
Associazioni biologiche
Associazioni biologiche
di Valerio Giacomini
Associazioni biologiche
sommario: 1. Generalità e definizioni: a) il concetto di individuo; b) l'individualità cellulare; c) l'individualità pluricellulare; d) l'individualità nel tempo; e) il concetto di associazione biologica. 2. Livelli di integrazione associativa: a) livelli di organizzazione e di integrazione; b) sistemi di interrelazioni associative; c) informazione e comunicazione. 3. Sistemi precellulari e protocellulari (livello prebiologico e procariotico). 4. I sistemi cellulari (livello eucariotico): a) dalla protocellula alla cellula; b) la teoria cellulare simbiotica; c) argomenti contro la teoria simbiotica; d) origine dei mitocondri; e) origine dei cloroplasti; f) origine dei microtubuli. 5. Sistemi pluricellulari preorganismici (livello cenobiale e plasmodiale): a) dagli Eucarioti ai Metafiti e Metazoi; b) dalla cellula al cenobio; c) plasmodi e pseudoplasmodi; d) i Mesozoi; e) gli organismi pluricellulari come sistemi di comunicazione. 6. Sistemi interorganismici intraspecifici (livello superorganismico): a) le associazioni biologiche omotipiche; b) l'associazione coloniale; c) le società animali; d) livelli biosociologici. 7. Sistemi interorganismici interspecifici (livello simbiotico propriamente detto): a) gradienti di interrelazioni; b) mutualismo e parassitismo; c) sistemi intermicrobici; d) sistemi microbiciprotozoici; e) sistema Rhizobium-Leguminose; f) sistema micorrizico; g) sistemi microbici-endozoici; h) sistema lichenico e sistemi algali-endofitici; i) sistemi algali-endozoici; l) sistemi Metazoi-Metafiti; m) sistemi intermetafitici; n) sistemi intermetazoici; o) sistemi polisimbiotici. 8. Sistemi ecologici (livello metabiotico): a) approssimazioni conoscitive; b) l'ecosistema; c) l'ecosfera. □ Bibliografia.
1. Generalità e definizioni
Questa trattazione, data la grande vastità dell'argomento e delle ricerche che sono state compiute, non può costituire che un tentativo di sintesi e di generalizzazione. Il concetto di ‛associazione biologica' viene qui inteso nel senso più largo, al quale tendono oggi molti autori. Già nel passato, del resto, si erano affacciate idee su un senso molto generale e molto vasto da dare al concetto di associazione biologica.
In genere si ammette che non esiste l'individuo isolato, in quanto la vita vegetale e animale presenta sempre aspetti di aggregazione: individui della stessa specie e individui di specie diverse si associano contraendo diversi gradi di rapporti o interrelazioni. Sono rapporti di vicendevole contenimento, di predazione, di commensalismo, di parassitismo, di mutualismo; possono essere effimeri, cioè temporanei, o permanenti; possono essere facoltativi o obbligati.
I tentativi di classificare i tipi di associazione biologica urtano sempre contro la constatazione che ogni tipo presenta transizioni graduali verso altri tipi, con una continuità quasi ininterrotta. Un esempio significativo è l'impossibilità di delimitare nettamente il parassitismo dal mutualismo.
a) Il concetto di individuo
Di fronte a un concetto così generale di associazione biologica diventa critico il concetto di ‛individualità' degli esseri viventi. Il senso più comune di individuo è: un essere distinto dagli altri e indivisibile (corpora individua), una ‟forma microcosmica indivisibile" (v. Schmid, 1943), ciò che vive per se stesso realizzando una tale centralizzazione e coordinazione di funzioni, da non poter essere diviso senza perdere il suo nome e le sue qualità distintive. Vi è tuttavia chi non ammette l'assolutezza del concetto di individuo in biologia. Già a suo tempo E. Haeckel (v., 1866) distingueva tre punti di vista: a) morfologico o formale: manifestazione unitaria di forma, di un tutto le cui parti non possono esser separate senza sopprimere il carattere essenziale; b) funzionale o fisiologico (il cosiddetto ‛bionte'): manifestazione unitaria di forma che può avere, per un tempo più o meno lungo, un'esistenza propria, che attua uno sviluppo, una riproduzione, un'autoconservazione; c) genealogica: manifestazione unitaria nel tempo, costituita da una serie ciclica chiusa di variazioni spaziali. E di Haeckel anche la distinzione fra ‛attualità' e ‛potenzialità'. È facile rendersi conto di questa importante distinzione, se si considera l'eloquente esempio ricordato da H. Vöchting: il ramo dell'albero ha la possibilità di diventare individuo, ma non lo è ancora; lo diverrà appena venga separato dall'albero e formi le sue proprie radici. F. Raffaele (v., 1943) nega l'assolutezza del concetto di individuo e asserisce che ‟l'individuo non rappresenta se non uno stadio, una forma transitoria" ed esistono quindi ‟individui di diverso ordine" (superiore e inferiore). Questo modo di vedere, molto comprensibile in uno zoologo, considera l'individuo in divenire nell'ontogenesi. Ma vi è possibilità di considerare l'individuo in divenire anche nella filogenesi, e in tal senso diventa anche meno assoluto, anzi difficilmente afferrabile, perché entra in un processo di gradualità evolutiva; è più agevole allora parlare di un processo di ‛individualizzazione', che si attua in successivi livelli di crescente organizzazione.
b) L'individualità cellulare
L'individualità della cellula è stata oggetto di discussione sin da tempi lontani. Fin dal 1827 P.-J.-F. Turpin affermava che come un albero è composto da germogli e i germogli da cellule, così queste ultime sono costituite a loro volta da ‟individualités inférieures, subordonnées globuleuses", cioè da particelle del protoplasma coincidenti con le granulazioni (protosphaeria simplex) o con elementi filiformi (protonema simplex). Alcuni anatomici del tempo non esitarono a fantasticare sull'esistenza di monadi di natura animale per spiegare i movimenti del plasma (così le ‛biosfere' dell'anatomico A. Meyer di Bonn, 1827). Aderirono a questa interpretazione di un compositum cellulare F. T. Ktitzing, F. Unger (con i suoi individualisierte Körper), K. W. Nägeli e altri (v. Fisch, 1880), fino ad arrivare ai tempi nostri: F. Raffaele (v., 1943) afferma che quell'individuo, quell'essere vivente che noi, per fissarne l'individualità, chiamiamo cellula, è, in ogni caso, una riunione, una colonia di altre individualità viventi, la cui condizione di vita è di stare insieme. Si ricollega quindi con la teoria di A. Weismann che ritiene esistano esseri viventi più elementari - i ‛biofori' - aggregati di molecole con caratteri di vita da cui deriverebbero altri individui di ordine superiore (determinanti, idi, idanti...).
Nonostante questi e altri punti di vista, non pare si possa abbandonare il concetto che ‟la cellula è il più semplice organismo integrato nel sistema vivente" (v. Paul, 1965). La ‛teoria cellulare' rimane quindi sostanzialmente valida nel senso di Th. Schwann: le cellule sono organismi e gli animali e le piante sono aggregati di questi organismi. Neppure il problema dei Virus può turbare questa concezione, sia che si considerino cellule degenerate o fasi primitive di organizzazione cellulare.
Anche la teoria simbiotica della cellula, di cui tratteremo più avanti, non si oppone a una concezione pratica che considera la cellula come individuo, perché non è concepibile ‛attualmente' o ‛potenzialmente' una sua suddivisione in parti autonome.
c) L'individualità pluricellulare
Fissata l'individualità della cellula, sia pure come momento o livello dell'evoluzione, si è prolungata per molti anni una discussione sulla individualità degli organismi costituiti da più cellule. R. Virchow con la sua teoria della Zellenstaat ha sostenuto che solo le cellule sono unità viventi: ciò che ci appare unitario in un organo o in un organismo è la risultante di attività di singole cellule che si integrano a vicenda; anche l'organismo più elevato è di carattere ‛sociale' (teoria che ha avuto incredibili estensioni in campo medico, sociale, politico). L'idea che la cellula sia la ‛pietra di costruzione' per unità più complesse che possono essere semplici ‛somme' senza individualità è stata sostenuta da molti autori. R. H. Lotze (1851) riteneva individui solo gli aggregati di cellule diversificati secondo un ‛piano', secondo un ordine centralizzato, come è il caso degli animali; le piante - opinione durata a lungo - non avrebbero individualità, ma sarebbero solo coesistenze di parti. H. Vöchting invece negava che la cellula isolata possa compiere tutte le funzioni dell'organismo, affermando che la funzionalità della cellula isolata dipende solo da se stessa, mentre nell'organismo la funzionalità è condizionata dalla totalità, anche se non si deve confondere l'attuale col potenziale, perché potenzialmente in ogni cellula singola è contenuta la totalità. In conclusione, secondo C. Fisch (v., 1880) è necessario distinguere semplici aggregati cellulari, in cui permane l'indipendenza delle cellule, e complessi organici dove ogni cellula è pietra di costruzione di un edificio complesso: l'organismo composto individualizzato. Distinzione che però non è nettamente tracciabile. Tipico è l'esempio dei Volvox, in cui non è facile decidere fra una individualità del complesso o dei singoli componenti.
d) L'individualità nel tempo
Già sin dal 1844 E. Forbes aveva proposto di considerare l'individuo come una positive reality (in contrapposizione con la specie relative reality) in una connessione non spaziale ma temporale, cioè come una realtà limitata nel tempo. Anche altri hanno insistito su questo concetto, considerando il concetto di ‛morte' consequenziale al concetto di individuo (ad es. J. M. Schleiden). Se questo è chiaro negli organismi dotati di riproduzione sessuale, diventa meno precisabile negli organismi dotati di riproduzione asessuata, per i quali è stata proposta la teoria della ‛immortalità del protoplasma'. A questo proposito recentemente M. Bates (1961) ha proposto di considerare che l'individuo abbia inizio con la scissione in cui si determina la sua distinta unità, e termini con la morte o con l'ulteriore scissione che fa cessare la sua unità; più generalmente egli afferma che ‟l'individuo è l'organismo compreso tra due scissioni o due altri processi riproduttivi". Conseguentemente Bates tende a negare individualità a un ciclo vitale di microrganismi (ad es. al ciclo del plasmodio della malaria), che considera piuttosto come una serie di individui diversi e non come metamorfosi di un singolo individuo.
Si può convenire su un piano di considerazioni molto generale, che la sostanza germinale è continua e immortale in tutti gli organismi. Scrive W. Morton Wheeler che tutti gli esseri viventi sono geneticamente correlati come membri di una grande famiglia, di un vasto simplasma vivo, che nonostante sia frammentato in individualità spaziali, tuttavia è assolutamente continuo nel tempo. Si accresce la ‛relatività' del concetto di individuo se al punto di vista morfologico si aggiunge quello fisiologico, e finalmente anche il punto di vista temporale. I tentativi di combinare questi diversi criteri hanno qualche successo solo se si abbandona il proposito di definire individualità assolute e si entra nell'ordine di idee di un processo di individualizzazione che si svolge nell'evoluzione della vita nel mondo.
e) Il concetto di associazione biologica
Avendo adottato il concetto di associazione biologica nel senso più ampio, intendiamo anche dare all'‛associazione', come all'‛individuazione', un senso dinamico-evolutivo.
Come non pochi autori hanno già osservato, nei tentativi di chiarire il meccanismo dell'evoluzione si è data troppa importanza ai processi di lotta e di selezione fra organismi, mentre troppo poco ci si è interessati ai processi di associazione e di integrazione. È certamente una delle tendenze più significative della biologia moderna la concezione di ‛livelli di integrazione' sia in campo ontogenetico, sia in campo filogenetico, sia - ancor più recentemente - in campo ecologico. I problemi di integrazione strutturale e funzionale riguardano l'individuo, la popolazione, la specie, ma noi desideriamo considerarli qui soprattutto come problemi di progressiva organizzazione di bionti in metabionti, di metabionti in associazioni sempre più complesse e più solidali, che possono dare origine a metabionti più altamente integrati.
Ci sembra non si possa sfuggire oggi a una esigenza ‛olistica' e organicistica, che accetta l'atteggiamento filosofico del ‛realismo pluralistico' nel momento analitico della ricerca, ma che eredita anche un impulso dal ‛monismo idealistico' per esigenze insopprimibili di coerenza del sapere scientifico. Quando il botanico A. Braun (1852-1853) afferma che la divisibilità esteriore non è di decisiva importanza e che più importa la connessione sostanziale di tutti i livelli dello sviluppo a una Idea della comune appartenenza del Tutto, propone tuttavia un dogmatismo teleologico, destinato a essere travolto dalla sconcertante molteplicità del reale. Più accettabile appare oggi una concezione olistica strumentale che si appaga di cogliere approssimazioni il meno possibile frammentarie alla conoscenza di ‛livelli di integrazione', considerati come risultati di un vasto divenire della vita nel mondo.
In questo senso si ammette una relatività dell'individuo e anche una relatività dell'associazione. Diventano, invece, realtà più accettata nella prassi della ricerca scientifica l'‛individualizzazione' e l'‛associazione' come processi di un più grande divenire evolutivo.
Non sembra tuttavia necessario giungere con M. Hartmann (v. Schmid, 1943) all'idea di livelli di integrazione discontinui, distanziati nettamente gli uni dagli altri (‛legge della stratificazione'), mentre pare più accettabile che la legalità dei livelli inferiori valga per quelli superiori e non viceversa, costituendo un novum specifico (‛legge del novum').
2. Livelli di integrazione associativa
a) Livelli di organizzazione e di integrazione
Un'antica concezione umana, che risale almeno ai tempi delle cosmologie presocratiche, persiste nella storia del pensiero scientifico e sta assumendo oggi una forte e rinnovata incidenza: si tratta di una visione delle realtà naturali e, in particolare, delle realtà biologiche, in un ordine di ascendente complessità, in una scala di gradienti di organizzazione. L'idea di una struttura scalare sta superando le fasi metafisiche di una più o meno vaga ‛scala naturale', per assumere una crescente solidità scientifica. Un principio di integrazione si impone sempre più oggi non soltanto come retaggio della teoria della discendenza, ma come esigenza fondamentale delle più recenti sintesi ontogenetiche, filogenetiche e anche ecologiche, che stanno offrendo nuove visioni compendiose del mondo vivente nel suo essere e nel suo divenire.
Le più recenti espressioni di questa concezione hanno assunto carattere chimico-energetico. Secondo W. Ostwald l'organismo di più elevata organizzazione è quello capace di massima assunzione di energia e di più economico consumo. In questo senso lo stesso mondo vivente costituisce un grande livello diversificato di organizzazione della materia. Questa considerazione ha indotto J. Needham (v., 1936) alla più vasta generalizzazione: tutto l'universo inorganico e organico è organizzato su livelli di diversa complessità.
È su questa base che, in questo tentativo di sintesi, si è voluto ordinare il materiale informativo su un'ampiezza che alcuni non ritengono giustificata (ad es. v. Schmid, 1943), pur non ignorando il problema di fondo posto da F. E. Egler (v., 1942) quando si chiedeva se esistono infraorganismi (organi, cellule, molecole) e superorganismi (società, specie) come unità per se stesse. Si tratta però di un problema più metafisico che scientifico, tale da non poter essere affrontato in questa sede. Non possiamo tuttavia non essere influenzati da una ‛continuità' e ‛similarità' che andiamo constatando sempre più fra entità organiche di diverso livello, che abbiamo in qualche modo individualizzato.
b) Sistemi di interrelazioni associative
Le unità organiche ordinate in livelli di organizzazione sono qui considerate come sistemi, intendendo il sistema nel senso più generale, cioè come un complesso di componenti in interazione in modo da costituire un tutto collegato. Il concetto di sistema riunisce due aspetti importanti, da cui derivano criteri di conoscibilità: le proprietà delle unità componenti (aspetto statico) e le interrelazioni fra queste unità (aspetto dinamico). Evidentemente questo modo di considerare realtà complesse ci pone in una posizione molto utile dal punto di vista pratico, quando si tratta di passare dalla pura osservazione alle scelte operative.
Siamo quindi indotti a riunire nella stessa concezione di livelli di organizzazione tanto i sistemi omotipici, quanto quelli eterotipici, perché li riteniamo collegati in un grande processo evolutivo condizionato non soltanto dal fattore selettivo della lotta per l'esistenza, ma anche dal fattore selettivo associativo. Ci sembra infatti di poter ammettere una tendenza generale degli individui ad associarsi in unità complesse, che tendono pure all'individuazione in successivi processi concatenati di associazione-individuazione-associazione, i quali implicano processi di transizione da sistemi omotipici a sistemi eterotipici e di nuovo a sistemi omotipici.
In questo senso crediamo di poter generalizzare il concetto di sistemi di interrelazioni di carattere associativo.
c) Informazione e comunicazione
I tentativi di misurare l'organizzazione e quindi di valutare i suoi accrescimenti lungo la catena ininterrotta di forme viventi, in base alla possibilità di esprimere l'organizzazione in termini di ‛informazione', sono ancora all'inizio.
È di L. von Bertalanffy il merito di aver ideato col suo ‛organismo' una teoria della totalità intelligibile in senso funzionale e dinamico, che trova felici corrispondenze cibernetiche. È un'impostazione storicistica che ci conduce a riaffermare più che mai una ‛storia naturale' globale in contrapposizione a ogni più frammentaria concezione specialistica. Bertalanffy afferma che non vi è per la biologia una forma organica rigida portatrice di processi vitali, ma un flusso di processi che si manifestano in forme apparentemente persistenti. È l'idea dei sistemi aperti (a scambi con l'ambiente) e del dinamismo costruttivo di crescente organizzazione.
Si esprime in questo modo efficacemente la capacità creativa e inventiva attribuita alla vita per la realizzazione di crescenti possibilità di adattamento e per una sempre più vasta conquista dell'ambiente. La capacità costruttiva di strutture più altamente organizzate è connessa a processi propri dei sistemi aperti, intercalati fra un input e un output, cioè fra un ingresso e un'uscita di informazioni, processi che sono inseparabili da una regolazione destinata sia a conservare equilibri vitali sia ad arricchire l'organizzazione. La regolazione organica ha quindi caratteri comuni con la regolazione cibernetica.
Non pare inaccettabile che in questa concezione di un divenire organico come processo di organizzazione ascendente e quindi di crescente informazione, rientrino anche processi di associazione. Ogni episodio di associazione è condizionato dalla possibilità di ‛comunicazione' come presupposto di trasmissione di ‛informazioni' da sistema a sistema, così da condurre a gerarchizzazione di sottosistemi e a creazioni di sistemi più complessi.
3. Sistemi precellulari e protocellulari (livello prebiologico e procariotico)
Dedichiamo pochi cenni ai primi sistemi che si son venuti organizzando nella fase della cosiddetta ‛evoluzione chimica' e nella fase dell'evoluzione biologica più primitiva, dal momento che questi argomenti sono connessi con il problema delle origini della vita, trattato in un altro articolo (v. vita, origine della). Ci sembra opportuno rilevare, tuttavia, che sin dal loro più remoto divenire i sistemi organici e protobiologici si sono organizzati con sempre più complicati processi di aggregazione di parti più semplici in unità progressivamente più complesse. Ci sembra infatti di poter interpretare in questo senso anche quei primi processi di ‛condensazione' e di ‛polimerizzazione' che hanno prodotto le prime molecole organiche.
