
Endocrinologia
Endocrinologia
di Jean Roche
Endocrinologia
sommario: 1. Origini e tappe iniziali dell'endocrinologia all'inizio del XX secolo (1900-1925). a) Origine dei concetti di ormone e di correlazione funzionale ormonale. b) Fisiologia endocrina e endocrinologia medica. c) Prime tappe dell'endocrinologia moderna. 2. Scoperta di nuove secrezioni ormonali e di correlazioni endocrine di diverso tipo negli ultimi cinquant'anni. a) Secrezione di ormoni diversi da parte di uno stesso organo e differenziamento cellulare. b) Regolazioni ormonali complesse e funzione endocrina del sistema ipotalamo-ipofisario. c) Atri tipi di attività ormonali: ormoni tessutali, ormoni di Invertebrati, feromoni. 3. Biochimica endocrina. a) Costituzione chimica degli ormoni. b) Biosintesi, metabolismo e modalità d'azione degli ormoni. c) Endocrinologia medica e biochimica. 4. Considerazioni conclusive. □ Bibliografia.
1. Origini e tappe iniziali dell'endocrinologia all'inizio del XX secolo (1900-1925)
L'endocrinologia si è differenziata nel quadro delle scienze biologiche durante il XX secolo. Ne ha determinato la nascita un piccolo numero di nozioni che via via si rendevano disponibili dallo studio anatomico e fisiologico di organi diversi. Queste nozioni meritano pertanto di essere ricordate prima di ogni altra cosa.
a) Origine dei concetti di ormone e di correlazione funzionale ormonale
La descrizione anatomica di organi sprovvisti di dotto escretore, come la tiroide o le capsule surrenali, e il fatto che alcuni di essi presentino una struttura che ricorda quella delle ghiandole secretorie ha condotto al concetto di ‛ghiandola chiusa'. La loro funzione è stata, all'inizio, spiegata solo attraverso ipotesi e H. J. Muller, F. O. J. Henle, R. A. Kölliker hanno dedotto dalla loro struttura che essi dovessero modificare la composizione del sangue. Il concetto di ‛secrezione interna' ha avuto origine dalla scoperta di Cl. Bernard, nel 1855, che il fegato secerne glucosio, proveniente dalla glicogenolisi, nel sangue delle vene sovraepatiche. Tuttavia l'idea del prodotto di un processo secretivo direttamente riversato nel sangue circolante fu inizialmente così circoscritta che soltanto trent'anni dopo la sua formulazione ebbe quell'effetto che ha assicurato il suo sviluppo. Infatti Bernard scriveva ancora nel 1867: ‟Le secrezioni interne sono generalmente delle secrezioni nutritive, che preparano sostanze a rapida utilizzazione destinate ai fenomeni di nutrizione di elementi istologici: glucosio, albumina, fibrina" (v. Gley, 1921, p. 21). Senza dubbio egli considerava, come i suoi contemporanei, che ogni regolazione funzionale non può essere che nervosa. Come scriveva Gley (v., 1921): ‟Con Brown-Sequard tutto cambia. La teoria, come addormentata per più di trent'anni, si rianima, quindi si evolve rapidamente". In effetti, dal 1889 al 1891, egli introdusse in questa teoria un'idea del tutto nuova, quella che ‟molti organi secernono nel sangue delle sostanze che agiscono in modo elettivo su altri organi. Le diverse cellule del sistema vengono così rese intimamente interdipendenti per mezzo di un particolare meccanismo, oltre che dall'azione del sistema nervoso" (ibid., p. 27). Era così nata l'idea di una correlazione funzionale umorale tra gli organi per mezzo dei prodotti di secrezione interna; essa si impose in pochi anni e da allora animò la fisiologia endocrina. Per illustrare l'importanza del periodo che va dal 1900 al 1925 circa, basti pensare che il termine di ‛ormone' fu proposto nel 1905 dal fisiologo inglese E. H. Starling e quello di ‛ghiandola endocrina' dal clinico italiano N. Pende nel 1912. È opportuno ricordare anche che il termine ormone designava inizialmente un prodotto di origine tissutale, la secretina, e non una sostanza prodotta da una ghiandola chiusa. W. M. Bayliss e E. H. Starling avevano infatti stabilito nel 1902 che, sotto l'azione del chimo gastrico acido, la mucosa duodenale secerne nel sangue questa sostanza, che stimola la secrezione del succo pancreatico. La scoperta della secretina e del suo trasporto per mezzo del sangue, più di dieci anni dopo che era stata formulata la teoria della correlazione funzionale umorale, ha dato a quest'ultima una nuova conferma, tanto più importante dal momento che il primo ormone puro, l'adrenalina, era stato isolato nel 1901 dalla midollare surrenale, da T. B. Aldrich e da Y. Takamine, senza che ne fosse ben chiara la funzione. L'endocrinologia si sviluppa quindi dalla correlazione della fisiologia all'istologia e allo studio delle sindromi cliniche dipendenti da disturbi funzionali delle ghiandole. Inoltre, la conoscenza di un primo ormone puro, l'adrenalina, dava il suo giusto valore a una nozione prima accettata implicitamente in mancanza di prove, quella di ormoni ‛messaggeri chimici' attivi a distanza e trasportati per via sanguigna in concentrazione molto bassa. Oggigiorno si tende ad applicare il termine di ormone essenzialmente ai prodotti di secrezione di ghiandole endocrine, sebbene si designino sotto il nome di ormoni tessutali sostanze che, come la serotonina o l'istamina, sono riversate nel sangue da tessuti e non da una ghiandola chiusa.
b) Fisiologia endocrina e endocrinologia medica
L'ablazione di una ghiandola, e la compensazione dei disturbi che da tale ablazione derivano per mezzo della somministrazione delle sostanze che essa produce, rappresenta uno dei metodi diretti per mettere in evidenza una secrezione endocrina. Una sperimentazione basata su questo principio fu realizzata molto tempo prima che fossero preparati ormoni puri o purificati. Durante i primi venticinque anni di questo secolo la maggior parte dei prodotti endocrini utilizzati nella sperimentazione e nella pratica fu ottenuta soltanto con mezzi di fortuna: ingestione di organi appena prelevati, iniezione di loro estratti o trasfusione del sangue efferente dalle ghiandole; in tal modo i risultati ottenuti non furono significativi se non in condizioni particolari per i singoli organi. Così, l'ipoglicemia conseguente all'ablazione totale del pancreas ovvero alla legatura del peduncolo vascolare del solo nodulo pancreatico rimasto dopo l'estirpazione quasi totale dell'organo è stata tra le poche prove indiscutibili della secrezione endocrina di insulina che si sono potute stabilire. Al tempo stesso, però, non ci si era resi conto che le proteinasi pancreatiche inattivano rapidamente questo ormone negli estratti dell'organo.
Diversi fattori hanno complicato l'interpretazione dei fatti osservati. Uno di essi è l'eterogeneità di organi come il pancreas o il testicolo che elaborano una secrezione esterna - succo pancreatico o sperma - e una secrezione interna, da cellule istologicamente differenziate. L'atrofia o la distruzione selettiva della parte ghiandolare esocrina e la conservazione del tessuto endocrino, come la realizzarono Ancel e Bouin nel testicolo - in cui poterono ledere la parte acinosa che produce gli spermatozoi conservando intatto il tessuto interstiziale che secerne gli ormoni androgeni - ha portato una dimostrazione diretta dell'attività endocrina di questo organo. Nel caso dell'ovaio, la presenza in uno stesso organo di due funzioni endocrine, localizzate l'una nei follicoli di Graaf l'altra nel corpo luteo, ha reso particolarmente incerta la prima fase della sperimentazione fisiologica, prima che E. W. Allen ed E. A. Doisy, e R. Courrier dimostrassero, nel 1923, la specializzazione delle due parti attive della ghiandola. L'istologia è stata e rimane di grande ausilio, in questo campo, alla fisiologia, non soltanto mettendo in evidenza l'eterogeneità di certe ghiandole, ma fornendo utili informazioni sull'attività delle cellule secernenti, in particolare nel caso dei due lobi dell'ipofisi, dell'ovaio, della tiroide. Un'altra difficoltà nell'interpretazione dei primi risultati scaturiva dal fatto che le azioni ormonali presentano un'estrema diversificazione nelle loro modalità e nella loro natura. Esse sono a volte assai rapide e poco persistenti, come quando partecipano a una regolazione funzionale quale quella della glicemia a opera dell'insulina e dell'adrenalina; altre volte sono lente e progressive, per esempio quando agiscono sullo sviluppo corporeo o sui caratteri morfologici di un organismo, come nel caso della somatotropina ipofisaria o talvolta degli ormoni sessuali. Infatti la diversità di azione dei prodotti endocrini dipende sia dalla loro natura, sia dalle cellule che ricevono il messaggio. Dobbiamo anche dire che l'intervento di più ormoni in un medesimo processo metabolico e l'antagonismo tra le azioni di alcuni di essi, metaboliche o morfogenetiche, hanno complicato in egual misura l'interpretazione dei dati, in particolar modo nel caso degli ormoni sessuali.
