
Orologi biologici circadiani
Orologi biologici circadiani
La vita sulla Terra si è evoluta in un contesto caratterizzato da imponenti variazioni ambientali, alcune di natura ciclica. È il caso, per esempio, dell'alternanza del giorno e della notte, dovuta alla rotazione terrestre attorno al proprio asse che si completa, incessantemente, ogni ventiquattro ore. Anche i moti di rivoluzione della Luna attorno alla Terra e della Terra attorno al Sole generano variazioni cicliche ambientali, come nel caso delle maree o delle stagioni. La maggior parte degli organismi viventi si è quindi evoluta in ambienti che hanno selezionato adattamenti a oscillazioni di parametri ambientali in diversi domini temporali. Tra le manifestazioni più evidenti di questi adattamenti vi sono comportamenti ritmici quali i fenomeni migratori stagionali, che caratterizzano un gran numero di specie di Uccelli, la maturazione delle gonadi e il raggiungimento della fertilità confinato in alcuni specifici periodi dell'anno, come avviene nelle pecore, che si riproducono solamente in autunno, garantendo alla progenie, che nasce in primavera, condizioni ambientali favorevoli nel delicato periodo che segue alla nascita.
Questi adattamenti alle oscillazioni di parametri ambientali sono in gran parte regolati da orologi biologici endogeni che 'dettano' il tempo agli organismi e consentono loro di anticipare variazioni periodiche dell'ambiente esterno, come il sorgere o il tramontare del Sole, l'alta marea o l'approssimarsi dell'inverno. Ciò permette agli organismi di 'prepararsi' per tempo ai cambiamenti che stanno per sopraggiungere, modificando il loro metabolismo e il loro comportamento ed evitando così di subire passivamente le fluttuazioni ritmiche dei parametri ambientali. Gli orologi biologici endogeni, costituiti da particolari geni e proteine, producono quindi variazioni molecolari cicliche durante tutta la vita di un organismo e vengono continuamente sincronizzati con i cicli astronomici, che sottolineano la loro periodicità, a opera di stimoli ambientali ritmici. Tra questi, particolare importanza per la sincronizzazione in natura degli orologi circadiani riveste l'alternarsi, ogni ventiquattro ore, delle fasi di luce e buio, mentre importante è la durata della fase luminosa (fotoperiodo) per la sincronizzazione degli orologi biologici circannuali. In relazione alla frequenza che caratterizza l'oscillazione generata da questi orologi endogeni, essi vengono classificati come orologi ultradiani (il periodo della loro oscillazione è compreso tra alcuni millisecondi e alcune ore), circadiani (il periodo approssima le ventiquattro ore) e infradiani (il periodo è di settimane, mesi o anni). La cronobiologia si occupa dello studio di questi oscillatori endogeni e in particolare della loro dissezione a livello genetico, molecolare e fisiologico. A partire dagli anni Novanta del XX sec. sono stati compiuti progressi straordinari, soprattutto per quanto riguarda la comprensione del funzionamento degli orologi circadiani, mentre rimangono ancora ampiamente sconosciute la natura e l'organizzazione molecolare degli orologi operanti in altri domini temporali, quali gli orologi 'circalunari' o 'circannuali'.
Ritmi circadiani
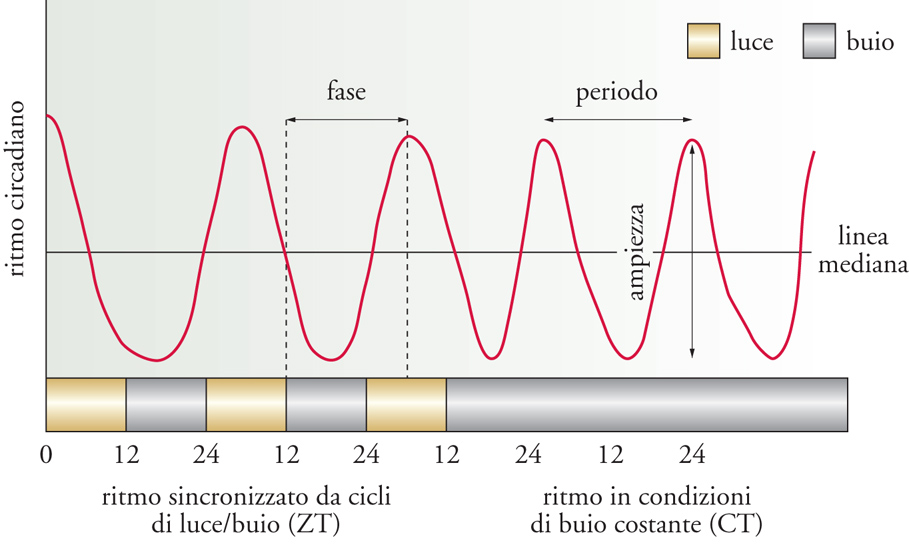
