoso. Questa teoria resta però al livello di semplice ipotesi, poiché, come risulta dall'esposizione precedente, ogni tipo di cellula può rispondere ugualmente bene sia alla diminuzione sia all'aumento della luce, e non abbiamo per ora nessuna informazione sul modo in cui gli aumenti o le diminuzioni della scarica sono interpretati a livello centrale. Resta però il fatto che l'organizzazione delle cellule gangliari sembra ottimale per la trasmissione dell'informazione chiaro su scuro o viceversa. Il contrasto visivo, che può essere definito come il rapporto tra le luminanze della zona più illuminata e della zona più in ombra di un oggetto, potrebbe trovare la sua base fisiologica in questa organizzazione antagonista dei campi recettivi.
I campi recettivi delle cellule gangliari variano grandemente nelle loro dimensioni. Per la regione foveale della scimmia, per esempio, il centro del campo recettivo può scendere addirittura alle grandezze di un cono, circa 20″ di arco; mentre nella periferia si possono trovare, nello stesso animale, centri di campi recettivi dell'ordine di 3-4°.
Si può arguire che i campi recettivi piccoli nella regione foveale diano un campionamento più accurato dell'oggetto, mentre i grossi campi recettivi potrebbero servire come meccanismi atti a raccogliere una maggiore quantità di energia luminosa nella visione crepuscolare. Anche se i campi recettivi più piccoli sono nella regione foveale e i campi più grandi in quella periferica, risulta però che ogni regione della retina ha campi recettivi di dimensioni molto diverse.
Nei Vertebrati inferiori, come Anfibi e Uccelli, e anche in alcuni Roditori, come il coniglio, le cellule gangliari retiniche presentano risposte più varie e complesse. È regola generale, del resto, che man mano che si scende nella scala dei Vertebrati la retina diventi anatomicamente più complicata, e le risposte presentino specificità per particolari tipi di stimoli, come sbarre di varie dimensioni, ecc. La caratteristica principale è che nella retina di questi animali vi è una classe di cellule che risponde solo a oggetti in movimento e spesso con specificità direzionale: la cellula dà una risposta allo stimolo se l'oggetto si muove in una determinata direzione e non in un'altra.
Nei Mammiferi, come il gatto e la scimmia, questa classe di cellule che analizza il movimento si ritrova solo a livello più centrale, come nel collicolo superiore o nella corteccia visiva.
Lo studio delle proprietà di analisi del movimento o di particolari caratteristiche dello stimolo da parte di cellule delle stazioni visive è stato iniziato da J. Y. Lettvin e altri (v., 1959). Studiando le risposte delle cellule gangliari della rana, questi autori ebbero la geniale idea di usare stimoli visivi che simulassero oggetti o animali dell'habitat naturale della rana. Trovarono così cellule retiniche che rispondono solo a oggetti in movimento o a particolari caratteristiche della loro forma. È da ritenere che anche le ricerche sulle cellule della corteccia visiva dei Mammiferi, delle quali tra breve tratteremo, si siano ispirate a questi esperimenti fondamentali.
Negli anni più recenti si è cercato di classificare le cellule gangliari rispetto ad altre proprietà funzionali. Esse sono state divise nelle classi X, Y e W.
Cellule X. - Rappresentano circa il 40% delle cellule gangliari (nel gatto). Sono presenti in tutte le parti della retina, ma principalmente nelle aree di maggiore acuità visiva, come nell'area centrale corrispondente alla fovea dell'uomo. Le loro fibre hanno velocità di conduzione media tra 10 e 20 m/s. Hanno proprietà lineari di integrazione nello spazio e rispondono in maniera tonica a uno stimolo stazionario (cellule sostenute) che illumina il centro del loro campo recettivo.
Cellule Y. - Rappresentano circa il 5-6% delle cellule gangliari. Le loro fibre hanno più alta velocità di conduzione (30-40 m/s) e rispondono in maniera fasica a stimoli stazionari (cellule transienti).
Cellule W. - Rappresentano più della metà delle cellule gangliari. Sono sparse dappertutto, ma sono maggiormente concentrate nella periferia della retina. Hanno bassa velocità di conduzione (5-10 m/s) e proprietà peculiari di risposta a stimolazioni del loro campo recettivo. Per esempio, ve ne sono alcune che rispondono solo a stimoli in movimento, o presentano proprietà on-off su tutto il loro campo recettivo, ecc.
Le fibre delle cellule X si dirigono al corpo genicolato laterale, dove prendono contatto con cellule che hanno proprietà funzionali simili a quelle X retiniche. Le cellule Y si dirigono al genicolato, ma soprattutto al collicolo superiore. Le cellule W si dirigono, nel gatto, agli strati profondi del genicolato e al collicolo superiore. Le cellule del genicolato X e Y proiettano all'area visiva striata primaria, le cellule Y e W alle altre aree (v. Stone e Fukuda, 1974). Il ruolo delle cellule W nel processo della visione non è ancora del tutto chiaro. Si suppone che tutta la linea delle cellule X, detta sistema X, abbia un ruolo importante nell'analisi delle forme, mentre per le cellule Y questo ruolo sarebbe qualitativamente secondario. Il loro ruolo predominante consisterebbe nell'analizzare le proprietà temporali dello stimolo visivo e quindi, probabilmente, le immagini in movimento.
d) Corpo genicolato laterale
Abbiamo visto dall'anatomia che gran parte delle fibre del nervo ottico prendono contatto con le cellule del corpo genicolato laterale. Le varie terminazioni di una singola fibra prendono contatto con diverse cellule, e d'altronde terminazioni di varie fibre ottiche, da 20 a 100, prendono contatto con la stessa cellula del corpo genicolato. Si ha quindi, a livello del genicolato, un processo di convergenza e di divergenza dell'informazione visiva. Questo processo sembra però avvenire all'interno dei gruppi delle cellule X, Y e W separatamente, cioè non ci sarebbero convergenze crociate tra i diversi tipi di cellule. L'organizzazione dei campi recettivi a livello del genicolato è molto simile a quella retinica: si trovano ancora due classi di cellule a centro on e a centro off.
Una differenza importante rispetto ai campi recettivi retinici è costituita dal fatto che la periferia del campo recettivo ha un'azione antagonista più potente sull'attività del centro, tantoché la cellula risponde scarsamente o non risponde affatto a un'illuminazione diffusa di tutto il suo campo recettivo.
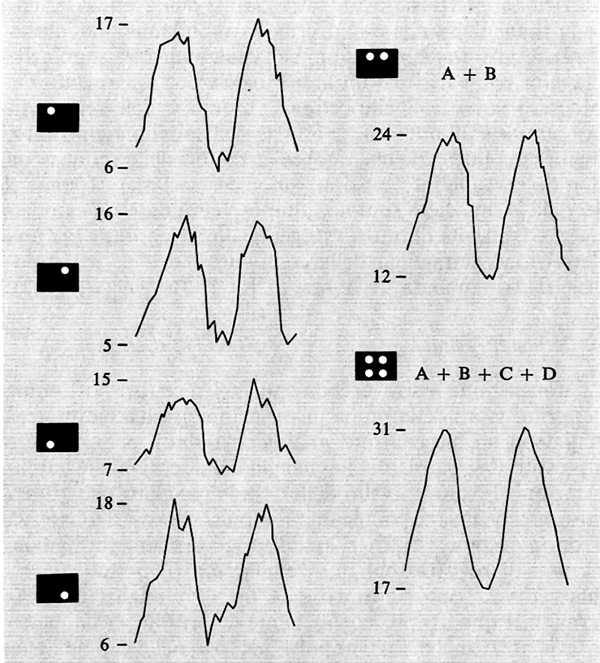
La somiglianza fra l'organizzazione del genicolato e quella della retina ha fatto credere per un certo tempo che il corpo genicolato fosse una stazione passiva non essenziale nell'elaborazione o nella trasmissione dell'informazione visiva. Man mano che gli esperimenti proseguono, questa concezione si rivela sempre più errata. Le cellule del corpo genicolato laterale presentano una scarica spontanea che è circa la metà di quella delle cellule retiniche. Ora, se il sistema, come abbiamo supposto, trasmette l'informazione tramite la modulazione della frequenza di scarica delle cellule, ciò significa una diminuzione della capacità del sistema di trasmettere l'informazione, a condizione che il numero dei canali non cambi rispetto al numero dei canali retinici. Poiché quest' ultima condizione è, in prima approssimazione, rispettata nella maggior parte dei Mammiferi, ne segue che al livello del genicolato avviene probabilmente una compressione dell'informazione con scarto di messaggi o di parti di messaggi ritenuti ridondanti. Una compressione dell'informazione, che peraltro significa sempre una sua riduzione, può avvenire in varia maniera: per esempio tramite un'integrazione temporale e spaziale dell'informazione retinica. È dimostrato che entrambi questi processi sono presenti a livello del genicolato. L'integrazione spaziale, cioè la convergenza di più fibre retiniche su una cellula del genicolato, può essere di fondamentale importanza nella trasmissione del segnale visivo. Infatti, se più canali che trasmettono in parallelo l'informazione dello stesso oggetto convergono su una singola cellula, questa viene a fare, praticamente, una media dell'informazione dei vari canali, con una diminuzione del rumore di fondo, e quindi un miglioramento del rapporto segnale-rumore, che sarà tanto maggiore quanto maggiore sarà il numero dei canali che convergono sulla cellula. Che questo meccanismo di riduzione del rumore di fondo operi attivamente a livello del genicolato è stato dimostrato paragonando le risposte a una luce modulata sinusoidalmente di una o più porzioni del centro del campo recettivo di una cellula del genicolato, controllando opportunamente l'effetto dell'aumento dell'energia luminosa. I risultati di questo esperimento sono illustrati nella fig. 6. La risposta alla luce modulata appare molto meno disturbata quando più stimoli luminosi sono usati simultaneamente.

Recenti studi comparativi dei campi recettivi retinici e di quelli del genicolato hanno potuto mettere in evidenza che non vi è corrispondenza spaziale tra la periferia dei campi recettivi retinici e quella dei campi recettivi del genicolato. Un esempio di collegamento è dato dallo schema della fig. 7. Nello schema il campo recettivo d'una cellula a centro on del genicolato corrisponde a quello d'una cellula a centro on retinica per la sola parte centrale; alla periferia esso corrisponde, per esempio, ai centri di quattro cellule retiniche a centro off. Teoricamente la periferia di cellule on può essere formata anche da altre cellule on, facendo l'ipotesi che queste subiscano un inversione di segno al livello del genicolato, per esempio tramite un interneurone; la stessa cosa può essere ovviamente detta per le cellule off del genicolato. Il genicolato, oltre a esercitare questo delicato gioco di rielaborazione dell'informazione visiva, ne esercita uno ben più appariscente di controllo su di essa, che dipende dalla situazione di veglia o di sonno dell'animale. Il ritmo veglia-sonno, com'è noto, è principalmente regolato dalla sostanza reticolare del tronco dell'encefalo (v. sonno). Le ricerche che dimostrano la variazione delle risposte del genicolato dalla veglia al sonno sono numerose sia nei classici preparati da esperimento, sia negli animali intatti. Verranno riportate solo alcune ricerche che appaiono sufficientemente illustrative.
Durante la veglia, la frequenza di scarica delle cellule retiniche, così come quella delle cellule del genicolato, è modulata in maniera quasi perfetta da uno stimolo luminoso la cui intensità varia sinusoidalmente. Almeno nel gatto, durante la fase di sonno cosiddetto ‛sincronizzato', la scarica retinica continua a essere modulata dallo stimolo come durante la veglia, mentre la scarica delle cellule del genicolato diventa irregolare e riproduce scarsamente o non riproduce affatto la modulazione dello stimolo luminoso.
