
Allergia
Allergia
Il termine 'allergia' fu proposto, nel 1904, dal viennese Clemens von Pirquet, per indicare una modificazione della capacità di reagire dell'organismo. Fu merito di due medici tedeschi, Otto Carl Prausnitz e Heinz Kustner (1921), la dimostrazione che una sensibilizzazione poteva essere trasferita passivamente da un soggetto allergico a uno sano. Con il termine di 'atopia', Arthur F. Coca e Robert A. Cooke designarono, nel 1923, quel gruppo di affezioni a incidenza familiare caratterizzate da sensibilizzazione nei confronti di sostanze comunemente presenti nell'ambiente. Le malattie allergiche rappresentano un gruppo di affezioni legate a una disregolazione del sistema immunitario nei confronti di particolari antigeni (allergeni) normalmente innocui per la maggior parte dei soggetti. Mentre il termine 'allergia' ha un significato abbastanza esteso, il termine 'atopia' viene utilizzato per indicare più specificamente quel gruppo di affezioni allergiche, caratterizzate dall'esistenza di familiarità e dalla propensione alla produzione di anticorpi (in particolare delle immunoglobuline E) nei confronti di allergeni ambientali. Gli allergeni sono di diversi tipi, e sono classificati in base alla modalità con cui vengono a contatto con il soggetto allergico: attraverso le vie respiratorie (allergie ai pollini e agli animali come cani e gatti), per ingestione (alimenti e farmaci), per iniezione parenterale (farmaci e insetti).
In genere le manifestazioni cliniche dei soggetti atopici sono la dermatite atopica, l'asma bronchiale e la rinite allergica e, meno frequentemente, l'orticaria e le manifestazioni gastrointestinali, che possono comparire in successione nel corso della vita, ma anche essere presenti contemporaneamente nello stesso soggetto, conducendo al concetto di atopia come condizione fisiopatologica a carattere sistemico. Per contro, le reazioni di tipo allergico o anafilattico nei confronti di farmaci o di componenti del veleno di alcuni insetti possono insorgere anche in soggetti senza stigmate di atopia. Infine, dal momento che almeno una parte delle manifestazioni cliniche dell'allergia e dell'atopia è caratterizzata dall'insorgenza a breve distanza di tempo (pochi minuti) dall'introduzione dell'antigene che ne è responsabile, esse sono anche note come 'reazioni di ipersensibilità immediata'. Tuttavia, le reazioni immediate costituiscono, almeno nella maggior parte dei soggetti allergici, solo una componente dell'intero quadro fisiopatologico, potendo essere seguite dalle cosiddette 'reazioni di fase tardiva', le quali contribuiscono in maniera determinante alla genesi delle manifestazioni cliniche e soprattutto alla loro cronicizzazione.
Oggi l'incidenza di queste malattie appare in continuo aumento, e la loro prevalenza è certamente superiore al 10%. La causa di questo marcato incremento, avvenuto soprattutto a partire dal secondo dopoguerra, è ancora ignota. Tuttavia, vi sono diverse ipotesi che sottolineano l'importanza delle condizioni ambientali in cui si sviluppa l'organismo: la minore esposizione a fattori allergenici durante l'infanzia potrebbe essere responsabile (interagendo con fattori genetici) della maggiore reattività allergica tipica dei Paesi occidentali.
Sommario: 1. Gli allergeni. 2. Patogenesi dell'allergia. 3. Ruolo complementare dei fattori genetici e ambientali nello sviluppo dell'allergia. 4. Diagnostica delle malattie allergiche. 5. Cenni di terapia delle malattie allergiche. □ Bibliografia.
Gli allergeni
Gli allergeni sono gli antigeni presenti comunemente nell'ambiente, dotati della proprietà di indurre, in determinati individui, una risposta immunologica prevalentemente caratterizzata dalla produzione di anticorpi IgE (Immunoglobuline E). Tale proprietà non è strettamente correlata con le caratteristiche fisico-chimiche di tali molecole, che spesso sono proteine o glicoproteine resistenti al calore, alle variazioni del pH e agli enzimi proteolitici, e hanno peso molecolare compreso tra 5 e 60 kDa (kiloDalton; un dalton è uguale a 1,6605655×10−27 kg). La maggior parte degli allergeni viene introdotta nell'organismo per via inalatoria (allergeni da inalazione), per ingestione (allergeni alimentari e farmaci) oppure per via parenterale (allergeni da iniezione).
Tra gli allergeni da inalazione sono compresi i pollini, gli acari della polvere domestica, i derivati epidermici di animali (cane, gatto, cavallo, ecc.), alcune spore fungine; in tale tipologia di allergeni rientrano inoltre alcuni composti volatili utilizzati nell'industria (isocianati, fluoruri, parafenilendiamina, ecc.) responsabili di forme allergiche legate all'ambiente di lavoro, nonché alcune proteine vegetali presenti nella farina di grano, nella soia e nel frumento). Gli allergeni alimentari sono più spesso responsabili di disturbi gastrointestinali e della sindrome orticarioide e angioedematosa; più raramente di gravi quadri di shock anafilattico. Infine, gli allergeni da iniezione includono i farmaci e il veleno degli Insetti, soprattutto gli Imenotteri (api, vespe, calabroni).