Nella medesima progressione vediamo succedersi anche quei processi di ‛coacervazione', nel senso di A. I. Oparin e di generazione di microstrutture (microsfere) nel senso di S. W. Fox, che hanno condotto ai primi abbozzi di un sistema biologico.
Si prospetta a questo punto il problema dell'‛organismo minimo', che dovrebbe costituire la fase conclusiva di una serie graduale di stadi, quale potrebbe essere l'‛emergenza progressiva' ipotizzata da C. Lecourier. Questo organismo minimo dovrebbe aver superato la fase dell'autoconservazione (stabilità di strutture termodinamiche) per soddisfare le esigenze di autoregolazione, di autoriproduzione e di variabilità ereditaria. R. W. Kaplan (1968) configura la necessità di un ‛aggregato eubiontico' con un gruppo basale di proteine, ma anche con un grande numero di molecole ‛inutili' non codificabili né rinnovabili, purché si instaurino sia uno sviluppo dei coenzimi in enzimi completi sia un meccanismo riproduttore che garantisca partizione in parti vitali.
Il problema dei Procarioti ancestrali può essere affrontato sulla base delle conoscenze sui Procarioti attualmente viventi, oppure fondandosi su documenti fossili, purtroppo abbastanza rari e problematici. Qualche maggiore chiarezza è stata invece acquisita in questi ultimi anni sul problema, peraltro molto difficile, del passaggio dalla cellula procariotica alla cellula eucariotica.
4. I sistemi cellulari (livello eucariotico)
a) Dalla protocellula alla cellula
Non sfugge ad alcuno la discontinuità non facilmente colmabile che esiste fra il livello strutturale della cellula procariotica e quello della cellula eucariotica; le diversità si possono riassumere in modo molto sintetico come nel prospetto riportato in questa pagina.
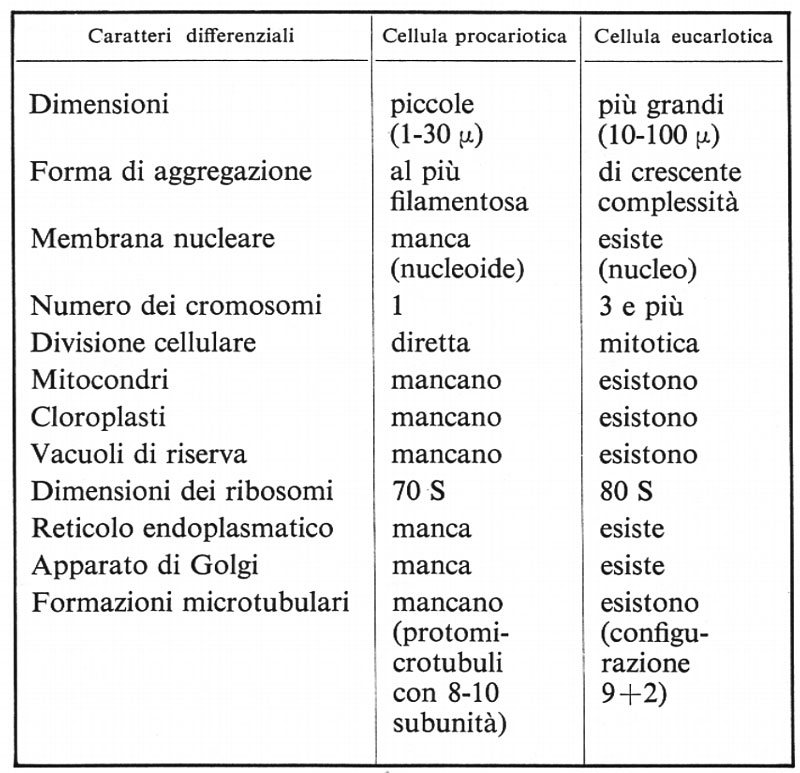
Queste numerose e importanti diversità permettono di dividere oggi gli organismi viventi in due vasti regni ben più giustificati di qualsiasi altra tradizionale suddivisione. Cade quindi anche l'alternativa animali-piante per lasciar posto a quella Procarioti-Eucarioti. Si comprendono quindi le recenti proposte di classificazione di H. F. Copeland (1956) che divide il mondo vivente in 4 regni: Monere (Procarioti), Protisti, Animali, Piante, e quella di R. H. Whittaker che considera 5 regni: Monere, Protisti, Piante, Funghi, Animali, sebbene queste divisioni non siano affatto di rango equivalente, ma siano comprensibili solo come ramificazioni di importanza molto diversa in schemi filogenetici.
Resta da interpretare il brusco passaggio fra Procarioti ed Eucarioti, che si verifica senza le transizioni che ci aspetteremmo in un continuum naturale. Non manca chi esprime dubbi sulla ‟santità dell'idea procariotica" (v. Bisset, 1973) ritenendo inconsistenti le diversità essenziali fra i Batteri e gli Eucarioti; i Batteri avrebbero solo semplificato strutture comuni. Ma questo rovesciamento di concezioni che possiedono un notevole corredo di argomenti richiederebbe motivazioni più persuasive.
La profonda diversità fra i due regni fondamentali induce a considerare la loro divergenza come la più importante di tutta l'evoluzione dei viventi. Una conferma è espressa quantitativamente da P. J. Mc Laughlin e M. O. Dayhoff (v., 1970): la divergenza fra Eucarioti e Procarioti è 2,6 volte più remota delle divergenze esistenti fra i regni in cui si sono distinti gli Eucarioti; è una constatazione che deriva dalla considerazione delle variazioni nel citocromo c e nell'RNA transfer (v. acidi nucleici).
Pertanto, la distinzione qui adottata di un livello procariotico e di un livello eucariotico supera per importanza tutte le altre successive distinzioni, che consideriamo condizionate e subordinate anche agli effetti dell'argomento dell'associazione biologica.
b) La teoria cellulare simbiotica
Non sono recenti le prime intuizioni e ipotesi di un'origine simbiotica della cellula eucariotica. Si può risalire a A. F. W. Schimper (1885), a R. Altmann (1894), a C. Merejkovsky (1905, 1920), a P. Portier (1918), a U. Pierantoni (1920), a I. E. Wallin (1927) e ad altri ancora, che più o meno esplicitamente si sono espressi in favore di una simbiosi di microrganismi (Batteri e Alghe unicellulari) entro le cellule di piante o animali, o di organismi di superiore organizzazione.
Una generalizzazione massima era raggiunta in un enunciato di P. Portier (v., 1918) quando affermava che tutti gli esseri viventi, tutti gli animali, dall'Ameba all'Uomo, tutte le piante, dalle Crittogame alle Dicotiledoni, sono costituiti dall'associazione, dall'inglobamento (emboitement) di due esseri differenti, che ogni cellula vivente racchiude nel suo protoplasma formazioni che gli istologi chiamano mitocondri, organuli che per lui non erano altro che Batteri simbiotici. Con la stessa enfasi si esprimeva C. Merejkovsky (v., 1920) a proposito delle piante, dicendo che una pianta, una quercia ad esempio, è un animale: un enorme animale nel quale vivono da parassiti, o piuttosto in simbiosi, una moltitudine infinita di piccoli organismi microscopici verdi, delle specie di Alghe unicellulari, delle Cianoficee, i granuli di clorofilla.
Ma soltanto in tempi più recenti si sono accumulati argomenti che possono servire di più valido sostegno. Una spinta decisiva è derivata dai progressi nella conoscenza delle strutture submicroscopiche e della biologia molecolare delle cellule. La sintesi più recente e più organica è offerta dalla teoria simbiotica di L. Margulis (1970), che utilizza e sviluppa concetti già maturati da S. Nass (1967), da L. Sagan (1967) e da altri studiosi. Riassumiamo da L. Margulis in uno schema comparativo le diverse implicazioni:
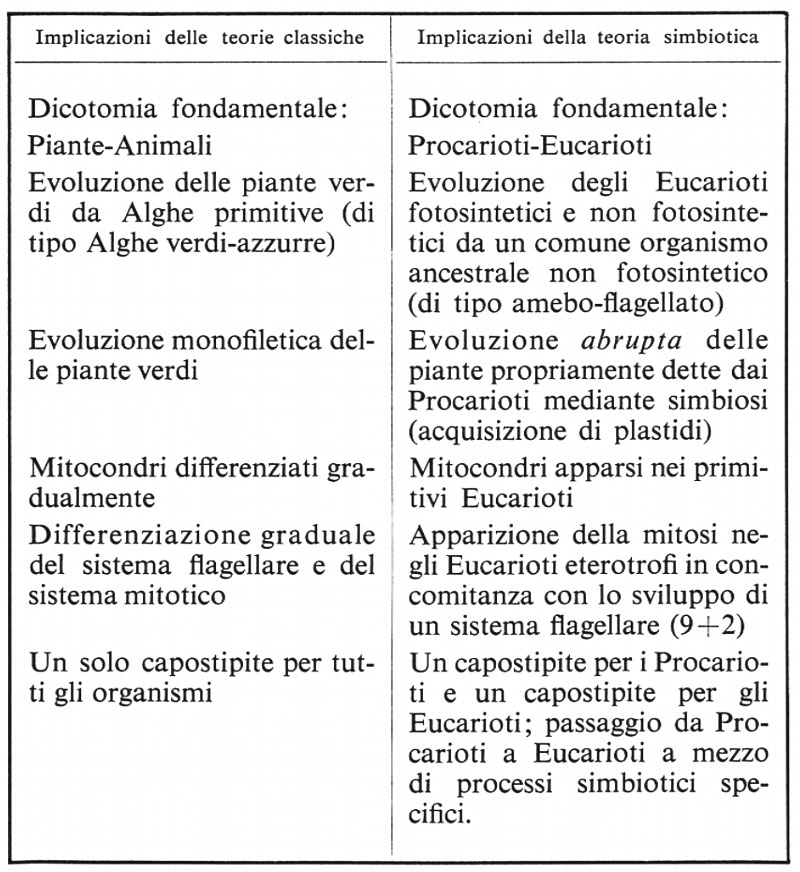
Possiamo altresì riassumere brevemente in alcuni punti principali gli argomenti sui quali viene fondata una teoria simbiotica dell'origine della cellula eucariotica.
1. Discontinuità strutturale fra Procarioti ed Eucarioti, già sopra evidenziata, e mancanza di organismi di transizione.
2. Discontinuità storico-geologica (senza altri esempi confrontabili) fra il Precambriano ‛microfossilifero' e il Fanerozoico ‛macrofossilifero', coincidente con profonde diversità ambientali: passaggio da atmosfera riducente (anossica) a ossidante, corrispondente a una ‛crisi dei Procanoti anaerobi'.
3. Affinità dei ribosomi dei cloroplasti e dei mitocondri con i ribosomi dei Procarioti (coefficiente di sedimentazione 70 S) e diversificazione dei ribosomi dal citoplasma delle cellule cui appartengono (caratterizzate da 80 S), con conseguenti diversità nel meccanismo di iniziazione della sintesi proteica (v. punto 7).
4. Similarità di moltiplicazione per semplice fissazione fra plastidi delle Alghe e organismi procariotici.
5. Presenza di DNA nei plastidi e nei mitocondri in forma di sottili filamenti, simili a quelli del nucleoplasma dei Procarioti; contenuto nel DNA dei mitocondri di molecole circolari del tipo noto soltanto per il DNA circolare di Virus e di Batteri.
6. Unimodalità dell'equilibrio nei gradienti di densità in soluzioni di CsCl nel DNA dei Procarioti e degli organuli cellulari in questione.
7. Concordanze di sensibilità ad antibiotici da parte dei sistemi che operano le sintesi proteiche (v. punto 3) nei mitocondri e nei Batteri; discordanze nei confronti del citoplasma: la cicloesimmide agisce solo sul citoplasma; il doramfenicolo e l'eritromicina agiscono sui mitocondri e sui Batteri, non sul citoplasma.
8. Presenza di microrganismi in molte cellule animali e vegetali (basterebbe ricordare la numerosissima casistica di P. Buchner, 1965, e di S. M. Henry, 1966-1967), con possibilità di sopravvivenza e di attività metabolica, spesso con sincronizzazione di divisione con l'ospite e con alto grado di coordinazione (per es. Mixotricha, protozoo con simbionti di tipo Spirochete e di tipo Batterio, studiato da A. V. Grimstone e L. R. Cleveland nel 1964).
9. Introduzione di nuove proprietà metaboliche a seguito di simbiosi: ad es. produzione di leghemoglobina da Rhizobium-Leguminose, di acidi organici e pigmenti nei Licheni, di derivati steroidi e terpeni in Celenterati-Zooxantelle, ecc.; introduzione anche di effetti morfogenetici (per es. formazione di noduli); dimostrazioni della possibilità di attivazione di meccanismi d'informazione aventi nella cellula una indipendenza l'uno dall'altro, come se la cellula non fosse un'unità genetica, ma un'accolta di organelli che vi si sono integrati.
c) Argomenti contro la teoria simbiotica
La teoria simbiotica non è accolta favorevolemente da tutti. La mancanza di dimostrazioni sperimentali dirette, l'insufficienza dei dati ottenuti con colture in vitro, inducono non pochi autori a respingerla, attribuendole solo una certa plausibilità suggestiva. Riassumiamo i principali argomenti che vengono portati contro di essa.
1. Esistono protoplastidi senza strutture lamellari, ma con ribosomi (evidenziabili ad es. in germinazione al buio) capaci di riprodurre alla luce le strutture del cloroplasto.
2. È possibile la derivazione dei cloroplasti da tre gruppi diversi di procarioti fotosintetici.
3. Sono problematici e molto discussi i risultati degli isolamenti e delle colture di organelli cellulari.
4. Sono presenti geni strutturali per la sintesi di enzimi fotosintetici e per la formazione di carotenoidi anche nel nucleo: i trasferimenti dal genoma endosimbionte al genoma nucleare implicano meccanismi difficili a concepirsi.
5. Non sembra così grande l'indipendenza fra Eucarioti e Procarioti, specialmente se si considerano i caratteri biochimici: infatti non è possibile escludere l'esistenza di ancestrali eucarioti aerobi (ad es. lieviti).
6. I più antichi microrganismi fossili interpretabili come Eucarioti risalirebbero a 1,4 miliardi di anni, cioè molto prima di un sufficiente accumulo di ossigeno nell'atmosfera.
7. È improbabile l'origine delle ciglia da Spirochete simbiotiche: una tale genesi implicherebbe un'inesistente omologia fra flagellina e proteine del microtubulo.
8. La cellula eucariotica avrebbe provato di possedere ben altra capacità di evoluzione, senza dover ricorrere ad apriorismi simbiotici.
Queste e altre obiezioni sono di diversa incidenza e più o meno facilmente discutibili. Dimostrano quanto sia ancora aperto il problema e come non sia consentito considerare del tutto consolidata una teoria simbiotica. Tuttavia non sono sufficienti a indurre l'abbandono di una concezione simbiotica che stimola notevolmente le ricerche sulle interrelazioni fra gli organismi lungo tutta la scala delle integrazioni morfologiche e fisiologiche.
d) Origine dei mitocondri
Tre problemi principali vengono affrontati dalla teoria simbiotica: l'origine dei mitocondri, dei cloroplasti e delle formazioni microtubulari. Tali problemi sono analizzati in base a un criterio evolutivo-selettivo, ammettendo una crescente mutua dipendenza e integrazione fra due simbionti.
L'origine dei mitocondri assume particolare interesse perché riguarda la totalità degli Eucarioti. Secondo la teoria simbiotica organismi eucariotici primitivi avrebbero ingerito, ma non digerito, protomitocondri aerobi. Si tratterebbe di Batteri (eu-Batteri) aerobi, gramnegativi, contenenti gli enzimi del ciclo di Krebs e il sistema citocromo-citocromossidasi per l'ossidazione dei carboidrati. Evidentemente questa simbiosi doveva richiedere un ambiente con ossigeno atmosferico, quindi doveva coincidere con la transizione, avvenuta nel Precambriano, fra atmosfera riducente e ossidante. In tale ambiente la simbiosi doveva diventare obbligata, eccetto che nel caso di alcuni Eucarioti primitivi (per es. lieviti tipo Saccharomyces) (v. Margulis, 1970), in cui si è avuta la sopravvivenza di una aerobiosi facoltativa molto specializzata. La produzione di ossigeno da parte di Procarioti clorofilliani doveva produrre una crisi: tutte le cellule che non potevano trovar rifugio in ambienti anaerobi specializzati dovevano adattarsi alla nuova atmosfera. D'altra parte, come già si è detto, dovevano esaurirsi le scorte di materia organica accumulate per processi abiogenici. Si ritiene che questa evoluzione ambientale e biologica abbia richiesto mezzo miliardo di anni; all'età di 1,2×109 anni tutta la vita della Terra dovette diventare tributaria direttamente o indirettamente della fotosintesi.
L'acquisizione dei mitocondri dovette costituire il primo momento dell'evoluzione simbiotica. Solo con la presenza dei mitocondri era possibile passare dal processo di scissione anaerobica del glucosio (processo di Embden-Meyerhof) al processo ossidativo dei carboidrati tipico degli Eucarioti (ciclo di Krebs). È noto che i mitocondri possiedono DNA e RNA specifici e possiedono capacità di autoduplicazione. L. Sagan (v., 1967) ammette che migliaia di mutazioni devono aver permesso l'evoluzione del mitocondrio.
Gli argomenti principali di affinità fra mitocondri e Batteri e di dissimiglianza nei confronti del sistema nucleo-citoplasma degli Eucarioti si possono più dettagliamente prospettare come segue: 1) dimensioni e massa; 2) membrane: presenza di subunità di 90 Å di diametro nei Batteri e nei mitocondri, omologie funzionali fra le cristae e i mesosomi, plasmalemmamesosomi e condriodi riscontrati nei Batteri; reazioni analoghe al dinitrofenolo, alla gramicidina e valinomicina; 3) accumulazione di ioni bivalenti legati a fosfati organici (granuli densi mitocondriali e granuli di volutina dei microrganismi); 4) similari proprietà di colorazione e fissazione delle fibrille di DNA; distribuzione unimodale dell'equilibrio in CsCl; presenza di molecole di DNA con configurazione circolare; analogie di peso molecolare; presenza di nucleasi diverse dalle nucleasi nucleari (per altri caratteri inerenti al DNA v. Nass, 1969); 5) ribosomi dei mitocondri più piccoli di quelli citoplasmatici; costante di sedimentazione rispettivamente di 70 S e 80 S (anche nei Batteri 70 S); 6) proprietà del tRNA comuni ai Batteri e mitocondri, diverse da quelle del tRNA dei sistemi eucariotici; 7) mRNA a vita più breve nei Batteri e mitocondri; 8) inibizione della sintesi proteica da cloramfenicolo nei Batteri, cloroplasti e mitocondri e altre analoghe concordanze.
e) Origine dei cloroplasti
Si sono già ricordati vari argomenti che suggeriscono la possibile derivazione dei cloroplasti per simbiosi di Alghe di vario tipo con le cellule eucariotiche. Sono particolarmente probanti le somiglianze strutturali (submicroscopiche) e biochimiche fra cloroplasti e Alghe verdi-azzurre in particore. Ma queste somiglianze si estendono in buona parte anche ai mitocondri, il che ha stimolato in vari autori l'idea di una comune origine dei plastidi e dei mitocondri (per es. v. Du Praw, 1968; v. Raven, 1970). L. Margulis (v., 1970) insiste sulla continuità genetica dei cloroplasti, essendo ben noto che essi contengono DNA, e che hanno capacità di dividersi e di distribuirsi, seguendo vari meccanismi, in concomitanza con la mitosi.