Anche la sensibilità della risposta delle cellule del genicolato a lampi di luce diminuisce enormemente durante il sonno rispetto alla veglia. Sembra proprio che durante il sonno la sostanza reticolare cerchi di bloccare, o almeno di attenuare, il passaggio dell'informazione visiva attraverso il genicolato, quasi a impedire l'analisi da parte dei centri superiori di un'informazione inutile o di scarsa importanza per la vita dell'animale.
e) Corteccia visiva
Negli ultimi venti anni sono stati fatti notevoli progressi nello studio elettrofisiologico della corteccia visiva, specie per merito di due fisiologi, D. H. Hubel e T. N. Wiesel, che per questi loro studi hanno ricevuto nel 1981 il premio Nobel. Le loro ricerche riguardano principalmente la corteccia del gatto e della scimmia.
Le cellule della corteccia visiva mostrano proprietà più complesse delle altre stazioni visive. Hubel e Wiesel (v., 1965 e 1968) le hanno divise schematicamente in tre gruppi: cellule semplici, complesse e ipercomplesse. Le cellule semplici presentano un'ulteriore suddivisione del campo recettivo in regioni eccitatorie e inibitorie. In alcuni casi, ad esempio nello strato corticale di arrivo delle scimmie, il campo recettivo presenta anche una struttura concentrica, come i campi recettivi del genicolato e della retina, ma negli altri strati corticali (in tutti quelli descritti da Hubel e Wiesel) esso ha una forma allungata: una striscia eccitatoria fiancheggiata da un lato o da entrambi da strisce inibitorie, o viceversa, cioè una striscia inibitoria fiancheggiata da strisce eccitatorie. Queste strisce eccitatorie e inibitorie del campo recettivo delle cellule semplici possono mostrare tutte le inclinazioni. Questa morfologia particolare dei campi recettivi favorisce la risposta a stimoli ottici costituiti da sbarre luminose su fondo scuro o sbarre scure su fondo luminoso, e la risposta è inoltre strettamente legata all'orientamento della sbarra. Per esempio, una cellula che mostra una risposta ottimale per una luce o sbarra verticale non risponde affatto se la sbarra è inclinata di 30° o anche di soli 20° dalla verticale.

Anche le cellule complesse, come le cellule semplici, rispondono di preferenza a sbarre o linee luminose di un determinato orientamento. Esse però non presentano una chiara distinzione tra regioni eccitatorie e inibitorie del loro campo recettivo e rispondono di preferenza a stimoli luminosi in movimento in una determinata direzione. La cellula dà una risposta che dura per tutto il tempo in cui lo stimolo in movimento si trova nel suo campo recettivo. Un esempio del comportamento delle cellule complesse è riportato nella fig. 11.
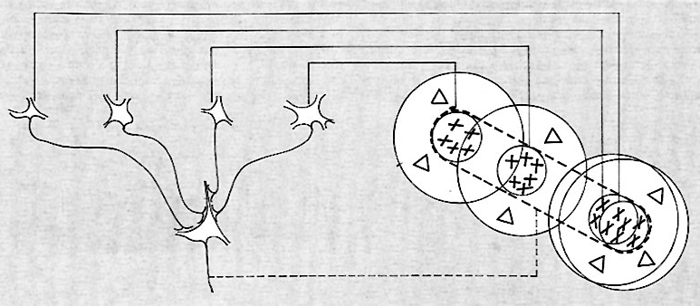
Le cellule ipercomplesse si trovano particolarmente nelle aree associative 18 e 19, ma anche negli strati superficiali e profondi dell'area 17. Anche questa classe di cellule, come le due precedenti, presenta una chiara specificità per l'orientamento dello stimolo luminoso. Queste cellule rispondono solo a stimoli luminosi in movimento e di regola rispondono di preferenza a una direzione del movimento. Il loro campo recettivo è, nella maggior parte dei casi, costituito da una zona di eccitazione fiancheggiata da zone antagoniste. Sia la zona di eccitazione che quella di inibizione presentano la stessa preferenza per l'orientamento dello stimolo. La principale caratteristica delle cellule ipercomplesse è che esse non hanno soltanto una specificità di orientamento: per avere la risposta ottimale, infatti, lo stimolo luminoso, oltre all'orientamento, deve avere anche dimensioni e spesso forma particolari. L'unica proprietà invariante nelle tre classi di cellule è dunque la loro specificità all'orientamento dello stimolo. Questa caratteristica ha anche un correlato anatomico. Infatti, risultati elettrofisiologici dimostrano che la corteccia è suddivisa funzionalmente in tante piccole colonne che si estendono dalla superficie della corteccia alla sostanza bianca (v. Hubel e Wiesel, 1963). La colonna è definita dal fatto che tutte le centinaia di cellule che contiene - semplici, complesse e ipercomplesse - hanno la stessa specificità di orientamento. Le colonne presentano una sezione trasversa irregolare, che è dell'ordine del centinaio di μm. Dalle cellule del genicolato a quelle della corteccia, cellule semplici, complesse, ipercomplesse, le proprietà di risposta allo stimolo luminoso diventano gradualmente più complicate. Hubel e Wiesel (v., 1962), sulla base dei loro risultati, hanno proposto dei modelli di connessioni fra le cellule, che spiegano come le proprietà di una certa classe di cellule non derivino che dalla convergenza organizzata di cellule della classe più semplice. Il diagramma della fig. 13 spiega, per esempio, come la proiezione di un gruppo di cellule del genicolato potrebbe dar luogo alla particolare distribuzione dell'eccitazione e dell'inibizione in una cellula semplice. La parte centrale, supponiamo eccitatoria, sarebbe formata dalla proiezione dei centri dei campi recettivi di un gruppo di cellule on del genicolato. Questi centri sarebbero tutti disposti lungo una striscia allungata, delimitata in figura dal tratteggio; i fianchi inibitori sarebbero costituiti dalle periferie degli stessi campi recettivi. Il modello schematizzato nella fig. 13 suggerisce che la grandezza della regione centrale delle cellule semplici dovrebbe essere dello stesso ordine del diametro dei centri delle cellule del genicolato. Tale fatto, nell'esperienza di Hubel e Wiesel, sembra essere vero, almeno in prima approssimazione. Alcune delle proprietà delle cellule complesse possono, con uguale semplicità, essere spiegate dalla convergenza organizzata di cellule semplici, come illustrato nella fig. 14. Si è schematizzata qui una cellula complessa che riceve afferenze di cellule semplici disposte su una linea orizzontale, che posseggono tutte una specificità per la direzione verticale. È chiaro che nel caso in cui una striscia luminosa verticale eccitasse le cellule semplici, poste nella situazione indicata dallo schema, la cellula complessa verrebbe a essere continuamente stimolata e risponderebbe al movimento della sbarra su tutto il suo campo recettivo: è questa una sua proprietà caratteristica, che abbiamo già visto.
Con schemi simili si possono ricostruire anche le proprietà delle cellule ipercomplesse da quelle del grado immediatamente inferiore, le cellule complesse.
Questi modelli di Hubel e Wiesel hanno solo un valore di ipotesi: essi non considerano, per esempio, l'informazione proveniente dalla via colliculotalamocorticale che, almeno nel gatto, è anatomicamente ben definita e termina nelle aree associative, principalmente nell'area 18. Le cellule colliculari presentano già risposte estremamente complesse, simili a quelle delle cellule corticali delle aree associative. Non è perciò da escludere che, almeno nel gatto, la preelaborazione dell'informazione visiva avvenga in parte a livello colliculare o del pulvinar. Sotto questo aspetto i modelli proposti da Hubel e Wiesel si presentano, se non altro, incompleti.
Nello studio delle varie stazioni della via visiva è stato facile notare le proprietà di analisi del sistema. Nella retina l'immagine viene finemente scomposta dal mosaico dei recettori e dei campi recettivi. Nel corpo genicolato l'immagine subisce una prima preelaborazione che cerca di mettere in evidenza l'oggetto rispetto allo sfondo, i suoi contorni, le differenze di contrasto. Nelle cellule semplici della corteccia è l'orientamento dello stimolo che viene analizzato. Nelle cellule complesse e ipercomplesse vengono probabilmente analizzati altri attributi dell'oggetto, come il movimento e forse la dimensione e la forma di certe parti dell'oggetto. Nella cellula corticale, infine, avviene una elaborazione di sintesi: qui si comincia a ricostruire l'immagine dell'oggetto nella sua interezza.
Cerchiamo di seguire il lavoro delle cellule corticali servendoci di un esempio. Supponiamo di avere davanti a noi una figura geometrica, per esempio un triangolo. Nella corteccia, le prime a entrare in gioco sarebbero le cellule semplici, con delimitazione dei lati e della loro inclinazione. L'informazione poi, secondo Hubel e Wiesel, passerebbe alle cellule complesse che, mantenendo la stessa risposta per tutta l'estensione del loro campo recettivo, rendono largamente indipendente l'attributo orientamento dai movimenti oculari; si passerebbe quindi alle cellule ipercomplesse, dove il tipo di angolo potrebbe venire astratto e catalogato, dando inizio a una parziale ricostruzione dell'immagine.
È chiaro, dal discorso che abbiamo fatto, che le nostre conoscenze a livello della sintesi dell'immagine sono del tutto insufficienti. Ancora non si è potuta trovare una classe di cellule dove l'intera immagine venga astratta e riconosciuta. Alcuni ricercatori stanno analizzando altre zone di corteccia che sembrano pure coinvolte nell'analisi del segnale visivo, come la corteccia infratemporale. Per ora non si hanno risultati incoraggianti. D'altronde potrebbe essere del tutto banale la ricerca di questo elemento finale di sintesi in una singola cellula. Esso potrebbe essere rappresentato, come suggerito da Hebb, da una infinità di connessioni tra le cellule che vengono modificate sulla base di precise influenze del mondo esterno esercitate sulle sinapsi ancora plastiche, nella nostra prima infanzia.
f) Architettura funzionale della corteccia visiva: colonne e ipercolonne
Tra le proprietà funzionali più importanti dei neuroni dell'area visiva primaria del gatto e della scimmia sono da annoverare le loro specificità per l'orientamento e per le dimensioni dello stimolo visivo. Altra caratteristica è che questi neuroni rispondono nella maggior parte alla stimolazione dei due occhi separatamente.
Riconfluisce così in analizzatori unici l'informazione visiva che fino al corpo genicolato laterale era stata trasmessa in canali separati. Rispetto a queste proprietà principali i neuroni corticali si raggruppano anatomicamente in maniera ordinata. Abbiamo già detto che i neuroni, che l'elettrodo incontra in una penetrazione perpendicolare alla superficie della corteccia, sono caratterizzati dal fatto di avere il loro campo recettivo nella stessa regione dello spazio e di avere lo stesso orientamento preferenziale per l'immagine visiva. Questo raggruppamento di neuroni viene chiamato colonna e va dalla superficie della corteccia alla sostanza bianca, una distanza di circa 2 mm. Hubel e Wiesel (v., 1977) con penetrazioni dell'elettrodo parallele alla superficie della corteccia hanno dimostrato altre caratteristiche di organizzazione anatomo-funzionale.