Pollini. I pollini anemofili, cioè diffusi nell'aria che si respira e trasportati dal vento, rappresentano gli allergeni inalanti più frequentemente responsabili di malattie allergiche a carico dell'apparato respiratorio. Si ritrovano nell'atmosfera in stagioni ben definite, caratteristiche per ciascuna pianta e area geografica. Gli allergeni pollinici, di natura proteica o glicoproteica, vengono liberati al contatto del granulo pollinico con la superficie umida e ricca di enzimi delle mucose respiratorie. I pollini più importanti dal punto di vista allergenico sono quelli delle Graminacee: tra questi i più studiati sono quelli di Lolium perenne, il cui allergene maggiore (Lol p1) e anche i due minori (Lol p2 e Lol p3) mostrano una notevole reattività crociata con i principali allergeni delle altre specie di Graminacee: chi è allergico a queste ultime, è reattivo anche agli allergeni di Lolium. Altri allergeni pollinici importanti in Italia e in Europa sono quelli di Artemisia vulgaris e di Parietaria judaica, mentre negli Stati Uniti oltre alle Graminacee rivestono notevole importanza i pollini di Ambrosia artimisiifolia e di A. trifida. I pollini delle piante ad alto fusto sono meno importanti, sia per la minore frequenza con cui inducono sensibilizzazione, sia perché la stagione di pollinazione è molto più breve di quella delle erbe. Ricordiamo comunque la crescente importanza dei pollini di olivo, betulla e cipresso.
Spore fungine
Le spore fungine si ritrovano in grande quantità nell'ambiente esterno e vengono trasportate da correnti aeree a grandi distanze, per poi depositarsi al suolo, anche in ambienti chiusi, sviluppandosi e dando origine a molte altre spore. L'importanza degli allergeni fungini è minore di quella dei pollini. I più rilevanti tra quelli ritrovabili nell'ambiente esterno sono Alternaria alternata e Cladosporium herbarium, Aspergillus fumigatus, Penicillum niger e Mucor si ritrovano in genere nelle zone più umide delle abitazioni (spugne, contenitori di immondizie, vasi da fiori). Va tenuto presente che i sintomi nei pazienti allergici alle spore fungine, a differenza di quelli dei pazienti con allergia ai pollini, si accentuano nei periodi umidi e piovosi.
Allergeni di derivazione animale
I più importanti sono quelli derivati dalla saliva, dal siero, dalle secrezioni sebacee e dall'epitelio di gatti e cani. L'allergia più notevole è quella legata ai derivati del gatto; si calcola che circa il 15-20% dei pazienti atopici possegga IgE specifiche per un antigene presente nella forfora del gatto, il Fel d1, che deriva principalmente dalle cellule sebacee e si accumula sugli strati superficiali dell'epidermide. Le particelle risultano di dimensioni molto piccole (2,5 μm) e quindi molto diffusibili con le correnti aeree. Per questa caratteristica tale allergene si ritrova anche in abitazioni dove non è presente l'animale. Gli allergeni derivati dal cane, i Can f1, per la diffusione di questo animale costituiscono un'altra importante fonte allergenica. Devono essere infine ricordati gli allergeni derivati dal cavallo per la loro grande potenza allergenica, capace di provocare gravi quadri clinici di asma bronchiale a insorgenza improvvisa; nonché gli allergeni del coniglio, che costituiscono un problema prevalentemente per gli allevatori ma che negli ultimi anni, per la diffusione di questo animale come animale da appartamento, sta diventando un problema clinico non trascurabile. Si deve inoltre ricordare che frequentemente si osserva una reattività crociata, almeno dal punto di vista clinico, tra i diversi animali, per cui soggetti sensibili agli allergeni del cane non tollerano soggiornare in ambienti in cui è presente il gatto e viceversa.
Acari
Gli acari delle polveri domestiche (Dermatophagoides pteronyssinus e farinae) costituiscono la principale sorgente di allergeni responsabili di malattie allergiche ad andamento subcontinuo o cronico. Nei pazienti asmatici la prevalenza della sensibilizzazione agli allergeni degli acari oscilla tra il 45 e l'85%. Gli acari si nutrono di scaglie epiteliali umane, per cui tendono a vivere nei materassi e in generale nei mobili con imbottiture (poltrone, cuscini, ecc.). Liberano allergeni sia in conseguenza della frammentazione dei loro tegumenti, sia attraverso le loro particelle fecali. Poiché l'atmosfera caldo-umida favorisce la crescita degli acari, nei pazienti con questo tipo di allergia si verifica un'accentuazione della sintomatologia durante il periodo dell'anno compreso tra la tarda estate e l'inizio dell'autunno; per contro, per gli stessi motivi, l'allergia agli acari è virtualmente inesistente alle altitudini superiori ai 1600 metri. Sono stati anche descritti altri acari definiti 'minori', ma la loro importanza come sorgente allergenica appare decisamente inferiore in quanto rappresentano circa il 10% della popolazione degli acari. Dobbiamo ricordare Acarus siro, che vive nelle farine e nei cereali, nonché sulla superficie dei formaggi in stagionatura; Tyrophagus putrescentiae, presente soprattutto nei magazzini alimentari e chiamato anche 'acaro del prosciutto crudo'; Glyciphagus domesticus, che predilige prodotti dolciari e farine; infine, Lepidoglyphus destructor, che vive soprattutto nei fienili e nei granai. Sembra esistere un certo grado di reattività crociata con gli allergeni derivati dagli acari maggiori. Del resto, sono abbastanza rari i casi clinici in cui è presente una sensibilizzazione isolata nei confronti degli acari minori.
