
Microbiologia
Microbiologia
di John F. Wilkinson
Microbiologia
sommario: 1. Introduzione. 2. Un organismo modello, Escherichia coli: a) La struttura di Escherichia coli. b) La crescita di Escherichia coli. c) La genetica di Escherichia coli. 3. I Procarioti. a) Variazioni strutturali dei Procarioti. b) La gamma dei processi fisiologici nei Procarioti. c) Lo sviluppo in ambienti proibitivi. 4. Un organismo modello, Saccharomyces cerevisiae. 5. I microrganismi eucarioti. 6. I Virus; sviluppo della virologia. a) Un organismo modello, i fagi T2 e T4 di Escherichia coli. b) Altri Virus. 7. Classificazione ed evoluzione dei microrganismi. 8. Le applicazioni della microbiologia. a) Microbiologia medica. b) Microbiologia ambientale e agricola. c) Microbiologia alimentare e industriale. □ Bibliografia.
1. Introduzione
Che cos'è la microbiologia? Convenzionalmente si potrebbe dire che è lo studio dei microrganismi. Che cosa sono i microrganismi? La risposta convenzionale potrebbe essere: sono gli organismi studiati dai microbiologi. Chi sono i microbiologi? Ovviamente si potrebbe rispondere che sono coloro che studiano la microbiologia. È chiaro che queste definizioni tautologiche sono prive di valore, ma riflettono la reale difficoltà che i microbiologi incontrano quando vogliano definire il tipo di organismi che osservano. Esse rispecchiano anche la difficoltà, progressivamente crescente, di scindere la biologia in una serie di discipline distinte e separate che, sebbene notevolmente aumentate di numero in questo secolo, in effetti sono divenute sempre meno significative a livello di ricerca.
Nel secolo scorso esistevano solamente due ‛regni' di organismi viventi: quello animale e quello vegetale, campi di ricerca rispettivamente degli zoologi e dei botanici. La microbiologia si venne sviluppando molto lentamente, legata al perfezionamento di un unico strumento, il microscopio. È noto che A. van Leeuwenhoek, all'inizio del XVII secolo, osservò per primo, utilizzando un microscopio semplice a una sola lente, ciò che oggi definiamo con il termine di microrganismi, ma le sue tecniche erano al di fuori della portata di ricercatori di abilità e pazienza medie. Solo nel Novecento sono stati messi a punto microscopi composti che hanno consentito di osservare i Batteri con relativa facilità e, sempre in questo secolo, sono state perfezionate anche altre due tecniche che si sono dimostrate basilari per lo sviluppo della microbiologia: la prima è consistita nella messa a punto dei metodi di sterilizzazione, che hanno consentito di eliminare dai terreni di coltura i microrganismi contaminanti; la seconda, sviluppatasi dalla precedente, è consistita nell'introduzione dei metodi per separare i diversi microrganismi tra di loro e per mantenerli in coltura pura.
In seguito all'applicazione di queste tecniche furono individuati i principali gruppi di microrganismi, eccezion fatta per i Virus; però l'attenzione di gran lunga maggiore venne rivolta ai microrganismi patogeni e tali ricerche fornirono l'impulso principale per lo sviluppo della microbiologia nella prima metà di questo secolo: dipartimenti di batteriologia o di microbiologia sono sorti in seno alle facoltà mediche universitarie. I principali gruppi di microrganismi studiati sono stati i Batteri e, dopo la loro scoperta nei decenni a noi più vicini, i Virus; ma anche le Alghe, i Funghi e i Protozoi hanno cominciato a uscire da un periodo in cui erano stati relativamente trascurati e vengono in genere considerati, più giustamente, microrganismi e fatti rientrare quindi nell'ambito della microbiologia. Pertanto oggi si è soliti distinguere cinque gruppi principali di microrganismi (Virus, Batteri, Alghe, Funghi e Protozoi) e dividere conseguentemente la microbiologia in virologia, batteriologia, ficologia, micologia e protozoologia. Tale ampliamento delle competenze della nostra disciplina ha determinato la creazione di dipartimenti di microbiologia nelle facoltà di scienze. Entro certi limiti questa separazione dei microrganismi dagli animali e dalle piante è basata, come suggerisce il termine microbiologia, sul fatto che la maggior parte degli organismi compresi in questo gruppo è di dimensioni microscopiche e, normalmente, unicellulare; quando sono organismi multicellulari non si riscontra un reale differenziamento rispetto ai tessuti degli animali e delle piante. Forse però la giustificazione più importante dell'esistenza della microbiologia come scienza biologica è che le tecniche adottate per lo studio di tutti i gruppi di microrganismi di sua competenza sono simili tra loro e ben diverse da quelle impiegate dai botanici e dagli zoologi.
All'inizio di questo secolo la microbiologia era ancora una scienza molto poco sviluppata, specialmente per ciò che riguardava gli aspetti non applicativi. Nell'ambito della microbiologia medica gli sforzi maggiori erano limitati alla ricerca dei metodi migliori per l'isolamento e l'identificazione dei microrganismi patogeni e conseguentemente al perfezionamento dei metodi diagnostici: sebbene i principi generali dell'igiene elementare e della disinfezione fossero ormai codificati, era opinione comune che ogni tentativo di eliminare i microrganismi patogeni, una volta che si fossero localizzati nell'organismo umano, fosse destinato a fallire. Al di fuori del campo medico si erano ottenuti alcuni dati sul ruolo dei microrganismi nel terreno e quindi sulla loro importanza per l'ecosistema generale e per l'agricoltura, mentre le conoscenze erano estremamente limitate per ciò che riguarda il ruolo dei microrganismi nell'acqua dolce o salata. In campo industriale estremamente modesti erano stati i progressi dal punto di vista microbiologico, eccezion fatta per l'acquisizione che procedimenti tradizionali, quali la produzione di bevande alcoliche, di pane, di formaggi e di altri prodotti del genere, erano effettivamente processi microbiologici. Quindi il progresso compiuto al di fuori della medicina era esiguo, se si considera la microbiologia una scienza applicata, e la situazione era anche peggiore se la si valuta come scienza di base. In altri termini, all'inizio del secolo la microbiologia era agli albori; oggi è divenuta una disciplina con un notevole margine d'azione e con molteplici scopi, in possesso di un'impressionante quantità di profonde conoscenze in molti campi. Noi oggi conosciamo un microrganismo - Escherichia coli - meglio di qualsiasi altro organismo, eccetto forse l'uomo.
In breve la microbiologia, al pari della biochimica e della biologia molecolare, con il cui sviluppo è intimamente connessa, è effettivamente una scienza di questo secolo: la descrizione del suo sviluppo nel corso del Novecento equivale pertanto a un compendio della storia della microbiologia.
2. Un organismo modello, Escherichia coli
Escherichia coli è un batterio molto tipico che si riscontra nell'intestino e che un quarto di secolo fa era noto principalmente come indice di contaminazione fecale delle riserve idriche. I motivi per cui è stato scelto come modello, per delucidare molti principi basilari di biologia molecolare e di genetica molecolare, sono in parte razionali e in parte casuali. È un microrganismo che cresce facilmente in terreni di coltura semplici, contenenti una sostanza organica qualsiasi, come fonte di carbonio e di energia, insieme a pochi sali inorganici. Come la maggior parte dei Batteri esso è in grado di adattarsi a molteplici condizioni ambientali; ciò è ben documentato dalla sua capacità di crescere utilizzando varie sostanze come fonti di carbonio e di energia ed entro intervalli abbastanza ampi di pH, di pressione osmotica e di tensione di ossigeno. Infine viene attaccato da numerosi virus.
La scelta di Escherichia coli come sistema modello in biologia derivò dall'estensione del concetto generale che, particolarmente a livello biochimico, genetico e di biologia molecolare, esiste un'uniformità nei principi che regolano gli organismi viventi. E con lo sviluppo delle ricerche biochimiche negli anni trenta e quaranta, divenne chiaro che tutti gli organismi viventi sono formati, per lo più, dagli stessi costituenti chimici: acidi nucleici, proteine, polisaccaridi e lipidi; che i costituenti elementari delle proteine e degli acidi nucleici sono sostanzialmente gli stessi e vengono sintetizzati con processi metabolici spesso identici; che le vitamine e i fattori essenziali per l'accrescimento sono simili, poiché necessari per costituire alcuni coenzimi e certi gruppi prostetici - l'acido nicotinico, la riboflavina, la biotina, la vitamina B12, il piridossale, la tiamina, ecc. - diffusi universalmente; che i meccanismi di produzione, trasferimento e utilizzazione dell'energia, implicanti tutti normalmente sistemi mediati dall'ATP e la formazione di legami fosforici altamente energetici, sebbene diversi nei dettagli, si esplicano attraverso gli stessi processi generali. Con lo sviluppo della biologia molecolare apparve altresì chiaro che la conservazione e il trasferimento dell'informazione genetica sono essenzialmente comuni a tutti gli organismi viventi (v. acidi nucleici; v. biologia). Nel corso di questo sviluppo, e in particolare per verificare l'ipotesi dell'‛unità biochimica', divenne usuale tentare di convalidare tale ipotesi con lo studio degli organismi più strani dal punto di vista fisiologico. Spesso si trattava di microrganismi e tra questi uno dei più diffusamente scelti fu Thiobacillus thiooxidans, batterio che si sviluppa su composti contenenti zolfo inorganico ridotto quale unica fonte di energia e di potere riducente necessari per ridurre il CO2 a materiale organico cellulare. Il processo può essere così schematizzato:
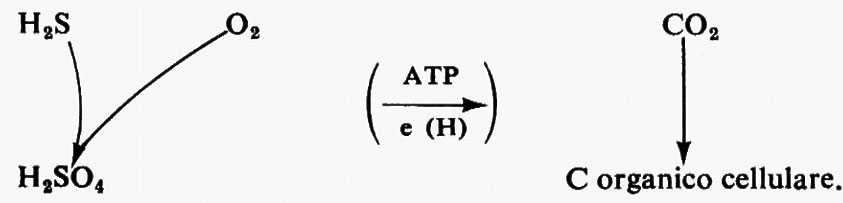
I normali metaboliti organici non possono essere utilizzati da questo batterio per la sua crescita, anzi spesso la inibiscono; inoltre il suo optimum di pH, per lo sviluppo, è di circa 3. Se un microrganismo di questo tipo è dotato di una struttura biochimica di base simile a quella di organismi più evoluti, sicuramente allora l'ipotesi dell'unità è una certezza. Ciò è stato dimostrato: in questo batterio, infatti, l'energia viene prodotta mediante un processo di fosforilazione ossidativa in un sistema di trasporto di elettroni del tutto convenzionale (v. fosforilazione ossidativa); incidentalmente il donatore di, elettroni può essere un composto inorganico anziché organico. Il CO2 viene ridotto a carbonio organico cellulare attraverso il ciclo di Calvin, processo fondamentalmente identico a quello presente negli organismi fotosintetici (v. fotosintesi); infine i processi biosintetici, i coenzimi e gli enzimi necessari sono simili o identici.
Una volta chiarito che l'unità biochimica esiste, ne deriva che, se si desidera studiare un particolare biologico fondamentale, si deve scegliere ovviamente l'organismo più adatto a tale ricerca, senza tener conto del fatto che sia un animale, una pianta o un microrganismo. In pratica si è visto che i Batteri sono spesso i modelli sui quali è più facile compiere tali indagini. Inoltre è utile impiegare quanto più è possibile un solo organismo e raccogliere su di esso il maggior numero possibile di dati; ciò consente di superare il problema del coordinamento dei risultati di ricercatori diversi nei vari laboratori del mondo e consente anche una migliore integrazione dei risultati da punti di vista biologici diversi. Escherichia coli è stato scelto come organismo modello.
a) La struttura di Escherichia coli
Il microrganismo si presenta sotto forma di cellule bastoncellari di circa 1 μm di lunghezza e 0,5 μm di diametro. Ben poco si sapeva sulla struttura interna di tali cellule sino all'avvento del microscopio elettronico, poiché il potere risolutivo del microscopio ottico è, ai fini pratici, solo di circa 0,2 μm. Lo sviluppo dei metodi delle sezioni ultrasottili, insieme a quello delle tecniche di frazionamento dei componenti cellulari, ha consentito di comporre un quadro più chiaro della struttura, quadro che è stato confermato e ampliato dalle tecniche d'indagine più recenti.
La cellula è di una semplicità essenziale. Il citoplasma è circondato da una membrana citoplasmatica che, nelle sezioni ultrasottili, risulta identica alle membrane di tutte le altre cellule, è cioè una ‛membrana unitaria' formata da due strati elettrondensi tra i quali è interposto uno strato trasparente agli elettroni, ed è costituita da proteine e da lipidi. Nella membrana citoplasmatica sono localizzate delle molecole di permeasi proteica che le conferiscono una caratteristica semipermeabilità, consentendo solo ad alcune molecole di entrare o di uscire dalla cellula e permettendo anche una maggior concentrazione di sostanze nutritizie all'interno della cellula rispetto all'ambiente circostante. Questa capacità di concentrazione è estremamente importante per molti microrganismi che si sviluppano in ambienti naturali dove le sostanze nutritizie sono in concentrazioni estremamente basse. Bisogna rilevare che la membrana citoplasmatica è l'unica membrana semipermeabile di Escherichia coli, che pertanto è una cellula costituita da un unico compartimento. Inoltre la membrana è sia la sede di produzione dell'energia (ATP), sia la struttura su cui è localizzata la maggior parte degli enzimi responsabili della sintesi della parete e della capsula e interviene probabilmente come punto di attacco per il cromosoma. La membrana può presentare speciali zone invaginate dette ‛mesosomi': queste formazioni sono rare in Escherichia coli, ma in alcuni batteri sono costantemente presenti e si ritiene siano in rapporto con la formazione della parete trasversa (crosswall) o con la divisione nucleare.
In seno al citoplasma la struttura più importante che si può osservare su sezioni ultrasottili è il nucleo amorfo. Esso non è circondato da una membrana, ma sembra costituito da un'unica molecola, in genere circolare, di DNA a doppia elica, che è l'unico cromosoma del microrganismo. Tale cromosoma è lungo anche mille volte più della stessa cellula ed è pertanto ripiegato in modo complesso a formare il nucleo amorfo. Il nucleo, a partire da un punto iniziale, si divide in due cromosomi che vengono segregati, probabilmente per la scissione del punto d'attacco sulla membrana citoplasmatica, nelle due metà della cellula, prima che questa si divida per scissione binaria. Non vi sono mitosi, né apparato mitotico. Le altre strutture che costantemente si possono osservare nel citoplasma sono i ribosomi, caratterizzati dalla consueta funzione protidosintetica. Inoltre, in determinate condizioni ambientali, possono anche essere presenti granuli di glicogeno, trasparenti agli elettroni, che esplicano una funzione di riserva, analoga a quella svolta nelle cellule animali.
Intorno alla membrana citoplasmatica c'è una parete cellulare che al microscopio elettronico appare come una struttura pluristratificata. Essa è il costituente più caratteristico della cellula, in quanto contiene non solo sostanze tipiche dei Batteri, per ciò che riguarda la composizione chimica generale, ma anche componenti specifici di Escherichia coli e di altri batteri con esso correlati, per ciò che si riferisce alla struttura in dettaglio. La forma della parete e la sua resistenza alle sollecitazioni meccaniche sono dovute a un polimero complesso costituito da catene polisaccaridiche derivate dalla polimerizzazione di unità alternantisi di N-acetilglucosammina e di acido N-acetilmurammico, collegate tra loro da brevi peptidi che contengono alcuni insoliti D-amminoacidi. Tale struttura, detta generalmente mucopeptide, sembra che sia una sola enorme molecola, tenuta insieme da legami covalenti, a foggia di sacchetto, con la forma della cellula. Probabilmente il meccanismo che controlla la sua sintesi e i processi di associazione delle catene polisaccaridiche determina la forma della cellula e la sua delucidazione offrirà un campo di ricerca sempre più importante nei prossimi decenni. Il possesso di una parete cellulare glicoproteica da parte di Escherichia coli e di altri batteri è risultato una circostanza estremamente favorevole per l'uomo, poiché la sua sintesi rappresenta il sito d'azione di molti antibiotici di uso comune, come ad esempio la penicillina: l'inibizione ditali processi sintetici provoca indebolimento della parete cellulare e, eventualmente, usi e morte della cellula, mentre l'uomo, che non possiede polimeri simili al mucopeptide, non subisce effetti tossici da antibiotici di questo tipo. Oltre al mucopeptide la parete cellulare contiene lipidi, polisaccaridi e proteine disposti in modo da formare una struttura pluristratificata. Il più caratteristico di questi costituenti è il lipopolisaccaride, che in genere è presente come strato superficiale e quindi rappresenta l'antigene di superficie; esso agisce anche comunemente come recettore per l'aggressione virale. Il lipopolisaccaride va incontro a rapide modificazioni evolutive in conseguenza della selezione operata da virus o da anticorpi specifici e ciò ha portato al riconoscimento di una serie di tipi (sierotipi o tipi batteriofagici). Tale delicata tipizzazione è risultata di notevole importanza in microbiologia medica per seguire l'andamento di un'infezione e quindi per gli studi epidemiologici.