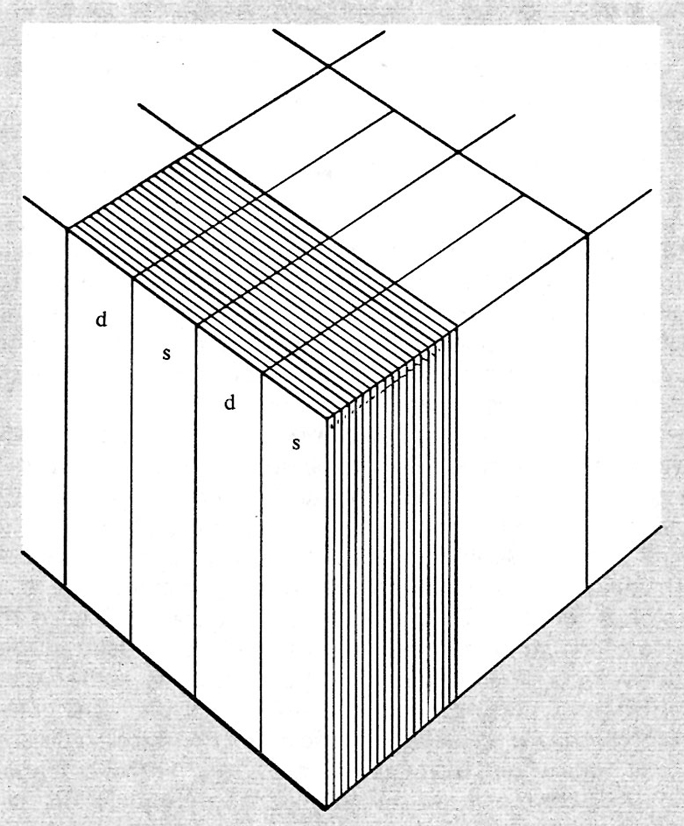
L'orientamento preferenziale dei campi recettivi delle cellule in queste penetrazioni cosiddette ‛tangenziali' varia ordinatamente da cellula a cellula, con salti discreti di 10-15°; raggiunge variazioni fino a 180° per un avanzamento dell'elettrodo di circa 1 mm. In questo spazio anatomico si trova un'altra organizzazione che si riferisce alla dominanza oculare. I neuroni della corteccia, infatti, o sono monoculari e rispondono quindi solo a un determinato occhio (questi neuroni si trovano principalmente in uno strato della corteccia chiamato IV c) o, pur essendo binoculari, rispondono più vigorosamente a uno dei due occhi. Si è trovato che le cellule che rispondono principalmente a un occhio sono raggruppate in colonne, dette ‛colonne della dominanza oculare'. Queste colonne, impropriamente chiamate così dato che la loro sezione trasversale non è circolare, hanno il lato maggiore lungo circa 500 μm. In penetrazioni tangenziali è stato trovato che le colonne della dominanza oculare dell'occhio destro e del sinistro si alternano regolarmente. In una penetrazione di 1 mm, le cellule variano il loro orientamento di circa 180° e si dispongono in due colonne di dominanza oculare, una per l'occhio destro e una per il sinistro. Hubel e Wiesel hanno chiamato ipercolonna questo parallelepipedo di tessuto corticale, nel quale sono rappresentati tutti i possibili orientamenti, e nel quale sono presenti due colonne di dominanza oculare, una per l'occhio destro e una per il sinistro. L'ipercolonna è quindi un raggruppamento di cellule che unisce i due campi visivi e possiede le proprietà per analizzare oggetti di tutti gli orientamenti in una determinata zona del campo visivo. La fig. 15 riassume schematicamente alcune delle proprietà anatomofunzionali della corteccia visiva.
Importanti informazioni sulle funzioni della corteccia visiva o di altre aree corticali, aventi influenza sulla visione, ci provengono da esperimenti di lesione o ablazione della corteccia stessa. Lesioni della corteccia occipitale dell'uomo, in seguito a operazioni chirurgiche o a ferite, portano a cecità permanente; nella maggior parte dei casi resta la possibilità di distinguere l'improvviso abbuiamento o illuminamento dell'ambiente. Osservazioni recenti nell'uomo (v. Weiskrantz e altri, 1974) dimostrano che l'ablazione della corteccia striata lascia un certo tipo di funzione sensoriale visiva che potremmo definire inconscia. Pazienti con grosse lesioni patologiche o chirurgiche della corteccia visiva presentano corrispondenti scotomi, cioè aree del campo visivo per le quali essi sono completamente ciechi. Tuttavia, se si presentano piccoli lampi di luce in queste aree del campo visivo, essi, pur affermando di non vedere nulla, sono capaci di indicare con la mano con buona approssimazione il punto da cui provengono i lampi. Essi sono anche capaci di congetturare con sorprendente precisione se le linee luminose, lampeggiate nello scotoma, sono verticali od orizzontali, sempre negando di vedere alcunché.
Probabilmente, risposte motorie di questo tipo sono legate ad aree visive sottocorticali, deputate a fornire informazioni utili per i movimenti delle mani o degli occhi. Evidentemente la corteccia potrebbe essere il centro della visione conscia, senza escludere però che certi comportamenti motori innescati dalla visione possano avere luogo senza di essa.
Anche in altri animali, come gatti e scimmie, lesioni dell'area visiva striata non producono abolizione delle funzioni visive, ma piuttosto disturbi visivi ai quali segue un buon recupero (per le scimmie, v. Humphrey e Weiskrantz, 1967).
Certamente la corteccia striata 17 e anche le aree associative 18 e 19 non sono le sole aventi funzioni visive e il loro contributo al processo visivo è tuttora oscuro. Basta del resto ricordare gli esperimenti di Sperry e altri (v., 1955), i quali trovarono che nel gatto numerose incisioni della corteccia 17, 18, 19 non disturbavano sostanzialmente la discriminazione visiva dell'animale. Fra le diverse interpretazioni di questo esperimento, una plausibile è che il processo conclusivo di ricostruzione dell'immagine visiva avvenga anche in aree diverse dalla 17, 18, 19. Numerosi sono gli esempi che suggeriscono che anche altre parti della corteccia, non propriamente visiva, siano correlate col processo della visione. Basta ricordare, per esempio, i classici esperimenti di ablazione dei lobi temporali nelle scimmie di Klüver e Bucy (v., 1939).
Queste lesioni producono una particolare sindrome comportamentale, nella quale i soggetti possono vedere gli oggetti, ma non riconoscerli. Klüver e Bucy chiamarono questa sindrome cecità psichica. Questi esperimenti sono stati recentemente confermati da altri autori che hanno messo in evidenza ulteriori dettagli del contributo del lobo temporale al processo visivo.
Anche la lesione dei lobi parietali porta nell'uomo a una certa incapacità di far uso dell'informazione visiva, come l'incapacità di copiare disegni od organizzare oggetti (‛aprassia costruzionale'), o di ritrovare luoghi e vie prima familiari (‛disorientamento spaziale').
4. Psicofisica della visione
La psicofisica della visione studia la percezione degli stimoli luminosi nell'uomo. Questo metodo di indagine è stato ed è tuttora fecondissimo di risultati. I grandi psicofisici del secolo scorso come Helmholtz, Hering, Mach, per nominare i più famosi, hanno fatto fare allo studio della visione notevoli, fondamentali progressi. Talvolta questi autori sono stati capaci di predire risultati di esperimenti che solo le tecniche dei tempi più moderni hanno potuto verificare. È proprio il caso di dire che questi grandi ricercatori usarono l'acume dell'ingegno nella soluzione di problemi oggi troppo spesso affidati all'elaborazione tecnica.
Gli esperimenti di psicofisica possono essere divisi in due classi (v. Brindley, 1970): quantitativi e a indirizzo più qualitativo. Fanno parte della prima classe gli esperimenti nei quali vengono effettuate precise misurazioni delle variabili fisiche in questione, come misura delle soglie assolute o incrementali della percezione della luce, uguaglianza della brillanza di due immagini. Nella seconda classe si considerano gli esperimenti nei quali al soggetto viene richiesto di valutare la qualità o l'intensità delle sue sensazioni.
Gli esperimenti che riportiamo appartengono principalmente alla prima classe.
a) Adattamento visivo
Questo fenomeno comprende l'adattamento alla luce e l'adattamento all'oscurità.
L'adattamento alla luce è rapido ed è certamente una caratteristica di funzionalità positiva del nostro sistema visivo. Esso permette di mantenere costanti o quasi costanti molti aspetti della percezione, anche se la reale intensità di illuminazione della retina varia di migliaia di volte. Questa costanza della percezione si verifica particolarmente per il contrasto relativo ad aree del campo visivo più o meno luminose. Le aree del campo visivo aventi luminanza massima (quelle che normalmente ci appaiono bianche) conservano praticamente lo stesso aspetto, sia a illuminazioni solari molto intense, sia a illuminazioni relativamente deboli, purché l'occhio sia adattato al rispettivo livello di illuminazione. Così pure si mantiene più o meno invariato l'aspetto di oggetti più scuri, grigi o neri. È questo il fenomeno della costanza della brillanza che è la valutazione soggettiva di una certa luminanza.
Mediante il processo di adattamento alla luce, quindi, la brillanza di un oggetto è determinata non dal valore assoluto dell'energia luminosa che esso riflette, bensì dal rapporto tra questa e il livello luminoso medio dell'ambiente. Nella via di trasmissione dell'informazione visiva c'è quindi un sistema di controllo automatico dell'ampiezza della risposta alla luce. La sensibilità del sistema viene automaticamente ridotta al crescere del livello medio dell'illuminazione della retina, in modo da mantenere costante la risposta a stimoli che differiscono dal livello medio per una quantità percentualmente costante.
L'adattamento all'oscurità è, invece, come diceva Rushton (v., 1965), una limitazione funzionale del sistema visivo, che tutti, del resto, abbiamo giornalmente occasione di sperimentare passando da ambienti ben illuminati ad altri più oscuri. Da principio si resta completamente privi dell'uso della vista, che viene poi recuperata lentamente, appunto con l'adattamento allo scuro. L'adattamento allo scuro consiste, quindi, in un miglioramento della sensibilità della retina, al quale contribuisce anche una dilatazione della pupilla, che avviene parallelamente all'adattamento allo scuro.
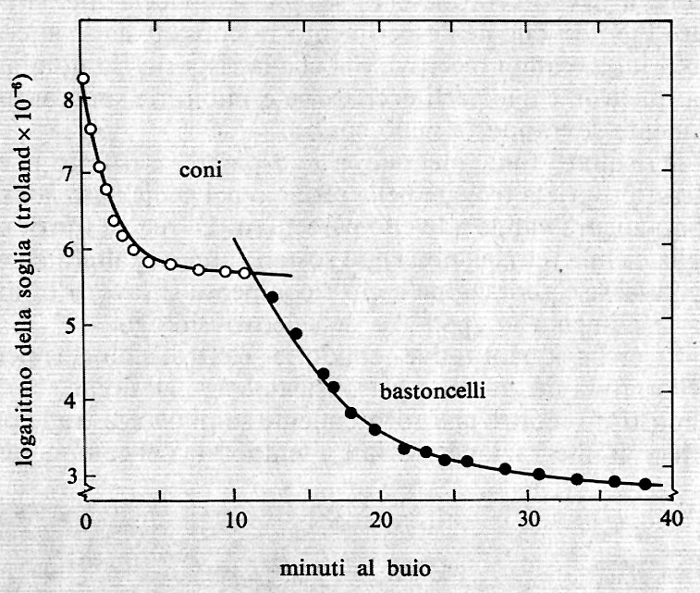
Nella fig. 16 è riportata una curva tipica di adattamento allo scuro determinata mediante la misura della minima quantità di luce visibile nell'oscurità (soglia) a vari intervalli di tempo dopo esposizione a una luce molto intensa. Sulle ordinate sono riportati i logaritmi della minima quantità di luce visibile ai diversi tempi di adattamento che sono riportati sulle ascisse. Si osservi che, in circa 30 minuti di adattamento allo scuro, il minimo della quantità di luce chiesta per essere vista diminuisce di circa 100.000 volte. La curva di adattamento allo scuro è composta di due rami. Il ramo superiore è dovuto ai coni, perché è presente anche quando la luce test viene mandata direttamente sulla fovea, che non contiene bastoncelli e inoltre ha la sensibilità spettrale della visione fotopica (coni). Il ramo più basso è dovuto ai bastoncelli, perché è assente quando la luce test viene mandata sulla fovea e la sua dipendenza dalla componente spettrale è simile a quella della visione scotopica e dell'assorbimento della rodopsina.