Nel primo terzo di questo secolo destarono grande interesse ricerche cliniche grazie alle quali si stabilì una relazione tra disturbi endocrini e alcune sindromi, di cui si poté chiarire il meccanismo patogenetico. Malattie da tempo note, come il gozzo semplice, l'iperfunzionalità tiroidea della malattia di Graves-Basedow furono così spiegate e al tempo stesso ne vennero identificate delle nuove. A questo proposito si deve far rilevare che i risultati della sperimentazione clinica frequentemente precedettero e completarono quelli delle ricerche fisiologiche e talvolta le orientarono. Ne sono esempi significativi l'opera di Kocher, di Parhon, Marañon, Marine, Pende, le ricerche cliniche su l'acromegalia, il diabete pancreatico, il morbo di Addison, quelle sulla terapia ormonale di sindromi multiple. Una parte delle ricerche svolte in tema di malattie endocrine ha avuto per primo scopo la terapia di queste in un'epoca in cui non si disponeva ancora di prodotti ormonali puri o purificati; si giunse così all'opoterapia. Questo tipo di terapia consiste nella somministrazione di ghiandole o di loro estratti; essa però non sopravvisse all'impiego di ormoni puri o purificati. Il principio a cui si ispirava l'opoterapia ha in ogni modo contribuito allo sviluppo dell'endocrinologia, sebbene per la maggior parte le preparazioni somministrate fossero poco attive, poiché per lo più le ghiandole endocrine, esclusa la tiroide, non hanno riserve ormonali. La nozione di assenza di specificità zoologica degli ormoni che l'opoterapia presupponeva ha permesso di trattare le malattie endocrine umane per mezzo della somministrazione di ghiandole di origine animale e ha portato all'estrazione di ormoni a partire da materiale disponibile su scala industriale.
c) Prime tappe dell'endocrinologia moderna
L'aver stabilito l'esistenza di una secrezione ormonale caratterizzò l'orientamento della prima fase della ricerca, che poté essere poi superata soltanto per mezzo della sperimentazione di tipo fisiologico. Per questa via si giunse all'isolamento degli ormoni e in seguito anche a stabilirne la struttura molecolare. Quest'ultimo obiettivo non rimase fine a se stesso nell'ambito dell'endocrinologia, ma le fornì anzi notevoli possibilità di sviluppo. L'endocrinologia si è inizialmente sviluppata sia secondo grandi correnti di ricerca, sia secondo un ritmo proprio a ciascun organo; per cui le tappe successive si sovrappongono allorché si cerca di definirne l'evoluzione generale. Alla fine del primo quarto di questo secolo (v. Cameron, 1933), si era certi dell'attività endocrina soltanto di un piccolo numero di tessuti differenziati nell'uomo e nei Vertebrati superiori: le zone corticali e midollari delle capsule surrenali, i lobi anteriore e posteriore dell'ipofisi, i follicoli e il corpo luteo dell'ovaio, il tessuto interstiziale del testicolo, gli isolotti di Langerhans del pancreas, le ghiandole paratiroidi, l'epitelio delle vescicole tiroidee. Un ormone puro, l'adrenalina, fu isolato dalla zona midollare delle ghiandole surrenali nel 1901 da Aldrich e Takamine. Un altro ormone ricco di iodio, la tiroxina, fu isolato, allo stato puro, nel 1915 da E. C. Kendall dalla tiroide, ma la sua struttura di 3,5,3′,5′-tetraiodotironina sarebbe poi stata stabilita soltanto nel 1926 da C. R. Harington. Un terzo ormone, l'insulina, fu estratto dal pancreas da F. G. Banting, C. H. Best e J. J. MacLeod nel 1921 e preparato allo stato cristallino da J. J. Abei nel 1926. All'inizio il suo isolamento fu reso possibile dalla realizzazione di condizioni operative tali per cui esso non veniva distrutto dagli enzimi proteolitici del pancreas esocrino; il suo isolamento ha anche fornito la prima prova dell'esistenza di un ormone proteico e ha contribuito con ciò ad ampliare i confini non ben definiti della biochimica degli ormoni. Questa non era però ancora pronta a uno sviluppo rapido, sia per la mancanza di mezzi tecnici appropriati all'isolamento e allo studio degli ormoni puri, sia per lo stato rudimentale delle conoscenze sulla struttura delle proteine. L'esistenza di vari steroidi ad azione ormonale negli estratti della parte corticale delle capsule surrenali non poteva essere sospettata nel 1925 e neppure dopo la preparazione, nel 1929, della ‛cortina', estratto surrenale depurato dall'adrenalina. J. B. Collip aveva già preparato estratti paratiroidei ipocalcemizzanti, ma il loro frazionamento non aveva ancora orientato chiaramente le ricerche verso la natura proteica delle loro parti attive.
L'endocrinologia moderna si è sviluppata a partire da questa prima serie di risultati, prima dei quali essa si era limitata a ricerche non coordinabili in un unico quadro. I suoi maggiori obiettivi, definiti verso la fine degli anni trenta, consistevano nell'identificazione di nuove funzioni endocrine, nello studio delle coordinazioni endocrine e in quello della biochimica degli ormoni. Questi tre temi sono complementari ed è difficile trattarli del tutto separatamente. I primi due, già differenziati nel primo quarto del secolo, conobbero poi un brillante sviluppo. Quanto alla biochimica, essa ha aperto all'endocrinologia una nuova era, sia stabilendo la struttura degli ormoni puri sia permettendo di correlarne l'attività alla struttura.
2. Scoperta di nuove secrezioni ormonali e di correlazioni endocrine di diverso tipo negli ultimi cinquant'anni
I progressi dell'endocrinologia dopo il 1925 sono stati realizzati grazie alla fisiologia endocrina; essi hanno rappresentato la condizione prima che ha determinato la nascita della biochimica degli ormoni. La tappa iniziale è stata la scoperta di nuove attività ormonali, di cui alcune si esercitano a carico di cellule bersaglio differenziate, recettrici dei messaggeri chimici. Lo studio di correlazioni endocrine tra organi, di cui l'uno è il recettore di un prodotto elaborato dall'altro, e quello di sistemi di coordinazioni di secrezioni ormonali complesse tra più organi endocrini hanno aperto successivamente alla fisiologia e alla patologia nuovi campi (v. Pincus, 1944-1972; v. Harris e Thimann, 1942- 1972; v. Pincus e altri, 1948-1964; v. Gabe e altri, 1964; v. Wolff, 1972; v. Karlson, 19644; v. Hermann e Cier, 1970; v. Régulations hormonales..., 1937; v. Hall e altri, 1968) tra i quali merita una particolare menzione la neuro-endocrinologia.
a) Secrezione di ormoni diversi da parte di uno stesso organo e differenziamento cellulare
Tutte le ghiandole endocrine si sono rivelate poco a poco secretrici di un numero più o meno elevato di ormoni dall'attività qualitativamente o quantitativamente diversa. Ci si può quindi chiedere se esistano ghiandole che elaborano un solo ormone. Le azioni di ormoni elaborati da una stessa ghiandola sono a volte qualitativamente identiche tra loro, come nel caso delle iodotironine, ma possono essere molto diverse, come nel caso degli ormoni ipofisari (v. Pincus e altri, 1948-1964; v. Régulations hormonales..., 1937).