Esternamente alla parete cellulare possono essere presenti altre formazioni: si può riscontrare uno strato capsulare, formato da un gel polisaccaridico, che esplica funzioni protettive contro la fagocitosi, l'aggressione virale e l'essiccamento e, contemporaneamente, è di ausilio per l'assorbimento e la concentrazione degli ioni. Infine, possono esservi due tipi di appendici filiformi. Il possesso di flagelli è in rapporto con la motilità cellulare: si tratta di tubuli elicoidali a loro volta formati da subunità proteiche disposte elicoidalmente; il moto si ha in conseguenza di un unico movimento rotatorio analogo a quello dell'elica di una nave. Tale movimento è soggetto a basilari modalità di comportamento, in quanto presenta risposte chemiotattiche; tuttavia, anche se su questa base sono state fatte valutazioni secondo le quali Escherichia coli, a parità di peso, sarebbe 106 volte più intelligente dell'uomo, non sembra verosimile che i Batteri possano essere impiegati come modelli per lo studio delle modalità di comportamento, eccezion fatta per la chemiotassi! Oltre ai flagelli possono riscontrarsi altri filamenti di diametro inferiore: questi peli o fimbrie non hanno forma elicoidale, sebbene sembri che anch'essi siano costituiti da subunità proteiche disposte elicoidalmente, e non esplicano funzioni locomotorie; intervengono invece nel processo di coniugazione oppure si comportano da apparati adesivi consentendo alle cellule di attaccarsi a superfici quale la parete intestinale.
La struttura di una cellula di Escherichia coli è quindi di una semplicità estrema, fatto questo correlato con le piccole dimensioni della cellula stessa.
b) La crescita di Escherichia coli
Uno dei motivi del successo dei microrganismi, e dei Batteri in particolare, nel mondo vivente è legato al loro esplosivo ritmo di crescita. Un organismo come Escherichia coli, in condizioni ambientali favorevoli, è in grado di dividersi ogni venti minuti: in altri termini raddoppia le proprie dimensioni ogni venti minuti e quindi si divide, per scissione binaria, in due metà uguali. Ne deriva che la crescita sarà esponenziale sino a quando l'ambiente sarà adatto alla conservazione del massimo ritmo di divisione: è facile calcolare che da un'unica cellula, del volume di circa 1 μm3, in venti ore si potrebbero sviluppare 1018 cellule, per un volume complessivo di 1 m3, in quaranta ore 1036 cellule pari a un volume di 1.000 km3 e che prima della cinquantesima ora il volume potrebbe essere superiore a quello della Terra. È questo potenziale di crescita esplosiva che consente ai Batteri di sfruttare un ambiente favorevole in un breve periodo di tempo, ma ciò ha anche permesso ai biologi di far sviluppare una coltura, per ricerche sperimentali, con un'incubazione protratta per una sola notte. Per mantenere questo elevato ritmo di crescita Escherichia coli deve possedere un metabolismo dal ritmo corrispondentemente intenso: per esempio la velocità di consumo dell'ossigeno e quella della sintesi proteica sono circa cento volte maggiori di quelle di una tipica cellula animale o vegetale e anche queste proprietà sono risultate di grande vantaggio per il biochimico che indaga i processi metabolici. Come riesce ad avere capacità metaboliche e di crescita così elevate Escherichia coli? La soluzione di questo problema sembra che risieda soprattutto nella sua piccola mole e quindi nell'elevato rapporto tra superficie e volume. La velocità di crescita di un organismo è in genere limitata dal ritmo di assorbimento di sostanze nutritizie, come ad esempio l'ossigeno, e dalla rapidità con cui vengono eliminati i prodotti catabolici. Negli animali e nelle piante il rapporto reale tra superficie e volume può essere incrementato dai sistemi circolatori, ecc., e tuttavia un piccolo batterio presenta ancora un vantaggio potenziale estremamente grande. Allora quanto può essere piccola una cellula, pur continuando a svolgere compiutamente le sue funzioni? Se essa sintetizza la maggior parte dei suoi costituenti cellulari, ha bisogno di enzimi in numero e in quantità sufficienti; ciò richiede un genoma opportunamente ampio associato a un efficiente apparato protidosintetico. Tutti questi fattori pongono dei limiti al volume minimo efficiente di una cellula, al di sotto del quale, infatti, è dubbio che sia possibile un qualsiasi apprezzabile incremento del ritmo di crescita. In Escherichia coli in fase di crescita al massimo ritmo, più di un quarto della cellula, in peso secco, è costituito dai ribosomi necessari per mantenere un'adeguata rapidità di sintesi proteica; un ulteriore raddoppio del ritmo di crescita richiederebbe un raddoppio del contenuto cellulare in ribosomi e questi di per sé imporrebbero alla cellula un impossibile tenore biosintetico. Anche se esistono cellule più piccole di Escherichia coli, quali ad esempio i micoplasmi, esse vivono in un ambiente di sviluppo molto specializzato, in cui, grazie alla loro esistenza parassitaria, si riforniscono di metaboliti complessi; ciò permette la funzione di un genoma più piccolo, che nei micoplasmi è della grandezza di circa 0,4×109 dalton, mentre in Escherichia coli è di 2,2×109 dalton. In altri termini, Escherichia coli rappresenta il limite inferiore di dimensione che consente di condurre in maniera efficiente un'esistenza non parassitaria e tale piccola mole è compatibile solo con una struttura cellulare semplice.
Una crescita al massimo ritmo possibile dipende da vari fattori, come la temperatura, il pH, la tensione di ossigeno, la pressione osmotica e sostanze nutritizie in concentrazione ottimale. Un'analisi dei materiali nutritizi necessari a Escherichia coli ha posto in risalto che, sebbene esso possa crescere su un terreno nutritivo complesso, come ad esempio un idrolizzato proteico arricchito con estratto di carne, puo anche svilupparsi in un terreno contenente una sola fonte di energia e di carbonio, insieme ad altre sostanze nutritizie soltanto inorganiche; ciò è dovuto alla sua capacità biosintetica di elaborare tutti i propri costituenti chimici a partire da questi semplici materiali costruttivi. Tuttavia, lo studio dell'effetto delle variazioni del tipo delle sostanze nutritizie, disponibili durante la crescita, sulla concentrazione intracellulare degli enzimi ha portato a un'acquisizione che è risultata d'importanza fondamentale per lo sviluppo della biologia molecolare: alcuni enzimi, detti costitutivi, non presentano alcun rapporto con l'ambiente, mentre altri, detti adattativi, vengono elaborati soltanto in risposta a particolari condizioni. Questa capacità di adattarsi a una grande varietà di condizioni ambientali diverse è stata di notevole importanza per i biologi ed è causa di una netta distinzione rispetto a molte cellule animali e vegetali che si sviluppano in condizioni ambientali molto controllate. Ulteriori indagini sugli enzimi adattativi hanno consentito di distinguerli in due tipi: 1) enzimi inducibili che prendono soprattutto parte a processi catabolici o di scissione; essi vengono elaborati soltanto in presenza di un induttore specifico che in genere è il substrato su cui agisce l'enzima. Un enzima inducibile, la β-galattosidasi di Escherichia coli, è divenuto un modello famoso per lo studio del controllo della sintesi degli enzimi; 2) enzimi reprimili che intervengono nei processi biosintetici; essi vengono sintetizzati soltanto se non è presente il prodotto finale di un dato processo biosintetico che reprime la loro sintesi.
Uno studio di questi enzimi ha portato alla teoria di Fr. Jacob e J. Monod del controllo della sintesi enzimatica; tale teoria dimostra come Escherichia coli, per la sua adattabilità all'ambiente, possa essere scelto come sistema modello per la dimostrazione di un principio biologico apparentemente generale: il controllo della sintesi enzimatica.
Un argomento correlato con la crescita batterica e che ha assunto notevole rilievo negli ultimi due decenni è la tecnica di coltura continua. Prima del suo sviluppo la coltura dei Batteri veniva effettuata in sistemi chiusi (a lotto o batch) nei quali veniva inoculato il microrganismo da coltivare. La crescita continuava sino a quando il terreno non era più adatto o per l'esaurimento delle sostanze nutritive o per l'accumulo di prodotti tossici dovuti alla crescita batterica stessa; la coltura allora entrava in una fase stazionaria, nella quale non si verificava una crescita evidente e, dopo un periodo variabile di tempo, si aveva l'estinzione. In condizioni di coltura continua si crea un sistema nel quale si ha un'aggiunta continua o pressoché continua di nuove sostanze nutritive nel recipiente di coltura (o fermentatore) e, con identico ritmo, in genere ottenuto mediante un apparato che mantiene costante il livello, si ha l'allontanamento delle cellule sviluppatesi e dei loro prodotti. Questo sistema può essere mantenuto per lunghi periodi di tempo durante i quali l'ambiente nel recipiente di coltura rimane in fase stazionaria: ciò è di notevole importanza per il biologo che voglia indagare gli effetti dell'ambiente, ma è utile anche in quanto assicura cellule identiche dal primo all'ultimo giorno, poiché si può ritenere che, fatta eccezione per gli eventi del ciclo cellulare, esse si trovino in una fase stazionaria. Un altro vantaggio che la coltura continua offre è rappresentato dal fatto che essa imita le situazioni che si verificano in sistemi di crescita naturali, per esempio nei processi che hanno luogo nel rumine, molto più di una coltura in sistema chiuso. Infine la coltura continua trova un'applicazione sempre più diffusa in ambito industriale non solo per la maggior redditività, espressa in termini di capitale fisso, che essa offre rispetto alla coltura a mescolamento, ma anche perché consente un più facile controllo dell'ambiente e quindi un aumento quantitativo del prodotto.
c) La genetica di Escherichia coli
Nei primi decenni di questo secolo si sapeva ben poco sulla genetica dei Batteri. Esistevano dubbi sia sulla presenza di un nucleo, come normalmente s'intende, sia sulla possibilità di mutazione spontanea, e, come spesso accade in campo scientifico, questa scarsa conoscenza si rifletteva nella veemenza delle diverse opinioni che si contendevano il campo. I dubbi cui abbiamo accennato vennero chiariti soltanto dopo il 1940.
È noto, come abbiamo già ricordato, che Escherichia coli non possiede un nucleo ‛classico' circondato da una membrana e che pertanto non presenta né mitosi né meiosi; nel batterio c'è invece un filamento circolare di DNA che rappresenta il nucleo formato da un unico cromosoma. Mutazioni spontanee si verificano con una frequenza simile a quella che si riscontra in altri organismi viventi e, poiché Escherichia coli, al pari di altri batteri, è normalmente aploide, la mutazione è facilmente individuabile. Una quantità sempre maggiore di mutageni ha trovato impiego nella ricerca dei principî del processo di mutazione e Escherichia coli è stato utilizzato anche in questo campo come materiale di scelta. Mutanti resistenti ai mutageni sono stati isolati e analizzati per delucidare la natura dei processi intracellulari che portano alla ristrutturazione del DNA. Per la facilità con cui si ottengono mutanti fenotipici e per la capacità di Escherichia coli di opporsi a un ampio spettro di condizioni ambientali e quindi a rilevanti pressioni selettive, che possono essere esercitate su una popolazione, è stato possibile isolare, con relativa facilità, un numero cospicuo di mutanti diversi. Un processo evolutivo che, in condizioni naturali, si compie nell'arco di migliaia di anni, si può far avvenire nel volgere di una notte; in tal modo, se un batterio sensibile alla penicillina muta nel senso di una resistenza all'antibiotico, si può porre a incubare in un terreno di coltura, contenente un'opportuna concentrazione di penicillina, un numero sufficientemente ampio di microrganismi, così che facilmente, fra di essi, possa trovarsi un mutante resistente. Conseguentemente, dopo un'incubazione di un giorno, si potrà ottenere una coltura relativamente pura del mutante resistente alla penicillina. Sfortunatamente cospicue pressioni selettive di questo tipo possono essere presenti anche in natura, specialmente quando l'uomo altera l'ambiente, e possono svilupparsi mutanti non desiderabili: l'antibiotico-resistenza ne è un ovvio esempio.
Se Escherichia coli doveva essere impiegato come modello per ricerche di genetica o per altre indagini biologiche che richiedono la presenza di mutanti, era essenziale disporre di adatte modalità di ricombinazione genetica. Negli anni intorno alla metà di questo secolo si è avuta la scoperta di questi meccanismi.
Coniugazione. - La scoperta di un trasferimento quasi sessuale di materiale genetico venne compiuta da J. Lederberg e E. L. Tatum nel 1946. Essi osservarono che se si coltivavano insieme due ceppi di Escherichia coli con differenti esigenze colturali (mutanti auxotrofi), si verificava una ricombinazione dei determinanti genetici. Tale processo presuppone il contatto fisico tra i due ceppi, uno dotato del fattore della fertilità (F+) e l'altro privo di esso (F-). La coniugazione è legata al passaggio di DNA dal ceppo F+ a quello F-. Sono state compiute ricerche sperimentali per chiarire che cosa determini questa primitiva forma di sesso: si è visto che esiste un fattore della fertilità (fattore F+) che può essere trasferito da una cellula a un'altra così che ne risulta la formazione di nuovi ceppi F+. Il fattore F+ di per sé è una piccola molecola circolare di DNA a doppia elica che normalmente si moltiplica autonomamente nel citoplasma batterico al quale conferisce la caratteristica ‛maschile'; però esso può essere anche integrato nel cromosoma principale della cellula, dando così ceppi che presentano una frequenza più elevata di ricombinazione sessuale (High-frequency of recombination o ceppi Hfr). Si può immaginare che il processo in realtà si compia in tre fasi: a) si ha un contatto fisico (coniugazione) tra il ceppo F+ o Hfr e quello F- per l'adesione di superfici complementari; il fattore più importante ai fini della complementarità è il possesso di speciali peli sessuali localizzati sulla superficie della cellula maschile; la produzione di questi peli è regolata dal fattore F-; b) il DNA viene trasferito sotto forma di filamento lineare dal ceppo Hfr o F+ a quello F-. L'intimo processo di trasferimento è ancora oggetto d'ipotesi, ma è un processo ordinato. La coniugazione può essere interrotta con un'agitazione violenta delle cellule a diversi intervalli di tempo dall'associazione iniziale e si è osservato che i geni penetrano a intervalli di tempo determinati dalla loro distanza dal punto di penetrazione del cromosoma. Questa tecnica d'interruzione della coniugazione è risultata estremamente importante per stabilire la mappa del cromosoma di Escherichia coli; c) in seguito al trasferimento del cromosoma si forma una cellula temporaneamente e spesso parzialmente diploide; ha luogo il crossing-over dei geni e infine si ricostituisce la forma aploide.
È dubbio se la coniugazione sia importante nei Batteri in genere: essa è stata infatti dimostrata in un gruppo di microrganismi strettamente correlati con Escherichia coli, ma bisogna rilevare che sono state compiute poche indagini per stabilire la massima diffusione del processo.