Gli orologi biologici circadiani (dal latino circa diem) oscillano con un periodo che si approssima alla durata di un giorno astronomico e regolano una grande varietà di fenotipi ritmici che caratterizzano il metabolismo, la fisiologia e il comportamento della maggior parte degli organismi che popolano il nostro pianeta. Il movimento delle foglie o l'apertura degli stomi di alcune piante nel corso della giornata, le variazioni cicliche della pressione sanguigna e della temperatura corporea nei Mammiferi, così come l'alternanza delle fasi di sonno e di veglia negli animali, sono tutti fenomeni controllati da uno o più oscillatori endogeni che sono comparsi, e si sono organizzati nel corso dell'evoluzione, in risposta alle variazioni ambientali cicliche prodotte dalla rotazione della Terra attorno al proprio asse. Un ritmo circadiano può essere descritto definendo il periodo che lo caratterizza, l'ampiezza del carattere che oscilla e la fase, cioè lo stato dell'oscillazione in un determinato momento rispetto a un punto di riferimento nel ciclo (fig. 2). Più in generale, il periodo associato a un ritmo corrisponde al tempo che impiega una determinata fase dell'oscillazione a ripresentarsi. La fase di un fenomeno biologico ritmico corrisponde allo stato 'istantaneo' di una oscillazione in relazione a un particolare punto di riferimento (per es., l'accensione della luce o il suo spegnimento). L'ampiezza di un ritmo è una misura del suo livello di espressione (corrisponde alla misura dell'intervallo compreso tra i valori massimi e minimi dell'attività ritmica esaminata). I ritmi circadiani generati da un oscillatore biologico persistono in condizioni costanti, cioè in assenza di stimoli ambientali, come l'alternanza del giorno e della notte. Inoltre, gli oscillatori circadiani endogeni che generano ritmicità biologica in natura vengono continuamente sincronizzati con il periodo di ventiquattro ore che caratterizza la rotazione terrestre, grazie agli effetti di segnali provenienti dall'ambiente, come la luce del giorno e variazioni di temperatura. Tali segnali sono stati denominati dai cronobiologi Zeitgeber, fattori che 'danno' il tempo.
Caratteristiche generali degli orologi circadiani
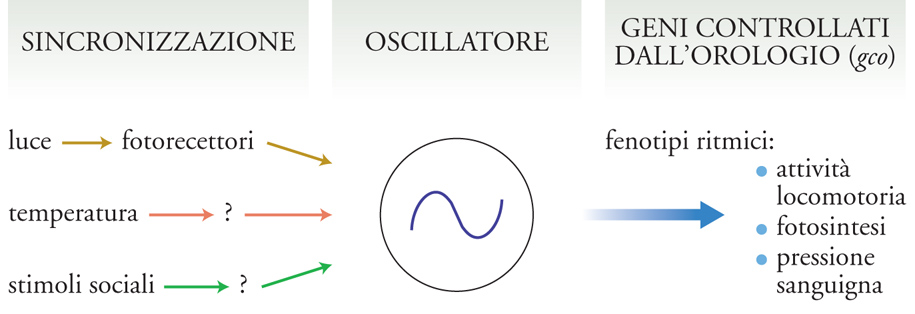
Studi condotti in diversi organismi modello, come il cianobatterio Synechococcus elongatus, il fungo unicellulare Neurospora crassa (la comune muffa del pane), il moscerino della frutta Drosophila melanogaster, la pianta Arabidopsis thaliana e il topo Mus musculus, hanno permesso, nonostante la grande eterogeneità che esiste tra queste specie a livello filogenetico, di identificare alcune proprietà generali che caratterizzano l'organizzazione degli orologi circadiani di questi organismi (fig. 3). In essi, infatti, l'orologio è costituito da un core (l'oscillatore) che rappresenta il meccanismo centrale, cioè l'insieme degli 'ingranaggi' che producono l'oscillazione molecolare circadiana. Questo core è tuttavia permeabile a informazioni provenienti dall'ambiente, i segnali ambientali Zeitgeber, che sincronizzano in continuazione l'oscillazione endogena e la mantengono in fase con le variazioni ambientali prodotte dalla rotazione terrestre, impedendo quindi che l'orologio, e i ritmi biologici da esso controllati, si sfasino determinando conseguenze negative per l'organismo. Infine, l'oscillatore endogeno regola, trasmettendo in uscita le informazioni provenienti dalla sua oscillazione, tutta una serie di fenotipi metabolici, fisiologici e comportamentali. Ciò avviene prevalentemente mediante la regolazione in cascata di numerosi geni implicati nella genesi di questi fenotipi. I meccanismi che caratterizzano la trasduzione delle informazioni ambientali al core dell'orologio (via di ingresso), quelli che operano nel core, e quelli che sono responsabili del controllo, in uscita, dell'attività di numerosi geni controllati dall'orologio (gco) non sono tuttavia da considerare componenti dell'orologio nettamente distinte tra loro e quindi autonome, bensì componenti interdipendenti, capaci di influenzare reciprocamente alcune loro caratteristiche.