Allergeni alimentari. Gli alimenti che più frequentemente causano reazioni allergiche sono il latte e i suoi derivati, il frumento, l'uovo, il pesce, la cipolla, l'aglio, la carne di maiale, il pomodoro, le noci e i semi. Il gruppo delle noci e dei semi comprende alimenti che hanno allergeni in comune, per cui è possibile una reattività crociata. In questo gruppo si ritrovano alimenti quali cacao, piselli, fagioli, fave, fagiolini, noci, nocciole, mandorle, arachidi, olive, soia, semi di girasole, lamponi, more, uva, agrumi, ciliegie, pesche, ecc. Inoltre, di grande interesse è la dimostrazione di reattività crociata tra pollini e alimenti (frutta e vegetali). Si ritiene comunemente che gli allergeni di origine vegetale siano sensibili alla temperatura, siano cioè termolabili, contrariamente a quelli di origine animale, che sono termostabili, in quanto la cottura non modifica l'allergenicità del pesce, delle uova o del latte. Tuttavia esistono delle eccezioni, in quanto le albumine di diversi tipi di carne sono termolabili e, per converso, gli allergeni delle arachidi e dei pomodori sono termostabili.
Patogenesi dell'allergia
Anche sul piano storico, una delle caratteristiche fondamentali dei soggetti allergici è quella di produrre anticorpi della classe IgE diretti verso gli allergeni. Le più recenti acquisizioni sul ruolo delle cellule coinvolte nella produzione degli anticorpi IgE e di quelle coinvolte nei processi infiammatori conseguenti (linfociti, mastociti, granulociti eosinofili, granulociti basofili), nonché la scoperta di numerose citochine e chemochine (citochine chemiotattiche), hanno modificato l'interpretazione dei meccanismi patogenetici, soprattutto con riferimento alle malattie allergiche caratterizzate da flogosi persistente con tendenza alla cronicizzazione.
I linfociti Th2
Lo studio delle varie componenti cellulari che partecipano alla risposta immunitaria ha permesso di dimostrare l'esistenza di differenti tipi di linfociti Th (T helper). I linfociti Th sono le cellule deputate, nel sistema immunitario, al riconoscimento di tutti i possibili agenti estranei capaci di penetrare nell'organismo e di dare inizio alle reazioni effettrici devolute alla loro rimozione o eliminazione. Infatti, i linfociti Th hanno una fine capacità discriminatrice garantita dalla presenza sulla loro superficie di un recettore che è specifico per ciascuno dei numerosissimi peptici costituenti le molecole proteiche degli antigeni. Il repertorio di variabilità del recettore dei linfociti T è così ampio da permettere il riconoscimento di tutti i potenziali antigeni. Sono stati distinti almeno due tipi di Th sulla base del loro profilo di produzione di citochine, che condiziona le modalità della susseguente reazione effettrice. II primo tipo di linfociti Th tipo 1 o Th1 produce IL-2 (interleuchina-2), IFN-γ (interferone-γ), TNF-β (Tumor necrosis factor-β) ma non IL-4, IL-5 e IL-13, mentre i linfociti Th di tipo 2 o Th2 producono IL-4, IL-5, IL-6 e IL-13, ma non IL-2, IFN-γ e TNF-β. In genere quella dei linfociti Th1 rappresenta un tipo di risposta altamente protettiva nei confronti delle infezioni, soprattutto quando sostenute da batteri a parassitismo intracellulare. Per contro, i linfociti Th2 pongono le premesse fisiopatologiche per il manifestarsi a livello dell'organo bersaglio, e/o a livello di sedi diverse dell'organismo, della reazione infiammatoria allergica. Infatti i linfociti Th2, attraverso meccanismi che coinvolgono le interazioni tra molecole di superficie (CD40 e CD40 ligando), nonché mediante la produzione di IL-4 e IL-13, che funzionano come fattori di switch isotipico, inducono i linfociti B a produrre anticorpi della classe IgE.
Gli anticorpi IgE
Le IgE sono gli anticorpi primariamente responsabili delle reazioni di ipersensibilità immediata che stanno alla base sia dell'anafilassi che dell'atopia. La dimostrazione che le malattie allergiche rappresentano la conseguenza dell'interazione di un antigene con lo specifico anticorpo venne fornita nel 1921 da O.C. Prausnitz e H. Kustner. Questi ricercatori osservarono che l'iniezione intradermica del siero di un soggetto allergico al pesce nella cute di un soggetto non allergico e la successiva inoculazione nella stessa sede di un estratto antigenico di pesce causavano la comparsa di una reazione pomfo-eritematosa analoga a quella ottenibile sulla cute del primo con l'inoculazione del solo antigene. Questo esperimento è passato alla storia con la definizione di 'trasporto passivo locale di Prausnitz-Kustner' (P-K) e prima che fosse resa nota la possibilità del rischio di trasmissione di agenti infettivi virali (virus epatitici, ecc.) per l'applicazione di questo test, esso è stato utilizzato per molti anni anche a scopo diagnostico. I fattori sierici responsabili del trasferimento passivo dell'ipersensibilità vennero denominati 'regaine'. La scoperta che le regaine appartenevano a una classe immunoglobulinica diversa dalle altre quattro fino allora conosciute (IgM, IgG, IgA e IgD) condusse alla designazione delle IgE.