Trasduzione. - Come diremo in un capitolo successivo esistono numerosi fagi che attaccano Escherichia coli: durante il processo di riproduzione del fago la sua testa contiene normalmente il genoma desossiribonucleico fagico, tuttavia una piccola percentuale di teste può accidentalmente comprendere un segmento del DNA dell'ospite, costituito da un numero variabile di geni, da trenta sino a centocinquanta. Quando un fago di questo tipo aggredisce una cellula di Escherichia coli, il segmento di DNA dell'ospite precedente penetra nel nuovo ospite e si dice che è stato trasdotto. Una volta all'interno della cellula può essere incorporato, secondo i normali processi di ricombinazione, nel cromosoma del nuovo ospite. Poiché un fago che opera la trasduzione può prelevare a caso un qualsiasi frammento del cromosoma dell'ospite, ne risulta che, grosso modo, può trasdurre qualsiasi gene con identica frequenza. Questo processo viene indicato come ‛trasduzione generalizzata', per distinguerlo dalla ‛trasduzione ristretta' di cui diremo successivamente (v. sotto, cap. 6). La trasduzione generalizzata sembra diffusa in natura e rispecchia quindi la notevole diffusione dei fagi; per conseguenza è probabilmente il più importante sistema naturale d'interscambio di geni tra Batteri. Essa è risultata altresì un processo di notevole importanza per il microbiologo interessato alla genetica.
Trasformazione. - Con una delle più importanti ricerche biologiche di questo secolo R. J. Dubos e O. T. Avery dimostrarono che nello pneumococco, agente eziologico della polmonite, si possono trasferire geni da una cellula all'altra mediante frammenti del DNA cromosomico purificato. Tale scoperta è stata la prima vera dimostrazione che il DNA è il depositario dell'informazione genetica. Il processo si compie nello pneumococco con una rapidità notevolmente maggiore che in altri batteri e un microrganismo come Escherichia coli può essere trasformato solo con difficoltà; tali differenze nel subire la trasformazione sembrano legate principalmente al problema della penetrazione di una grossa molecola, come il DNA, nel citoplasma e poi nel nucleo, poiché la parete cellulare e la membrana citoplasmatica si comportano normalmente come una vera e propria barriera contro un simile trasferimento. È pertanto difficile valutare l'importanza in natura della trasformazione.
Bisogna tener presente che sia la trasduzione, sia la trasformazione comportano il trasferimento di una parte soltanto del cromosoma batterico nella cellula ospite; ne deriva la formazione di un diploide parziale, prima che avvenga il crossing-over e la ricostituzione di un normale aploide. Perché avvenga la ricombinazione, la cellula donatrice e quella recettrice devono essere correlate abbastanza strettamente.
Le tecniche di ricombinazione, di cui abbiamo parlato, hanno consentito di stabilire la disposizione nel cromosoma di un notevole numero di geni di Escherichia coli: una recente rassegna della mappa cromosomica di questo batterio riporta più di seicentocinquanta loci, di ciascuno dei quali è stata stabilita la disposizione nella mappa. Come si può rilevare, alcune zone sono delucidate chiaramente, mentre altre non presentano determinanti noti; è interessante il fatto che la distribuzione dei diversi tipi di determinanti presenta un certo grado di simmetria e ciò induce a supporre che il cromosoma di Escherichia coli possa aver avuto origine da un processo di duplicazione avvenuto a un certo momento in passato. È chiaro che, con l'approfondirsi delle conoscenze sulla più intima struttura genetica, si potranno ottenere importanti informazioni sui processi evolutivi.
Bisogna tener conto di un altro fenomeno che è divenuto d'importanza rilevante come fattore additivo in grado di determinare il genotipo di Escherichia coli e di altri batteri: esso si ricollega con la presenza di DNA extracromosomico. Con frequenza sempre maggiore, recentemente, sono stati scoperti esempi della presenza di DNA sotto forma di plasmidi localizzati al di fuori del cromosoma principale. I plasmidi sono in genere frammenti circolari di DNA a doppia elica che si riproducono autonomamente all'interno della cellula; possiedono molte proprietà di un piccolo cromosoma e contengono informazioni genetiche dello stesso tipo, per il controllo della propria replicazione e per assicurare la segregazione di ciascuna copia in ogni cellula figlia al momento della divisione cellulare; si differenziano però da un cromosoma in quanto sono molto più piccoli (contengono cioè da 20 a 100 geni) e non sono essenziali per la normale vitalità della cellula. Talora un plasmidio può essere integrato nel cromosoma principale della cellula batterica e viene indicato con il termine di ‛episoma', tuttavia tale distinzione non è sempre facile a farsi e si tende sempre più a operare una restrizione, indicando con il termine di plasmidi tutte le forme di DNA extracromosomico. Abbiamo già menzionato un esempio di plasmidio, il fattore F che interviene nella coniugazione; un altro esempio è rappresentato da alcuni fattori di resistenza a farmaci che possono essere presenti nei plasmidi piuttosto che nel cromosoma principale: un unico plasmidio può codificare la resistenza a numerosi antibiotici di uso comune. Sfortunatamente i plasmidi possono essere trasferiti con notevole facilità da una cellula all'altra tramite un processo di adesione cellulare analogo alla coniugazione sessuale e possono determinare la rapida diffusione di un'antibiotico-resistenza in seno a una popolazione batterica. I fini di questo processo devono essere ancora in gran parte delucidati, in particolare per ciò che riguarda ceppi batterici relativamente poco correlati: essi sono infatti tema di ricerche sempre più numerose.
In rapporto con le modalità di trasferimento dei geni, di cui abbiamo detto in precedenza, si è giunti a ipotizzare un'estesa ‛ingegneria genetica' in grado di dare origine a batteri completamente nuovi: sebbene non esistano dubbi sul valore potenziale che avrebbe per l'industria lo sviluppo di batteri più efficienti, recentemente si è sviluppata una polemica molto intensa sul danno che può derivare da una ricerca di questo tipo, condotta indiscriminatamente, e sui principî morali che essa implica. Anche se è auspicabile che l'ingegneria genetica non possa mai essere applicata all'uomo, il quale del resto possiede molte barriere biologiche che i Batteri non hanno, e sebbene evidentemente esista il pericolo di una diffusione ancora maggiore della resistenza ai farmaci e di altre proprietà non desiderabili, come ad esempio i fattori di virulenza, forse il danno reale sarebbe esiguo se essa venisse applicata da ricercatori qualificati in campi circoscritti (v. genetica: Applicazioni della genetica).
3. I Procarioti
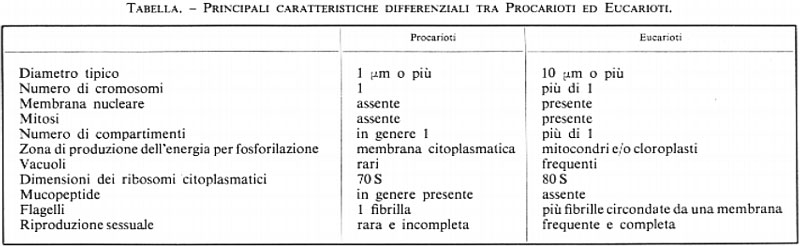
Stanier e van Neil (v., 1962) proposero che tutte le cellule venissero comprese in due gruppi ben definiti: il primo, di cui è prototipo Escherichia coli, venne detto dei Procarioti e il secondo, che è esemplificato da Saccharomyces cerevisiae (di cui parleremo in un capitolo successivo), degli Eucarioti. Le differenze tra questi due tipi di cellule sono riportate nella tabella e, sebbene alcune caratteristiche elencate da Stanier e van Neil siano state modificate, la distinzione fondamentale, sino a oggi, è stata confermata. Evidentemente è questa una delle più importanù acquisizioni della microbiologia: appartengono ai Procarioti i Batteri e le Alghe verdi-azzurre (Cianofite), mentre il tipo cellulare cucariotico si riscontra nelle altre Alghe, nei Funghi e nei Protozoi, tra i microorganismi, e in tutti gli altri organismi animali e vegetali.
a) Variazioni strutturali dei Procarioti
La cellula procariotica non presenta grandi variazioni della struttura, per lo meno negli organismi unicellulari che rappresentano la grande maggioranza dei Procarioti. La forma può essere varia e va da quella sferica a quella bastoncellare a quella a spirale, mentre le dimensioni oscillano da 0,2 μm a più di 10 μm. Nella cellula si possono osservare alcune variazioni strutturali, particolarmente per ciò che riguarda elementi che si trovano all'esterno della membrana citoplasmatica, quali la parete cellulare, le capsule, i flagelli e i peli; all'interno dell'elemento, le diversità sono rappresentate dalla presenza d'invaginazioni della membrana citoplasmatica, che può essere abbastanza complessa, e dalla presenza o dalla mancanza di granuli di deposito. Talora alcuni procarioti sono multicellulari, anche se in essi non si riscontra un vero differenziamento dei tessuti: per esempio alcuni batteri possono formare lunghe catene cellulari, come gli Attinomiceti, un gruppo che presenta somiglianze superficiali con i Funghi e che è più generalmente noto per la notevole capacità di produrre antibiotici. Nei Procarioti può riscontrarsi una forma semplice di differenziamento, con comparsa di cellule specializzate, processo che negli ultimi anni ha suscitato notevole interesse, nell'ipotesi che possa rappresentare un modello del differenziamento presente negli organismi superiori. L'esempio più elementare è costituito dalla comparsa di uno stadio di quiescenza, la spora. Nei Procarioti il processo di sporulazione si compie con il differenziamento di un'unica cellula in un'unica endospora, in risposta a condizioni ambientali sfavorevoli; quando queste divengono nuovamente favorevoli, la spora germina e dà origine a un'unica cellula vegetativa. Durante questo processo i geni vengono attivati o meno a seconda delle differenze fra i componenti della spora e della cellula vegetativa: per esempio, la spora contiene generalmente un composto, l'acido dipicolinico, che non si riscontra nella forma vegetativa; i geni strutturali preposti alla formazione degli enzimi che partecipano alla biosintesi dell'acido dipicolinico vengono attivati durante la sporulazione, mentre sono nuovamente inattivati nel corso della germinazione. D'altro lato, la maggior parte degli enzimi che intervengono nei normali processi di crescita sono repressi durante la sporulazione e derepressi durante la germinazione. I meccanismi che partecipano al controllo di questi processi e le modalità con cui sono determinati dalle condizioni ambientali possono riflettere i processi che hanno luogo nelle cellule più evolute. Un esempio assai più complicato di sistema primitivo che si differenzia è costituito dai Mixobatteri fruttificanti, un gruppo di batteri presente sul legno in decomposizione e sugli escrementi animali: sebbene essi sembrino di scarso interesse per il biologo generale, si è visto che, in risposta ad appropriate condizioni ambientali, le forme vegetative si riuniscono, producendo un corpo fruttificante complesso e multicellulare, all'interno del quale, successivamente, si formano cellule in stato di latenza o cisti. È questo un altro sistema che può essere preso a modello per lo studio del differenziamento.
b) La gamma dei processi fisiologici nei Procarioti
Abbiamo ricordato in precedenza che Procarioti ed Eucanoti sono formati dagli stessi componenti chimici fondamentali e che le modalità con cui essi sviluppano e trasferiscono l'energia, insieme ai principali processi catabolici (come la glicolisi e il ciclo degli acidi tricarbossilici) e ai più importanti meccanismi biosintetici degli amminoacidi dei nucleotidi e dei lipidi, presentano notevoli somiglianze anche se non sono identici. In altri termini l'unità biochimica si estende anche ai Procarioti; tuttavia, nell'ambito di questa generalizzazione, si riscontrano risposte notevolmente diverse nei confronti di ambienti e di ecosistemi particolari.
Trasferimento dell'energia. - Sebbene il trasferimento dell'energia tra le reazioni di scissione o cataboliche e i processi che richiedono energia, quali quelli biosintetici e di trasporto, si compia normalmente tramite legami fosforici altamente energetici utilizzando l'ATP, l'ADP, l'AMP, nei Procarioti rispetto agli Eucarioti è presente un più ampio spettro di meccanismi specifici. Scegliendo come esempio i processi di ossidazione per l'utilizzazione dei composti organici, si è visto che batteri come Pseudomonas sono in grado di ossidare più di cento sostanze diverse, ciascuna delle quali può essere l'unica fonte di carbonio e di energia necessaria per la crescita. Questo ampio spettro di attività cataboliche ha determinato lo sviluppo di una serie di enzimi ugualmente estesa e ciò, di per sé, ha contribuito a perfezionare i meccanismi di controllo dell'induzione che abbiamo ricordato in precedenza. Se si considerano globalmente i Procarioti, è possibile rilevare che essi possiedono la capacità di ossidare tutti i composti organici naturali insieme a molti altri artificiali, proprietà d'importanza fondamentale per il processo di mineralizzazione che è essenziale per il ciclo del carbonio in natura. Le ossidazioni avvengono tramite un sistema di trasporto di elettroni come negli Eucarioti, anche se i trasportatori in causa presentano notevoli differenze di complessità: alcuni batteri possiedono un sistema analogo a quello mitocondriale degli Eucarioti, altri invece hanno una serie ridotta di citocromi e di altri trasportatori di elettroni.
Un processo generale di produzione di energia ossidativa, caratteristico dei Procarioti, consiste nell'utilizzazione di composti inorganici ridotti come fonte di energia. Questi composti, attraverso un sistema di trasporto degli elettroni, vengono ossidati, liberando energia per fosforilazione ossidativa: così i solfuri e lo zolfo vengono ossidati a solfato, l'ammoniaca e i nitriti a nitrato, l'idrogeno ad acqua e il metano a CO2. Tali batteri, indicati con denominazioni diverse, autotrofi chemiosintetici o chemiolitotrofi, assumono il carbonio loro necessario sotto forma di CO2 e lo riducono a carbonio cellulare con gli stessi procedimenti presenti negli organismi fotosintetici.
Nei Procarioti l'ossidazione di un composto organico a CO2 non è necessariamente legata all'impiego dell'ossigeno come accettore finale di elettroni: può verificarsi un processo, detto respirazione anaerobica, in cui un composto inorganico ridotto funziona da donatore di elettroni; in altri termini le vie metaboliche sono identiche a quelle della respirazione aerobica e l'energia si forma per fosforilazione ossidativa con l'intervento di un sistema di trasporto di elettroni modificato. Ne deriva che vengono invertite le trasformazioni inorganiche effettuate dai batteri chemiolitotrofi: il solfato viene ridotto a solfuro, il nitrato ad azoto molecolare e il CO2 a metano. Queste interconversioni sono di notevole importanza in natura per i cicli degli elementi. Un altro e probabilmente più comune metodo di produzione anaerobica di energia, al buio, è rappresentato dalla fermentazione, processo con il quale il carbonio organico e la fonte di energia danno luogo a prodotti terminali in cui si ha una ristrutturazione degli atomi operata da una reazione esoergonica con produzione di ATP per fosforilazione a livello del substrato. Non si ha una netta ossidazione. La composizione chimica effettiva dei prodotti terminali varia notevolmente da composti che si riscontrano anche negli Eucarioti, come l'acido lattico, l'alcool etilico e il CO2, a una serie di acidi e di alcoli caratteristici dei Procarioti, come l'acido formico, l'acetico, il propionico, il butirrico, il succinico, oltre all'acetone e all'alcool butilico. Una simile varietà è stata sfruttata per classificare i Batteri, ma è anche servita all'industria fermentativa per la produzione commerciale delle sostanze che abbiamo citato.
I tre processi di scissione delle sostanze organiche sono riassunti schematicamente qui di seguito, considerando sempre il glucosio come substrato:
1 - Respirazione aerobica:
C6H12O6+6O2→6CO2+6H2O
2 - Respirazione anaerobica:
C6H12O6+3H2SO4→6CO2+3H2S+6H2O
3 - Fermentazione:
C6H12O6→2C2H5OH+2CO2.
Altri procarioti ottengono l'energia con un tipo di foto- sintesi nel corso della quale non si produce ossigeno e che, evolutivamente, è più primitiva di quella presente negli Eucarioti. Nei batteri fotosintetici il CO2 viene ridotto a materiale cellulare con l'ATP prodotto per fosforilazione fotosintetica, ma come ultimo donatore di elettroni per il processo di riduzione invece dell'acqua viene utilizzata o una forma ridotta di zolfo inorganico, come il solfuro d'idrogeno (acido solfidrico), o un composto organico. Van Neil e collaboratori hanno dimostrato che questa via fotosintetica è importante per chiarire i processi fondamentali della fotosintesi; d'altro canto, nelle Alghe verdi-azzurre l'ossigeno viene prodotto con sistemi non molto diversi da quelli delle Alghe eucariote e delle piante.
c) Lo sviluppo in ambienti proibitivi
Quando i biologi cominciarono a studiare le forme di vita presenti in ambienti proibitivi per temperatura, pH, pressione osmotica, tasso di radiazioni, ecc., risultò chiaro che man mano che l'ambiente diventava più ostile, una popolazione composita, comprendente tutti i principali gruppi di organismi viventi, veniva sostituita dai soli Procarioti. Questo fatto si può constatare facilmente nelle zone circostanti le fonti di acqua calda, nelle quali v'è un gradiente di temperatura fino alla sorgente. Un altro esempio è offerto dalle zone in cui il suolo ha un'elevata concentrazione salina, zone che possono spesso diventare rosa quando i sali vanno incontro a saturazione per la presenza di batteri alofili pigmentati: i microrganismi responsabili possono svilupparsi solo in presenza di elevate concentrazioni saline. Un ultimo esempio l'abbiamo già ricordato prima: Thiobacillus thiooxidans, batterio chemiolitotrofo che ossida lo zolfo, elabora acido solforico come prodotto terminale del suo metabolismo energetico e si sviluppa molto bene a un pH inferiore a 3.