Gli orologi biologici circadiani, in tutti gli organismi in cui sono stati identificati e caratterizzati, possiedono alcune proprietà fondamentali: (a) la ritmicità endogena da loro generata e i fenotipi ritmici evocati da quest'ultima persistono anche in assenza di Zeitgeber; (b) gli orologi circadiani sono permeabili a stimoli sincronizzanti provenienti dall'esterno (in natura la luce rappresenta uno dei fattori sincronizzanti più efficaci, ma anche variazioni di temperatura o stimoli sociali possono risultare importanti per la sincronizzazione degli oscillatori endogeni); (c) gli orologi circadiani sono 'temperatura-compensati': infatti, il periodo che caratterizza l'oscillazione endogena non è di fatto influenzato da variazioni di temperatura comprese entro intervalli compatibili con la vita dell'organismo in questione, e ciò anche nel caso di organismi che non sono in grado di mantenere un controllo omeostatico della temperatura del loro corpo, come gli Insetti e i Rettili.
Le conoscenze attuali circa l'organizzazione molecolare e il funzionamento degli orologi circadiani sono il risultato di una straordinaria accelerazione della ricerca in questo settore, che si è prodotta a partire dalla metà degli anni Novanta e che, in pochissimi anni, ha portato la cronobiologia molecolare a conquistare una posizione di grande attualità e di primaria importanza nel panorama delle attività sperimentali che caratterizzano la moderna biologia. L'analisi molecolare dei geni coinvolti nell'organizzazione degli oscillatori circadiani di batteri, funghi, piante e animali ha messo in evidenza come essi non siano omologhi (non derivino da geni ancestrali comuni) e suggerisce quindi che, nel corso dell'evoluzione, alcuni orologi circadiani si siano organizzati più volte e in modo indipendente. Tuttavia, nonostante le diverse collocazioni filogenetiche e le enormi differenze in termini di complessità che caratterizzano le specie finora analizzate, l'organizzazione generale degli oscillatori, da quelli batterici a quelli dei Mammiferi, appare sostanzialmente la stessa.
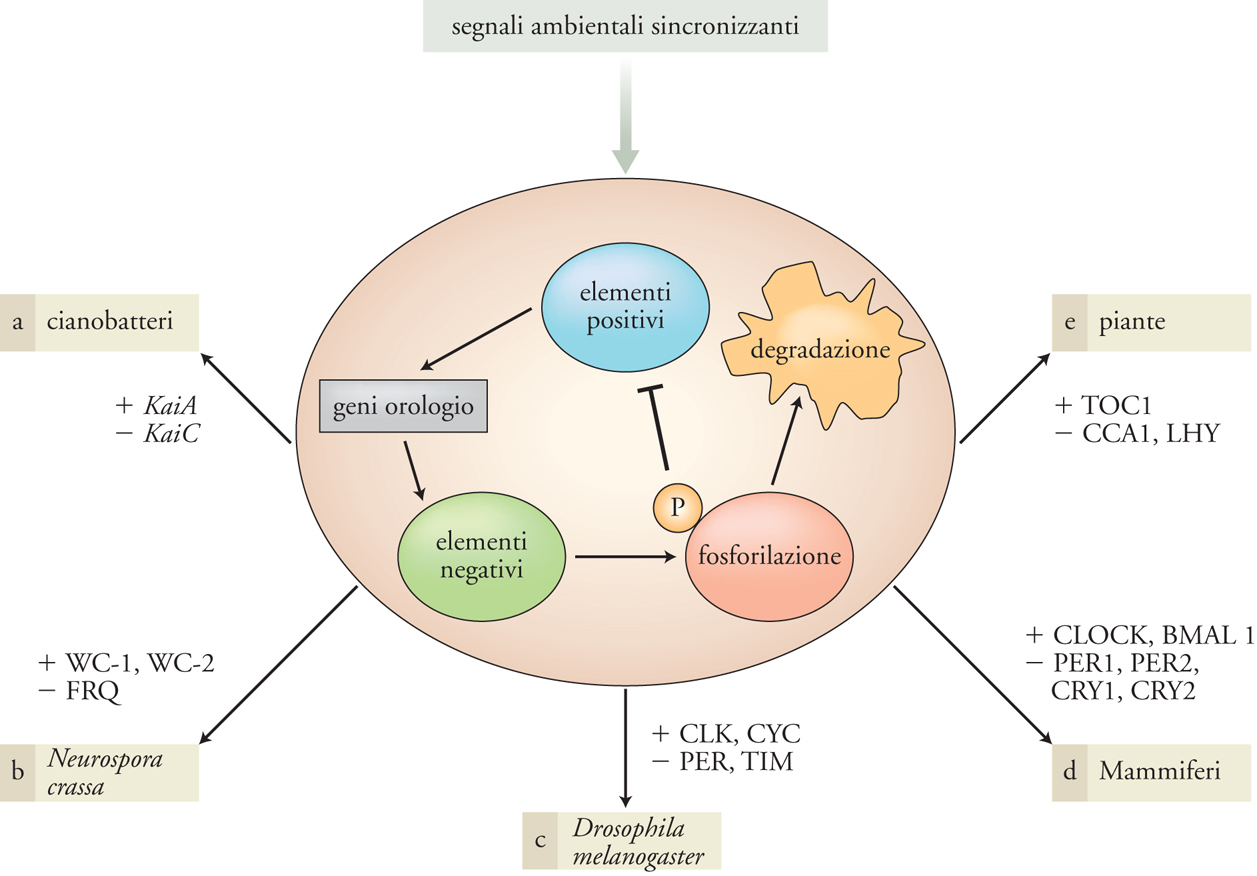
Gli oscillatori endogeni di tutti questi organismi sono infatti costituiti da 'geni orologio' e dalle corrispondenti 'proteine orologio' che possiamo classificare in elementi 'positivi' e 'negativi', sulla base dell'azione che esercitano a livello del core dell'orologio e che formano cicli di autoregolazione mediante fenomeni di retroazione; tali fenomeni generano circuiti