È particolarmente importante considerare la possibilità della derivazione dei cloroplasti dalle Alghe verdi-azzurre, le più somiglianti ai cloroplasti di molti Eucarioti. Sono frequenti gli episodi di simbiosi di questi Procarioti in cellule di Amebe, di Protozoi flagellati e anche in Alghe verdi che hanno perduto i cloroplasti (Glaucocystis, Cyanoptyche, Gloeochaete), e perfino in Ficomiceti (Geosiphon pyriforme). È indiscusso che in queste simbiosi le Alghe verdi-azzurre assumono una funzione analoga a quella dei cloroplasti. Un esempio di cloroplasti estremamente semplici, che denunciano una probabile origine da antichi simbionti procarioti (Alghe verdi-azzurre), è costituito dai cloroplasti delle Alghe rosse, che tuttavia sono assai più recenti (650 milioni di anni in confronto ai 3 miliardi almeno delle Alghe verdi-azzurre). I cloroplasti sarebbero originati da Alghe verdi-azzurre che hanno perduto la parete cellulare (sono noti però anche esempi di Cianofite nude).
È stato ipotizzato che si debbano ammettere almeno tre gruppi di Procarioti fotosintetici che avrebbero dato origine a simbiosi per vie fra loro indipendenti: a) Alghe verdi- azzurre, che possiedono clorofilla a, carotenoidi e ficobiline; b) Procarioti verdi ipotetici con clorofilla a e b, con carotenoidi accessori, ma senza ficobiline; c) Procarioti gialli pure ipotetici, con clorofilla a e c, vari carotenoidi, senza ficobiline (v. Raven, 1970). È un modo per esprimere la proposta di una multipla origine dei plastidi, estensibile anche ai mitocondri.
f) Origine dei microtubuli
Una terza categoria di organelli cellulari eucariotici è costituita da fini strutture, rivelate dal microscopio elettronico, che prendono il nome di microtubuli per la forma allungata e apparentemente cilindrica dei singoli elementi. Si concorda nel ritenerli organi ubiquitari ed essenziali per tutte le cellule eucariotiche.
Si tratta di filamenti del diametro di 210-250 Å riuniti in gruppi di 12-13, composti da unità globulari di circa 40 Å di diametro, tendenti a una certa rigidità. Esempi particolarmente noti sono le ciglia e i flagelli, le fibre del fuso mitotico, gli axopodi degli Eliozoi, le centrosfere. Questi e altri organuli costituiscono omologhi unificati da una struttura comune, visibile in sezione, caratterizzata dal modulo (9+2), cioè da 9 doppiette intorno a 1 paio centrale; è omologa anche la variante somatica (9+0) dei corpi basali dei flagelli.
La presenza degli omologhi (9+2) nella cellula eucariotica viene messa in relazione con l'insorgere di una ‛simmetria' spaziale della cellula, di una anisotropia che si manifesta nell'estensione o allungamento in direzione parallela agli omologhi stessi. Ne deriva un'importante implicazione nella produzione e conservazione della forma cellulare. Gli omologhi sarebbero quindi responsabili di importanti differenziamenti morfologici, connessi con differenziamenti chimici dipendenti da traslazione e trascrizione del codice genetico mediante produzione di particolari gruppi di proteine; si tratta di differenziamenti cellulari molto vari: ad es. i centrioli funzionali delle Alghe e di molti Funghi, le strutture corticali dei Protozoi, i tentacoli succhiatori di vari Protozoi, le increspature della pellicola dei Flagellati e dei Ciliati, gli axopodi, e simili.
Un caso particolarmente importante è la formazione del fuso mitotico (fibre cromosomiali, interzonali, astrali, tutte composte da microtubuli con diametro di 250 Å), il che indica la stretta connessione dell'evoluzione degli omologhi (9+2) con l'evoluzione della mitosi. Le fasi possibili di questa evoluzione sono dettagliamente proposte da Sagan (v., 1967).
La presenza di queste strutture negli Eucarioti viene attribuita, secondo la teoria simbiotica, all'acquisizione di un simbionte mobile spirocheta-simile da parte di una cellula ameboide aerobica (tipico l'esempio di Spirocheta in Myxotricha). I vantaggi acquisiti dalla simbiosi sarebbero stati rappresentati dall'acquisizione di mobilità, vantaggiosa per un'attiva ricerca di nutrimento, e dall'avviamento di processi mitotici.
L'origine di questo tipo di endosimbiosi viene collocata nel Precambriano dopo il passaggio dall'atmosfera riducente all'atmosfera ossidante. L'induzione di meccanismi di segregazione di acido nucleico avrebbe costituito una pressione selettiva: solo le cellule contenenti gli endosimbionti con il completo genoma (9+2) potevano essere avvantaggiate.
Si sta facendo strada la convinzione che gli organismi eucariotici siano complessi multigenomici; gli Eucarioti eterotrofi sarebbero trigenomici (nucleo-citoplasma, mitocondri, e omologhi 9+2), le piante sarebbero quadrigenomiche, perché contengono cloroplasti.
Alla luce della teoria simbiotica si prospetta una successione filogenetica di indubbio interesse (v. Margulis, 1970): a) organismi monogenomici: tutti i Procarioti con un genoma (genoforo-citoplasma); b) organismi digenomici: gli Eucarioti con due genomi (nucleo-citoplasma e mitocondri): forse le Amebe primitive, o nessuno? c) organismi trigenomici: gli Eucarioti con tre genomi (nucleo-citoplasma, mitocondri, complesso flagellare 9+2): Protozoi, Funghi, Animali; d) organismi quadrigenomici: gli Eucarioti clorofilliani con quattro genomi (in più i cloroplasti): Alghe eucariotiche e Piante superiori.
5. Sistemi pluricellulari preorganismici (livello cenobiale e plasmodiale)
a) Dagli Eucarioti ai Metafiti e Metazoi
Un momento importante dell'evoluzione è certamente quello della comparsa dei primi organismi pluricellulari. È un momento ancora carico di interrogativi che non trovano risposta. I documenti fossili sono molto controversi; i più semplici Metafiti e Metazoi viventi si prestano a interpretazioni discordanti. È difficile anche ricostruire le condizioni ambientali in cui si è realizzato questo trapasso evolutivo, che si considera determinante per il passaggio dall'era criptozoica all'era fanerozoica.
Sembra assai probabile che il trapasso da organismi unicellulari a organismi pluricellulari sia avvenuto gradualmente, attraverso stadi di vita coloniale, cioè attraverso aggregazioni più o meno lasse di individui monocellulari. Il problema assume aspetti diversi a seconda che si consideri l'origine dei Metafiti o dei Metazoi. Questi ultimi in special modo sono stati oggetto di diverse teorie, che hanno cercato di spiegare la comparsa di strutture multicellulari costituite da più tipi di cellule somatiche, adatte sempre più distintamente a un metabolismo eterotrofico ossidativo compatibile con l'accrescimento in dimensioni (ad es. il differenziarsi di una cavità digestiva e di una circolazione per l'utilizzazione dell'ossigeno).
Poiché non si conoscono Metazoi che siano adattati a condizioni permanenti di assenza di ossigeno, si è indotti ad ammettere che essi abbiano avuto origine in ambienti in cui si erano già estesamente imposti processi fondamentali di produzione di energia per opera di organismi aerobi procariotici ed eucariotici. Doveva essersi verificato un incremento di ossigeno nell'atmosfera, almeno nella misura dell'1% rispetto al livello attuale (indicato con PAL: Present Atmospheric Level), raggiungendo cioè quel livello che è stato chiamato ‛punto di Pasteur'. Pasteur aveva infatti notato che a tale punto si verificava in microrganismi un mutamento da metabolismo fermentativo a metabolismo respiratorio (v. Berkner e Marshall, 1964).
Sarebbe quindi fuori luogo esaminare i dati paleontologici che sono oggetto di una rassegna critica di P. E. Cloud jr. (v., 1968). I vari organismi medusiformi (Pseudomeduse) o anellidiformi (Pseudoanellidi) ritenuti Metazoi precambriani sono di molto dubbia interpretazione. Più impressionante appare la fauna di Ediacara: una comparsa massiccia di organismi indubbiamente metazoici planctonici o pelagici. L'età attribuita a questa fauna è di 684 milioni di anni; corrisponderebbe quindi al tardo Precambriano. Vi prevalgono ancora forme medusoidi, forme simili ad Anellidi attuali, e altre che parrebbero Echinodermi ancestrali, nonché organismi singolarissimi arborescenti e pennati.
Particolarmente importante e significativo è quel gruppo di forme che recentemente ha preso il nome di Petaloidi (Petalonamae). Sono stati trovati nella formazione Nama dell'Africa di SO che è tipica formazione Ediacariana. Costituiscono una transizione fra colonie di animali di tipo tubiforme e animali coloniali di struttura laminare-fogliacea. Da un tipo primitivo Fibularix a colonie a ramificazione dicotomica, si passa a un tipo ben più sviluppato ad Arborea con sistema simpodiale concresciuto con ramificazione pennata giacente in un piano, poi a un tipo Rangea con un sistema di tubi ramificati imbutiformi, quindi con formazione di un corpo cavo: è la tipica fase petaloide (v. Pflug, 1971). E stata constatata un'alternanza di generazione (Fibularix) con una forma vegetativa fissata al substrato e una forma generativa pelagica.
Per ciò che riguarda in special modo l'origine dei Metazoi si avanzano diverse ipotesi: 1) per evoluzione a partire da una colonia di Protozoi secondo l'ipotesi della Gastraea di Haeckel (1866); tale sarebbe l'origine di organismi scarsamente integrati come i Parazoi; la teoria è ripresa più recentemente da Jägersten (1972) col concetto di Bilaterogastraea; 2) per moltiplicazione e differenziazione di cellule, quindi partendo da un organismo unicellulare; tale il caso di Metazoi derivati da Protozoi ciliati o flagellati (Hyman, 1940), o da Protisti plasmodiali per ‛cellularizzazione' (Steinböck, 1937; Hadzi, 1953); 3) partendo dalle Metafite; opinione suggerita da fossili simili a fronde di Alghe (Charnia) e in genere dalle forme pennate della fauna di Ediacara (Hardy, 1953; Ford, 1958).
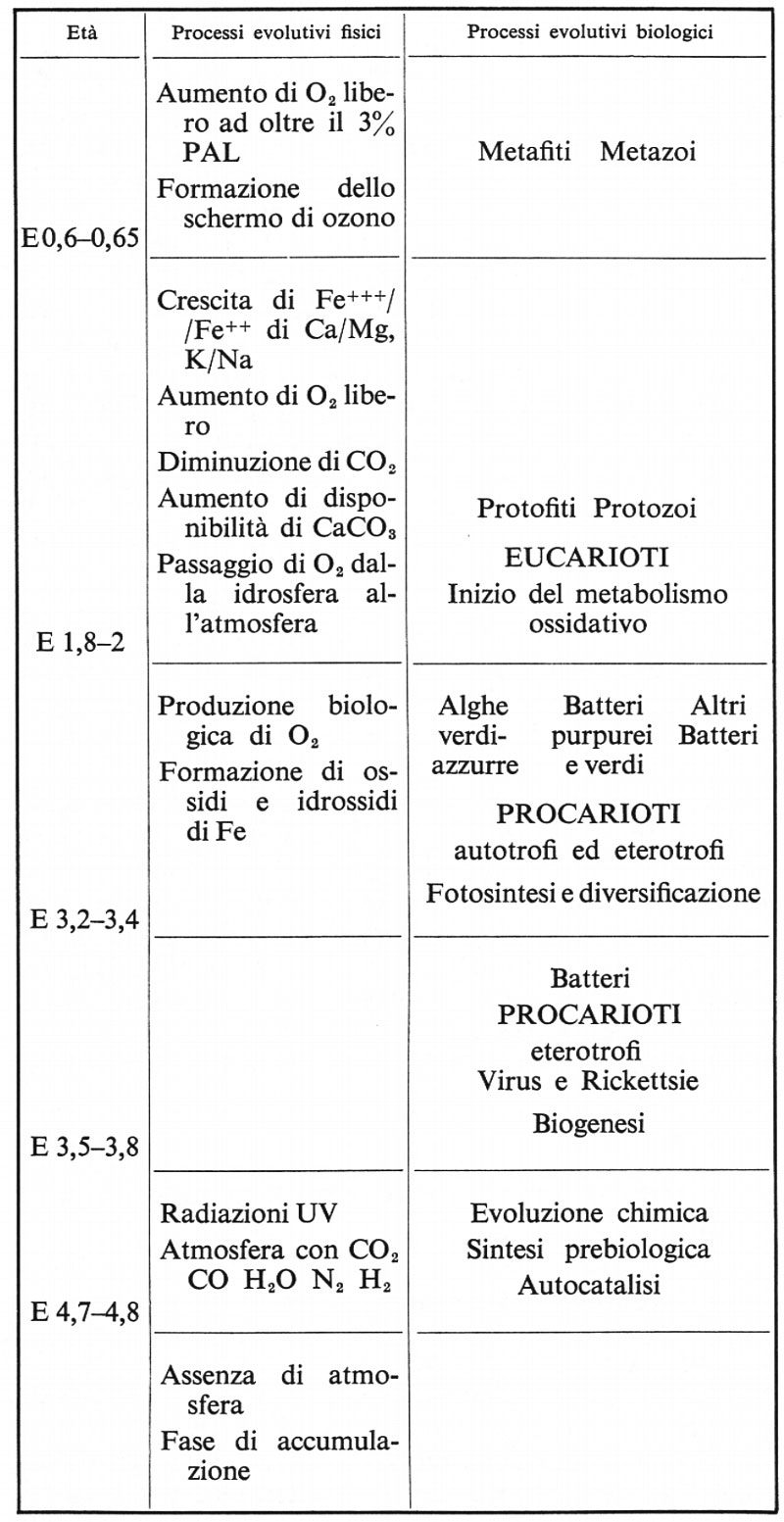
Cloud jr. (v., 1968) dà una sintesi filogenetica polifiletica dei più probabili processi evolutivi, fisici e biologici, dalla formazione della crosta terrestre fino alla comparsa dei Metafiti e dei Metazoi (v. oltre, tabella).
b) Dalla cellula al cenobio
Già Haeckel (v., 1866) trattando dei Protisti distingueva due tipi di organizzazione: una tipicamente individuale, unicellulare, il monobio (Monobion); e una costituita da gruppi più o meno coerenti di cellule omospecifiche, il cenobio (Coenobium). Fra i Cenobi distingueva diversi tipi, che potrebbero ancora aver qualche significato: 1) ‛Cenobi gregari', ammassi di cellule tondeggianti o senza forma determinata: ad es. le zooglee di Batteri flagellati, i Cenobi di Palmellacee, ecc.; 2) ‛Cenobi sferali', con ordinamento delle cellule su superfici sferiche o elissoidali: ad es. in molte Volvocali, nelle Spumellarie (Radiolari), ecc.; 3) ‛Cenobi arborali' o dendroidi, con cellule ordinate in complessi ramosi fissati al substrato; ad es. alcune Diatomee (per es. Gomphonema), alcuni Rizopodi, Flagellati e molte Vorticelle; 4) ‛Cenobi catenali' filiformi, con cellule ordinate in catene (seriate); ad es. Desmidiacee, Zignemacee e altre Alghe; molti Batteri; vari Protozoi (Rizopodi e Infusori). È una delle vie più comunemente seguite nell'evoluzione dei Metafiti.
Queste distinzioni sono peraltro fondate soltanto su un criterio morfologico-spaziale e fanno apparire i primi complessi cellulari soltanto come ‛somme' o addizioni di parti. Dipende da criteri di interrelazione funzionale se possiamo interpretarli come qualche cosa che è più di una somma di parti. Il più elementare di questi criteri può essere quello della separabilità più o meno facile delle cellule e la loro capacità di riprendere esistenza indipendente. Un ulteriore elemento è offerto dalla differenziazione delle cellule, ad esempio in una polarità del complesso (cellule basali e cellule terminali) o in evidenti divisioni di lavoro.
Un esempio celebre di cenobio interessa l'evoluzione dei Metafiti: il cenobio sferico dei Volvox, costituito da numerose cellule (fino a 25.000 e più), riunite fra loro da briglie plasmatiche, avvolte ognuna in una guaina separatrice gelatinosa. Le cellule sono biflagellate come nei Chlamydomonas e i flagelli sporgono alla periferia del cenobio e servono a imprimergli un movimento. È importante considerare che questo movimento è d'insieme, perché è coordinato in modo da diventare unidirezionale. Probabilmente interviene un segnale di natura chimica (analogo alla acrasina dei Mixomiceti), che determina una risposta univoca. È un esempio molto significativo di correlazione funzionale.
Analoghe coordinazioni intervengono anche presso microcolonie ruotanti di Procarioti; ad es. in Bacillus circulans, le cui colonie rispondendo a uno stimolo, si muovono sul terreno di coltura ora in verso sinistrorso ora in verso destrorso.
Altro esempio di cenobio è fornito dall'Alga verde Hydrodictyon utriculatum con struttura reticolata a maglie esagonali o pentagonali, costituita da migliaia di cellule (fino a 7-10 mila). È un materiale che si presta all'osservazione di fatti di correlazione intercellulare nelle fasi di sviluppo del cenobio e dell'allungamento delle cellule (v. Harper, 1908).
c) Plasmodi e pseudoplasmodi
Due singolari categorie di Protisti hanno attirato l'attenzione come possibili precursori di organismi pluricellulari: le Acrasiali e i Mixomiceti.
Le Acrasiali, dette simple slime molds o anche amoeboid slime molds, offrono l'interessante esempio di un'aggregazione di individui cellulari a costituire uno ‛pseudoplasmodio' o plasmodio di aggregazione. Poiché si moltiplicano asessualmente è stato detto che l'aggregazione sostituirebbe parzialmente la sessualità; ma è un riunirsi di cellule vegetali senza ricombinazioni genetiche (v. Larpent, 1970). L'integrazione nello pseudoplasmodio costituirebbe un'autentica unità fisiologica perché si manifesta un'organizzazione polarizzata (un polo anteriore della massa determina una proliferazione vegetativa, un polo posteriore dà luogo alle spore), un differenziamento cellulare (formazione del sorocarpo). Risulta evidente una capacità globale di regolazione che si manifesta nell'integrazione d'individui isolati in un aggregato, mediante un segnale chimico sperimentato con l'acrasina. Non è ancora raggiunto un livello pluricellulare, ma è una prima tappa verso di esso: cellule che conservano ancora la loro individualità entrano in autentica correlazione.
Un caso analogo è offerto, fra i Procarioti, dai Mixobatteri (Myxobacteriales), che attuano un'aggregazione in una sorta di corrente batterica in cui si differenziano microcisti e macrocisti. Alcuni considerano anche questo tipo di colonia uno pseudoplasmodio.
