
Apprendimento. Basi molecolari dell'apprendimento
Apprendimento. Basi molecolari dell'apprendimento
I moderni studi di psicologia cognitiva hanno chiaramente dimostrato che l'apprendimento e la memoria non sono facoltà unitarie della mente; al contrario, è divenuto sempre più evidente che tali processi intellettivi sono costituiti da stadi di elaborazione differenti. Questi processi sono distinguibili funzionalmente sia in virtù della natura specifica dell'informazione elaborata, sia in base al decorso temporale del consolidamento (o, al contrario, del decadimento) dell'informazione acquisita. Le forme di apprendimento esplicito o dichiarativo sono connesse all'acquisizione di informazioni relative agli altri individui, ai luoghi, alle cose. Per 'apprendimento implicito' o 'procedurale' gli studiosi intendono l'apprendimento di abilità motorie o di altro genere, tra cui i processi associativi semplici, come per esempio il condizionamento classico, e i processi non associativi, quali la sensibilizzazione e l'assuefazione. Come è stato dimostrato per la prima volta attraverso studi di neuropsicologia condotti su pazienti cerebrolesi, l'apprendimento esplicito dipende fondamentalmente dall'integrità delle strutture del lobo temporale mediale, una porzione della corteccia cerebrale all'interno della quale sicuramente la struttura più importante è la formazione ippocampale. L'apprendimento implicito interessa invece soltanto i percorsi sensoriali, motori o associativi reclutati per le abilità percettive o motorie particolari, utilizzate durante il processo di apprendimento. Di conseguenza, l'apprendimento implicito è studiato con maggiore facilità in vari sistemi a riflessi semplici, inclusi quelli degli invertebrati superiori, mentre le forme esplicite di apprendimento sono studiate soprattutto nei Mammiferi.
Studi clinici condotti su esseri umani, nonché i numerosi studi sperimentali su differenti modelli animali, dai Molluschi ai Roditori e alle scimmie, suggeriscono che ogni forma di memoria è caratterizzata da stadi distinti: uno stadio a breve termine, che si protrae per alcuni secondi, minuti o ore, e uno a lungo termine, che permane per giorni, settimane e talvolta per tutta la vita. Gli studi sulla memoria umana compiuti all'inizio del XX sec. descrivevano un periodo di transizione dalla memoria a breve termine a quella a lungo termine. Durante questo periodo di consolidamento la registrazione mnemonica è labile e altamente suscettibile di cancellazione (cioè, facilmente destinata al cosiddetto 'oblio'). Studi molecolari recenti sulle forme implicite ed esplicite di apprendimento suggeriscono che questa transizione corrisponde a un programma di espressione genica specificamente differente da quella di norma in corso. Tale programma richiede l'intervento di una lunga e complessa sequenza di mediatori chimici all'interno della cellula nervosa e del suo nucleo, che ne modificano notevolmente il funzionamento generale. Questa cascata di eventi molecolari trasforma quello che sarebbe di per sé un processo transitorio, consistente nella modificazione covalente delle proteine preesistenti e in un cambiamento dell'efficienza delle sinapsi già funzionanti nel sistema nervoso, in un processo stabile di autoconservazione che si accompagna alla crescita di nuove connessioni sinaptiche e, quindi, di vera e propria ristrutturazione fisica della rete neurale cerebrale.
sommario
1. Biologia molecolare dell'apprendimento implicito in Aplysia. 2. Il ruolo dei fattori di trascrizione genica. 3. Induzione di geni precoci. 4. Stabilizzazione della facilitazione a lungo termine. 5. Consolidamento della memoria a lungo termine. 6. La memoria a lungo termine come sequenza di stadi molecolari. □ Bibliografia.
