
Sonno
Sonno
di Giuseppe Moruzzi
Sonno
sommario: 1. Introduzione storica. 2. Fenomenologia del sonno: a) il ciclo sonno-veglia; b) sonno sincronizzato e sonno desincronizzato. 3. Effetti di lesioni: a) la formazione reticolare ascendente; b) le regioni deattivanti del tronco dell'encefalo; c) alternanza di attività e connessioni reciproche tra sistemi antagonisti. 4. Effetti delle stimolazioni periferiche e centrali: a) stimolazioni periferiche; b) stimolazioni centrali. 5. Ricerche neurochimiche: a) l'approccio umorale alle ricerche sul sonno; b) i mediatori chimici del tronco dell'encefalo; c) la dottrina monoamminergica del ciclo sonno-veglia. 6. Registrazioni elettrofisiologiche da singoli neuroni del tronco dell'encefalo. 7. Il significato funzionale del sonno: a) le teorie del ristoro ipnico; b) significato del sonno desincronizzato. 8. Sonno e comportamento istintivo. 9. Interpretazioni neurofisiologiche nella patologia del sonno: a) narcolessia e cataplessia; b) letargo della malattia del sonno e coma; c) le insonnie. 10. Stati simili al sonno: a) l'ipnosi animale; b) l'ibernazione dei Mammiferi. □ Bibliografia.
1. Introduzione storica
È passato poco più di mezzo secolo da quando C. von Economo (v., 1918) riferiva che nell'encefalite letargica epidemica, o malattia del sonno, le lesioni sono localizzate soprattutto nella regione del mesencefalo che circonda l'acquedotto di Silvio e nell'ipotalamo. L'interpretazione da lui data delle sue osservazioni (v. von Economo, 1929) era errata, ma egli era certo nel giusto quando pensava che le strutture nervose lese dalla malattia epidemica avevano un'importanza critica nella regolazione del ciclo sonno-veglia.
Non molti anni dopo W. R. Hess (v., 1927) trovava che il sonno poteva essere ottenuto, nel gatto senza narcosi e libero nei suoi movimenti, con stimolazioni elettriche appropriate della regione mediale del talamo. Malgrado la possibilità di errori (v. cap. 4), questo indirizzo di ricerca ebbe un influsso profondo sullo sviluppo della fisiologia del sonno. Era la prima dimostrazione che il sonno fisiologico può essere ottenuto con stimolazioni elettriche localizzate del cervello, e i risultati dovevano essere più volte confermati stimolando altre regioni del diencefalo e del tronco dell'encefalo. Inoltre gli esperimenti di Hess aprivano la strada a una feconda linea di ricerca in cui lo studio del comportamento era associato a diversi tipi di stimolazione e di registrazione elettrofisiologica. Naturalmente era impossibile individuare, in quei tempi, un qualsiasi rapporto fra le osservazioni di von Economo e gli esperimenti di Hess. Tutto sembrava tanto differente: casi clinici ed esperimenti su animali, lesioni e stimolazioni, letargia e sonno fisiologico.
La scoperta di H. Berger (v., 1929) che l'attività elettrica della corteccia cerebrale poteva essere registrata, nell'uomo, attraverso i tegumenti cranici (elettroencefalogramma, EEG) apriva l'era degli studi elettrofisiologici nell'uomo e negli animali, in assenza di narcosi. Lo stesso Berger diede un contributo fondamentale agli studi sul sonno quando dimostrò che l'EEG dell'uomo presenta durante questo stato oscillazioni di potenziale lente e di ampio voltaggio, ed eliminò in tal modo dalla discussione l'ingenua ipotesi che i centri superiori dovrebbero essere silenti quando manca la coscienza. Egli dimostrò inoltre che questo tipo d'attività elettrica è nettamente diverso dalle onde rapide di basso voltaggio (onde β) che caratterizzano la veglia attiva; e anche dalle lente oscillazioni di potenziale a 10/s (onde α) che compaiono durante la veglia rilassata. Infine Berger trovò che sia le onde α sia i ritmi del sonno potevano essere aboliti da stimolazioni sensitive o sensoriali e rimpiazzati dalle onde β: due fenomeni designati, rispettivamente, reazione d'arresto di Berger e risveglio elettroencefalografico. Tutti questi effetti erano generalizzati non limitati quindi all'area corticale corrispondente alla modalità sensitiva stimolata. Il loro brusco insorgere impedì allora di riconoscere i legami con i fenomeni tonici responsabili del mantenimento della veglia. Toccherà a E. D. Adrian (v., 1934) spiegare le osservazioni di Berger sulla base di modelli d'attività dei neuroni corticali. Egli suggerì l'ipotesi che i neuroni della corteccia cerebrale pulsino insieme durante il sonno e la veglia rilassata, e che i ritmi sincronizzati vengano disorganizzati (desincronizzazione) durante la veglia attiva.
Da questi primi lavori sono nate due linee di ricerca. La linea di ‛indagine fenomenologica', essenzialmente basata su osservazioni, divenne estremamente feconda con l'introduzione delle tecniche di registrazione elettroencefalografica e microelettrodica in Mammiferi senza narcosi, liberi nei loro movimenti. Essa portò alla dimostrazione che sono presenti, durante il sonno, nel gatto (v. Klaue, 1937) e nell'uomo (v. Dement e Kleitman, 1957), periodi caratterizzati da desincronizzazione dell'EEG e agli studi sui rapporti fra questa fase di sonno, chiamata sonno desincronizzato o paradosso, e i sogni (ibid.). Lungo le stesse linee di ricerca fenomenologica vanno situate le indagini sul comportamento di singole unità della corteccia cerebrale (v. Evarts, 1962 e 1964); e anche i lavori elettrofisiologici sul tono posturale (v. Jouvet, 1962), sui riflessi spinali (v. Pompeiano, 1965, 1966 e The neurophysiological..., 1967) e sulla trasmissione sensitiva durante il sonno desincronizzato (v. Pompeiano, Sensory inhibition..., 1967).
La seconda linea di ricerca, ‛l'indagine sperimentale', si basa sullo studio dei cambiamenti del ciclo veglia-sonno prodotti da lesioni o da stimolazioni. Molti aspetti di questo campo d'indagine sono stati esaminati da F. Bremer nell'articolo sistema reticolare ascendente. Basti ricordare qui le ricerche sull'encéphale isolé e sul cerveau isolé (v. Bremer, 1935, 1937 e 1938), il lavoro di G. Moruzzi e H. W. Magoun (v., 1949) sul sistema reticolare ascendente e gli esperimenti di D. B. Lindsley e altri (v., 1949 e 1950) sugli effetti di lesioni reticolari; e infine la dimostrazione dell'esistenza nella parte inferiore del tronco dell'encefalo di un sistema deattivante, probabilmente ipnogeno (v. Moruzzi, 1963, per la letteratura). Il sistema reticolare ascendente ha permesso di dare una spiegazione unitaria di osservazioni fra loro apparentemente senza rapporto, come quelle che abbiamo riferito nella prima parte di questa introduzione.
Gli sviluppi sperimentali più recenti riguardano lo studio neurochimico delle strutture del tronco dell'encefalo che controllano il ciclo veglia-sonno e l'indagine elettrofisiologica del comportamento di singole cellule nervose del ponte e della corteccia cerebrale.
2. Fenomenologia del sonno
Lo studio dei meccanismi neurofisiologici e neurochimici del ciclo sonno-veglia, che verranno trattati nei successivi capitoli, deve essere preceduto dalla descrizione dei fenomeni che si osservano quando non si fa alcun tentativo di modificare il sonno o la veglia né il loro ritmico alternarsi. ‟Si dà il nome di osservatore a colui che applica i procedimenti di investigazione semplici o complessi allo studio di fenomeni che egli non modifica e che, di conseguenza, raccoglie così come glieli offre la natura" (v. Bernard, 1865, p. 29). Questo è sempre il primo approccio in ogni campo delle scienze naturali.
Mezzo secolo fa la sola strada aperta ai ricercatori era lo studio del comportamento animale e questa linea d'attacco è tuttora seguita in molte ricerche di fisiologia comparata e in un campo delle scienze naturali, l'etologia. Il comportamento animale è basato soprattutto su movimenti o su posizioni, quindi sull'attività fasica e tonica di muscoli scheletrici e dei corrispondenti motoneuroni. Gli sviluppi moderni delle tecniche elettrofisiologiche hanno reso possibile lo studio del comportamento non solo di grandi popolazioni di neuroni cerebrali (elettroencefalografia), ma anche dell'attività di singole cellule nervose nell'animale senza narcosi, libero nei suoi movimenti. Tuttavia lo scopo della ricerca è sempre lo stesso, sia che noi studiamo il comportamento di fibre muscolari sia quello dei neuroni cerebrali. Noi osserviamo semplicemente quello che succede, senza fare alcun tentativo d'influenzare il ciclo fisiologico sonno-veglia. Naturalmente qualche esperimento, nel senso ristretto della parola (v. cap. 3), è inevitabile anche in questo tipo di ricerca. Può essere utile, infatti, porre specificamente qualche domanda alla natura, come facciamo, ad esempio, quando osserviamo gli effetti di stimoli sensitivi o sensoriali. Il punto fondamentale, tuttavia, è che non si cerca minimamente di modificare il ritmo fisiologico dell'alternanza fra sonno e veglia. Nostro scopo è solo quello di descrivere con cura fenomeni naturali e le loro relazioni nel tempo.
Questo approccio ha portato a quella che può essere chiamata la ‛fenomenologia del sonno'. Noi limiteremo la nostra esposizione a due campi di studio: a) l'alternanza fra sonno e veglia, cioè il ciclo sonno-veglia; b) l'alternarsi durante il sonno comportamentale di due stati diversi, caratterizzati rispettivamente da sincronizzazione e desincronizzazione dell'EEG.
a) Il ciclo sonno-veglia
Per ciò che riguarda il ciclo sonno-veglia gli animali possono essere divisi in due classi. Gli animali monociclici sono caratterizzati da una periodicità circadiana di circa 24 ore, mentre quelli policiclici presentano molte alternanze fra riposo e attività, in modo del tutto indipendente dall'alternanza fra giorno e notte. L'uomo adulto e molti mammiferi e uccelli sono esseri monociclici, mentre topi e ratti presentano molti cicli in un giorno. Il bambino neonato presenta un ritmo policiclico, caratterizzato da risvegli periodici legati soprattutto a necessità di nutrizione: è la veglia di necessità di N. Kleitman. L'evoluzione ontogenetica da questa periodicità primitiva al ritmo monociclico è in rapporto con lo sviluppo della corteccia cerebrale e della capacità di reagire in modo critico, mentre il tempo trascorso nel sonno diminuisce continuamente. La veglia protratta dell'uomo adulto è stata chiamata da Kleitman (v., 1963) veglia evoluta. I brevi periodi di sonno che caratterizzano i primi periodi, policiclici, di vita, si consolidano successivamente in un'unica fase di sonno notturno.
Ognuno conosce le differenze comportamentali fra sonno e veglia. Alcune manifestazioni del sonno nell'uomo che tanto hanno impressionato poeti, scultori e pittori - come la chiusura degli occhi, l'incapacità di mantenere la stazione eretta, il rilassamento dei muscoli - sono epifenomeni fisiologici. I buoi dormono con gli occhi aperti e gli Uccelli dormono appollaiati sui rami. La differenza principale fra sonno e veglia è la mancanza di coscienza durante il sonno, o la presenza di una forma rudimentale di coscienza, se prendiamo in considerazione i sogni. Se invece partiamo dal punto di vista obiettivo, comportamentale, dell'etologia, vediamo che il sonno può essere definito, negativamente, come quel periodo in cui tutti i comportamenti di veglia sono impossibili. Un animale reagisce di solito come un tutto, come un individuo, al mondo fisico o biologico che lo circonda, in particolare a certi animali. Può presentare manifestazioni tipiche di difesa o d'aggressione, o i comportamenti legati alla riproduzione o alla cura dei nati; può pulirsi o nutrirsi. Ciò che chiamiamo veglia è uno stato generale del sistema nervoso centrale che è comune a tutti questi tipi di comportamento, e ciò che chiamiamo sonno è la scomparsa, completa e del tutto reversibile, dello stato di veglia.
Va ricordato, infine, che l'approccio obiettivo dell'elettrofisiologia che è stato brevemente ricordato nell'introduzione storica ci permette lo studio dei tracciati elettroencefalografici, desincronizzati e sincronizzati, che si osservano, rispettivamente, nella veglia e nel sonno.
b) Sonno sincronizzato e sonno desincronizzato
Un altro ciclo si svolge durante il sonno: una fase caratterizzata da oscillazioni di potenziale ampie e lente (sincronizzazione dell'EFO), e inoltre da immobilità del corpo e degli occhi, si alterna con un'altra fase caratterizzata da onde rapide e di basso voltaggio (desincronizzazione dell'EEG) e inoltre da scosse cloniche degli arti e da rapidi movimenti oculari (REM: Rapid Eye Movements). La prima fase è il sonno chiamato sincronizzato o anche classico, perché per molti anni esso è stato la sola forma riconosciuta di sonno; la seconda fase è chiamata sonno desincronizzato o REM, o anche sonno paradosso, con un'espressione che indica la nostra sorpresa per il fatto che uno stadio caratterizzato, dal punto di vista comportamentale, da segni manifesti di sonno profondo possa presentare lo stesso tipo di onde rapide e a basso voltaggio che si osserva durante la veglia. In realtà questo paragone non è più giustificato, giacché J. F. Tönnies (v., 1969) ha dimostrato, con il suo analizzatore per EEG, che i tracciati sono di fatto differenti durante il sonno paradosso e durante la veglia. D'altronde anche l'attività di singole unità corticali è differente nei due tipi di sonno, come vedremo fra poco.