Secondo le vecchie teorie, sorprendentemente riportate anche in alcuni dei testi più moderni, il guadagno di sensibilità alla luce durante l'adattamento allo scuro era dovuto soltanto alla progressiva ricomposizione della rodopsina dai suoi prodotti di scissione. Si credeva, inoltre, che vi fosse una proporzionalità diretta tra la quantità di rodopsina presente nei recettori e la sensibilità alla luce. Il fenomeno, a uno studio più accurato, si è rivelato assai più complesso. Rushton (v., 1962) ha infatti potuto misurare la quantità di rodopsina nella retina dell'uomo con una tecnica basata sul principio dell'oftalmoscopio: viene misurata, cioè, la quantità di luce riflessa dalla retina illuminata da una determinata sorgente, che sarà tanto minore quanto maggiore è la quantità di luce assorbita dal pigmento, restando costanti le altre cause di diffusione e assorbimento della luce nell'occhio. Con questa tecnica il Rushton ha potuto dimostrare che ai normali adattamenti alla luce, a cui siamo esposti giornalmente, la quantità di rodopsina scissa nei nostri recettori è minima, anche se questi stessi adattamenti sono capaci di diminuire di migliaia di volte la nostra sensibilità alla luce.
Queste ricerche e altre di natura elettrofisiologica hanno dimostrato che la soglia non è determinata a livello dei recettori visivi, almeno fino a che non sia raggiunto il completo adattamento all'oscurità, bensì a livello del punto in cui i messaggi dei recettori convergono sui neuroni della retina. Il luogo di convergenza, detto da Rushton summation pool, è stato individuato con ricerche elettrofisiologiche a livello delle cellule bipolari. Risulta così che la soglia visiva durante l'adattamento allo scuro è funzione non solo di fattori fotochimici (quantità di rodopsina presente), ma anche di fattori nervosi. Nei casi di esposizione a luce particolarmente intensa, l'adattamento allo scuro che segue mostra che non vi è affatto una proporzionalità tra soglia visiva e quantità di rodopsina presente nei recettori. Si è trovato, invece, che il logaritmo della soglia è una funzione lineare della quantità di rodopsina ancora scissa e, quindi, della quantità di opsina ancora libera.
b) Psicofisiologia della brillanza
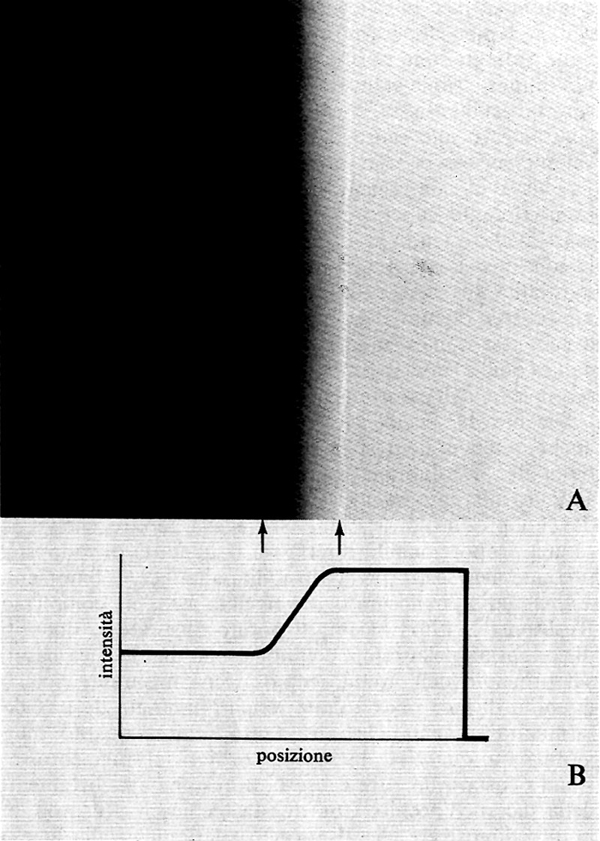
La brillanza è il correlato percettivo della luminanza, che è il valore dell'intensità luminosa per unità di superficie. La brillanza non è sempre una funzione monotonica della luminanza e quindi dell'eccitazione dei recettori e delle vie visive. Questa mancanza di correlazione monotonica tra stimolo e percezione è dimostrata dai fenomeni cosiddetti ‛al contorno', che si hanno quando l'immagine ha zone d'ombra e zone di luce adiacenti. L'effetto è particolarmente evidente quando da zone d'ombra si passa con gradualità a zone di luce. Un esempio è riportato nella fig. 17. La distribuzione spaziale dell'intensità luminosa oggettiva è riportata in B; la distribuzione della brillanza, cioè della intensità luminosa soggettiva, in A. Si noti che nei punti di passaggio dalla porzione chiara a quella più scura della figura si sono formate due strisce, rispettivamente più chiara e più scura, che non corrispondono alla distribuzione della intensità luminosa. Sono queste le bande o righe di Mach, dal nome dell'eminente fisico che le scoprì oltre 100 anni or sono.

Altro fenomeno in cui la brillanza non è correlata monotonicamente all'intensità luminosa è il fenomeno del ‛contrasto simultaneo'. Questo fenomeno è illustrato nella fig. 18: i quadrati più piccoli hanno tutti la stessa intensità luminosa, ma evidentemente non la stessa brillanza in dipendenza dalle diverse intensità luminose dei quadrati più grandi che fanno da sfondo: più lo sfondo è luminoso e più i quadrati piccoli appaiono scuri. Aumentando notevolmente lo sfondo luminoso si arriva a ottenere dei gradi di nero che è impossibile avere altrimenti. Questo fenomeno è stato studiato quantitativamente da E. G. Heinemann (v., 1955).
c) Fenomeni di interazione tra zone eccitatorie e inibitorie del sistema visivo rilevati con metodi psicofisici
L'analisi elettrofisiologica a livello delle varie stazioni del sistema visivo ha dato e dà, come abbiamo visto nel precedente capitolo, un'enorme quantità di informazioni sul modo in cui il segnale visivo viene analizzato e trasmesso. Questa tecnica, però, si è rivelata fino a ora insufficiente a farci conoscere come l'immagine visiva venga ricostruita nel nostro cervello. Per questa ragione è interessante valutare alcune ricerche psicofisiche nell'uomo, per studiare fino a che punto l'elettrofisiologia è in accordo con i risultati dell'esperienza subbiettiva. In questi ultimi anni alcuni ricercatori hanno sistematicamente unito i due approcci di ricerca nell'intento di correlare i risultati di una tecnica con quelli dell'altra. Questa metodologia sembra essere stata, ed essere tuttora, assai fruttuosa (v. Fiorentini e altri, 1971).
Tali ricerche hanno messo in evidenza che anche nell'uomo è possibile delimitare campi recettivi, cioè aree concentriche con influenze eccitatorie e inibitorie, nella visione di un determinato stimolo luminoso. Questi esperimenti possono essere condotti in vari modi. Si può determinare la soglia di percezione di un dischetto luminoso con un diametro che sottende 2-3 minuti di arco, e vedere come questa soglia vari in funzione del diametro di un anello luminoso di spessore costante, disposto concentricamente al dischetto. I risultati di questi esperimenti dimostrano che per i diametri più piccoli dell'anello la soglia di percezione del dischetto diminuisce: evidentemente entrambe le immagini cadono in una zona di sommazione spaziale di effetti eccitatori. Per diametri più ampi la soglia del dischetto luminoso aumenta, e ciò può essere spiegato con l'ipotesi che l'anello cada su una zona periferica inibitrice del campo recettivo; per diametri ancora più ampi non si ha alcun effetto, perché l'immagine dell'anello cadrebbe al di fuori del campo recettivo.
Le dimensioni di questi campi recettivi, che chiameremo psicofisici, variano sensibilmente con il livello di adattamento alla luce: il loro diametro diminuisce con l'aumentare dell'intensità della luce. Per dare un'idea delle loro dimensioni si può ricordare che nella visione foveale, con l'adattamento a una luce di 4 cd/m2, il centro del campo recettivo ha un diametro dell'ordine di 5-7 minuti di arco, circondato da una zona antagonista all'incirca uguale o leggermente più grande. Il termine di campo recettivo psicofisico, sopra introdotto, non deve essere confuso con il campo recettivo delle cellule delle stazioni visive. Esso vuole richiamare soltanto una somiglianza di proprietà spaziali e funzionali, in particolare con il campo recettivo delle cellule gangliari retiniche o di quelle del genicolato: si noti che anche le dimensioni sono dello stesso ordine di grandezza. È chiaro che stimoli luminosi come quelli usati nell'esperimento psicofisico riescono a stimolare una moltitudine di campi recettivi retinici variamente sovrapposti. Riesce perciò difficile capire le proprietà del campo recettivo psicofisico, così simili a quelle del campo recettivo di una sola cellula gangliare o del genicolato. Si può pensare a riorganizzazioni complesse dell'informazione a livello centrale o forse, più semplicemente, si può fare l'ipotesi che nell'esperimento psicofisico siano sì coinvolti molti campi recettivi, ma che la risposta sia dominata da quelli in cui le parti eccitatorie e inibitorie sono ordinatamente sovrapposte nello spazio.
Il risultato fondamentale, a mio parere, è il fatto che le stesse proprietà trovate nella retina o nel genicolato laterale sono ancora valide a livello percettivo. Le regole funzionali della cellula retinica non sono sostanzialmente alterate nella successiva elaborazione e ciò ricompensa il lavoro di analisi e alimenta la speranza dell'elettrofisiologo.
Ricerche quantitative condotte elettrofisiologicamente nella retina del gatto, e psicofisicamente nell'uomo, hanno dimostrato inoltre una sorprendente somiglianza tra le proprietà di integrazione spaziale e temporale nel campo recettivo delle cellule gangliari e le proprietà corrispondenti nel campo recettivo psicofisico. Queste ricerche sono state recentemente vieppiù incoraggiate dal fatto che registrazioni elettrofisiologiche della retina isolata dell'uomo hanno dimostrato che l'organizzazione dei campi recettivi della retina umana è del tutto simile a quella degli altri mammiferi (v. Weinstein e altri, 1971).
d) Caratteristiche temporali della visione
L'occhio è capace di risolvere stimoli luminosi separati nel tempo solo fino a una certa frequenza di ripetizione. Si definisce ‛frequenza critica di fusione' la frequenza di ripetizione dello stimolo, oltre la quale lo stimolo non ci appare più variante nel tempo, ma continuo. Questa frequenza dipende dalla luminanza dello stimolo ed è al massimo dell'ordine di 50-60 stimoli al secondo. C'è una relazione lineare tra la frequenza di fusione e il logaritmo della luminanza dello stimolo. Questa relazione va sotto il nome di legge di Ferry-Porter:
f=a log l+b,
dove f è la frequenza critica di fusione, l la luminanza dello stimolo e a e b sono due costanti. In questi ultimi tempi le proprietà temporali dell'occhio sono state studiate in maniera quantitativa, usando principalmente i metodi dell'analisi lineare e talvolta anche dell'analisi non lineare. Si è considerato il sistema visivo come una serie di filtri in cascata e si è cercato di descrivere matematicamente il funzionamento di questi filtri. Un semplice metodo che permette di risalire alla descrizione matematica di un sistema lineare è quello che fa uso dell'analisi sinusoidale. Praticamente per l'occhio si procede come segue: si presenta al soggetto umano o all'animale una luce la cui intensità è modulata sinusoidalmente nel tempo, a varie frequenze. Nel caso dell'animale si considerano poi l'ampiezza e la fase delle risposte elettriche, che per quanto riguarda la retina sono, nelle adatte condizioni sperimentali, in prima approssimazione sinusoidali.