Biologia molecolare dell'apprendimento implicito in Aplysia
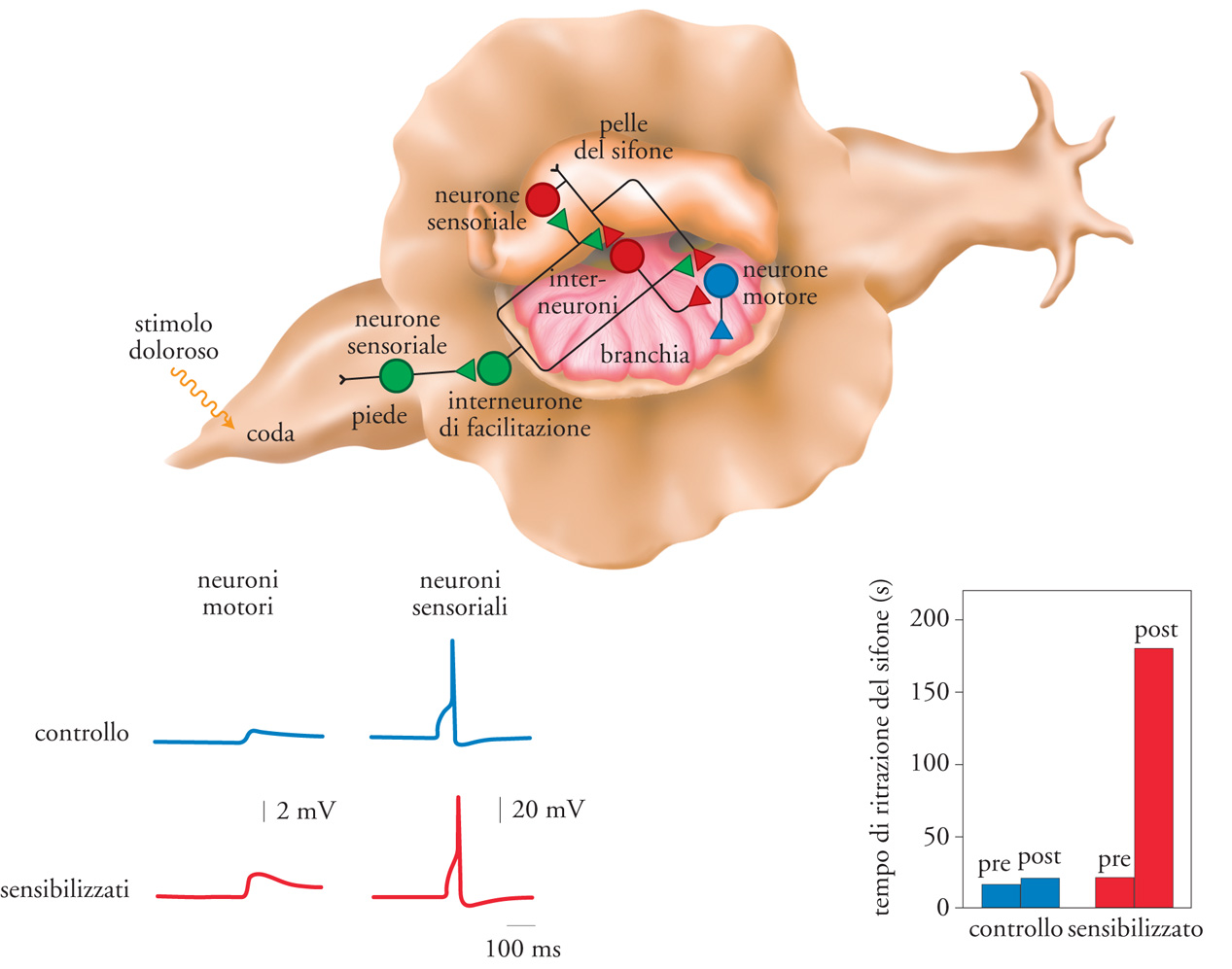
Il sistema nervoso della lumaca marina Aplysia californica si è dimostrato utile come modello per lo studio delle basi cellulari e molecolari dell'apprendimento e della memoria. Questo sistema contiene soltanto circa 20.000 grandi cellule nervose ben evidenti, raggruppate in dieci gangli principali. La possibilità di identificare i singoli neuroni e di registrarne l'attività ha permesso di definire i principali componenti dei circuiti neuronali connessi a comportamenti specifici. I meccanismi molecolari che contribuiscono all'immagazzinamento della memoria implicita sono stati studiati in modo particolarmente esteso per il riflesso di ritrazione della branchia in Aplysia, che può venire modificato da numerose forme di apprendimento implicito. Verrà qui considerata la sensibilizzazione, una forma elementare di apprendimento grazie alla quale, dopo essere stato esposto a uno stimolo potenzialmente pericoloso, l'animale impara a rafforzare i propri riflessi difensivi e a rispondere vigorosamente a stimoli che in precedenza davano risposte neutre. In Aplysia è possibile sensibilizzare il riflesso di ritrazione della branchia mediante uno stimolo forte a livello della coda (fig. 2); un singolo stimolo alla coda dà luogo a una sensibilizzazione a breve termine che si protrae per minuti o ore. La ripetizione di questo stimolo produce apprendimento a lungo termine che può protrarsi per giorni o settimane. In questo modo si attivano gli interneuroni di facilitazione che formano collegamenti sinaptici con determinati neuroni sensoriali e rafforzano la connessione sinaptica tra i neuroni sensoriali e i loro neuroni motori (motoneuroni) bersaglio.
