
Rappresentazione dello spazio
Rappresentazione dello spazio
Un problema ricorrente nella vita di molte specie animali è il ritrovamento di un luogo specifico, quale un nido o un nascondiglio. La sopravvivenza e la continuazione della specie spesso dipendono dalla soluzione di questo problema. In questo saggio illustreremo alcune delle strategie utilizzate da vari organismi per risolvere tale problema e, in particolare, due diversi meccanismi ubiquitari: l'integrazione del percorso e la memoria spaziale basata su punti di riferimento. Utilizzando l'integrazione del percorso, o navigazione inerziale, l'animale durante il movimento mantiene traccia del vettore che indica la direzione e la distanza rispetto al punto da cui è partito, e ritorna al punto di partenza invertendo il vettore così calcolato. Utilizzando la memoria basata su punti di riferimento, o pilotaggio, la posizione del/' obiettivo viene localizzata mediante le sue relazioni spaziali con i punti di riferimento circostanti. Descriveremo numerosi esperimenti, condotti su formiche, api, uccelli, roditori e uomini, che hanno permesso di scoprire analogie e differenze tra le diverse specie.
L'importanza di ritrovare un luogo
La vespa scavatrice deve il suo nome al fatto che la femmina scava buchi nel terreno nei quali in seguito depone le sue uova. Per nutrire la sua prole la vespa deve affrontare il problema di ritrovare i suoi nidi dopo che si è procurata le prede. L'informazione sulla localizzazione di un nido viene a quanto pare acquisita nel caratteristico volo di ricognizione che la vespa compie non appena lascia il nido. La vespa vola in cerchio descrivendo archi sempre più ampi prima di allontanarsi. La continuità della progenie di una vespa scavatrice dipende dall'abilità di ritrovare il nido.
Un parente stretto della vespa scavatrice, l'ape domestica, è in grado di ritrovare il percorso tra il suo alveare e un gruppo di fiori preferito. Nei climi temperati trovare il cibo, e quindi localizzare con successo il posto dove esso si trova, è cruciale per la sopravvivenza dell'alveare. Soltanto il 25% degli alveari di nuovo impianto riesce a superare l'inverno. L'abilità spaziale delle api domestiche è stata molto studiata e il loro uso dei punti di riferimento sarà discusso in seguito.
Molte specie di uccelli conservano il cibo in posti diversi per far fruttare al massimo i brevi periodi di abbondanza. L'immagazzinatore di cibo più efficiente è la nocciolaia di Clark, un uccello delle dimensioni di un pettirosso che vive nell'America Settentrionale, nella parte orientale delle Montagne Rocciose. l semi di pino costituiscono una parte considerevole della dieta di questi uccelli, che immagazzinano molte provviste di semi per sopravvivere durante i mesi invernali freddi e carenti di cibo. Ogni inverno essi devono rintracciare circa 2200÷3000 nascondigli. Da studi sul campo si stima che possono ricordare il 70% dei siti in cui hanno nascosto il cibo. Questo dato è probabilmente una sotto stima in quanto spesso i roditori rubano le loro scorte nascoste. In laboratorio, fino al 90% dei loro tentativi di ricerca vengono condotti nei siti in cui hanno immagazzinato il cibo, anche dopo un ritardo di 30 giorni e dopo che il cibo stesso è stato volutamente rimosso (Balda e Turek, 1984). Gli uccelli ricordano la localizzazione dei loro nascondigli per almeno nove mesi, una capacità che forse li rende i migliori organismi in quanto a memoria spaziale. Tra gli uccelli che hanno la caratteristica di nascondere il cibo vi sono altri membri della famiglia dei corvidi, cui appartiene la nocciolaia di Clark, come le ghiandaie, e alcuni membri della famiglia dei paridi, tra i quali le cince del continente europeo e quelle del continente americano. Sebbene nessuna di queste specie sia così abile come la nocciolaia di Clark, la memoria per la localizzazione del cibo immagazzinato svolge un ruolo importante nella vita di tutti questi uccelli.
Il ritrovamento di un luogo come servomeccanismo
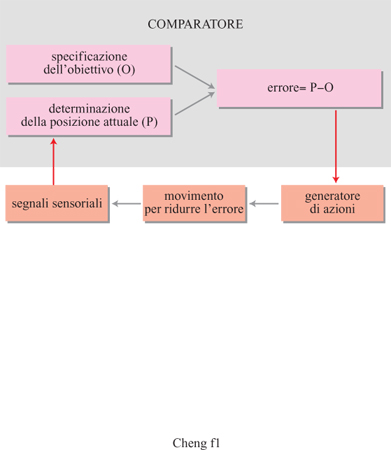
Affinché un organismo sopravviva, molte variabili devono essere mantenute entro intervalli limitati. I servomeccanismi fisiologici aiutano a mantenere entro intervalli accettabili i livelli degli zuccheri nel sangue e la temperatura basale. I meccanismi coinvolti nella ricerca di un sito possono essere concepiti come servomeccanismi comportamentali. Un servomeccanismo rintraccia gli errori e li riduce. In pratica è un comparatore che confronta lo standard ottimale di una variabile con una lettura della misura effettiva di questa variabile (fig. 1). La differenza tra la lettura effettiva e lo standard costituisce un segnale di errore. I segnali di errore guidano il servomeccanismo a generare azioni che riducono l'errore, completando così il cerchio di un feedback negativo.
Nel caso della localizzazione di un sito, lo standard è il ritorno al sito obiettivo. In questo saggio verranno discussi due meccanismi ubiquitari, l'integrazione del percorso e la memoria spaziale basata su punti di riferimento. Questi meccanismi sono utilizzati nell'ultimo stadio della navigazione quando un organismo deve ritornare con precisione in un punto esatto. Per approfondimenti sulla navigazione su lunghe distanze si rimanda al saggio di F. Papi sulla navigazione animale e a quello sulla migrazione degli uccelli di E. Gwinner, in questo volume.