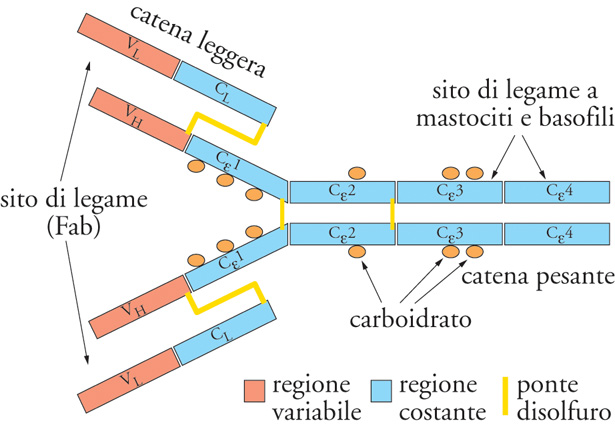
La molecola IgE presenta la classica struttura a quattro catene polipeptidiche (due leggere κ e λ e due pesanti ε). I circa 110 residui aminoacidici della catena ε dal lato NH-terminale differiscono da catena a catena originando la regione variabile (V) responsabile della specificità per l'antigene. La rimanente parte della catena ε è identica in tutte le IgE e costituisce la parte costante (C) della molecola. Essa comprende, a differenza delle altre Ig, che ne comprendono tre, quattro regioni omologhe o domini, denominati rispettivamente Cε1, Cε2, Cε3 e Cε4. Ricordiamo che le IgE non sono capaci di attivare il complemento per la via classica, di passare la barriera placentare e di dare reazioni di precipitazione con l'antigene; per questa ragione è stato difficile dimostrare la presenza di tali anticorpi con i metodi tradizionali di rilevazione. La peculiarità delle IgE risiede nella regione C dove sono situate le strutture responsabili della cosiddetta 'attività omocitotropica', cioè della capacità di questo tipo di Ig di interagire con i recettori ad alta affinità (FcεRI) dei mastociti e dei granulociti basofili (fig. 3).
Recettori per le IgE
Esistono due diversi tipi di recettori cellulari per le IgE designati rispettivamente di tipo 1 (FcεRI) e di tipo 2 (FcεRII). Gli FceRI sono presenti sui mastociti e sui granulociti basofili ed eosinofili, nonché sulle cellule di Langerhans della cute e sono caratterizzati da un'affinità molto elevata per le molecole delle IgE. Ciò rende ragione della prolungata persistenza, che può essere anche di alcune settimane, della sensibilità verso l'antigene, dimostrabile nella cute normale dopo il trasporto passivo locale secondo P-K, nonostante questi anticorpi presentino una breve emivita e una bassa concentrazione sierica. Il numero degli FcεRI sulla superficie dei mastociti e dei basofili è molto elevato (tra 4000 e 500.000 per cellula) e appare correlato, così come il loro grado di saturazione da parte delle molecole IgE, con la concentrazione di queste ultime nel siero. È stato dimostrato infatti che il numero dei recettori ad alta affinità aumenta con l'incremento dei livelli sierici di IgE. Gli FcεRI contengono quattro catene transmembrana, una α, una β e due γ. Il legame con la molecola IgE è ovviamente indipendente dalla sua specificità, per cui su una stessa cellula possono coesistere molteplici molecole IgE. Gli FcεRII differiscono dagli FcεRI per la diversa struttura, per la minore affinità nei confronti delle IgE (107÷108 M−1) e perché presenti su tipi cellulari diversi (monociti/macrofagi, eosinofili, linfociti e piastrine). Il legame delle IgE con gli FceRII attiva una varietà di funzioni effettrici da parte di queste cellule (citotossicità IgE-dipendente nei confronti di parassiti, fagocitosi di particelle rivestite di IgE, liberazione di mediatori).
La flogosi allergica
A causa della presenza sulla loro superficie di recettori ad alta affinità per le IgE (FcεRI), i mastociti e i granulociti basofili rappresentano le cellule effettrici più importanti delle reazioni immediate di tipo allergico. Infatti, l'interazione simultanea di due molecole adiacenti di IgE (legame a ponte) con l'allergene specifico determina il cosiddetto cross-linking recettoriale e induce la rapida secrezione di mediatori. Inoltre, attraverso la liberazione di mediatori chemiotattici e di citochine, i mastociti determinano un'amplificazione del fenomeno infiammatorio grazie al reclutamento di altre cellule nella sede della reazione immediata, quali granulociti neutrofili ed eosinofili, i macrofagi, che partecipano attivamente all'instaurarsi delle reazioni di fase tardiva e quindi del perpetuarsi della flogosi allergica. In seguito alla loro attivazione, i mastociti e i basofili secernono nell'ambiente esterno un'ampia gamma di mediatori distinguibili in quattro gruppi principali: (a) preformati, rilasciati rapidamente; (b) secondari, generati ex novo; (c) preformati, associati ai granuli; (d) citochine. Le reazioni immediate sono il risultato dell'azione vasodilatatrice e broncocostrittrice svolta dall'istamina, dalle prostaglandine (PGD2), dai leucotrieni e dal PAF (Platelet activating factor), i quali per tali motivi sono stati anche definiti 'mediatori effettori primari'.
In aggiunta ai linfociti e ai mastociti, i granulociti eosinofili rappresentano un altro elemento cellulare di importanza centrale nella genesi della flogosi allergica. Gli eosinofili originano nel midollo da precursori cellulari midollari identificati dalla molecola CD34 sotto l'influenza di fattori differenziativi, quali IL-3, IL-5 e GM-CSF. IL-5 costituisce senza dubbio il fattore di crescita più importante per queste cellule, che esprimono già dalle prime fasi di differenziazione il recettore specifico (IL-5R) nella sua struttura completa. È noto che IL-5 non solo stimola la crescita degli eosinofili ma ne rallenta anche il processo di apoptosi (morte cellulare programmata) a livello dei tessuti. In condizioni normali si trova uno scarso numero di eosinofili a livello dei tessuti, per esempio della parete bronchiale o intestinale, e tali cellule sono virtualmente assenti dalla cute; in condizioni patologiche, tuttavia, molti organi possono essere infiltrati dagli eosinofili e la cinetica della loro produzione risulta alterata. Ne sono esempi classici la cute in corso di dermatite atopica, la mucosa bronchiale in corso di asma bronchiale, la mucosa intestinale in corso di allergia alimentare. Recentemente è stato dimostrato che nel processo di reclutamento selettivo degli eosinofili nei tessuti sede di flogosi hanno un ruolo determinante alcune CC-chemochine (β-chemochine) tra le quali eotassina-1 e -2, RANTES, MCP-3 e MCP-4. Infatti, questi fattori richiamano gli eosinofili nella sede grazie all'interazione con lo specifico recettore CCR-3.