I Mixomiceti, detti true slime molds, formano invece veri plasmodi di fusione, perché le cellule invece di aggregarsi si fondono diventando degli ‛energidi'; sono caratterizzati da un'aplofase sessuata e da una diplofase sporifera. Il plasmodio presenta un movimento orientato corrispondente a una polarità: una proteina contrattile, la mixomiosina, affine all'actomiosina dei muscoli, ne sembra responsabile reagendo all'ATP con la contrazione. Plasmodi distinti possono fondersi sincronizzando, allo stesso modo in cui si fondono individui cellulari integrandosi fino a perdita dell'individualità, almeno spaziale, delle cellule. I Mixomiceti insieme con le Acrasiali e i Mixobatteri costituiscono materiali ideali per cogliere le prime fasi di quei processi morfogenetici che si risolvono in interazioni cellulari di relativa semplicità, ma che diverranno progressivamente più complesse negli organismi superiori.
d) I Mesozoi
Recentemente l'attenzione degli studiosi è stata attratta da un gruppo di organismi vermiformi costituiti da un numero abbastanza piccolo di cellule (da 20 a 30 circa), rimasti per lungo tempo misconosciuti. Si deve a F. Cavolini una prima sommaria segnalazione verso la fine del Settecento. Li descrisse meglio e diede loro il nome di Diciemidi il tedesco A. Krohn nel 1839. P. van Beneden li considerava organismi intermedi fra i Protozoi e i Metazoi e per questo diede loro il nome di Mesozoi. Caduta l'opinione che si trattasse di organismi semplificati, a causa della loro vita parassitaria nel rene dei Cefalopodi bentonici, si tende oggi (v. Lapan e Morowitz, 1973) a considerarli effettivamente rappresentanti di uno stadio evolutivo della vita pluricellulare.
Sono particolarmente interessanti i confronti operati su valori percentuali G+C (guanina+citosina) del DNA di Mesozoi e di altri organismi primitivi. È assai basso, circa 23%, nel genere mesozoico Dicyemennea, in Protozoi ciliati genere Tetrahymena (22-23%), ed anche nelle Acrasiali (22%), mentre nei Protozoi flagellati raggiunge il 45-60% e nei Platelminti il 35-50%.
I Mesozoi sono organismi differenziati con una distinta polarità, con un ciclo vitale complesso, e sono provvisti di abbondanti ciglia. Si collocherebbero verosimilmente in posizione affine ai Protozoi ciliati, rientrando nell'ipotesi evolutiva già ricordata (v. sopra, § a).
Rientrerebbero nello stesso tipo evolutivo anche i Parazoi (le Spugne), organismi a basso grado di coordinazione che dovrebbero aver avuto origine già nel Paleozoico, e secondo alcuni già nel Precambriano, da Protozoi Flagellati, verosimilmente Coanoflagellati (v. Hyman, 1940), per crescente specializzazione e organizzazione di una simmetria assiale. È una particolare via evolutiva già intuita da Haeckel quando osservò che le cellule coanocitiche delle Spugne potevano trasmutarsi in ameboidi, osservazione ripresa poi da altri e specialmente da E. N. Willmer (v., 1956), che descrive un dimorfismo opzionale da forma ameboide a forma flagellata in Naegleria gruberi. Quello dei Parazoi pare, tuttavia, costituisca un caso particolare non generalizzabile all'origine dei Metazoi.
e) Gli organismi pluricellulari come sistemi di comunicazione
Diventano sempre più numerosi e significativi gli esperimenti di riaggregrazione di cellule di vari organi animali, e in particolare di embrioni di Metazoi. Particolarmente indicativi per il nostro assunto sono quelli compiuti su Metazoi inferiori, per es. su Hydra (Gierer, 1972), che è diventato un organismo modello per lo studio del processo di riaggregazione. Cellule dissociate ricompongono aggregati polari con struttura assiale che non dipende da orientazione delle singole cellule, ma da un meccanismo d'informazione, come se le cellule dissociate conservassero una memoria della loro posizione originaria. Una sostanza inibitrice fornisce col suo gradiente di concentrazione un'informazione di posizione. Si tratta di un sistema ormonale costituito da cellule capaci di emettere segnali nello spazio extracellulare, segnali che possono raggiungere processi recettori di altre cellule.
È il caso della riaggregazione di cellule delle Spugne marine che avviene per opera di un ‛fattore di aggregazione' AF (Adhesive Factor di Humphrey e di Moscona) liberato dalla superficie delle cellule. Questo tipo di aggregazione è specifico, come ha dimostrato da molto tempo l'esperimento di E. B. Wilson che ha ottenuto da mescolanze in sospensione di cellule gialle e rosse di Spugne marine aggregati specifici dei colori separati. Lo sviluppo dei Metazoi viene quindi delineandosi come un processo di coordinate interazioni cellulari dovute a gradienti molecolari (v. Turner e Burger, 1973).
Recentemente ha assunto crescente interesse anche l'esistenza di specifiche aderenze cellulari. Le cellule dei Metazoi variamente organizzati possono comunicare fra loro non soltanto con sistemi ormonali, ma anche mediante trasmissione di segnali attraverso vari tipi di comunicazioni congiunzionali: membrane di congiunzione strutturate in modo da non permettere perdite esterne, passaggio diretto, ad esempio mediante plasmodesmi. Le membrane di congiunzione in particolare costituiscono zone di maggiore permeabilità che si creano nelle zone di contatto fra cellula e cellula, come nell'aggregazione già ricordata delle Spugne.
Un sistema di cellule così comunicanti permette un flusso da cellula a cellula di sostanze regolatrici dell'attività cellulare e della stessa attività genetica. Si tratta di un sistema con flusso incanalato che caratterizza organizzazioni chiuse; perciò ogni sistema cellulare interconnesso costituisce un canale di comunicazione.
I Metazoi sono quindi sistemi di comunicazione capaci di accumulare informazioni ordinatrici. Ciò che si ammetteva tempo addietro solo a livello umano, o al più a livello dei Vertebrati, viene riconosciuto ormai valido anche per i Metazoi inferiori. Alcuni autori insistono sulla facoltà di apprendimento proprio nel caso dei fatti associativi degli Invertebrati (apprendimento associativo). Si tratta di una condizione ambientale che può sovraimporsi allo stesso codice genetico e che costituisce una sorta di ‛cemento' fra i simbionti.
6. Sistemi interorganismici intraspecifici (livello superorganismico)
a) Le associazioni biologiche omotipiche
Vi sono autori che danno valore concreto di categorie o livelli ‛microcosmici' soltanto alle associazioni costituite da individui o corpi omotipici o omotetici, perché solo in questo caso è possibile concepire ‛totalità' aventi carattere ‛sociale'. Solo aggregati omotipici permetterebbero di considerare il tutto risultante come una macchina, in cui le singole parti realizzano una divisione del lavoro. Solo in aggregati omotipici si attuerebbe una centralizzazione e una correlazione funzionale.
La stessa concezione dinamico-evolutiva sarebbe applicabile solo nell'omogeneità di un livello di integrazione microcosmica. Solamente a un determinato livello avente questo carattere potrebbe essere accettabile una sistematica ‛naturale'.
Sembra tuttavia altrettanto degno di attenzione scientifica anche il confronto fra diversi livelli microcosmici, e fra questi e il macrocosmo. Si tratta di due aspetti diversi che tuttavia non sono oggetto di discipline così diverse, come si vuol credere, ma possono convergere utilmente in un medesimo ordine di ricerche integrate. Proprio i più recenti sviluppi dell'ecologia dimostrano quali profonde interferenze possono verificarsi fra fisiologia, genetica, biochimica, biofisica - discipline riferite ai microcosmi proto e metabiontici - e geografia, biocenologia, ecologia - discipline riferite al confronto fra microcosmi eterogenei e macrocosmo. Ma è molto più opportuno abbandonare il concetto di microcosmi e adottare quello più correntemente valido di sistemi.
b) L'associazione coloniale
L'aggregazione di individui prodotti per via agamica costituisce le cosiddette ‛colonie' e ‛cormi', nei quali si attua un collegamento materiale fra parti del tutto equivalenti, che possono staccarsi e vivere indipendenti. Esempi tipici sono le colonie di polipi dei Coralli, che hanno origine per gemmazione da un solo individuo nato da un uovo. Gli individui formatisi possono tuttavia restare permanentemente congiunti (tali le colonie permanenti di Cordylophora, Corydendrium, ecc.); possono anche differenziarsi realizzando una divisione del lavoro (tali le ben note colonie eteromorfe dei Sifonofori).
È significativo che organismi come questi fossero tempo addietro confusi coi vegetali, da cui il vecchio nome di zoofiti. Ha perdurato a lungo, del resto, un'analogia fra colonie di polipi e piante ramose, nelle interminabili discussioni sulla individualità o non individualità dei singoli polipi da un lato e dei rami e germogli delle piante dall'altro. A parte la poca opportunità di stabilire un'analogia fra oggetti tanto profondamente diversi, il problema ritorna a quella distinzione fra attualità e potenzialità che si è già ricordata (v. sopra, cap. 1, § b).
Risale a W. Goethe l'idea che le piante sono associazioni di parti elementari, che però possono presentarsi variamente metamorfosate, ora come foglie, ora come spine, petali, ecc.; onde L. Oken giunse a dire che ‟la foglia è un albero di forma speciale, i cui rami o fasci di fibre sono disposti in un piano [...]". Risale addirittura a Teofrasto l'idea che siano individui le gemme (‟la gemma cresce sull'albero come una pianta sulla terra"). E si potrebbe continuare con gli esempi, fino alle più recenti teorie sulla ‛pianta elementare' terrestre. Si tratta di punti di vista giustificati dall'ordine di ricerche cui appartengono, e dall'idea - come dice A. De Candolle - da cui ci si lascia dominare.
Giova sempre conservare coscienza della relatività e strumentalità di queste concezioni. Le stesse classificazioni in livelli di cui è un esempio quella data da Haeckel (cellule - organi - antimeri - metameri - persone - cormi - e vegetali) è una dimostrazione delle difficoltà che si incontrano a voler irrigidire e schematizzare una materia così fluida nella quale dobbiamo cogliere piuttosto momenti di un divenire continuo.
c) Le società animali
Accenniamo solo brevemente all'importante realtà biologica delle aggregazioni di animali in diverso grado solidali sia nello spazio sia nel tempo. Si tratta di passare dalla considerazione dei cosiddetti ‛infraorganismi' (cellule, organi) ai ‛superorganismi', fra cui figurano aspetti abbastanza concreti come le società animali e, secondo alcuni autori, anche aspetti astratti come le specie.
L'espressione più integrata di società animale implica la verifica di alcune condizioni essenziali: l'omotipia dei componenti, la divisione del lavoro, un ordine centralizzato e mezzi di comunicazione; implica anche una necessità o obbligatorietà, nel senso che un componente non può vivere separato permanentemente dalla struttura sociale cui appartiene. Ma ciò che si verifica, ad esempio, nel caso di certe categorie di Insetti sociali (api, formiche, termiti) rappresenta l'aspetto più tipico. Esistono infatti società ‛temporanee' (vespe, bombi) o annuali. E per gradi di più bassa gregarietà si giunge ad associazioni familiari (ad es. le ‛famiglie matriarcali' di Artropodi, realizzate per la protezione dei piccoli) e anche ad associazioni individualistiche pure temporanee (come i branchi, le mandrie, ecc.). Si tratta di un vastissimo argomento, su cui non possiamo soffermarci, ma che dimostra una grande ricchezza di adattamenti morfologici, fisiologici e di comportamento, che scuotono alquanto la pretesa di dare una definizione categorica a questo ordine di associazioni biologiche.
Considerazioni più generali trapassano al livello delle ‛popolazioni' (insiemi di individui che si moltiplicano fra loro in un determinato spazio), livello che diventa argomento di un'apposita branca della genetica, ma che interessa anche un'importante integrazione ecologica (fluttuazioni delle popolazioni e rapporti trofici) oltre che l'aspetto dinamico-evolutivo.
d) Livelli biosociologici
Essendo d'accordo sul principio che non esiste alcuna pianta e alcun animale solitari, e che l'isolamento di un individuo è un fatto accidentale e temporaneo, la regola è il raggrupparsi di individui fra loro eguali o diversi, in associazioni lasse o più o meno solidali. Si tratta di un fenomeno biologico universale che presenta tuttavia aspetti estremamente vari. Anche se consideriamo soltanto le associazioni omotipiche, i gradi di integrazione sono molto diversi nelle piante, negli animali e nei diversi gruppi vegetali e animali.
Le più semplici aggregazioni di piante sono costituite da individui che si sono moltiplicati vegetativamente e si sono resi indipendenti pur restando contigui: ad esempio pianticelle di Sphagnum a costituire cespi serrati in una torbiera. In questo caso l'aggregazione costituisce una difesa dal disseccamento. Si tratta di una vaga relazione ecologica.
Per gli animali, W. C. Allee elenca una serie di condizioni ‛subsociali' in ordine ascendente verso condizioni ‛sociali': a) assenza di vicendevole attrazione, tolleranza di una stretta prossimità (livello di tolleranza); b) reazioni vicendevoli in un sistema più o meno unitario (livello di integrazione); c) divisione fisiologica del lavoro; d) differenziazioni morfologiche associate a divisione del lavoro (società di Insetti); e) organizzazione in gran parte a mezzo di comunicazione simbolica (società umane).
Si tratta di un elenco di situazioni che non hanno fra loro alcun legame evolutivo; ognuna ha significato nella linea evolutiva cui appartiene: ad es. il punto d) a quella degli Insetti. Servono a dimostrare come una tendenza associativa, sia pur in diversi gradi, non sia un'eccezione, ma un fatto molto generale, anzi fondamentale.
7. Sistemi interorganismici interspecifici (livello simbiotico propriamente detto)
a) Gradienti di interrelazioni
Come a rigor di termini non esiste l'individuo isolato, si può dire che non esiste neppure un aggregato assolutamente puro, cioè rigorosamente omotipico. Anche una pianta che cresca isolata nel deserto costituisce in realtà un'associazione biologica con organismi vegetali o animali incospicui o almeno con microrganismi della sua rizosfera. I sistemi ‛eterotipici' costituiscono dunque la casistica più ricca e varia, che è stata oggetto di molte proposte di classificazione, ma che ha sempre dimostrato di adattarsi male a tutte le più ingegnose classificazioni. Per questo preferiamo riunire tutti i casi più salienti in un'unica trattazione, in un ‛livello simbiotico' da intendersi in senso lato.
Infatti si presentano all'osservazione tutte le gradazioni di situazioni, se si considerano i modi di aggregarsi di specie diverse. Un modo di esprimere i possibili rapporti fra gli organismi riuniti in sistemi può esser quello già proposto da Haskell (1949) e da Burkholder (1952) e ripreso più recentemente da Prat (v., 1961) e da Levy e altri (v., 1973), e che si può fondare sulla combinazione di tre tipi di interrelazioni: effetto non percettibile 0, effetto favorevole +, effetto nocivo −. Se si considerano anche le diverse possibilità che gli effetti siano esercitati da un organismo piccolo su un organismo grande o viceversa, si può giungere alla seguente tabella riassuntiva nella quale l'organismo debole è tra parentesi: (0), (−), (+). È inutile dire che si tratta di una tabella teorica, che prospetta tutti i casi teoricamente possibili.
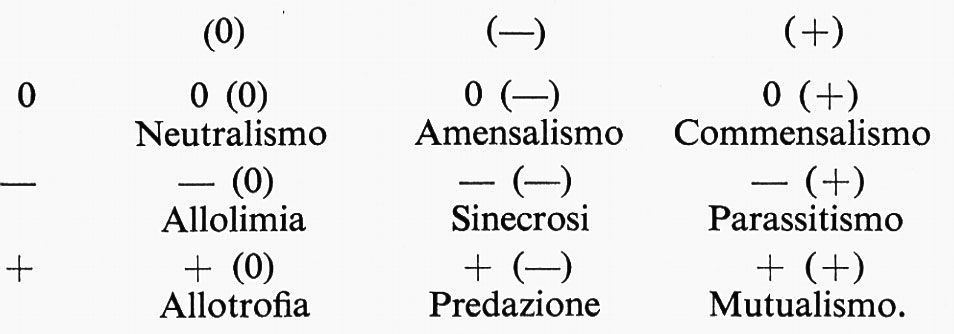
Queste combinazioni possono tuttavia costituire punti di riferimento, in nessun modo distinzioni assolute. Ognuna di esse infatti, considerata in esempi concreti, presenta transizioni verso le altre. È bene tuttavia diffidare da ordinamenti tabellari di questo tipo se si pretende, come alcuni autori vogliono, che essi rappresentino anche collegamenti evolutivi: essi possono al più rappresentare alcune affinità di situazioni. Una breve analisi della tabella può offrire occasione a considerazioni sulla relatività di tutte le possibili distinzioni.
0 (0): Neutralismo. - In assoluto si può dire che non esiste in natura, come del resto non esiste neppure la situazione diametralmente estrema del mutualismo. Si potrebbe porre il problema se l'‛epifitismo' può rientrare in questa categoria, dato che esistono minime interrelazioni fra i due componenti di questo modo di associazione. Ma anche in questo caso abbiamo una notevole varietà di situazioni: esiste un ‛epifitismo accidentale', non obbligato, che si attua quando specie indifferentemente capaci anche di vivere su roccia o sul terreno possono diventare ‛arboricole' (esempio tipico e universale il Polypodium vulgare; altri esempi varie Fanerogame disseminate dagli uccelli); esistono poi tipi di ‛epifitismo obbligato' o vero ‛epifitismo', in cui si attuano vari gradi di adattamento morfologico (per es. Orchidee con radici aeree, Platycerium, liane epifite, ecc.) o fisiologico (resistenza al disseccamento di Felci e Licheni). Ma si presenta allora la possibilità che l'epifita tragga qualche vantaggio dalla struttura della corteccia e quindi si verifichi una transizione al commensalismo, oppure che l'ingombro di epifite sia tale da produrre qualche danno all'albero: ad es. nel caso di Licheni sovrabbondanti nelle foreste troppo fitte o su alberi sofferenti; in questi casi si delinea un ‛parassitismo dello spazio' nel senso degli autori tedeschi e quindi un passaggio alla condizione −(+), sebbene sorga il dubbio che si tratti perfino di un ‛saprofitismo', se le piante ospiti sono così danneggiate da potersi considerare alla stregua dei tronchi caduti al suolo che sono spesso anche più ingombri da vegetazione di Licheni, Muschi, Epatiche. Vanno ricordati i casi analoghi di ‛epifillismo' e di ‛epitallismo' (per es. di Diatomee su Alghe laminari, di Epatiche su Licheni frondosi, ecc.). Non è escluso che possano rientrare qui anche casi di ‛epizoismo' (per es. Alghe epizoiche su Tartarughe, ecc.), ma anche questo è un modo di associazione che può condurre fino a casi di simbiosi di tipo comportamentale (behavioral symbioses) quando si attua fra animali.