Integrazione del percorso
L'integrazione del percorso è nota anche come navigazione inerzia le o metodo della stima della posizione (dead reckoning). Nell'integrazione del percorso il sito obiettivo è il punto di partenza, tipicamente la tana o il nido di un organismo, e i siti sono definiti in termini di un vettore con origine nel punto di partenza. Un vettore è la classica grandezza algebrica composta di una direzione, un verso e una distanza. Quello che deve fare un organismo quando utilizza questo meccanismo è mantenere, durante il viaggio di andata, la traccia della distanza e della direzione rispetto al punto da cui è partito. Deve fare una vera e propria somma, utilizzando il calcolo vettoriale, di tutte le curve e le svolte che compie durante il percorso. In termini matematici, la continua addizione di minuscole porzioni di tragitto è l'operazione di integrazione, o integrale, da cui il nome del meccanismo. Un'altra similitudine utile è quella di un filo tenuto teso e attaccato al punto di partenza (Wehner e Wehner, 1990). Dovunque l'organismo si sposti, quel filo, computato nel cervello, punta diritto al punto di partenza. Il vettore calcolato (vettore rotta) specifica il punto in cui si trova rispetto al punto di partenza. Quest'ultimo è rappresentato dal vettore 0, per cui il vettore rotta costituisce l'errore. Quando è il momento di dirigersi verso il nido, l'organismo si muove nella direzione opposta al vettore rotta calcolato frnché questo si riduce a 0. Tale processo non utilizza punti di riferimento per localizzare un sito e ciò costituisce un limite. Infatti, gli errori non sistematici si accumulano quanto più lungo è il tragitto e, se non viene calcolato il punto in cui l'individuo si trova rispetto a dei punti di riferimento, gli errori non vengono corretti. Quando gli errori sono troppo grandi, il meccanismo diventa inutile. Nonostante ciò l'integrazione del percorso è un metodo ubiquitario ed è estremamente utile quando mancano punti di riferimento o quando un individuo si trova in un territorio nuovo e le relazioni tra punti di riferimento e obiettivo sono ignote.
Memoria spaziale basata su punti di riferimento
La memoria spaziale basata su punti di riferimento è nota anche come pilotaggio o mnemotassi (il prefisso rnnemosta per memoria, in questo caso memoria di punti di riferimento). Il sito obiettivo è definito rispetto alle sue relazioni spaziali con i punti di riferimento circostanti. Differenti organismi definiscono differenti relazioni spaziali, come sarà discusso più avanti. Anche la posizione di un individuo in un dato momento è definita rispetto ai punti di riferimento. L'organismo si sposta in modo da ridurre la discrepanza tra come i punti di riferimento appaiono in un dato momento e come dovrebbero apparire nel sito obiettivo. Questo meccanismo non è soggetto a errori cumulativi e 'fare il punto' basandosi sui punti di riferimento spesso aiuta un organismo ad aggiornare i calcoli basati sull'integrazione del percorso. In tal modo il pilotaggio e l'integrazione del percorso sono utilizzati spesso in tandem.
Integrazione del percorso nelle formiche del deserto
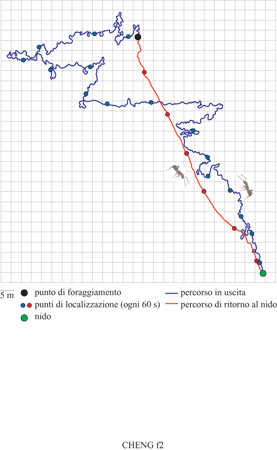
Nel torrido e arido deserto della Tunisia vive la formica Cataglyphis fortis, un minuscolo organismo della massa di 12 mg che può essere considerata il campione dell'integrazione del percorso. Il nido della formica è un piccolo buco nel terreno visibile a malapena in una pianura in gran parte priva di caratteristiche particolari. Il caldo implacabile costituisce un buon motivo per un pronto ritorno al nido. Non sorprende dunque che l'integrazione del percorso sia un meccanismo ben sviluppato in questo insetto. R. Wehner e i suoi colleghi dell'Università di Zurigo hanno studiato questa specie in dettaglio. Nei viaggi di foraggiamento l'operaia può vagare in cerca di cibo effettuando tragitti curvilinei. Tuttavia, essa torna al nido seguendo un tragitto più o meno lineare simile a quello dell'ape. La figura (fig. 2) mostra un esempio spettacolare in cui una foraggiatrice percorse un tragitto sinuoso di 592 m prima di trovare un boccone di cibo, mentre nel suo viaggio di ritorno coprì un percorso di 140,2 m, pari solamente a 1,06 volte la distanza lineare.
La formica può utilizzare anche dei punti di riferimento qualora essi vengano posti artificialmente vicino al suo nido (Wehner et al., 1996), ma la modalità principale resta l'integrazione del percorso. Ciò è stato verificato con un esperimento nel quale alla formica fu messo a disposizione un sito di alimentazione artificiale a una particolare distanza e direzione dal nido (Wehner e Srinivasan, 1981). Dopo il raggiungimento del sito di alimentazione l'operaia fu catturata in una scatola nera e trasferita in un altro luogo. Se la formica si fosse basata su punti di riferimento per dirigersi verso il nido, avrebbe dovuto orientarsi verso il nido effettivo. Essa non si comportò in tal modo, e si diresse nella direzione che l'avrebbe portata al nido se non fosse stata spostata. Essa viaggiò per una distanza simile a quella che l'avrebbe portata al nido e, non avendo lo trovato, effettuò una netta virata che indicava l'inizio di una ricerca sistematica del nido. L'integrazione del percorso continuò durante questa ricerca che consisteva nel descrivere un pattern a forma di spirale verso l'esterno occasionalmente interrotta da un ritorno all'inizio della spirale (Müller e Wehner, 1994) con giri via via più ampi. Per ritornare al punto di inizio della ricerca la formica doveva conservare il tracciato del proprio percorso. Di nuovo non utilizzò punti di riferimento locali. Durante la ricerca sistematica la formica poteva essere catturata e spostata un'altra volta (Wehner e Srinivasan, 1981). Il centro della ricerca veniva il tal caso spostato del vettore da cui la formica era stata dislocata. L'insetto avrebbe potuto proseguire in tal modo fino a morire di caldo nel giro di un paio di ore.
Indicazione della direzione
Per integrare un percorso un organismo deve mantenere la traccia della direzione e della distanza. Negli imenotteri come le formiche e le api il meccanismo con cui viene indicata la direzione è ben noto. Essi utilizzano una bussola solare, che indica la direzione grazie a due proprietà del Sole per loro visibili. Una è il Sole stesso. L'altra è la luce polarizzata creata dalla dispersione della luce solare da parte delle molecole presenti nell'aria (Wehner e Wehner, 1990; Wehner et al., 1996). La specie umana non è sensibile alla luce polarizzata, ma molti insetti e uccelli lo sono. Perciò per questi organismi, anche quando le nubi coprono il Sole, un pezzo di cielo azzurro è sufficiente per indicare la direzione del Sole.
La direzione del Sole non è l'unica informazione necessaria. Dato che il Sole cambia la sua posizione nel cielo col trascorrere del tempo ne II 'arco del giorno, è necessario tener conto di queste variazioni. Per far ciò gli organismi devono essere dotati di un orologio che registra l'ora del giorno, detto orologio circadiano. Gran parte degli organismi multicellulari possiede orologi di questo tipo (v. il saggio di G. Tosini e M. Menaker, Orologi biologici).