temporali caratterizzati da una periodicità di circa ventiquattro ore. Nella fig. 4 è riassunto un modello generale di funzionamento degli orologi circadiani e vengono fornite informazioni sulla natura molecolare degli elementi positivi e negativi identificati finora in diversi organismi modello. La persistenza della ciclicità e della periodicità circadiana a livello molecolare, anche in condizioni ambientali costanti, dipende da modificazioni post-traduzionali a carico di alcune di queste proteine orologio, che introducono nel sistema, a ogni ciclo, un ritardo tra trascrizione, sintesi proteica e retroazione negativa che impedisce al sistema il raggiungimento di condizioni di equilibrio e determina invece condizioni indispensabili per sostenere l'oscillazione nel tempo. Queste modificazioni post-traduzionali sono per lo più dovute all'attività di proteine ad azione enzimatica del tipo delle chinasi, enzimi che catalizzano la fosforilazione di residui amminoacidici specifici nelle proteine bersaglio, e delle fosfatasi, che svolgono invece un'azione opposta, ossia catalizzano la rimozione dei gruppi fosfato. Questi processi di fosforilazione e defosforilazione regolano le dinamiche di stabilizzazione, mobilità intracellulare e degradazione di componenti fondamentali degli orologi biologici circadiani.
L'orologio circadiano in organismi unicellulari
A dispetto della loro apparente semplicità, se confrontati con la complessità degli organismi pluricellulari, gli organismi unicellulari procarioti ed eucarioti per i quali sono oggi disponibili informazioni sull'organizzazione molecolare degli orologi circadiani, rispettivamente il cianobatterio Sinechococcus elongatus e il fungo filamentoso Neurospora crassa, mostrano un'organizzazione dell'orologio circadiano piuttosto complessa. Nel procariote S. elongatus il meccanismo che genera l'oscillazione è basato sull'interazione tra le proteine codificate dai geni orologio KaiA, KaiB e KaiC, il cui assemblaggio ritmico ‒ mediato da fosforilazioni transienti ‒ a formare un complesso a elevato peso molecolare, il periodosoma, genera una ritmicità circadiana. Questa oscillazione regola, in cascata, il funzionamento dell'intero trascrittoma batterico (ossia l'insieme degli RNA messaggeri trascritti da tutti i geni presenti sul cromosoma batterico) e fenomeni quali la divisione cellulare e la collocazione temporale distinta di processi fondamentali, e tuttavia tra loro incompatibili, quali la fotosintesi e la fissazione dell'azoto.