È stato introdotto da alcuni il termine di ‛inquilinismo' come sinonimo del ‛parassitismo dello spazio', quindi per indicare un termine di passaggio verso la situazione −(+). Siamo tuttavia sempre di fronte a gradazioni talora inafferrabili: tale il caso dei rapporti fra Copepodi e Ascidie a seconda che la sede di Copepodi sia nelle cavità branchiali o invece nell'apparato digerente o nei tubi epicardici dell'ospite. Ci sembra che un eventuale criterio per distinguere l'inquilinismo dal parassitismo o dal mutualismo consista nell'obbligatorietà legata a modificazioni morfologiche e nel danno permanente all'ospite.
− (0): Allolimia. - Si tratta di una situazione piuttosto improbabile e rimane di significato teorico. L'organismo più forte verrebbe danneggiato senza che il più debole ne tragga vantaggio. Prat (v., 1961) ricorda il caso di un'ape che punge un uomo, ma si tratta di un esempio troppo occasionale, che resta ai margini se non al di fuori del problema della associazione biologica.
0 (−): Amensalismo. - Si tratta ancora di una situazione in cui l'organismo più piccolo viene danneggiato o eliminato senza vantaggi per il più grande. Potrebbe essere il caso dell'emissione di metaboliti tossici da parte di un organismo come un Penicillium a danno di Batteri. Potrebbe anche essere il caso di alberi di Eucalyptus, che accumulano al suolo una lettiera nociva al normale sviluppo della flora erbacea e anche allo sviluppo della flora batterica più ricca dei normali suoli forestali nelle regioni temperate. Non può sfuggire l'interesse ecologico di questo tipo di rapporti.
+ (0): Allotrofia. - Si tratta del caso in cui l'organismo più grande trae vantaggio senza danni per il più piccolo. I casi che sono ricordati a questo riguardo rientrano piuttosto in situazioni di ‛metabiosi'.
0 (+): Commensalismo. - Quando l'organismo minore convive con l'organismo maggiore in modo che il minore trova vantaggi di sostegno, di moto (‛foresi'), di protezione, di nutrizione e il maggiore non ne risente in alcun modo, si ha un'associazione con effetti unilaterali che prende il nome di ‛commensalismo'. Nel senso originario datogli da P. van Beneden (1876), è commensale un organismo che condivide col suo vicino lo spazio, non le risorse nutritive. Più recentemente il commensalismo è stato definito un'associazione di organismi, uno dei quali si nutre dei residui lasciati dall'altro. È tale la varietà delle definizioni che si impone un chiarimento ogni volta che questo termine viene usato. Sembra comunque che il commensalismo possa ritenersi, nel senso più corrente, un'associazione che realizza l'utilizzazione di risorse (non solo nutritive) in comune e che implica sempre effetti unilaterali.
Gli effetti si manifestano per lo più nell'organismo più piccolo in adattamenti più o meno complessi che possono essere morfologici (deformazioni, organi adesivi, ecc.), fisiologici (acquisizione d'immunità) e di comportamento (per es. il classico adattamento del Pilota - Naucrates ductor - ai movimenti degli squali con cui si accompagna).
Si verificano oscillazioni verso il mutualismo e il parassitismo, ma anche verso l'inquilinismo.
− (−): Sinecrosi. - Un caso estremo, che costituisce in realtà la negazione dell'associazione in senso costruttivo, è quello di una convivenza che conduce ambedue i partners alla degradazione e alla distruzione. Un esempio può essere costituito da germi patogeni che uccidono l'ospite e quindi anche se stessi, esempio che può esser interpretato come un caso estremo di parassitismo o come una situazione squilibrata di parassitismo.
− (+): Parassitismo. - Il parassitismo consiste in uno sfruttamento operato dall'organismo più piccolo a danno più o meno sensibile del più grande. Si tratta tuttavia di un danno che si mantiene entro certi limiti e che evita aggrava- menti che possono uccidere l'ospite; si instaura perciò un certo equilibrio, del quale analizzeremo gli aspetti e i significati. Un ‛parassitismo necrotrofico' è, come vedremo, un aspetto estremo, che conduce praticamente alla ‛sinecrosi'.
(+) −: Predazione. - La predazione costituisce pure un caso estremo, che si colloca ai margini del nostro interesse, per entrare piuttosto nei rapporti della ‛metabiosi'. La cattura e distruzione dell'organismo più piccolo da parte del più grande rientra, quindi, nella logica delle catene alimentari e nelle strutture dell'ecosistema come un aspetto di fondamentale importanza. La predazione è attività animale, salvo le poche eccezioni delle piante cosiddette carnivore.
Alcuni autori si svincolano dal concetto che il predatore sia l'organismo più grande e considerano predazione anche il comportamento della zanzara, che si nutre del sangue di Vertebrati. Evidentemente è eccessiva la nostra pretesa di voler inquadrare tutte le manifestazioni naturali in alcune categorie del nostro pensiero.
Un aspetto di predazione è la fagocitosi, che può esser considerata come l'epilogo di un parassitismo che provoca reazione violenta nell'ospite fino a eliminazione totale del parassita (per es. fagocitosi di endofiti fungini entro i tessuti di piante superiori). Un altro caso abbastanza singolare è quello di miceli fungini, che catturano con ingegnose trappole i Nematodi.
Diventano particolarmente delicati in molti casi i confronti della predazione col parassitismo. Vi è chi ritiene che il parassitismo possa derivare anche dalla predazione, ma anche chi crede che la predazione sia un aspetto estremo e drammatico del parassitismo (v. Raper, 1970).
Ultimamente si è parlato di ‛sistemi predatore-preda' (v. May, 1972) e si sono proposti modelli di alto interesse ecologico.
+ (+): Mutualismo. - Il mutualismo o associazione con aspetti di vicendevole vantaggio coincide con la ‛simbiosi in senso stretto' ed è, quindi, il culmine di un'evoluzione associativa, al di là della quale può collocarsi soltanto un fatto di ‛individuazione'.
b) Mutualismo e parassitismo
Risulta chiaro dallo schema precedente che mutualismo e parassitismo costituiscono aspetti di associazione molto vicini e culminanti. Prat (v., 1961) propone le serie evolutive:
predazione → allotrofia → simbiosi
sinecrosi → allolimia → parassitismo
e collega la simbiosi (=mutualismo) al parassitismo con l'ulteriore serie evolutiva:
parassitismo → commensalismo → simbiosi
su cui non ci sentiamo molto d'accordo, perché preferiamo considerare piuttosto oscillazioni dirette secondo il seguente schema:
parassitismo ⇄ mutualismo,
che rappresenta del resto i motivi di incertezza che spesso abbiamo quando cerchiamo di distinguere nettamente l'una condizione dall'altra. L'incertezza dei limiti ha indotto alcuni (v. Trager, 1970) a definire il mutualismo un parassitismo controllato: ma già A. De Bary a suo tempo riteneva il parassitismo un aspetto del fenomeno più generale della simbiosi, una simbiosi, secondo vari autori, con adattamento unilaterale, di uno solo dei due simbionti. Certamente è più logico pensare a una complessità di momenti adattativi della vita animale e vegetale che mal si lascia schematizzare entro il sistema di concetti: parassitismo-predazione-commensalismo-mutualismo; sistema che tuttavia è necessario continuare ad accettare per poter disporre di riferimenti semantici. Infatti nessun aspetto di mutualismo, neppure quello dei Licheni, è sfuggito a interpretazioni in senso parassitario.
Senza pretesa di assolutezza, che sarebbe contraria all'idea di un continuum di situazioni in largo senso simbiotiche, confrontiamo i caratteri che tendono a prevalere nel parassitismo e nella simbiosi e che in senso molto relativo si possono considerare distintivi.

Non è da attendersi una coesistenza di tutti questi caratteri, ma una prevalenza degli uni o degli altri, mentre non potrà sorprendere l'esistenza di situazioni intermedie o di transizione se, come pensiamo, non esistono casi di totale e definitivo parassitismo o mutualismo. Trager (v., 1970) pensa che in tutti i casi si abbia un equilibrio più o meno labile o solidificato fra forze potenzialmente distruttive ed evidenzia i vantaggi generali che ne derivano in senso selettivo. Questa situazione, latente anche nelle più evolute forme di mutualismo, è assai più minacciosa nel parassitismo.
È significativo che il parassitismo degeneri in ‛malattia' e in effetti distruttivi, specialmente a carico di piante coltivate e che si trovano in condizioni di squilibrio con l'ambiente. In condizioni naturali le piante, e anche gli animali, sono in grado di realizzare coesistenze equilibrate, sia pure limitatamente. I parassiti non hanno alcun interesse a danneggiare eccessivamente e tanto meno a uccidere l'ospite, perché nella maggior parte dei casi - in natura - finirebbero col danneggiare o distruggere se stessi.
Io vedo il parassitismo come un vastissimo processo di associazione biologica che pervade, più di quanto non si creda, tutto il mondo vivente, obbedendo a un impulso irresistibile di creazione di equilibri di coesistenza. Non si può sottovalutare questo processo perché ha certo esercitato un fortissimo influsso sulla stessa evoluzione degli organismi.
c) Sistemi intermicrobici
Le associazioni che si stabiliscono fra microrganismi, e in speciai modo fra Procarioti, sono di enorme importanza ecologica specialmente nell'ambiente del suolo e delle acque; ma non sempre si verificano in esse delle simbiosi nel senso pieno; sovente si tratta di episodi di metabiosi o di associazione extraorganismica (meno usato il termine ‛parasimbiosi' o quello ‛parabiosi').
Offrono esempi importanti anzitutto le ‛simbiosi interbatteriche'. La microbiologia è da tempo fortemente interessata agli antagonismi e sinergismi che si manifestano fra popolazioni di Batteri che crescono commisti. L'attenzione si rivolge specialmente alla produzione di sostanze di crescita oppure di antibiotici, da cui derivano fatti unilaterali 6 bilaterali (e multilaterali) di stimolazione o di inibizione. I fatti di ‛sintropia' sono numerosissimi: alcuni ceppi batterici producono fattori di crescita che compensano le deficienze di altri ceppi, che a loro volta sono in grado di compensare le deficienze dei primi. Esempi diventati ormai classici sono offerti dai Lactobacillus: un ceppo di L. leichmannii necessita di vitamina B12 e produce il fattore citrovorum; un ceppo di Leuconostoc citrovorum necessita del fattore citrovorum e produce vitamina B12: i due ceppi si compensano e possono vivere in ambienti privi di queste due sostanze. Numerose sono poi le associazioni compensatrici di Batteri anaerobi e aerobi nel suolo e in diversi substrati: gli aerobi sottraggono l'ossigeno favorendo lo sviluppo normale degli anaerobi.
Cresce anche l'attenzione sulle associazioni batteriche cavitarie di interesse medico. È opportuno osservare che si è giunti a considerare queste associazioni in senso fitosociologico e, quindi, a sviluppare anche concetti di micro-ecosistemi batterici (v. Wurch e Linder, 1968).
Diversamente dallo schema offerto (v. sopra, È a), in genere le associazioni batteriche tra diversi microrganismi presentano forme di dimensioni equivalenti. Potrebbe quindi valere uno schema come il seguente, in cui utilizziamo la terminologia più specifica proposta da Levy e altri (v., 1973):
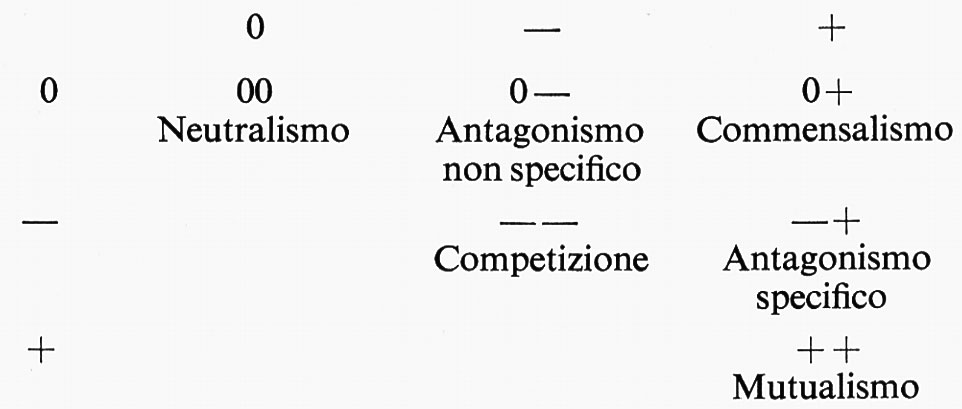
Sono possibili infatti anche casi di parassitismo (per es. Mixobatteri su Cianofite, Streptomyces su Cytophaga e Bacillus subtills su Caulobacter), di commensalismo, ecc. Tuttavia l'‛antagonismo specifico' è la situazione più ricca di aspetti e di graduazioni. Gli antibiotici costituiscono gli strumenti più validi e di uso più generale e nello stesso tempo più specifici (per es. il complesso della ‛bacteriocina').
Se tuttavia consideriamo i microrganismi in senso più lato, non limitato ai Procarioti, ma esteso ai Protozoi e Protofiti, può valere ancora lo schema precedente (v. sopra, È a).
Anche tra i Funghi si stabiliscono associazioni in cui dominano diverse gradazioni di parassitismo. Alcuni aspetti di transizione al saprofitismo introducono nuove difficoltà di giudizio, soprattutto si presentano aspetti di parassitismo obbligato, facoltativo, e anche occasionale, nonché casi di iperparassitismo (parassiti su parassiti: per es. il Cincinnobolus cesatii su Erysiphe); inoltre si verificano tutte le gradazioni dall'ectoparassitismo all'endoparassitismo: parassitismo esterno (Erysiphe), sotto la cuticola (alcuni Funghi epifilli), negli spazi intercellulari, con austori nel lume cellulare. A complicare ulteriormente le situazioni interviene anche l'eteroicismo, cioè il passaggio regolare da un ospite a un altro di specie diversa (per es. Uredinali), legato a vicende stagionali ma ancora oscuro soprattutto per ciò che riguarda la scelta degli ospiti; forse si tratta di una notevole sensibilità a fattori chimici.
Aspetti di simbiosi fungina sono piuttosto di carattere metasimbiotico o parasimbiotico, senza contatti fra gli organismi, ma con mutui vantaggi assicurati dall'emissione di sostanze di cui sono produttori o carenti (per es. Nematospora gossypii produce tiamina e necessita di biotina, mentre alcuni Polyporus producono biotina e necessitano di tiamina).
Un'altra categoria di simbiosi fra microrganismi è costituita dalle ‛sincianosi', associazioni di Alghe verdi-azzurre con altri microrganismi (per es. Chroastipes linearis nella capsula gelatinosa del flagellato aclorico Oikomonas syncyanoticus). A. Pascher (v., 1929) distingue nel comportamento delle Cyanellae una ‛ectocianosi' e una ‛endocianosi'; quest'ultima si verifica in cellule di Alghe e Protozoi di acqua dolce. Il mutualismo si realizza col conferimento della proprietà della fotosintesi da parte delle Cianelle e probabilmente con fornitura di composti azotati da parte dell'ospite. Quando manca un accumulo di materiali di riserva, secondo Pascher avrebbe già inizio una digestione e quindi una rottura dell'equilibrio. Non sfugge l'interesse di questo tipo di mutualismo come argomento in favore della teoria simbiotica della cellula eucariotica.
Le associazioni fra Batteri e Alghe sono meno ben conosciute. In special modo è stato evidenziato un possibile mutualismo fra Nostoc e Batteri azotofissatori. Sono stati trovati Batteri e Attinomiceti in colonie di Volvox, e pare sia stato accertato che sono necessari per il normale sviluppo dell'Alga.
d) Sistemi microbici-protozoici
Particolare importanza ed estensione assume l'associazione fra Batteri e Protozoi, perché si colloca pure alle possibili più remote origini delle simbiosi e perché viene invocata come argomento importante a favore della teoria simbiotica dell'origine della cellula eucariotica.
Un esempio di associazione è costituito dalla presenza di Batteri capaci di digerire la cellulosa in Flagellati endobiotici delle termiti, ma permangono dubbi sulla reale natura batterica dei corpuscoli osservati, e anche sul carattere obbligato di questa simbiosi.
Ricerche recenti (J. A. Breznak, 1973) riconfermano il ruolo azoto-fissatore di Batteri contenuti in Coptotermes formosanus, essendo stata rilevata la loro capacità di ridurre C2H2 a C2H4, catalisi caratteristica dei sistemi fissatori di azoto atmosferico. Più nota perché utilizzata in favore della teoria simbiotica della cellula eucariotica è la complessa situazione che si attua nel protozoo dell'intestino delle termiti Mixotricha paradoxa. Questo protozoo ospita alla periferia della cellula numerose Spirochete, che protendono all'esterno i loro prolungamenti sottili, i quali producono il movimento ondulatorio uniforme e regolato caratteristico della Mixotricha. Esistono, inoltre, Batteri endocellulari, la cui funzione non è conosciuta.
È interessante pure la presenza di Clorobatteri in Ciliati sapropelici: l'associazione è equilibrata se l'ambiente endocellulare - anzi più propriamente vacuolare - è anerobio, diventa squilibrata, con conseguente digestione dei Batteri, se insorge l'aerobiosi. Altro caso di simbiosi è quello di cosiddette particelle kappa, mi, lambda, epsilon, di indubbio carattere microrganismico, somiglianti alle Rickettsie, entro le cellule di Paramecium; questa associazione conferisce ai ceppi di Paramecium che sono interessati la proprietà di uccidere altri ceppi con cui si trovino a convivere. Le particelle in questione sono evidentemente di origine esogena e non fanno parte degli elementi congeniti dell'endoplasma; per molti caratteri assomigliano ai Batteri (dimensioni, scissione, composizione chimica, sensibilità agli antibiotici). Alcuni autori pensano che questa associazione realizzi condizioni simili a quelle del sistema lisigeno dei Batteri. Il problema è complesso e ancora aperto.
Corpi batterici o batterio-simili sono stati segnalati anche in un flagellato parassita, Crithidia oncopeltii; questi corpi si trovano appaiati nella parte posteriore del citoplasma e sono pure capaci di sintetizzare la lisina con processi batterici, proprietà di cui il flagellato è sprovvisto.
Sono analoghe le associazioni batteriche dei Mixomiceti e delle Plasmodioforacee. È stato dimostrato che la presenza di certi Batteri, come Bacillus subtilis, Escherichia coli, rende possibile una coltura durevole dei plasmodi dei Mixomiceti; la sterilizzazione li rende invece effimeri. Accade che la colorazione stessa del plasmodio sia dovuta alla presenza di Batteri cromogeni come Bacillus luteus, e altri. Se l'equilibrio si rompe il Batterio è digerito dal Mixomicete.
È singolare anche l'associazione di Batteri con Plasmodiophora brassicae, parassita delle radici delle Crucifere; i Batteri in questo caso determinano la putrefazione della radice infestata, favorendo la liberazione delle spore della Plasmodiofora e assicurandone quindi la dispersione.
e) Sistema Rhizobium-Leguminose
I Batteri attuano associazioni anche con le piante (Metafiti), con carattere simbiotico non sempre accertato, almeno in senso naturalistico. Rimane per molti aspetti oscuro il problema della presenza normale di Batteri, come di Attinomiceti e di Funghi, nelle cellule delle piante. Il loro ruolo è sconosciuto nonostante si stiano moltiplicando le ricerche. Sembra che l'origine da semplice contaminazione dell'aria sia poco rilevante, ma che si debbano considerare vari fattori: come l'inoculazione per opera di Insetti, o la presenza di ferite naturali, ecc. Non di rado si hanno florule di Batteri abbastanza complesse. Dai peduncoli dei frutti di pomodoro sono stati isolati ceppi di Enterobatteri, Corinebatteri, Micrococcacee e anche Pseudomonadacee.