In condizioni fisiologiche il sonno comincia sempre con un periodo sincronizzato, che è interrotto più volte durante la notte da brevi episodi di desincronizzazione. Ciascuno di essi dura circa 6 minuti nel gatto adulto, libero nei suoi movimenti; la percentuale di sonno desincronizzato rispetto a tutto il sonno oscilla fra il 20 e il 25%. Nell'uomo adulto la percentuale varia tra il 18 e il 22%, ma può arrivare fino al 50% nel neonato (v. Roffwarg e altri, 1966).
Dal punto di vista dell'esperienza soggettiva la differenza fra i due stadi del sonno è data dal fatto che in genere abbiamo il ricordo di sogni quando siamo risvegliati durante o subito dopo un periodo desincronizzato.
Dal punto di vista obiettivo, il sonno desincronizzato è caratterizzato da molti fenomeni che colpiscono per la loro evidenza; essi meritano di essere ricordati, per quanto noi siamo ancora incapaci di comprenderne il significato fisiologico. V'è completa flaccidità o atonia di tutti quei muscoli che vengono chiamati ‛antigravitari', perché permettono le posizioni di veglia che fanno ostacolo alla forza di gravità (v. Jouvet, 1962); i riflessi spinali e i movimenti prodotti dalla stimolazione elettrica del fascio piramidale sono bloccati da scariche discendenti di impulsi inibitori provenienti dal tronco dell'encefalo (v. Pompeiano 1965, 1966, e The neurophysiological..., 1967); infine, e ancora una volta paradossalmente, queste potenti scariche inibitrici sono incapaci di bloccare l'eruzione di attività motorie fasiche, rappresentate dalle scosse cloniche degli arti e dalle esplosioni di movimenti rapidi oculari (v. Jouvet, 1967, per la letteratura).
È facile, naturalmente, vedere il significato funzionale dei movimenti osservati durante la veglia: come quelli per la difesa o l'aggressione, o in rapporto alla pulizia o alla nutrizione; ma non siamo ancora in grado di comprendere il significato delle attività fasiche che caratterizzano lo stadio desincronizzato del sonno. Forse ciò è dovuto al fatto che partiamo da un punto di vista sbagliato. Quello che importa nell'animale sveglio è il risultato finale, motore o posturale, dell'attività integrata dei muscoli scheletrici. Naturalmente ci rendiamo conto del fatto che posizioni o movimenti altro non sono che l'espressione periferica di scariche ben determinate all'interno del sistema nervoso centrale. Ma il nostro interesse converge soprattutto su questo effetto periferico, il cui significato fisiologico per la sopravvivenza dell'animale o della specie è sempre facile a comprendersi. I modelli di scarica neuronale che si hanno durante il sonno possono avere il loro significato funzionale per il ristoro. Le scosse cloniche del sonno paradosso sono fenomeni che colpiscono dal punto di vista comportamentale, ma possono essere la conseguenza di fenomeni di liberazione prodotti dall'inattività di neuroni inibitori. Anche il sonno, naturalmente, è un comportamento; ma è essenzialmente un tipo di comportamento introverso. Ciò che importa nelle ricerche sul sonno è l'attività dei neuroni, forse solo di certi tipi di neuroni; assai più dei loro effetti periferici, la contrazione delle fibre muscolari.
3. Effetti di lesioni
La parola ‛esperimento' verrà usata in senso ristretto, allo scopo di fare una netta distinzione concettuale tra la fenomenologia del sonno e i lavori sui meccanismi neurofisiologici del ciclo sonno-veglia. ‟Si dà il nome di sperimentatore a colui che usa i procedimenti di investigazione semplici o complessi per far variare o per modificare, a qualsiasi scopo, i fenomeni naturali e farli comparire in circostanze o in condizioni in cui la natura non glieli presenta. In questo senso l'‛osservazione' è l'esame di un fenomeno naturale e l'‛esperimento' è l'esame di un fenomeno modificato dall'esaminatore" (v. Bernard, 1865, p. 29). In questo capitolo riassumeremo i risultati di esperimenti in cui coma, letargia o insonnia, o qualsiasi cambiamento nel ciclo sonno-veglia, vennero ottenuti mediante lesioni nel tronco dell'encefalo o nel cervello.
a) La formazione reticolare ascendente
Questa linea di ricerca è strettamente legata alla fisiologia del sistema reticolare ascendente. Si rimanda, quindi, all'omonimo articolo di Bremer di cui riassumeremo in breve le principali conclusioni come introduzione a questa parte.
Dobbiamo partire dalla desincronizzazione elettroencefalografica prodotta da stimolazioni sensitive o sensoriali (‟reazione di arresto" di Berger, reazione di risveglio), che abbiamo ricordato nella parte storica di questo articolo. Il cambiamento del tracciato elettroencefalografico viene di solito considerato come il segno di un aumento del livello generale di attività del cervello, e il fenomeno è quindi chiamato attivazione. Moruzzi e Magoun (v., 1949) hanno dimostrato che la stessa attivazione può essere prodotta con la stimolazione elettrica della formazione reticolare del tronco dell'encefalo e hanno provato che questa influenza attivante è mediata da vie differenti da quelle dei classici sistemi sensitivi o sensoriali, che rappresentano il substrato anatomico delle percezioni. Si è visto in seguito che anche gli aspetti comportamentali della reazione di risveglio possono essere riprodotti con la stimolazione della formazione reticolare, in animali senza narcosi e liberi nei loro movimenti (v. Moruzzi, 1972, per la letteratura). Questi sono effetti fasici, di breve durata.
Il passo successivo fatto da Moruzzi e Magoun (v., 1949) fu di suggerire l'ipotesi che il sistema reticolare ascendente fosse continuamente, e cioè tonicamente, attivo e che la sua influenza sul cervello dovesse essere al di sopra di un determinato livello critico per mantenere la veglia. L'interruzione di questa influenza ascendente sarebbe la causa del coma che compare nel gatto dopo la sezione del mesencefalo, nel preparato cerveau isolé di Bremer (v. È c). Questa sindrome si osserverebbe anche nell'uomo dopo una lesione del mesencefalo prodotta da trauma. Interruzioni meno complete del mesencefalo, combinate a lesioni ipotalamiche, produrrebbero la letargia, spiegando così le osservazioni di von Economo. La dimostrazione che un'attività continua, ‛tonica', era presente nel sistema reticolare fu ottenuta seguendo due diverse linee di ricerca: a) riproduzione della sindrome di corna del cerveau isolé con interruzione delle proiezioni reticolari ascendenti (v. Magoun, 19632; v. Moruzzi, 1972, per la letteratura); b) dimostrazione mediante registrazione microelettrodica dell'esistenza di una scarica continua, irregolare, nei neuroni reticolari (v. Moruzzi, 1954, per la letteratura; v. sistema reticolare ascendente).
Un terzo passo in avanti fu compiuto da C. Batini e altri (v., 1959), quando dimostrarono che una sindrome comportamentale ed elettroencefalografica d'insonnia, opposta dunque alla sindrome di coma del cerveau isolé, poteva essere ottenuta sezionando il tronco dell'encefalo pochi millimetri all'indietro, a livello pontino. Questo ‛preparato mediopontino pretrigeminale' è caratterizzato da EEG desincronizzato e dal comportamento alerte degli occhi. Questa osservazione e molte altre fatte seguendo differenti vie di ricerca (v. Moruzzi, 1963, 1972; v. Bonvallet, 1966, per la letteratura) condussero alla conclusione che nel tronco dell'encefalo c'è anche un sistema che può essere chiamato ‛deattivante', perché è antagonista al sistema reticolare ascendente che abbiamo visto invece essere attivante. Si tratta di popolazioni di neuroni dotate di effetti sincronizzanti l'EEG e ipnogeni.
Riassumendo, alla fine degli anni quaranta appariva possibile una spiegazione unitaria di osservazioni fra loro apparentemente non legate, quali quelle di von Economo e di Berger. Questo risultato era dovuto alla dimostrazione di un sistema reticolare ascendente, dotato di un'influenza attivante, tonica e fasica, sul cervello. Alla fine degli anni cinquanta, infine, gli effetti ipnogeni classici, ottenuti con la stimolazione elettrica (v. cap. 4), potevano essere messi in qualche modo in rapporto con influenze deattivanti esercitate da altre strutture del tronco dell'encefalo.
b) Le regioni deattivanti del tronco dell'encefalo
Tutti questi risultati erano stati ottenuti con esperimenti acuti e gli effetti cronici di sezioni del tronco dell'encefalo venivano usati solo per controllo. La sperimentazione cronica cominciò a essere usata soprattutto durante gli anni sessanta. Questi esperimenti portarono a risultati importanti per il problema dell'origine del ciclo sonno-veglia. La dimostrazione che sul cervello vengono esercitate due influenze opposte condusse all'ipotesi che il ciclo stesso, e cioè l'alternarsi del sonno e della veglia, potesse nascere nel tronco dell'encefalo.
La storia degli esperimenti di decerebrazione cronica è lunga (v. Moruzzi, 1972, per la letteratura). Tuttavia noi discuteremo solo i risultati ottenuti da J. Villablanca (v., 1966) su gatti in cui il tronco dell'encefalo era stato separato dal cervello con una sezione fatta a livello più alto, e cioè proprio in fronte ai collicoli superiori (decerebrazione precollicolare). Questi gatti mesencefalici alti venivano seguiti per un lungo periodo di tempo. È naturalmente impossibile definire come sonno o come veglia qualsiasi stato osservato in animali senza cervello. Tutto quello che possiamo dire è che, dopo la decerebrazione cronica, è possibile osservare comportamenti che ricordano quelli dell'animale normale durante il sonno e la veglia. Dopo 15-20 giorni i gatti venivano trovati mentre stavano strisciando o sedevano o addirittura facevano tentativi di camminare; le palpebre erano aperte e le pupille dilatate. Questi erano manifestamente sintomi di veglia e tale impressione veniva rafforzata dal fatto che questi periodi si alternavano con stati caratterizzati da manifestazioni corporee di sonno. Villablanca (ibid.) fece una distinzione fra uno stato caratterizzato dalla chiusura delle palpebre, dal sollevamento della membrana nittitante e, soprattutto, dalla fluttuazione del diametro pupillare (miosi fluttuante) - che corrisponde al sonno sincronizzato dell'animale intatto - e uno stato caratterizzato dal restringimento estremo delle pupille (miosi fessurata) e dal collasso generalizzato del tono posturale, che corrisponde al sonno desincronizzato dell'animale intatto. I periodi caratterizzati dalla scomparsa reversibile della rigidità da decerebrazione sono di solito chiamati ‛episodi cataplessici', dal nome di una sindrome clinica che esamineremo in seguito (v. cap. 9, § a).
Il risultato principale ottenuto da Villablanca con i suoi esperimenti non è la dimostrazione che frammenti di comportamento di sonno o di veglia possono essere osservati in assenza del cervello - questo era già stato visto da altri prima di lui - ma piuttosto la dimostrazione del fatto che sia il ciclo sonno-veglia sia il ritmico alternarsi dei due stadi di sonno possono nascere quando il tronco dell'encefalo è separato dal cervello. Naturalmente solo i nervi cranici e il midollo spinale sono disponibili per le manifestazioni di sonno e di veglia, quando il cervello è assente. Ma è un risultato fondamentale l'aver dimostrato che ritmi di questo tipo possono nascere, in un animale senza cervello, nel tronco dell'encefalo.
La spiegazione ovvia di questi risultati, se si trascura il problema del sonno paradosso, è che vi sia un'alternanza di attività fra due sistemi: il sistema reticolare ascendente o attivante e le regioni deattivanti della parte caudale del tronco dell'encefalo. Ricerche recenti hanno permesso di localizzare almeno due di queste regioni deattivanti: 1) la regione del tratto solitario, che è dotata di attività fasiche, come è dimostrato da esperimenti di lesione e di stimolazione (v. Moruzzi, 1963; v. Bonvallet, 1966, per la letteratura); 2) i nuclei del rafe, che sono tonicamente attivi, come è dimostrato dall'insonnia prolungata prodotta dalla loro lesione (v. Jouvet e Renault, 1966). Si tratta di due sistemi indipendenti, e lo dimostra il fatto che è ancora possibile produrre sincronizzazione dell'FEO e miosi mediante la stimolazione di fibre afferenti vagoaortiche che si portano alla regione del fascio solitario. Questo effetto deattivante è presente anche quando l'incrociamento del rafe è stato interrotto da una sezione sagittale (v. Puizillout e Ternaux, 1974).
c) Alternanza di attività e connessioni reciproche tra sistemi antagonisti
Esperimenti cronici sul cerveau isolé mostrano, d'altra parte, che un ciclo sonno-veglia può nascere anche in un cervello isolato, dopo la sezione completa del mesencefalo. I classici esperimenti di Bremer erano acuti e il risultato principale fu la scoperta che il ciclo sonno-veglia era presente dopo la sezione del midollo cervicale a C1, e cioè quando il cervello era ancora connesso al tronco dell'encefalo (encéphale isolé), mentre il ciclo scompariva, ed era sostituito da un coma ‛permanente' dopo la sezione del mesencefalo (cerveau isolé). Manifestamente o l'abolizione del flusso di impulsi sensitivi e sensoriali attraverso i nervi cranici, secondo quanto aveva suggerito originariamente Bremer (v., 1937 e 1938), o la soppressione di un'influenza ascendente che nasce fra le due sezioni potevano spiegare differenze così evidenti fra questi due preparati acuti. Sappiamo oggi che l'eliminazione dell'influenza tonica del sistema reticolare ascendente è responsabile del coma del cerveau isolé acuto.