A differenza degli altri granulociti, gli eosinofili mostrano la presenza di granuli secondari caratteristici, contenenti quattro distinte proteine cationiche: la proteina basica maggiore (MBP), la proteina cationica eosinofila (ECP), la perossidasi eosinofila (EPO) e la neurotossina derivata dagli eosinofili (EDN). In aggiunta alle proteine cationiche, gli eosinofili attivati secernono anche mediatori lipidici, PAF, LTC4, lo ione superossido O2− e alcune citochine, quali IL-1, IL-3, IL-4, IL-5, IL-6, GM-CSF e TGF-α e -β. Inoltre, gli eosinofili esprimono una serie di molecole di adesione, quali ICAM-1, VCAM-1, LFA-1 e le selectine, indispensabili per la loro fuoriuscita dal circolo ematico. I prodotti di derivazione eosinofila svolgono probabilmente anche un ruolo fisiologico nella difesa contro i parassiti, ma intervengono certamente come effettori primari delle alterazioni istopatologiche nei tessuti sede di flogosi allergica.
È stato posto in evidenza da tempo che la flogosi allergica non si esprime solamente attraverso l'interazione tra le molecole allergeniche e gli anticorpi IgE fissati ai recettori ad alta affinità dei mastociti e dei basofili, e la successiva liberazione di istamina e degli altri mediatori. Nella maggioranza dei soggetti allergici intervengono anche meccanismi più complessi. Nelle fasi tardive delle risposte primarie e nelle risposte secondarie agli allergeni, ossia quelle che si realizzano quando l'organismo ha già incontrato l'allergene e ha avuto ormai inizio un'efficiente sintesi di anticorpi IgE, gli allergeni sono captati molto facilmente dalle cellule dendritiche con IgE specifiche fissate ai FcεRI di membrana e anche dai linfociti B con IgE specifiche fissate ai recettori a bassa affinità (FcεRII). I linfociti B sono in grado di cooperare con i linfociti Th2 della memoria specifici per gli allergeni così da amplificare la risposta immunitaria. Questo costituisce un sistema di amplificazione e di mantenimento importante per la continuità delle risposte verso gli allergeni e per la flogosi allergica.
Le conoscenze attuali hanno dunque permesso di prospettare una visione integrata dei meccanismi patogenetici delle malattie allergiche, sostenuta anche da meccanismi di amplificazione e di mantenimento che originano da complesse interazioni cellulari e molecolari e che conducono alla fase tardiva di cronicizzazione della flogosi stessa. Del resto, sia i mastociti che i granulociti basofili sono in grado di liberare citochine, quali IL-4, IL-5, GM-CSF, e TNF-α, capaci di indurre la crescita, la differenziazione e l'attivazione di cellule direttamente coinvolte nella flogosi tissutale allergica, anche nelle fasi più avanzate di 'cronicizzazione'. L'espansione dei linfociti Th2 attraverso la liberazione di citochine (IL-3, IL-4, IL-5, IL-13, GM-CSF), rappresenta un fattore comune capace di collegare l'iperproduzione di IgE e il coinvolgimento di cellule quali gli eosinofili, i mastociti, i basofili, e i macrofagi.
Va sottolineata l'importanza degli eosinofili quali cellule effettrici responsabili di buona parte delle lesioni tissutali riscontrabili in corso di malattie allergiche. Gli eosinofili possono danneggiare le mucose fondamentalmente mediante il rilascio di proteine basiche, di radicali liberi dell'ossigeno e di mediatori lipidici. Gli eosinofili esercitano la loro azione proinfiammatoria anche tramite la liberazione di molteplici citochine, quali IL-3, IL-4, TGF-β, TGF-α, GM-CSF, IL-1, IL-6 e IL-5. Particolare attenzione è stata rivolta allo studio dei fenomeni coinvolti nella migrazione degli eosinofili dal circolo ematico ai tessuti sede di flogosi allergica. Uno dei primi eventi è legato all'attivazione delle cellule endoteliali a opera di citochine (IL-1, IL-4, IL-13, TNF-α, IFN-γ) con conseguente aumentata espressione di molecole di adesione. Infatti, gli eosinofili aderiscono inizialmente alla selettina E presente sull'endotelio dei piccoli vasi (rolling cellulare) e successivamente le integrine, quali LFA-1 e MAC-1, mediano l'ancoraggio endoteliale legandosi a ICAM-1.