I Batteri si presentano entro le cellule alquanto deformati (‛batteroidi'), ma introdotti in coltura riprendono gradualmente la forma normale attraverso stadi di forme a L o a X. Questi microrganismi, secondo H. Shanderl (v., 1948) e altri, non costituirebbero casi fortuiti o eccezionali ma avrebbero in ‛tutte' le piante - dai Funghi, alle Briofite, alle Tracheofite - la funzione di fissare l'azoto per la loro sopravvivenza. È una teoria generale che viene suffragata da numerose esperienze che si stanno moltiplicando. Il concetto di ‛istosfera' di Perotti e Zaffuto (v., 1923) già poneva in termini generali il problema di una simbiosi pianta-batterio in ambiente endocellulare.
Diventa più specifico il problema nel caso di Batteri che vengono raggruppati col termine di ‛rizobi' in quanto produttori di noduli a carico delle radici delle Leguminose e di altri gruppi di piante.
Le Leguminose sono, com'è noto, particolarmente ricche di azoto oltre che di altri elementi; esse sono quindi diventate importanti per la loro antica utilizzazione in agricoltura. I Batteri ‛rizobi' appartengono solo al genere Rhizobium (altri generi di Rizobiacee come Agrobacterium e Chromobacterium non sono distinti che per la mancanza di attività simbiotica formatrice di noduli).
I noduli radicali variano di forma e dimensione secondo le specie ospiti; sono per lo più tondeggianti, regolari, facilmente staccabili, da qualche millimetro possono raggiungere le dimensioni di un paio di centimetri. La loro struttura partecipa di quella delle radici secondarie ma è resa abbastanza complessa; però l'accumulo di cellule parenchimatose ipertrofiche affollate di Batteri costituisce la massa essenziale. Si tratta tuttavia di modificazioni a carico di tessuti radicali originari, non di nuovi elementi istologici. Si ammette che i noduli siano il risultato di un'azione bilaterale: la pianta favorisce selettivamente lo sviluppo dei Batteri; questi reagiscono e, valendosi di un meccanismo chimico ancora non chiaro, penetrano nei peli radicali. Procede allora l'infezione con moltiplicazione dei Batteri e loro chiusura in una struttura nodulare, che prende inizio da cellule di cui è stimolata la divisione e che sono tetraploidi o ottaploidi. Il nodulo nel suo complesso sviluppo presenta una suddivisione in zone con un addensamento mediano colorato più vivacemente in rosso per l'ammassarsi di rizobi: è questa la zona ove si attua la fissazione dell'azoto; il colore è dovuto a presenza di leghemoglobina, che è tipico prodotto del complesso simbiotico. Non è chiara la funzione biochimica di questa sostanza in rapporto con la fissazione dell'azoto.
L'associazione è mutualistica, ma non obbligata, perché i rizobi possono vivere liberi nel suolo; tuttavia, in condizioni di scarsità di azoto nel suolo, diventa necessaria alla vita del micro- e del macrosimbionte.
Anche in piante non Leguminose si verificano casi di nodulazione radicale: per es. in Alnus, Myrica, Hippophae, ecc. e anche in Gimnosperme come Agathis, Podocarpus, ecc. Tuttavia si verificano diversità strutturali e funzionali. Tende a verificarsi, ad esempio, la digestione del microsimbionte, analogamente a quanto accade nella simbiosi micorrizica. Si verificano anche (per es. in Agathis australis) casi di polisimbiosi per la presenza di due microsimbionti, probabilmente un Attinomicete e una Endogone.
f) Sistema micorrizico
L'associazione di ife fungine con tessuti delle radici di piante superiori, che ha preso nome di ‛micorriza', è stata oggetto di numerose interpretazioni ed è ancora oggi molto problematica. Si parla per lo più di simbiosi fra un micobionte e un ospite, ma anche di un parassitismo benevolo. L'interesse è anche di ordine pratico, data la grandissima importanza che questa associazione assume per gli alberi forestali che vengono notevolmente avvantaggiati per quanto riguarda la loro capacità di assorbimento di minerali (P, N e K in particolare) e dell'acqua stessa.
Resta valida la distinzione di sistemi ectotrofici ed endotrofici, sebbene abbia perduto un carattere assoluto. Nei sistemi ectotrofici le ife di funghi di vari generi (Agaricus, Amanita, Russula, Cortinarius, Lactarius, Tricholoma, Suillus, e anche Gasteromiceti e Tuberali) penetrano nella corteccia e si diffondono negli spazi intercellulari, creando un mantello fungino pseudoparenchimatoso (Hartig net) spugnoso. Si tratta di un'integrazione morfologica modesta in confronto a quella dei noduli delle Leguminose. Le radici con micorriza differiscono tuttavia da quelle normali per la loro forma raccorciata densamente ramosa-coralloide. La specificità è piccola specialmente per quanto riguarda i micobionti Basidiomiceti che si contano a centinaia, talora a più di cento su una medesima specie arborea.
L'integrazione fisiologica fra tessuto e micobionte si attua soprattutto con cessione di carboidrati dal primo al secondo, come è stato ampiamente provato con l'uso del 14C, e anche, sembra, con cessione di vitamine (tiamina e biotina) necessarie allo sviluppo del fungo. Il micobionte col suo mantello spugnoso facilita l'assunzione e l'assorbimento dell'acqua; esso fornirebbe anche indolo e sostanze con proprietà auxinosimili, che favoriscono lo sviluppo delle radici. Ma assume sempre più interesse anche un complesso di secrezioni, che non sono più soltanto della radice, ma anche del Fungo micorrizogeno (v. Rambelli, 1973), che costituirebbe quindi un'attività del complesso. Si parla quindi di una ‛micorrizosfera' con caratteristico effetto, diverso da quello di una comune ‛rizosfera'. I Funghi micobionti provengono dalla rizosfera entro la quale sono in competizione con altri Funghi del suolo sia saprofiti sia parassiti potenziali; favoriti nello sviluppo competitivo dalle secrezioni delle radici, si addensano intorno a queste e vi penetrano. Una volta che si sia creata la struttura micorrizica, continua un influsso più complesso che potremmo chiamare ‛effetto micorrizosferico' che regola la flora di microrganismi intorno alla radice, creando una protezione contro specie patogene, e probabilmente favorendo aspetti di biochimismo utile alla nutrizione delle piante.
I sistemi endotrofici si instaurano quando le ife fungine non creano un coerente rivestimento esterno alle radici, ma penetrano più addentro fra le cellule ed entro le cellule con caratteristici ‛arbuscoli' e ‛vescicole'. Sono assai pochi i Funghi che fanno parte di questi sistemi: Ficomiceti del genere Endogone, Basidiomiceti del genere Rhizoctonia. Si tratta di un'associazione a carattere spiccatamente antagonistico che può ammettere anche la distruzione delle ife per digestione da parte del macrobionte. Un caso particolarmente noto per il suo felice livello di integrazione è costituito dal sistema Rhizoctonia-Orchidee. Lo sviluppo dei semi delle Orchidee, che sono piccolissmi con poca riserva ed embrione poco differenziato, è consentito dall'endosimbionte, che pare comportarsi aggressivamente, mentre in realtà viene sfruttato dall'ospite cui cede sostanze stimolanti la germinazione (acido nicotinico, piridossina, riboflavina), e anche composti di carbonio, necessari a specie di Orchidee che sono notoriamente saprofite. Proprio perché consente l'utilizzazione di sostanze organiche complesse e difficili ad assimilarsi, come la cellulosa e la lignina, questo tipo di associazione differisce notevolmente dai tipi di micorriza degli alberi forestali. L'attività digestiva del macrobionte controlla efficacemente lo sviluppo del microbionte, al punto che alcuni hanno considerato quest'ultimo come parassitato dal primo (v. Harley, 1968). Sono state però isolate da bulbi di Orchidee due sostanze fungistatiche di natura fenantrenica - l'orchinolo e l'ircinolo - sintetizzate nelle cellule dell'ospite sotto lo stimolo dell'invasione fungina; sembra però che le sostanze antifungine elaborate siano poco specifiche.
La distinzione fra sistemi ectotrofici ed endotrofici è resa relativa da episodi di coesistenza dei due tipi come in Monotropa, o di transizioni come nelle Ericales. Non è facile l'interpretazione della micorrizia sia nell'uno che nell'altro caso. Secondo alcuni si tratta di casi di ‛parassitismo attenuato'; secondo altri di uno stato non patologico prolungato simile a quello che è proprio dei cicli di molti parassiti obbligati; secondo E. Hacskaylo (v., 1972) si tratterebbe di un ‛parassitismo reciproco', quindi di una sorta di reciproco sfruttamento, che è pervenuto a un certo grado di equilibrio e di interdipendenza caratterizzato da scambio ininterrotto di metaboliti. È significativa l'insistenza sul carattere parassitario più che sul carattere simbiotico-mutualistico.
g) Sistemi microbici-endozoici
La considerazione di sistemi microbici endozoici presuppone l'esistenza di sistemi di rapporti più lassi che alcuni hanno chiamato di ‛ectosimbiosi'. Esempi suggestivi sono stati ricordati più volte a questo proposito; ma troppo spesso sono inficiati da interpretazioni antropomorfiche. È il caso dei Funghi mirmecofili come le Rozites, Basiodiomiceti che costruiscono i cosiddetti ‛giardini delle formiche' nei formicai delle Atta del Sudamerica: le ife vengono tosate quotidianamente dalle formiche e reagiscono crescendo in forma di minuscoli cavolfiori (i Kohlrabihäufchen di F. H. Moeller). Analogo esempio è quello della flora fungina dei termitai (Basidiomiceti ‛termitofili' in special modo), che ha dato adito a supposizioni ingiustificate circa una loro coltivazione da parte delle termiti. Gli studi sui Termitomyces hanno messo in evidenza al più una flora caratteristica di cui è notevole la specializzazione ecologica. Altre associazioni lasse sono costituite dai Funghi del gruppo biologico cui è stato dato il nome di ‛Ambrosina', perché produttori di un micelio alimentare nelle gallerie che vari insetti (Scolitidi, Bostrichidi, ecc.) scavano nel legno di alberi o entro gli entomocecidi di varie piante. Si tratta di una mistura di specie fungine molto varie dei generi Dematium, Endomyces, Cephalosporium, Monilia, ecc. In tutti questi casi manca ogni accenno a integrazione fisiologica; tuttavia in non pochi casi si verifica una temporanea endosimbiosi non senza adattamenti morfogenetici, come le ‛camere di fermentazione' dei Tipulidi, le evaginazioni con lieviti dei Cerambicidi, dei Crisomelidi, come vari ‛micetomi' nei Silvanidi, ecc.
Esempi assai più significativi sono costituiti da fenomeni di simbiosi endozoica di Batteri e di Funghi. Si apre a questo punto un vastissimo capitolo che qui è impossibile anche soltanto sintetizzare, e che trova ampia trattazione nelle opere di P. Buchner e in speciale modo nell'ampia sintesi del 1965. Il grande capitolo si apre alle più moderne indagini dal 1909-1910, quando, indipendentemente, U. Pierantoni in Italia e K. Sulc in Moravia interpretarono come organo simbiotico il cosiddetto pseudovitellus, nome dato a formazioni a lungo rimaste enigmatiche, da molti anni segnalate negli Omotteri e in altri Insetti. Proprio il Sulc introdusse il nome di ‛micetomi' che era destinato a indicare durevolmente questi organi simbiotici. Da allora in poi gli studi si sono moltiplicati, rilevando molteplicità di casi, tendenza a polisimbiosi, implicazioni filogenetiche, modi di trasmissione ereditaria e via dicendo. Buchner offre una documentazione particolarmente ricca di endosimbiosi in Insetti che si nutrono di succhi vegetali (per es. Eterotteri con cripte ed escrescenze tubulari intestinali per Batteri simbiotici, Coccidi con micetociti, Fulgoridi con grande complessità di organi additivi, ecc.); in animali che si nutrono di sangue di Vertebrati, pure con micetomi, diverticoli esofagei, ispessimenti intestinali, ecc. Un esempio assai noto è quello dell'associazione di Batteri coi Blattidi caratterizzata da cellule ipertrofiche ripiene di ‛batteroidi', la cui presenza risulta necessaria al normale sviluppo dell'Insetto ospite, mentre lo stesso batteroide è incapace di sopravvivere indipendentemente dalla simbiosi. Gli esperimenti hanno dimostrato un singolare aspetto di integrazione: le Blatte possiedono la capacità, più propria di animali superiori, di utilizzare zolfo inorganico come sorgente di zolfo proteico, che però perdono se sono private dei loro Batteri simbiotici. È un fatto significativo che Blatte aposimbiotiche presentano tuttavia micetociti, alquanto più piccoli, evidente risultato di acquisizione della risposta cellulare all'infezione nella loro costituzione genetica.
Secondo Buchner si può ipotizzare una sequenza di complessità nelle risposte morfogenetiche che conducono ai più tipici micetomi: all'origine si potrebbero collocare i micetociti uninucleati (tipo Tachardiella), cui sarebbero seguiti micetociti multinucleati (tipo Orthezia), i gruppi di micetociti con organizzazione lassa e i pseudomicetomi (tipo Balaninus) e finalmente i micetomi propriamente detti, coperti da un epitelio, che pure si prestano a ulteriori suddivisioni a seconda che siano costituiti da micetociti monoo bi-nucleati o sinciziali, a seconda che siano mono-, di- o anche tri-simbiotici.
Un caso particolarmente interessante è costituito da episodi di luminosità animale attribuita da alcuni autori a organismi simbiotici. Pierantoni ha segnalato ammassi di granulazioni, da lui interpretate come Batteri, nei tubuli delle ghiandole nidamentarie accessorie di seppia; alcuni di questi, coltivati, davano luminescenza. Ma microrganismi simbionti isolati da organi effettivamente luminescenti di altre seppie non mostravano capacità di bioluminescenza; le ricerche sui Cefalopodi si sono moltiplicate, ma con esiti contraddittori. Buchner tuttavia difende l'ipotesi di Pierantoni con diversi argomenti, anche filogenetici. Le ricerche sono state estese da altri autori ai Tunicati luminosi, i Pirosomidi, che si ammassano dando impressionante spettacolo nelle acque del Pacifico australe, ai Pesci luminosi (per es. Anomalopidi), nonché ad animali terrestri come Lampyris noctiluca (v. bioluminiscenza).
L'integrazione fra simbionti nelle endosimbiosi si manifesta in alcuni fatti di regolazione da parte dell'ospite: il numero definito di cellule dei micetomi diventato specifico, la partecipazione della simbiosi alla differenziazione sessuale (prevale nelle femmine), il controllo della trasmissione in funzione dell'infezione delle uova. Un problema aperto a molte ricerche è costituito dal pleomorfismo dei Batteri, cioè dalle profonde modificazioni e combinazioni morfologiche con cui si presentano nei micetomi: la stessa rallentata riproduzione, l'inibita produzione di spore, sono pure argomenti che dimostrano una regolazione. Buchner giunge a generalizzare: il sito simbiotico è per se stesso un tipo generale di fattore di inibizione. La stessa limitazione spaziale in una sezione specializzata implica una severa distinzione fra cellule con simbionti e cellule sterili. Circa l'integrazione fisiologica, si è ancora sul terreno delle ipotesi: pare che nel caso di lieviti gli Insetti provvedano a fornire loro biotina e tiamina; pare che alcuni microsimbionti fungini (da Blastomiceti) possano provvedere alla decomposizione degli urati, sembra anche che possano produrre enzimi capaci di digerire gli zuccheri abbondantemente ingenti specialmente da Insetti che si nutrono di succhi vegetali. Si è parlato anche di possibile fissazione dell'azoto da parte dei Batteri tipo Azotobacter. Si è anche parlato di un'attivazione simbiotica della partenogenesi (v. Peleg e Morris, 1972). Pierantoni in base a questi dati sostiene l'esistenza di una ‛simbiosi fisiologica ereditaria', mentre altri ravvisano casi di ‛moderato parassitismo ereditario' (v. Müller, 1934).
h) Sistema lichenico e sistemi algali-endofitici
Il più classico esempio su cui si sono fondate le prime definizioni di simbiosi (v. De Bary, 1879) è costituito dall'associazione Alghe-Funghi che dà origine alla complessa struttura dei Licheni. L'argomento, vastissimo, ci consente di cogliere in questa sede solo alcuni aspetti che particolarmente interessano le attuali ricerche. Il primo aspetto più immediato è l'integrazione morfologica che si presenta in un tipo più primitivo omomero (mistura non differenziata di ife e cellule algali) e uno più evoluto eteromero (con una ordinata distinzione di zone con diversa struttura); a questo proposito G. D. Scott richiama l'attenzione sulla somiglianza impressionante fra struttura fogliare e struttura di una sezione di tallo lichenico (per es. di Sticta), e giunge perfino a proporre l'ipotesi che il modello fogliare delle Angiosperme possa aver avuto origine da un sistema simbiotico algale, che si sarebbe trasmutato in sistema di cloroplasti, estensione forse troppo ardita e prematura della teoria simbiotica della cellula eucariotica.
Più complesso è il problema dei rapporti a livello citologico che ha dato origine alle diverse interpretazioni sul sostanziale significato dell'associazione. A lungo si è data importanza alla presenza di austori aderenti o penetranti nelle cellule. La microscopia elettronica ha permesso di distinguere più propriamente austori intracellulari e intramembranosi, ma oggi si pensa che sia sufficiente in molti casi un contatto, un'aderenza delle ife alle cellule algali, per spiegare una diffusione di sostanze che genera lo sviluppo, del resto lentissimo, di un tallo lichenico.
Le Alghe che entrano nell'associazione appartengono a 26 generi: più numerose le Cloroficee (70 generi), seguono le Cianofite (8 generi) e le Xantoficee (1 genere di Eterotricali) (v. Ahmadjian, 1970). Il caso più interessante è costituito dal genere Trebouxia (Chlorococcales), la cui esistenza è accertata solo nella simbiosi lichenica e di cui si sono descritti due gruppi di forme fondati su caratteri del cloroplasto e dei tilacoidi; in particolare i tilacoidi sono connessi con lipidi, il che fa pensare che proprio in Trebouxia si sia evoluto un tipo di riserva caratteristico o più proprio della simbiosi lichenica (v. Ahmadjian, 1970). In coltura queste Alghe hanno dato produzione di zoospore e anche episodi di isogamia, fatti che, come è ben noto, sono inibiti entro il tallo del Lichene. Il problema della specificità delle Alghe simbionti sembra risolto in senso positivo; la presenza di Alghe in associazione secondaria va interpretata diversamente, cioè nel senso di formazioni di tipo galligeno.
Anche i Funghi sono stati isolati e coltivati e si è notato che differiscono assai poco dai Funghi liberi. Le colture progrediscono fino a creare talli molto simili a quelli del lichene da cui derivano (per es. Acarospora). Viene studiata con particolare attenzione la capacità che i micobionti in coltura hanno di produrre i loro fattori di crescita, e si è constatato che ciò avviene quanto più sono coltivati in condizioni che riproducono quelle prevalenti nel tallo del Lichene.