Le ricerche su questi ecosistemi ‛limite' hanno permesso di ritenere che i Procarioti siano gli organismi maggiormente in grado di compiere lo sviluppo evolutivo necessario per tali colonizzazioni. Presumibilmente ciò è dovuto a diversi fattori che abbiamo già citato: in primo luogo i Procarioti presentano la caratteristica generale di adattarsi a condizioni ambientali diverse, in secondo luogo sono in grado di modificare rapidamente il proprio genotipo, infine la loro struttura elementare può ridurre notevolmente le modificazioni necessarie per la colonizzazione di questi ambienti proibitivi.
4. Un organismo modello, Saccharomyces cerevisiae
Tra i microrganismi eucarioti quello di cui conosciamo meglio struttura e fisiologia è un lievito, Saccharomyces cerevisiae: i motivi di questa più approfondita conoscenza sono facilmente comprensibili, poiché sin dai tempi più antichi Saccharomyces cerevisiae è stato impiegato per produrre bevande alcoliche con la fermentazione e per far lievitare il pane con la formazione di CO2. Tuttavia, rispetto a Escherichia coli, la nostra conoscenza della struttura di questa cellula è molto meno particolareggiata, in parte anche per la sua maggiore complessità. Per la somiglianza che esiste tra le cellule dei microrganismi eucarioti e quelle degli animali e delle piante, e poiché in questa Enciclopedia altri articoli trattano questo argomento, ci limiteremo a un'esposizione succinta, ponendo in risalto le differenze strutturali riscontrabili tra le cellule di Saccharomyces cerevisiae e quelle dei Procarioti.
La differenza di una tipica cellula di Saccharomyces cerevisiae rispetto a Escherichia coli consiste nelle dimensioni che, in genere, raggiungono il diametro di 5 μm; le dimensioni più cospicue si associano a una struttura interna a più compartimenti, ciascuno dei quali è delimitato da una membrana semipermeabile. Intorno al citoplasma c'è una membrana plasmatica che assolve a una funzione simile a quella della membrana citoplasmatica di un procariote, salvo che non è la sede di produzione dell'energia, né elabora un punto di attacco per il cromosoma; presenta altresì una differenza fondamentale dal punto di vista chimico, nel senso che partecipano alla sua composizione gli steroli, molto rari nei Procarioti. Membrane unitarie delimitano anche vacuoli, mitocondri e nucleo; la membrana endoplasmatica è costituita da altre strutture membranose. Lo stesso nucleo, ove il DNA è segregato nei cromosomi, ha una struttura molto più complessa: si divide per mitosi e meiosi, processi che assicurano una serie completa di cromosomi a ciascuna cellula figlia e che non sono necessari al nucleo del procariote che è costituito da un unico cromosoma. Nelle cellule degli Eucarioti all'interno del nucleo è in genere presente un corpicciolo contenente RNA, detto nucleolo, struttura specializzata collegata con la sintesi ribosomale dell'RNA, compito questo che nei Procarioti viene assolto dal cromosoma principale. I cromosomi degli Eucarioti si differenziano anche perché possono esistere sotto una duplice forma, estesa o contratta, e perché contengono proteine basiche dette istoni (v. cellula: Fisiologia della cellula; v. genetica: Citogenetica).
La genetica del lievito è un campo relativamente inesplorato se confrontata a quella di Escherichia coli, anche se sono disponibili tecniche di ricombinazione genetica per appaiamento. Il DNA contenuto in un nucleo aploide di Saccharomyces cerevisiae è circa quattro volte quello del nucleo di Escherichia coli ed è suddiviso in diciassette cromosomi relativamente piccoli: tale maggior quantità fa supporre una complessità e un numero di loci considerevolmente maggiori, ma è difficile individuare la causa di una differenza così cospicua. Attualmente solo con estrema lentezza si sta stabilendo la mappa cromosomica di Saccharomyces cerevisiae: si conoscono meno di cento loci, a differenza dei più di seicento noti di Escherichia coli. Tra i microrganismi eucarioti Neurospora crassa è stato quello più comunemente utilizzato come modello per indagini genetiche e venne impiegato, negli anni intorno al 1940, per stabilire la correlazione ‛un gene - un enzima'.
I ribosomi degli Eucarioti hanno dimensioni leggermente superiori (costante di sedimentazione 80S) rispetto a quelli dei Procarioti, sebbene, per il resto, esplichino funzioni del tutto simili; esistono tuttavia alcune differenze che tornano a vantaggio dell'uomo, in quanto nel ribosoma dei Procanoti si trovano componenti caratteristici che lo rendono suscettibile all'azione di alcuni antibiotici comunemente impiegati in terapia.
Nel citoplasma si può riscontrare una varietà di formazioni strutturali molto più ampia che nei Procarioti: le più note sono i mitocondri, ove sono localizzati i sistemi enzimatici deputati alla produzione di energia che invece nei Procarioti si riscontrano a livello della membrana citoplasmatica. Negli Eucarioti fotosintetici i corrispondenti sistemi di produzione dell'energia si trovano nei cloroplasti membranosi. Il reticolo endoplasmatico rappresenta il sito di attacco dei ribosomi e costituisce un sistema di canali di comunicazione che in alcune zone può essere organizzato a formare corpi di Golgi. Infine sono presenti granuli di deposito costituiti da glicogeno o da lipidi neutri.
Esternamente alla membrana plasmatica la cellula di lievito è circondata da una parete contenente diversi polisaccaridi che le conferiscono resistenza alle sollecitazioni meccaniche: i componenti principali sono un glucano (un poliglucosio congiunto da legami 1-β→3 e 1-β→6), un mannano (un polimannosio) e una proteina. La moltiplicazione avviene per gemmazione e perciò la parete può presentare una serie di cicatrici prodotte dalle gemme, zone a composizione chimica diversa rispetto al resto della parete in quanto contengono il polisaccaride chitina. Poiché la superficie della parete è limitata e dato che le cicatrici dovute alla gemmazione non possono essere utilizzate per formare altre gemme, si verifica una specie di ‛invecchiamento': l'età è determinata dal numero delle cicatrici. Al contrario i batteri che si riproducono per scissione binaria non presentano un processo del genere e l'età della cellula può essere espressa solo in correlazione con le precedenti divisioni cellulari e quindi in rapporto al ciclo cellulare. Del resto è dubbio anche che l'‛invecchiamento' dei lieviti sia realmente analogo a quello che si riscontra negli organismi superiori: infatti si può dimostrare che l'intero processo d'invecchiamento negli animali e nelle piante adempie a una funzione utile ed è stato selezionato in senso evolutivo, mentre in un organismo unicellulare che si riproduce per scissione binaria o per gemmazione non si riscontra una simile finalità. È possibile trovare un modello per il processo d'invecchiamento tra i microrganismi?
5.I microrganismi eucarioti
Se tra i microrganismi eucarioti si comprendono le Alghe, i Funghi e i Protozoi, le loro dimensioni possono variare notevolmente, oscillando da quelle di organismi unicellulari poco più grandi di un batterio a quelle delle gigantesche alghe marine. Tentare di riassumere le nostre conoscenze su questo argomento in uno spazio ristretto sarebbe assurdo, anche se in pratica l'incremento delle nostre acquisizioni su questi organismi non ha seguito l'andamento esponenziale che hanno fatto registrare negli ultimi anni quelle sui Procarioti. Uno dei motivi che hanno fatto sì che i microrganismi eucarioti siano stati relativamente trascurati è sicuramente la loro minore importanza per la medicina, per lo meno per ciò che riguarda il numero di condizioni morbose da loro provocate, anche se i Protozoi in particolare sono responsabili della diffusione di rovina e miseria nelle zone tropicali del globo. Il fatto che gli Eucarioti non siano stati diffusamente impiegati come modelli al pari dei Procarioti costituisce un altro di tali motivi, solo in parte attribuibile alla loro maggiore complessità; a esasperarlo ha infatti concorso la progressiva tendenza a escluderli dai propri campi di ricerca mostrata dalla biologia, che si è focalizzata sui Batteri e sui Virus, e dalla botanica e dalla zoologia, che si sono decisamente orientate sulle piante e sugli animali più evoluti.
Se si analizza la struttura delle singole cellule degli Eucarioti è possibile osservare una notevole serie di differenze, nell'ambito dell'architettura fondamentale che abbiamo descritto nel capitolo precedente. Consideriamo gli strati superficiali. Nelle Alghe la struttura fondamentale è mantenuta da microfibrille formate dall'intreccio di lunghe molecole di cellulosa associate ad altri omopolisaccaridi, come mannani, xilani (polixilosi) e pectine (acidi poliuronici); in alcune di esse, tuttavia, le pareti possono essere plasmate dalla silice o dal carbonato di calcio in forme strane e leggiadre; nei Funghi le pareti presentano una struttura simile a quella dei lieviti. Nei Protozoi i costituenti di queste strutture sono condizionati dal fatto che l'assorbimento delle sostanze nutritizie si compie prevalentemente con l'ingestione di particelle solide e quindi le cellule spesso non possiedono pareti ben definite, o tutt'al più ne presentano una flessibile. Tuttavia alcuni protozoi possono avere pareti complesse, composte da cellulosa, carbonato di calcio, silice o solfato di stronzio, con pori sufficientemente ampi da consentire il passaggio di particelle di cibo.
I microrganismi eucarioti presentano numerose modalità di locomozione: quelli sprovvisti di parete sono in grado di compiere movimenti ameboidi, ma più comunemente il movimento avviene con l'ausilio di flagelli o di ciglia, strutture che, se presenti, hanno una complessità notevolmente superiore a quella dei Procarioti, poiché sono formate da una guaina che avvolge numerose fibrille, ciascuna delle quali ha le dimensioni del flagello dei Procarioti. Anche le modalità di comportamento degli Eucarioti sono relativamente più complesse: talora presentano movimenti di avvicinamento o di allontanamento rispetto alla luce, al calore o a talune sostanze chimiche.
In un eucariote unicellulare esiste una chiara tendenza alla specializzazione di alcune zone intracellulari per particolari funzioni, processo che raggiunge la massima espressione in alcuni protozoi ciliati, come il paramecio. Questo organismo possiede un complesso apparato ciliare che necessita di un fine sistema di coordinazione e di controllo sia per la locomozione sia per l'assunzione di cibo che si compie attraverso una particolare zona incavata della parete; i vacuoli che si formano e che contengono il cibo vengono quindi trasportati, attraverso il citoplasma, in un'altra area specializzata della parete, attraverso la quale sono espulsi all'esterno. Nella parete sono localizzate anche strutture specializzate - le tricocisti - che servono per l'immobilizzazione della preda o dei predatori. Nel citoplasma si trovano vacuoli contrattili deputati alla regolazione della pressione osmotica, due tipi di nucleo - il macro e il micronucleo e una serie di altre strutture membranose e granulari. I Ciliati sembra che rappresentino quindi l'ultimo termine di una catena evolutiva per ciò che riguarda la specializzazione intracellulare o il differenziamento e, più di ogni altro prganismo, pongono un problema fondamentale in biologia cellulare: che cosa determina la distribuzione delle molecole nella cellula? È chiaro che le proteine elaborate nei ribosomi devono possedere siti per il riconoscimento di particolari strutture cellulari, ad esempio le permeasi per la membrana, le DNA-polimerasi per il nucleo, le depolimerasi idrolitiche per i liposomi, le proteine flagellari per i flagelli, e così via. È evidente che questo è un campo di ricerca che diverrà molto importante nell'ultimo quarto di questo secolo, dato che è già disponibile la maggior parte delle tecniche che esso richiede; l'organismo di scelta potrà essere un microrganismo, ma non necessariamente un ciliato che presenta il fenomeno in forma estrema.
Il modo più diffuso negli organismi viventi per conseguire una specializzazione ulteriore è il differenziamento di una serie di tipi cellulari in un organismo multicellulare; nei microrganismi eucarioti si può seguire l'attuarsi di questo processo in misura notevolmente superiore che nei sistemi primitivi dei Procarioti. Questo differenziamento è spesso diretto verso l'evoluzione di complessi cicli biologici derivati dall'adozione di nicchie ecologiche molto delimitate: esempi di tale differenziamento si osservano con estrema facilità nei Funghi, ma l'organismo che negli ultimi anni sembra aver maggiormente attratto l'attenzione fa parte di un gruppo con molte caratteristiche intermedie tra i Funghi e i Protozoi, quello delle muffe mucose (slime moulds) cellulari o Mixomiceti. Si tratta di eucarioti non fotosintetici che assomigliano ai Protozoi, in quanto presentano una forma vegetativa ameboide, e ai Funghi nei loro stadi di sporificazione. Le forme ameboidi vegetative si nutrono di materiale particolato e, quando le sostanze nutritive sono terminate, si aggregano in gruppi pseudoplasmodiali, processo determinato dalla secrezione di una sostanza similormonale, detta acrosina, che per chemiotassi fa accostare le cellule. Lo pseudoplasmodio si muove verso la luce come una singola unità circondata da una guaina mucoide. Si può avere un differenziamento con produzione di corpi fruttiferi, processo che implica l'elaborazione di una guaina di cellulosa da parte delle cellule superficiali; altre cellule superano questa guaina e all'estremità formano spore. Bisogna osservare che questo processo, anche se notevolmente più sofisticato, è simile alla formazione dei corpi fruttiferi nei Mixobatteri ed è un esempio di evoluzione convergente. Entrambi i suddetti gruppi possono essere scelti come sistemi modello per l'analisi dei principi del differenziamento.
6. I Virus; sviluppo della virologia
Lo studio dei Virus o virologia è effettivamente una scienza del XX secolo; prima dell'avvento della microscopia elettronica l'esistenza dei Virus poteva essere solo ipotizzata, poiché le loro dimensioni sono inferiori al potere risolutivo del microscopio ottico. Comunque, nei primi decenni di questo secolo, era apparsa evidente l'esistenza di un gruppo di parassiti obbligati capaci di autoriprodursi e di aumentare il proprio potere infettivo con il passaggio in ospiti infettati: tali agenti dovevano essere in grado di passare attraverso filtri con pori di diametro inferiore a quello necessario per arrestare gli organismi cellulari (circa 0,2 μm). Quindi prima della microscopia elettronica il termine ‛virus' era sinonimo di ‛agente infettivo filtrabile' e si riteneva possibile ottenere virus capaci di aggredire una notevole serie di organismi, dai Batteri da un lato, agli animali più evoluti e alle piante dal lato opposto. Fu possibile valutare il potere infettivo di un particolare virus e in molti casi si è successivamente dimostrata la corrispondenza fra l'unità infettiva e una particella virale morfologica. La determinazione del potere infettivo portò alla concentrazione e alla purificazione dei Virus, processo che consentì a W. M. Stanley, nel 1935, di dare la sensazionale comunicazione che il virus del mosaico del tabacco poteva essere ottenuto in forma cristallina. In effetti i metodi da lui adottati erano essenzialmente quelli perfezionati dai biochimici per la purificazione e la cristallizzazione delle proteine ed egli riteneva che il virus cristallizzato fosse formato solo da due componenti: proteina e RNA. È difficile rendersi conto oggi della profonda sensazione prodotta sui biologi dalla scoperta di un organismo, apparentemente provvisto di vita, che poteva essere trattato come un polimero complesso di elevato peso molecolare e che era analogo, per dimensioni e complessità, a molte sostanze chimiche presenti nelle cellule viventi. Una simile entità si può dire vivente? Ciò solleva il problema più generale di che cos'è un organismo vivente: è chiaro che in senso scientifico ogni definizione deve essere solo applicativa e in questo senso va intesa anche quella che stabilisce se una particolare cellula microbica è vivente o meno. In questo caso l'unica definizione adatta dal punto di vista applicativo dev'essere basata sulla capacità di riprodursi: un organismo vive se cresce e si divide in due nuovi organismi. Allo stesso modo qualsiasi definizione di organismo vivente deve comprendere i Virus, che possiedono la maggior parte degli attributi di cui un biologo deve necessariamente tener conto in una siffatta definizione. Il problema sta nel differenziamento delle strutture cellulari che si autoriproducono, come i mitocondri, ed è ovvio che ogni definizione racchiude in sé elementi di artificiosità.