Il coma, tuttavia, è un sintomo prolungato, ma certo non permanente, del cerveau isolé, come è dimostrato dagli esperimenti cronici. La prima letteratura, dai lavori di U. Genovesi e altri (v., 1956) a quelli di H. L. Batsel (v., 1960 e 1964), è stata già esposta (v. Moruzzi, 1972). Qui parleremo solo degli esperimenti di Villablanca (v., 1965) sul preparato cerveau isolé alto; questa espressione indica che ogni influenza esercitata sul cervello dal tronco dell'encefalo è stata eliminata con una sezione alta (precollicolare) nel mesencefalo. In questo preparato periodi di desincronizzazione comparivano spontaneamente da 7 a 10 giorni dopo la sezione mesencefalica. Molte osservazioni e molti controlli, discussi a fondo in due rassegne critiche (v. Moruzzi, 1972 e 1974), conducono alla conclusione che in un cervello cronicamente isolato possono essere mantenuti veri periodi di veglia, e può apparire un ciclo sonno-veglia.
Le argomentazioni seguite nella discussione fatta sopra, a proposito dei risultati della decerebrazione cronica, condurrebbero all'ipotesi di un'alternanza fra strutture attivanti e deattivanti localizzate nel cervello. Gli esperimenti classici di S. W Ranson (v., 1939) sembrerebbero suggerire che le strutture attivanti siano localizzate nell'ipotalamo. Una sindrome di letargo, che ricorda la malattia del sonno della patologia umana, può essere ottenuta, nella scimmia, con lesioni dell'ipotalamo posteriore. D'altra parte, vi sono nel cervello molte regioni che producono il sonno con la stimolazione elettrica. Molte di esse, tuttavia, non hanno probabilmente importanza critica, giacché il ritmo sonno-veglia non è abolito dalla decorticazione o dalla talamectomia. Forse la struttura deattivante del cervello che più facilmente contribuisce al ritmo è l'area preottica e la parte basale del cervello anteriore, la cui lesione è seguita da insonnia (v. McGinty e Sterman, 1968; v. Moruzzi, 1972, per la letteratura).
Riassumendo, sia nel tronco dell'encefalo sia nel cervello vi sono coppie di sistemi antagonisti che producono effetti opposti sul ciclo sonno-veglia. Nel preparato cronico decerebrato un'alternanza fra i due tipi di attività è dovuta a meccanismi intrinseci al tronco dell'encefalo; nel preparato cerveau isolé cronico la stessa alternanza è dovuta unicamente a meccanismi cerebrali. Questi risultati, naturalmente, dimostrano solo una potenzialità di comportamento ritmico; tuttavia entrambi questi effetti implicano l'esistenza di connessioni reciproche fra, sistemi antagonisti. Il problema principale, ovviamente, e di sapere cosa avviene quando il tronco dell'encefalo è connesso al cervello, come nell'animale normale. Si possono fare due ipotesi. Il ciclo sonno-veglia potrebbe nascere nel tronco dell'encefalo ed essere imposto al cervello attraverso vie ascendenti. Questo è il nucleo della dottrina monoamminergica di Jouvet, che esamineremo nella parte dedicata alla neurochimica. Secondo un'altra ipotesi alternativa, il ciclo sonno-veglia potrebbe nascere nel cervello ed essere semplicemente controllato dall'afflusso di impulsi ascendenti provenienti dal tronco dell'encefalo. Naturalmente si possono verificare molte interazioni fra tronco dell'encefalo e cervello, quando l'animale è intatto. Questi sono tra i maggiori problemi della fisiologia del sonno e verranno di nuovo esaminati alla fine di quest'articolo.
4. Effetti delle stimolazioni periferiche e centrali
a) Stimolazioni periferiche
L'azione ipnogena di stimolazioni sensitive e sensoriali monotone è ben conosciuta. Pavlov aveva spesso osservato la comparsa del sonno negli esperimenti di condizionamento, ma la sua spiegazione del sonno ha ormai solo interesse storico (v. Moruzzi, 1960, 1964 e 1972) Secondo J. Konorski (v., 1967) possono condurre alla sonnolenza e al sonno sia meccanismi innati sia meccanismi condizionati. Tutti i tipi di stimolazione sensitiva e sensoriale monotona appartengono agli stimoli innati. Essi sono stati riprodotti da O. Pompeiano e J. E. Swett (v. i contributi del 1962) con la stimolazione elettrica ripetuta di fibre nervose cutanee del gruppo Il, probabilmente in rapporto a recettori del tatto, della pressione e dei peli. Anche gli stimoli condizionati sono importanti per la nascita del sonno, soprattutto nell'uomo; essi ‟si stabiliscono generalmente in rapporto al tempo, secondo le modalità stereotipe che si verificano ogni giorno, quando ci si addormenta" (v. Konorski, 1967, p 300). Le abitudini individuali riguardanti il modo di addormentarsi, tanto importanti in molti uomini, sono probabilmente in rapporto a meccanismi condizionati.
Impulsi prodotti dalla stimolazione naturale dei barocettori senocarotidei o cardioaortici (v Dell, 1971) o dalla stimolazione elettrica di fibre barocettive vagoaortiche (v. Puizillout e Ternaux, 1974; v. Puizillout e Foutz, 1977) scatenano in via riflessa il sonno. Nel preparato ‛encefalo isolato' la stimolazione di fibre afferenti barocettive vagoaortiche permette di riprodurre a volontà il sonno sincronizzato seguito da episodi di sonno desincronizzato.
È stata prospettata l'ipotesi (v. Moruzzi, 1960) che meccanismi innati, come quelli messi in moto da stimolazioni sensitive e sensoriali monotone, possano agire reclutando progressivamente i neuroni deattivanti della parte caudale del tronco dell'encefalo. I meccanismi condizionati possono interessare le aree basali del cervello anteriore e l'area preottica, come vedremo fra poco. L'effetto opposto, il risveglio, è prodotto dall'attivazione fasica del sistema reticolare ascendente, che è trascinato più facilmente da stimolazioni sensitive o sensoriali, caratterizzate dall'insorgere rapido e inaspettato. Il sistema reticolare ascendente cessa ben presto di rispondere a stimolazioni sensitive ripetute, come dimostra la scomparsa delle manifestazioni elettroencefalografiche e comportamentali del risveglio, un fenomeno chiamato ‛abituazione' (v. Sharpless e Jasper, 1956). Le strutture deattivanti si comportano in modo esattamente opposto: i loro neuroni vengono reclutati a poco a poco da stimolazioni ripetute (v. Moruzzi, 1960) con un meccanismo ben noto nella neurofisiologia classica, la ‛sommazione temporale'. (Il lettore viene rinviato all'articolo sinapsi: Fisiologia della sinapsi centrale, per ciò che riguarda i meccanismi cellulari della ‛sommazione').
b) Stimolazioni centrali
L'opera di W. R. Hess (v. i contributi in bibliografia) è l'introduzione naturale agli esperimenti di stimolazione centrale. Hess dimostrò che la stimolazione elettrica a bassa frequenza di una regione talamica situata lateralmente alla massa intermedia era seguita dalla comparsa del sonno: un genuino fenomeno fisiologico, come dimostra il risveglio che si osserva quando si mette della carne sotto al naso dell'animale. N. Tinbergen (v., 19552) ha osservato che sia il sonno naturale sia quello indotto con il metodo di Hess vengono annunziati da uno stato che ricorda la ‛fase appetitiva' che dà inizio a ogni tipo di comportamento istintivo (v. Craig, 1918): il gatto si guarda attorno, cerca un posto per andare a dormire e infine si acciambella nella posizione naturale del sonno. Manifestamente l'animale tende a raggiungere una situazione che gli permetta, o gli faciliti, l'insorgenza del sonno.
Molte critiche che erano state sollevate contro gli esperimenti di Hess sono state adeguatamente controbattute (v. Moruzzi, 1972). La sola obiezione che merita d'essere discussa, soprattutto in ragione della lunga latenza del fenomeno, è che l'animale avrebbe potuto addormentarsi anche senza stimoli. Che questa sia una causa d'errore nessuno potrebbe negare. Tuttavia gli esperimenti sulla stimolazione dell'area preottica e delle aree basali del cervello anteriore - che verranno discussi fra poco - non lasciano dubbio alcuno che il sonno può essere prodotto, con un meccanismo attivo, dalla stimolazione del cervello. Il decorso nel tempo del sonno indotto da Hess, infatti, è inusitato solo se lo si paragona alle risposte classiche della fisiologia sherringtoniana, che cominciano subito dopo l'inizio della stimolazione (tempo latente) e sopravvivono allo stimolo elettrico che le aveva provocate solo per breve tempo (scarica o effetto postumo). Anche i movimenti prodotti dalla stimolazione della corteccia motoria sono un esempio tipico di questo tipo di risposte: l'effetto periferico è guidato dagli impulsi piramidali prodotti dalla stimolazione elettrica. Tuttavia se il sonno indotto è fenomeno genuinamente fisiologico, come sembra essere, esso deve necessariamente iniziare lentamente e sopravvivere allo stimolo. Il sonno di Hess è in realtà non prodotto, ma piuttosto ‛avviato' dalla stimolazione talamica. In altre parole, le salve d'impulsi nervosi prodotte dagli stimoli elettrici mettono in moto processi nervosi innati, responsabili del comportamento di sonno, purché sia disponibile un tempo sufficiente per i lenti processi di facilitazione centrale. Quando questo risultato è stato ottenuto, probabilmente grazie a un rilassamento dell'attività tonica del sistema reticolare ascendente, il processo va avanti da sé, fino a che è interrotto dal risveglio.
L'importanza dei recenti risultati sull'influenza ipnogena della regione basale del cervello anteriore e sull'area preottica è dovuta a diversi fattori: a) il sonno può essere indotto anche da stimolazioni ad alta frequenza (v. i contributi di Sterman e Clemente, 1962); b) il tempo latente è molto più breve, anche di soli 5 s (ibid.); c) associando al solito modo la stimolazione centrale, utilizzata come stimolo incondizionato, a un tono acustico usato come stimolo condizionato, si vede che è possibile condizionare lo stimolo uditivo, dando così avvio a manifestazioni comportamentali ed elettroencefalografiche di sonno (v. Clemente e altri, 1963); d) lesioni della stessa area producono insonnia (v. McGinty e Sterman, 1968). Bremer ha dato una spiegazione di tutti questi effetti, e forse di tutti i casi di sonno indotto, con la dimostrazione che segni elettrofisiologici d'inibizione appaiono nella formazione reticolare in seguito alla stimolazione elettrica dell'area basale del cervello anteriore e dell'area preottica (v. Bremer, 1970, 1975 e 1977). Queste importanti ricerche sono esposte e documentate nell'articolo sistema reticolare ascendente.
5. Ricerche neurochimiche
a) L'approccio umorale alle ricerche sul sonno
Gli esperimenti sull'ipnotossina (v. Piéron, 1913) possono essere considerati come i precursori del moderno approccio neurochimico. Iniezioni di liquido cerebrospinale (LCS), prelevato da cani privati del sonno, dentro la cisterna magna di un altro cane, ovviamente dopo l'aspirazione di una uguale quantità di LCS, producevano un sonno comportamentale della durata di 2-6 ore. Iniezioni di controllo di LCS normale erano senza effetto. Il fattore ipnogeno venne chiamato ipnotossina. Questi risultati furono confermati da J. R. Pappenheimer e altri (v., 1967 e 1971). Perfusioni intraventricolari di LCS ottenuto da capre private di sonno eseguite su ratti prolungavano i periodi di sincronizzazione dell'EEG e deprimevano l'attività locomotoria per circa 6 ore. Altri fattori sono stati trovati da M. Monner nel plasma del sangue durante il sonno prodotto con la stimolazione elettrica del talamo (v. Jouvet, 1972, per la letteratura).
Non abbiamo finora prova alcuna che sostanze originate dalle strutture cerebrali durante la veglia raggiungano i neuroni attivanti o deattivanti del tronco dell'encefalo attraverso le vie umorali e che il loro accumulo nel sangue o nel LCS, in conseguenza di un periodo fisiologico di veglia, sia sufficiente a produrre o a mettere in marcia un sonno genuino. Le prove ora disponibili sono anzi piuttosto contrarie a tale ipotesi, almeno per ciò che riguarda il sangue, giacché non è stato trovato alcun rapporto fra i cicli di sonno-veglia di gemelli siamesi (v. Alekseyeva, 1958). Questo però non esclude che, quando s'impedisce a un animale di dormire per un lungo periodo di tempo, prodotti endogeni che normalmente eserciterebbero solo azioni locali possano passare nel sangue o nel LCS, dando origine ai fenomeni che abbiamo sopra descritto. Ma non vi sono prove che questo meccanismo entri in azione nella regolazione del ciclo veglia-sonno normale.