Indicazione della distanza
In origine si pensava che gli insetti misurassero la distanza tramite la quantità di energia che avevano speso durante il tragitto. Questa ipotesi è stata recentemente scartata. B. Ronacher e Wehner (1995) hanno scoperto che in C. fortis la dimensione del pezzo di cibo da trasportare al nido non influenza la distanza percorsa nel viaggio di ritorno, un dato che smentisce l'ipotesi energetica. Funzionalmente l'energia spesa non è una misura affidabile della distanza, perché il carico trasportato potrebbe differire da un viaggio all'altro. In altri esperimenti fu posto un nastro mobile sotto le formiche in viaggio verso il nido. Le formiche viaggiavano su una superficie plastica stazionaria, con un pattern visivo che si spostava sotto di loro. In confronto a pattern stazionari, un pattern che si muoveva nella stessa direzione della formica rendeva il viaggio più lungo. Un pattern che si spostava nella direzione opposta faceva viaggiare la formica di meno. Così, una misura di distanza che gli insetti utilizzano è il flusso visivo. Quando le foraggiatrici si spostano, i pattern sul terreno sembrano muoversi all'indietro sotto di loro e questo flusso visivo può essere utilizzato per calcolare la distanza percorsa. Tuttavia, questo fattore non spiega completamente la prestazione della formica e così Ronacher e Wehner ipotizzarono che anche alcuni segnali cinestetici, generati dai movimenti attivi dei muscoli, contribuissero alla misura della distanza.
H.E. Esch e J.E. Bums (1995) effettuarono un ingegnoso esperimento utilizzando le api domestiche per scartare l'ipotesi energetica. Le api furono allenate a trovare il cibo attaccato a un pallone con elio. Gradualmente gli sperimentatori sollevavano il pallone in aria. A mano a mano che questo saliva, le api dovevano viaggiare più a lungo per raggiungerlo. Tuttavia, sorprendentemente, le foraggiatrici, tramite la loro danza, comunicarono alle loro compagne di alveare che la distanza era più breve. Questo risultato paradossale può essere spiegato assumendo che le api misurino la distanza percorsa mediante la quantità di flusso visivo che scorre al di sotto di loro mentre volano. Più in alto volano, minore è il flusso visivo e di conseguenza minore è la distanza calcolata.
Integrazione del percorso nei criceti
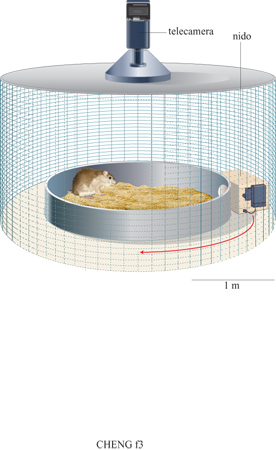
Il criceto dorato (Mesocricetus auratus) ripone il cibo in nascondigli. Questo comportamento è stato sfruttato in esperimenti di laboratorio sull'integrazione del percorso effettuati da A. Etienne dell'Università di Ginevra. Il soggetto sperimentale viveva in un nido al margine di un'arena circolare (fig. 3). Nell'esperimento principale i criceti venivano attirati nell' arena per raccogliere una certa quantità di cibo da nascondere e veniva poi misurata la direzione percorsa dal criceto per tornare al nido (Etienne et al., 1998). In questi esperimenti è stato fatto il possibile per evitare che i criceti utilizzassero strategie diverse dall'integrazione del percorso. Gli esperimenti furono condotti al buio con una telecamera a raggi infrarossi che registrava i percorsi effettuati dagli animali, fu rimossa la sabbia sul pavimento per eliminare i segnali odorosi sul substrato, e fu emesso un suono costante per eliminare possibili segnali uditivi. La scatola del nido veniva ruotata mentre il criceto si trovava nell'arena per eliminare segnali provenienti dall'obiettivo stesso. Nonostante il controllo di tutte queste possibili fonti di informazione, il criceto che trovava il cibo nel centro dell'arena si dirigeva più o meno verso la direzione del nido (se non era stato spostato) o verso la direzione in cui il nido si trovava prima di essere spostato (se questo veniva spostato durante una prova).
Il tracciamento della direzione
I roditori, essendo attivi di notte, ovviamente non utilizzano il Sole come segnale indicatore della direzione. Essi utilizzano due fonti di informazioni. Una è costituita dal loro sistema vestibolare nell'orecchio interno. Questo sistema consiste di canali semicircolari pieni di liquido. La rotazione, sia essa attiva o passiva, sposta il liquido nei canali facendoli collidere contro ciglia sensoriali. L'altra fonte di informazione proviene dal movimento stesso dell'animale. I movimenti muscolari e le informazioni cinestetiche provenienti dai muscoli possono entrambi essere registrati per conservare la traccia del percorso che viene coperto. Conservare la traccia quando si effettuano curve è un'impresa che presenta dei limiti poiché gli errori si accumulano senza possibilità di correzione. Quando gli errori, sono abbastanza grandi, il sistema di integrazione del percorso diviene non funzionale. Gli esperimenti condotti nellaboratorio di Etienne mostrano che dopo tre rotazioni passive di 360⁰ o cinque rotazioni attive il criceto sceglie la direzione giusta solo in maniera casuale.
Sperimentalmente la rotazione passiva veniva ottenuta ruotando il criceto su una piattaforma mentre raccoglieva cibo nel centro di un'arena per nasconderlo. La rotazione attiva veniva ottenuta trascinando di fronte al criceto un pezzo di cibo appeso a una corda. Il cibo girava in circolo inducendo il criceto a girarsi per seguirlo.
Il tracciamento della distanza
Esperimenti condotti da Etienne e collaboratori mostrano che i criceti sono in grado di mantenere la traccia della distanza coperta col loro movimento attivo (Séguinot et al., 1993). Non sono richiesti segnali visivi dato che sono in grado di compiere queste prestazioni nell' oscurità. Nelle giovani oche, invece, è stato mostrato che il flusso visivo costituisce un modo per conservare il tracciato della distanza (von St. Paul, 1982). Le oche subirono dapprima un imprinting su uno sperimentatore (v. il saggio di J.J. Bolhuis, in questo volume). Esse furono poi allenate a ritrovarlo in casa ogni volta che questo compariva esprimendo così un comportamento di homing (ritorno a casa). Successivamente furono trasportate via da casa o con un movimento attivo o passivamente su un carretto. Fintantoché le oche potevano vedere gli oggetti che scorrevano fuori dal carretto, esse mantenevano la traccia sia di quanto erano state spostate (cosa che richiede una stima della distanza), sia di quanto erano state ruotate. L'informazione visiva non era necessaria per mantenere la traccia della rotazione: anche in un carretto completamente coperto le oche mantennero la traccia della rotazione, utilizzando presumibilmente il loro sistema vestibolare.