Per l'analisi psicofisica si considera invece la soglia di modulazione, cioè la più piccola ampiezza di modulazione dello stimolo luminoso che ancora lo fa apparire modulato nel tempo, alle diverse frequenze dello stimolo.
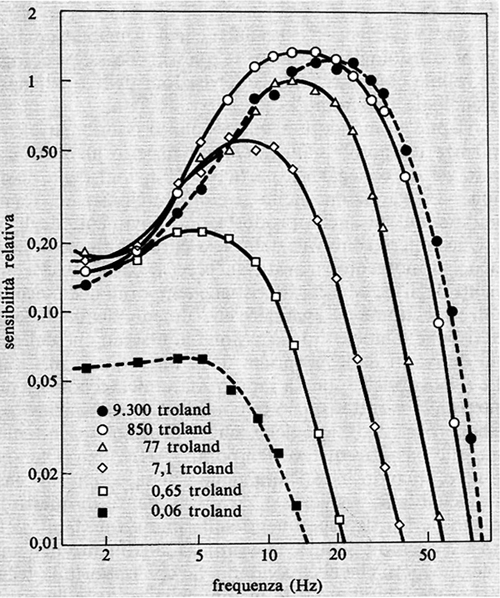
Già nel 1952 il De Lange, usando la tecnica sopra accennata, descrisse le caratteristiche psicofisiche del sistema visivo dell'uomo. Queste caratteristiche sono riportate, così come risultano da esperimenti successivi del Kelly (v., 1961), nella fig. 19. Si noti che per alte e medie luminanze dello stimolo le basse frequenze, intorno a 1-2 cicli per secondo (Hz), sono percepite peggio di un gruppo di frequenze intorno a 10 Hz, in corrispondenza delle quali la curva mostra un picco. Al di sopra di queste frequenze centrali, lo stimolo è percepito via via con maggiore difficoltà fino alla frequenza critica di fusione.
Le caratteristiche del filtro sono, come si dice in linguaggio ingegneristico, di tipo passa-banda. A luminosità mesopiche o scotopiche le curve presentano scarsa o nessuna tendenza a formare un picco. Queste curve sono chiamate di tipo passa-basso. Le stesse caratteristiche temporali della visione umana le ritroviamo a livello dell'attività elettrica retinica o del genicolato negli animali.
Recenti ricerche a livello delle cellule gangliari del gatto hanno chiarito quali siano i meccanismi che rendono la nostra percezione migliore intorno a 8-10 Hz, nell'ipotesi, d'altronde molto probabile, che la retina dell'uomo possegga meccanismi simili a quelli del gatto. Si è visto, innanzitutto, che le curve di trasferimento del centro e della periferia del campo recettivo, analizzate isolatamente, non mostrano tendenza a formare picchi nell'ambito delle frequenze centrali (5-10 Hz); inoltre la curva della periferia indica una minore capacità nell'analisi delle alte frequenze. Alle basse frequenze di stimolazione le risposte del centro e della periferia sono praticamente in antifase e, se si illuminano simultaneamente le due regioni antagoniste, l'ampiezza della risposta è, in prima approssimazione, uguale alla differenza delle due risposte prese separatamente. Poiché il centro e la periferia hanno caratteristiche di trasferimento diverse e anche la latenza delle loro risposte è diversa, la differenza di fase tra le risposte del centro e della periferia non è costante, ma è funzione della frequenza dello stimolo. Accade così che intorno a 8-10 Hz le risposte delle due regioni siano praticamente in fase e che la risposta a una loro stimolazione simultanea diventi praticamente uguale alla somma delle due prese separatamente, dando luogo così al picco nelle caratteristiche di trasferimento temporali.
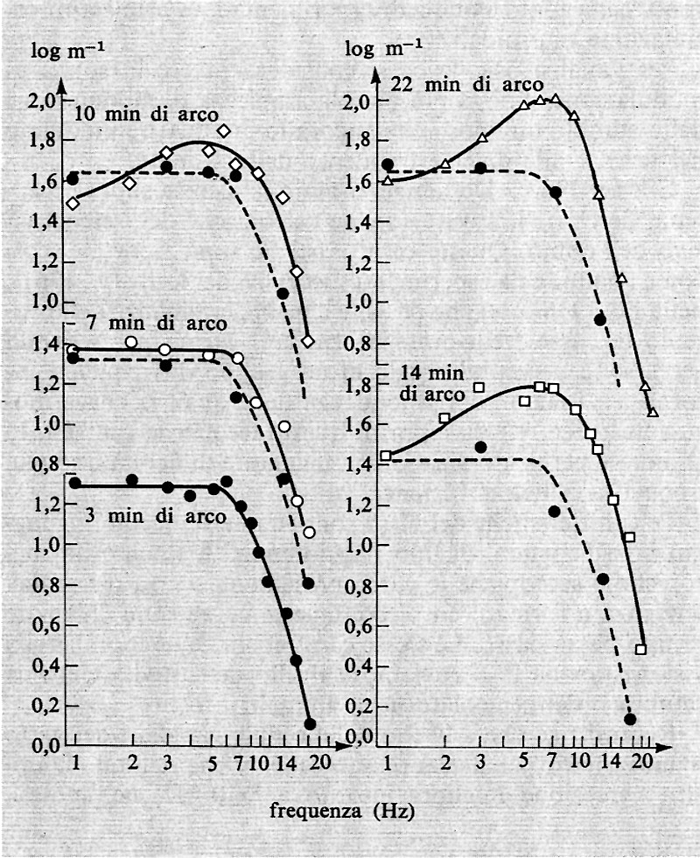
Contemporanei esperimenti condotti sull'uomo con la stessa metodologia, stimolando soltanto il centro del campo recettivo psicofisico, o centro e periferia simultaneamente, hanno dato risultati concordanti con quelli ottenuti nell'animale. Questi risultati sono riportati nella fig. 21. Si noti che, quando il diametro del disco luminoso è inferiore a 7 minuti di arco, le curve sono del tipo passa-basso e non presentano nessun picco. Come precedentemente detto, il diametro del centro del campo recettivo psicofisico è dell'ordine di 7-8 minuti. Appena il disco supera questo diametro, le curve mostrano chiaramente un picco intorno alle frequenze medie di stimolazione.
Il paragone col comportamento delle cellule gangliari del gatto mostra somiglianze sorprendenti. Mach già più di cento anni fa scriveva: ‟Può darsi che il principale compito della psicofisiologia sia di capire questi processi psicologici nelle cellule gangliari, dalle interreazioni delle quali nasce la psicologia dell'uomo".
Ci si può domandare a questo punto come mai alcune frequenze temporali siano facilitate rispetto ad altre e se questo sia da collegarsi a particolari funzioni della nostra visione. La variazione dello stimolo luminoso nel tempo tende a simulare quella prodotta dai movimenti degli occhi. Questi però sono vari e di varia natura e riesce difficile, almeno al momento, trovare un preciso correlato fra movimenti degli occhi e caratteristiche temporali della visione. D'altronde, non è da escludere che almeno alcune delle caratteristiche temporali della visione non siano che epifenomeni derivanti dalle proprietà della linea di trasmissione.
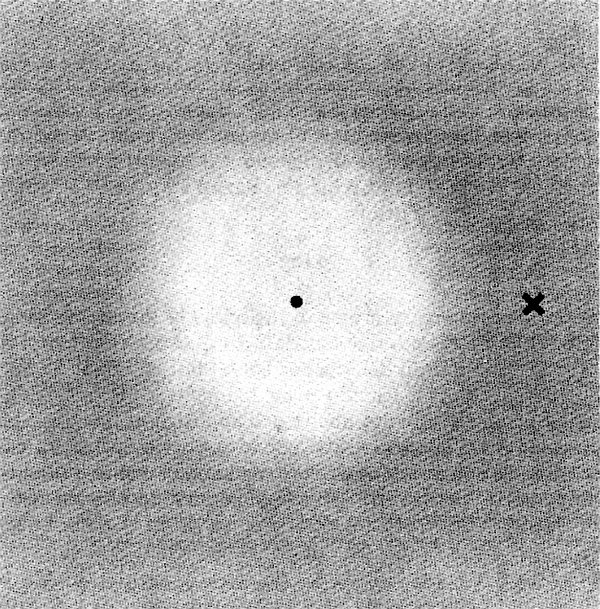
Le risposte visive sono a rapido adattamento, come si può notare dalle caratteristiche temporali di trasferimento psicofisiche o elettrofisiologiche, che mostrano una attenuazione alle basse frequenze temporali. A frequenza temporale zero, quando cioè l'immagine è fissa, anche la sensibilità del sistema visivo va a zero. L'evidenza di questo fenomeno deriva da esperimenti sulle immagini stabilizzate. Il nostro occhio è sempre in continuo movimento nell'analisi degli oggetti che ci circondano. Se con opportune tecniche si riesce a bloccare le immagini nella retina, si hanno le cosiddette ‛immagini stabilizzate'. In queste condizioni sperimentali l'immagine nell'ambito di pochi secondi perde i colori e i contorni fino a svanire completamente. Del resto è noto che nella visione usuale noi non siamo capaci di vedere l'ombra dei vasi della nostra retina, e ciò perché essi sono fissi rispetto alla retina e si muovono con essa. Un'elegante dimostrazione dell'importanza dei movimenti dell'occhio può essere fatta con l'aiuto della fig. 22. Se in visione monoculare fissiamo attentamente il punto nel mezzo del disco, questo, dopo alcuni secondi, scompare. Basta battere gli occhi o spostare lo sguardo al punto X che il disco ricompare. La scomparsa del disco è favorita in questo caso dal fatto che il disco ha un contorno sfumato, e quindi gli effetti di eventuali movimenti involontari degli occhi vengono ridotti.
A questo punto è logico domandarsi quali siano i movimenti oculari che mantengono la visione. L'occhio possiede tutta una varietà di movimenti volontari e involontari che qui non abbiamo lo spazio di analizzare. Sembra che a mantenere la visione siano sufficienti dei rapidi micromovimenti involontari, dell'ordine di qualche minuto di arco, che il nostro occhio compie continuamente anche quando noi tentiamo di fissare il nostro sguardo col massimo dell'attenzione. Ugualmente importanti sarebbero movimenti più ampi e lenti chiamati drifts.
e) Caratteristiche spaziali della visione
Fino a questi ultimi anni l'analisi del fattore spazio nella visione era limitata, dal punto di vista psicofisico, alla misura dell'acuità visiva. Quest'ultima è una funzione della posizione dell'immagine sulla retina ed è di gran lunga maggiore nella regione foveale, dove solo i coni sono presenti.
L'acuità visiva può essere misurata in diverse maniere, per esempio come reciproco della minima distanza per cui due punti o due linee si percepiscono ancora come separati (minimo separabile). L'acuità visiva misurata con due linee è maggiore che non quella tra due punti: la distanza minima separabile per due linee è 40″, fra due punti da 1′ a 1′ 5″ di arco. Si può determinare anche il minimo visibile, ossia le dimensioni minime d'un oggetto che possa ancora essere percepito. Per un punto nero o un filo nero su fondo luminoso il minimo visibile è rispettivamente dell'ordine di 14″ e di 1″ di arco. Si è discusso molto in passato su quali fossero i fattori limitanti l'acuità visiva. Per molto tempo si è pensato che l'unico limite alla discriminazione visiva fosse posto dalle dimensioni dei recettori, dei coni in particolare. Teoricamente le dimensioni dei recettori svolgono al riguardo un ruolo fondamentale, se non si considera il fatto che altri fattori riguardanti l'ottica dell'occhio o il sistema nervoso possono intervenire a limitare più drasticamente l'acuità visiva, prima che il limite delle dimensioni dei recettori possa entrare in gioco. Il diametro dei coni della fovea è dell'ordine di 18-20″ di arco e la distanza tra due coni vicini è dell'ordine di 10-20″ di arco. È chiaro che se le immagini di due punti cadono nello stesso recettore, i punti non possono essere visti come separati. Se le immagini cadono su due recettori vicini, i punti possono essere visti come distinti solo se hanno diverse luminanze. Teoricamente se un recettore a riposo è interposto fra quelli eccitati, i due punti luminosi possono essere sempre visti come separati. Nel peggiore dei casi, l'acuità visiva, determinata con due punti luminosi, dovrebbe essere quindi dell'ordine di 50-55″. Risulta, invece, che è superiore al minuto di arco.