Il problema dell'integrazione fisiologica è stato da tempo risolto nel senso di un mutualismo molto ovvio: l'Alga cede al Fungo prodotti della fotosintesi, il Fungo provvede all'Alga sali minerali e protezione dal disseccamento e probabilmente dalle radiazioni luminose troppo intense. Questo mutualismo sarebbe equilibrato ‛nel senso più esteticamente attraente'. Ma molte sono le riserve al proposito. Alcuni attribuiscono una prevalenza al micobionte che si comporterebbe come parassita attenuato; infatti il Fungo impone la sua forma al tallo del Lichene e mantiene, al contrario dell'Alga, la sua capacità di riproduzione. È stato coniato anche il termine ‛ilotismo' (Warming, 1902) per esprimere una condizione di schiavitù benevola, come quella degli Iloti. Altri hanno considerato il micobionte come saprofita perché si nutrirebbe delle sostanze delle Alghe che muoiono progressivamente entro il tallo del Lichene (Elenkin, 1904). Finalmente F. e M. Moreau (1927) hanno ritenuto parassitario il comportamento dell'Alga, e pertanto hanno ritenuto che il Lichene costituisca una sorta di ‛cecidio' algale, quindi una formazione patologica; opinione che tuttavia si regge difficilmente.
È comunque accertata la cessione di composti del carbonio dall'Alga al Fungo; è probabile anche la cessione di prodotti di escrezione (oligodinamici), ma non è accertata quella di ormoni di crescita. Ahmadjian ammette anche la possibilità di fissazione dell'azoto atmosferico da parte di Nostoc a vantaggio del micobionte. Quest'ultimo fornisce protezione all'Alga, come generalmente si ammette, ma soprattutto in certe condizioni estreme, e potrebbe anche cedere acido ascorbico per favorire la fotosintesi. Dalle prove in coltura risulterebbe la possibilità di produzione di vitamine (biotina, tiamina) da parte del fitobionte, perché risultano necessarie allo sviluppo di un tallo lichenico.
Ricerche recenti non escludono aspetti di saprofitismo di Trebouxia e di Nostoc, che sono capaci di vivere a lungo su substrati glucosati, all'oscurità. Attira sempre più attenzione, poi, la preferenza dei Licheni per substrati ricchi di azoto (esempio tipico di florule di rocce con escrementi di uccelli); sembra che i ‛cefalodi', grumi di Alghe verdi-azzurre estranee che si localizzano a costituire simbiosi secondarie sul tallo lichenico, possano rappresentare organi di fissazione dell'azoto. Secondo alcuni i Licheni a Cianofite si assicurerebbero pure questa proprietà.
I problemi che i Licheni pongono a future ricerche sono quindi molto numerosi. In particolare si attendono interessanti risultati da un confronto più approfondito fra fisiologia dei simbionti singoli e fisiologia del complesso, e da una ricerca di specifici morfogeni chimici.
Un problema che richiede ulteriore supporto di indagini è la filogenesi dei Licheni. Questo problema ci conduce a considerare associazioni accidentali ed effimere di Funghi con Alghe, come quelle con micobionti Basidiomiceti, che prendono il nome di Coriscium viride e di Botrydina vulgaris, o ancora i Licheni leprosi del genere Lepraria, i cosiddetti ‛emilicheni'.
Ma esistono associazioni endofitiche di Alghe con altri vegetali, per es. con Epatiche dei generi Anthocero e Blasia, su cui i Nostoc formano cefalodi. Si tratta di una simbiosi che assicurerebbe alle Epatiche i vantaggi derivanti dalla fissazione dell'azoto atmosferico. Altro caso è la simbiosi di Nostoc con le radici di Cycas, caratterizzata da formazione di noduli simili a quelli delle Leguminose. Notevole è pure la presenza di Nostoc in rizomi di Gunnera; le cellule dell'Alga restano verdi anche sottoterra e rendono fotosintetici questi organi. Altri casi di Alghe verdi-azzurre si presentano in Lemna e in foglie di Aracee tropicali su cui si generano formazioni simili a galle.
i) Sistemi algali-endozoici
Molto ampia è la gamma di associazioni di Alghe con animali. Di particolare interesse sono i sistemi endozoici che riguardano i Protisti. Da molto tempo corpi gialli presenti in animali marini e corpi verdi presenti specialmente in animali di acque dolci sono stati riconosciuti di natura algale e denominati rispettivamente ‛zooxantelle' (Criptomonadinee) e ‛zooclorelle' (Clorelle e altre Alghe verdi). È ben noto il caso del Paramecium bursaria che ospita zooclorelle nel suo citoplasma; ma l'associazione cessa al buio e se viene dato abbondante nutrimento; la Clorella isolata in coltura differisce dalle specie libere per il metabolismo, perché libera una più abbondante quantità di carboidrati (maltosio) e per la capacità di infettare i Parameci. L'infezione avviene per ingestione da parte del Paramecio; osservazioni al microscopio elettronico evidenziano cellule di Chiorella dotate di una spessa membrana entro vacuoli pure con membrana. Un esempio di simbiosi con Criptomonadinee è rappresentato dall'associazione del ciliato marino Mesodinium rubrum con corpi rosso-bruni, che tuttavia vengono considerati piuttosto degli autentici cloroplasti; in questo caso l'origine da simbiosi con zooxantelle sembra verosimile, ma lo stato endosimbiotico nel senso più comune è superato da una integrazione funzionale che rende il ciliato un organismo autotrofo. Il riferimento alla teoria simbiotica della cellula eucariotica ricorre ovviamente (v. Taylor e Blackbourn; 1969).
L'esempio più classico di endosimbiosi algali di questo tipo è costituito dal celenterato Chlorohydra viridissima che ospita nelle cellule endodermiche delle cellule di Chlorella; si è osservata assunzione di prodotti della fotosintesi da parte dell'Idra in presenza di CO2. L'infezione si è dimostrata ereditaria perché si verifica spesso anche nell'uovo. La moltiplicazione delle cellule dell'Alga è contemporanea a quella delle cellule dell'Idra.
Un'ulteriore categoria particolare di ficobionti è costituita dalle Cianelle, Alghe verdi-azzurre. Dalla Cyanella paradoxa si pensa possano pure aver origine cloroplasti. Questa e altre ‛sincianosi' offrono esempi di associazioni molto solidali.
L'origine di questi tipi di cosiddetta ‛clorofilla animale' è identificata con l'ingestione da parte di animali erbivori. Le Alghe possono essere digerite o resistere e permanere ‛imprigionate' nell'ospite. Le situazioni sono molto varie: da uno sfruttamento parziale dell'Alga, si passa al suo sfruttamento totale, e anche alla sua digestione e distruzione. Casi di simbiosi pare si instaurino più difficilmente, ma più tipicamente, in animali carnivori. Si può dire, comunque, che si presentano tutte le gradazioni in integrazione, da associazioni labilissime, fino a pochi ma autentici casi di obbligato mutualismo.
L'olobionte che risulta dalle associazioni più strette acquista proprietà significative, come la produzione di grandi quantità di amido. L'integrazione diventa notevole in Convoluta roscoffiensis, in cui si attua un'infezione del- l'uovo che si è dimostrata necessaria allo sviluppo ulteriore.
Enorme interesse è stato attribuito all'associazione di zooxantelle nelle formazioni coralline degli atolli e della Grande Barriera. Ricerche sull'utilizzazione del bicarbonato delle acque e del CO2 metabolico per la formazione dello scheletro dei coralli hanno condotto a evidenziare la funzione delle zooxantelle nella rimozione dei prodotti catabolici, il che produrrebbe restrizioni fisiologiche e conseguenze negative sull'evoluzione che superi le più semplici forme ‛aermatipe' (v. Woodhead e Weber, 1970). Se si pensa che i tessuti dei coralli possono contenere decine di migliaia di cellule di zooxantelle per mm3, si possono intuire le vaste conseguenze di ordine ecologico.
l) Sistemi Metazoi-Metafiti
I rapporti fra Metazoi e Metafiti, di fondamentale interesse ecologico, includono una notevole varietà di aspetti associativi, quando i Metafiti diventano gli ospiti (macrosimbionti) in confronto ai Metazoi (microsimbionti).
Un primo vasto caso è quello delle galle prodotte da animali sulle piante (entomocecidi o zoocecidi), strutture con cui l'ospite tende a isolare, mediante barriere cellulari, il corpo estraneo costituito dall'organismo infestante. Sembra che si possa in questi casi parlare senz'altro di un parassitismo confinato, che tuttavia solleva interessanti problemi fisiologici e morfologici. Gli animali (‛cecidiozoi') sono prevalentemente Insetti, ma possono essere anche Nematodi e Acari; provocano la produzione di galle sugli organi più diversi della pianta, ma soltanto se interessano con punture e inoculazioni un tessuto meristematico. Si tratta di un processo altamente specifico con localizzazioni di sorprendente precisione e che conduce a forme di alta differenziazione. Ricordiamo questi tipi di parassitismo più che altro come estremo esempio di contrapposizione ai parassitismi che tendono a realizzare effetti di reciproca utilità. È il tipico caso della capacità, da parte del macrosimbionte, di realizzare una difesa invalicabile e quindi di rifiutare ulteriori rapporti favorevoli o nocivi.
Non possiamo a questo punto omettere un accesso al vasto campo di ricerche che da tempo si è aperto su interazioni chimiche fra piante e Insetti fitofagi; interazioni che implicano complessi fenomeni di stimolazione, di attrazione, di apprendimento, di memoria e in generale di comportamento. Si parla di sistemi chemiorecettivi degli Insetti da correlare con le diversità di sistemi vegetali, da cui derivano diversi modi di associazione. Basterebbe ricordare un caso ben noto: quello dell'ormone denominato ‛ecdisone', necessario per lo sviluppo della metamorfosi degli Insetti e che è sintetizzato nell'estremità anteriore del loro corpo (v. ormoni: ormoni negli invertebrati); recentemente autori giapponesi hanno scoperto in numerose piante varie sostanze consimili (‛fitoecdisoni'), fra cui un autentico B-ecdisone. La presenza è stata confermata anche in piante comunemente note, come il Taxus baccata e il Polypodium vulgare. I fitoecdisoni hanno dimostrato attività anche superiore agli ecdisoni animali, comportandosi come ‛superormoni' (v. Sondheimer e Simeone, 1970).
Siamo in campo di considerazioni più propriamente ecologiche (in particolare chemio-ecologiche), e ancor più vi entreremmo se prendessimo in considerazione i più complessi rapporti che si instaurano fra piante e animali nei fenomeni di impollinazione e di disseminazione; fenomeni che parrebbero appartenere, come vedremo, al vastissimo campo delle metasimbiosi, ma che in alcuni casi implicano strutture molto strettamente congiunte, come nell'esempio della Blastophaga collegata alla complessa biologia fiorale di Ficus carica.
m) Sistemi intermetafitici
Le associazioni fra Metafiti e Metafiti hanno pure carattere parassitario unilaterale −(+), cioè fra un microbionte parassita e un macrosimbionte ospite. Le ricordiamo molto brevemente per ricollegarci con fatti precedentemente considerati (per es. epifitismo) e quindi per completare il quadro, a nostro parere inscindibile, dei rapporti associativi fra pianta e animali. È oggetto di discussioni complesse il problema enigmatico dell'origine del parassitismo nei vegetali, e più propriamente dell'origine degli austori e dei tipi evolutivi di austori. Non pare sostenibile l'opinione che il parassitismo vegetale abbia avuto origine da causali rapporti fra pianta e pianta che non implicano solo complessi adattamenti anatomico-morfologici, ma anche aspetti di genetica delle popolazioni. È necessario infatti affrontare la spiegazione del meccanismo per mezzo del quale popolazioni di piante nei diversi phyla acquisiscono l'informazione genetica necessaria per la produzione diretta di austori. Problema che si allarga del resto a tutto il campo degli adattamenti simbiotici e parassitari (v. Kuijt, 1969).
Vi sono coloro che ipotizzano la derivazione di un ‛parassitismo epifitoide' dall'epifitismo propriamente detto che già si è ricordato più sopra; analogamente da un epirizoidismo deriverebbe un ‛parassitismo epirizoide'. Alcuni fanno derivare dall'‛emiparassitismo' un parassitismo totale o ‛oloparassitismo' come quello delle Lorantacee. Si tratta di ipotesi ben lontane da una solida giustificazione, ma che aprono un campo affascinante di indagini.
n) Sistemi intermetazoici
Anche le associazioni di Metazoi con Metazoi possono essere in buona parte tipici casi di parassitismo unilaterale. Basterebbe considerare i Vermi parassiti che occupano tanto spazio nei trattati di parassitologia con i loro cicli vitali spesso assai complicati. Ci limitiamo ad accennare i casi di simbiosi più o meno chiaramente riconoscibili.
Un esempio significativo di tendenza verso una simbiosi di carattere nutrizionale è stato recentemente illustrato fra il Crostaceo Echinoecus pentagonus e l'Echinoideo Echinothrix calamaris (v. Castro, 1969). È uno dei molti casi consimili noti a proposito dei Crostacei; in moltissimi di essi è stata riconosciuta una interdipendenza metabolica in forma di scambio di nutrimento, di enzimi e di fattori stimolanti lo sviluppo. L'Echinoecus utilizza come nutrimento, quindi come sorgente di energia, il micro-habitat dell'ospite (regione rettale), ingerendo i celomiceti, nonché i resti parzialmente digeriti delle alghe di cui si ciba l'Echinothrix e che si accumulano nel retto e nell'epitelio peristomiale. È un comportamento spiccatamente parassitario, i cui danni sono tuttavia equilibrati dalla notevole capacità di rigenerazione dell'ospite, che forse viene con qualche meccanismo stimolata.
Un altro esempio è fornito dalle cosiddette ‛simbiosi di pulizia' (cleaning symbioses). Esistono pesci capaci spesso di mutare vivacemente il loro colore, in modo da evidenziarsi fortemente nell'ambiente in cui vivono (Acanthurus achilles, Parupeneus trifasciatus, ecc.) all'avvicinarsi di pesci più piccoli (come Labroides dimidiatus) che si incaricano di ripulire il corpo dei primi da ectoparassiti, animali batterici o fungini che possono recare danno. L'ospite accetta la pulizia arrestando il movimento e passando quasi a una sorta di ipnosi. Anche feroci predatori, come i barracuda, si sottopongono a questo processo, astenendosi dal divorare i pulitori. Fatto questo non ancora esplicato, forse dipendente dalla natura talora non commestibile, talora velenosa delle carni del pulitore. Viene attribuita a questo tipo di associazione importanza evolutiva sia per i pulitori, sia per gli ectoparassiti; ma soprattutto costituisce uno dei più significativi esempi di adattamenti di comportamento, così numerosi nelle simbiosi animali. Anche nel caso di queste behavioral symbioses entrano in gioco fattori chimici attrattivi, che sono ancora scarsamente noti, e anche comportamenti attrattivi, come quelli che descrive Gohar (v., 1948) per le anemoni giganti del Pacifico, i cui tentacoli vengono ‛accarezzati' dai pesci associati (simpathetically caressing).
o) Sistemi polisimbiotici
Abbiamo più volte ricordato esempi di associazioni che annoverano più di due simbionti. Tipico il caso dei Licheni cefalodi che ospitano un terzo simbionte del genere Nostoc o Scytonema, o Stigonema; sistema multiplo che è stato interpretato come un apparato che garantisce anche la fissazione dell'azoto atmosferico.
Sono noti anche sistemi micorrizici multipli ectotrofici, la cui interdipendenza è tuttavia meno facilmente interpretabile. È il caso, ad esempio, della presenza di Batteri associati alla micorrizosfera di Pinus radiata in Australia. Sembra che i Batteri in questione siano azotofissatori e ricevano dal Fungo sostanze nutritive, come il mannitolo.
Un esempio di associazione ‛simultanea' su piante superiori è offerto da specie di Cuscuta, parassite di varie piante verdi, che sono associate in permanenza a Insetti, per es. del genere Smicronyx nel caso illustrato recentemente da D. M. Anderson (1970). Gli esempi si potrebbero moltiplicare.
Ma un aumento notevole di specie associate si riscontra in sistemi più lassi e innegabilmente capaci di realizzare certi equilibri antagonistici, come nel caso dei microrganismi (Batteri, Funghi e anche Alghe) che si possono trovare a convivere nella rizosfera di una determinata specie arborea.
A questo punto è facile proporsi la domanda se non possa esservi un passaggio insensibile verso associazioni multiple di Metafiti e Metazoi.
8. Sistemi ecologici (livello metabiotico)
a) Approssimazioni conoscitive
È uso corrente parlare di associazioni vegetali o fitocenosi, di associazioni animali o zoocenosi e anche di più complesse associazioni animali e vegetali insieme o biocenosi. Non si tratta in nessun modo di aspetti oggettivi che esauriscano un fatto associativo: sono approssimazioni conoscitive che conducono gradualmente a concezioni ecologiche caratterizzate da più valida integrazione. Hanno quindi un significato pratico e spesso costituiscono uno strumento che permette di rivelare e di indicare diversità ambientali, nel senso più lato, attuale e storico, determinate da fattori che spesso sfuggono all'analisi compiuta con strumenti microclimatici, con reattivi chimici e simili. La fitocenosi, in particolare, è per sua natura molto indicatrice; assai più che non lo siano le tradizionali specie indicatrici dell'autoecologia. Molto si è detto sui rapporti che intercorrono fra specie vegetali associate in un determinato ambiente. Non sono mancate interpretazioni organicistiche specialmente di autori americani e di forestali europei, che hanno rappresentato l'associazione vegetale come uno stato di organizzazione paragonabile a un'entità tassonomica. Specialmente la complessità e relativa stabilità di certe foreste ha offerto l'immagine di un'armonica composizione di strutture e di funzioni coordinata in una finalità diventata comune. All'altro estremo si colloca un'interpretazione ‛individualistica', che considera la fitocenosi come un mosaico di individui che si congiungono a comporre e scomporre tappeti erbacei, foreste e ogni altro aspetto collettivo del paesaggio vegetale. La realtà è certo più vicina a questa seconda interpretazione, ma è innegabile che in natura si ripetano, comunque, assetti che hanno una loro fisionomia ben distinguibile e un loro significato ecologico. Studiare sciami di individualità è molto impegnativo e si può realizzare soltanto con complesse trattazioni matematiche o con l'uso di modelli convenienti; si tratta di un metodo che si impone specialmente in formazioni, che presentano graduali transizioni nel tempo e nello spazio, inafferabili con i comuni rilevamenti statistico-fioristici (per es. la prateria nordamericana).
Ma nella stragrande maggioranza dei casi di vegetazione naturale e seminaturale le cosiddette associazioni vegetali si impongono all'attenzione e quindi si prestano a legittima individuazione, purché si considerino nel loro più sostanziale significato, cioè come stadi evolutivi, talora labili, talora praticamente (a misura di tempo umana) stabilizzati. Proprio nelle associazioni meglio definite si rivelano interrelazioni che alcuni autori considerano più o meno determinanti per il costruirsi e conservarsi di una struttura ‛associativa'; molto si è insistito specialmente sulle secrezioni radicali che condizionerebbero con fatti di antibiosi determinate coesistenze di specie vegetali, se non direttamente attraverso le più complesse microcenosi delle rizosfere. Ma sembra più plausibile che prevalga un gioco di antagonismi di carattere ecologico; ogni specie possiede una sua ampiezza ecologica rispetto a un determinato fattore ambientale, ma più specie che convivono modificano questa ampiezza, perché possono spostare altre specie o esser spostate esse stesse da un optimum specifico. Non potremmo tuttavia escludere fatti di reciproci influssi chimici (allelochimici e più in generale allelopatici).