Errori sistematici nell'integrazione del percorso
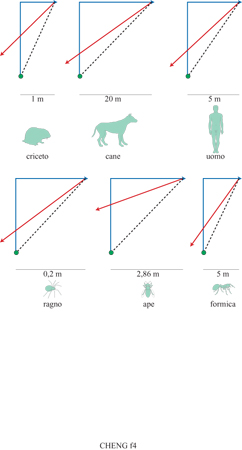
Gli errori sistematici sono quelli che influiscono sulla risposta media facendola deviare in una certa direzione rispetto al valore corretto. Mentre gli errori non sistematici determinano con uguale probabilità uno scarto in eccesso o in difetto rispetto al valore corretto, quelli sistematici si verificano con maggiore probabilità in una direzione piuttosto che nell'altra. Ciò significa che, in media, essi sono compiuti in una certa direzione; viceversa, la media di molti valori affetti da errori non sistematici porta a un valore che si avvicina a quello corretto. In precedenza abbiamo discusso degli errori non sistematici, il cui accumulo rende alla fine l'integrazione del percorso non funzionale. In una vasta gamma di organismi, che vanno dalle formiche alla specie umana, nell'integrazione del percorso si riscontra anche un errore sistematico (fig. 4). L'errore si verifica quando l'animale che sta effettuando l'integrazione di un percorso viene forzato a compiere una virata netta prima di intraprendere il percorso di ritorno verso il nido. In questi casi l'individuo, nel tornare indietro, esegue una virata eccessiva, passando oltre la direzione del nido. Tale errore indica che gli animali non effettuano veri calcoli trigonometrici per risolvere il problema. Veri calcoli trigonometrici per direzione e distanze porterebbero, in media, alla direzione precisa; non si verificherebbe alcun errore sistematico. Possono essere utilizzate alcune scorciatoie aritmetiche, fino a veri e propri calcoli trigonometrici (Wehner et al., 1996; Etienne et al., 1998).
Memoria spaziale basata su punti di riferimento nelle api domestiche
Il premio Nobel K. von Frisch (1953) dimostrò per primo l'utilizzo di punti di riferimento, che consistevano in differenti tipi di colorazioni, nelle api domestiche. I suoi soggetti sperimentali vivevano in un alveare, facente parte di una fila di alveari. La parte frontale di ciascun alveare era colorata in modo differente. L'esperimento di von Frisch consistette nel modificare i colori di tre alveari: quello in cui i suoi soggetti sperimentali vivevano e i due alveari vicini. L'intero pattern di colorazione venne 'spostato' di una unità attribuendo il colore dell'alveare sperimentale a quello alla sua destra o sinistra. In tali condizioni ciascuna ape che usciva dall'alveare sperimentale entrava in quello vicino il cui colore corrispondeva a quello del suo alveare usuale. Dunque le api seguivano il pattern di colorazione della parte frontale dell' alveare ignorando odori e suoni provenienti dal proprio.
Gli studi più sistematici sull'utilizzo di punti di riferimento negli invertebrati provengono da esperimenti sulle api effettuati principalmente da T. Collett dell 'Università del Sussex (Cartwright e Collett, 1982; 1983; Collett e Baron, 1994). In questi studi api libere furono indotte a entrare in una stanza di laboratorio dove trovavano acqua molto zuccherata. Questa ricompensa sperimentale era tale da invogliare le api a tornare ripetutamente nel laboratorio. In esso fu allestita una disposizione sperimentale con punti di riferimento costituiti da tubi cilindrici. La piccola porzione di cibo da trovare era sistemata in un punto costante rispetto alla disposizione. Tuttavia, dopo ciascuna visita l'apparato veniva spostato, senza rotazione, nel laboratorio. Ciò significa che l'obiettivo non si trovava in un punto costante rispetto a qualsiasi punto di riferimento diverso dai tubi. Inoltre, occasionalmente gli individui venivano sottoposti alla prova in assenza dell'acqua zuccherata. Il loro comportamento di ricerca fu videoregistrato e in seguito un'analisi eseguita fotogramma per fotogramma permise di ricavare la quantità di tempo che un individuo aveva trascorso in ciascuna piccola unità di spazio.
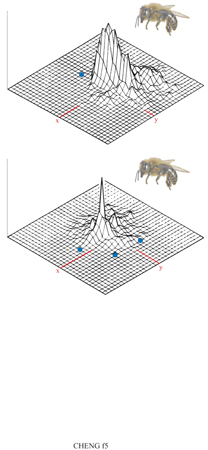
Due di questi tipi di comportamento di ricerca sono presentati nella figura (fig. 5). In entrambi i casi la disposizione con cui le api furono sottoposte alla prova corrispondeva a quella che incontravano durante l'allenamento (eccetto per il fatto che l'acqua zuccherata era assente). Quando furono allenate e sottoposte alla prova con un singolo cilindro come punto di riferimento, la distribuzione di ricerca risultò sparpagliata, ma centrata nel punto in cui il cibo sarebbe stato usualmente trovato. Ciò significa che le api domestiche possono indicare le direzioni della bussola sulla base di qualcosa di diverso rispetto alla disposizione dei punti di riferimento, dato che un singolo cilindro non offre indicazioni di direzione. Infatti, come verrà precisato in seguito, ulteriori esperimenti mostrano che le api utilizzano solo segnali esterni per indicare la direzione e non la disposizione dei punti di riferimento come tale. Con tre tubi cilindrici come punti di riferimento la distribuzione della ricerca risultava più raggruppata intorno all'obiettivo, mostrando il fatto non sorprendente che tre punti di riferimento localizzano un sito con precisione maggiore di uno solo.
Codifica della distanza
Interessanti approfondimenti sui meccanismi della memoria basata su punti di riferimento possono essere ottenuti trasformando il sistema di punti di riferimento. Con una disposizione a un solo cilindro si può variare la dimensione di questo (Cartwright e Collett, 1983). Ingrandire il punto di riferimento fa sì che la ricerca delle api sia effettuata da più lontano, mentre rimpicciolirlo la rende più vicina. Inoltre, le api giudicano la distanza rispetto a un punto di riferimento dalla dimensione (angolo) che un oggetto proietta sugli occhi. Al di fuori degli esperimenti ciò funziona bene dato che gli oggetti non cambiano dimensione 'capricciosamente'. Le api possono anche utilizzare il moto parallasse per giudicare la distanza. Il moto parallasse deriva dal movimento del soggetto. Quando muoviamo la nostra testa da destra a sinistra, la scena visiva appare spostarsi da sinistra a destra. L'ampiezza e la velocità del movimento, tuttavia, dipendono da quanto un oggetto è lontano dai nostri occhi. Oggetti vicini si spostano velocemente e molto, oggetti lontani si spostano lentamente e poco. Questo moto parallasse fornisce dunque un segnale visivo di distanza. Per l'occhio composto di un'ape questi sono gli unici segnali a disposizione per giudicare la distanza.
Mantenimento della traccia della direzione di bussola
Utilizzare la direzione di una bussola pone alcuni problemi. Ogni volta che l'ape gira, la posizione del punto di riferimento esterno rispetto all'insetto stesso cambia. La mappa mentale dei punti di riferimento dovrebbe in un certo senso ruotare con l'ape e questo sembra un processo complicato. Di fatto non è necessaria una rotazione. La soluzione è semplice e forse è evolutivamente la più elegante possibile: le api volano seguendo la stessa direzione quando compiono una ricerca basata su punti di riferimento (Collett e Baron, 1994). In primo luogo esse si avvicinano ai punti di riferimento da utilizzare. In qualche punto vicino all'obiettivo interrompono il volo mentre girano verso la stessa direzione e cominciano una ricerca basata su punti di riferimento. La direzione di bussola è stata ora ridotta alla parte dell'occhio su cui dovrebbe apparire un punto di riferimento.