La midollare surrenale secerne due catecolammine: l'adrenalina e la noradrenalina, isolate rispettivamente nel 1901 e nel 1949. Entrambe sono dei potenti vasocostrittori e attivano la glicogenolisi nei tessuti. L'azione dell'adrenalina, che sotto questo aspetto è più energica, dà luogo a un'iperglicemia. La tiroide secerne due iodotironine, la tiroxina (tetraiodata) e la sua omologa triiodata, la 3, 5, 3′-triiodotironina, rispettivamente isolate nel 1915 e nel 1952. Le cellule interstiziali del testicolo elaborano due prodotti, l'androsterone, isolato nel 1931, e il testosterone, isolato nel 1935. Questi due ormoni regolano nel maschio lo sviluppo del tratto genitale e i caratteri sessuali secondari; essi sono antagonisti di ormoni sessuali femminili. Si sono potute identificare istologicamente, nel contesto di queste tre ghiandole, le cellule che sono responsabili dell'elaborazione degli ormoni: le cellule cromaffini della midollare surrenale, l'epitelio che riveste le vescicole tiroidee, le cellule di Leydig del testicolo. La differenza di affinità di ciascuna delle due catecolammine della midollare surrenale rispetto ai loro recettori periferici α e β spiega come la loro azione su questi ultimi sia quantitativamente diversa. Lo stesso dicasi delle prostaglandine, ultime nate fra gli ormoni, e di cui quattordici sono state isolate da ghiandole seminali di diversi Mammiferi, da vescicole seminali e da sperma umano, a partire dal 1957. Queste prostaglandine hanno strutture molto simili ed esercitano la loro azione sul tono della muscolatura liscia e sulla sua regolazione in diversi organi; esse sono sintetizzate da cellule istologicamente identiche. Lo stesso vale per gli ormoni paratiroidei che regolano il metabolismo del calcio e la calcemia.
Il pancreas endocrino elabora negli isolotti di Langerhans, distinti dagli acini ghiandolari che sintetizzano gli enzimi digestivi del succo pancreatico, due ormoni antagonisti: l'insulina e il glucagone. Gli estratti pancreatici impuri contengono entrambe queste due sostanze, ma in misura assai maggiore insulina; per cui dopo l'iniezione di questi estratti si manifesta soltanto l'azione ipoglicemizzante di quest'ultima. Ovvero, l'ipoglicemia insulinica è seguita da una debole iperglicemia di breve durata; era quindi legittimo pensare che l'iniezione aveva messo in circolo una sostanza antagonista dell'insulina, che si è poi rivelata essere un nuovo ormone, il glucagone. Estratto dal pancreas insieme all'insulina, esso è stato separato da questa durante la sua purificazione. Entrambi sono elaborati nelle cellule specializzate degli isolotti di Langerhans, che prendono il nome di cellule α e β. Le cellule β, produttrici di insulina, sono selettivamente distrutte dalla somministrazione di allossana. In queste condizioni, l'insulina è assente dal pancreas degli animali trattati, che presentano quindi un diabete sperimentale detto ‛da allossana'; gli estratti della ghiandola in questo caso sono iperglicemizzanti, poiché non contengono altro che il glucagone.
Particolari problemi fisiologici sono scaturiti dallo studio di organi che, come la corteccia surrenale e l'ovaio, secernono svariati ormoni che svolgono la loro azione a livello di diversi recettori. Nel caso della corteccia surrenale, uno stesso tessuto produce un gran numero di molecole attive che hanno funzioni biologiche diverse; nel caso dell'ovaio, regioni specializzate di uno stesso organo esplicano una funzione endocrina loro caratteristica.
Le capsule surrenali, la cui esistenza fu scoperta da Leonardo da Vinci, contengono due ghiandole distinte, rispettivamente nelle loro parti midollare e corticale. La corteccia surrenale, il cui volume corrisponde ai 4/5 dell'organo, comprende tre zone istologicamente non omogenee, ma alle quali è stato possibile attribuire una specializzazione funzionale ben determinata. Tralasciando la funzione della midollare surrenale, di cui abbiamo già parlato, quella della corteccia surrenale è rimasta per molto tempo oscura. La surrenectomia bilaterale, sempre mortale, provoca una sindrome di squilibrio idro-elettrolitico caratterizzata da un aumento di escrezione urinaria di sodio e una diminuzione di quella di potassio; contemporaneamente si verificano disturbi del metabolismo glucidico cellulare e un'ipoglicemia. Questi disturbi si potevano parzialmente compensare con la somministrazione di un estratto della corteccia surrenale detto cortina. In seguito sono state messe in evidenza nell'uomo insufficienze cortico-surrenali ed è stata stabilita la loro indipendenza dal morbo di Addison, che colpisce selettivamente la midollare surrenale. D'altra parte non si poteva nemmeno prevedere l'esistenza del considerevole numero di ormoni corticali; oggi ne conosciamo più di cinquanta specie, a partire dalla scoperta del corticosterone nel 1934, e quindi del cortisone e del cortisolo a opera di una notevole serie di lavori inaugurata da E. C. Kendall e T. Reichstein.
Gli ormoni corticali, i corticosteroidi, sono stati suddivisi in ‛mineralcorticoidi' e ‛glucocorticoidi' il più importante dei primi è l'aldosterone, attivo nell'uomo alla concentrazione di 0,1-0,2 μ/kg (v. corteccia surrenale). Questo ormone, secreto nella zona glomerulare periferica della corteccia surrenale, esercita la sua azione sul movimento degli elettroliti a livello renale producendo una diminuzione della escrezione del Na+ e un aumento di quella del K+. Anche la secrezione dell'aldosterone è regolata da un meccanismo ormonale, poiché dipende da un sistema endocrino renale a due costituenti, il sistema renina-angiotensina. La conoscenza di questo sistema ha introdotto negli studi sull'aldosterone una nuova concezione, quella della regolazione ormonale della secrezione di questo mineralcorticoide. In effetti, una α-globulina del siero, l'angiotensinogeno, dà origine sotto l'azione della renina, enzima proteolitico secreto dalle cellule iuxtaglomerulari della parete delle arteriole dei glomeruli renali, a un decapeptide, l'angiotensina I, che si trasforma a sua volta per idrolisi in un ottopeptide, l'angiotensina II, che stimola la secrezione cortico-surrenale di aldosterone. I glucocorticoidi, fra cui il cortisone fu scoperto prima del più importante di essi, il cortisolo, sono secreti dalla zona fascicolata della ghiandola. La loro azione si esercita sul metabolismo dei glucidi, favorendo la sintesi epatica e muscolare del glicogeno a partire dal glucosio del sangue. Nell'uomo, alcune sindromi sono state associate a disturbi della secrezione corticosurrenale specificamente dell'uno o dell'altro gruppo di ormoni. Questo è il caso degli iperaldosteronismi, in particolare della sindrone ipertensiva di Conn, o della sindrome di Cushing che è legata a un'ipersecrezione di glucocorticoidi. Inoltre, poiché la stessa ghiandola secerne una serie di ormoni identici o simili agli androgeni, si è anche identificata una sindrome adreno-genitale da iperandrogenia.
L'endocrinologia dell'ovaio si è imbattuta in un altro tipo di difficoltà. In effetti, da una parte gli ormoni della gonade femminile sono più di uno e la loro secrezione è localizzata in diverse regioni morfologiche molto particolari dell'organo, dall'altra essi regolano l'ovulazione e i caratteri sessuali secondari e partecipano all'evoluzione della gestazione. L'evidenziazione delle rispettive funzioni dei follicoli secernenti gli estrogeni e del corpo luteo, che elabora il progesterone, è stata la tappa che ha preparato lo sviluppo della biochimica degli steroidi ovarici; essa è stata superata in gran parte grazie ai lavori di E. W. Allen, E. A. Doisy e R. Courrier poco prima del 1925. L'estrone, o follicolina, isolato nel 1929 da A. F. J. Butenandt, è, insieme al 17 β-estradiolo, il prodotto delle secrezioni follicolari; il 20 α- e il β-idrossiprogesterone e il progesterone sono secreti dal corpo luteo. Il ritmo della liberazione di questi ormoni durante il ciclo estrale è stato oggetto di numerosi lavori; così pure quello della secrezione degli ormoni del corpo luteo nel corso del suo sviluppo durante la gravidanza e della sua evoluzione durante il ciclo estrale; lo stesso dicasi del tasso plasmatico e urinario di questi ormoni durante la gravidanza. Tali ormoni regolano non soltanto l'ovulazione e la gestazione, ma anche i caratteri sessuali secondari femminili e si è scritto che ‟gli ormoni follicolari erano quelli delle innamorate e il progesterone l'ormone delle madri". Questa espressione evoca, inoltre, l'antagonismo degli estrogeni e degli ormoni luteinici nei confronti dell'annidamento dell'uovo. Durante la gestazione, il corpo luteo, che presenta in questo periodo una notevole ipertrofia, e la placenta sono la principale sorgente di tutti questi ormoni, che controllano allora lo sviluppo del feto, particolarmente del suo tratto genitale. La preparazione al parto e la sua realizzazione sono tributari di altri messaggi endocrini, di cui uno proviene dall'ovaio. Questo elabora alla fine della gravidanza un ormone che provoca un rilasciamento della sinfisi pubica e permette la sua apertura, la relaxina. Sebbene messo in evidenza nel 1934 da Hissaw, questo ormone, probabilmente proteico, non è stato ancora ottenuto altamente purificato.