Con lo sviluppo della microscopia elettronica e delle tecniche di colorazione negativa è divenuto possibile studiare la struttura delle particelle virali e ben presto è stata delineata un'immagine genetica, morfologicamente uniforme, consistente in una porzione centrale formata da un acido nucleico, DNA o RNA, circondata da un rivestimento proteico, detto capside, a sua volta costituito da singole unità proteiche che si ripetono, dette capsomeri. Nell'ambito di questa struttura generale, denominata nucleocapside, si riscontra una variabilità di forme e di dimensioni, che possono andare da quelle del batteriofago f2 di Escherichia coli, che è sferico e ha un diametro di 20 nm, a quelle di batteriofagi complessi, come il T2 di Escherichia coli, descritto nel paragrafo seguente, e a quelle dei vaccini a forma di mattone delle dimensioni di 250×300 nm. In alcuni virus il nucleocapside può essere circondato da un rivestimento fornito dalla cellula ospite. (V. anche virus).
a) Un organismo modello, fagi T2 e T4 di Escherichia coli
È stato nel secondo decennio di questo secolo che F. W. Twort e F. H. d'Herelle indipendentemente diedero una descrizione di infezioni virali dei Batteri: essi dimostrarono che cellule batteriche possono essere infettate e distrutte da agenti filtrabili. Questi virus batterici furono chiamati batteriofagi o, più semplicemente, fagi. Sebbene la maggior parte dei fagi presentino una struttura più complessa dei virus degli animali o delle piante, sono stati studiati profondamente, dato l'impiego degli organismi da loro infettati come sistemi modello per le ragioni che abbiamo già detto. Inevitabilmente il maggior numero di notizie sui meccanismi di riproduzione fagica e sull'influenza dell'ospite si è ottenuto dai virus che infettano Escherichia coli: in particolare un gruppo di fagi detto T (e specialmente i fagi T-pari, T2 e T4) è stato impiegato come modello insieme a pochi altri fagi di Escherichia coli. Da molti punti di vista i fagi T2 e T4 possono essere considerati estremamente simili, se non identici, e le notizie che esporremo possono essere riferite a ciascuno di essi.
Struttura. - Nella struttura del virus si possono osservare due parti principali, una testa e una coda. La testa contiene un'unica molecola circolare di DNA a doppia elica, o cromosoma, del peso molecolare di 1,3×108 dalton. Essa è circondata da un capside proteico a struttura icosaedrica di circa 125 nm di lunghezza e 81 nm di larghezza (un icosaedro è un poliedro regolare con 20 facce triangolari e 12 vertici): più precisamente ciascuna metà della testa è costituita da mezzo icosaedro e le due metà sono collegate da un breve prisma esagonale. Il capside è formato da singole subunità proteiche o capsomeri, la cui struttura ripetitiva determina la simmetria cristallina della testa.
La coda del fago è una struttura complessa di circa 95 nm di lunghezza: il suo componente più interno è un tubulo formato da subunità proteiche a disposizione elicoidale e rivestito da una guaina, costituita da altre subunità, che si estende da un collare a una placca basale e ha la capacità di contrarsi sino a meno della metà della sua estensione. Anche il collare e la placca basale sono composti da altre subunità proteiche e sulla placca vi sono piccoli perni ai quali sono attaccate sei lunghe e sottili fibre caudali.
In definitiva quindi un virus è costituito da un'unica molecola di DNA e da una serie di unità strutturali proteiche, probabilmente venti o trenta, disposte a formare le strutture proprie della testa o della coda.
Moltiplicazione. - La semplicità della struttura virale fa supporre un processo di moltiplicazione anomalo, ipotesi confermata dall'importante rilievo che una popolazione di un dato virus, se analizzata al microscopio elettronico, risulta costituita da particelle tutte delle stesse dimensioni; come può verificarsi un qualsiasi processo di riproduzione, con meccanismi analoghi a quelli presenti nelle forme cellulari di vita, senza una crescita del virus? La risposta a questa domanda si è ottenuta con lo studio degli eventi che si verificano in una cellula infettata e che si possono distinguere in tre fasi: adsorbimento, moltiplicazione, liberazione.
Inizialmente la particella fagica si adsorbe a una cellula di Escherichia coli per mezzo di un processo specifico di adesione tra la placca basale e le fibrille del fago e il lipopolisaccaride della parete della cellula batterica: il grado di specificità di questo processo è pari a quello che si ha nell'interazione di un anticorpo con l'antigene corrispondente. Immediatamente dopo l'adsorbimento si verifica un fatto sorprendente, la scomparsa delle particelle fagiche per la maggior parte del ciclo compreso tra l'infezione e la lisi. Cosa avviene in questo periodo di latenza? La soluzione venne fornita da uno degli esperimenti più importanti per lo sviluppo della biologia molecolare: A. D. Hershey e M. Chase nel 1952 marcarono le proteine del fago infettante con 35S e il DNA con 32P e seguirono quindi il comportamento dei traccianti. Risultò che mentre il DNA penetrava nel citoplasma batterico, la maggior parte del materiale proteico rimaneva invece sulla superficie della parete cellulare batterica; ciò fu confermato con la microscopia elettronica, che dimostrò che dopo l'infezione la testa vuota e la coda del fago rimanevano sulla superficie batterica. La deduzione obbligata fu che la successiva moltiplicazione del virus dipendeva esclusivamente dalla presenza del DNA virale nell'ospite: ciò fu confermato dall'osservazione che, se la permeabilità della parete batterica viene aumentata con un trattamento appropriato, il batterio può essere infettato con il DNA fagico purificato, esattamente con lo stesso risultato che si avrebbe se a infettarlo fosse il virus in toto. Divenne chiaro altresì che la funzione delle componenti proteiche del virus consiste nel proteggere l'acido nucleico durante il suo passaggio da una cellula all'altra e nel facilitare una specificità di penetrazione in un citoplasma ospite.
Dopo l'adsorbimento della coda del fago si ha la contrazione della sua guaina che fa penetrare la parte centrale della coda, attraverso la parete cellulare e la membrana citoplasmatica, nel citoplasma dell'ospite. Questo processo è seguito dal passaggio del DNA virale nell'ospite, attraverso il tubulo cavo centrale della coda: il meccanismo è grosso modo analogo a quello di una siringa ipodermica, salvo che non vi è contrazione della testa del fago; quindi il DNA fagico penetra nell'ospite con modalità ancora oscure.
La penetrazione del DNA fagico nell'ospite determina in questo una pressoché immediata sospensione della biosintesi di DNA, RNA e proteine: la sola biosintesi de novo di questi polimeri è codificata dal genoma fagico. Dopo un periodo di latenza, riprende la sintesi di DNA, ma completamente limitata alla produzione del DNA virale, come può facilmente essere dimostrato dall'osservazione che il DNA del fago T2 contiene come unica base la 5-idrossimetilcitosina. Dopo l'infezione viene elaborata solo una piccola quantità di RNA e sembra che si tratti esclusivamente di mRNA fagico: esso trascrive l'informazione per la sintesi delle proteine specifiche del virus, che vengono prodotte secondo una sequenza temporale ordinata. Le prime sono enzimi che partecipano alla produzione del DNA fagico, come ad esempio quelli che regolano la sintesi della 5-idrossimetilcitosina; successivamente vengono elaborate le proteine strutturali necessarie per la maturazione delle nuove particelle fagiche, insieme ai lisozimi fagici che partecipano all'eventuale lisi della cellula ospite. Vi è quindi un completo viraggio, dalla biosintesi dei polimeri dell'ospite a quella dei polimeri del virus, utilizzando tuttavia sempre lo stesso apparato biosintetico, cioè i ribosomi e gli enzimi dell'ospite presenti prima dell'infezione. Tale viraggio è fondamentale per il parassitismo virale che sostanzialmente è una sorta di parassitismo genetico in cui il genoma del virus si sostituisce a quello dell'ospite. In conseguenza di tutti questi processi, nel citoplasma dell'ospite si accumulano tutti i costituenti delle particelle mature del virus. La prima tappa del ‛montaggio' dei nuovi virus consiste nella condensazione del DNA fagico in elementi strutturali con la forma di teste fagiche, processo indotto da una speciale proteina ‛condensante'. Tale condensazione è seguita dalla cristallizzazione delle subunità proteiche della testa intorno al DNA, così da formare teste mature; la coda e la placca basale vengono poi elaborate per condensazione di appropriate subunità proteiche e, infine, le code e le teste si uniscono. Lo stadio finale sembra che consista nella sintesi e nel montaggio delle fibrille della coda. Ciascuno stadio dell'aggregazione di un componente strutturale dipende dal completamento del precedente. Quando l'elaborazione di nuove particelle fagiche è completata, si ha la lisi della cellula ospite, determinata in parte dall'azione di lisine indotte dal fago: ciò porta alla liberazione di alcune centinaia di particelle fagiche che possono infettare altre cellule ospiti. Poiché l'intero processo di moltiplicazione dura solamente dieci o venti minuti, il potenziale riproduttivo è evidentemente immenso e limitato esclusivamente dalla disponibilità di un adeguato numero di cellule ospiti. Utilizzando mutanti per i vari stadi che abbiamo ricordato è già stata puntualizzata dettagliatamente la mappa genetica del fago T4 e ne sono stati identificati da 90 a 100 geni.
La sequenza di eventi che abbiamo descritto, a partire dall'infezione operata da una particella virale, è il ciclo litico provocato da un fago virulento. A opera di un fago temperato può aversi una sequenza alternativa rispetto alla precedente, detta ‛lisogenia'; un altro fago di Escherichia coli, il fago lambda, è stato il microrganismo di scelta per l'analisi di questo processo. Il DNA virale penetra nell'ospite con le modalità che abbiamo già descritto ma, invece di causare la morte cellulare, è incorporato come un componente del genoma dell'ospite ed è detto ‛profago'. Si replica quindi come se fosse una parte del normale genoma batterico e solo occasionalmente si scatena spontaneamente un ciclo di sviluppo litico, la cui incidenza può essere incrementata da alcuni agenti induttori. La presenza di un profago, quale componente del genoma batterico, non lascia inalterato l'ospite: innanzi tutto il batterio lisogeno diviene immune dall'aggressione di altri fagi, ma, dato ancor più importante, si possono verificare alterazioni specifiche del genotipo batterico, dette conversioni lisogeniche, che non presentano rapporti evidenti con il ciclo biologico del fago. Per esempio possono essere modificati gli antigeni superficiali del batterio oppure può essere alterata la produzione di tossine. Quando si comprese che in natura la maggior parte dei batteri sono lisogeni, risultò chiara l'importanza del fenomeno. La lisogenia è responsabile anche di un utile processo genetico: la trasduzione restrittiva. In questa un batterio lisogeno viene attivato a compiere un ciclo litico e, in una piccola percentuale di cellule, quando il profago è indotto, viene escisso in modo non netto dal cromosoma così che si ha la rimozione di una porzione del DNA dell'ospite prossima a esso. Il fago difettivo può quindi aggredire un'altra cellula, ma non è in grado di duplicarsi normalmente e di formare altri fagi maturi; viene invece incorporato come profago e ne deriva che al cromosoma viene aggiunto un frammento di DNA batterico. Poiché un fago temperato ha un sito d'inserzione specifico e può prelevare solo DNA batterico prossimo a esso, il risultato viene detto trasduzione ristretta. Il fago lambda è sempre inserito tra i geni del catabolismo del galattosio (gal) e della sintesi della biotina (bio) e quindi trasduce loci prossimi a questi siti.
b) Altri Virus
Abbiamo detto che tutti i Virus presentano la stessa struttura nucleocapsidica generale. Possono contenere DNA o RNA, ma mai entrambi e l'acido nucleico può essere sia a singola sia a doppia elica. La quantità di acido nucleico può variare da 2×106 dalton a 160×106 dalton. La capacità di codificare è estremamente varia, oscillando da tre proteine a parecchie centinaia nei virus di dimensioni maggiori. Alcuni virus presentano un capside a struttura icosaedrica e il numero dei capsomeri è determinato dalle leggi della geometria: il più semplice comprende dodici capsomeri proteici identici che costituiscono i dodici vertici (per es. il fago ϕX di Escherichia coli); il numero successivo possibile è trentadue (dodici vertici e venti facce triangolari), l'altro è settantadue (dodici vertici e sessanta facce triangolari) e così via sino a virus che possiedono più di ottocentododici capsomeri (dodici vertici e ottocento facce triangolari). È interessante osservare che gli stessi principi sono sfruttati nella costruzione di cupole geodesiche da R. Buckminster Fuller e collaboratori: è possibile ottenere una struttura molto resistente associando fra loro subunità assai semplici e facilmente collegabili. Un altro tipo di disposizione dei capsomeri è quello elicoidale, in cui le singole molecole proteiche sono disposte a spirale, così che l'intera particella virale si presenta come un cilindro cavo che protegge l'acido nucleico, il quale, in genere, è localizzato in un solco elicoidale all'interno del capside. Infine sia il virus a struttura elicoidale, sia quello a icosaedro possono avere un rivestimento membranoso lasso: molti virus animali (così si chiamano i virus parassiti di cellule animali, per distinguerli dai virus vegetali, parassiti di cellule di piante) sono di questo tipo e si è visto che il rivestimento è in parte formato da componenti dell'ospite derivati dalla membrana plasmatica della cellula infettata.
Per ciò che riguarda la moltiplicazione degli altri virus, il meccanismo di base è molto simile. Esistono differenze circa le modalità di penetrazione dell'acido nucleico virale, le quali in genere sono più semplici di quelle del fago T2. Per quanto concerne i virus delle piante, le particelle generalmente penetrano attraverso una lesione meccanica dell'ospite o con la puntura di un insetto, mentre per i virus degli animali la fagocitosi è la normale modalità di penetrazione.
Un problema particolare si pone per la moltiplicazione dell'RNA dei virus a RNA: essi necessitano della codificazione di una polimerasi ribonucleica RNA-dipendente, così che l'RNA virale agisce sia come se fosse autoriproducentesi al pari del DNA, sia come messaggero per la sintesi proteica; se un virus a RNA provoca la lisogenia è richiesta anche una transcriptasi inversa dall'RNA al DNA (v. neoplasie: Oncologia sperimentale; v. sangue: Leucemie).
Infine sono numerosi i meccanismi di liberazione del virus dall'ospite: per esempio nei virus animali il nucleocapside viene normalmente avvolto in una membrana plasmatica alterata che costituisce per esso un involucro quando esce dalla cellula.
Concludendo dobbiamo sottolineare che la struttura e la riproduzione dei Virus sono ben diverse da quelle delle cellule eurocariotiche o procariotiche. La composizione di tali microrganismi è molto più semplice: in essi non esistono membrane semipermeabili, citoplasma o ribosomi, non hanno un metabolismo intermedio, né possiedono sistemi per la produzione di energia, e i soli enzimi che hanno sono in funzione della loro penetrazione o fuoruscita dalla cellula ospite. Il Virus dipende pressoché completamente dai processi metabolici dell'ospite e, per conseguenza, può essere considerato la forma più perfetta di parassitismo. Del resto, contrariamente a quanto si riteneva in passato, non sembra che esistano forme di vita intermedie che possano rappresentare stadi di transizione evolutiva tra quella virale e quella cellulare.
7. Classificazione ed evoluzione dei microrganismi
La classificazione dei microrganismi può essere effettuata secondo due diversi criteri. Il primo è un mezzo per riconoscere e identificare l'organismo ai fini pratici e di conseguenza le prove utilizzate devono essere semplici da effettuare e facili da riprodurre nei diversi laboratori di tutto il mondo. Taluni caratteri sono inevitabilmente considerati più importanti perché più utili in pratica: per esempio la capacità di provocare un particolare quadro morboso con determinati sintomi è un carattere importante per il microbiologo medico e pertanto gli organismi che provocano la stessa affezione vengono riuniti nello stesso gruppo, sebbene possano presentare altre caratteristiche che li differenziano gli uni dagli altri; allo stesso modo l'intero procedimento di identificazione è basato sull'utilizzazione di caratteristiche che portano a questo risultato. Un secondo criterio di classificazione può consistere nel tentativo di definire i raggruppamenti naturali dei microrganismi, rispecchiando quindi le loro interrelazioni evolutive: in altri termini può venire inteso come metodo di sviluppo di una tassonomia filogenetica.