Un sistema elementare: assenza di codifica delle configurazioni di punti di riferimento
Quando codifichiamo un luogo in termini di punti di riferimento, pensiamo naturalmente a configurazioni di punti di riferimento. Supponiamo che ci venga mostrato un recipiente contenente acqua zuccherata appoggiato su un tavolo. Un cilindro blu si trova sulla parte orientale del tavolo, mentre uno giallo su quella occidentale. Di prova in prova l'intera disposizione di recipiente e cilindri si sposta un poco sul tavolo, ma sempre nella stessa configurazione. Naturalmente pensiamo al cibo come posto tra i cilindri. Se in una prova le posizioni dei cilindri fossero invertite, quello blu a ovest e quello giallo a est, e il recipiente fosse assente, probabilmente penseremmo che esso dovrebbe ancora essere tra i cilindri. In seguito si presenteranno prove a favore di questa asserzione riguardanti la specie umana. Non è così per le api domestiche.
Questo esperimento sulle api domestiche è stato condotto di recente nel mio laboratorio. Le api venivano allenate a cercare il cibo tra due cilindri colorati. La metà delle api aveva il cilindro giallo sempre nella parte a est e quello blu in quella a ovest, mentre l'altra metà aveva il cilindro blu sempre a est e quello giallo a ovest. Nell'esperimento cruciale, in cui le posizioni dei punti di riferimento venivano invertite, le api non cercarono quasi mai nel mezzo. Al contrario, esse cercarono o a ovest del cilindro occidentale o a est di quello orientale. In queste posizioni esse mantennero la corretta direzione di bussola e la corretta distanza da uno dei cilindri colorati.
In generale quando una disposizione sperimentale di punti di riferimento viene ruotata, le api non seguono la rotazione. Esse provano ad appaiare le direzioni di bussola e le distanze dei punti di riferimento individuati con modalità di servomeccanica (Cartwright e Collett, 1982; 1983). Per ciascun punto di riferimento, e i punti di riferimento sono probabilmente definiti dal loro colore, una direzione di bussola e una distanza sono definite come standard. Lo standard specifica come dovrebbe apparire il punto di riferimento una volta raggiunto l'obiettivo. Come indicato in precedenza, la distanza è specificata dalla dimensione retinica proiettata e dal moto parallasse, e la direzione di bussola è equivalente alla parte dell'occhio su cui il punto di riferimento dovrebbe apparire. Quando si compie la ricerca basata su punti di riferimento, un punto di riferimento che appare a sinistra, rispetto a dove dovrebbe essere, induce l'ape a voltare a sinistra, mentre un punto di riferimento che appare a destra, rispetto a dove dovrebbe essere, induce l'ape a girare a destra. Un punto di riferimento che appare troppo vicino induce l'ape ad allontanarsi da esso e un punto di riferimento che appare troppo lontano induce l'ape a spostarsi verso di esso. L'ape fa la media dei dettami di tutti questi calcoli separati e indipendenti. Possiamo pensare che il sistema utilizzi molti servomeccanismi differenti, due per ciascun punto di riferimento, uno basato sulla distanza e uno basato sulla direzione. Ciascuno prova a mantenere solo uno standard: la distanza o la direzione giuste (ma non entrambe) rispetto a un punto di riferimento. E il totale è semplicemente la somma delle parti indipendenti. In questo sistema non vi è nulla che codifica qualcosa riguardante due o più punti di riferimento presi insieme (cioè una configurazione di punti di riferimento). Tale sistema è detto elementare.
Memoria spaziale basata su punti di riferimento nei vertebrati
Anche molti vertebrati utilizzano punti di riferimento per ritrovare un luogo. La lista comprende gli uccelli che fanno provviste di cibo quali le cince bigie americane, le ghiandaie europee, le nocciolaie di Clark; uccelli che non le fanno, quali galli formi e piccioni; tra i roditori i topi, i gerbilli, i criceti e i ratti; tra i primati le scimmie, gli scimpanzé e la specie umana (Cheng e Spetch, 1998). La lista è certamente molto più lunga. Come le api domestiche, i vertebrati utilizzano le proprietà metriche di distanza e direzione. Diversamente dalle api, le specie di vertebrati studiate in dettaglio (ratti, gerbilli, piccioni e uomini) utilizzano talvolta la configurazione dei punti di riferimento.
Analogie e differenze tra colombo e uomo
Il paragone tra la specie umana e i colombi nasce in gran parte da una collaborazione tra M. Spetch dell'Università di Alberta e l'autore di questo saggio dell'Università di Toronto (Spetch et al., 1997). Abbiamo cominciato a studiare i colombi includendo poi la specie umana per effettuare un confronto. Sono stati compiuti dapprima esperimenti in cui i colombi cercavano il cibo sotterrato in un'arena coperta con trucioli di legno. Tale esperimento è stato denominato la prova di campo. I colombi domestici furono dapprima allenati a camminare nell'arena per cercare un po' di granaglie o pezzi di cibo visibili. Il cibo poteva essere all'interno di un tappo di bottiglia oppure in un buco esposto nel piano dell' arena. Gradualmente nel corso delle prove fu coperto sempre più cibo finché i trucioli di legno non coprirono uniformemente tutto il cibo. Furono disposti punti di riferimento che i colombi potessero utilizzare. Essi potevano essere le pareti dell' arena, bottiglie o blocchi. Dopo che gli uccelli avevano imparato a trovare il cibo sotto i trucioli furono effettuati dei test occasionali. Come nell'esperimento effettuato con le api, in queste prove il cibo e il suo contenitore erano assenti. In alcuni casi l'intera arena fu ruotata in modo che l'area di ricerca non presentasse alcun odore o altri segnali lasciati durante le ricerche in prove con ricompense. Ciascun punto di riferimento nell'arena fu riordinato in modo che in tali prove apparisse come se fosse nella disposizione della prova di allenamento. Una telecamera registrava dall'alto il comportamento di ricerca e dall'analisi fotogramma per fotogramma sono state ricavate le distribuzioni di ricerca.
Nelle prove in cui i punti di riferimento erano disposti come nelle condizioni di allenamento gli uccelli hanno cercato il cibo soprattutto dove veniva normalmente trovato. Quando i punti di riferimento furono spostati, anche il punto di massima attività di ricerca da parte degli uccelli si spostava mostrando così, in conclusione, che i soggetti stavano utilizzando punti di riferimento per guidare il loro comportamento di ricerca.