L'insieme dei progressi realizzati durante il XX secolo dall'endocrinologia ovarica mostra la fecondità della collaborazione in questo campo di medici, fisiologi e biochimici.
b) Regolazioni ormonali complesse e funzione endocrina del sistema ipotalamo-ipofisario
Un tipo più complesso di regolazione endocrina è stato messo in evidenza nella secrezione di ormoni da parte dell'ipofisi, appendice ghiandolare del cervello, e dell'ipotalamo cerebrale, collegati tra loro dal peduncolo infundibulare e per mezzo di un particolare sistema vascolare (v. Pincus e altri, 1948-1964; v. Karlson 19644). La scoperta dell'esistenza di una regolazione ipotalamo-ipofisaria ha segnato una tappa fondamentale dell'endocrinologia. Le stesse strutture ipotalamiche possono, in effetti, influire su tre tipi di cellule: i motoneuroni, i neuroni pregangliari del sistema nervoso autonomo e le cellule di ghiandole endocrine, tramite l'ipofisi.
L'ipofisi o ghiandola pituitaria è costituita dall'associazione di due lobi, di cui ciascuno è una ghiandola endocrina che secerne suoi specifici ormoni. Esiste anche un lobo intermedio, sviluppato pochissimo nei Mammiferi, maggiormente negli Anfibi e nei Pesci. Nei capitoli successivi sarà illustrata l'endocrinologia dell'ipofisi, mentre in questo ci limiteremo a mettere in rilievo il particolare interesse che ha rappresentato lo studio della formazione e della secrezione degli ormoni pituitari.
L'ipofisectomia non è letale ma provoca una serie di turbe delle quali le più significative sono il nanismo, l'atrofia delle capsule surrenali e della tiroide e l'assenza di maturazione sessuale. Fin dal 1886, P. Marie aveva ricollegato certe sindromi umane a un'alterazione ipofisaria, associando alcuni nanismi all'ipoattività della ghiandola e l'acromegalia e il gigantismo alla sua iperattività. La via così aperta alle ricerche fisiologiche venne confermata dallo studio di queste malattie e successivamente della sindrome di Cushing, dovuta all'iperincrezione di glucocorticoidi surrenali provocata da uno squilibrio nella secrezione di una stimolina ipofisaria spesso conseguente a un tumore.
L'ipofisi, organo endocrino trofico, controlla il funzionamento di ghiandole endocrine bersaglio per mezzo delle stimoline, secrete dal suo lobo anteriore. Tali stimoline attivano la secrezione dalle ghiandole bersaglio, e l'aumento del tasso sanguigno di uno degli ormoni di ciascuna di queste ultime inibisce a sua volta la secrezione della stimolina ipofisaria corrispondente; è questo un meccanismo elementare di retroazione negativa, spesso chiamato feedback. La sostanza secreta dalla ghiandola bersaglio agisce, quindi, da un lato sulle cellule periferiche effettrici, dall'altro, per feedback, sulla secrezione della stimolina ipofisaria che specificamente le corrisponde.
L'ipofisi risponde quindi solo a variazioni del tasso ematico dell'ormone secreto dalla ghiandola bersaglio, mentre il suo funzionamento esige un sistema di regolazione più complesso, più preciso, al quale partecipa il sistema nervoso. Una serie di lavori biologici e clinici ha consentito di stabilire che diverse stimolazioni sensoriali, come quelle tattili, uditive, olfattive e termiche, influenzano l'equilibrio endocrino; lo stesso dicasi per modificazioni dell'ambiente cellulare, come la disidratazione o gli edemi. La regolazione ormonale da parte del sistema nervoso centrale si attua in due modi: direttamente sull'organo bersaglio o sul tessuto effettore, come nel caso degli ormoni postipofisari; indirettamente, come hanno mostrato nel 1953 J. Benoit e Assenmacher, tramite le stimoline del lobo anteriore. In questa condizione entra in gioco una correlazione funzionale ipotalamoipofisaria che si basa sul trasporto di polipeptidi designati con il nome di releasing factors o RF: questi sono ormoni di comando della secrezione di stimoline da parte dell'ipofisi anteriore, individuati inizialmente da Schally nel 1957. Gli RF, che sono specifici per ogni stimolina, sono secreti da una particolare zona dei nuclei basali del tuber cinereum ipotalamico propria a ciascuno di essi. Vengono trasportati per via sanguigna, nel sistema portale ipotalamo-ipofisario di Popa, all'ipofisi anteriore ove provocano la secrezione della stimolina corrispondente, che agisce a sua volta sul suo organo bersaglio periferico. Un aumento della concentrazione nel sangue dell'ormone prodotto dall'organo bersaglio interviene a ostacolare, per feedback, la secrezione di questa ghiandola, sia direttamente inibendo la liberazione della stimolina ipofisaria, sia indirettamente tramite l'ipotalamo. È anche possibile che la secrezione degli RF ipotalamici sia regolata da quella delle stimoline stesse, in un processo di feedback ipotalamo-ipofisario.
La neuroipofisi o postipofisi secerne ormoni prodotti dalle cellule neurosecretrici dell'ipotalamo che sono trasportati al lobo posteriore pituitario da fibre nervose. La neurosecrezione ha potuto essere messa in evidenza con le tecniche istologiche e con l'esame ultramicroscopico del movimento, lungo gli assoni, di goccioline di secrezione nelle quali gli ormoni sono probabilmente fissati a lipoproteine. Due polipeptidi endocrini di origine ipotalamica sono secreti dalla postipofisi: la vasopressina o adiuretina, o ADH, e l'ossitocina, i cui organi bersaglio sono rispettivamente il rene e l'utero. L'ADH regola l'escrezione renale dell'acqua, e una diminuzione della sua secrezione provoca un aumento della diuresi: questa si manifesta, in patologia umana, con una forma di diabete, detto insipido, nei cui casi più gravi si è osservata l'emissione di urina fino a cinquanta litri al giorno. L'ossitocina stimola la contrazione uterina durante il parto; essa provoca inoltre l'eiezione del latte per contrazione delle fibre lisce della mammella durante l'allattamento. Questo fenomeno di neurosecrezione ormonale, la cui teoria è stata elaborata nel 1904 da T. R. Elliott, è stato dimostrato e studiato da Scharrer a partire dal 1928.
L'adenoipofisi, o lobo anteriore della ghiandola, secerne stimoline in cellule la cui specializzazione è stata dimostrata con metodi istologici e citologici. Si distinguono infatti, per mezzo di reazioni appropriate, cellule somatotrope, corticotrope, tireotrope e cellule gonadotrope che contengono o gonadostimoline o la prolattina. Si sono isolate e caratterizzate, in base alla specificità d'azione, 6 stimoline, per ognuna delle quali esiste un fattore di liberazione di origine ipotalamica (RF). La somatotropina o ormone della crescita (SH) esercita un'azione generale sull'accrescimento e si è già detto che la sua assenza provoca il nanismo. La tireostimolina (TSH) controlla la secrezione ormonale della tiroide, la corticostimolina (ACTH) quella della corteccia surrenale. Sono anche secreti tre ormoni gonadotropi: la gonadotropina A, che agisce sullo sviluppo dei follicoli ovarici e delle cellule germinali testicolari spermatogene, alla quale, per la sua azione follicolare, si dà anche il nome di follicolostimolina (FSH); la gonadotropina B o luteotropina (LH), che stimola la trasformazione del follicolo di Graaf in corpo luteo e la produzione a livello dei testicoli di ormoni maschili; infine, la prolattina o ormone lattotropo (LTH), che stimola l'attività del corpo luteo e provoca lo sviluppo della ghiandola mammaria e la sua secrezione lattea. Durante la gravidanza la placenta elabora e secerne in abbondanza una sostanza molto vicina alla gonadotropina B (HCG), la cui presenza nell'urina delle donne gravide permette una diagnosi precoce di gravidanza, secondo un principio stabilito da S. Aschheim e B. Zondek da oltre quaranta anni.