In passato il primo di questi orientamenti è ampiamente prevalso, specie per ciò che riguarda i Batteri: la nomenclatura era basata sul sistema binomiale di C. Linneo e i microrganismi venivano indicati con il nome del genere e quello della specie. Inizialmente i caratteri utilizzati riguardavano la morfologia grossolana e le caratteristiche di colorazione, ma, con il passare degli anni, sono stati sfruttati con sempre maggiore frequenza test biochimici. Gli organismi vennero riuniti in famiglie, ordini e così via, spesso in base a criteri di scarso valore. Il Bergey's manual of determinative bacteriology stabilisce dal punto di vista batteriologico i criteri attuali per la classificazione e l'identificazione e ogni nuova edizione di questo testo fondamentale è causa di sconforto: inevitabilmente i metodi utilizzati per caratterizzare specie e generi variano ampiamente e i tentativi di uniformarli, in mancanza di un accordo sui principi generali, provocano confusione.
Molti sono stati i tentativi di elaborare un sistema di tassonomia più significativo e ancor più attinente ai principi filogenetici: nell'ultimo decennio è divenuto sempre più popolare quello della tassonomia numerica, basato sul principio di M. Adanson introdotto in biologia, per le piante, nel XVIII secolo. In esso vengono impiegati quanti più test (caratteri) possibili, a ciascuno dei quali viene attribuita un'uguale importanza; il grado di correlazione si valuta quindi analizzando mediante un elaboratore il numero dei caratteri comuni che i microrganismi presentano. In considerazione dell'adattabilità dei microrganismi è sorprendente che tale metodo abbia confermato i raggruppamenti naturali, sebbene questi rispecchino semplicemente l'appartenenza comune a una particolare nicchia ecologica. Così, benché la classificazione numerica possa essere un importante contributo alla tassonomia, è in definitiva probabile che gli indizi più importanti delle correlazioni evolutive tra i microrganismi vengano dalle ricerche di biologia molecolare: per esempio il DNA può essere analizzato a tre livelli, dimensioni, composizione in basi (rapporto G/C) e sequenza nucleotidica. È chiaro che il terzo metodo è l'unico che potrà fornire valide indicazioni correlative. In mancanza di tecniche adatte per stabilire la sequenza nucleotidica del DNA, si è prestata l'attenzione maggiore al metodo indiretto del grado di ibridazione del DNA di due organismi diversi, sebbene le opinioni sulla validità di tali indagini non siano concordi. In alcuni casi informazioni più utili possono essere ottenute mediante lo studio delle sequenze enzimatiche.
Negli ultimi anni si è registrato un incremento dell'interesse per l'evoluzione dei microrganismi, provocato in parte dalle ipotesi sull'origine della vita. Si ritiene comunemente che la cellula procariotica sia stata un precursore di quella eucariotica, ma la generale accettazione dei concetti esposti da A. I. Oparin e J. B. S. Haldane negli anni intorno al 1930, che cioè la vita sul nostro pianeta sia stata preceduta da circa duemila milioni di anni di sintesi chimiche di composti organici, ha portato a teorie sul probabile orientamento in senso procariotico dell'evoluzione. È possibile che nell'ambiente anaerobico presente prima della comparsa della vita si siano succedute le seguenti tappe: 1) circa 2-3×109 anni or sono comparve un anaerobio eterotrofo in grado di soddisfare le sue complesse necessità nutritizie con il magma primordiale e di ricavare l'energia necessaria dalla fosforilazione a livello del substrato; 2) non appena il tasso ambientale di sostanze nutritive organiche venne abbassato dagli eterotrofi, si svilupparono nuovi processi biosintetici che portarono a organismi con necessità nutritive estremamente semplici; 3) probabilmente venne elaborato un sistema di trasporto di elettroni in grado di utilizzare accettori inorganici di elettroni, corrispondente al processo di respirazione anaerobica; 4) si sviluppò poi un sistema di trasporto di elettroni in processi correlati per la fosforilazione fotosintetica, simili a quello che oggi si riscontra nei batteri fotosintetici, evoluto forse verso un sistema in grado di liberare ossigeno; 5) con l'accumulo di ossigeno si sarebbero infine sviluppati i procarioti aerobi.
Potrebbe essere particolarmente interessante verificare come queste ipotesi siano in accordo con i dati sempre più numerosi sulle correlazioni filogenetiche tra i Procarioti. Supponendo che la cellula eucariotica si sia sviluppata in un periodo successivo a circa 1×109 anni or sono, come si può essere determinato questo sviluppo? Non lo sappiamo, ma le somiglianze morfologiche e funzionali tra mitocondri e cloroplasti da un lato e Procarioti dall'altro hanno indotto a formulare l'ipotesi che un eucariote primitivo, in grado di ottenere l'energia necessaria per fosforilazione a livello del substrato, abbia stabilito un rapporto di simbiosi intracellulare con un procariote, rapporto che gradualmente ha determinato la perdita da parte del procariote della maggior parte delle funzioni, salvo quella della produzione di energia. Il fatto che mitocondri e cloroplasti contengano DNA e ribosomi 70 S (le dimensioni del procariote) ha dato valore a questo concetto, e del resto non sono insolite talune correlazioni simbiotiche tra microrganismi procarioti ed eucarioti.
Qual è la posizione dei Virus? In base a quanto abbiamo detto è possibile che non siano del tutto primitivi, ma piuttosto rappresentino un'evoluzione di alcune strutture contenenti RNA e DNA in grado di autoriprodursi in sede intracellulare.
8. Le applicazioni della microbiologia
All'inizio di questo secolo la microbiologia si presentava come una scienza ampiamente applicativa; negli ultimi tre decenni però si è sviluppata prevalentemente come scienza pura e solo recentemente le conoscenze risultanti da questo esplosivo incremento di attività hanno cominciato a essere applicate. Per opportunità descriveremo le applicazioni della microbiologia suddividendole in tre parti principali: innanzi tutto la microbiologia medica, poi quella ambientale e agricola, infine la microbiologia alimentare e industriale.
a) Microbiologia medica
Abbiamo già detto che lo sviluppo iniziale della microbiologia si è avuto con la scoperta del potere patogeno dei microrganismi. In effetti la maggior parte dei progressi più importanti in campo microbiologico medico si erano potuti presagire nel secolo scorso: erano state descritte le tecniche d'isolamento di molti microrganismi che provocano malattie infettive e bisognava solo perfezionarle. Tuttavia il progresso maggiore in questo campo è rappresentato dall'isolamento dei virus umani e dalla delucidazione dell'ampio spettro di malattie che possono provocare; in alcuni casi l'eziologia delle affezioni è stata chiarita molto recentemente e un esempio è la genesi virale di alcuni tipi di neoplasie (v. neoplasie: Oncologia sperimentale). Un virus tumorale o oncogeno era stato descritto prima del 1890 nei polli e l'esistenza di vari altri virus che determinano neoplasie era stata dimostrata prima dell'inizio di questo secolo. Non è stato però provato definitivamente che le neoplasie maligne umane possano essere indotte da virus, sebbene si sia osservato che molti virus dell'uomo provocano neoplasie maligne nello hamster e nel topo. Invero esiste una mole cospicua di dati indiretti che depongono a favore della genesi virale di alcune neoplasie umane. Negli animali sono stati descritti due tipi principali di neoplasie provocate da virus: in uno la neoplasia secerne attivamente il virus oncogeno che può aggredire altre cellule animali al pari di virus infettivi, provocando però la comparsa di cellule neoplastiche; altre neoplasie indotte da virus, al contrario, non presentano particelle virali libere, che sono invece integrate nella cellula ospite. È evidente la somiglianza con il fenomeno della lisogenia, di cui abbiamo parlato a proposito dei virus batterici; infatti il processo di conversione lisogenica è simile a quello della trasformazione cellulare che si verifica nella formazione di una cellula neoplastica. Non vi è dubbio che future ricerche chiariranno il meccanismo che sta alla base di questa trasformazione indotta da virus e il significato che essa ha nel contesto del problema generale del cancro (v. neoplasie: Oncologia sperimentale).
A cavallo tra i due secoli R. Koch, con la creazione di ‛stazioni batteriologiche' in zone della Germania ove il tifo era diffusissimo, dimostrò l'importanza dei portatori umani per la diffusione delle infezioni: studi combinati batteriologici ed epidemiologici permisero di rilevare che convalescenti e portatori eliminavano il bacillo del tifo con le feci ed erano quindi fonte di ulteriori infezioni. In tal modo venne data una congrua base scientifica all'epidemiologia e, con l'analisi delle modalità di diffusione delle malattie, si definirono i metodi per la loro eliminazione. A questo fine sono stati d'ausilio i più recenti metodi di tipizzazione serica e fagica per l'individuazione rapida e soprattutto specifica dei microrganismi patogeni. Bisogna rilevare l'effetto dello sviluppo della microbiologia medica sull'incidenza delle malattie infettive: all'inizio del secolo queste erano la più importante causa di morte in tutti i paesi ma, da allora, si è verificata, per lo meno nelle nazioni economicamente più avanzate, una considerevole diminuzione della loro incidenza e dei decessi che esse provocano. Prendendo come esempio la Scozia, i decessi causati dalle malattie infettive più importanti sono diminuiti da circa ottocento per centomila individui nel 1900 a molto meno di cento ai nostri giorni. Le principali cause di morte erano tubercolosi, polmonite, bronchite, forme dissenteriche e scarlattina; oggi solo la polmonite e la bronchite sono ancora di un certo rilievo e generalmente soltanto negli individui anziani. Tali modificazioni si sono verificate in seguito al miglioramento dell'alimentazione, delle condizioni igieniche ambientali e del tenore di vita, ma negli ultimi trenta anni sono state accelerate dall'impiego di sostanze antimicrobiche e dalla maggior diffusione delle tecniche d'inoculazione. Alcune malattie, come il vaiolo, sembrano completamente scomparse, ma sebbene tale progresso sia motivo di notevole soddisfazione, non deve indurci ad arrestare i nostri sforzi. Molto rimane da fare per applicare le nostre conoscenze nei paesi sottosviluppati, che si trovano in questa condizione per la povertà e per l'elevato tasso di natalità. Non sono state messe a punto terapie realmente valide delle infezioni virali, situazione che non sorprende dato l'intimo rapporto tra moltiplicazione del virus e metabolismo della cellula ospite. Probabilmente sarà necessaria una conoscenza molto più dettagliata di questa correlazione prima che si possa conseguire un qualsiasi reale successo.
Un altro problema è rappresentato dalla frequenza sempre maggiore di ceppi di batteri patogeni resistenti agli agenti chemioterapici. Come abbiamo già detto, una resistenza a molti farmaci può essere facilmente trasferita per lo meno tra organismi correlati. Del resto la possibilità di disporre di nuovi antibiotici è probabilmente esaurita: una maggiore fiducia deve essere riposta quindi nel logico sviluppo di nuovi agenti chemioterapici derivati dalle nostre maggiori conoscenze sulla biochimica e sulla biologia molecolare dell'agente patogeno. Sebbene il passaggio dalla microbiologia di base alla chemioterapia sia stato sino a oggi trascurato, esso in futuro dovrà essere sviluppato perché esplichi tutta la propria efficacia.
Una parte della microbiologia che si è fortemente sviluppata negli ultimi decenni è quella che s'interessa ai meccanismi del potere patogeno dei microbi. I processi di difesa dell'ospite vengono esposti altrove (v. cellula: Patologia della cellula; v. febbre; v. infiammazione); qui dobbiamo prendere in considerazione l'aspetto microbico del rapporto parassitario. Che cosa determina il potere patogeno di un organismo e l'assoluta innocuità di un altro organismo, apparentemente in stretta correlazione con il primo? La capacità di provocare una malattia, cioè il potere patogeno, è una proprietà relativamente rara tra i microrganismi e dipende da tre componenti fondamentali: trasmissibilità, potere infettivo e virulenza. La trasmissibilità, o capacità di passare da un ospite all'altro, spesso implica la produzione in notevole quantità del microrganismo patogeno da parte dell'ospite infettato, oppure la capacità di sopravvivere in condizioni ambientali avverse. In altri casi le modalità di trasferimento possono proteggere il microrganismo nei suoi passaggi da un ospite all'altro, come si verifica nella trasmissione sessuale delle malattie veneree o quando c'è un insetto vettore. Il potere infettivo è indice della capacità dell'agente patogeno di aprire una breccia nella prima linea di difesa dell'ospite, a livello cioè della cute o delle mucose, mentre la virulenza è una misura delle manifestazioni del processo morboso ed è un fattore variabile che può fare aumentare o diminuire il potere patogeno. Tali fattori possono far variare molto la quantità degli organismi necessaria perché si determini infezione: cosi, per ciò che riguarda Salmonella typhi, agente eziologico della febbre tifoide, la dose infettante può essere molto piccola, mentre nel caso di altre salmonelle che alterano i cibi sono necessarie quantità più rilevanti.
L'infezione dell'ospite determina la comparsa di uno stato morboso che può portare a morte. Due fattori principali intervengono a questo proposito: innanzi tutto l'agente patogeno può produrre una o più tossine. Negli ultimi decenni sono state compiute molte ricerche sull'isolamento e la purificazione di queste tossine: si tratta in genere di proteine e sono tra i composti tossici più noti. Per esempio Clostridium botulinum, agente del botulismo, produce una tossina: 1 mg di questa proteina purificata può uccidere più di un milione di cavie e si è calcolato che una quantità di 3 kg potrebbe sterminare l'intera popolazione mondiale. Per provocare la comparsa dei sintomi della malattia evidentemente non è necessaria un'infezione massiva e sfortunatamente è facile da immaginare la minaccia che tali tossine rappresentano se vengono usate come arma di guerra biologica, minaccia notevolmente accresciuta dalla relativa facilità e dal basso costo con cui si può produrne quantità cospicue in forma purificata. Per fortuna esistono ancora problemi per ciò che riguarda una diffusione controllata a scopo bellico di microrganismi patogeni o di loro prodotti e fortunatamente c'è ancora il preconcetto, per quanto illogico, che il loro impiego sia disumano. La modalità d'azione di molte tossine è oggi ben nota; la maggior parte di esse sono enzimi o inibitori di enzimi dell'ospite. Così l'α-tossina di Clostridium welchii è una fosfolipasi che agisce sulle membrane cellulari dell'ospite, mentre la tossina difterica, attiva su una transferasi, inibisce la sintesi proteica dell'ospite.
Il secondo fattore che determina la gravità di una malattia è la capacità che l'agente patogeno ha d'invadere e di moltiplicarsi considerevolmente nei tessuti sani, cioè l'invasività. Un fattore d'invasività è chiaramente la capacità di superare i meccanismi di difesa dell'ospite e, in particolare, di resistere alla fagocitosi e alle sostanze con potere battericida presenti nei liquidi dell'organismo ospite. Un fattore importante per annullare la fagocitosi è il possesso di una capsula: così gli streptococchi spesso presentano una capsula costituita da un polisaccaride, l'acido ialuronico, e se questo viene scisso dall'enzima ialuronidasi, la virulenza del microrganismo può essere attenuata anche più di diecimila volte. Un altro fattore che facilita l'invasività è la capacità dell'agente patogeno di produrre aggressine che sono, in genere, enzimi che attaccano i tessuti dell'ospite: ne sono esempi molte depolimerasi extracellulari quali le polisaccaridasi, le proteasi e le lipasi.