Gli uccelli possono anche essere addestrati a cercare sulla superficie del monitor di un computer. Sebbene i colombi siano stati sperimentati in prove del genere in studi precedenti, l'esperimento pionieristico fu effettuato nellaboratorio di M.L. Spetch. Il monitor veniva coperto con un pannello di materiale plastico per rilevare i colpi dati dalle beccate del colombo ed era circondato da una cornice sensibile al tocco. Nell'esperimento, raggi infrarossi si incrociavano verticalmente e orizzontalmente. Quando qualcosa toccava lo schermo, si interponeva ai raggi interrompendoli. Il momento e il punto del tocco venivano registrati e immagazzinati nel computer. Tale apparato è detto touch-screen (schermi per computer sensibili al tocco) e l'esperimento prova del touch-screen.
Nell'esperimento, al colombo viene presentato dapprima un oggetto obiettivo sullo schermo, seguito subito dopo da una ricompensa in cibo. Un tipico obiettivo è costituito da un quadrato di 2 cm che appare sullo schermo. n cibo può consistere in una tavoletta di cibo che scivola in un contenitore o in un recipiente di granaglie sollevato dal basso, in modo che l'uccello abbia accesso al cibo per pochi secondi. In tali condizioni esso impara ad anticipare l'ottenimento del cibo quando appare l'oggetto e inizia a beccare quest'ultimo prima che arrivi il cibo, un processo noto come autoshaping (automodellamento). In seguito possiamo disporre i punti di riferimento intorno all'obiettivo richiedendo che l'uccello becchi l'obiettivo per ottenere il cibo. L'obiettivo è localizzato in un punto costante rispetto ai punti di riferimento. Tuttavia, l'intera disposizione è spostata intorno allo schermo da prova a prova forzando così l'uccello a effettuare la localizzazione utilizzando la disposizione obiettivopunti di riferimento. In seguito, con un processo detto fading (affievolimento), nel corso delle prove l'obiettivo viene reso sempre meno visibile fino a sparire del tutto. Il colombo è a questo punto allenato a beccare per il cibo in un punto non marcato, ossia in un sito definito solamente dalle sue relazioni spaziali con punti di riferimento disegnati dal computer sullo schermo.
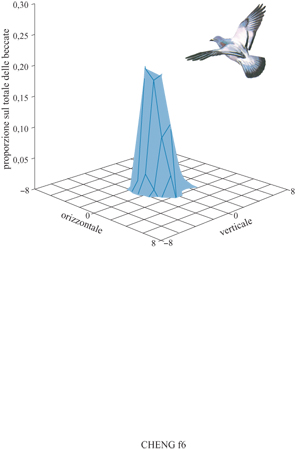
Dopo che l'uccello è stato allenato possono essere effettuate prove senza ricompensa. In tali prove appare sullo schermo una disposizione di punti di riferimento, ma la consegna del cibo è sospesa. l punti in cui il colombo becca sono registrati sul computer, risparmiando il lavoro dell'estrazione video. Un esempio di una distribuzione di ricerca in una prova del touch-screen è riportato nella figura (fig. 6). La distribuzione fu ottenuta con una disposizione di punti di riferimento corrispondente a quella che gli uccelli incontravano durante l'allenamento. Distribuzioni di ricerca simili si ottengono con la prova di campo.
La prova di campo e la prova del touch-screen differiscono per molti aspetti. Il touch-screen è verticale e più piccolo mentre il piano (prova di campo) è orizzontale e su di esso l'uccello può camminare nello spazio. Nella prova di campo il cibo è nel sito obiettivo e i punti di riferimento sono in genere sparpagliati sul piano e distanti. Nella prova del touchscreen il cibo viene fornito a una certa distanza dal sito che costituisce l'obiettivo, i punti di riferimento sono molto più vicini (all'obiettivo e tra di loro) e l'uccello può vedere l'intera disposizione a prima vista. Tuttavia, nonostante tutte queste differenze, nelle due prove si ottengono essenzialmente gli stessi risultati.
Abbiamo effettuato esperimenti analoghi anche su studenti universitari (Spetch et al., 1997). Di nuovo furono presentati i due tipi di prove. In quella del touch-screen gli studenti toccavano lo schermo con una gomma da matita e ottenevano punti per trovare l'obiettivo. Il campo per la specie umana consisteva in spazi sulla superficie di un tavolo e in un caso uno spazio erboso all'aperto. In queste prove gli studenti cercavano un oggetto nascosto. Anche per la specie umana i due metodi producevano lo stesso tipo di risultato.
Utilizzando le strategie trasformazionali descritte precedentemente in alcuni esperimenti, si è passati poi a modificare la disposizione dei punti di riferimento per esaminare quali segnali utilizzassero le due specie. Una manipolazione informativa consisteva nell'allargare la disposizione dei punti di riferimento spostando i punti di riferimento allontanandoli l'uno dall'altro. In una versione di questi esperimenti quattro punti di riferimento che apparivano identici furono collocati in una disposizione quadrata. L'obiettivo si trovava al centro della disposizione. Le due versioni di questo esperimento, la prova di campo e quella del touchscreen, furono effettuate su entrambe le specie. Quando la disposizione fu allargata, gli individui della specie umana continuarono a cercare al centro della disposizione. Gli studenti ci raccontarono che nell'effettuare la prova utilizzavano una sorta di regola di centro. Essi codificavano l'obiettivo come se fosse al centro della disposizione. Nella stessa situazione, invece, i colombi non effettuavano la ricerca al centro della disposizione. Essi cercavano in un sito che fosse alla giusta distanza e direzione da uno dei punti di riferimento. Per esempio, quando i punti di riferimento erano spostati 25 cm a NO, NE, SO e SE, un uccello avrebbe potuto effettuare la ricerca 25 cm a SE del punto di riferimento posto a NO. Uccelli differenti sceglievano punti di riferimento differenti, ma ciascun individuo manteneva lo stesso comportamento da un test all'altro.
Si può dunque concludere che nella ricerca basata su punti di riferimento sia i colombi, sia la specie umana utilizzano informazione metrica, ossia distanze e direzioni, ma nel fare questo si servono di regole differenti. Nella specie umana, la ricerca del sito condotta dopo che si era allargata la disposizione dei punti di riferimento corrispondeva alla situazione di allenamento nelle direzioni dei punti di riferimento, ma non nella distanza. La ricerca del sito nei colombi, al contrario, corrispondeva per distanza e direzione, ma solo rispetto a uno dei punti di riferimento.
Utilizzo delle configurazioni di punti di riferimento
l vertebrati talora utilizzano configurazioni di punti di riferimento. Tutte le specie studiate in sufficiente dettaglio si comportano in tal modo e comprendono gerbilli, ratti, colombi e specie umana.