Nel fungo eucariote N. crassa, la ritmicità endogena più importante è generata dall'interazione dei prodotti dei geni orologio frequency (frq), white collar1 (wc1) e white collar2 (wc2), che sono componenti di quello che viene denominato 'oscillatore FWO' (Frequency-white collar oscillator); quest'ultimo svolge un ruolo importante nel controllo circadiano dell'attività di numerosi geni e di un processo fondamentale come quello della maturazione circadiana delle spore asessuali (conidi). Le proteine WHITE COLLAR-1 (WC-1) e WHITE COLLAR-2 (WC-2) funzionano da elementi positivi, che promuovono la trascrizione del gene orologio frq, mentre WC-1 funziona anche da fotorecettore della luce blu e media quindi la sincronizzazione dell'oscillatore circadiano con le informazioni che con continuità provengono dall'ambiente. Molto di recente è stata dimostrata l'esistenza in Neurospora di un altro oscillatore, indipendente dall'azione della proteina orologio FREQUENCY (FRQ) e perciò definito FLO (Frequency less oscillator), che risponde, con meccanismi ancora ignoti, all'azione sincronizzante indotta da variazioni di temperatura o da alcune molecole implicate nella sintesi di steroli quali il farnesolo e il geraniolo, dirigendo l'espressione ritmica di specifici geni. Gli oscillatori FWO e FLO formano un sistema accoppiato, nel quale l'oscillatore FWO svolge un ruolo guida nei processi di sincronizzazione dell'intero organismo con le variazioni ambientali, mentre l'oscillatore FLO concorre, in modo subordinato, a rafforzare la ritmicità biologica circadiana esibita da questo organismo.
L'orologio circadiano nelle piante
Nonostante gli organismi vegetali siano stati tra i primi nei quali sono stati identificati e studiati fenotipi che esibiscono una ritmicità circadiana, le informazioni relative all'organizzazione molecolare dell'oscillatore circadiano dell'organismo modello Arabidopsis thaliana sono piuttosto limitate. Sono stati identificati finora tre geni orologio, LHY (Late elongated hypocotyl), CCA1 (Circadian clock associated) e TOC1 (Timing of cab expression 1), che codificano componenti del core dell'oscillatore circadiano coinvolte nell'organizzazione di un ciclo di autoregolazione a retroazione negativa nel quale le proteine CCA1 e LHY svolgono il ruolo di elementi negativi mentre TOC1, probabilmente in associazione con altre proteine non ancora identificate, attiva la trascrizione dei geni CCA1 e LHY. Sono stati identificati, invece, numerosi geni controllati direttamente dall'orologio, che modulano in uscita, con periodicità circadiana, importanti modificazioni fisiologiche e metaboliche. Infine, sono stati descritti e caratterizzati alcuni fotorecettori, proteine appartenenti alle famiglie dei fitocromi e dei criptocromi, coinvolti nella fototrasduzione del segnale luminoso e nella sincronizzazione dell'orologio circadiano.
L'orologio circadiano negli Insetti
Il primo gene implicato nell'organizzazione di un orologio circadiano, period, è stato identificato nel 1972 nel moscerino della frutta Drosophila melanogaster e successivamente, nel 1985, clonato e caratterizzato dal punto di vista molecolare. Da allora, la dissezione genetica e molecolare dell'architettura degli oscillatori endogeni degli Insetti ha compiuto progressi straordinari e oggi le informazioni disponibili sull'organizzazione e il funzionamento dell'orologio circadiano nell'organismo modello D. melanogaster sono sicuramente le più dettagliate e integrate. Ciò è particolarmente importante dal momento che gli orologi circadiani degli animali, e quindi quelli di Insetti e Mammiferi, sono costruiti in gran parte con gli stessi 'ingranaggi', cioè con proteine omologhe prodotte da geni (omologhi) che si sono evoluti a partire da geni ancestrali comuni. La comprensione del funzionamento dell'orologio di un moscerino è quindi di enorme utilità per indagare, per esempio, il funzionamento dell'orologio circadiano dell'uomo.