L'approccio umorale alle ricerche sul sonno può essere nondimeno importante, perché fornisce, in un certo senso, l'immagine ingrandita di fenomeni che in condizioni normali si svolgono solo nella sostanza grigia. I ‛mediatori chimici' sono per definizione molecole che vengono liberate, agiscono e sono distrutte localmente. Il punto centrale è sapere se dentro i sistemi regolatori del tronco dell'encefalo vi sia un accumulo progressivo di molecole ipnogene per effetto di un periodo fisiologico di veglia; se queste sostanze scompaiano per effetto del sonno fisiologico; se infine questi prodotti ipnogeni coincidano con i mediatori chimici dei sistemi deattivanti.
b) I mediatori chimici del tronco dell'encefalo
I mediatori chimici dei sistemi del tronco dell'encefalo che regolano il ciclo sonno-veglia sono stati identificati durante gli ultimi anni, soprattutto per opera di Jouvet e collaboratori. I risultati sono riassunti in rassegne sintetiche recenti, per ciò che riguarda il problema delle monoammine del cervello (v. Axelrod, 1971), con particolare riferimento al sonno (v. Jouvet, 1972).
Noi conosciamo la maggior parte dei mediatori al servizio dei sistemi che nascono nel tronco dell'encefalo, e i metodi di istofluorescenza hanno permesso di fare una mappa dei neuroni che posseggono lo stesso mediatore. Ci occuperemo in questo paragrafo solo del ciclo vegliasonno sincronizzato. Del ciclo sonno sincronizzato-sonno desincronizzato diremo nel capitolo successivo, esponendo i risultati delle moderne ricerche elettrofisiologiche eseguite su singoli neuroni. Pochi esempi di esperimenti su due sistemi monoamminergici, che utilizzano rispettivamente la 5-idrossitriptammina (5-HT) e la noradrenalina (NA) come mediatore, daranno un'idea delle potenzialità dell'approccio neurochimico, anche senza dettagliate esposizioni e discussioni.
È regola generale per tutti i sistemi monoamminergici che le molecole del mediatore siano sintetizzate dentro il soma del neurone da enzimi specifici, che siano trasportate per flusso assonico alle terminazioni del cilindrasse e che quivi siano immagazzinate, per essere liberate al momento opportuno e messe in condizione di legarsi con molecole recettrici. (Il neurone è costituito da molti dendriti, da un soma e da un assone o cilindrasse. Il soma è la parte del citoplasma che circonda il nucleo: per questa ragione si parla anche di pericari delle cellule nervose). Mentre gli enzimi e le vescicole che immagazzinano le monoammine sono sempre prodotti all'interno del soma, sembra che le terminazioni del cilindrasse possano captare le molecole precorritrici del mediatore e che la sintesi di questo avvenga anche (e forse principalmente) in esse. In ogni modo, dopo la distruzione di pericari monoamminergici, la fluorescenza specifica delle rispettive terminazioni assoniche diminuisce, per scomparire entro 8-10 giorni. Ovviamente i neuroni del tronco dell'encefalo che emettono lunghi cilindrassi ascendenti possono agire su membrane postsinaptiche di cellule nervose localizzate nel cervello; pertanto mediatori sintetizzati nel tronco dell'encefalo e liberati da impulsi nervosi che quivi prendono origine possono combinarsi con appropriate molecole recettrici situate sulle membrane postsinaptiche di neuroni del cervello, esercitando così la loro azione specifica a livello cerebrale. Queste terminazioni di vie ascendenti dal tronco dell'encefalo saranno, ovviamente, degenerate nel cerveau isolé cronico e la stessa situazione si verificherà per le terminazioni di vie cerebrali discendenti, nel preparato decerebrato cronico. Pertanto la tendenza all'attività ritmica che si osserva in questo preparato cronico sarà dovuta a neuroni i cui pericari si trovano, rispettivamente, nel cervello o nel tronco dell'encefalo. Per semplicità solo due sistemi ascendenti verranno presi in esame: i nuclei del rafe, uno dei sistemi deattivanti della parte caudale del tronco dell'encefalo, e il sistema reticolare ascendente, che è di natura attivante. (V. anche sinapsi; sistema nervoso autonomo; neurosecrezione).
5-Idrossitriptammina. - I metodi di fluorescenza istochimica mostrano che la maggior parte dei neuroni il cui soma contiene 5-HT sono localizzati nei ‛nuclei del rafe'. Queste osservazioni suggeriscono che l'azione deattivante, che abbiamo visto nascere in questi nuclei del tronco dell'encefalo, è legata alla liberazione di un mediatore chimico identificato come 5-HT.
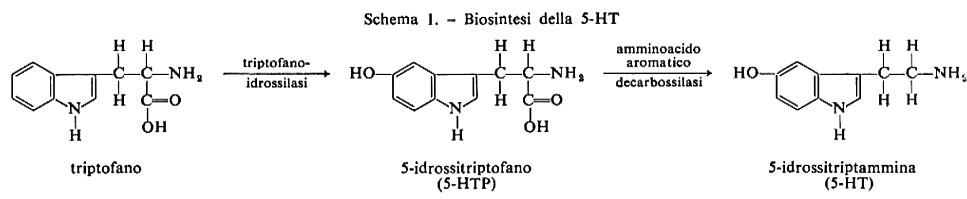
La biosintesi della 5-HT cerebrale comincia con l'assunzione dal plasma sanguigno dell'amminoacido triptofano; la prima trasformazione biochimica consiste nella 5-idrossilazione del triptofano a opera della triptofanoidrossilasi. L'amminoacido 5-idrossitriptofano (5-HTP) viene decarbossilato dando origine alla 5-idrossitriptammina (5-HT), una indolammina che è probabilmente il mediatore chimico. Lo schema 1 dà le tappe della biosintesi della 5-HT.
Dopo iniezione di p-clorofenilalanina, che blocca la triptofanoidrossilasi, si ha nel cervello una diminuzione della 5-HT. Una sola iniezione di 400 mg/kg di questa sostanza, nel gatto, è senza alcun effetto sul ciclo sonno-veglia durante le prime 18-24 ore. Ma questo periodo iniziale è seguito dalla diminuzione del sonno e quindi dalla completa insonnia. Il sonno riappare solo dopo 40 ore, ma il normale ciclo sonno-veglia ritorna solo dopo 2.000 ore (v. Jouvet, 1972).
Il fatto che l'insonnia cominci con un ritardo di circa un giorno fa pensare che la p-clorofenilalanina non abbia un'azione diretta sul ciclo sonno-veglia, che verrebbe alterato solo come conseguenza dell'esaurimento delle riserve di 5-HT nel sistema deattivante del rafe. Ulteriori prove del significato fisiologico di queste osservazioni sono date dalla correlazione, nel ratto, fra la diminuzione del sonno e il calo della 5-HT cerebrale e dall'osservazione che l'insonnia viene sostituita, nel gatto, da un normale ciclo sonno-veglia se l'azione di blocco è aggirata con iniezione di 5-HTP (v. Jouvet, 1972).
Noradrenalina. - I metodi di fluorescenza istochimica dimostrano che i pericari del tronco dell'encefalo che contengono NA sono in massima parte localizzati nel tegmento del mesencefalo, ma anche nel ponte e nel bulbo, e cioè nei luoghi d'origine del sistema reticolare ascendente. Questa osservazione fa pensare che la liberazione di un mediatore chimico identificato come NA sia legata all'azione attivante che il tronco dell'encefalo esercita sul cervello.
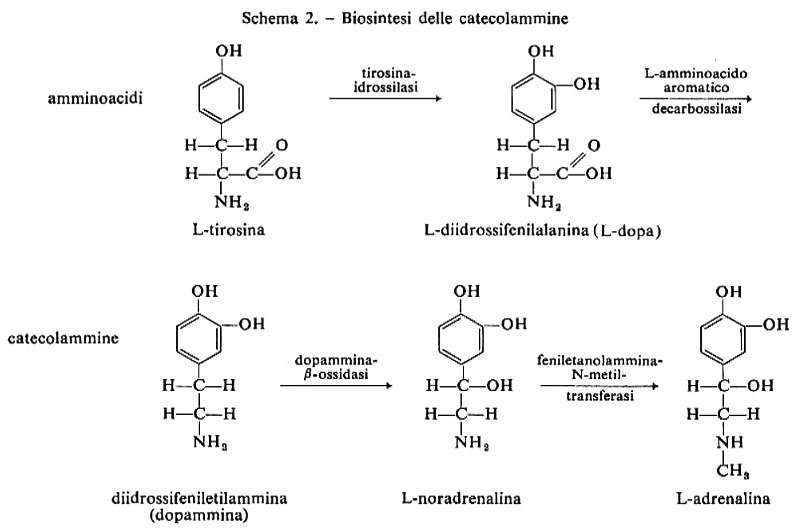
La biosintesi della NA comincia con l'assunzione dell'amminoacido tirosina dal sangue circolante. La prima trasformazione biochimica è rappresentata dalla metaidrossilazione a L-diidrossifenilalanina (L-dopa) per opera della tirosinaidrossilasi. La decarbossilazione di questo amminoacido, per opera della dopadecarbossilasi, porta alla formazione della prima catecolammina, la 3,4-diidrossifeniletilammina o dopammina (DA), che è il mediatore chimico per alcuni neuroni. La DA viene trasformata in un'altra catecolammina, la L-noradrenalina, da una dopammina-β-ossidasi, che è localizzata nelle vescichette sinaptiche in cui s'immagazzina la NA. Un'ulteriore trasformazione, che per altro avviene soprattutto nelle cellule secretorie della midolla surrenale, è la conversione nella catecolammina terminale, la L-adrenalina, per opera di una feniletanolammina-N-metiltransferasi Lo schema 2 dà le tappe della biosintesi della NA.
c) La dottrina monoamminergica del ciclo sonno-veglia
Questi che abbiamo citati sono solo pochi esempi dei risultati che possono essere ottenuti con l'approccio neurochimico. In complesso la natura 5-idrossitriptamminergica del sistema deattivante dei nuclei del rafe e la natura noradrenergica del sistema attivante reticolare sembrano assodate. La ‛teoria monoamminergica' di M. Jouvet (v., 1972) parte dal presupposto che l'alternanza fra veglia e sonno sincronizzato sia dovuta a un ciclo di attività che nasce nel tronco dell'encefalo, essendo legato al risultato di un conflitto fra i due sistemi monoamminergici antagonisti. L'attività dei neuroni cerebrali modificati dalla veglia e dal sonno si limiterebbe a registrare l'esito d'una lotta che è decisa a livello del tronco dell'encefalo.
6. Registrazioni elettrofisiologiche da singoli neuroni del tronco dell'encefalo
I metodi di lesione e di stimolazione e le indagini neurochimiche portano alla conclusione che il ciclo veglia sonno e le fasi stesse del sonno, sincronizzata e desincronizzata, sono legati al ritmico alternarsi dell'attività di strutture nervose localizzate nel tronco dell'encefalo e nel diencefalo. In questi ultimi anni i progressi della tecnica hanno permesso di registrare la scarica di singoli neuroni nell'animale senza narcosi e libero nei suoi movimenti e quindi hanno consentito di mettere in rapporto la scarica unitaria con il comportamento, con l'elettroencefalogramma e con l'elettromiogramma durante la veglia e le fasi sincronizzata e desincronizzata del sonno. Non tutte le strutture nervose il cui significato funzionale è stato chiarito con i metodi di lesione o di stimolazione sono state studiate a livello cellulare, ma per alcune di esse i progressi conseguiti sono tali da giustificare un'esposizione a parte dei risultati di questo nuovo metodo d'indagine.
Abbiamo visto che il ciclo veglia-sonno è legato all'attività di strutture antagoniste, dette attivanti e deattivanti. Recentemente M. Steriade e altri (v., 1980) sono riusciti a registrare le scariche di singoli neuroni del tegmento mesencefalico e a dimostrarne con metodi fisiologici l'appartenenza al sistema reticolare ascendente. La scarica di queste cellule nervose si accelera infatti quando si passa dal sonno sincronizzato alla veglia, mentre diminuisce poco prima dell'inizio del sonno sincronizzato. Il passo successivo sarà verosimilmente quello di analizzare a livello cellulare le strutture deattivanti localizzate nelle regioni basali del cervello anteriore, nel rafe e nella regione del tratto solitario.
Le nostre conoscenze sono molto più soddisfacenti per ciò che riguarda lo studio a livello cellulare delle strutture pontine che assicurano l'alternarsi delle fasi sincronizzata e desincronizzata del sonno.