Nell'esperimento precedente gli studenti che raccontarono di avere utilizzato una regola di centro stavano certamente indicando l'utilizzo di una configurazione di punti di riferimento: il centro è definito solo rispetto a una configurazione di un numero di punti di riferimento. Prove più convincenti dei resoconti verbali derivarono da una manipolazione con rimescolamento della configurazione (Spetch et al., 1996). I soggetti umani furono allenati a cercare al centro di una disposizione quadrata di quattro punti di riferimento distinti. In una prova con rimescolamento, le posizioni dei quattro punti di riferimento furono scambiate a caso, pur mantenendo la disposizione a quadrato. In tal modo ciascun punto di riferimento indicava qualche altro sito obiettivo diverso dal centro. Per esempio, quando il punto di riferimento in alto a sinistra veniva spostato nella posizione in alto a destra, indicava un obiettivo alla destra della disposizione. In un esperimento di questo tipo, gli studenti continuarono a cercare al centro della disposizione mostrando che stavano utilizzando la configurazione della disposizione per decidere dove cercare.
Consideriamo di nuovo la reazione dei colombi all'allargamento di una disposizione di quattro punti di riferimento identici con l'obiettivo al centro. Gli uccelli cercarono nel sito suggerito da un particolare punto di riferimento. Per far ciò in modo coerente essi avrebbero dovuto scegliere quel punto di riferimento, ma non potevano fado in base alle sue caratteristiche perché tutti i punti di riferimento apparivano identici. Essi non potevano neppure scegliere il punto di riferimento in base alla sua posizione assoluta nello spazio perché l'intera disposizione veniva cambiata da prova a prova. Potevano scegliere un punto di riferimento solo in base alla sua posizione relativa agli altri. In altre parole essi lo identificavano in base alla sua posizione rispetto alla configurazione dei punti di riferimento.
T.S. Collett e collaboratori (1986) effettuarono esperimenti sui gerbilli. I gerbilli dovevano cercare il cibo in un'arena circolare coperta con trucioli di legno. L'obiettivo era un seme di girasole nascosto. Come negli esperimenti sulle api, alcuni tubi cilindrici fungevano da punti di riferimento. In un esperimento i gerbilli furono allenati a trovare un sito obiettivo definito in relazione a due punti di riferimento distinti. In una prova successiva la disposizione fu ruotata. I gerbilli cercarono nel posto appropriato in accordo alla rotazione della disposizione. Dunque, essi avevano definito delle direzioni rispetto alla disposizione, ma non potevano aver definito le direzioni rispetto a ciascun singolo punto di riferimento in quanto esso era cilindrico, e non dava informazioni direzionali. Pertanto, per definire la direzione dovevano aver usato la configurazione dei punti di riferimento. Va ricordato che le api domestiche, al contrario, non seguono la rotazione di una disposizione di punti di riferimento nella maniera dei gerbilli. Le api provano a mantenere la direzione di bussola rispetto a un sotto gruppo di punti di riferimento. In un altro esperimento che mostrava l'utilizzo della configurazione nei gerbilli, la disposizione di allenamento consisteva di due punti di riferimento identici. Il sito obiettivo si trovava tra i due punti, ma a una certa distanza dalla linea congiungente i due punti di riferimento. Come nei nostri esperimenti sui colombi, in alcune prove i due punti di riferimento furono allontanati. In modo analogo ai colombi, i gerbilli effettuarono la ricerca basandosi su uno dei punti di riferimento. Applicando la stessa logica, si può concludere anche in questo caso che essi devono identificare il punto di riferimento scelto in base alle relazioni spaziali con l'altro. Dunque devono codificare la configurazione di due punti di riferimento.
Modulo geometrico nei ratti e nei bambini
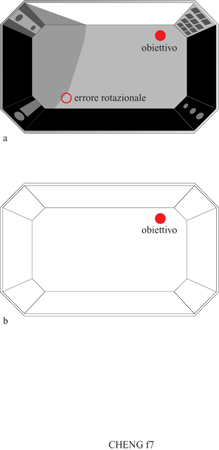
La prova che i ratti usano la configurazione proviene dall'osservazione di un errore sistematico che essi commettono (fig. 7). In un esperimento un ratto aveva il compito di ricordare un sito in cui a esso erano stati mostrati alcuni cereali dolci (un cibo per esso molto prelibato). L'arena di ricerca era rettangolare e conteneva molte caratteristiche che potevano fungere da segnali. Una delle due pareti lunghe era bianca mentre le altre tre pareti erano nere. Negli angoli vi erano pannelli rivestiti con tessuti di differenti trame, caratterizzati da differenti pattern visivi e da luci diverse che riflettevano da essi. Odori di anice e menta peperita venivano emanati da due angoli. Il sito obiettivo differiva da un giorno all'altro cosicché il ratto doveva ogni giorno apprendere un sito nuovo. In una prova un ratto fu messo nell'arena con il cibo esposto su un piattino. Gli fu concesso di mangiare tre tavolette, dopo di che il cibo fu rimosso. Mentre il ratto era fuori dell'arena, il cibo fu sotterrato nello stesso sito sotto i trucioli di legno che coprivano il piano dell' arena. Dopo un ritardo di 90 s, il compito del ratto era trovare il cibo sotterrato di nuovo nello stesso posto.
l ratti talora scavavano nel sito corretto per cercare il cibo, ma altre volte commettevano un notevole errore (Cheng e Spetch, 1998). Essi, in particolare, scavavano nel sito indicato nella figura 7 come errore rotazionale. Rispetto al sito corretto, questo sito è ruotato di 180⁰ attraverso il centro dell'arena. Essi commisero questo errore con una frequenza pari quasi a quella della scelta corretta. L'errore non è peculiare dei ratti. Bambini di due anni commettono lo stesso errore. Per spiegarlo dobbiamo supporre che il ratto codifichi qualche rappresentazione come quella mostrata nella figura 7. Questo modulo, detto modulo geometrico, registra la forma generale dell'ambiente, ma non le caratteristiche discrete di esso, come per esempio, la brillantezza delle pareti, i pattern visivi dei pannelli, gli odori, e così via. È una registrazione di una configurazione pura, di una forma estrapolata dalle sue caratteristiche. Su questa registrazione viene marcata la localizzazione del sito.
Nell'ambiente sperimentale che, a differenza degli ambienti naturali, è perfettamente rettangolare, utilizzare il modulo geometrico determina un' ambiguità. Il ratto ha due modi per far corrispondere la sua mappa geometrica con l'ambiente: l'uno porta a individuare il sito corretto, l'altro lo induce a commettere l'errore rotazionale (v. figura 7). L'esistenza dell' errore rotazionale ci porta a questa conclusione.
Altri esperimenti dimostrano che i ratti possono in realtà discriminare le molte caratteristiche a disposizione nell'ambiente. Nel commettere l'errore rotazionale i ratti e i bambini agiscono in questa sequenza: in primo luogo, essi misurano le giuste distanze e direzioni dalle pareti, cioè utilizzano informazioni metriche nella memoria basata su punti di riferimento; in secondo luogo, utilizzano un'immagine della configurazione pura della forma (modo 'forte' di utilizzare una configurazione).