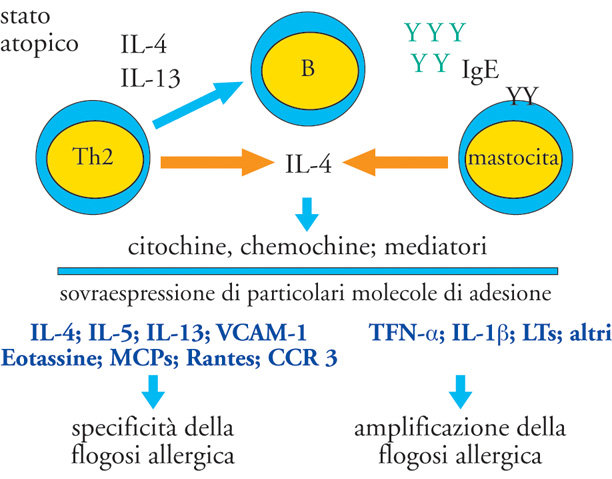
Inoltre, gli eosinofili si legano tramite le molecole VLA-4, espresse costitutivamente sulla loro membrana, alle molecole VCAM-1 delle cellule endoteliali. L'espressione di VCAM-1 sulle cellule endoteliali, importante per la migrazione selettiva degli eosinofili, è indotta dalla produzione locale, oltre che di IL-4, anche di IL-13. La migrazione verso la mucosa è decisamente favorita da IL-5, ma anche da PAF. Va notato che IL-5 e GM-CSF prolungano la sopravvivenza degli eosinofili nel tessuto sede della flogosi allergica, inibendo la loro apoptosi. Gli eosinofili in tal modo sono in grado di migrare verso l'endotelio, al quale possono aderire grazie, nuovamente, alle molecole di adesione ICAM-1 espresse dalle cellule epiteliali ed esercitare le proprie funzioni di danno. Come già riferito, nel processo di migrazione cellulare hanno assunto rilievo citochine dotate di potere chemiotattico e denominate 'chemochine' (RANTES, MIP-1α, MCP-3 e -4, eotassine). Esse inducono una forte e selettiva chemiotassi degli eosinofili, soprattutto attraverso il recettore CCR-3, che è presente anche, come già ricordato, sui basofili e sui mastociti (fig. 4).
Ruolo complementare dei fattori genetici e ambientali nello sviluppo dell'allergia
Le caratteristiche genetiche sono molto importanti nel condizionare la produzione delle IgE nei confronti degli allergeni; la presenza di IgE rappresenta l'alterazione fisiopatologica fondamentale della cosiddetta 'condizione atopica'. È noto da tempo che quando entrambi i genitori sono atopici le manifestazioni allergiche sono presenti nel 60-80% dei figli; qualora invece sia allergico un solo genitore la condizione si realizza nel 20-40% dei casi. La madre sembra condizionare più del padre la suscettibilità allo stato atopico. Per quanto riguarda l'analisi delle basi molecolari della predisposizione allo stato atopico, sono stati posti in evidenza come possibili 'candidati' i geni presenti nel cromosoma 5; i geni del cromosoma 6 correlati con gli antigeni delle subregioni DR e DQ del sistema HLA e con la citochina TNF-α; un gene localizzato nel cromosoma 11q13 correlato con la catena β di FcεRI; i geni localizzati nel cromosoma 12q correlati con IFN-γ; i geni localizzati nel cromosoma 14q correlati con la catena α del TCR e con l'importante fattore di trascrizione NF-kB; un gene localizzato nel cromosoma 16p12 correlato con la catena α del recettore di IL-4; infine, i geni che codificano la subfamiglia delle β-chemochine, localizzati nel cromosoma 17q11-12.
Negli ultimi anni è stata attribuita molta importanza all'esistenza nei soggetti atopici di una possibile disregolazione dei geni delle citochine presenti sul cromosoma 5 (IL-4, IL-5, IL-3, IL-9, IL-13, GM-CSF) appartenenti alla 'famiglia di IL-4'. IL-4 rappresenta infatti la citochina fondamentale in quanto costituisce non solo uno dei due fattori di attivazione della produzione di IgE da parte dei linfociti B, ma anche il fattore più importante per la differenziazione dei linfociti Th vergini in linfociti di tipo Th2. A tale proposito sono stati identificati fattori di trascrizione specifici per la differenziazione dei linfociti Th2, tra cui i più importanti sono quelli designati rispettivamente GATA-3, c-maf, Stat-6. Alterazioni della funzione regolatoria di questi fattori potrebbero essere responsabili della iperespressione del gene di IL-4. Inoltre, i fattori di trascrizione della differenziazione Th2 giocano un ruolo antagonizzante reciproco con quelli della differenziazione Th1 (in particolare quello designato T-bet capace di attivare il 'promoter' di IFN-γ) per cui, per esempio, l'attivazione di GATA-3 non solo favorisce la differenziazione dei linfociti Th2, ma inibisce anche quella dei linfociti Th1, e viceversa.
Recentemente è stata anche identificata l'esistenza di cellule T regolatorie (Treg), per la cui funzione appare determinante l'epressione del gene Foxp3, fondamentali sia nel mantenimento della tolleranza periferica nei confronti di autoantigeni che nella regolazione della risposta immunologica nei confronti di antigeni esogeni tra i quali anche gli allergeni. È stato suggerito che nei soggetti atopici esista una minore efficacia di funzione soppressiva delle cellule Treg rispetto ai soggetti non atopici. Tuttavia, a prescindere dalle alterazioni genetiche di Foxp3, le quali determinano l'insorgenza di una grave malattia congenita caratterizzata dalla comparsa sia di malattie autoimmuni che di manifestazioni allergiche designata IPEX, le possibili alterazioni genetiche della funzione delle cellule Treg, eventualmente responsabili delle forme più comuni di atopia, sono del tutto sconosciute. Appare comunque evidente che le malattie allergiche rappresentano sempre il risultato di alterazioni multigeniche.