Un'altra osservazione, la quale rende poco probabile che le dimensioni dei recettori siano un fattore essenziale nel limitare l'acuità visiva, è che essa varia con la luminanza media e, precisamente, è maggiore a luminanze maggiori. Le variazioni piu notevoli si hanno nell'ambito delle luminanze fotopiche basse e medie. Poiché a questi livelli di luminanza non si può attribuire il miglioramento della discriminazione visiva alla maggiore disponibilità di fotoni per luminanze maggiori, e poiché è assurdo pensare che le dimensioni dei recettori varino con la luminanza, non resta che pensare a variazioni di influenze nervose.
Per quanto riguarda il ‛minimo visibile', per un punto nero su fondo chiaro esso è, come già detto, intorno ai 14″, misura che approssima il diametro dei più piccoli coni. Sarebbe però assurdo pensare che la visione di un singolo punto sia legata alle dimensioni dei recettori. Le dimensioni dei recettori sono in questo caso completamente fuori questione. Il fatto fondamentale è che un recettore viene eccitato mentre altri vicini non lo sono o lo sono meno, o viceversa. Quello che si misura è dunque una soglia differenziale. Teoricamente anche recettori di dimensioni molto maggiori avrebbero la stessa possibilità discriminativa, a patto che esistesse un sufficiente contrasto fra il punto test e lo sfondo.
Sperimentalmente si vede infatti che l'acuità visiva diminuisce se il contrasto tra il punto e lo sfondo diminuisce. Il minimo visibile per una linea è addirittura dell'ordine del secondo di arco, a patto che la linea sia lunga almeno 30′. Questo risultato non è in contrasto con il precedente. Una linea di 30′ di lunghezza si estende su 60-70 coni e, quindi, l'informazione della larghezza della linea è portata da molti canali in parallelo che, convergendo, almeno parzialmente, al livello centrale, potrebbero migliorare di molto il rapporto segnale-rumore del messaggio. Simili considerazioni si possono fare per spiegare la maggiore acuità visiva fra due linee, che, come già detto, è di circa 40″, rispetto a quella tra due punti, che è di circa 60-65″.
Secondo chi scrive, il problema dei limiti posti all'acuità visiva dalle dimensioni dei recettori ha senso solo nella distinzione tra due punti di uguale luminosità, come detto sopra; infatti già nella determinazione dell'acuità visiva tra due linee giocano fattori di integrazione nervosa. Nella percezione del minimo visibile, punto o riga nera su sfondo luminoso, la dimensione dei recettori non può giocare, neppure teoricamente, alcun ruolo.
D'altronde, riguardo all'acuità visiva tra due punti, abbiamo osservato che questa è leggermente peggiore di quella predetta in base alle dimensioni dei recettori. È probabile che i limiti essenziali siano qui posti dall'ottica dell'occhio, sulle cui caratteristiche funzionali ritorneremo tra poco.
Lo studio dell'acuità visiva ci dà solo il limite superiore di funzionamento del sistema, raramente impegnato a quel limite nella visione normale del mondo esterno. Esso non ci dà invece nessuna informazione su come i vari oggetti di diverse dimensioni vengano analizzati e percepiti. D'altronde, il problema essenziale è proprio capire se oggetti di certe dimensioni sono percepiti meglio di oggetti di dimensioni diverse, più grandi o più piccoli, anche in funzione dei vari gradi di contrasto dell' immagine, fattore non considerato negli esperimenti classici sull'acuità visiva.

Un recente studio ha chiarito in gran parte quali siano i fattori limitanti la visione delle diverse frequenze spaziali. Per definire la ‛frequenza spaziale' ci si riferisce a una mira periodica, per esempio un reticolo a righe parallele chiare e scure. Il numero delle sbarre chiare o scure, contenute nell'unità di angolo visivo, viene definito come la frequenza spaziale di quel reticolo. Per ogni frequenza spaziale esiste un contrasto minimo tra le sbarre chiare e scure, per cui il soggetto è ancora capace di risolvere il reticolo. Questo contrasto minimo è definito come la soglia di contrasto per vedere il reticolo di quella data frequenza spaziale. Il fatto che un certo ambito di frequenze spaziali ha soglia di contrasto minore di altre può essere subito rilevato osservando la fig. 23. In questa figura il contrasto varia logaritmicamente lungo le ordinate e la frequenza spaziale logaritmicamente lungo le ascisse. Si può chiaramente vedere che le alte e basse frequenze spaziali (estremi destro e sinistro della figura) sono percepite a contrasti più alti che non le frequenze spaziali intermedie (al centro della figura).
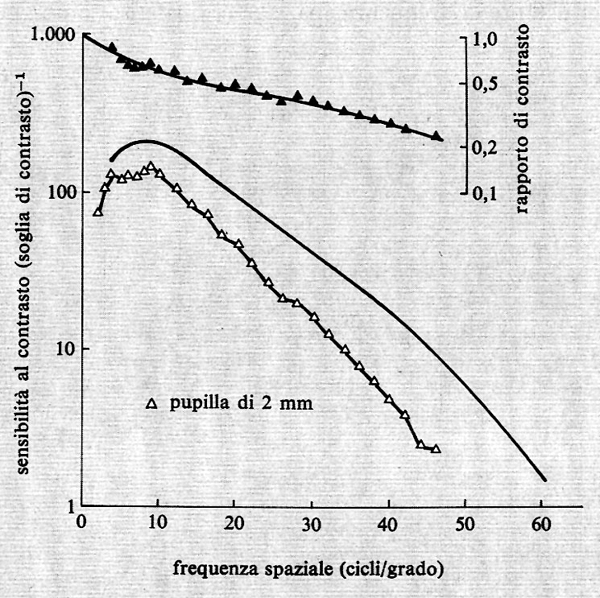
Le caratteristiche di trasferimento della sensibilità al contrasto alle varie frequenze spaziali sono riportate nella fig. 24 con linee continue. Queste caratteristiche sono quelle di un filtro passa-banda.
Le frequenze spaziali meglio percepite, per le quali cioè la soglia di contrasto è più bassa, sono intorno a 3-6 c/grado, mentre il limite risolutivo superiore arriva a 40-50 c/grado. L'esistenza di questa percezione differenziale delle varie frequenze spaziali, il fatto cioè che alcune frequenze intermedie vengano percepite meglio di altre e pertanto l'osservazione che la curva non è monotonica, è già di per sé la dimostrazione che le dimensioni dei recettori non sono l'unico fattore nell'elaborazione della variabile spazio e, quindi, dell'acuità visiva.
I fattori limitanti la visione alle diverse frequenze spaziali possono essere divisi essenzialmente in ottici e nervosi. Teoricamente i primi devono presentare senza dubbio una qualche influenza, perché non esiste nessuna trasformazione ottica, dall'oggetto all'immagine, o di qualsiasi altro tipo, che non comporti una perdita di informazione. In generale, se un messaggio ha la stessa quantità di informazione in due punti di una linea di trasmissione, ciò significa che non ha subito nessuna trasformazione.
Fino a poco tempo fa si credeva che gran parte dell'attenuazione alle alte frequenze spaziali derivasse da fattori ottici. Vi sono molte ricerche, vecchie e recenti, intese a determinare i limiti posti alla nostra visione dall'ottica dell'occhio. La più recente ed eloquente di queste mi sembra quella del gruppo di Cambridge (v. Campbell e Green, 1965). Questi ricercatori hanno proiettato direttamente sulla retina delle frange di interferenza chiaro-scura di varie frequenze. Tale metodo consente di ottenere le caratteristiche di trasferimento spaziali, evitando le trasformazioni dell'ottica dell'occhio. I risultati sono riportati nella fig. 24 (linea continua). Si nota subito che anche le caratteristiche di trasferimento spaziali, che chiameremo nervose, presentano un'attenuazione alle alte e alle basse frequenze spaziali che non appare sostanzialmente diversa da quella presentata dalle caratteristiche di trasferimento quando l'ottica dell'occhio è inclusa (triangoli vuoti). Poiché la parte ottica e la parte nervosa del sistema visivo sono due sistemi in serie, è possibile risalire alle caratteristiche di trasferimento proprie solo dell'ottica dell'occhio (triangoli pieni). Risulta, in realtà, che l'attenuazione provocata dall'ottica dell'occhio è abbastanza piccola e riguarda particolarmente le frequenze medie e alte.
Cade così il vecchio preconcetto che la nostra acuità visiva e, in generale, la nostra visione della variabile spazio, sia sostanzialmente limitata dall'imperfezione della nostra ottica. Di conseguenza, cade anche un'altra teoria che sosteneva che l'organizzazione dei campi recettivi retinici in zone antagoniste non fosse che un meccanismo del sistema nervoso atto a correggere, specie nella visione dei margini e dei chiaroscuri, le imperfezioni introdotte dall'apparato ottico dell'occhio.
Riassumendo, i fattori nervosi sono quelli che caratterizzano maggiormente la visione della variabile spazio.
Nelle esperienze sopra citate, il contrasto dei reticoli usati, supponiamo a sbarre verticali, era modulato sinusoidalmente lungo la direzione orizzontale. Questi segnali sono i più semplici possibili e ogni altro segnale può essere pensato come composto dalla sintesi di sinusoidi di opportuna frequenza e ampiezza (sintesi di Fourier). Le curve delle figure, poiché riportano risultati di esperimenti nei quali si usavano reticoli il cui contrasto era modulato sinusoidalmente e poiché si comportano come sistemi lineari, rappresentano le caratteristiche di trasferimento spaziali del sistema ottico o nervoso.
Resta da risolvere il problema di come la variabile spazio venga analizzata dalle varie stazioni nervose, e di come l'immagine venga ricostruita a livello dei centri superiori. Su questo argomento non si possono ricordare che delle ipotesi. La più recente di queste trae spunto da esperimenti psicofisici ed elettrofisiologici (una sintesi della letteratura si trova in Blakemore e Campbell: v., 1969). Si è fatta l'ipotesi che nella visione delle forme (sono considerate solo due dimensioni) le stazioni più periferiche del sistema visivo funzionino da analizzatori di frequenze spaziali. Ogni frequenza o banda di frequenze spaziali sarebbe portata ai centri in canali separati. Si postula, poi, che la ricostruzione dell'immagine avvenga per la convergenza delle informazioni da vari canali.

Fra gli svariati esperimenti psicofisici che dimostrano come l'informazione visiva sia codificata in canali di frequenze spaziali a banda limitata, il più facilmente ripetibile dallo stesso lettore è quello di Blakemore e Campbell (v., 1969). Essi hanno dimostrato che la prolungata osservazione di un reticolo di alto contrasto riduce o temporaneamente annulla la visibilità di un reticolo di basso contrasto della stessa frequenza spaziale e dello stesso orientamento. Il lettore può verificare questo fenomeno guardando il reticolo in alto a sinistra della fig. 25. Dopo un certo periodo di osservazione, che chiameremo di adattamento, il reticolo centrale sembra apparire uniforme per 10-20 s.