Jouvet (v., 1962) aveva dimostrato che la distruzione bilaterale e simmetrica della parte rostrale del nucleus reticulapis pontis caudalis e della parte caudale del nucleus reticularis pontis oralis sopprimeva il sonno paradosso. I neuroni ‛generatori' di questa fase del sonno si trovano dunque nel tegmento pontino. Hobson e altri (v., 1975), con ricerche sul gatto, hanno dimostrato che nel nucleo tegmentale gigantocellulare del ponte esistono neuroni la cui frequenza di scarica aumenta selettivamente durante il sonno desincronizzato. Altre ricerche eseguite da Pivik e altri (v., 1977) hanno dimostrato che i movimenti rapidi oculari che caratterizzano il sonno desincronizzato sono preceduti, e verosimilmente causati, da uno spiccato aumento selettivo della scarica degli stessi neuroni. Le cellule nervose del nucleo tegmentale gigantocellulare del ponte sono dunque i generatori delle manifestazioni toniche e fasiche del sonno desincronizzato. Questo non significa, naturalmente, che i predetti neuroni non svolgano la loro azione anche durante la veglia.
Il problema che ora si pone è quello del meccanismo fisiologico che dà il via alla scarica dei neuroni del nucleo reticolare pontino e quindi alla comparsa delle manifestazioni toniche e fasiche del sonno desincronizzato. Hobson e altri (v., 1975) hanno trovato, nel locus coeruleus e nel nucleus subcoeruleus, neuroni che hanno un comportamento reciproco a quello del nucleo gigantocellulare. Durante il passaggio dal sonno sincronizzato a quello desincronizzato la loro frequenza di scarica si riduce, e questo avviene proprio mentre la scarica dei neuroni del nucleo gigantocellulare si accelera. Queste osservazioni suggeriscono l'ipotesi che per queste ultime cellule nervose si verifichi in quel momento la liberazione da un freno inibitore.
Esperimenti recenti fanno ritenere che i neuroni del nucleo gigantocellulare siano colinergici, mentre quelli del locus coeruleus e del nucleus subcoeruleus sarebbero noradrenergici. T. T. Amatruda e altri (v., 1975) hanno dimostrato infatti che l'iniezione locale nelle regioni del nucleo gigantocellulare di un agente colinergico a lunga durata, il carbacolo, aumenta fortemente le manifestazioni del sonno paradosso del gatto normale, quali l'atonia muscolare e la desincronizzazione del tracciato elettroencefalografico. Se il gatto è stato decerebrato si possono avere solo le manifestazioni del sonno paradosso legate a scariche discendenti dei neuroni generatori pontini. Hoshino e Pompeiano (v., 1976) hanno dimostrato che l'iniezione endovenosa di un farmaco anticolinesterasico, quale l'eserina, fa comparire nel gatto decerebrato acuto fenomeni cataplessici caratterizzati da atonia, sempre preceduti da un aumento rilevante delle scariche dei neuroni del nucleo gigantocellulare. Quando la rigidità ricompare, spontaneamente o per effetto di stimoli cutanei, essa si accompagna a una marcata diminuzione della frequenza di scarica di questi neuroni. Pompeiano e Hoshino (v., 1976) hanno anche osservato che neuroni del nucleo del locus coeruleus si comportano in modo reciproco, ossia riducono la loro frequenza di scarica durante gli episodi cataplessici: quando la rigidità ricompate, la scarica dei neuroni del locus coeruleus si intensifica di nuovo. Questi esperimenti e altri ricordati nei lavori originali fanno ritenere che i neuroni del nucleo gigantocellulare siano colinocettivi e colinergici e scatenino con la loro scarica l'insorgenza delle manifestazioni del sonno paradosso, e che invece i neuroni del locus coeruleus, colinocettivi ma noradrenergici, inibiscano periodicamente i neuroni generatori del nucleo gigantocellulare.
7. Il significato funzionale del sonno
Fino a pochi anni or sono il sonno veniva considerato come un fenomeno omogeneo. Si diceva che compito del sonno era di ristabilire, con meccanismi che potevano essere genericamente definiti ‛di ristoro', le condizioni che si avevano nel cervello all'inizio del precedente periodo di veglia. La teoria del ristoro presupponeva che durante la veglia si verificasse un logoramento nei centri nervosi (v. Sherrington, 1946, p. 252). Si trattava forse di alterazioni limitate a un gruppo o ad alcuni gruppi di neuroni, ma la loro caratteristica fondamentale era che esse richiedevano un lungo tempo (alcune ore) per essere riparate.
Sappiamo ora che vi sono due periodi di sonno, profondamente diversi nelle loro manifestazioni e nel loro significato funzionale. Questo diverso significato funzionale è dimostrato dal fatto che nella notte successiva a quella in cui un soggetto era stato svegliato ogniqualvolta il tracciato elettroencefalografico si desincronizzava, e quindi dopo una privazione selettiva del sonno paradosso, la percentuale di tempo trascorsa nel sonno desincronizzato si accresceva. L'aumento compensatorio del sonno paradosso dimostra che questa fase di sonno ha un significato funzionale suo proprio. Le azioni di ristoro svolte durante il sonno desincronizzato non possono essere sostituite da meccanismi fisiologici che si svolgono in altre fasi del sonno.
Esamineremo separatamente le teorie del ristoro ipnico e in seguito il significato del sonno desincronizzato.
a) Le teorie del ristoro ipnico
Pressoché tutte le attività del cervello e del corpo sono influenzate dal ciclo sonno-veglia. Tuttavia alcune manifestazioni corporee del sonno sono chiaramente epifenomeni, non particolarmente interessanti per ciò che riguarda il problema fisiologico principale. Il rilassamento muscolare è assente nel treno anteriore e nel collo del bestiame bovino. La chiusura degli occhi è estremamente rara nel bestiame bovino, e d'altra parte brevi manifestazioni di lagoftalmo sono state descritte persino nell'uomo. L'abolizione dei riflessi di raddrizzamento, infine, non compare durante il sonno degli Uccelli, come è dimostrato dal fatto che questi animali possono dormire appollaiati su un ramo senza mai cadere. Possiamo terminare con una semplice considerazione. Quando giaciamo insonni su un letto, sappiamo perfettamente che scopo principale del sonno non è dare un periodo di riposo al nostro corpo.
Scopo del sonno è il ristoro del sistema nervoso centrale. Ma occorre fare una distinzione fra i neuroni la cui attività è modificata come conseguenza dello stato di sonno e quelli il cui ristoro è il principale compito del sonno. Il sonno influenza fortemente, per esempio, il respiro, il ritmo cardiaco, il tono dei muscoli scheletrici; ma tutti sappiamo che il ristoro del centro respiratorio o dei neuroni cardioregolatori o del cervelletto o del midollo spinale non rappresenta lo scopo principale del sonno. In realtà sintomo principale del sonno è la mancanza di coscienza o la sua grave alterazione. Come giustamente ha messo in evidenza Evarts (v., Relation of cell..., 1965, p. 89) ‟le vivide esperienze dei sogni non lasciano tracce permanenti nella memoria, a meno che il soggetto non sia svegliato durante il sogno o poco dopo. Pertanto le scariche neuroniche necessarie per la percezione immediata permangono durante il sogno, mentre invece cessano i processi necessari al consolidamento delle tracce della memoria". E, infine, è un fatto ben conosciuto che i sintomi prodotti dalla privazione del sonno appartengono alla sfera psichica (v. Kleitman, 1963, pp. 215-229).
È stata operata una distinzione fra processi rapidi e lenti di ristoro delle attività nervose. I processi di ristoro rapidi sono in rapporto alla conduzione e alla trasmissione sinaptica degli impulsi nervosi, e quindi, essenzialmente, alla permeabilità agli ioni della membrana (v. elettrofisiologia; v. sinapsi). Questo è il compito principale del ristoro nel cervelletto, ad esempio, o nel centro respiratorio bulbare o nel midollo spinale. I processi di ristoro lenti, invece, occupano una parte molto maggiore della vita, almeno in molti Mammiferi e negli Uccelli: quasi un terzo della vita umana viene speso nel sonno. Un andamento nel tempo tanto differente deve essere in rapporto a meccanismi del tutto differenti di ristoro. È stata prospettata l'ipotesi che il sonno non riguardi tutto il cervello, e neppure tutta la neocorteccia (la parte filogeneticamente nuova della corteccia cerebrale, che contiene aree di proiezione - motorie e sensitive - e di associazione), ma solo quei neuroni e quelle sinapsi, e forse quelle cellule gliali, che durante la veglia sono responsabili delle funzioni cerebrali a cui è dovuto il comportamento cosciente, o sono in qualche modo in rapporto con esse (v. Moruzzi, 1972, p. 121). Cambiamenti di natura plastica, probabilmente macromolecolare, è verosimile avvengano durante la veglia nelle strutture nervose, in rapporto con i processi di apprendimento.
I. Feinberg ed E. V. Evarts (v., 1969, p. 338) hanno giustamente rilevato che l'idea di una funzione positiva del sonno ‟implica la nozione di complementarità, e cioè che la quantità e l'intensità del sonno debbano essere in rapporto all'intensità di quei processi di veglia che richiedono, appunto, un'azione di completamento da svolgersi durante il sonno". Tra le funzioni positive che riconoscono al sonno, anche in base ai loro esperimenti di psicofisiologia nell'uomo, essi citano non solo il consolidamento degli engrammi, ma anche la scomparsa di certe categorie di memorie. Questa è indubbiamente un'ipotesi stimolante, se si tiene presente l'importanza ben conosciuta dell'oblio selettivo dei ricordi non significativi. È chiaro che la parola ‛ristoro' ha significati differenti per i muscoli scheletrici e per il cervello. Non può essere scopo del sonno il ristabilire o il restaurare esattamente le stesse condizioni che erano presenti all'inizio del precedente periodo di veglia: ciò impedirebbe l'apprendimento. L'ottenere condizioni costanti mediante il riposo può essere possibile per le fibre muscolari, non per il cervello, anche qualora si prescinda dal problema della possibilità di periodi caratterizzati dall'assenza di attività specifica nel cervello, cosa invece che accade nel caso delle fibre di un muscolo scheletrico completamente rilassato.
D'altra parte la durata del sonno non ci dà una misura soddisfacente dell'ammontare dei processi neurochimici che si svolgono in esso. Ciò è dimostrato dal fatto che la sindrome imponente prodotta in un uomo da un'insonnia prolungata per ben 200 ore può scomparire pressoché completamente solo dopo 13 ore di sonno (v. Luce, 1965, p. 20). Non v'è dubbio, inoltre, che molti animali, quali il gatto domestico, dormano in eccesso rispetto ai loro bisogni. Secondo Jouvet (v., 1972) questa mancanza di una relazione lineare fra le durate della veglia e del sonno riguarda solo la fase caratterizzata da EEG sincronizzato, che sarebbe funzione del log della durata della veglia. La durata del sonno paradosso, invece, è direttamente proporzionale alla durata della veglia, nella misura in cui la veglia implica deprivazione di sonno paradosso. Abbiamo detto sopra che la privazione selettiva della fase paradossale è seguita da un debito specifico per questa fase del sonno, che può essere pagato solo con un aumento della durata del tempo trascorso nei periodi di sonno desincronizzato durante la notte successiva.
Durante il periodo di ristoro il muscolo scheletrico è rilassato; esso quindi è inattivo, almeno per ciò che riguarda la sua funzione specifica, la contrazione. Per analogia si potrebbe pensare che durante il sonno i neuroni della corteccia cerebrale, dediti alle complesse operazioni metaboliche necessarie al ristoro, siano essi pure fisiologicamente inattivi e quindi non scarichino impulsi. Questa era effettivamente la concezione di Pavlov, che presupponeva un silenzio generalizzato della corteccia cerebrale durante il sonno. L'obliterazione della coscienza, che caratterizza il sonno, poteva spiegarsi con l'inattivazione funzionale del mantello neocorticale. La registrazione elettrofisiologica di singoli neuroni della corteccia cerebrale ha reso insostenibile questa ipotesi. Sappiamo con certezza che i neuroni corticali sono spontaneamente attivi durante il sonno, anche se la loro attività appare nettamente diversa da quella che si osserva in condizioni di veglia.
b) Significato del sonno desincronizzato
Evarts è stato il primo a registrare sistematicamente durante le varie fasi della veglia e del sonno le scariche dei neuroni della corteccia visiva e anche di quelli della corteccia motoria che danno origine al fascio piramidale (v. Evarts, 1962, 1964 e 1965). Si sapeva da tempo che i neuroni motori corticospinali sono tonicamente attivi durante la narcosi barbiturica, e quindi in assenza di movimenti (v. Adrian e Moruzzi, 1939); la scarica d'impulsi piramidali diventa sopraliminare per i motoneuroni spinali solo quando raggiunge una frequenza critica e solo allora appaiono i movimenti. Nella scimmia senza narcosi, Evarts (v., 1964) ha dimostrato che i neuroni del fascio piramidale non sono affatto inattivi durante le fasi sincronizzata e desincronizzata del sonno, ma semplicemente si scaricano in modo diverso rispetto alla veglia rilassata. Le modalità di scarica che si osservano in questi esperimenti di Evarts non hanno però validità generale, a differenza di quanto si riteneva un tempo. Lavori successivi di G. Berlucchi (v., 1965) e di L. M. Mukhametov e altri (v., 1970), e infine diverse ricerche ricordate da Steriade (v., 1978) hanno dimostrato che per altri tipi di neuroni le modalità di scarica durante le fasi del sonno sono diverse. V'è però un punto fondamentale su cui tutti sono d'accordo: una scarica cellulifuga è sempre presente nel sonno in tutti i neuroni corticali studiati in queste condizioni sperimentali.