L'oscillatore circadiano di Drosophila è costituito da due cicli sovrapposti di autoregolazione, nei quali la trascrizione ritmica di particolari geni orologio è controllata dalla retroazione esercitata dai loro stessi prodotti proteici (figg. I.A, I.B). Inoltre, l'abbondanza delle proteine orologio e la loro localizzazione subcellulare sono controllate da meccanismi di modificazione post-traduzionale (fosforilazioni, defosforilazioni, ecc.) che stabilizzano l'oscillazione su un periodo circadiano e collocano le retroazioni trascrizionali in momenti opportuni nel corso della giornata. In Drosophila sono stati identificati numerosi oscillatori circadiani 'periferici', presenti in diversi distretti tessutali, come nei tubuli malpighiani, nel cardium, nelle antenne, nella proboscide, in aggiunta all'oscillatore 'centrale' costituito da un certo numero di gruppi di neuroni localizzati nel cervello che sono implicati nella genesi di fenotipi ritmici come l'attività locomotoria del moscerino. In Drosophila, a differenza di quanto avviene per esempio nei Mammiferi, gli orologi circadiani periferici sembrano essere sostanzialmente autonomi dall'oscillatore centrale per quanto riguarda la sincronizzazione con l'ambiente esterno, poiché vengono direttamente sincronizzati dalla luce. Nel caso degli Insetti, l'evoluzione avrebbe quindi favorito l'organizzazione di specializzazioni tessuto-specifiche degli oscillatori circadiani. La sincronizzazione dell'orologio a opera della luce è mediata dal fotorecettore per la luce blu CRYPTOCHROME (CRY), una proteina appartenente alla famiglia delle fotoliasi che, attivata dalla luce, forma un complesso con la proteina TIM (fotolabile) e ne avvia la degradazione (fig. I.C). Gli effetti della luce sulla fase della ritmicità endogena, che possono comportare un avanzamento, un ritardo o nessuna variazione, sono in relazione all'abbondanza di TIM e a quella del suo messaggero nel momento in cui, durante le ventiquattro ore, viene fornito lo stimolo luminoso. Impulsi luminosi somministrati nella prima parte della notte si traducono in un ritardo dell'orologio, mentre causano un avanzamento se forniti nella parte terminale della notte. Impulsi luminosi somministrati durante il giorno (quando la proteina TIM è virtualmente assente) non producono alcun effetto.
L'orologio circadiano nei Mammiferi
Nei Mammiferi, a differenza degli Insetti, l'organizzazione degli oscillatori circadiani è gerarchica e consiste di numerosi oscillatori periferici distribuiti in vari organi come il fegato, i polmoni, la milza, il cuore, e di un orologio circadiano centrale, localizzato nel cervello a livello dei nuclei suprachiasmatici dell'ipotalamo, che svolge un ruolo importante di coordinamento delle attività degli oscillatori periferici ed è essenziale per convertire Zeitgeber di natura luminosa, provenienti dal tratto retino-ipotalamico, in segnali in cascata che sincronizzano gli oscillatori periferici (benché non necessariamente tutti con la stessa fase). Si ritiene che ciò avvenga sia mediante proiezioni nervose dirette dai nuclei suprachiasmatici dell'ipotalamo verso altre regioni bersaglio nel cervello, sia mediante segnali di natura umorale.
I geni la cui espressione è controllata dall'orologio centrale dei Mammiferi e dagli oscillatori periferici rappresentano una porzione significativa del genoma e sono in gran parte differenti nei diversi oscillatori. Infatti, analisi molto recenti sulla natura dei trascrittomi in diversi tessuti di Mammiferi hanno messo in evidenza come la frazione di RNA messaggeri che viene trascritta con una cinetica circadiana rappresenti fino al 10% dei messaggeri totali, mentre i trascritti la cui oscillazione avviene in più di un tessuto non raggiungono generalmente l'1% del totale. Queste informazioni, benché indirettamente, suggeriscono che oscillatori periferici diversi contribuiscono nei Mammiferi alla regolazione di fenotipi ritmici tessuto-specifici, che probabilmente riflettono la peculiarità delle funzioni esplicate da organi diversi. Per quanto riguarda la natura dei geni e delle proteine orologio implicate nella genesi della ritmicità circadiana nei Mammiferi, abbiamo già ricordato come la maggior parte delle componenti si siano conservate, nel corso dell'evoluzione, negli animali.