La sensibilizzazione a breve o a lungo termine intensifica la trasmissione sinaptica a livello del collegamento monosinaptico tra i meccanorecettori dei neuroni sensoriali e quelli dei neuroni motori. Sebbene sia attribuibile a questa componente solo una parte della modificazione comportamentale misurata nell'animale integro, la sua semplicità ha facilitato l'analisi cellulare e molecolare sia della forma di sensibilizzazione a breve termine sia di quella a lungo termine. Il percorso monosinaptico può essere ricostituito in coltura cellulare dissociata, dove la serotonina (5-HT, 5-idrossitriptammina), un importante neurotrasmettitore, sostituisce la scossa inviata alla coda durante l'addestramento comportamentale dell'animale integro. Analogamente alla sensibilizzazione, una singola applicazione di 5-HT produce cambiamenti a breve termine nell'efficacia sinaptica, mentre cinque applicazioni distanziate, somministrate durante un periodo di 1,5 h, determinano cambiamenti che durano uno o più giorni. Gli studi biofisici e biochimici delle connessioni che collegano i neuroni sensoriali a quelli motori, sia nell'animale integro sia nelle cellule in coltura, indicano che nei cambiamenti a breve e a lungo termine si ritrovano aspetti di un meccanismo molecolare comune. Ambedue i processi vengono infatti avviati dalla 5-HT e l'aumento della forza sinaptica, osservato sia durante il cambiamento a breve termine sia durante quello a lungo termine, deriva in parte da un aumento del rilascio di trasmettitori dai neuroni sensoriali. Questo aumento presinaptico dell'eccitabilità è dovuto all'allungamento nel tempo del potenziale d'azione, determinato dalla modulazione, per mezzo della 5-HT, delle correnti di potassio.
Nonostante le numerose analogie, i cambiamenti cellulari a breve termine differiscono da quelli a lungo termine per due aspetti importanti. In primo luogo, a breve termine avviene soltanto la modificazione covalente di proteine preesistenti e un'alterazione delle preesistenti connessioni. Sia la sensibilizzazione comportamentale a breve termine nell'animale, sia la facilitazione a breve termine in coltura cellulare dissociata non richiedono una sintesi macromolecolare continua. Per contro, gli inibitori della trascrizione o della traduzione bloccano l'induzione dei cambiamenti a lungo termine sia nella preparazione semintatta sia nella coltura cellulare primaria. Particolarmente interessante è la scoperta che l'induzione della facilitazione a lungo termine, a livello di questa singola sinapsi di Aplysia, richiede la sintesi di proteine e di RNA durante la finestra temporale critica (o periodo di consolidamento). Numerose forme di memoria, sia nei Vertebrati sia negli invertebrati, condividono, durante il periodo di consolidamento, questa necessità di sintesi di macromolecole. Da un punto di vista molecolare, questi studi indicano che per i cambiamenti comportamentali e cellulari a lungo termine è necessaria l'espressione di geni e proteine, diversamente che per i processi a breve termine. L'identificazione dei prodotti genici richiesti per questo consolidamento rimane il principale obiettivo della ricerca molecolare sui processi della memoria. In secondo luogo, il processo a lungo termine, ma non quello a breve termine, comporta un cambiamento strutturale. Craig H. Bailey e Mary Chen (1983, 1988) hanno da tempo dimostrato che in Aplysia l'addestramento della sensibilizzazione a lungo termine si associa alla crescita di nuove connessioni sinaptiche tra i neuroni sensoriali e i neuroni motori. Cambiamenti analoghi possono essere ricostituiti nelle cocolture di neuroni sensoriali e motori per mezzo di presentazioni ripetute di 5-HT.
Il ruolo dei fattori di trascrizione genica
Nelle cellule dei Mammiferi, la 5-HT induce una serie di eventi biochimici a cascata, mediati dal cAMP (adenosinmonofosfato ciclico) e dalla proteina-chinasi A (PKA); quest'ultima, a sua volta, attiva l'espressione genica attraverso la fosforilazione dei fattori di trascrizione che si legano a CRE (cAMP responsive element). CRE è una sequenza di DNA presente all'interno della regione di controllo del gene. Il legame di differenti fattori di trascrizione a questi elementi di risposta regola l'attività dell'RNA-polimerasi, determinando così il momento e il livello dell'espressione genica. Uno dei principali fattori di trascrizione che riconoscono la sequenza CRE è CREB1 (CRE binding protein 1), che funziona da attivatore trascrizionale solo dopo essere stata fosforilata dalla PKA. Mediante esperimenti in coltura, si è visto che CREB1 è essenziale per la facilitazione a lungo termine: infatti, bloccando il legame di questa proteina alla sequenza di DNA responsabile della risposta al cAMP si elimina selettivamente il processo a lungo termine.