Resta il problema di vedere come il ristoro possa intervenire se i neuroni che dovrebbero beneficiarne sono sempre attivi. In un tentativo di risolvere queste difficoltà, Evarts (v., 1964) e Moruzzi (v., 1966) avevano suggerito che questa incessante attività fosse limitata ai soli neuroni del I tipo, a cilindrasse lungo. I neuroni del Il tipo o interneuroni, che assicurano i circuiti nervosi all'interno della sostanza grigia, erano sfuggiti fino allora allo studio microelettrodico, almeno nelle condizioni sperimentali. Queste cellule nervose avrebbero potuto essere silenti durante il sonno. Recentemente Steriade e collaboratori sono riusciti a identificare entrambi i tipi di neuroni nella corteccia cerebrale e a registrarne separatamente l'attività nell'animale senza narcosi, libero nei suoi movimenti (v. Steriade, 1978). Essi hanno visto che la scarica dei neuroni corticali del Il tipo è addirittura maggiore nel sonno sincronizzato rispetto alla veglia. Questi risultati rimettono in discussione tutte le teorie sui meccanismi nervosi che si svolgono nella corteccia cerebrale durante il sonno. È certo, in ogni caso, che durante il sonno le cellule nervose corticali non riposano, ma presentano un'organizzazione diversa della loro attività.
Sembra dunque chiaro che durante il sonno non si possano avere fenomeni di ristoro secondo lo schema classico del ristoro del muscolo scheletrico. Almeno per il sonno desincronizzato sembra addirittura che esso sia necessario per permettere attività del sistema nervoso che nulla hanno a che fare con i fenomeni di ristoro, nel senso che in fisiologia si dà a questa parola. Secondo Jouvet (v., 1978) il sonno desincronizzato, con i suoi potenti meccanismi inibitori che isolano il cervello dalla periferia motoria e forse (in parte almeno) dalla periferia sensitiva, potrebbe fornire un quadro ideale per una programmazione genetica e periodica del cervello. J. P. Sastre (v., 1978) ha continuato le ricerche di Jouvet e Delorme (v., 1965) sugli effetti di lesioni bilaterali delle strutture nervose responsabili della comparsa delle manifestazioni inibitrici negli episodi di sonno desincronizzato. In queste condizioni vengono alla luce i comportamenti motori corrispondenti ai sogni che, come è noto, compaiono nell'uomo durante il sonno desincronizzato. L'animale sembra allora cercare un oggetto immaginario o combattere un nemico immaginario. Il cervello eccita dunque, direttamente o indirettamente, le sinapsi, ‛provando' in tal modo i circuiti sensitivi e motori messi in gioco (durante la veglia) nei comportamenti innati. La soppressione dei fenomeni inibitori porta alla luce gli effetti motori della scarica programmata che agiscono sui motoneuroni spinali. In condizioni normali solo frammenti di attività motoria sfuggono all'inibizione e possono venire alla luce. Il resto dell'attività onirica dei centri motori è bloccato dalle scariche inibitrici che discendono al midollo spinale dal tronco dell'encefalo. Il sogno si riduce così, in gran parte almeno, ad allucinazioni visive, con scarse manifestazioni motorie.
8. Sonno e comportamento istintivo
La regolazione del ciclo sonno-veglia è dovuta a meccanismi nervosi innati, che possono essere modificati sia da impulsi incondizionati sia dal condizionamento (v. condizionamento, meccanismo del).
Molti etologi (v. Holzapfel, 1940; v. Tinbergen, 1952 e 19552; v. Hediger, 1959 e 1969) hanno rilevato che il comportamento degli animali nel periodo che precede il sonno ricorda la fase appetitiva che preannunzia ogni tipo di comportamento istintivo.
Ai fini del presente articolo possiamo trascurare qui il problema delle componenti acquisite del comportamento animale o quello della differenza fra attività istintive e attività puramente riflesse (v. istinto).
Secondo Tinbergen (v., 1952) il sonno potrebbe essere considerato come la fase finale, o atto consummatorio, di un comportamento istintivo. Come ha osservato W. Craig (v., 1918), quando il comportamento istintivo è stato completato con un atto consummatorio, l'animale appare ‛sazio' o ‛soddisfatto'.
Questi concetti etologici sono stati sviluppati da un punto di vista neurofisiologico. È stato detto (v. Moruzzi, 1969) che il sonno, con le sue fasi sincronizzata e desincronizzata, può essere piuttosto paragonato a una catena di atti consummatori. Questa catena di risposte inizierebbe sempre con la fase caratterizzata da sincronizzazione dell'EEG, non appena l'attività tonica del sistema reticolare ascendente cade al di sotto del livello critico.
In tutti i tipi di comportamento istintivo che si osservano durante la veglia, la fase consummatoria è caratterizzata da movimenti stereotipati; essi implicano l'esistenza di modelli neuronici di scarica stereotipata che regolano, nello spazio e nel tempo, l'attività dei motoneuroni corrispondenti. Scariche stereotipate di neuroni cerebrali caratterizzano le due fasi di sonno, peraltro in modo del tutto indipendente dagli effetti periferici sui muscoli.
Dal punto di vista del tema di quest'articolo abbiamo la scelta fra due ipotesi: 1) vi potrebbe essere un rapporto di causalità tra la scarica stereotipata di neuroni corticali e il compito fondamentale del sonno, il ristoro del cervello. Abbiamo definito questo modello di comportamento neuronico un tipo introverso di comportamento, per mettere l'accento sulla differenza fondamentale rispetto al comportamento di veglia. Nell'animale sveglio lo scopo principale della scarica dei neuroni è la produzione di determinati movimenti, e cioè di effetti periferici di cui è facile comprendere il significato rispetto al mondo che circonda l'animale. I modelli di scarica stereotipata che si hanno nel sonno sarebbero necessari, per ragioni ancora sconosciute, allo scopo di avere il ristoro ipnico, o di assicurare la programmazione periodica dei circuiti nervosi, mentre i loro effetti periferici, quali le scosse cloniche della fase paradossa, sarebbero solo epifenomeni; 2) vi potrebbe anche essere un'associazione fra manifestazioni del sonno, di natura istintiva, e i processi neurochimici e neurofisiologici di ristoro, che di per sé non sarebbero istintivi. La situazione sarebbe analoga all'associazione fra il comportamento istintivo che porta all'assunzione di acqua e i meccanismi neuroendocrini, puramente fisiologici, che regolano la diuresi. Due tipi diversi di attività, un comportamento istintivo e meccanismi neurochimici o neuroendocrini, potrebbero convergere nella produzione dello stesso risultato: rispettivamente, il ristoro ipnico dei neuroni cerebrali e la costanza della pressione osmotica nel mezzo interno.
Indubbiamente il sonno è una forma unica di regolazione fisiologica, perché implica la perdita temporanea della coscienza o un suo forte indebolimento (se si considerano i sogni). Tutte le forme conosciute di comportamento istintivo scompaiono durante il sonno, e quindi gli animali devono adattare il ciclo sonno-veglia alle loro esigenze per ciò che riguarda la difesa o l'approvvigionamento di cibo o acqua. Di conseguenza ogni animale deve pagare un forte scotto allo scopo di beneficiare degli effetti del sonno, come appare chiaro a chi consideri cosa può apportare la perdita della coscienza negli erbivori allo stato selvaggio. Pertanto la regolazione dei periodi dedicati al sonno ha un grande valore per la sopravvivenza, e non è sorprendente che tale controllo diventi parte di un modello di attività istintiva. Esso è infatti il risultato di un conflitto fra gli istinti della veglia, da una parte, e il comportamento istintivo del sonno dall'altra. Le differenze tanto marcate fra i vari Mammiferi, per ciò che riguarda il ritmo sonno-veglia, possono essere spiegate in questo modo. È improbabile che il fatto che in molti erbivori si osservano rari e brevi episodi di sonno, mentre il gatto può dedicarvi il 72% del suo tempo, possa essere spiegato da differenze nell'esigenza di ristoro ipnico (v. Moruzzi, 1972, per la letteratura etologica).
Queste considerazioni portano a una concezione più ampia del significato fisiologico del controllo del cervello da parte del tronco dell'encefalo (ibid.). I risultati delle ricerche sul sonno, in grande sviluppo nell'ultimo quarto del secolo, potrebbero dare l'impressione che compito principale dei sistemi ascendenti del tronco dell'encefalo sia il controllo del ciclo sonno-veglia. Ma in realtà appare improbabile che siano richieste tante strutture nervose per un compito comparativamente così semplice.
9. Interpretazioni neurofisiologiche nella patologia del sonno
Le principali sindromi della patologia del sonno possono ora essere reinterpretate in base alle nostre conoscenze di fisiologia reticolare.
a) Narcolessia e cataplessia
La narcolessia è una malattia caratterizzata da un bisogno irresistibile di dormire. Ricerche elettroencefalografiche (v. Dement e Rechtschaffen, 1968) hanno dimostrato che si tratta di casi di sonno paradosso, non preannunciati da una fase sincronizzata. La cataplessia, invece, è caratterizzata da episodi brevi (da 3 a 5 minuti) di atonia completa, senza perdita di coscienza. Una scarica improvvisa del centro pontino responsabile degli episodi paradossi potrebbe spiegare entrambi i fenomeni. Durante la cataplessia, tuttavia, sarebbe presente, per ragioni ignote, solo l'influenza discendente inibitrice esercitata dal tronco dell'encefalo sul midollo spinale (v. Pompeiano, 1965 e The neurophysiological..., 1967). La sensazione di essere paralizzati durante gli incubi notturni e la paralisi da sonno che si osserva al momento del risveglio potrebbero essere pure in rapporto con episodi di sonno paradosso, o con una scarica postuma delle salve d'impulsi discendenti inibitori di solito legate a questi episodi. L'ipotesi è suggerita dall'arresto dei movimenti prodotti dalla stimolazione elettrica del fascio piramidale, fenomeno che si osserva costantemente nel gatto durante gli episodi di sonno desincronizzato (v. Marchiafava e Pompeiano, 1964).
b) Letargo della malattia del sonno e coma
Il letargo che si osserva nella malattia del sonno (encefalite letargica epidemica) differisce nettamente dal coma, perché il paziente può essere facilmente risvegliato, e allora appare ben orientato rispetto all'ambiente. La sindrome è stata riprodotta nella scimmia con lesioni ipotalamiche (v. Ranson, 1939).
Nei casi clinici questo risultato è probabilmente dovuto alla distruzione parziale del tegmento del mesencefalo, che determina un minore afflusso di impulsi reticolari attivanti combinato alla lesione delle strutture ipotalamiche in rapporto con la veglia. Il coma è uno stato caratterizzato dalla completa perdita di coscienza, che non può essere interrotto neppure con le più forti stimolazioni sensitive o sensoriali (v. coscienza: Alterazioni della coscienza). Esso compare ogni qualvolta il sistema reticolare ascendente è interrotto, a livello del mesencefalo, da traumi o tumori, o è inattivato funzionalmente da dosi narcotiche di barbiturici. Questo stato è riprodotto sperimentalmente nel preparato cerveau isolé (v. Bremer, 1935, 1937 e 1938). H. Orthner (v., 1969) ha dimostrato che le sindromi sperimentali sia del cerveau isolé sia del preparato pretrigeminale sono riprodotte nella patologia umana. (Per la patologia del sonno v. Kleitman, 1963; v. Orthner, 1969; v. Otter, 1970; per la farmacologia del sonno, v. Holm, 1970).
c) Le insonnie
Gli sviluppi recenti della neurochimica hanno gettato luce sulla patologia e sulla farmacologia delle insonnie. Di notevole interesse è la marcata insonnia osservata da Jouvet e collaboratori in seguito alla distruzione dei nuclei del rafe (v. cap. 3) o al blocco farmacologico della biosintesi di 5-HT (v. cap. 5). Oltre che dalla depressione del sistema deattivante indolamminergico, l'insonnia può risultare anche dall'iperattività primaria dei neuroni attivanti, catecolamminergici. Un esempio farmacologico di questo tipo di squilibrio del ciclo sonno-veglia è rappresentato dall'azione dell'anfetammina. Il lettore viene rinviato alla rassegna critica di Jouvet (v., 1972) per la discussione e per l'esposizione di molti altri effetti neurochimici e neurofarmacologici sul sonno e sulla veglia.
10. Stati simili al sonno
Cercheremo ora di riassumere brevemente i principali dati fisiologici sull'ipnosi animale e sull'ibernazione. Entrambi questi fenomeni sono nettamente differenti dal sonno fisiologico. Tuttavia in essi potrebbero intervenire alcuni aspetti dei meccanismi neurofisiologici e neurochimici interessati nel ciclo sonno-veglia. In particolare vedremo che il sistema inibitore discendente, attivo durante la fase paradossale del sonno fisiologico, può essere selettivamente messo in azione durante l'ipnosi animale, mentre l'ibernazione potrebbe essere messa in rapporto all'arresto del turnover di catecolammine nel sistema reticolare ascendente. Pertanto una breve analisi di tali problemi può essere giustificata alla fine di questo articolo.
a) L'ipnosi animale
L' ipnosi animale è caratterizzata dall'assenza di movimenti e dall'abolizione dei riflessi di raddrizzamento. Viene facilmente ottenuta ponendo sul dorso conigli, galline o altri animali che, immobilizzati per un tempo molto breve, rimangono senza muoversi nella nuova, inusitata posizione. La vecchia letteratura su questo argomento - e anche quella sull'ipnosi umana, tanto meno soddisfacente dal punto di vista scientifico - è data da Kleitman (v., 1963), mentre la letteratura più moderna si trova in un lavoro di G. Carli (v., 1971).