Il lobo intermedio dell'ipofisi secerne la melanotropina (MSH), analoga per la sua natura alla corticotropina e attiva sulla contrazione dei melanofori cutanei negli Anfibi e nei Pesci. Nell'uomo, l'MSH sembra che intervenga solamente sulla melanogenesi cutanea e sulla steroidogenesi corticosurrenale.
Pochi campi dell'endocrinologia hanno conosciuto un così ampio e rapido sviluppo come lo studio dell'ipofisi, ma non si potevano ricordare qui altro che le linee generali della sua evoluzione. Basterà rammentare che concetti così generali come quelli di ghiandole-bersaglio, di neurosecrezione, di ormoni di comando della secrezione di stimoline e l'evidenziazione della coordinazione ipotalamo-ipofisaria costituiscono scoperte fondamentali, che dimostrano l'eccezionale importanza che ha rivestito lo studio di questa ghiandola nel corso del secolo (v. ormoni nei vertebrati).
c) Altri tipi di attività ormonali: ormoni tessutali, ormoni di Invertebrati, feromoni
Si designa sotto il nome di ormoni tessutali un gruppo di sostanze trasportate, per mezzo del sangue, a cellule diverse da quelle che le elaborano (v. Gabe e altri, 1964; v. Wolff, 1972). Questa definizione, molto generale, ha condotto a una certa confusione, perché talvolta è stata estesa a ogni sostanza endogena biologicamente attiva che subisce un trasporto umorale, senza prendere in considerazione la specificità dei suoi effetti e dei suoi recettori. Infatti, una sostanza come l'angiotensina, la cui formazione si verifica sotto l'azione della renina nel rene e la cui azione si esercita sulla liberazione di aldosterone nella corteccia surrenale (v. corteccia surrenale), è incontestabilmente un ormone. È meno evidente che un carattere endocrino debba essere attribuito a composti presenti in tessuti diversi, come la serotonina e l'istamina; o a sostanze liberate a livello delle terminazioni nervose, come l'acetilcolina, mediatore chimico della maggior parte dei nervi parasimpatici o colinergici, e la tirammina, precursore delle catecolammine liberate dalle terminazioni dei nervi simpatici o adrenergici. Si è anche ipotizzata l'esistenza di una serie di ormoni del tratto digestivo analoghi alla secretina, la sola tra essi attualmente ben definita.
Un campo nuovo e vasto è stato aperto dallo studio degli ormoni degli Invertebrati, particolarmente degli Insetti e dei Crostacei (v. Gabe e altri, 1964; v. Wolff, 1972; v. ormoni negli invertebrati). La regolazione della metamorfosi degli Insetti, studiata soprattutto da Wiggleworth e da P. Karlson, ne ha fornito esempi significativi nei Lepidotteri (Bombyx) e nei Ditteri (Calliphora). Il susseguirsi di tre mute che trasforma la larva, il bruco, prima in crisalide, quindi in immagine e in insetto completo, farfalla o mosca, è regolato da tre ormoni. Le cellule neurosecretrici del cervello controllano tutte le mute producendo un primo ormone detto adrenotropo, che stimola la secrezione, dalle ghiandole protoraciche, di un ormone detto ecdisone e dai corpora allata di un altro ormone, detto ormone giovanile o neotenina. La prima muta, nel corso della quale la larva si trasforma in crisalide, richiede la presenza della neotenina e dell'ecdisone, mentre le due seguenti, la muta da crisalide in immagine e quindi in insetto completo, avvengono sotto l'azione del solo ecdisone, ormone steroide isolato allo stato puro nel 1954 da Butenandt e Karlson. Si può inoltre menzionare, nel quadro generale dell'endocrinologia degli Invertebrati, che certi ormoni di Vertebrati, come le iodotironine, esistono anche nei Protocordati, che tuttavia non possiedono neppure un abbozzo della ghiandola tiroide presente nei Vertebrati superiori dei quali sono i precursori evolutivi, mentre non si trovano nei gruppi zoologicamente meno evoluti.
L'esistenza di correlazioni umorali tra individui della stessa specie è stata posta in evidenza in particolare ne- gli Insetti; esse si esplicano tramite sostanze che Karlson e Lüscher hanno chiamato nel 1954 ‛feromoni' o ‛ormoni di gruppo'. La funzione di comunicazione a distanza di questi agenti chimici era stata sospettata fin dall'Ottocento dall'entomologo J. H. Fabre, in base all'osservazione che femmine di farfalle attirano i maschi quando sono prigioniere in sacchetti di garza, ma non quando sono chiuse in vasi di vetro.
Responsabile dell'attrazione dei maschi verso le femmine è una sostanza volatile secreta da ghiandole endocrine delle femmine vergini e captata elettivamente dalle antenne dei maschi, che, isolata e sintetizzata, fu chiamata bombicolo da E. Beckmann e J. F. K. Hecker nel 1967. Chemorecettori presenti nelle antenne dei maschi captano lo stimolo prodotto dal bombicolo e trasmettono per via nervosa un eccitamento ai centri che presiedono al comportamento sessuale del maschio, già a una concentrazione bassissima, che, espressa in unità fisiologiche, è di 10-10 μg per ml di soluzione.
Anche altri insetti sociali, le api e le termiti, elaborano feromoni. Nelle api un alveare ha una sola ape regina, qualche centinaio di maschi e decine di migliaia di operaie, le cui ovaie sono atrofizzate. Un feromone secreto dalla regina provoca l'involuzione ovarica delle operaie. Questa sostanza, isolata da Butler nel 1959, possiede tre notevoli proprietà biologiche: attira i maschi per via olfattiva durante il volo nuziale; agisce per via gustativa sulle operaie che l'assorbono preparando la corte alla loro regina; esercita quindi una doppia inibizione ostacolando lo sviluppo delle ovaie e neutralizzando l'istinto che spinge le operaie a costruire alveoli regali. In definitiva, la regina è in grado, grazie al feromone che elabora, di attirare i maschi durante il volo nuziale e, nello stesso tempo, di assicurarsi la supremazia sulle operaie che essa stessa sterilizza, evitando così di avere tra loro delle rivali che si costruiscano un proprio habitat regale.
L'esistenza di azioni di tipo ormonale è stata messa in evidenza in vegetali, da cui si sono isolati dei fitormoni. Tra questi, le auxine sono responsabili dell'allungamento di cellule e di diversi tropismi di piante inferiori; altri, come le gibberelline e la chinetina, stimolano la divisione e la moltiplicazione delle cellule (v. botanica).
3. Biochimica endocrina
La biochimica endocrina comprende lo studio della chimica degli ormoni puri e del loro metabolismo normale e patologico.
Il suo sviluppo, relativamente recente, è oggi considerevole e le ‛applicazioni pratiche che ne derivano in diversi campi sono molto importanti (v. Pincus e altri, 1948-1964; v. Karlson, 19644; v. Herman e Cier, 1970).
a) Costituzione chimica degli ormoni
La diversità delle funzioni caratteristiche delle sostanze endocrine è necessariamente collegata a quella della loro struttura; la tab. I fornisce, a questo riguardo, qualche indicazione generale.