Durante la facilitazione a lungo termine, la PKA sembra attivare l'espressione genica nei neuroni sensoriali di Aplysia per mezzo di un omologo di CREB1 presente in questo mollusco. A questo punto occorre chiedersi se CREB1 agisce da solo o con altri fattori di trascrizione: uno di questi, ApCREB2, è stato studiato in dettaglio. ApCREB2 è espresso a livello basale (senza induzione da parte della 5-HT) nei neuroni sensoriali di Aplysia e non è indotto dalla 5-HT. Nella sua struttura primaria generale, ApCREB2 è omologo tanto a CREB2 umano quanto a ATF4 murino, fattori di trascrizione che fungono da repressori dell'espressione genica mediata da CREB1. Curiosamente, in determinate circostanze, ApCREB2 può fungere da attivatore trascrizionale PKA-dipendente. Poiché ApCREB2 non contiene alcuna sequenza di consenso di fosforilazione da parte di PKA, questo enzima può regolare l'attività di CREB2 modificando altri fattori di trascrizione che interagiscono con CREB2, tra cui CREB1. In questo modo il partner di dimerizzazione di CREB2 (e di CREB1) può essere fondamentale per determinare l'effetto che si produce sulla regolazione trascrizionale.
Mediante esperimenti di inattivazione del fattore ApCREB2, si è visto che esso è un repressore funzionale della facilitazione a lungo termine e l'attenuazione di questa repressione potenzia il processo di attivazione. Di conseguenza l'espressione genica indotta da cAMP può comportare due stadi collegati: l'attivazione di CREB1 e l'attenuazione della repressione di CREB2. Gli studi su Aplysia sono di particolare interesse in quanto indicano la possibilità che l'eliminazione della repressione ApCREB2-mediata possa limitare la regolazione dell'aumento a lungo termine della forza sinaptica. Consideriamo come sia possibile attenuare la repressione esercitata da ApCREB2 su ApCREB1. Poiché dopo l'esposizione a 5-HT non si osserva alcuna distruzione della proteina ApCREB2, si deve dedurre che le immissioni ripetute di 5-HT inducano modificazioni covalenti di ApCREB2. In effetti, sono stati osservati dei cambiamenti nello stato di fosforilazione di ApCREB2, in seguito all'esposizione ripetuta a 5-HT. È interessante osservare che ApCREB2 condivide i siti di fosforilazione della proteina-chinasi C e della MAP-chinasi con i propri omologhi umani e murini. Inoltre, la MAP-chinasi è attivata dalla 5-HT nei neuroni di Aplysia e, come la PKA, con un trattamento prolungato di 5-HT si trasferisce nel nucleo. Il trasferimento della PKA e della MAP-chinasi nel nucleo può in parte spiegare le ragioni per le quali la facilitazione a lungo termine richiede ripetute immissioni di 5-HT. È possibile che queste siano necessarie per permettere l'attivazione persistente della PKA e della MAP-chinasi in modo da rendere operanti gli attivatori (CREB1) e attenuare i repressori (CREB2).
Induzione di geni precoci
Gli inibitori della sintesi dell'RNA o delle proteine non producono alcun effetto sull'attivazione dell'adenilatociclasi da parte della 5-HT, sulla generazione del cAMP, sull'attivazione della PKA, sul trasferimento della subunità catalitica della PKA al nucleo e sulla fosforilazione di CREB1. È chiaro che, per quanto riguarda il punto in cui si manifesta la sintesi dell'RNA e delle proteine che caratterizza la fase di consolidamento, è necessaria una fase ulteriore, cioè la sintesi delle proteine codificate dai geni la cui espressione è indotta da CREB1 e repressa da CREB2. Per esaminare i geni che si trovano a valle di CREB1, C. Alberini e collaboratori (1994) hanno analizzato i fattori di trascrizione regolati da cAMP, in modo da identificare i geni espressi durante il consolidamento della facilitazione a lungo termine. Alcuni fattori di trascrizione, noti per essere attivati da cAMP, appartengono alla famiglia di proteine che si legano alla sequenza enhancer CCAAT (C/EBPs, C/enhancer binding proteins). Un membro di questa famiglia, C/EBP, è espresso nella linea cellulare PC 12 del feocromocitoma di ratto dove, come è stato dimostrato, viene attivato da cAMP e regola l'espressione del gene c-fos legandosi a ERE (Enhancer response element) nel promotore c-fos. Poiché i neuroni di Aplysia sono caratterizzati da un'attività specifica di legame con ERE, Alberini e i suoi collaboratori hanno usato la sequenza di legame di ERE per isolare un clone che codifica una proteina la quale interagisce in modo specifico con i siti di legame al DNA di C/EBP. In Aplysia, l'mRNA di C/EBP è espresso a bassi livelli allo stato basale, ma viene indotto rapidamente e in modo transitorio dalla 5-HT e dal cAMP. Questa regolazione dell'espressione ApC/EBP, cAMP-dipendente, può essere mediata direttamente da CREB1 poiché, a monte del gene ApC/EBP, si trova un sito CRE.