Lo studio casuale delle fitocenosi conduce inevitabilmente a coinvolgere presenze animali, che possono essere studiate a parte come zoocenosi. Le zoocenosi possono pure essere altamente caratteristiche e quindi indicatrici di determinate situazioni ambientali, fino al punto di essere più espressive delle stesse fitocenosi. Accade purtroppo che le zoocenosi siano spesso così complesse e così difficili a conoscersi nella loro composizione faunistica - si pensi ad esempio alle faune del suolo - da rendere praticamente impossibile un loro esauriente rilevamento. Tuttavia anche il rilevamento di alcuni gruppi più significativi può permettere utili tipologie.
Ma la coesistenza immancabile di piante e di animali pone ben altri problemi di correlazione. Basterebbe ricordare i rapporti che le piante contraggono con Insetti pronubi o con altri animali impollinatori. Le piante dispongono di strutture adatte, gli Insetti oltre a strutture adatte allo scopo assumono comportamenti specializzati. Ricerca di nutrimento da parte degli Insetti, esigenza di un vettore del polline da parte delle piante, costituiscono le utilità scambievoli di questo mutualismo metabiotico. Se aggiungiamo i rapporti analoghi connessi alla disseminazione, parassitismi, saprofitismi, sulle piante, nella lettiera e entro il suolo, ne risulta un complesso di interrelazioni che giustificano l'integrazione conoscitiva a livello della biocenosi, cioè dell'insieme delle piante e degli animali in un determinato ambiente.
b) L'ecosistema
Le approssimazioni conoscitive dei sistemi parziali o parzialmente integrati tendono a raggiungere il livello dell'ecosistema, che è ormai una ben più oggettiva realtà ecologica, anzi l'unità fondamentale del discorso ecologico (v. ecologia).
Si giunge all'ecosistema integrando la biocenosi con le microcenosi del suolo, composte da Batteri, Funghi, Alghe, animali microscopici e macroscopici, e riunendo tutto il complesso vivente con il complesso non vivente costituito dalla parte di litosfera, idrosfera e atmosfera entro cui il complesso vivente si sviluppa. L'ecosistema costituisce, quindi, un sistema aperto in cui circola un flusso di energia ordinatrice e regolatrice, che parte dal sole, è captata dalle piante verdi (costruttrici), è trasmessa agli animali (consumatori), è restituita dai microrganismi (decompositori). Se si volesse pensare a un complesso organizzato, e sia pure di carattere prevalentemente metabolico, si potrebbe effettivamente farlo coincidere con un autentico ecosistema. Degli ecosistemi fanno parte integrante tutti i fatti di associazione simbiotica a tutti i livelli che abbiamo sin qui considerati.
c) L'ecosfera
L'insieme di tutti gli ecosistemi terrestri e acquatici compone il più grande e integrato dei sistemi: l'ecosfera. Preferiamo, con altri autori recenti, questo termine a quello di biosfera, perché quest'ultimo ha permesso, con la sua stessa etimologia, troppi equivoci; non pochi hanno considerato la biosfera come il complesso di tutti i viventi, prescindendo dal componente non vivo cui sono strettamente connesse tutte le forme viventi. Conserverei effettivamente il termine di biosfera per indicare solo la totalità vivente in questo ultimo senso.
Nell'ecosfera si attua la più grande integrazione che possa associare i viventi, e in tal modo, con tale fitta rete di interrelazioni, da far pensare veramente a una grande indivisibile organizzazione unitaria. L'indivisibilità è obiettiva: nessun essere vivente si sottrae a sia pur infinitesimi influssi da parte di tutti gli altri esseri viventi e da tutto il mondo fisico, entro una totalità planetaria. Basterebbe pensare ai fondamentali cicli biogeochimici che collegano tutti i viventi dal più piccolo Batterio, all'animale, alla pianta più gigantesca, a tutta la litosfera, idrosfera e atmosfera connesse alla vita.
A questo punto viene spontaneo riprendere il riferimento alla concezione di J. Needham (v., 1936): ‟Un netto cambiamento del livello di organizzazione spesso significa che ciò che erano delle totalità a un livello inferiore diventano parti del nuovo livello". È quanto si verifica nell'organismo individuale in senso stretto e quanto si realizza nell'organizzazione più generale a cui siamo pervenuti in questa sintesi. È quanto si è verificato in una continuità storica della natura, nella quale si innesta una continuità di storia umana.
bibliografia
Ahmadjian, V., Lichens in symbiosis, in Symbiosis (a cura di S. M. Henry), vol. I, New York 1966, pp. 35-97.
Ahmadjian, V., The Lichen symbiosis. Its origin and evolution, in Evolutionary biology (a cura di Th. Dobzhansky e altri), vol. IV, Amsterdam 1970, pp. 163-183.
Ainsworth, G. C., Sussman, A. S., The Fungi, 3 voll., New York-London 1968.
Allee, W. C., Animal sociology, in Encyclopaedia Britannica, vol. I, Chicago-London-Toronto 1959.
Allsopp, A., Phylogenetic relationships of the Procaryota and the origin of the Eucaryotic cell, in ‟New phytologist", 1969, LXVIII, pp. 591-612.
Altmann, R., Die Elementarorganismen und ihre Beziehungen zu den Zellen, Leipzig 1890.
Berkner, L. V., Marshall, L. C., History of oxygenic concentration in the earth's atmosphere, in ‟Discussions of the Faraday Society", 1964, XXXVII, pp. 122-141.
Bernard, N., Recherches expérimentales sur les Orchidées, in ‟Revue générale de botanique", 1904, XVI, pp. 405-451.
Bertalanffy, L. von, Théorie générale des systèmes, Paris 1973.
Bisset, K., This procaryotic-eucaryotic business, in ‟New scientist", 1973, XLV, pp. 296-298.
Buchner, P., Tier und Pflanze in Symbiose, Berlin 19302.
Buchner, P., Historische Probleme der Endosymbiose bei Insekten, in ‟Tijdschrift voor Entomologie", 1952, XCV, pp. 143-163.
Buchner, P., Endosymbiosis of animals with microorganisms, New York 1965.
Byrde, R. J. W., Cutting, C. V., Fungal pathogenicity and the plant's response, London-New York 1973.
Castro, P., Symbiosis between Echinocereus pentagonus and its host in Hawaii, Echinothrix calamaris, in Contribution n. 384 of the Hawaii Institute of Marine Biology, Hawaii 1969, pp. 229-247.
Caullery, M., Le parasitisme et la symbiose, Paris 1950.
Charles, H. P., Knight, B. C. J. G. (a cura di), Organization and control in Prokaryotic and Eukaryotic cells, Cambride 1970.
Cizek, F., Hodanova, D., Evolution als Selbstregulation, Jena 1971.
Cloud, P. E. Jr. Pre-metazoan evolution and the origins of the Metazoa, in Evolution and environment (a cura di E. T. Drake), New Haven-London 1968, pp. 1-72.
Cohen, S. S., Are/were mitochondria and chloroplasts microorganisms?, in ‟American scientist", 1970, LVIII, pp. 281-299.
Colosi, G., Le correlazioni nel mondo vivente, in ‟Bollettino del laboratorio di zoologia generale e agraria", 1954, XXXIII, pp. 83-99.
Davenport, D., Specificity and behavior in symbiose, in ‟Quarterly review of biology", 1955, XXX, pp. 29-46.
De Bary, A., Die Erscheinung der Symbiose, Strassburg 1879.
Deverall, B., Fungal parasitism. Studies in biology, London 1969.
Dobzhansky, Th., Hecht, M. K., Steere, W. C., Molecular biology and bacterial phylogeny, in Evolutionary biology (a cura di Th. Dobzhansky e altri), vol. II, Amsterdam 1970, pp. 104-155.
Drake, E. T. (a cura di), Evolution and environment, New Haven 1968.
Du Praw, E. J., Cell and molecular biology, New York 1968.
Egler, F. E., Vegetation as an object of study, in ‟Philosophy of science", 1942, CCXLV, pp. 245-260.
Ehrlich, P. R., Holm, R. H., The process of evolution, New York 1963.
Fisch, C., Aufzahlung und Kritik der verschiedenen Ansichten über das pflanzliche Individuum, Rostock 1880.
Fox, S. W., L'origine de la vie, in ‟Atomes", 1968, XXIII, pp. 348-355.
Giacomini, V., Considerazioni sul concetto di ‛associazione vegetale', in ‟Archivio botanico", 1952, XXVIII, pp. 1-25.
Gohar, H. W. F., Commensalism between fish and anemone, in Publications of the marine biology station, Chardaqa 1948, pp. 35-44.
Gola, G., La simbiosi cloromicetica, in ‟Nuovo giornale botanico italiano", 1948, LV, pp. 421-432.
Hacskaylo, E., Mycorrhiza: the ultimate in reciprocal parasitism?, in ‟Bioscence", 1972, XXII, pp. 577-583.
Hadzi, J., Die Abteilung der vielzelligen Tiere von den Einzelligen, in ‟Bioloski Glasnik", 1953, VII, pp. 3-10.
Haeckel, E., Generelle Morphologie der Organismen, Berlin 1866.
Hardy, A. C., On the origin of the Metazoa, in ‟Quarterly journal of microscopical sciences", 1953, XCIV, pp. 441-443.
Harley, J. L., The biology of Mycorrhiza, London 1959.
Harley, J. L., Mycorrhiza, in The Fungi (a cura di G. C. Ainsworth e A. S. Sussman), vol. III, New York-London 1968, pp. 139-171.
Harper, R. A., The organization of certain coenobic plants, in ‟Bulletin of the University of Wisconsin", 1908, CCVII, pp. 270-334.
Harris, H., Cell fusion, Oxford 1970.
Henry, S. M. (a cura di), Symbiosis, voll. I-II, New York 1966-1967.
Hyman, L. H., The Invertebrates, in The earth as a planet (a cura di G. P. Kuiper), vol. I, Chicago 1940, pp. 371-433.
Imms, A. D., A general textbook of entomology, London 1957.
Imshenetskii, A. A. (a cura di), Mycotrophy in plants, Jerusalem 1967.
Jaag, O., Symbiose und Krankheit bei den Pflanzen, in Verhandlungen Schweizerischen Naturforschenden Gesellschaft, Schaffhausen 1943, pp. 52-62.
Jägersten, G., Evolution of the Metazoan life cycle, New York 1972.
Kelley, A. P., Mycotrophy in plants, Waltham 1950.
Kitály, Z., Szalay-Marzsó, L. (a cura di), Symposium on the biochemical and ecological aspects of plant-parasite relations, Budapest 1971.
Knerer, G., Atwood, C. E., Diprionidsawflies. Polymorphism and speciation, in ‟Science", 1973, CLXXIX, pp. 1090-1099.
Kujit, J., The biology of parasitic flowering plants, Berkeley 1969.
Lapan, E. A., Morowitz, H., I Mesozoi, in ‟Le scienze", 1973, LV, pp. 26-33.
Larpent, J. P., De la cellule à l'organisme, Paris 1970.
Lecourier, C., L'origine de la vie, Paris 1970.
Levy, J., Campbell, J. J. R., Blackburn, T. H., Introductory microbiology, New York 1973.
Loewenstein, W. R., Communication through cell junctions, in The emergence of order in tissues and organs (a cura di M. Locke), New York-London 1968, pp. 151-183.
Mc Laughlin, P. J., Dayhoff, M. O., Eucaryotes versus Procaryotes. An estimate evolutionary distance, in ‟Science", 1970, CLXVIII, pp. 1469-1471.
Mani, M. S., The ecology of plant galls, den Haag 1963.
Margulis, L., Origin of eucaryotic cells, New Haven 1970.
Mattirolo, A., La simbiosi nei vegetali, Torino 1883.
May, R. M., Limit cycles in predator-prey community, in ‟Science", 1972, CLXXVII, pp. 900-902.
Merejkovsky, C., Über Natur und Ursprung der Chromatophoren im Pflanzenreiche, in ‟Biologische Zentralblatt", 1905, XXV, p. 593.
Merejkovsky, C., La plante considérée comme un complexe symbiotique, in ‟Bulletin de la Société des Sciences Naturelles de l'Ouest" (3e série), 1920, IV, pp. 17-98.
Metzner, H., Die Zelle. Elementarorganismus oder Symbiose?, in ‟Naturwissenschaften", 1973, LX, pp. 507-515.
Moeller, F. H., Mycorrhiza and other plant symbioses, in Symbiosis (a cura di S. M. Henry), vol. I, New York 1966.
Moreau, F., Les Champignons, Paris 1952.
Morton Wheeler, W., Social life among the Insects, Harcourt 1923.
Muller, W., Über die Pilzsymbiose holzfressender Insektenlarven, in ‟Archiv für Mikrobiologie", 1934, V, pp. 84-147.
Nass, S., The significance of the structural and functional similitarities of Bacteria and Mitochondria, in ‟International review of cytology", 1969, XXV, pp. 55-117.
Needham, J., Order and life, New Haven 1936.
Nutman, P. S., Mosse, B., Symbiotic associations, Cambridge 1963.
Oparin, A. I., L'origine de la vie sur la Terre, Paris 1965.
Oparin, A. I., Biogenesis and early development of life, New York 1969.
Ozenda, P., Clauzade, G., Les Lichens, Paris 1970.
Pascher, A., Über einige Endosymbiosen von Blaualgen in Einzellen, in ‟Jahrbuch wissenschaftlichen Botanik", 1929, LXXI, p. 386.
Paul, J., Cell biology, London 1965.
Peleg, B., Morris, D. M., Bacterial symbiote activation of Insect parthenogenetic reproduction, in ‟Nature new biology", 1972, CCXXXVI, pp. 111-112.
Perotti, R., Zaffuto, G., Sui bacilli radicali della ‛Calendula officinalis L.', in ‟Rendiconti dell'Accademia Nazionale dei Lincei", 1923, XXXII, pp. 94-98.
Pflug, H. D., Neue Zeugnisse zum Ursprung der höheren Tiere, in ‟Naturwissenschaft", 1971, LVIII, pp. 348-352.
Piaget, J., Biologie et connaissance, Paris 1967.
Pierantoni, U., Le simbiosi e le attività dei plasmi cellulari, in ‟Rivista di biologia", 1919, I, pp. 213-248.
Pierantoni, U., I recenti studi sulla simbiosi fisiologica ereditaria, in ‟Atti della Società Italiana per il Progresso delle Scienze", XIII riunione, Napoli 1924, pp. 209-226.
Pierantoni, U., De Lerma, B., Le radiazioni luminose di origine biologica, Modena 1939.
Ponnamperuma, C., Exobiology, Amsterdam 1972.
Portier, P., Les symbiotes, Paris 1918.
Prat, H., Metamorphose explosive de l'humanité, Paris 1961.
Preston, E. C. Jr., Pre-metazoan evolution and the origins of the Metazoa, in Evolution and environment (a cura di E. T. Drake), New Haven 1968.
Raff, R. A., Mahler, H. R., The non symbiotic origin of mitochondria, in ‟Science", 1972, CLXXVII, pp. 575-582.
Raffaele, F., L'individuo e la specie, Firenze 1943.
Rambelli, A., The Rhizosphere of Mycorrhizae, in Ectomycorrhizae, New York-London 1973, pp. 229-349.
Raper, J. R., Chemical ecology among lower plants, in Chemical ecology (a cura di E. Sondheimer e J. E. Simeone), New York-London 1970, pp. 21-37.
Raven, H., A multiple origin for plastids and mitochondria, in ‟Science", 1970, CLXIX, pp. 641-646.
Redfield, R., Levels of integration in biological and social systems, 8 voll., London 1942.
Reinert, J., Ursprung, H. Y. (a cura di), Origin and continuity of cell organelles, New York 1971.
Sabatini, D. D., Bensch, K., Barnett, R. J., Cytochemistry and electron microscope. The preservation of cellular ultrastructure and enzymatic activity by aldehyde fixation, in ‟Journal of cell biology", 1963, XVII, pp. 19-58.
Sagan, L., On the origin of mitosing cells, in ‟Journal of theoretical biology", 1967, XIV, pp. 225-274.
Schmid, E., Der Stufenbau der Materie und die biologischen Wissenschaften, in ‟Vierteljahresschrift der Naturforschenden Gesellschaft in Zürich", 1943, LXXXVII, pp. 261-269.
Scott, G. D., Plant symbiosis, London 1971.
Shanderl, H., Botanische Bakteriologie und Stickstoffhaushalt der Pflanzen, Stuttgart 1948.
Smith, D. C., Symbiosis of Algae with Invertebrates, London 1973.
Smith, F., The biology of Mycoplasmas, New York-London 1971.
Smith, N. G., The advantage of being parasitized, in ‟Nature", 1968, CCXIX, pp. 690-694.
Sondheimer, E., Simeone, J. B., Chemical ecology, New York-London 1970.
Stanier, R. Y., Some aspects of the biology of cells and their possible evolutionary significance, in Organization and control in Prokaryotic and Eukaryotic cells (a cura di H. P. Charles e B. C. J. G. Knight), Cambridge 1970, pp. 1-38.
Taylor, F. J. R., Blackbourn, D. J., Ultrastructure of the Chloroplasts and associated structures within the marine ciliate ‛Mesodinium rubrum', in ‟Nature", 1969, CCXXIV, pp. 819-821.
Trager, W., Intracellular parasitism and symbiosis, in The cell (a cura di J. Brachet e A. E. Mirsky), vol. IV, New York-London 1960, pp. 151-213.
Trager, W., Symbiosis, New York 1970.
Tribe, M., Whittaker, P., Chloroplasts and mitochondria. Studies in biology, London 1972.
Turner, R. S., Burger, M. M., Involvement of a carbohydrate group in the active site for surface guided reassociation of animal cells, in ‟Nature", 1973, CCXXIV, pp. 509-510.
Wallin, I. E., On the nature of mitochondria, in ‟American journal of anatomy", 1922, XXX, p. 203.
Wallin, I. E., Symbioticism and the origin of species, Baltimore 1927.
Whittaker, R. H., New concepts of kingdoms of organisms, in ‟Science", 1969, CLXIII, pp. 150-160.
Willmer, E. N., Factors which influence the acquisition of flagella by the amoeba Naegleria grubrei, in ‟Journal of experimental biology", 1956, XXXIII, pp. 583-603.
Wilson, E. O., The Insect societies, London 1971.
Woodhead, P. M. J., Weber, J., Evolution of the reefbuilding corals and the significance of their association with zooxanthellae, in International symposium on hydrogeochemistry and biogeochemistry, Tōkyō 1970.
Wurch, Th., Linder, R., Concept et dynamique des associations dans les biocénoses vaginales, in ‟Strasbourg médical", 1968, I, pp. 70-83.
Yonge, C. M., Symbiosis, in ‟Geological Society of America memoirs", 1957, LXVII, pp. 429-442.