È stato necessario isolare gli ormoni allo stato puro per studiarne la struttura, in quanto la loro azione si manifesta specificamente quando sono presenti nei mezzi naturali complessi da cui si estraggono. L'alto tenore di estrogeni dell'urina delle femmine gestanti ha permesso a Butenandt di isolare la prima di queste sostanze nel 1929 e di aprire così la via alla chimica degli ormoni steroidei. Molti altri prodotti endocrini hanno potuto essere isolati soltanto dalle ghiandole stesse, dove non si trovano che in quantità minime; l'accumulo di ormoni tiroidei, grazie al loro legame con la tireoglobulina, proteina depositata nelle vescicole ricolme di colloide, è, infatti, un'eccezione a questo riguardo. Così la biochimica degli ormoni non ha potuto svilupparsi che grazie ai recenti progressi della chimica analitica e organica. Alcuni ormoni sono degli steroidi (v. tab. I), altri derivano da acidi grassi o da amminoacidi e un gran numero di essi sono peptidi o proteine. Solo alcuni dell'ultimo gruppo presentano una specificità di origine, a seconda dell'organismo animale da cui derivano.
Gli ormoni steroidei sono derivati del nucleo policiclico ciclopentanfenantrenico, come il colesterolo e gli acidi biliari. La differenza della loro struttura si manifesta a livello delle catene laterali dell'idrocarburo ciclico fondamentale del colesterolo, il colestano, nel grado d'insaturazione dei suoi cicli e nella presenza o assenza di funzioni ossigenate - alcooliche, aldeidiche, chetoniche - in diverse posizioni. La tab. I illustra le relazioni strutturali tra il Colesterolo e diversi steroidi caratteristici di varie secrezioni ormonali menzionate nella tab. II: il corticosterone e l'aldosterone della corteccia surrenale, il progesterone e l'estrone del corpo luteo e dei follicoli ovarici, l'androsterone e il testosterone testicolari. L'ecdisone degli Insetti appartiene alla stessa serie.
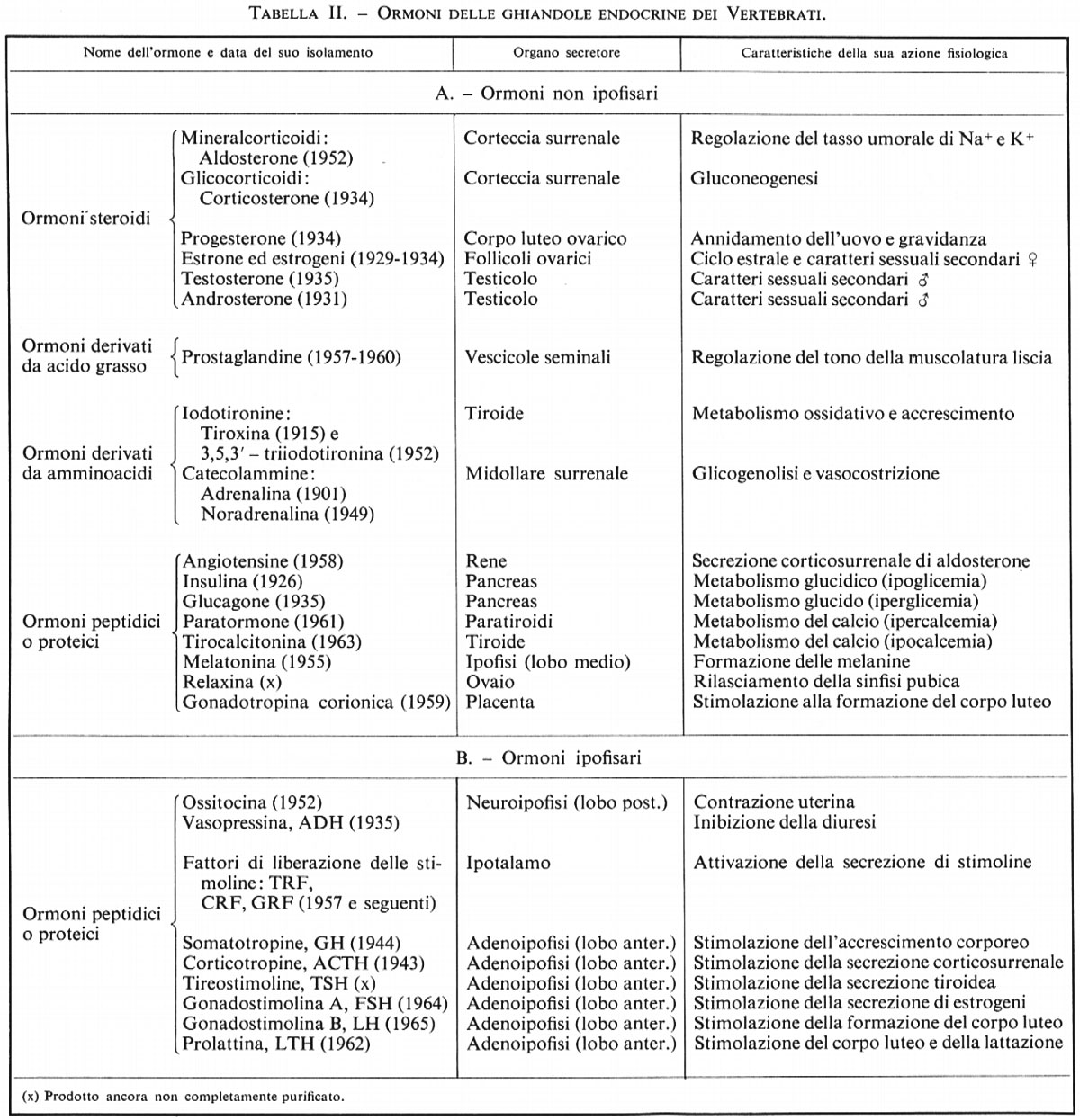
Le prostaglandine derivano tutte da uno stesso acido grasso, l'acido prostanoico, dalla formula schematica:

Esse differiscono per l'esistenza di uno o più legami etilenici o per quella di funzioni ossigenate, alcooliche o chetoniche, in diverse posizioni.
I feromoni degli Insetti fin qui identificati hanno strutture vicine a quelle di acidi grassi. In effetti il bombicolo delle farfalle è il ∆11-12- esadecadienolo e l'ormone regale delle api è un acido 9-chetodecanoico insaturo.
Gli ormoni che derivano dalla tirosina o p-idrossifenilalanina sono le catecolammine della midollare surrenale e le iodotironine della tiroide. La loro parentela di struttura si vede dalle formule dei più importanti tra essi e dell'amminoacido.
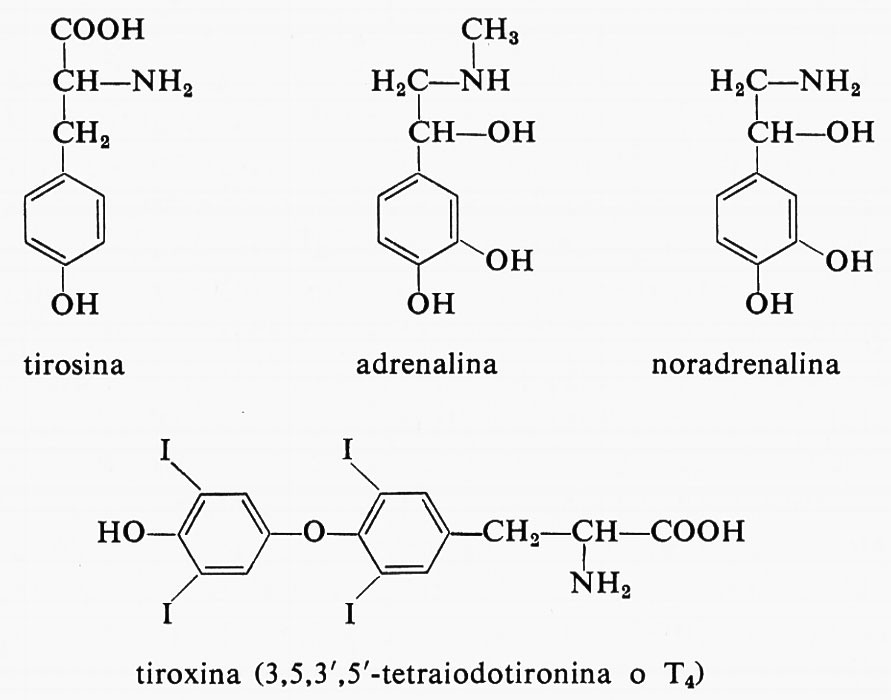
La presenza di iodio negli ormoni tiroidei T3 e T4, come nei loro precursori biologici, le iodotirosine, determina l'attività dei primi, poiché la loro struttura non iodata, la tironina, è inattiva. Un amminoacido indolico, il triptofano, è il precursore di ormoni vegetali, le auxine.