I fattori di trascrizione vengono comunemente denominati 'costitutivi', quando sono sempre presenti, e 'inducibili', qualora la loro induzione richieda l'attivazione dei rispettivi geni. I fattori di trascrizione inducibili si dividono ulteriormente in precoci e tardivi, a seconda della rapidità di attivazione in risposta a un dato stimolo; CREB1 è un fattore di trascrizione espresso in termini costitutivi. I cosiddetti 'geni precoci', descritti originariamente negli studi sulla regolazione virale, sono attivati dalla fosforilazione dei fattori di trascrizione costitutivi; ne risulta che essi vengono indotti rapidamente e transitoriamente da meccanismi che non dipendono dalla sintesi proteica. ApC/EBP è indotto come gene precoce che svolge la funzione di interruttore molecolare per il passaggio della memoria a breve termine nella memoria a lungo termine: dopo 15 min è individuabile l'mRNA di ApC/EBP, che raggiunge il livello massimo dopo 2 h e quindi decresce; inoltre viene indotto in presenza di inibitori della sintesi proteica. Per quanto riguarda la durata dell'attivazione di questo fattore di trascrizione, Alberini e collaboratori hanno dimostrato, iniettando oligonucleotidi ERE in cellule sensoriali in tempi diversi dopo trattamento con 5-HT, che l'attività di ApC/EBP era necessaria solo durante le prime 12 h successive all'addestramento. Quindi, l'induzione di ApC/EBP, durante il trattamento con 5-HT, produce l'attivazione di una cascata di eventi autoperpetuantisi, essenziali per la fase tardiva della facilitazione a lungo termine.
Stabilizzazione della facilitazione a lungo termine
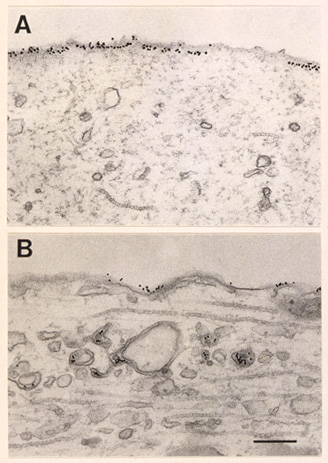
I prodotti della rete di geni, solo in parte identificati, conducono alla crescita di ulteriori sinapsi tra i neuroni sensoriali e i neuroni motori che caratterizzano la fase tardiva dell'immagazzinamento mnemonico. In effetti la stabilità della facilitazione a lungo termine sembra essere il risultato della persistenza di questi cambiamenti strutturali, il cui decadimento corre parallelamente a quello della traccia di memoria. Consideriamo come si formano le nuove connessioni sinaptiche. In cocolture di neuroni sensoriali e motori, la formazione di nuove sinapsi, indotta da 5-HT, è associata all'abbassamento della concentrazione delle molecole di adesione neuronale apCAM, strutturalmente simili a NCAM (Neuronal cell adhesion molecule), sulla superficie della membrana dei neuroni sensoriali. Il fenomeno della diminuzione di concentrazione è particolarmente cospicuo nei siti presso i quali i processi dei neuroni sensoriali vengono a contatto l'uno con l'altro e si realizza grazie all'attivazione, dipendente dalla sintesi proteica, di un programma coordinato di endocitosi mediata dalla clatrina, che conduce all'internalizzazione e all'apparente degradazione di apCAM (fig. 3). I neuroni di Aplysia esprimono due isoforme di apCAM, una transmembrana e una legata al fosfoinositolo. Solo l'isoforma transmembrana viene internalizzata in seguito a esposizione a 5-HT.
L'internalizzazione selettiva della forma transmembrana mette in evidenza la potenziale importanza di regolazione del suo dominio intracellulare in cui è presente una sequenza ricca in prolina, acido glutammico, serina e treonina (sequenza PEST), che sembra mediare la degradazione proteica e che possiede due sequenze di consenso per la fosforilazione della MAP-chinasi. Mediante la sovraespressione in neuroni sensoriali di costrutti chelati o mutanti a livello transmembrana, è stato dimostrato che la 5-HT induce un abbassamento di concentrazione a livello della membrana citoplasmatica ed è stato possibile bloccare la concomitante internalizzazione eliminando l'intera coda citoplasmatica della proteina, oppure unicamente la regione intermedia contenente la sequenza PEST, o ancora sostituendo semplicemente la treonina con l'alanina nelle sequenze di consenso della MAP-chinasi. Questi risultati suggeriscono un possibile ruolo della MAP-chinasi nella modulazione di apCAM. Si ritiene che la diminuzione di apCAM indotta da 5-HT comporti almeno due conseguenze principali: la defascicolazione, cioè la disarticolazione dei fascicoli associati in maniera omofilica dei neuroni sensoriali, un processo che può destabilizzare i contatti adesivi e inibire, di norma, la crescita; l'attivazione endocitotica, che può condurre a una ridistribuzione dei componenti di membrana nei siti di formazione delle nuove sinapsi. Di conseguenza, gli aspetti delle fasi iniziali della crescita delle connessioni sinaptiche associate all'apprendimento, che costituiscono una caratteristica del processo a lungo termine, potranno essere compresi in futuro nel contesto di una forma nuova e mirata di endocitosi mediata da recettori.