Un importante settore ancora ampiamente inesplorato è quello della natura della specificità dell'ospite. La maggior parte dei microrganismi aggredisce solo una serie ristretta di ospiti, almeno per quanto riguarda l'induzione di malattia in senso clinico, e tuttavia organismi a essi correlati e che colpiscono altri ospiti sembrano pressoché identici anche per ciò che riguarda il possesso di antigeni di superficie comuni. Questa è risultata una circostanza molto fortunata per la produzione di vaccini contro organismi sia vivi sia uccisi, come dimostrato dal successo delle vaccinazioni antivaiolosa e antipoliomielitica. La natura di questa specificità è stata indagata dettagliatamente solo nel sistema batterio/batteriofago, in cui possono verificarsi o una reazione specifica con la superficie cellulare oppure reazioni intracellulari. In un ospite sensibile vi è un ulteriore elemento di specificità, nel senso che vi è spesso un'affinità per determinati organi o tessuti: così i meningococchi aggrediscono membrane sierose, gli pneumococchi hanno una predilezione per il tessuto polmonare, i gonococchi per le mucose delle vie genito-urinarie e così via. Un altro fattore può essere costituito da agenti specifici che stimolano o inibiscono lo sviluppo del microrganismo patogeno: per esempio si è dimostrato che l'eritritolo può essere un fattore di crescita per un microrganismo che provoca l'aborto nel bestiame, Brucella abortus, e sembra che esso si trovi solamente nella placenta bovina. Tuttavia in genere la risposta o le risposte a questo organotropismo rimangono oscure e, come per molti altri argomenti, possono richiedere una conoscenza più dettagliata sia dell'agente patogeno sia dell'ospite: tale conoscenza potrà portare, inoltre, a metodi di lotta contro le infezioni completamente nuovi. È questa una considerazione che deriva dall'idea che il futuro della microbiologia medica sarà sempre più legato alla conoscenza dettagliata dei meccanismi che sono alla base del potere patogeno; tale conoscenza probabilmente potrà essere acquisita solo con gli sforzi congiunti di gruppi di ricerca interdisciplinari.
Questa esposizione è stata sostanzialmente limitata a quei microrganismi che possono provocare una malattia in un ospite umano. Tuttawia un gran numero di microrganismi è quasi sempre presente nelle zone corporee che sono a contatto con l'ambiente esterno, come la pelle, la cavità orale, l'apparato respiratorio, quello intestinale e quello genito-urinario. Se si eccettua il processo della carie dentaria, questi microrganismi non hanno un effetto significativo sull'ospite, però è risultato evidente che i numerosisssimi microrganismi presenti nel tratto intestinale e soprattutto nelle sue regioni più basse, che hanno pH neutro, possono svolgere un ruolo importante dando luogo a una correlazione simbiotica. È sorprendente che questi microrganismi siano stati studiati accuratamente solo negli ultimi anni; si era pensato che essi fossero per lo più anaerobi facoltativi, come Escherichia coli, ma nuove tecniche di coltura degli anaerobi hanno posto in risalto che gli organismi prevalenti sono anaerobi obbligati. In due modi diversi si è visto che questi organismi possono essere importanti per l'ospite: in primo luogo gli antibiotici ad ampio spettro somministrati per via orale possono praticamente sterilizzare il tratto intestinale, almeno per quanto riguarda la maggior parte dei batteri che vi si trovano e, se ciò accade, possono svilupparsi organismi insoliti, come i lieviti, che non assolvono gli stessi compiti della flora normale e determinano quindi un'alterazione nociva della funzione digestiva; in secondo luogo gli effetti della presenza nell'ospite della normale flora batterica possono essere studiati con tecniche sviluppate recentemente che si avvalgono dell'uso di animali sterili (germ-free). La conclusione che in generale si è tratta da queste ricerche è che tali animali germ-free, se ricevono una dieta appropriata, sono del tutto vitali, pur presentando, sotto molti punti di vista, peculiarità fisiologiche e anatomiche; per esempio il loro intestino non è sviluppato, possono necessitare di sostanze nutritizie accessorie, come la vitamina K, e gli apparati che partecipano ai normali meccanismi di difesa dell'ospite sono poco sviluppati. È chiaro che bisognerà prestare maggiore attenzione all'intimo funzionamento della nostra normale flora microbica.
b) Microbiologia ambientale e agricola
I microrganismi rappresentano una componente significativa della massa vivente universale e, se si prendono in considerazione anche i loro elevati ritmi metabolici e di crescita e la loro adattabilità, il contributo che essi danno all'ecosistema in toto è notevolmente superiore a quanto potrebbero indicare le loro dimensioni.
Durante i primi decenni di questo secolo furono raccolti molti dati sulla diffusione e sul ruolo dei microrganismi nel suolo e nell'acqua. Ne derivò il concetto generale dei cicli degli elementi, concetto che è stato ampiamente elaborato nei decenni a noi più vicini: l'importanza di questi cicli per la conservazione della vita degli animali, delle piante e dell'uomo è divenuta sempre più chiara.
I cicli degli elementi. - L'accrescimento, inteso in funzione della massa vivente complessiva della Terra, consiste nella conversione di alcuni elementi da una forma inorganica a una organica strutturata che costituisce gli organismi viventi. La quota di gran lunga maggiore di energia necessaria per tale processo viene ottenuta dalla luce solare mediante la fotosintesi. Perché queste conversioni possano continuare in presenza di quantità limitate di elementi in uno stato organico, deve esistere un corrispondente ritmo di riconversione: è questo il processo di mineralizzazione. Un'ulteriore complicazione è rappresentata dal fatto che le forme inorganiche degli elementi possono riscontrarsi sia allo stato ossidato, sia a quello ridotto, ed è essenziale, per la conservazione della vita, che essi non vengano accumulati soltanto sotto una di queste forme.
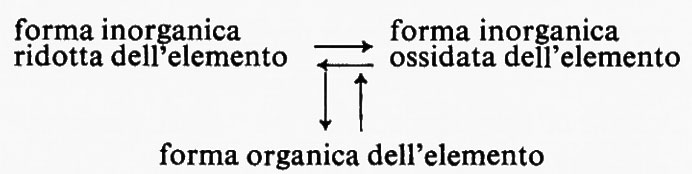
Consideriamo il ciclo del carbonio e l'importanza dei microrganismi nella sua attuazione. Esso, nella sua forma più semplice, può venire schematizzato come segue:
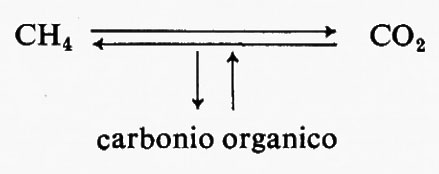
Il principale metodo di formazione del carbonio organico è la fotosintesi delle piante superiori e delle Alghe eucariotiche; in alcuni ambienti, anche organismi procariotici fotosintetici, come le Alghe verdi-azzurre, possono contribuire in maniera rilevante. Una piccola quantità di CO2 può anche essere convertita in carbonio organico da batteri chemiolitotrofi che utilizzano l'energia di ossidazioni inorganiche. Anche il metano può essere assimilato in carbonio organico e poi trasformato in CO2 da un gruppo di batteri che lo metabolizzano: l'isolamento di una serie di questi procarioti e l'accertamento della loro importanza per l'ecosistema sono conquiste recenti della microbiologia. Comunque, il contributo di gran lunga più importante dei microrganismi, e in particolare dei Procarioti, al ciclo del carbonio consiste nella mineralizzazione. Abbiamo già ricordato che i Batteri sono capaci di ossidare a CO2 tutti i composti organici naturali: se così non fosse ben presto tutto il carbonio della Terra verrebbe convertito in composti non biodegradabili. In condizioni anaerobiche la mineralizzazione è possibile solo parzialmente, tramite il processo di respirazione anaerobica, con formazione di metano e di CO2 insieme a una varietà di composti, parzialmente degradati, contenenti carbonio organico.
Un ciclo più complesso, per ciò che riguarda le interconversioni delle forme inorganiche dell'elemento che vi partecipano, è quello dell'azoto. In molti ecosistemi naturali, e certamente in quelli artificiali creati dall'agricoltura, un sufficiente livello di composti azotati utilizzabili è spesso il fattore che limita l'incremento della massa vivente. Lo sviluppo di organismi viventi determina la conversione dell'azoto inorganico in azoto organico (proteine, acidi nucleici, ecc.). I composti inorganici azotati più adatti per la crescita delle piante sono i nitrati, mentre il principale prodotto del processo di mineralizzazione microbica è l'ammoniaca liberata con le reazioni di deamminazione; in ambienti aerobici l'ammoniaca può essere ossidata a nitrato da un gruppo di batteri chemiolitotrofi, detti batteri nitrificanti. Sfortunatamente però esiste un altro gruppo di batteri che utilizzano i nitrati come accettori di elettroni per la respirazione anaerobica e tale processo porta alla riduzione del nitrato ad azoto molecolare. Se non si verificasse un'eventuale perdita di tutto l'azoto fissato disponibile per la crescita degli organismi viventi, dovrebbe aver luogo il processo inverso della fissazione dell'azoto. Sebbene la fissazione dell'azoto possa in parte attuarsi con semplici processi chimici, tuttavia la fonte principale è biologica ed è caratteristica dei Procarioti. I procarioti azotofissatori possono esistere sia liberi, sia come simbionti, per esempio nell'associazione tra Batteri e noduli radicali delle Leguminose. Si riteneva molto limitata, nell'ambito dei Procarioti, la capacità di fissare azoto ma nuovi metodi di saggio hanno posto in evidenza che essa non è insolita, in adatte condizioni ambientali.
Il processo di fissazione dell'azoto è stato molto studiato negli ultimi anni, poiché rappresenta un campo in cui sussiste la reale possibilità d'incrementare le rese agricole senza aumentare l'impiego di costosi fertilizzanti azotati. Le tappe biochimiche alla base del processo sono state chiarite e si è visto che i sistemi enzimatici, al di fuori della cellula, sono estremamente sensibili all'ossigeno: il meccanismo che, in sede intracellulare, protegge dall'ossidazione può essere un fattore critico che limita l'ampliamento della sua diffusione. Quali metodi si possono impiegare in agricoltura per aumentare la fissazione biologica? Si stanno attualmente indagando diverse possibilità: 1) la scoperta di nuovi metodi biologici naturali di fissazione dell'azoto; per esempio recenti indagini hanno portato alla scoperta di un tipo di rapporto simbiotico tra batteri azotofissatori ed erbe naturali e altre piante del Brasile, che è meno specializzato di quello presente nei noduli delle Leguminose. Non si conosce ancora bene, però, la diffusione di tali sistemi né la possibilità di una loro applicazione ai raccolti agricoli; 2) l'inoculazione di batteri azotofissatori nei terreni. Questo metodo può non incrementare significativamente la resa dei raccolti, poiché in molte zone sono già naturalmente presenti microrganismi azotofissatori, ma può portare alla selezione di ceppi più redditizi; 3) la selezione di nuovi procarioti azotofissatori con l'ausilio dell'ingegneria genetica. La genetica dei batteri azotofissatori è stata compresa recentemente e si è visto che l'operon in causa può essere trasferito a Escherichia coli da un batterio ad esso correlato. Non è noto se questo processo possa essere esteso; 4) il trasferimento genetico del sistema di fissazione dell'azoto nelle piante stesse. La realizzazione di questo metodo sembra ancora molto lontana e potrebbe presentare problemi insormontabili.
Queste diverse possibilità sono esempi del modo in cui la conoscenza dei meccanismi su cui si fondano i cicli degli elementi può essere di ausilio per l'uomo. Ma allora, a livello d'interrelazione tra i singoli organismi o tra gruppi di organismi, come si svolgono questi processi e quali sono i principi ecologici più delicati implicati in essi? Sfortunatamente l'ecologia microbica si è limitata all'enumerazione dei microrganismi presenti in un particolare ambiente, anche perché lo stabilire le interazioni fra questi microrganismi è un problema di formidabile complessità. La difficoltà dipende in parte dal livello d'informazioni sui rapporti distributivi delle cellule in un ambiente, se cioè sono distribuite casualmente o se sono associate in microambienti ristretti, quali si riscontrano nel suolo. Sono stati compiuti alcuni tentativi per progredire, in base a studi di colture pure, di dubbia importanza per l'analisi di sistemi di coltura mista. In effetti bisogna rilevare che l'analisi anche di sistemi semplici di coltura mista è estremamente difficile e che il microbiologo tende tuttora a creare, in queste ricerche, condizioni di sviluppo del tutto innaturali: per esempio, la maggior parte dei microrganismi cresce sulla superficie di particelle di argilla o di detriti organici del suolo, su materiale organico in sospensione e sugli scogli degli oceani, dei laghi e dei fiumi, sulla pelle, sui denti e sui tessuti epiteliali degli animali e sulle radici o sulle foglie delle piante; invece in laboratorio la crescita avviene di solito in mezzi liquidi con una concentrazione innaturalmente elevata di sostanze nutritive. Lo studio dell'effetto delle superfici sulla crescita, ancor oggi in fase iniziale, diverrà sempre più importante. Un altro fenomeno frequente in natura, e che invece è stato studiato molto poco in laboratorio, è la tendenza dei microrganismi a rimanere uniti per flocculazione. Ne è un esempio, presente ogni giorno nel nostro organismo, l'associazione che si ha a livello dell'apparato dentario: alcuni batteri elaborano sulla superficie del dente un destrano extracellulare, che viene prodotto specificamente a partire dal saccarosio e che mantiene unito un gruppo di organismi diversi, tenendoli saldamente legati all'idrossiapatite dello smalto.
Bisogna augurarsi che gli studiosi interessati all'ecologia microbica, il cui numero va sempre crescendo, non siano attratti dall'apparente semplicità dell'argomento, rispetto ai campi di ricerca biochimico, genetico o di biologia molecolare: tale apparente semplicità è del tutto illusoria ed è solo con l'ausilio di tecniche sempre più fini e complesse che sarà possibile un'effettiva conoscenza degli ecosistemi naturali.
Inquinamento e microbiologia. - Oggi si presta un'attenzione sempre maggiore all'inquinamento del nostro ambiente, sia esso terrestre o idrico. Due tipi d'inquinamento interessano il microbiologo: in primo luogo quello biologico, con il quale microrganismi potenzialmente dannosi vengono introdotti nelle riserve idriche utilizzate per bere o per bagnarsi; in secondo luogo l'inquinamento biodegradabile, che si compie riversando nell'acqua o nel suolo sostanze in grado di essere utilizzate come nutrimento dai microrganismi, i quali possono quindi crescere e spostare l'equilibrio ecologico dell'ambiente ponendo in pericolo l'esistenza delle piante più evolute, degli animali e, infine, dell'uomo stesso.
La contaminazione delle riserve idriche a opera di microrganismi e, in particolare, di quelli presenti nelle deiezioni umane, è stata considerata innanzitutto un problema di salute pubblica per l'introduzione in acqua di agenti patogeni potenziali. Però con lo sviluppo dell'urbanizzazione e quindi con l'aumentare dell'inquinamento dovuto agli scarichi, essa è divenuta anche un problema ecologico. Il tasso elevato di materiale organico che viene scaricato in un fiume o in un lago, a opera degli organismi aerobi eterotrofi, può rendere l'acqua povera di ossigeno con conseguenze sgradevoli. La soluzione di entrambi i problemi sta nell'applicazione di metodi di trattamento delle acque di scolo, in modo da renderle prive degli agenti patogeni potenziali e della maggior parte dei materiali biodegradabili che contengono. In genere questo trattamento viene compiuto in due fasi: la prima consiste nella sedimentazione delle particelle solide in ampie vasche di decantazione e nella decomposizione anaerobica del materiale sedimentato; nella seconda il materiale in soluzione viene sottoposto a una mineralizzazione batterica aerobica o in un filtro a gocciolamento, ove il materiale viene spruzzato sulla superficie di uno strato di pietre, oppure per gorgogliamento d'aria in una vasca nel processo dei ‛fanghi attivati'. Tali metodi sono stati sviluppati in gran parte empiricamente e sono legati all'arricchimento naturale in microrganismi flocculanti che contemporaneamente operano la mineralizzazione. I dettagli microbiologici di questo processo, che è di fondamentale importanza per la nostra civiltà moderna, devono essere ancora in gran parte elaborati. Gli stessi metodi generali possono essere impiegati per il trattamento dei rifiuti organici biodegradabili dell'industria e dell'agricoltura che inquinano sempre di più l'ambiente.