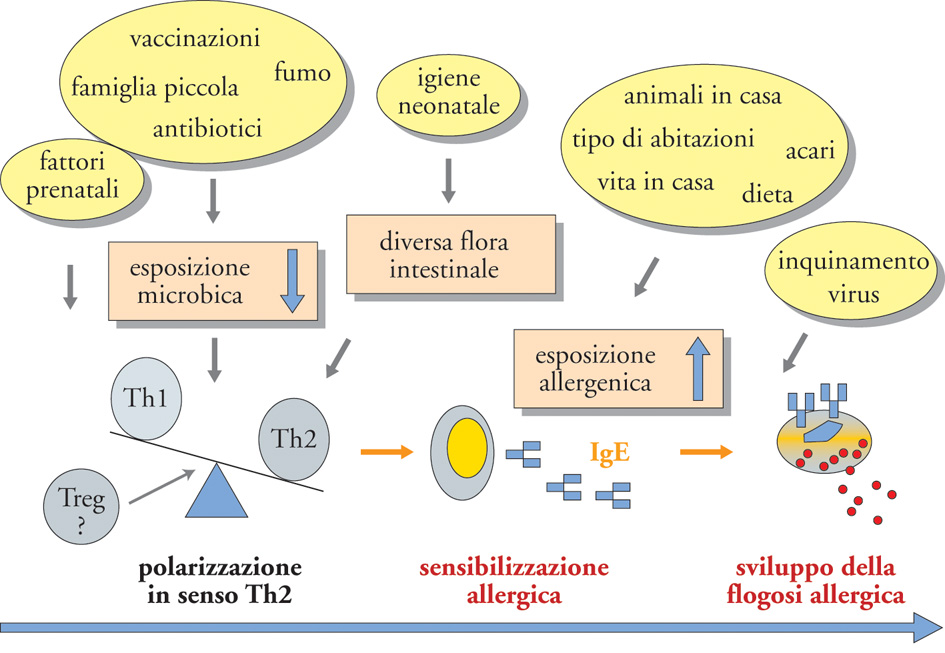
Il ruolo concomitante dei fattori ambientali nello sviluppo delle malattie atopiche e, soprattutto, nell'incremento della loro prevalenza nelle comunità con stile di vita occidentale ha indotto a formulare alcune ipotesi, tra le quali la cosiddetta 'ipotesi igienica' (fig. 5). Tra i vari fattori ambientali quelli più importanti sembrano essere rappresentati dai cambiamenti avvenuti nei Paesi occidentali a partire dalla fine della Seconda guerra mondiale, soprattutto per quanto riguarda l'esposizione microbica e agli allergeni ambientali domestici durante l'infanzia. Cambiamenti radicali nell'esposizione microbica si sono infatti realizzati per il verificarsi di alcune condizioni e circostanze concomitanti: (a) migliori standard igienici dei cibi e delle bevande e conseguente modificazione della flora intestinale; (b) diminuzione dei componenti delle famiglie con ridotto rischio di trasmissione di infezioni; (c) decorso dei quadri infettivi come conseguenza della migliore nutrizione, delle vaccinazioni e dell'uso di antibiotici.
L'interpretazione biologica di tali osservazioni si basa sul fatto che in questa fase della vita la riduzione dello stimolo microbico può influenzare la polarizzazione della risposta immunitaria a favore dei linfociti Th2. Il lipopolisaccaride (LPS), come probabilmente altri componenti della parete batterica, costituisce uno stimolo potente per l'induzione della produzione di IL-12 da parte delle cellule dendritiche che favorisce la differenziazione dei linfociti Th1. La mancanza o, più probabilmente, la riduzione di tali stimoli potrebbe favorire la polarizzazione della risposta immunitaria in senso Th2. Un'altra possibilità sarebbe rappresentata dalla minore attivazione delle cellule Treg legata alla ridotta stimolazione del sistema immunitario con minore produzione di citochine regolatrici quali IL-10 e TGF-β (Transforming growth factor-β), capaci di sopprimere la risposta immune sia agli agenti batterici che agli allergeni. Tra gli altri cambiamenti nello stile di vita verificatisi negli ultimi trenta-quarant'anni, hanno un ruolo significativo le modalità di costruzione delle abitazioni con minore ventilazione degli ambienti; il riscaldamento e l'arredamento delle abitazioni che facilitano lo sviluppo degli acari della polvere domestica. Molto controverso appare invece il ruolo degli inquinanti atmosferici. Verosimilmente, in soggetti geneticamente predisposti e in coloro che hanno manifestazioni allergiche subcliniche, l'incremento dell'esposizione agli inquinanti, tipo anidride solforosa e ozono, e al fumo di tabacco può favorire la sensibilizzazione allergica, scatenare i sintomi di alcune patologie allergiche quali l'asma bronchiale e la rinite.
Diagnostica delle malattie allergiche
La diagnosi eziologica delle malattie allergiche consiste nella ricerca dei fattori specifici che determinano la malattia in un particolare soggetto. L'individuazione dell'allergene (o degli allergeni) verso cui il soggetto è sensibile appare di estrema importanza, non soltanto quale convalida della diagnosi clinica, ma anche per i suoi riflessi terapeutici. L'individuazione degli allergeni responsabili dei sintomi può essere operata agevolmente per mezzo delle prove allergologiche cutanee. In alcuni casi è necessario ricorrere anche a esami sieroimmunologici, quali, per esempio, il RAST (Radioallergosorbent test). Le prove allergologiche cutanee, che risultano di facile esecuzione e consentono di testare in breve tempo un considerevole numero di allergeni sospetti, sono molto attendibili, a condizione che la loro esecuzione sia corretta e i risultati vengano valutati attentamente. Le prove allergologiche dovrebbero costituire il primo accertamento diagnostico, almeno nelle forme cliniche più comuni (per es., nelle manifestazioni allergiche a carico dell'apparato respiratorio). Tali prove sono in genere ben accettate dai pazienti; inoltre, sono di rapida esecuzione, sicure, sensibili ed economiche. I test cutanei vengono ormai eseguiti in tutto il mondo, prevalentemente con la metodica del Prick test.