Questo fenomeno è selettivo sia per l'orientamento che per la frequenza spaziale, come il lettore può facilmente verificare adattandosi agli altri reticoli di alto contrasto della figura. Blakemore e Campbell, adattandosi a reticoli di svariate frequenze spaziali, sono riusciti a determinare l'ampiezza della banda del canale delle frequenze spaziali, che è circa una ottava.
È stato possibile trovare anche il correlato elettrofisiologico dei canali delle frequenze spaziali nelle cellule della corteccia visiva striata sia del gatto che della scimmia (v. Maffei, 1978).
Questi neuroni, e in particolare le cellule semplici, presentano altissima selettività per un reticolo di una data frequenza spaziale, dando risposte minori man mano che ci si allontana dalla frequenza spaziale ottimale dello stimolo. Questa selettività alle frequenze spaziali aumenta progressivamente dalle cellule gangliari della retina alle cellule del corpo genicolato laterale, alle cellule corticali.
f) Sensibilità della visione per diversi orientamenti dell'immagine
Abbiamo visto, a proposito dell'elettrofisiologia delle vie visive, che le risposte delle cellule corticali mostrano una notevole specificità all'orientamento dello stimolo. Ricerche quantitative hanno dimostrato che le cellule corticali del gatto danno risposte solo nell'ambito di +14° intorno all'orientamento ottimale. Nell'uomo l'informazione orientamento è trasmessa in canali ancora più ristretti, come hanno messo in evidenza ricerche del gruppo di Cambridge.
È un fatto ben noto che il potere risolutivo del sistema visivo dell'uomo è migliore per mire verticali e orizzontali che per mire oblique, e si sa anche che questa caratteristica non è dovuta a proprietà astigmatiche dell'ottica dell'occhio. La sensibilità al contrasto, per esempio, ha una soglia nel meridiano a 45° che è circa il doppio rispetto a quella nei meridiani verticale e orizzontale. È risultato, inoltre, che questa è una elaborazione probabilmente corticale, in quanto i potenziali evocati da reticoli dello stesso contrasto mostrano ampiezze molto maggiori per i meridiani orizzontale e verticale che non per i meridiani obliqui. D'altronde queste differenze non sono osservate a livello delle risposte elettroretinografiche.
Si pensa che questa maggiore abilità nei meridiani verticali e orizzontali sia dovuta alla nostra maggiore esperienza visiva in questi assi del piano, prevalendo nel mondo esterno principalmente linee verticali e orizzontali.
g) Percezione della distanza
È difficile comprendere come si possano estrarre dall'immagine retinica bidimensionale indizi per la percezione della terza dimensione dello spazio visivo. Ne deriva logicamente che nella visione monoculare la proiezione della terza dimensione non può che derivare da una interpretazione dell'immagine bidimensionale retinica. Negli animali, come l'uomo, che hanno i campi visivi dei due occhi largamente sovrapposti, esiste una possibilità di valutazione molto più accurata e diretta della terza dimensione, basata sull'elaborazione delle due immagini retiniche e, precisamente, sulla differenza che esiste nelle proiezioni sulle due retine dello stesso oggetto (parallasse binoculare).
La parallasse binoculare, o dispapità, sorge poiché sulle due retine si formano proiezioni della stessa scena visiva da due punti di vista leggermente diversi.
Il lettore può rendersi conto di questo fenomeno tenendo un dito davanti ai propri occhi mentre guarda una determinata scena visiva. Si accorgerà che se prima chiude un occhio e poi un altro il dito appare in posizione diversa rispetto allo sfondo. Ciò deriva dal fatto che l'immagine del dito sulle due retine non cade in punti corrispondenti di esse.

Per capire meglio il ruolo giocato dalla parallasse binoculare, può essere d'aiuto lo schema riportato nella fig. 27, dove Fd e Fs sono la fovea destra e sinistra. Per definizione tutti i punti che giacciono sul circolo che passa per il punto di fissazione e i punti nodali degli occhi hanno parallasse binoculare nulla. Punti giacenti all'interno o all'esterno di questo circolo presentano disparità, cioè, per essi,
αs, βs≠αd, βd.
Poiché gli occhi sono posti su una linea orizzontale, le disparità rilevanti sono quelle su questo piano dello spazio. É chiaro che la proiezione della stessa immagine su punti non esattamente corrispondenti delle due retine dà informazioni sulla distanza delle diverse parti dell'oggetto o degli oggetti dai nostri occhi. La nostra abilità nel distinguere e valutare queste disparità è notevole. L'acuità stereoscopica è dell'ordine di 10″, cioè, riferendosi alla figura, basta che l'angolo αs (o βs) sia diverso di una quantità dell'ordine dei 10″ dall'angolo αd (o βd) per provocare la percezione della profondità. Nell'uomo la cosiddetta ‛area Panum foveale', l'area foveale, cioè, all'interno della quale è possibile fondere le immagini provenienti dai due occhi, è di circa 6′. L'estrema accuratezza con cui, grazie alla visione binoculare, noi valutiamo le distanze degli oggetti non riguarda la loro distanza assoluta dai nostri occhi, bensì le distanze relative, cioè le differenze delle distanze tra i diversi oggetti. La distanza assoluta è valutata in massima parte per mezzo di indizi monoculari che prendono spunto da esperienze precedenti. Fino a non molto tempo fa si pensava che la valutazione delle distanze relative in scene complesse (folla, fogliame di un albero, ecc.) avvenisse tra forme già individuate, tra elaborate Gestalten: nel caso della folla, per esempio, tra una determinata faccia e un'altra. I recenti esperimenti di Julesz mostrano che la valutazione delle disparità può avvenire anche senza alcun riferimento a forme o contorni. Questo autore ha generato con l'uso del calcolatore stereogrammi puntiformi completamente privi, quando visti con un solo occhio, di forme o contorni, e che tuttavia hanno gruppi di punti aventi la stessa disparità. Quando i due stereogrammi sono fusi binocularmente con l'uso di uno stereoscopio, per esempio, i punti aventi la stessa disparità appaiono chiaramente in rilievo. Solo in questi ultimi anni sono state intraprese ricerche intese a studiare i meccanismi neurofisiologici della visione binoculare. Questi studi sono stati fatti per la massima parte al livello della corteccia visiva del gatto e della scimmia. A questo livello, infatti, la maggior parte delle cellule riceve informazioni da entrambi gli occhi. Si è trovato che l'attività di una classe di cellule corticali, se sottoposte a stimolazione binoculare, dà risposte massime non quando gli stimoli cadono in punti corrispondenti delle due retine, bensì quando le loro immagini sulle due retine presentano una certa disparità.
h) Indizi monoculari
Anche se chiudiamo un occhio, tuttavia il nostro spazio percettivo non diventa bidimensionale e ben poco sembra succedere. Questo è già di per sé dimostrazione dell'esistenza di indizi monoculari nella percezione della distanza. Ne elenchiamo alcuni di seguito: 1) dimensione di oggetti familiari: se si conoscono le dimensioni relative degli oggetti, è chiaro che allora l'informazione sulla loro reale dimensione nella immagine retinica può essere usata per valutare le loro relative distanze; 2) ombre: nota per altri indizi la posizione della sorgente luminosa, la posizione delle ombre di certi oggetti rispetto ad altri ci dà informazioni sulla loro relativa posizione; 3) parallasse monoculare: visione di un oggetto da diversi punti di vista, tecnica sfruttata in pittura, in particolare dai cubisti analitici; 4) sovrapposizione: ogni oggetto non trasparente impedisce la vista di altri oggetti posti dietro di esso; 5) presenze di gradienti regolari: ogni regolarità nella struttura di una superficie (ad es. le mattonelle di un pavimento) appare tanto più grande quanto più vicina è all'occhio dell'osservatore; 6) dimensioni di oggetti o esseri familiari: oggetti o animali di dimensioni note sono di riferimento per la valutazione di altri oggetti più vicini o lontani; 7) prospettiva lineare: nota in arte fin dai tempi antichi ma portata a teoria nel Rinascimento, in particolare a opera del Brunelleschi. Essa ci insegna che a partire da una certa distanza le cose ci appaiono di dimensioni inversamente proporzionali alla distanza. Questo indizio è largamente usato in arte: si pensi, ad es., alla Trinità del Masaccio in Santa Maria Novella a Firenze; 8) prospettiva aerea: le minuscole particelle dell'atmosfera diffondono la luce riducendo così il contrasto di oggetti lontani, che appaiono anche più bluastri poiché la luce di breve lunghezza d'onda è più diffusa dalle particelle atmosferiche.
i) Visione dei colori
La fusione dei tre colori fondamentali dello spettro è sufficiente a riprodurre tutti gli altri colori. Biologicamente, almeno nei Primati, la visione dei colori è realizzata con un meccanismo molto simile. Infatti nella nostra retina esistono tre diverse classi di coni con spettro di assorbimento diverso. Questa scoperta è del tutto recente, anche se la probabile esistenza nella retina di tre sostanze sensibili in maniera diversa alla luce era già stata ipotizzata da Th. Young nel 1802.
La scoperta di tre pigmenti, diversamente sensibili alla luce di diversa lunghezza d'onda nella retina dell'uomo, è frutto di due brillanti serie di ricerche effettuate negli anni sessanta. Da un lato ci sono infatti le ricerche di Marks, MacNichol, Brown e Wald (v. Brindley, 1970), che sono riusciti a misurare, con metodo microspettrofotometrico, l'assorbimento di singoli coni della retina dell'uomo e della scimmia. D'altro lato vi sono le ricerche di Rushton, che ha potuto mettere in evidenza tre diversi tipi di spettri di assorbimento in vivo nell'uomo con una raffinatissima tecnica di oftalmoscopia, e cioè misurando la luce riflessa dalla retina. Anche registrazioni elettrofisiologiche da singoli coni hanno dato risultati nello stesso senso.
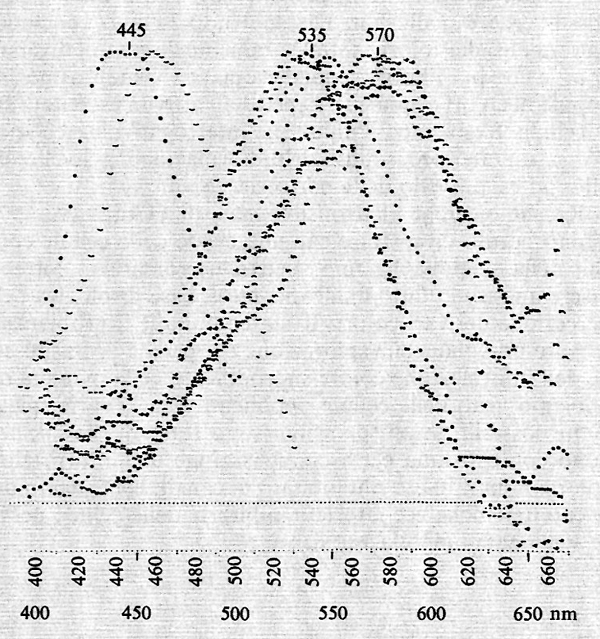
Nella fig. 29 sono riportati i risultati delle ricerche di Marks e MacNichol; si può vedere che i tre diversi spettri di assorbimento hanno i loro picchi intorno a 450 nm (blu), a 540 nm (verde) e a 570 nm (giallo).