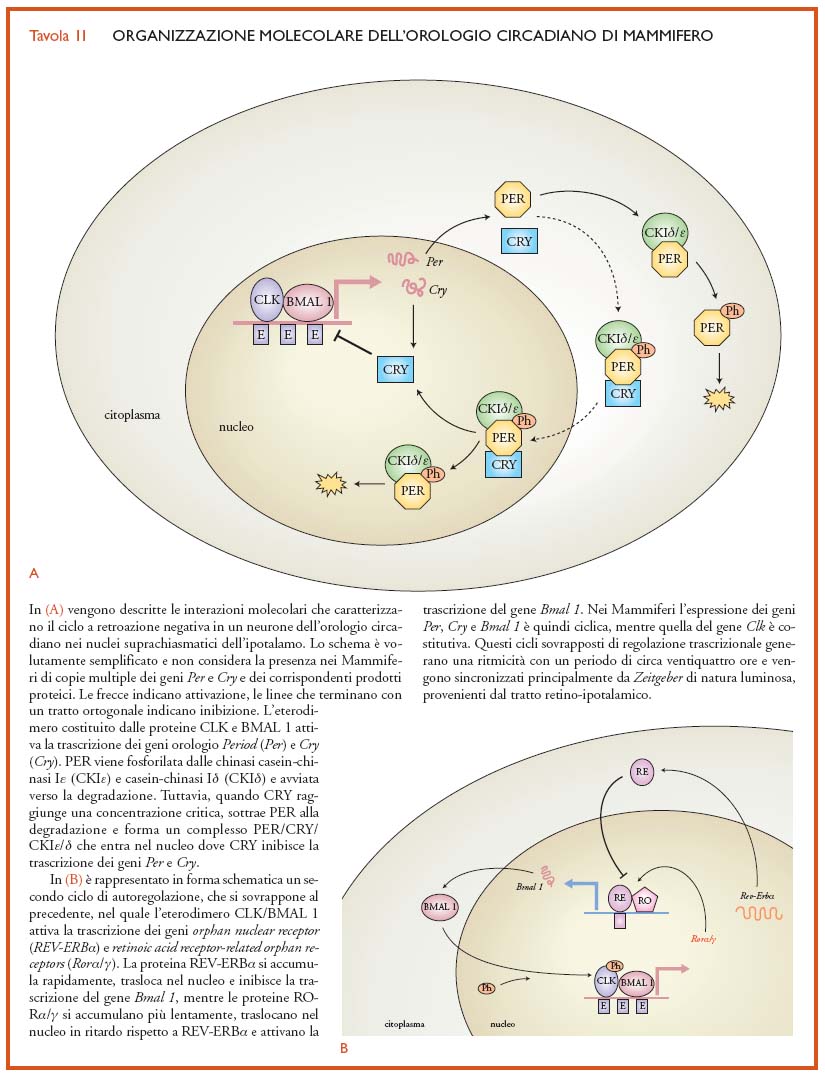
Tuttavia, nei Mammiferi, eventi di duplicazione genica hanno prodotto una ridondanza di informazione genetica, con la conseguente comparsa di copie multiple di numerosi geni orologio (per es., Per1, Per2 e Per3, Cry1 e Cry2). Questo fenomeno ha comportato un aumento di complessità nell'organizzazione degli oscillatori circadiani dei Mammiferi. Inoltre, benché conservate nel corso dell'evoluzione, alcune proteine orologio sembrano avere assunto ruoli diversi all'interno del congegno molecolare che genera l'oscillazione. È il caso della proteina CRY, che nell'oscillatore centrale è il fotorecettore per la luce blu ed è coinvolta nella sincronizzazione dell'orologio, mentre nei Mammiferi stabilizza le proteine PER nel citoplasma (al pari di TIM in Drosophila) formando dei complessi che entrano nel nucleo e regolano, mediante retroazione negativa, i geni orologio Per e Cry. CRY non sembra quindi implicata nella fotorecezione circadiana dei Mammiferi; si ritiene che questo ruolo venga svolto da altri fotopigmenti presenti nella retina. Inoltre, è attualmente ancora in discussione il ruolo del gene Timeless nell'orologio circadiano dei Mammiferi. Nella Tav.II viene descritto un modello di funzionamento dell'orologio dei Mammiferi nel quale, per semplicità, non viene considerata la ridondanza genica.
Alterazioni dell'orologio circadiano nell'uomo
Alterazioni significative della funzionalità dell'orologio circadiano nell'uomo possono determinare alcune cronopatologie. Attività occupazionali che prevedono variazioni sistematiche dei turni di lavoro, come avviene per un consistente numero di individui nelle società industrializzate, possono comportare sfasamenti cronici dell'orologio endogeno rispetto alle variazioni circadiane ambientali e produrre effetti negativi per la salute dei lavoratori coinvolti che, a loro volta, determinano costi economici e sociali importanti. Situazioni di jet lag cronico possono essere sperimentate da piloti d'aereo e personale di bordo, se impegnati con continuità e per lunghi periodi su rotte transmeridianiche. Studi recenti hanno messo in evidenza come in questi casi possano manifestarsi addirittura alterazioni anatomiche a livello del sistema nervoso centrale e deficit cognitivi. Sono noti inoltre alcuni disturbi stagionali dell'umore, caratterizzati da stati depressivi (SAD, Seasonal affective disorder), correlati all'attenuarsi dell'effetto sincronizzante della luce naturale, che accompagna l'avvento dell'autunno e dell'inverno e che diventa particolarmente importante nelle popolazioni che vivono a latitudini elevate.