Gli esami sieroimmunologici vengono utilizzati per dosare la quantità degli anticorpi IgE presenti nel siero e in altri liquidi biologici. La determinazione dei valori degli anticorpi IgE totali con metodiche di radioimmunoassorbimento (RIST, Radioimmunosorbent test) o di radioassorbimento su carta (PRIST, Paper radioimmunosorbent test), in caso di risultato fortemente positivo è ritenuta probativa per uno stato allergico. Ciò, naturalmente, dopo avere escluso le altre cause di aumento delle IgE totali. Oltre che nelle malattie allergiche, infatti, un aumento delle IgE totali nel siero può essere riscontrato nella aspergillosi broncopolmonare e nelle infestazioni da parassiti, nonché in alcuni linfomi. Il ricorso a questo metodo per la ricerca mirata di anticorpi IgE specifici verso un numero selezionato di allergeni è giustificato nelle seguenti condizioni: quando esista una discordanza tra storia clinica e prove allergologiche cutanee; nei casi di prove allergologiche dubbie; nei bambini molto piccoli, nei quali non è possibile effettuare le prove allergologiche; nei soggetti trattati con antistaminici, i quali sono in grado di diminuire la reattività cutanea agli allergeni utilizzati per le prove cutanee; infine, nei pazienti con gravi ed estese manifestazioni dermatologiche, ove l'esecuzione dei test cutanei non è tecnicamente possibile.
Cenni di terapia delle malattie allergiche
La terapia delle malattie allergiche si basa, quando possibile, sulla rimozione o sulla riduzione della concentrazione nell'ambiente dell'allergene responsabile della sensibilizzazione. Quando questo provvedimento risulta impossibile è necessario l'impiego di farmaci, associati o meno alla cosiddetta 'immunoterapia specifica'. I farmaci più attivi sono certamente gli antistaminici nelle forme rinitiche e in quelle cutanee caratterizzate dalla presenza di prurito (orticaria, dermatite atopica). Nelle forme di asma bronchiale risultano più efficaci i corticosteroidi per uso topico, ma nelle forme asmatiche più gravi è necessario ricorrere anche ai corticosteroidi per via sistemica. Infatti l'asma bronchiale rappresenta una malattia infiammatoria cronica delle vie aeree, che può anche provocare modificazioni irreversibili a carico di queste ultime. Tale fenomeno, noto come 'rimodellamento delle vie aeree', può determinare una notevole riduzione delle capacità funzionali respiratorie e deve quindi essere prevenuto. La immunoterapia specifica si basa sulla somministrazione per alcuni anni dello stesso allergene responsabile dei sintomi, che può essere effettuata per via iniettiva o per via orale. Ciò allo scopo di determinare una iposensibilizzazione o uno stato di tolleranza verso l'allergene che viene normalmente introdotto spontaneamente per via inalatoria. La immunoterapia specifica per via iniettiva deve essere limitata ai casi resistenti ai farmaci, in quanto potenzialmente in grado di determinare effetti collaterali anche gravi, quale lo shock anafilattico. La immunoterapia specifica per via orale è più sicura, anche se non altrettanto efficace di quella per via iniettiva. Sono attualmente in studio nuove modalità di immunoterapia specifica, che potrebbero combinare la sicurezza di quelle per via orale con l'efficacia di quelle effettuate per via iniettiva.
Bibliografia
Annunziato 2002: Annunziato, Francesco e altri, Phenotype, localization and mechanism of suppression of CD4(+)CD5(+) human thymocytes, "Journal of experimental medicine", 196, 2002, pp. 379-387.
Bluestone, Abbas 2003: Bluestone, Jeffrey A. - Abbas, Abal K., Natural versus adaptive regulatory T cells, "Nature reviews. Immunology", 3, 2003, pp. 253-257.
Coca, Cooke 1923: Coca, Arthur F. - Cooke, Robert A., On the classification of the phenomena of hypersensitiveness, "Journal of immunology", 8, 1923, pp. 163-182.
D'Amato 1998: D'Amato, Giuseppe e altri, Pollen-related allergy in Europe, "Allergy", 53, 1998, pp. 567-568.
Del Prete 1991: Del Prete, Gianfranco e altri, Purified protein derivative of Mycobacterium tuberculosis and excretory-secretory antigen(s) of Toxocara canis expand in vitro human T cells with stable and opposite (type 1 T helper or type 2 T helper) profile of cytokine production, "Journal of clinical investigation", 88, 1991, pp. 346-350.
Mosmann 1986: Mosmann, Tim R. e altri, Two types of murine helper T cell clone. Definition according to profiles of lymphokine activities and secreted proteins, "Journal of immunology", 136, 1986, pp. 2348-2357.
Romagnani 1995: Romagnani, Sergio, Biology of human Th1 and Th2 cells, "Journal of clinical immunology", 15, 1995, pp. 121-129.
Romagnani 2004: Romagnani, Sergio, Immunologic influences on allergy and the TH1/TH2 balance, "Journal of allergy and clinical immunology", 113, 2004, pp. 395-400.
Romagnani 2004: Romagnani, Sergio, The increased prevalence of allergy and the hygiene hypothesis: missing immune deviation, reduced immune suppression, or both?, "Immunology", 112, 2004, pp. 352-363.