Misura della distanza
Non è stato studiato sistematicamente il modo in cui i vertebrati misurano le distanze dai punti di riferimento. l sistemi visivi di uccelli e mammiferi possono registrare molti indici per misurare le distanze. Essi comprendono indici monoculari, che richiedono l'uso di un solo occhio, e indici binoculari, che richiedono l'uso di entrambi gli occhi. Gli indici monoculari comprendono l'accomodamento (come i muscoli oculari devono essere utilizzati per mettere a fuoco l'occhio sull'oggetto), il moto parallasse, le trame dei tessuti e altri gradienti, e la dimensione che un oggetto familiare proietta sulla retina. Gli indici binoculari comprendono la convergenza (l'angolo a cui devono trovarsi i due occhi perché entrambi mettano a fuoco un oggetto) e la disparità binoculare. Quest'ultima si basa sul fatto che i due occhi posti frontalmente ricevono visioni leggermente diverse del mondo. Sulla base di queste differenze può essere calcolata la profondità. Una ragionevole ipotesi è che sia utilizzata una combinazione di tutti i sistemi a disposizione.
Conclusioni
In questo saggio abbiamo esaminato l'integrazione del percorso e la memoria spaziale basata su punti di riferimento, trattando entrambi i casi come servomeccanismi. Nell'integrazione del percorso un individuo mantiene la traccia del vettore coperto durante il cammino da un punto di partenza. Il vettore indica la direzione e la distanza rispetto a quest'ultimo. La strategia richiede di mantenere simultaneamente la traccia della distanza e della direzione percorse con continuità. Una vasta gamma di organismi, vertebrati e invertebrati, utilizza l'integrazione del percorso. Nella memoria spaziale basata su punti di riferimento il luogo obiettivo è definito in base alle sue relazioni spaziali con dei punti di riferimento circostanti. Organismi studiati in dettaglio hanno mostrato di utilizzare le proprietà metriche di distanza e direzione per sfruttare i punti di riferimento. l vertebrati studiati utilizzano talora la configurazione dei punti di riferimento, cioè la loro disposizione relativa. Le api domestiche, d'altra parte, sembrano utilizzare i punti di riferimento solo in maniera elementare, ciascuno individualmente e in maniera indipendente per specificare illuogo obiettivo. Non è noto al momento il modo in cui le modalità configurazionale ed elementare nell 'uso dei punti di riferimento sono distribuite tra le specie.
Bibliografia citata
BALDA, R.P., TUREK, R.J. (1984) The cache-recovery system as an example of memory capabilities in Clark's nutcracker. In AnimaI cognition, a c. di Roitblat H.L., Bever T.G., Terrace H.S., Hillsdale, Lawrence Erlbaum Associates, pp. 513-532.
CARTWRIGHT, B.A., COLLETT, T.S. (1982) How honeybees use landmarks to guide their retum to a food source. Nature, 295, 560-564.
CARTWRIGHT, B.A., COLLETT, T.S. (1983) Landmark learning in bees. J. Comp. Physiol. A, 151, 521-543.
CHENG, K., SPETCH, M.L. (1998) Mechanisms of landmark use in mammals and birds. In Spatial representation in animals, a c. di Healy S.D., Oxford, Oxford University Press, pp. 1-17.
COLLETT, T.S., BARoN, J. (1994) Biological compasses and the coordinate frame of landmark memories in honeybees. Nature, 368, 137-140.
COLLETT, T.S., CARTWRIGHT, B.A., SMITH, B.A. (1986) Landmark leaming and visuo-spatial memories in gerbils. J. Comp. Physiol. A, 158, 835-851.
ESCH, H.E., BURNS, J.E. (1995) Honeybees use optic flow to measure the distance of food source. Naturwissenschaft, 82, 38-40.
ETIENNE, A.S., BERLIE, J., GEORGAKOPOULOS, J., MAuRER, R. (1998) Role of dead reckoning in navigation. In Spatial representation in animals, a c. di Healy S.D., Oxford, Oxford University Press, pp. 54-68.
ETIENNE, A.S., JORIS-LAMBERT, S., MAuRER, R., REVERDIN, B., SITBON, S. (1995) Optimizing distallandmarks: horizontal versus vertical structures and relation to background. Behav. Brain Res., 68, 103-116.
FRISCH, K. VON (1953) The dancing bees. San Diego, Harcourt Brace Jovanovich.
MÜLLER, M., WEHNER, R. (1994) The hidden spiraI: systematic search and path integration in desert ants, CatagIyphis lortis. J. Comp. Physiol. A, 175, 525-530.
RONACHER, B., WEHNER, R. (1995) Desert ants CatagIyphislortis use self-induced optic flow to measure distances travelled. J. Comp. Physiol. A, 177, 21-27.
SÉGUlNOT, V., MAURER, R., ETIENNE, A.S. (1993) Dead reckoning in a small mammal: the evaluation of distance. J. Comp. Physiol. A, 173, 103-113.
SPETCH, M.L., CHENG, K, MAcDONALD, S.E. (1996) Learning the configuration of a landmark array: I. Touch-screen studies with pigeons and humans. J. Comp. Psychol., 110, 55-68.
SPETCH, M.L., CHENG, K, MAcDONALD, S.E., LINKENHOKER, B., KELLY, D., DOERKsoN, S. (1997) Use of landmark configuration in pigeons and humans: II. Generality across search tasks. J. Comp. Psychol., 111, 14-24.
SPETCH, M.L., MONDLOCH, M.V. (1993) Control of pigeons' spatial search by graphic landmarks in a touch-screen task. J. Exp. Psychol.: Anim. Behav. Proc., 19, 353-372.
ST. PAUL, U. VON (1982) Do geese use path integration for walking home? In Avian navigation, a c. di Papi F., Wallraff H.G., Berlin, Springer Verlag, pp. 298-307.
WEHNER, R., MICHEL, B., ANTONSEN, P. (1996) Visual navigation in insects: coupling of egocentric information. J. Exp. Biol., 199, 129-140.
WEHNER, R., SRINIVASAN, M.V. (1981) Searching behaviour of desert ants, genus CatagIyphis (Formicidae, Hymenoptera). J. Comp. Physiol., 142, 315-338.
WEHNER, R., WEHNER, S. (1990) Insect navigation: use of maps or Ariadne's thread? Ethol. Ecol. & Evol., 2, 27-48.
Bibliografia generale
CHENG, K Landmark-based spatial memory in the pigeon. In The psychology of learning and motivation, a c. di Medin D.L., voI. 33, New York, Academic Press, 1995, pp. 1-21.
COLLETT, T.S. (1992) Landmark leaming and guidance in insects. Phil. Trans. R. Soc. Lond. B., 337, 295-303.
GALLISTEL, C.R. The organization of Iearning. Cambridge, Mass., MIT Press, 1990.
KAMIL, A.C., BALDA, R.P. In The psychology of Iearning and motivation, a c. di Bower G., voI. 26, New York, Academic Press, 1990, pp. 1-25.