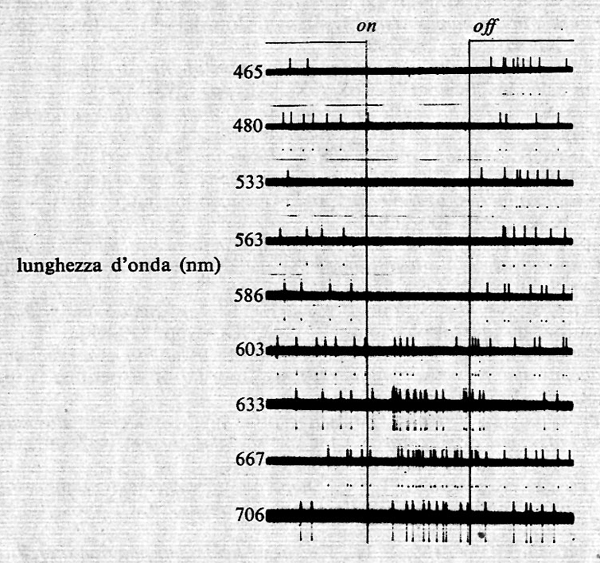
È indubbio che questi risultati danno supporto sperimentale alla vecchia teoria tricromatica della visione dei colori di Young-Helmholtz, a livello dei recettori retinici. I tre tipi di recettori però, sorprendentemente, non hanno una via privata ai centri superiori cerebrali. Infatti, registrazioni elettrofisiologiche delle cellule gangliari della retina dei Pesci e anche delle scimmie hanno messo in evidenza che gran parte delle cellule gangliari rispondono in maniera diversa a regioni diverse dello spettro, e cioè con risposte eccitatorie a certi colori e inibitorie a certi altri. Per esempio, una cellula può dare risposte eccitatorie per il verde e inibitorie per il rosso, o viceversa. Nel corpo genicolato della scimmia De Valois e Jacobs distinguono le seguenti classi di cellule, cosiddette ‛opponenti': rosso eccitatorio e verde inibitorio e viceversa, giallo eccitatorio e blu inibitorio e viceversa. La risposta unitaria di una cellula opponente del corpo genicolato laterale della scimmia Macaca è riportata nella fig. 30. Insieme a queste cellule si distinguono anche un gruppo di cellule non opponenti, che mostrano reazioni eccitatorie o inibitorie su tutto lo spettro. È ipotizzato dagli autori che quest'ultimo gruppo di cellule potrebbe essere atto a trasmettere la luminanza degli oggetti anziché il loro colore, e comunque un'informazione di tipo acromatico.
Questi risultati elettrofisiologici sembrano in una certa relazione con la vecchia teoria di Ewald Hering che sosteneva che il processo della visione dei colori è attribuibile a tre canali portanti informazione sul colore bianco-nero, rosso-verde e giallo-blu. Riassumendo, si può concludere che l'analisi dei colori è fatta certamente da tre tipi diversi di coni, e che poi l'informazione cromatica viene trasmessa a coppie di colori opponenti rispetto all'attività esercitata sulla cellula. Resta ancora un ultimo interrogativo e cioè se avvenga un'ulteriore sintesi dell'informazione a livello più alto, ad es. a livello della corteccia visiva.
Recenti ricerche elettrofisiologiche di Gouras (v., 1970) hanno potuto dimostrare che una classe delle cellule corticali della scimmia ha uno spettro tricromatico. Potrebbe essere questo il punto finale di sintesi, base fisiologica della percezione dei vari colori. Resta comunque il problema di trovare una plausibile spiegazione del perché la trasmissione dell'informazione colore sia codificata nella maniera che abbiamo visto nelle cellule opponenti. Tra le varie spiegazioni possibili, le più plausibili sono forse quelle che tengono conto di una certa condensazione dell'informazione colore in un numero più limitato di canali, usufruendo delle caratteristiche eccitatorie e inibitorie delle cellule nervose stazionate lungo la via visiva.
Bibliografia
Barlow, H. B., Blakemore, C., Pettigrew, J. D., The neural mechanism of binocular depth discrimination, in ‟Journal of physiology", 1967, CXCIII, pp. 327-342.
Blakemore, C., Campbell, F. W., On the existence of neurones in the human visual system selectively sensitive to orientation and size of retinal images, in ‟Journal of physiology", 1969, CCIII, pp. 237-260.
Brindley, G. G., The physiology of the retina and the visual pathway, London 1970.
Brown, K. T., The electroretinogram: its components and their origins, in ‟Vision research", 1968, VIII, pp. 633-677.
Campbell, F. W., The human eye as an optical filter, in ‟Proceedings of the IEEE", 1968, LVI, pp. 1009-1014.
Campbell, F. W., Green, D. G., Optical and retinal factors affecting visual resolution, in ‟Journal of physiology", 1965, CLXXXI, pp. 576-593.
De Valois, R. L., Jacobs, G. H., Primate color vision, in ‟Science", 1968, CLXII, pp. 533-540.
Dowling, J. E. Boycott, B. B., Organization of the Primate retina: electron microscopy, in ‟Proceedings of the Royal Society" (Series B), 1967, CLXVI, pp. 80-111.
Fiorentini, A., Bayly, J. E., Maffei, L., Peripheral and central contributions to psychophysical spatial interactions, in ‟Vision research", 1971, XII, pp. 253-259.
Fiorentini, A., Maffei, L., Transfer characteristics of excitation and inhibition in the human visual system, in ‟Journal of neurophysiology", 1970, XXXIII, pp. 285-292.
Gouras, P., Primate retina: duplex function of the dark-adapted ganglion cells, in ‟Science", 1965, CXLVII, pp. 1593-1594.
Gouras, P., Trichromatic mechanisms in single cortical neurons, in ‟Science", 1970, CLXVIII, pp. 489-491.
Granit, R., Sensory mechanism of the retina, Oxford-London 1947.
Hartline, H. K., The response of single optic fibers of the Vertebrate eye to illumination of the retina, in ‟American journal of physiology", 1938, CXXI, pp. 400-415.
Hecht, S., Haig, C., Chase, A. M., The influence of light adaptation on subsequent dark adaptation of the eye, in ‟Journal of general physiology", 1937, XX, pp. 831-850.
Hecht, S., Shlaer, S., Pirenne, M. H., Energy, quanta and vision, in ‟Journal of general physiology", 1942, XXV, pp. 819-840.
Heinemann, E. G., Simultaneous brightness induction as a function of inducing-and test-field luminances, in ‟Journal of experimental psychology", 1955, L, pp. 89-96.
Hubel, D. H., Wiesel, T. N., Receptive fields, binocular interaction and functional architecture in the cat's visual cortex, in ‟Journal of physiology", 1962, CLX, pp. 106-154.
Hubel, D. H., Wiesel, T. N., Shape and arrangement of columns in the cat's striate cortex, in ‟Journal of physiology", 1963, CLXV, pp. 559-568.
Hubel, D. H., Wiesel, T. N., Receptive fields and functional architecture in two nonstriate visual areas (18 and 19) of the cat, in ‟Journal of neurophysiology", 1965, XXVIII, pp. 229-289.
Hubel, D. H., Wiesel, T. N., Receptive fields and functional architecture of monkey striate cortex, in ‟Journal of physiology", 1968, CXCV, pp. 215-243.
Hubel, D. H., Wiesel, T. N., Functional architecture of Macaque monkey visual cortex, in ‟Proceedings of the Royal Society" (Series B), 1977, CXCVIII, pp. 1-59.
Humphrey, N. K., Weiskrantz, L., Vision in monkeys after removal of the striate cortex, in ‟Nature", 1967, CCXV, pp. 595-597.
Kelly, D. H., Visual responses to time-dependent stimuli I. Amplitude sensitivity measurements, in ‟Journal of the Optical Society of America", 1961, LI, pp. 422-429.
Klüver, H., Bucy, P. C., Preliminary analysis of functions of the temporal lobes in monkeys, in ‟Archives of neurology and psychiatry", 1939, XLII, pp. 979-1000.
Kuffler, S. W., Discharge patterns and functional organization of mammalian retina, in ‟Journal of neurophysiology", 1953, XVI, pp. 37-68.
Lettvin, J. Y., Maturana, H. R., McCulloch, W. S., Pitts, W. H., What the frog's eye tells the frog's brain, in ‟Proceedings of the Institute of Radio Engineers", 1959, XLVII, pp. 1940-1951.
Maffei, L., Spatial and temporal averages in retinal channels, in ‟Journal of neurophysiology", 1968, XXXI, pp. 283-297.
Maffei, L., Spatial frequency channels: neural mechanism, in Handbook of sensory physiology, vol. VIII, Perception (a cura di R. Held, H. W. Leibowitze e H. L. Teubner), Berlin-New York 1978, pp. 39-66.
Maffei, L., Campbell, F. W., Neurophysiological localization of the vertical and horizontal visual coordinates in man, in ‟Science", 1970, CLXVII, pp. 386-387.
Maffei, L., Cervetto, A., Fiorentini, A., Transfer characteristics of excitation and inhibition in cat retinal ganglion cells, in ‟Journal of neurophysiology", 1970, XXXIII, pp. 276-284.
Maffei, L., Fiorentini, A., Retino-geniculate convergence and the analysis of contrast, in ‟Journal of neurophysiology", 1972, XXXV, pp. 65-72.
Maffei, L., Fiorentini, A., The visual cortex as a spatial frequency analyser, in ‟Vision research", 1973, XIII, pp. 1255-1267.
Maffei, L., Moruzzi, G., Rizzolatti, G., Influence of sleep and wakefulness on the response of lateral geniculate units to sinewave photic stimulation, in ‟Archives italiennes de biologie", 1965, CIII, pp. 596-608.
Maffei, L., Rizzolatti, G., Effect of synchronized sleep on the response of lateral geniculate units to flashes of light, in ‟Archives italiennes de biologie", 1965, CIII, pp. 609-622.
Marks, W. B., Dobelle, W. H., MacNichol, E. F. Jr., Visual pigments of single Primate cones, in ‟Science", 1964, CXLIII, pp. 1181-1183.
Miller, R. F., Dowling, J. E., Intracellular responses of the Müller (glial) cells of mudpuppy retina: their relation to b-wave of the electroretinogram, in ‟Journal of neurophysiology", 1970, XXXIII, pp. 323-341.
Penn, R. D., Hagins, W. A., Signal transmission along retinal roads and the origin of the electroretinographic a-wave, in ‟Nature", 1969, CCXXIII, pp. 201-205.
Polyak, S., The Vertebrate visual system (a cura di H. Klüver), Chicago 1957.
Rushton, W. A. H., Visual pigments in man, in The Sherrington lectures, vol. VI, Springfield 1962.
Rushton, W. A. H., Visual adaptation. The Ferrier lecture, in ‟Proceedings of the Royal Society" (Series B), 1965, CLXII, pp. 20-46.
Sperry, R. W., Miner, N., Myers, R. E., Visual pattern perception following subpial slicing and tantalum wire implantation in the visual cortex, in ‟Journal of comparative and physiological psychology", 1955, XLVIII, pp. 50-58.
Stone, J., Fukuda, Y., Properties of cat retinal ganglion cells: a comparison of W cells with X and Y cells, in ‟Journal of neurophysiology", 1974, XXXVII, pp. 722-748.
Tomita, T., Kaneko, A., Murakami, M., Pautler, E. L., Spectral response curves of single cones in the carp, in ‟Vision research", 1967, VII, pp. 519-531.
Weinstein, G. W., Hobson, R. R., Baker, F. H., Extracellular recordings from human retinal ganglion cells, in ‟Science", 1971, CLXXI, pp. 1021-1022.
Weiskrantz, L., Warrington, E. K., Sanders, M. D., Marshall, J., Visual capacity in the hemianopic field following a restricted occipital ablation, in ‟Brain research", 1974, XCVII, pp. 709-728.
Werblin, F. S., Dowling, J. E., Organization of the retina of the mudpuppy Necturus maculosus. II. Intracellular recording, in ‟Journal of neurophysiology", 1969, XXXII, pp. 339-355.