Infine, la funzionalità dell'orologio circadiano nell'uomo può subire profonde alterazioni a opera di mutazioni che si verificano a livello dei geni orologio responsabili della genesi dell'oscillazione circadiana. È piuttosto recente, infatti, la scoperta che una mutazione a livello di un particolare sito nucleotidico nel gene Per2 determina una variazione nella composizione amminoacidica della proteina corrispondente, PER2, abolendo un importante sito di fosforilazione. Ciò causa profonde modificazioni della cinetica di accumulo e nella successiva traslocazione nel nucleo del complesso PER/CRY, coinvolto nella regolazione per retroazione negativa dell'attività degli stessi geni Per e Cry. Come risultato, i soggetti che portano nel loro genoma questa mutazione autosomica dominante sono affetti da una vera e propria sindrome (FASPS, sindrome familiare di avanzamento della fase del sonno) che altera, accorciandolo, il periodo dell'oscillazione endogena e anticipa al tardo pomeriggio l'inizio della fase del sonno e il risveglio nel mezzo della notte.
Il trattamento delle disfunzioni dell'orologio circadiano si basa oggi in larga misura sulla luce-terapia e sull'adozione di tabelle temporali di attività fisiche e sociali adatte a rafforzare la ritmicità endogena e a sincronizzarla con l'ambiente esterno. Un approccio cronofarmacologico è attualmente in una fase ancora sperimentale e basato su molecole quali la melatonina (un ormone, prodotto dalla ghiandola pineale ‒ epifisi ‒ particolarmente abbondante durante la fase del sonno) e i suoi analoghi. La comprensione dei meccanismi d'azione degli orologi circadiani e dei loro effetti fisiologici e metabolici riveste oggi un'importanza sempre maggiore nei trattamenti farmacologici di molte malattie, per le quali vengono individuati gli intervalli temporali ottimali, nel corso delle ventiquattro ore, per la somministrazione dei farmaci, in modo da aumentare la loro efficacia e, contestualmente, di ridurre al minimo eventuali effetti tossici per l'organismo.
Bibliografia
Antle, Silver 2005: Antle, Michael C. - Silver, Rae, Orchestrating time: arrangements of the brain circadian clock, "Trends in neurosciences", 28, 2005, pp. 145-151.
Bell-Pedersen 2005: Bell-Pedersen, Deborah e altri, Circadian rhythms from multiple oscillators: lessons from diverse organisms, "Nature reviews. Genetics", 6, 2005, pp. 544-556.
Cho 2001: Cho, Kwangwook, Chronic 'jet lag' produces temporal lobe athrophy and spatial cognitive deficits, "Nature neuroscience", 4, 2001, pp. 567-568.
Dunlap 2004: Dunlap, Jay C. - Loros, Jennifer - Decoursey, Patricia, Chronobiology: biological timekeeping, Sunderland (Mass.), Sinauer Associates, 2004.
Dunlap, Loros 2004: Dunlap, Jay C. - Loros, Jennifer J., The Neurospora circadian system, "Journal of biological rhythms", 19, 2004, pp. 414-424.
Hall 2003: Hall, Jeffrey C., Genetics and molecular biology of rhythms in Drosophila and other insects, San Diego-London, Academic Press, 2003.
Hardin 2005: Hardin, Paul E., The circadian timekeeping system of Drosophila, "Current biology", 15, 2005, pp. R714-R722.
Iwasaki, Kondo 2004: Iwasaki, Hideo - Kondo, Takeshi, Circadian timing mechanisms in the prokaryotic clock system of cyanobacteria, "Journal of biological rhythms", 19, 2004, pp. 436-444.
Russell 2005: Russell, Foster G. - Peirson, Stuart - Whitmore, David, Rhythmic temporal processes in biology, in: Encyclopedia of molecular cell biology and molecular medicine, 2. ed., edited by Robert A. Meyers, Weinheim, Wiley-VHC, 2004-2005, 16 v.; v. XII, 2005, pp. 413-482.
Salomé, McClung 2004: Salomé, Patrice A. - McClung, C. Robertson, The Arabidopsis thaliana clock, "Journal of biological rhythms", 19, 2004, pp. 425-435.
Schibler, Naef 2005: Schibler, Ueli - Naef, Felix, Cellular oscillators: rhythmic gene expression and metabolism, "Current opinion in cell biology", 17, 2005, pp. 223-229.
Sernagor 2005: Sernagor, Evelyne, Retinal development: second sight comes first, "Current biology", 15, 2005, pp. R556-R559.
Toh 2001: Toh, Kong L. e altri, An hPer2 phosphorylation site mutation in familial advanced sleep phase syndrome, "Science", 291, 2001, pp. 1040-1043.