Quando le nostre conoscenze sui processi di trattamento degli scarichi e di altri rifiuti organici saranno più progredite, sarà possibile utilizzarle meglio. In un mondo in cui vi è carenza di fonti d'energia e di fonti di carbonio organico, diviene sempre più dispendioso sviluppare processi che dissipano progressivamente il potenziale rappresentato dai prodotti di rifiuto della nostra civiltà. I microbiologi sono sempre più consci delle possibilità dei metodi microbiologici di trattamento per trasformare i prodotti di rifiuto in materiali utilizzabili: per esempio il metano, prodotto con il trattamento anaerobico del materiale di scarico, comincia già a essere impiegato come fonte d'energia per l'intero impianto di depurazione. Contemporaneamente si compiono sforzi per eliminare l'impiego di sostanze non biodegradabili per la presenza di componenti instabili, sia mediante la riformulazione delle sostanze chimiche utilizzate, così che possano esse stesse essere attaccate da agenti microbici (detergenti biodegradabili), sia controllando l'evoluzione di determinati microrganismi che possano essere in grado di attaccare sostanze non presenti in natura. Quest'ultimo metodo è solo agli inizi, ma è già chiaro che è possibile modificare la specificità di certi enzimi selezionando appropriati mutanti: in tal senso è stato solo possibile provocare modificazioni relativamente modeste della specificità di substrato di enzimi noti, ma il potenziale è enorme.
Microbiologia agricola. - Abbiamo già ricordato molte applicazioni della microbiologia all'agricoltura: i raccolti dipendono dall'appropriato svolgimento dei cicli degli elementi e la conoscenza delle modificazioni chimiche causate dai microrganismi nel terreno può portare a metodi più efficienti di produzione dei raccolti.
Per ciò che riguarda l'allevamento degli animali i microrganismi esplicano un ruolo fondamentale nel processo di nutrizione dei Ruminanti. La microbiologia del rumine, per la difficoltà d'isolare e di far crescere gli anaerobi obbligati in esso presenti, è stata studiata in modo appropriato solo negli ultimi vent'anni. Oggi è chiaro che esiste un gruppo cospicuo di Batteri e di Protozoi che determina la scissione della cellulosa, ed eventualmente di altri costituenti vegetali, sino ad acidi grassi, che vengono assorbiti dal ruminante, per il quale rappresentano la fonte principale di carbonio e di energia; gli altri prodotti sono il metano, che viene allontanato con le eruttazioni, e le cellule microbiche, che vengono digerite nel tratto terminale dell'intestino per fornire amminoacidi, vitamine e altre sostanze nutritive. Oggi è possibile analizzare questi processi in chemostati artificiali, utilizzando sistemi di coltura mista, e indagare alcune modificazioni che in essi si verificano; tali indagini porteranno a metodi pratici d'incremento dell'efficienza del rumine.
Le malattie possono tuttora provocare gravi danni sia agli allevamenti sia ai raccolti. La microbiologia veterinaria ha mostrato la tendenza a seguire gli indirizzi della microbiologia medica, dato che l'economia agricola ha consentito di applicare agli animali le acquisizioni della medicina. Fattori completamente diversi intervengono nelle malattie delle piante, ma le nostre conoscenze dei meccanismi delle condizioni patologiche e della reazione della pianta all'aggressione microbica sono ancora agli inizi. La coltivazione di piante in grado di resistere alle malattie di origine microbica è prevalentemente un processo empirico ed è necessario ancora che microbiologi e agricoltori sviluppino una più chiara conoscenza dei meccanismi che vi partecipano, prima che si possano compiere ulteriori progressi.
c) Microbiologia alimentare e industriale
Con la scoperta, verso la fine del secolo scorso, dei metodi d'isolamento e d'identificazione dei microrganismi, apparve chiaro che la maggior parte degli alimenti ospita una popolazione microbica naturale che può accrescersi durante i processi di conservazione degli alimenti stessi, oppure può aumentare per effetto della contaminazione atmosferica o di quella da utensili, da animali e da uomini. La presenza di questi microrganismi può far variare semplicemente le caratteristiche organolettiche dell'alimento, oppure può determinare lo sviluppo di agenti patogeni, causando quindi l'avvelenamento del cibo. Da questo punto di partenza la microbiologia alimentare si è sviluppata, nel corso del secolo, in numerose direzioni.
1. La diminuzione del numero dei microrganismi inizialmente presenti nell'alimento e la riduzione della contaminazione. Sfortunatamente è difficile allontanare tutti i microrganismi con mezzi fisici, se si eccettua la filtrazione dei liquidi, e i risultati che si ottengono con il lavaggio sono modesti. La prevenzione della contaminazione, soprattutto da agenti patogeni potenziali, consiste prevalentemente nell'adozione di presidi igienici ottimali e a questo fine sono state gradualmente elaborate leggi sempre più rigide.
2. La distruzione dei microrganismi presenti nell'alimento. Lo sviluppo delle tecniche per tale distruzione prese origine naturalmente dai metodi di sterilizzazione e, tra questi, il calore è stato di gran lunga il metodo più comune. Esso può essere impiegato per ottenere una distruzione selettiva, come in una serie di tecniche che vanno sotto il nome di pastorizzazione, oppure per conseguire una più completa sterilizzazione, al fine di una conservazione a lungo termine dell'alimento, come nell'inscatolamento. Sono stati effettuati anche tentativi di utilizzare radiazioni ad alta energia, ma sfortunatamente gli alti dosaggi necessari provocano alterazioni chimiche degli alimenti con conseguente modificazione del sapore.
3. Per un periodo di tempo breve può essere sufficiente modificare l'ambiente rappresentato dall'alimento, così da inibire lo sviluppo di Batteri. I metodi impiegati sono derivati spesso da procedimenti tradizionali e implicano la disidratazione (diretta o per aumento della pressione osmotica dovuto all'aggiunta di sale o di zucchero), la refrigerazione, l'acidificazione, o l'aggiunta di un inibitore della crescita batterica come il nitrato, il nitrito, il benzoato, ecc. Ricerche effettuate su tali fattori ambientali ne hanno evidenziato i possibili effetti sinergici, la cui adeguata conoscenza può portare a metodi più economici di conservazione del cibo. Gli stessi microrganismi possono essere impiegati per modificare l'alimento così da prevenire lo sviluppo di altri organismi indesiderabili, accrescendo contemporaneamente le proprietà nutritive del cibo: esempi tradizionali sono il formaggio e lo yoghurt, derivati dal latte, i crauti e, come alimento animale, il foraggio insilato. Molti di questi metodi sono basati sull'arricchimento in batteri acido-lattici che, in condizioni anaerobiche, riducono rapidamente il pH a circa 3,5 e quindi impediscono lo sviluppo di altri microrganismi.
L'impiego vantaggioso di microrganismi in processi industriali su larga scala è stato sviluppato in questo secolo a partire da altri procedimenti tradizionali, quali la produzione di bevande alcoliche, del lievito per panificazione e dell'aceto. Tale industria viene in genere detta ‛delle fermentazioni', sebbene si debba notare che molti dei metodi più recenti sono in effetti aerobici e perciò non implicano un processo fermentativo come tale. Esula dai nostri scopi esporre la tecnologia del fermentatore, sebbene esso esplichi un ruolo fondamentale nell' economia di procedimenti moderni che richiedono aerazione adeguata e conveniente. Tuttavia uno sviluppo che dev'essere ricordato è il progressivo incremento dei metodi di coltura continua in campo industriale, con conseguente considerevole risparmio nel costo degli impianti e con un'accresciuta efficienza e un maggior controllo del processo di fermentazione. È opportuno considerare i procedimenti industriali sotto due aspetti principali a seconda che trattino di cellule o dei loro prodotti.
Produzione di cellule. - All'inizio di questo secolo la produzione industriale di cellule era in gran parte limitata al lievito per panificazione, ma negli ultimi dieci anni si è registrato un incremento notevole dell'interesse per la produzione su larga scala di microrganismi proprio come alimenti, interesse che è stato molto stimolato dall'attuale carenza mondiale di proteine. Alcuni vantaggi della produzione di proteine facenti parte di microrganismi (le cosiddette single-cell proteins o S.C.P.), rispetto ai metodi agricoli convenzionali, consistono nel ritmo di crescita notevolmente più rapido e quindi nella maggiore sintesi proteica, nell'alto valore nutritivo delle proteine microbiche, nell'elevato grado di controllo che può essere esercitato sulla produzione, nello spazio modesto richiesto, nella facilità delle modificazioni fenotipiche e genotipiche dei produttori di proteine e nel modesto livello d'inquinamento determinato dal processo produttivo. Dovendo scegliere una fonte di carbonio e di energia per la produzione di proteine da parte di organismi unicellulari, è soprattutto necessario che sia disponibile materiale in quantità considerevoli, di basso costo e non soggetto a variazioni stagionali. Al momento attuale i metodi su scala industriale dipendono dall'impiego di combustibili fossili, siano essi n-alcani derivati dal petrolio o metano o metanolo derivati dal gas naturale. È stato dimostrato che è possibile ottenere una conversione di queste sostanze in proteine in una percentuale superiore al 50% in peso, ma purtroppo il notevole aumento del costo dei combustibili fossili ha reso l'economicità del processo molto meno competitiva che all'inizio. Di conseguenza è difficile prevedere quale sarà il ruolo della produzione di proteine da organismi unicellulari, come materiale alimentare per gli animali o direttamente per l'uomo, ma può darsi che i microrganismi fotosintetici, che utilizzano la luce solare come fonte di energia, costituiscano un migliore investimento a lungo termine.
Vi è anche interesse per un gruppo di batteri chemiolitotrofi che ricavano l'energia loro necessaria dall'ossidazione dell'idrogeno ad acqua: sebbene tale interesse derivi dalla possibilità d'impiegare tali organismi quale fonte di cibo per viaggi spaziali di lunga durata, nel corso dei quali l'idrogeno può essere prodotto per elettrolisi dell'acqua utilizzando batterie solari, il processo diverrà importante, da un punto di vista generale, se sarà disponibile una fonte di energia elettrica a buon mercato.
Produzione di sostanze chimiche. - I più importanti prodotti dell'industria fermentativa erano, sono e si può ritenere che saranno le bevande alcoliche. Le tecniche di produzione sono state scarsamente modificate per le tendenze conservatrici dell'industria e dei suoi clienti. Nei primi decenni di questo secolo la capacità dei microrganismi di sintetizzare un'ampia gamma di prodotti fermentativi diversi venne applicata alla preparazione di sostanze come l'alcool etilico, l'acido lattico, l'acido acetico, l'alcool butilico e l'acetone. Però l'aumento dei costi delle fonti di energia e di carbonio necessarie per la fermentazione e lo sviluppo di processi petrolchimici più efficienti resero questi procedimenti microbiologici progressivamente meno economici e nel 1940 l'industria fermentativa era quasi ferma ai procedimenti tradizionali. In quel periodo vennero scoperti gli antibiotici e nei decenni successivi si ebbe una ripresa dell'industria, poiché, salvo poche eccezioni, è più conveniente produrre le complesse molecole antibiotiche con procedimenti biologici piuttosto che con quelli chimici. Con la scoperta di antibiotici nuovi, altre ricerche portarono all'individuazione di mutanti provvisti di un più elevato livello produttivo. Divenne altresì chiaro che gli antibiotici erano esempi di un gruppo di sostanze, dette metaboliti secondari, prodotte principalmente dopo la fine della fase di crescita esponenziale in colture a lotto discontinuo (batch cultures). Ciò ha creato difficoltà all'introduzione di colture continue per la produzione di antibiotici, ma il problema potrà essere risolto in base a una migliore conoscenza del processo cellulare che controlla la produzione del metabolita secondario, oppure dall'uso di sistemi di coltura continua più complessi, a molti stadi.
L'impulso dato all'industria della fermentazione dai metodi di produzione degli antibiotici e la conoscenza sempre più progredita della biologia molecolare e della genetica hanno indotto a progettare una nuova vasta serie di processi potenziali, che ora possono essere economici, e tale tendenza è stata ulteriormente stimolata dal costo crescente delle sostanze petrolchimiche. Solo pochi però dei processi potenziali sono stati realizzati: per esempio possono essere ottenuti mutanti multipli di microrganismi che non presentano più un'inibizione o una repressione per feedback della biosintesi di un particolare amminoacido o di una vitamina, e ciò porta a un aumento notevole della produzione. Tali mutanti sono stati impiegati commercialmente in misura sempre maggiore.
Esiste anche un notevole interesse per la produzione di enzimi su scala industriale, interesse che ha avuto inizio con l'uso di proteasi nell'industria dei detergenti e di amilasi nell'idrolisi industriale dell'amido. Anche in questo caso è necessario isolare mutanti per i quali la sintesi dell'enzima richiesto non è più sottoposta alla repressione e un ulteriore incremento della produzione di enzimi si potrà verificare con l'introduzione di copie multiple degli appropriati geni strutturali. Il potenziale della produzione industriale degli enzimi è cospicuo e si è solo all'inizio della sua realizzazione. La gamma degli enzimi disponibili è enorme e, con i metodi ricordati precedentemente, è possibile modificare la specificità degli enzimi, così che questi possano favorire l'espletamento di nuovi processi che non si verificano in natura. Non vi è dubbio che si registrerà un incremento dell'utilizzazione degli enzimi per i processi chimici industriali di sintesi e di scissione. Questo è anche un altro esempio del modo in cui le maggiori conoscenze della microbiologia di base potranno essere eventualmente applicate a beneficio del genere umano. Bisogna solo sperare che la maggiore attenzione per la microbiologia applicata, insieme all'inevitabile passaggio a organismi viventi più evoluti come sistemi modello, non porti a una diminuzione dell'impulso a indagare i campi fondamentali della microbiologia, che continuano a offrire un panorama sempre più ampio di affascinanti problemi.
bibliografia
Ainswort, G. C., Sussman, A. S. (a cura di), The Fungi, voll. I, II, III, IV, New York 1965, 1966, 1968, 1973.
Ajl, S. J., Kadis, S., Montie, T. C. (a cura di), Microbial toxins, New York 1970.
Alexander, M., Microbial ecology, New York 1971.
Bachmann, B. J., Low, K. B., Taylor, A. L., Recalibrated linkage map of Escherichia coli K-12, in ‟Bacteriological reviews", 1976, XL, p. 116.
Bonner, J. T., The cellular slime moulds, Princeton 19672.
Brock, T. D. (a cura di), Milestones in microbiology, Englewood Cliffs, N. J., 1961.
Brock, T. D., Biology of microorganisms, Englewood Cliffs, N. J., 19742.
Buchanan, R. E., Gibbons, N. E. (a cura di), Bergey's manual of determinative bacteriology, Baltimore 19748.
Burnet, F. M., White, D. O., Natural history of infectious diseases, Cambridge 19724.
Collee, J. G., Applied medical microbiology, Oxford 1976.
Davis, B. D., Dulbecco, R., Elsen, H. N., Ginsberg, H. S., Wood, W. B. Jr., Microbiology, New York 19732 (tr. it.: Microbiologia, Roma 1971).
Dawes, I. W., Sutherland, I. W., Microbial physiology, Oxford 1976.
Hall, J. B., Evolution of prokaryotes, in ‟Journal of theoretical biology", 1971, XXX, p. 429.
Hayes, W., Genetics of bacteria and their viruses, Oxford 19682.
Hershey, A. D. (a cura di), The bacteriophage lambda, Cold Spring Harbour 1971.
Hungate, R. E., The rumen and its microbes, New York 1966.
Lechevalier, H. A., Solotorovsky, M., Three centuries of microbiology, New York 19672.
Luria, S. E., Darnell, J. G., General virology, New York 19672.
Mandel, M., New approaches to bacterial toxonomy, in ‟Annual review of microbiology", 1969, XXIII, p. 239.
Mandelstam, J., McQuillen, K., Biochemistry of bacterial growth, Oxford 19722.
Miller, S. L., Orgel, L. E., The origin of life on the earth, Englewood Cliffs, N. J., 1974.
Postgate, J., Microbes and man, Harmondsworth 19752.
Smith, H., Pearce, J. H., Microbial pathogenicity in man and animals, Symposium 22 of Society for General Microbiology, Cambridge 1972.
Stanier, R. Y., Doudoroff, M., Adelberg, E. A., The microbial world, Englewood Cliffs, N. J., 19703 (tr. it.: Il mondo dei microrganismi, Bologna 1975).
Stanier, R. Y., Neil, C. B. van, The concept of a bacterium, in ‟Archiv für Mikrobiologie", 1962, XLII, p. 17.
Watson, J. D., Molecular biology of the gene, New York 19763 (tr. it.: Biologia molecolare del gene, Bologna 19722).
Wilkinson, J. F., Introduction to microbiology, Oxford 19752.