Consolidamento della memoria a lungo termine
Gli studi condotti su Aplysia indicano che parte dell'interruttore molecolare necessario per la fase di consolidamento della memoria a lungo termine, durante le forme implicite ed elementari di apprendimento, comporta l'induzione mediata da cAMP dei geni precoci. La domanda da porsi è se nel cervello esista, allo stesso modo, un insieme di eventi molecolari capace di contribuire alla formazione di un magazzino mnemonico a lungo termine, per forme più complesse di apprendimento esplicito. Le forme esplicite di apprendimento, tanto negli esseri umani quanto negli animali da esperimento, dipendono in larga misura dalle strutture del lobo temporale, compreso l'ippocampo. Il problema di quali siano i meccanismi cellulari che vengono utilizzati all'interno dell'ippocampo per l'immagazzinamento della memoria esplicita è stato posto per la prima volta quando Timothy V.P. Bliss e Terje Lømo (1973) dimostrarono che i neuroni dell'ippocampo possiedono le caratteristiche di plasticità e durata necessarie per l'immagazzinamento della memoria a lungo termine. Una breve stimolazione ad alta frequenza in una delle tre vie neuronali meglio caratterizzate all'interno dell'ippocampo produce un aumento dell'efficienza sinaptica che può permanere per ore o giorni. Questo rafforzamento viene definito potenziamento a lungo termine o LTP (Long term potentiation).

Nell'ippocampo dei Mammiferi, l'LTP possiede alcuni dei meccanismi propri della facilitazione sinaptica di Aplysia. Per esempio (fig. I.A), l'LTP delle fibre muscoidi, che si manifesta a livello delle sinapsi tra le cellule granulari della regione dentata e le cellule piramidali della regione CA3, comporta un rafforzamento, dipendente da cAMP, del rilascio del trasmettitore dai terminali presinaptici. Per contro, l'LTP della via collaterale di Schaffer nella regione CA1 è molto più complesso; in questo caso l'induzione è un evento postsinaptico che comporta l'ingresso del calcio attraverso il canale del recettore NMDA (N-metil-D-aspartato) e l'utilizzazione di numerose vie modulate da secondi messaggeri che coinvolgono la tirosina-chinasi, la proteina-chinasi C e la chinasi II Ca2+/calmodulina-dipendente. In aggiunta a queste prime fasi che si verificano nella cellula postsinaptica, l'LTP della via collaterale di Schaffer comporta un rafforzamento del rilascio del trasmettitore dal neurone presinaptico che può essere mediato da messaggeri retrogradi, forse monossido di azoto o monossido di carbonio, i quali si diffondono dalla cellula postsinaptica.
Nei Roditori l'ippocampo riveste un ruolo fondamentale per la memoria spaziale, come è stato verificato per mezzo di numerosi modelli. L'uso di tecniche di ingegneria genetica per ottenere ceppi di topi knock out, nei quali è stato soppresso un gene specifico, o transgenici nei quali si è introdotto un gene nuovo, ha permesso ai ricercatori di esplorare, dal punto di vista genetico, il ruolo di diverse vie metaboliche di trasduzione del segnale nell'LTP e nella memoria a lungo termine. Anche le conclusioni raggiunte con studi farmacologici precedenti, che hanno suggerito l'importanza della proteina_-chinasi C, della chinasi II Ca2+/calmodulina-dipendente e della tirosina-chinasi per l'induzione dell'LTP, sono state convalidate ampiamente da questi studi genetici. Tali ricerche hanno sottolineato l'importanza dell'LTP nella via collaterale di Schaffer per l'apprendimento spaziale. Infatti, nei topi mutanti con gravi menomazioni all'LTP della via collaterale di Schaffer si ha un apprendimento spaziale ridotto. Per contro, nei topi mutanti con una menomazione selettiva alla via delle fibre muscoidi si ha un apprendimento normale.