Polipeptidi e proteine costituiscono una categoria di ormoni di cui molti, dotati di attività le più diverse, sono attualmente noti, ma indubbiamente altri restano da scoprire. Contrariamente a quelli già menzionati, gli ormoni peptidici manifestano, secondo la loro origine zoologica, una specificità nella loro struttura e nel loro grado d'attività. Quest'ultima è stata particolarmente ben evidenziata nel caso degli ormoni neuroipofisari, l'ossitocina e la vasopressina, peptidi che contengono nove amminoacidi, e nelle melanotropine, che ne contengono tredici. Nei primi, per esempio, du Vigneaud e Acher hanno constatato che gli amminoacidi che occupano le posizioni 3, 4, 8 nella catena peptidica non sono uguali nei diversi gruppi di animali: l'isoleucina può essere sostituita con la leucina o la fenilalanina e l'arginina con la lisina. Lo stesso vale per la vasopressina, in cui un resto d'arginina presente nell'ormone del maiale è sostituito da un resto di lisina in quello di altri Mammiferi; la prima è di gran lunga più attiva come inibitore della diuresi. Tra gli altri ormoni peptidici, alcuni, come le angiotensine, sono deca- e ottopeptidi, e i releasing factors, RF, hanno una struttura simile. Altri presentano un peso molecolare di gran lunga più elevato: il glucagone pancreatico è un polipeptide con 29 amminoacidi e l'ormone corticotropo dell'ipofisi anteriore o ACTH ne comprende 39. In quest'ultimo è stata rivelata una differenza di struttura tra l'uomo, il bue, la pecora e il maiale. Le altre stimoline sono proteine di peso molecolare variabile tra 20.000 e 30.000; anche l'insulina è costituita da subunità di peso molecolare di circa 6.000, associate per 2, 6, o 8 in molecole di peso molecolare 12.000, 36.000 e 48.000, e in diversi Mammiferi mostra differenze in un piccolo numero di amminoacidi delle sue due catene.
La conoscenza della struttura delle sostanze naturali ha permesso di produrne la maggior parte per sintesi e di preparare composti organici che contengono la parte biologicamente attiva della loro struttura. La sintesi degli ormoni steroidei, quella dell'adrenalina, dei peptidi postipofisari, hanno reso grandi servigi alla terapia, per la possibilità che hanno di essere utilizzate correntemente. Quella dell'insulina, realizzata dal 1965, non può mettere in concorrenza i prodotti naturali e sintetici per ragioni di ordine economico. Uno dei motivi di interesse nella preparazione chimica di ormoni peptidici o proteici è la possibilità di arrivare a produrre per sintesi la loro sola regione attiva, e ciò ha già permesso di realizzare gli analoghi strutturali dei più attivi peptidi naturali e permetterà senza dubbio di preparare omologhi inferiori di ormoni proteici privi di specificità zoologica e di potere antigene.
b) Biosintesi, metabolismo e modalità d'azione degli ormoni
La preparazione di ormoni puri e l'accertamento della loro struttura hanno permesso di affrontare lo studio della loro biosintesi e del loro metabolismo periferico, studio dal quale si sono potuti trarre alcuni concetti generali. La biosintesi di ormoni nell'interno di cellule differenziate esige la presenza in queste ultime di meccanismi enzimatici particolari. Sono stati identificati numerosi enzimi specifici di una reazione che dà origine a un ormone a partire da un precursore. Così, la formazione dei glucocorticoidi surrenali risulta da azioni successive di una 3β-olo-deidrogenasi specifica del 5-pregnenolone, che dà origine al progesterone, a partire dal quale l'azione di una 17-idrossilasi conduce al 17-idrossiprogesterone; successivamente le azioni di una 21- idrossilasi e di una 11-idrossilasi conducono al 17-idrossi-corticosterone o cortisolo, che è fisiologicamente il più importante dei derivati di questa serie di composti. Nello stesso modo, l'azione successiva di una 18-idrossilasi, poi di una 18-aldeidasi sul cortisolo conduce alla formazione dell'aldosterone. Così, l'intervento di enzimi specifici conduce non solamente a trasformazioni molecolari in una stessa serie di ormoni, quella dei corticosteroidi nell'esempio citato, ma anche alla formazione, a partire da certi ormoni, di altri che appartengono a una serie biologicamente diversa, per esempio di androgeni a partire dal corticosterone o dal cortisolo.
Il trasporto di ormoni e il loro fissarsi agli organi bersaglio sono assicurati da meccanismi specifici per ognuno o particolari a un piccolo gruppo di essi. Si sono potute mettere in evidenza nel siero α-globuline che assicurano il trasporto degli ormoni tiroidei grazie all'affinità elettiva che esse presentano per questi ultimi e lipoproteine che veicolano gli estrogeni. La penetrazione degli ormoni nella cellula può nello stesso modo fare entrare in gioco l'elevata affinità delle proteine che formano con essi complessi dissociabili. I recettori cellulari captano gli ormoni che esercitano la loro attività o nelle immediate vicinanze o al loro livello, ma queste sostanze non si accumulano nelle cellule, dove la loro degradazione richiede un rinnovamento continuo affinché sia assicurata la regolazione del loro tasso. Il catabolismo degli ormoni conduce all'inattivazione di tali sostanze con un meccanismo che mette in azione enzimi appropriati alla loro struttura. Per esempio, la tiroxina è degradata sia da un'amminoacidossidasi che assicura la deamminazione ossidativa del suo residuo di alanina, sia da una deiodasi che stacca lo iodio dai suoi anelli benzenici.
La regolazione del tasso cellulare degli ormoni e il loro meccanismo d'azione hanno posto problemi il cui studio è stato possibile affrontare nel quadro della biologia molecolare. L'azione degli ormoni non si svolgerebbe secondo un meccanismo unico; essa, ciò nonostante, ha potuto essere, collegata in un gran numero di casi a una regolazione della sintesi cellulare di proteine enzimatiche, ammettendo la possibilità dell'induzione o della repressione del processo. Karlson ha schematizzato questo fenomeno nel seguente modo: nella cellula bersaglio, il DNA cromosomico è caricato di repressori enzimatici; esso cede all'ormone, che lo capta, il repressore specifico della biosintesi dell'enzima di cui il gene corrispondente produce l'RNA messaggero (m-RNA); questo messaggero specifico migra allora nel citoplasma dove dirige la biosintesi della proteina enzimatica, della quale la regolazione ormonale controlla l'attività e il tasso. Poiché la maggior parte dei messaggeri endocrini mettono in gioco l'adenosin-3′,5′-monofosfato ciclico (c-AMP), di cui Sutherland ha mostrato la funzione generalizzata, si è pensato che l'azione di numerosissimi ormoni si eserciti mediante la regolazione cellulare dell'attività dell'enzima ciclizzante l'AMP in c-AMP, l'adenilciclasi; oppure della fosfodiesterasi specifica, che trasforma il c-AMP in acido adenosin-3′-fosforico (AMP) inattivo.
c) Endocrinologia medica e biochimica
La pratica clinica ha saputo collegare con una precisione sempre maggiore numerose sindromi all'esistenza di disturbi endocrini e istituirne una terapia (v. Hermann e Cier, 1970; v. Régulations hormonales..., 1937; v. Hall e altri, 1968; v. Les troubles congénitaux..., 1965). La spiegazione di sintomi che all'inizio ci si è limitati a descrivere senza poterne interpretare il meccanismo di produzione ha permesso l'identificazione di diverse sindromi endocrine. Così, il virilismo che è stato visto svilupparsi in alcuni soggetti si è potuto collegare, attraverso una serie di dati biochimici precisi, a un'iperproduzione patologica di aldosterone a spese del cortisone e dei glucocorticoidi surrenalici. Ora, poiché questo fenomeno era stato dapprima evidenziato in donne portatrici di tumori dell'ipofisi anteriore, lo si era attribuito, mancando ogni possibilità di controllo, a uno squilibrio della secrezione di ormoni sessuali da parte delle gonadi.