Risultati contraddittori sono stati ottenuti con studi elettroencefalografici sui conigli (v. Gerebtzoff, 1941; v. Svorad, 1957), ma sembra comunque assai improbabile che l'ipnosi animale possa essere paragonata a una forma qualsiasi di sonno (v. Klemm, 1966). Per ciò che riguarda il midollo spinale, invece, è stato dimostrato nel coniglio che i comuni riflessi sono molto depressi e che questi effetti non trovano rispondenza nel tracciato dell'EEG (v. Carli, 1969). Si è visto inoltre che l'ipnosi animale può ancora essere ottenuta, sempre nel coniglio, dopo decerebrazione precollicolare (v. Carli, 1971).
P. Buser e G. Viala (v., 1968) hanno avanzato l'ipotesi che l'arresto dei movimenti sia dovuto all'influenza inibitoria del sistema reticolo-spinale di Magoun, che probabilmente è anche responsabile dell'inibizione dei riflessi spinali e dei movimenti prodotti dalla stimolazione del fascio piramidale (v. Pompeiano, 1965, 1966 e The neurophysiological..., 1967). Pertanto meccanismi inibitori discendenti interessati al ciclo sonno-veglia possono essere messi selettivamente in attività durante l'ipnosi animale. Questo fenomeno è manifestamente in rapporto con i casi clinici di cataplessia (v. cap. 9, § a) e con gli episodi cataplessici del gatto decerebrato (v. cap. 3, § b).
b) L'ibernazione dei Mammiferi
L' ibernazione è un fenomeno stagionale che può essere osservato durante l'inverno in alcuni mammiferi, quali lo scoiattolo e la marmotta. L'argomento è stato oggetto di una rassegna critica di Sàtinoff (v., 1970).
Dopo un periodo di preparazione che si svolge nella tarda estate e nel primo autunno, durante il quale aumenta l'ingestione di cibo e si accresce il peso dell'animale, la temperatura corporea dello scoiattolo comincia a cadere e può giungere a valori assai bassi, fino a 4-5 °C. L'instaurarsi dello stato di letargo è in rapporto a questa caduta della temperatura. Dopo circa due settimane d'ibernazione l'animale si sveglia completamente, pur nell'assenza di qualsiasi cambiamento apprezzabile nell'ambiente, mentre la temperatura sale agli abituali livelli omeotermici di 37 °C. Il livello di vigilanza è eccellente durante questi periodi di veglia, che possono durare anche qualche giorno all'inizio o alla fine dell'ibernazione, mentre a mezzo inverno durano solo poche ore. In questi periodi l'animale orina e può anche mangiare, bere o defecare, per poi cadere nuovamente nello stato d'ibernazione. Solo dall'inizio della primavera all'autunno l'animale ritorna al normale ciclo sonno-veglia.
L'ibernazione non può essere considerata come uno stato di coma, giacché gli animali possono risvegliarsi, quando vengono stimolati, e rispondere a stimoli acustici voltando la testa in direzione del suono. Si tratta tuttavia di un tipo differente di sonno, almeno per ciò che riguarda le fasi sincronizzata e paradossale che si osservano nello scoiattolo non ibernante. Gli studi elettroencefalografici hanno dato risultati contrastanti, che possono anche essere spiegati con differenze di specie. È certo comunque che quando lo scoiattolo raggiunge lo stadio finale dell'ibernazione, caratterizzato da temperature che possono scendere a 6-8 °C e da una caduta della frequenza cardiaca a 6-7 pulsazioni al minuto, vi possono essere lunghi periodi di silenzio elettrico nella corteccia cerebrale. È molto improbabile che l'azione ristoratrice del sonno possa svolgersi nei neuroni corticali a temperatura così bassa. Il significato funzionale della letargia dell'animale ibernante sembrerebbe quindi essere del tutto differente da quello del sonno fisiologico.
Vi è tuttavia un lento ritmo ciclico fra i periodi di risveglio che interrompono l'ibernazione e le lunghe fasi di letargo. Questo ritmo è però del tutto differente dal ciclo dell'animale non ibernante. A mezzo inverno l'animale può rimanere in letargo per un periodo che dura fino a 14 giorni ed essere sveglio per sole 12-14 ore. Questo lento ciclo deve essere dovuto a meccanismi ipotalamici o del tronco dell'encefalo, giacché non è verosimile che la corteccia cerebrale silente possa intervenire nel risveglio.
bibliografia
Adrian, E. D., Electrical activity of the nervous system, in ‟Archives of neurology and psychiatry", 1934, XXXII, pp. 1125-1136.
Adrian, E. D., Moruzzi, G., Impulses in the pyramidal tract, in ‟Journal of physiology", 1939, XCVII, pp. 153-199.
Alekseyeva, T. T., Correlation of nervous and humoral factors in the development of sleep in non-disjointed twins, in ‟Zhurnal vysshei nervnoi deyatel'nosti imeni I. P. Pavlova", 1958, VIII, pp. 835-844.
Amatruda, T. T. III, Black, D. A., McKenna, T. M., McCarley, R. W., Hobson, J. A., Sleep cycle control and cholinergic mechanisms: differential effects of carbachol injections at pontien brain stem sites, in ‟Brain research", 1975, XCVIII, pp. 501-515.
Axelrod, J., Brain monoamines, in ‟Neurosciences research program bulletin", 1971, IX, pp. 188-196.
Batini, C., Moruzzi, G., Palestini, M., Rossi, G. F., Zanchetti, A., Effects of complete pontine transections on the sleep-wakefulness rhythm: the midpontine pretrigeminal preparation, in ‟Archives italiennes de biologie", 1959, XCVII, pp. 1-12.
Batsel, H. L., Electroencephalographic synchronization and desynchronization in the chronic ‛cerveau isolé' of the dog, in ‟Electroencephalography and clinical neurophysiology", 1960, XII, pp. 421-430.
Batsel, H. L., Spontaneous desynchronization in the chronic cat ‛cerveau isolé', in ‟Archives italiennes de biologie", 1964, CII, pp. 547-566.
Baust, W., Die Phänomenologie des Schafles, in Ermüdung, Schlaf und Traum (a cura di W. Baust), Stuttgart 1970, pp. 99-144.
Belugou, J. L., Benoit, O., Leygonie, F., Décharges neuronales de l'hippocampe au cours de la veille et du sommeil, in ‟Journal de physiologie", 1968, LX, suppl. 2, p. 399.
Berger, H., Über das Elektroenzephalogramm des Menschen, in ‟Archiv für Psychiatrie", 1929, LXXXVII, pp. 527-570.
Berlucchi, G., Callosal activity in unrestrained, unanesthetized cats, in ‟Archives italiennes de biologie", 1965, CIII, pp. 623-634.
Berlucchi, G., Mechanismes von Schlafen und Wachen, in Ermüdung, Schlaf und Traum (a cura di W. Baust), Stuttgart 1970, pp. 145-203.
Bernard, Cl., Introduction à l'étude de la médecine expérimentale, Paris 1865.
Bonvallet, M., Système nerveux et vigilance, Paris 1966.
Bremer, F., Cerveau ‛isolé' et physiologie du sommeil, in ‟Comptes rendus des séances de la Société de Biologie", 1935, CXVIII, pp. 1235-1241.
Bremer, F., L'activité cérébrale au cours du sommeil et de la narcose. Contribution à l'étude du mécanisme du sommeil, in ‟Bulletin de l'Académie Royale de Médecine de Belgique", 1937, IV, pp. 68-86.
Bremer, F., L'activité électrique de l'écorce cérébrale et le problème physiologique du sommeil, in ‟Bollettino della Società Italiana di Biologia Sperimentale", 1938, XIII, pp. 271-290.
Bremer, F., Preoptic hypnogenic focus and mesencephalic reticular formation, in ‟Brain research", 1970, XXI, pp. 132-134.
Bremer, F., Existence of a mutual tonic inhibitory interaction between the preoptic hypnogenic structures and the midbrain reticular formation, in ‟Brain research", 1975, XCVI, pp. 71-75.
Bremer, F., Cerebral hypnogenic centers, in ‟Archives of neurology", 1977, II, pp. 1-6.
Brunelli, M., Magni, F., Moruzzi, G., Musumeci, D., Effetti di stimolazioni elettriche localizzate del ponte sulle attività istintive del piccione talamico e del piccione integro, in ‟Atti dell'Accademia Nazionale dei Lincei" (serie VIII), 1971, II, pp. 603-606.
Brunelli, M., Magni, F., Moruzzi, G., Musumeci, D., Effects of pontine stimulations on sleep and waking behaviors of the pigeon, in Basic sleep mechanisms (a cura di O. Petre-Quadens e J. D. Schlag), New York-London 1974, pp. 33-49.
Buser, P., Viala, G., Analyse du mécanisme de l'akinésie induite ‛hypnotique' chez le lapin, in Actualités neurophysiologiques (a cura di A. M. Monnier), vol. VIII, Paris 1968, pp. 179-196.
Carli, G., Dissociation of electrocortical activity and somatic reflexes during rabbit hypnosis, in ‟Archives italiennes de biologie", 1969, CVII, pp. 219-234.
Carli, G., Subcortical mechanisms of rabbit hypnosis, in ‟Archives italiennes de biologie", 1971, CIX, pp. 15-26.
Clemente, C. D., Sterman, M. B., Wyricka, W., Forebrain inhibitory mechanisms: conditioning of basal forebrain induced EEG synchronization and sleep, in ‟Experimental neurology", 1963, VII, pp. 404-417.
Craig, W., Appetites and aversions as constituents of instincts, in ‟Biological bulletin", 1918, XXXIV, pp. 91-107.
Dell, P., Lower brain stem mechanisms and sleep induction, in Proceedings of the XXV international congress of physiology, München 1971, pp. 29-30.
Dement, W. C., Kleitman, N., Cyclic variations in EEG during sleep and their relation to eye movements, body motility, and dreaming, in ‟Electroencephalography and clinical neurophysiology", 1957, IX, pp. 673-690.
Dement, W. C., Rechtschaffen, A., Narcolepsy: polygraphic aspects, experimental and theoretical considerations, in The abnormalities of sleep in man (a curadi H. Gastaut, E. Lugaresi, G. Berti Ceroni e G. Coccagna), Bologna 1968, pp. 147-164.
Draskóczy, P. R., Lyman, C. P., Turnover of catecholamines in active and hibernating ground squirrels, in ‟Journal of pharmacology and experimental therapeutics", 1967, CLV, pp. 101-111.
Economo, C. von, Die Encephalitis lethargica, Wien 1918.
Economo, C. von, Schlaftheorie, in ‟Ergebnisse der Physiologie", 1929, XXVIII, pp. 312-339.
Evarts, E. V., Activity of neurons in visual cortex of the cat during sleep with low voltage fast EEG activity, in ‟Journal of neurophysiology", 1962, XXV, pp. 812-816.
Evarts, E. V., Temporal patterns of discharge of pyramidal tract neurons during sleep and waking in the monkey, in ‟Journal of neurophysiology", 1964, XXVII, pp. 152-171.
Evarts, E. V., Relation of cell size to effects of sleep in pyramidal tract neurons, in Progress in brain research, vol. XVIII, Sleep mechanisms (a cura di K. Akert, C. Bally e J. P. Schadé), Amsterdam 1965, pp. 81-91.
Evarts, E. V., Relation of discharge frequency to conduction velocity in pyramidal tract neurons, in ‟Journal of neurophysiology", 1965, XXVIII, pp. 215-228.
Evarts, E. V., Activity of individual cerebral neurons during sleep and arousal, in Association for Research in Nervous and Mental Disease, vol. XLV, Sleep and altered states of consciousness, Baltimore 1967, pp. 319-337.
Feinberg, I., Evarts, E. V., Changing concepts of the function of sleep: discovery of intense brain activity during sleep calls for revision of hypotheses as to its function, in ‟Biological psychiatry", 1969, I, pp. 331-348.
Genovesi, U., Moruzzi, G., Palestini, M., Rossi, G. P., Zanchetti, A., EEG and behavioral patterns following lesions of the mesencephalic reticular formation in chronic cats with implanted electrodes, in Abstracts of communications. 20th international physiological congress, Bruxelles 1956, pp. 335-336.
Gerebtzoff, M., État fonctionnel de l'écorce cérébrale au cours de l'hypnose animale, in ‟Archives internationales de physiologie", 1941, LI, pp. 365-378.
Hediger, H., Wie Tiere schlafen, in ‟Medizinische Klinik", 1959, LIV, pp. 938-946, 965-968.
Hediger, H., Comparative observations on sleep, in ‟Proceedings of the Royal Society of Medicine", 1969, LXII, pp. 153-156.
Heller, H. C., Glotzbach, S. F., Thermoregulation during sleep and hibernation, in ‟International review of physiology", 1977, XV, pp. 147-188.