Analogamente alla facilitazione a lungo termine di Aplysia, l'LTP che coinvolge la via collaterale di Schaffer è caratterizzato da fasi temporali distinte. La fase precoce è prodotta da una stimolazione tetanica singola, ha una durata di 1÷3 h e richiede solo la modificazione covalente delle proteine preesistenti. Per contro, la fase tardiva è indotta da una stimolazione tetanica ripetuta, persiste per varie ore ed è dipendente da una nuova sintesi proteica e di RNA. Come per la memoria a lungo termine di Aplysia, anche nell'LTP la trascrizione richiede una finestra temporale critica. Inoltre, la fase tardiva, dipendente dalla trascrizione (figg. I.D, E) dell'LTP nella via collaterale di Schaffer, è bloccata dagli inibitori della PKA. Importanti studi condotti da P.V. Nguyen e E.R. Kandel (1996) indicano che questi aspetti dell'LTP sono individuabili anche in una terza via ippocampale principale, la via perforante mediale che origina nella corteccia entorinale e termina nelle cellule granulari della regione dentata. In questa via l'LTP presenta sia una fase precoce, transitoria, che non necessita di sintesi di proteine o di RNA ed è indipendente dall'attivazione della PKA, sia una fase tardiva, nella quale è necessaria la sintesi di proteine e di RNA e che può essere bloccata selettivamente dagli inibitori della PKA (figg. I. B, C). Quindi, così come nella facilitazione presinaptica di Aplysia, la trascrizione mediata da cAMP sembra essere un meccanismo comune nella forma tardiva di LTP in tutte e tre le vie all'interno dell'ippocampo.
La memoria a lungo termine come sequenza di stadi molecolari
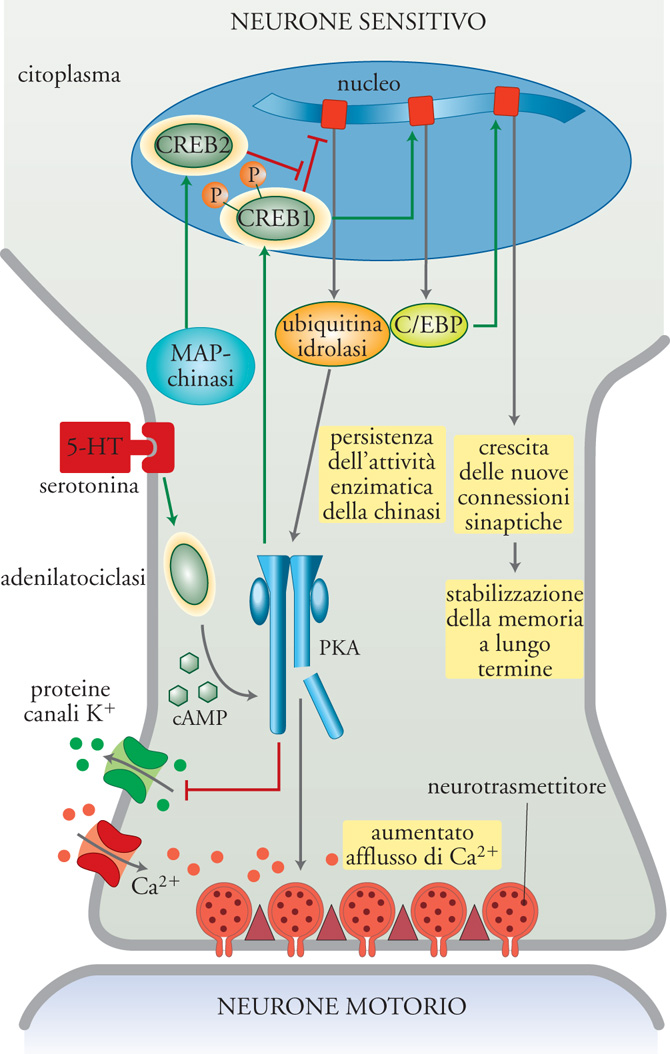
In termini molecolari, la Tav. II illustra schematicamente i tre stadi molecolari, che abbiamo delineato nella conversione della facilitazione presinaptica da breve a lungo termine, in Aplysia. In questo modello la 5-HT, un trasmettitore modulatorio rilasciato dagli interneuroni facilitanti che estendono sinapsi sui neuroni sensoriali, agisce in modo da avviare processi mnemonici separati con durata differente. La facilitazione a breve termine, che ha una durata dell'ordine di minuti, inizia con il legame della 5-HT ai suoi recettori di membrana. Ciò attiva l'adenilatociclasi che catalizza la sintesi di cAMP; questo si lega alla subunità regolatrice della PKA conducendo alla liberazione e all'attivazione della sua subunità catalitica. La PKA agisce su almeno due classi di substrati per aumentare il rilascio del trasmettitore; in primo luogo procede alla fosforilazione dei canali per K+ o di proteine legate a essi, provocando una riduzione della corrente di uscita di K+ e producendo un allungamento nel tempo del potenziale di azione e un aumentato ingresso di Ca2+ nel neurone presinaptico. Inoltre, la PKA sembra agire direttamente sul sistema molecolare coinvolto nel meccanismo di esocitosi del trasmettitore. Queste modificazioni si verificano nei terminali presinaptici, sono indipendenti dalla nuova sintesi macromolecolare e la loro durata determina la persistenza della facilitazione a breve termine.