L'identificazione di stati patologici caratteristici di disturbi endocrini particolari e la conoscenza della trasmissione genetica di alcuni di essi sono diventate sempre più precise man mano che nuove scoperte biochimiche hanno apportato alla medicina più ampie possibilità di diagnosi e terapia (v. Hermann e Cier, 1970; v. Régulations hormonales..., 1937; v. Hall e altri, 1968). I progressi nella tecnica del dosaggio degli ormoni circolanti nel sangue o escreti con l'urina sono stati, a questo proposito, particolarmente utili. Sarebbe stato impossibile identificare per mezzo dell'analisi dei liquidi dell'organismo un disturbo della secrezione corticosurrenale dei mineralcorticoidi e dei glucocorticoidi prima di poter dosare gli uni o gli altri selettivamente e anche alcuni di essi isolatamente. Nel campo della chimica clinica, il caso del diabete pancreatico, di cui alcune manifestazioni si erano potute mettere in luce studiando la glicosuria e la glicemia, ha costituito per lungo tempo una eccezione straordinariamente favorevole. Allo stato attuale della biochimica, il clinico è in grado di mettere in evidenza non solo gravi disturbi, ma anche lievi squilibri endocrini molto prima che essi raggiungano stati gravi, a volte difficilmente reversibili, e di compensarli. Infine, le possibilità terapeutiche sono diventate molto più ampie grazie alla preparazione di prodotti naturali puri o di prodotti di sintesi identici facilmente disponibili.
La chimica organica ha fornito in questo campo al clinico nuove armi per la preparazione di molecole non naturali, dotate specificamente di particolari attività ormonali, di loro antagonisti o di inibitori di processi ormonogenici. Il cancro della prostata regredisce sotto l'azione degli estrogeni, ma i prodotti naturali di questa serie provocano nell'uomo un certo numero di manifestazioni che rendono difficile il loro impiego terapeutico. Fu E. C. Dodds a sintetizzare estrogeni di un nuovo tipo strutturale il cui uso non determina tali inconvenienti, così che la terapia del cancro della prostata per mezzo del dietilstilbestrolo produce da quasi trenta anni la guarigione del tumore, e anche delle sue metastasi, nella maggior parte dei casi. Antagonisti ormonali, i cui effetti sono stati messi in evidenza nella cancerologia sperimentale, sono senza dubbio rivelatori di possibilità terapeutiche future. Così A. Lacassagne ha stabilito che la somministrazione a forti dosi di estrone provoca la comparsa di un carcinoma mammario nei topi maschi e di un epitelioma uterino nei topi femmina o nelle coniglie: orbene, il progesterone o gli ormoni testicolari, antagonisti fisiologici degli estrogeni, inibiscono anche la loro azione cancerogena (v. neoplasie: Oncologia sperimentale).
Gli antagonismi si manifestano sul piano fisiologico e possono anche esercitarsi attraverso meccanismi diversi che non comportano necessariamente un'azione inversa a quella di un ormone. Per es., alcuni solfammidici sono utilizzati come ipoglicenizzanti nei diabetici, ma la loro azione si esercita attraverso la stimolazione della secrezione pancreatica di insulina e la liberazione di quest'ultima a partire da complessi insulino-proteici presenti nelle cellule β degli isolotti di Langerhans. L'impiego di inibitori diretti o indiretti della secrezione o della biosintesi di ormoni sembrava destinata a un grande avvenire terapeutico e già attualmente è entrata nella pratica corrente per il controllo delle nascite, per mezzo di prodotti di sintesi che ostacolano l'ovulazione nella donna e vengono chiamati antiovulatori.
Un importante progresso analitico per la valutazione quantitativa degli ormoni circolanti è stato determinato recentemente dalla messa a punto dei metodi radioimmunologici. Il principio del metodo è fondato sull'inibizione competitiva esercitata da parte di una molecola ormonale marcata con un isotopo radioattivo sul complesso antigeneanticorpo costituito dall'ormone da misurare cimentato con il suo antisiero specifico. Questo metodo è particolarmente adatto per la determinazione degli ormoni proteici o peptidici. Più di recente il metodo radioimmunologico è stato anche utilizzato per ormoni non proteici legati a molecole proteiche, come ad esempio per la triiodotironina resa antigenicamente attiva mediante incorporazione nella molecola sieroalbuminica.
4. Considerazioni conclusive
I progressi dell'endocrinologia, di cui si è tentato di cogliere le tappe successive, sono oggi lontani dal rivelare una battuta d'arresto, poiché le grandi vie di ricerca che hanno aperto resteranno indubbiamente ancora a lungo feconde. La scoperta recente delle prostaglandine nei Mammiferi e quella dei feromoni negli Insetti lasciano sperare che molti prodotti endocrini siano ancora da identificare. Anche l'evidenziazione di meccanismi regolatori così complessi come quello che assicura la coordinazione ipotalamo-ipofisaria o la regolazione dell'immissione in circolo dell'aldosterone da parte dell'angiotensina illustrano le possibilità future di ricerca della fisiologia e della patologia delle secrezioni endocrine. Lo studio del meccanismo d'azione degli ormoni, poi, non è che all'inizio, poiché è certo che nella maggior parte dei casi ci si è finora limitati a descrivere gli effetti di queste sostanze senza aver potuto capire i fenomeni cellulari che li traducono. Il fatto che un insieme di ormoni partecipi al metabolismo del glucosio e alla regolazione del suo tasso sanguigno è, a questo proposito, un esempio caratteristico. È la biochimica endocrina che ha, in ogni caso, un avvenire pieno di promesse.
Conoscere la costituzione degli ormoni permette di correlare l'attività della loro molecola alla loro struttura, di preparare prodotti analoghi a quelli naturali e, a volte, di scoprirne di nuovi. La sintesi operata da Reichstein di un corticosteroide allora nuovo ha preceduto l'evidenziazione di tale composto come ormone corticosurrenale. Composti che possiedono nella loro molecola la zona attiva possono essere preparati per sostituire i prodotti naturali o per essere utilizzati come loro antagonisti in terapia, per la loro azione sia a livello delle cellule bersaglio sia a quello delle cellule ormonogene. Le frasi che seguono sono di R. Courrier (lettera personale all'autore datata 18 luglio 1972) e meritano di essere utilizzate come conclusione a questo capitolo: ‟È la biochimica che darà la prova che l'endocrinologia non arriverà mai alla fine della sua via. Quando tutti gli ormoni naturali saranno descritti e sintetizzati, i chimici costruiranno senza tregua nuove molecole, sostanze artificiali con le proprietà di ormoni naturali senza possederne la struttura". Queste frasi mettono in rilievo il ruolo che i biochimici continueranno ad assumere, in stretta collaborazione con i fisiologi, nello sviluppo dell'endocrinologia.
bibliografia
Cameron, A. T., Recent advances in endocrinology, London 1933.
Gabe, M., Karlson, P., Roche, J., Hormones in invertebrates, in Comparative biochemistry (a cura di M. Florkin e H. S. Mason), vol. VI, Cells and organisms, New York 1964, pp. 245-298.
Gley, E., Quatre leçons sur les sécrétions internes, Paris 1921.
Hall, R. J., Anderson, J., Smart, G. A., Fundamentals of clinical endocrinology, London 1968.
Harris, R. S., Thimann, K. V. (a cura di), Vitamins and hormones, 31 voll., New York 1942-1972.
Hermann, H., Cier, J. F., Endocrinologie, in Précis de physiologie, vol. IV, Paris 1970, pp. 1-390.
Karlson, P., Hormone, in Kurzes Lehrbuch der Biochemie für Mediziner und Naturwissenschaftler, Stuttgart 19644, pp. 321-344 (tr. it.: Biochimica, Milano 1966, pp. 379-410).
Les troubles congénitaux de l'hormonogénèse. Rapports présentés à la VIIIe réunion des endocrinologistes de langue française, Paris 1965.
Pincus, G. (a cura di), Recent progress in hormone research, 29 voll., New York 1944-1972.
Pincus, G., Thimann, K. V., Astwood, E. B. (a cura di), The hormones, 5 voll., New York 1948-1964.
Régulations hormonales en biologie, en clinique et en thérapeutique. Série de rapports d'intérêt historique présentés aux journées médicales internationales de Paris, 1937, Paris 1937.
Wolff, E., Hormones et différentiation sexuelle chez les invertébrés, Paris-London 1972.