Hess, W. R., Stammganglien-Reizversuche. Tagung der deutschen physiologischen Gesellschaft, in ‟Berichte über die gesamte Physiologie und experimentelle Pharmakologie", 1927, XLII, pp. 554-555.
Hess, W. R., Das Schlafsyndrom als Folge dienzephaler Reizung, in ‟Helvetica physiologicva et pharmacologica acta", 1944, II, pp. 305-344.
Hess, W. R., Das Zwischenhirn. Syndrome, Lokalisationen, Funktionen, Basel 1949.
Hess, W. R., Psychologie in biologisher Sicht, Stuttgart 1968.
Hess, W. R., Hypothalamus und Thalamus, Stuttgart 1969.
Hobson, J. A., McCarley, R. W., Discharge patterns of cat pontine brain stem neurons during desynchronized sleep, in ‟Journal of neurophysiology", 1975, XXXVIII, pp. 751-766.
Hobson, J. A., McCarley, R. W., Wyzinski, P. W., Sleep cycle oscillation: reciprocal discharge by two brain stem neuronal groups, in ‟Science", 1975, CLXXXIX, pp. 55-58.
Holm, E., Die pharmakologische induzierte Schlaf, in Ermüdung, Schlaf und Traum (a cura di W. Baust), Stuttgart 1970, pp. 247-308.
Holzapfel, M., Triebbedingte Ruhezustände als Ziel von Appetenzhandlungen, in ‟Naturwissenschaften", 1940, XXVIII, pp. 273-280.
Hoshino, K., Pompeiano, O., Selective discharge of pontine neurons during the postural atonia produced by an anticholinesterase in the decerebrate cat, in ‟Archives italiennes de biologie", 1976, CXIV, pp. 244-277.
Jouvet, M., Recherches sur les structures nerveuses et les mécanismes responsables des différentes phases du sommeil physiologique, in ‟Archives italiennes de biologie", 1962, C, pp. 125-206.
Jouvet, M., Neurophysiology of the states of sleep, in ‟Physiological reviews", 1967, XLVII, pp. 117-177.
Jouvet, M., The role of monoamine and acetylcholine containing neurons in the regulation of the sleep-waking cycle, in ‟Ergebnisse der Physiologie", 1972, LXIV, pp. 166-307.
Jouvet, M., Le sommeil paradoxal est-il responsable d'une programmation génétique du cerveau?, in ‟Comptes rendus des séances de la Société de Biologie", 1978, CLXXII, pp. 9-32.
Jouvet, M., Delorme, F., Locus coeruleus et sommeil paradoxal, in ‟Comptes rendus des séances de la Société de Biologie", 1965, CLIX, pp. 895-899.
Jouvet, M., Renault, J., Insomnie persistante après lésions des noyaux du raphé chez le chat, in ‟Comptes rendus des séances de la Société de Biologie", 1966, CLX, pp. 1461-1465.
Klaue, R., Die bioelektrische Tätigkeit der Grosshirnrinde in normalen Schlaf und in der Narkose durch Schlafmittel, in ‟Journal für Psychologie und Neurologie", 1937, XLVII, pp. 510-531.
Kleitman, N., Sleep and wakefulness, Chicago 1963.
Klemm, W. R., Electroencephalographic-behavioral dissociations during animal hypnosis, in ‟Electroencephalography and clinical neurophysiology", 1966, XXI, pp. 365-372.
Konorski, J., Integrative activity of the brain, Chicago-London 1967.
Lindsley, D. B., Bowden, J. W., Magoun, H. W., Effect upon the EEG of acute injury to the brain stem activating system, in ‟Electroencephalography and clinical neurophysiology", 1949, I, pp. 475-486.
Lindsley, D. B., Schreiner, L. H., Knowles, W. B., Magoun, H. W., Behavioral and EEG changes following chronic brain stem lesions in the cat, in ‟Electroencephalography and clinical neurophysiology", 1950, II, pp. 483-498.
Luce, G. G., Current research on sleep and dreams, Public Health Service Publication No. 1389, Bethesda 1965.
McGinty, D. J., Sterman, M. B., Sleep suppression after basal forebrain lesions in the cat, in ‟Science", 1968, CLX, pp. 1253-1255.
Magoun, H. W., The waking brain, Springfield 19632.
Mantegazzini, P., Glässer, A., Action de la DL-3,4-dioxyphenylalanine (Dopa) et de la dopamine sur l'activité électrique du chat ‛cerveau isolé', in ‟Archives italiennes de biologie", 1960, XCVIII, pp. 367-374.
Marchiafava, P. L., Pompeiano, O., Pyramidal influences on spinal cord during desynchronized sleep, in ‟Archives italiennes de biologie", 1964, CII, pp. 500-529.
Moruzzi, G., The physiological properties of the brain stem reticular system, in Brain mechanisms and consciousness (a cura di E. D. Adrian, F. Bremer e H. H. Jasper), Oxford 1954, pp. 21-53.
Moruzzi, G., Synchronizing influences of the brain stem and the inhibitory mechanisms underlying the production of sleep by sensory stimulation, in ‟Electroencephalography and clinical neurophysiology", 1960, suppl. 13, pp. 231-256.
Moruzzi, G., Active processes in the brain stem during sleep, in ‟Harvey lectures", 1963, LVIII, pp. 233---297.
Moruzzi, G., The historical development of the deafferentation hypothesis of sleep, in ‟Proceedings of the American Philosophical Society", 1964, CVIII, pp. 19-28.
Moruzzi, G., Summary statement, in Progress in brain research, vol. XVIII, Sleep mechanisms (a cura di K. Akert, C. Bally e J. P. Schadé), Amsterdam 1965, pp. 241-243.
Moruzzi, G., The functional significance of sleep with particular regard to the brain mechanisms underlying consciousness, in Brain and conscious experience (a cura di J. C. Eccles), Berlin-Heidelberg-New York 1966, pp. 345-388.
Moruzzi, G., Sleep and instinctive behavior, in ‟Archives italiennes de biologie", 1969, CVII, pp. 175-216.
Moruzzi, G., The sleep-waking cycle, in ‟Ergebnisse der Physiologie", 1972, LXIV, pp. 1-165.
Moruzzi, G., Neural mechanisms of the sleep-waking cycle. Basic sleep mechanism (a cura di O. Petre-Quadens e J. D. Schlag), New York-London 1974, pp. 13-31.
Moruzzi, G., Magoun, H. W., Brain stem reticular formation and activation of the EEG, in ‟Electroencephalography and clinical neurophysiology", 1949, I, pp. 455-473.
Mukhametov, L. M., Rizzolatti, G., Tradardi, V., Spontaneous activity of neurones of nucleus reticularis thalami in freely moving cats, in ‟Journal of physiology", 1970, CCX, pp. 651-667.
Orthner, H., Neuroanatomische Gesichtspunkte der Schlaf-Wach-Regelung, in Der Schlaf (a cura di U. J. Jovanović), München 1969, pp. 49-84.
Otter, H. P., Die pathologische Physiologie des Schlafes, in Ermüdung, Schlaf und Traum (a cura di W. Baust), Stuttgart 1970, pp. 217-245.
Pappenheimer, J. R., Koski, G., Fench, V., Induction of slow wave sleep by a factor released into cerebrospinal fluid (CSF) during sleep deprivation, in Proceedings of the XXV international congress of physiology. Abstracts of volunteer papers and films, vol. IX, München 1971, p. 439.
Pappenheimer, J. R., Miller, T. B., Goodrich, C. A., Sleep-promoting effects of cerebro-spinal fluid from sleep-deprived goats, in ‟Proceedings of the National Academy of Sciences", 1967, LVIII, pp. 513-517.
Parmeggiani, P. L., Rabini, C., Shivering and panting during sleep, in ‟Brain research", 1967, VI, pp. 789-791.
Piéron, H., Le problème physiologique du sommeil, Paris 1913.
Pivik, R. T., McCarley, R. W., Hobson, J. A., Eye movement-associated discharge in brain stem neurons during desynchronized sleep, in ‟Brain research", 1977, CXXI, pp. 59-76.
Pompeiano, O., Ascending and descending influences of somatic afferent volleys in unrestrained cats; supraspinal inhibitory control of spinal reflexes during natural and reflexly induced sleep, in Aspects anatomofonctionnels de la physiologie du sommeil (a cura di M. Jouvet), Paris 1965, pp. 309-395.
Pompeiano, O., Muscular afferents and motor control during sleep, in Muscular afferents and motor control (a cura di R. Granit), Stockholm 1966, pp. 415-436.
Pompeiano, O., Sensory inhibition during motor activity in sleep, in Neurophysiological basis of normal and abnormal motor activities (a cura di M. D. Yahr e D. P. Purpura), New York 1967, pp. 323-375.
Pompeiano, O., The neurophysiological mechanisms of the postural and motor events during desynchronized sleep, in Association for Research in Nervous and Mental Diseases, vol. XLV, Sleep and altered states of consciousness, Baltimore 1967, pp. 351-423.
Pompeiano, O., Hoshino, K., Tonic inhibition of dorsal pontine neurons during the postural atonia produced by an anticholinesterase in the decerebrate cat, in ‟Archives italiennes de biologie", 1976, CXIV, pp. 310-340.
Pompeiano, O., Swett, J. E., EEG and behavioral manifestations of sleep induced by cutaneous nerve stimulation in normal cats, in ‟Archives italiennes de biologie", 1962, C, pp. 311-342.
Pompeiano, O., Swett, J. E., Identification of cutaneous and muscular afferent fibers producing EEG synchronization or arousal in normal catas, in ‟Archives italiennes de biologie", 1962, C, pp. 343-380.
Puzillout, J. J., Foutz, A. S., Characteristic of the experimental reflex sleep induced by vago-aortic nerve stimulation, in ‟Journal of electroencephalography and clinical neurophysiology", 1977, XLII, pp. 552-563.
Puizillout, J. J., Ternaux, J. P., Endormement vago-aortique après section sagittale médiane du tronc cérébral et après administration de p-chlorophenylalanine ou destruction des noyaux du raphé, in ‟Brain research", 1974, LXX, pp. 19-42.
Purpura, D. P., Frigyesi, T. L., McMurtry, J. C., Scarf, T., Synaptic mechanisms in thalamic regulation of cerebello-cortical projection activity, in The talamus (a cura di D. P. Purpura e M. D. Yahr), New York-London 1966, pp. 153-172.
Ranson, S. W., Somnolence caused by hypothalamic lesions in the monkey, in ‟Archives of neurology and psychiatry", 1939, XLI, pp. 1-23.
Roffwarg, H. P., Muzio, J. N., Dement, W. C., Ontogenetic development of the human sleep-dream cycle, in ‟Science", 1966, CLII, pp. 604-619.
Sastre, J. P., Effets des lésions du tegmentum pontique sur l'organisation des états de sommeil chez le chat, Lyon 1978.
Sàtinoff, E., Hibernation and the central nervous system, in Progress in physiological psychology (a cura di E. Stellar e J. M. Sprague), vol. III, New York 1970, pp. 201-236.
Sharpless, S., Jasper, H., Habituation of the arousal reaction, in ‟Brain", 1956, LXXIX, pp. 655-680.
Sherrington, C. S., Man on his nature, Cambridge 1946.
Steriade, M., Cortical long-axoned cells and putative interneurons during the sleep-waking cycle, in ‟The behavioral and brain sciences", 1978, III, pp. 465-514.
Steriade, M., Ropert, N., Kitsikis, A., Oakson, G., Ascending activating neuronal networks in midbrain reticular core and related rostral systems in The reticular formation revisited (a cura di J. A. Hobson e M. A. Brazier), New York 1980, pp. 125-167.
Sterman, M. B., Clemente, C. D., Forebrain inhibitory mechanisms: cortical synchronization induced by basal forebrain stimulation, in ‟Experimental neurology", 1962, VI, pp. 91-102.
Sterman, M. B., Clemente, C. D., Forebrain inhibitory mechanisms: sleep patterns induced by basal forebrain stimulation in the behaving cat, in ‟Experimental neurology", 1962, VI, pp. 103-117.
Svorad, D., ‛Animal hypnosis' (Totstellreflex) as experimental model for psychiatry. Electroencephalographic and evolutionary aspects, in ‟American Medical Association. Archives of neurology and psychiatry", 1957, LXXVII, pp. 533-539.
Tinbergen, N., ‛Derived' activities: their causation, biological significance, origin, and emancipation during evolution, in ‟Quarterly review of biology", 1952, XXVII, pp. 1-32.
Tinbergen, N., The study of instinct, Oxford 19552.
Tönnies, J. F., Automatische EEG-Intervall-Spektrumanalyse (EISA) zur Langzeitdarstellung der Schlafperiodik und Narkose, in ‟Archiv für Psychiatrie und Nervenkrankheiten", 1969, CCXII, pp. 423-445.
Villablanca, J., The electrocortigram in the chronic cerveau isolé cat, in ‟Electroencephalography and clinical neurophysiology", 1965, XIX, pp. 576-586.
Villablanca, J., Behavioral and polygraphic study of ‛sleep' and ‛wakefulness' in chronic decerebrate cats, in ‟Electroencephalography and clinical neurophysiology", 1966, XXI, pp. 562-577.