Contrariamente a quanto si verifica per gli effetti a breve termine, l'attivazione ripetuta di interneuroni serotoninergici avvia una facilitazione a lungo termine di durata superiore a un giorno, per la quale è necessaria la sintesi di nuove proteine. Con l'applicazione ripetuta o prolungata di 5-HT, la subunità catalitica della PKA si trasferisce nel nucleo, dove agisce su substrati nucleari che includono fattori di trascrizione della famiglia CREB. La facilitazione a lungo termine si attiva grazie alla regolazione coordinata di almeno due fattori di trascrizione espressi costitutivamente: la PKA conduce all'attivazione di CREB1; altri sistemi di chinasi, quali la MAP-chinasi, possono esercitare un'azione di derepressione per mezzo di CREB2. Il coordinamento di attivazione e derepressione produce la rapida induzione di una rete di geni precoci durante il periodo di consolidamento della facilitazione a lungo termine. Alcuni di questi geni precoci codificano proteine regolatrici trascrizionali che, a loro volta, sembrano essere indotte a esprimere geni tardivi responsabili del periodo di stabilizzazione. Oltre ai fattori di regolazione, anche gli effettori precoci vengono sintetizzati durante la fase di consolidamento. Tra questi effettori si trovano l'ubiquitinaidrolasi carbossiterminale, che sembra partecipare alla divisione proteolitica della subunità regolatrice della PKA, in quanto mantiene l'attività enzimatica della subunità catalitica in assenza di aumento del cAMP, e le catene leggere di clatrina, che possono partecipare alla rimozione della apCAM dalla superficie cellulare per mezzo dell'attivazione del percorso endocitotico, contribuendo così agli stadi iniziali della crescita sinaptica.
Pur essendo necessaria per le prime dieci ore successive all'applicazione ripetuta di 5-HT, l'attività enzimatica della PKA non viene conservata. Al contrario, la fase tardiva è caratterizzata da cambi strutturali che si manifestano nel giro di 1 h dopo il trattamento con 5-HT o dopo l'addestramento per mezzo di scossa alla coda, e persistono per giorni o settimane. Infatti i geni regolatori indotti possono, a loro volta, avviare ulteriori cicli di attivazione trascrizionale, generando una cascata di espressione genica sequenziale che interessa le proteine di codificazione genica, quali la BiP (Binding protein) e la calreticulina, che possono riflettere una risposta generale destinata ad andare incontro alle richieste post-traduzionali di un aumento della sintesi proteica, nonché di proteine strutturali necessarie alla costruzione di nuove ramificazioni sinaptiche. La memoria che permane per ore è conservata per mezzo di proteine effettrici o di loro modificazioni funzionali, per esempio quando vengono fosforilate. Alcune proteine effettrici precoci possono anche servire a rinforzare e a conservare la risposta iniziale, per esempio per mezzo dell'attivazione proteolitica di una proteina-chinasi. La memoria che permane per giorni, settimane o mesi (più a lungo di metà della vita delle proteine effettrici) viene attivata da geni regolatori precoci i cui prodotti proteici attivano l'espressione conservata dei geni effettori tardivi necessari alla crescita delle nuove connessioni sinaptiche che stabilizzano le fasi finali della memoria a lungo termine.
Bibliografia
Alberini 1994: Alberini, Cristina e altri, C/EBP is an immediate-early gene required for the consolidation of long-term facilitation in Aplysia, "Cell", 76, 1994, pp. 1099-1114.
Bailey, Chen 1983: Bailey, Craig H. - Chen, Mary, Morphological basis of long-term habituation and sensitization in Aplysia, "Science", 220, 1983, pp. 91-93.
Bailey, Chen 1988: Bailey, Craig H. - Chen, Mary, Long-term memory in Aplysia modulates the total number of varicosities of single identified sensory neurons, "Proceedings of the National Academy of Sciences USA", 85, 1988, pp. 2373-2377.
Bliss, Lømo 1973: Bliss, Timothy V.P. - Lømo, Terje, Long-lasting potentiation of synaptic transmission in the dentate area of the anaesthetized rabbit following stimulation of the perforant path, "Journal of physiology", 232, 1973, pp. 331-356.
Nguyen, Kandel 1996: Nguyen, Phu V. - Kandel, Eric R., A macromolecular synthesis-dependent late phase of long-term potentiation requiring cAMP in the medial perforant pathway of rat hippocampal slices, "Journal of neuroscience", 16, 1996, pp. 3189-3198.
Squire, Zola-Morgan 1991: Squire, Larry R. - Zola-Morgan, Stuart, The medial temporal lobe memory system, "Science", 253, 1991, pp. 1380-1386.