
Cellula
Cellula
di Jean Brachet, Massimo Aloisi
Cellula
Fisiologia della cellula
di Jean Brachet
Sommario: 1. Introduzione storica. Metodi per lo studio della cellula. 2. Il citoplasma: a) membrana cellulare; b) reticolo endoplasmatico; c) mitocondri; d) lisosomi; e) perossisomi; f) corpi di Golgi; g) granuli di secrezione (zimogeno); h) centrosomi (centrioli) e cinetosomi (corpi basali); i) ciglia e flagelli; l) microtubuli; m) materiali di riserva; n) cloroplasti. 3. Il nucleo: a) composizione chimica del nucleo; b) membrana nucleare; c) cromatina; d) nucleoli; e) succo nucleare; f) il nucleo batterico. 4. Divisione cellulare (mitosi): a) cromosomi; b) apparato mitotico; c) anormalità mitotiche; d) variazioni biochimiche durante la mitosi; e) inibizione e stimolazione della mitosi; f) meccanismi operanti nella mitosi. 5. Interazioni nucleocitoplasmatiche: a) lavori su cellule integre; b) lavori su frammenti nucleati e anucleati; c) conclusioni. 6. Differenziamento della cellula. □ Bibliografia.
1. Introduzione storica. Metodi per lo studio della cellula
È cognizione comune che la cellula e l'unità fondamentale di cui sono costituiti tutti gli organismi viventi (dai Batteri agli uomini). Ma questo concetto è relativamente nuovo se paragonato alla teoria atomica elaborata da filosofi come Democrito, Epicuro, Lucrezio: non esisteva ragione filosofica per supporre l'esistenza di unità biologiche, mentre concetti come forza vitale, entelechia e altri fornivano una facile spiegazione delle differenze fra gli organismi viventi e la materia inerte. Si può dire che il concetto di ‛cellula' ebbe origine dal progressivo sviluppo degli strumenti ottici, particolarmente dei microscopi, che consentono ingrandimenti fino a 3.000 diametri: esso è quindi piuttosto il risultato del progresso tecnologico che del pensiero filosofico.
La parola ‛cellula' fu coniata da R. Hooke nel 1665; dopo aver osservato dei sugheri con una semplice lente, egli chiamò cellule le cavità che formano il sughero poroso, cavità che non hanno niente a che vedere con quelle che noi oggi chiamiamo cellule. Si attribuisce generalmente la paternità della ‛teoria cellulare', nel suo significato moderno, al botanico M. J. Schleiden e allo zoologo Th. Schwann, che la formularono nel 1838-1839. Le loro osservazioni al microscopio dimostravano che sia le piante, sia gli animali erano costituiti da cellule: queste erano sempre divise in due parti, un nucleo centrale (già visto da R. Brown nel 1833) e un citoplasma circostante. In realtà, cellule furono osservate molto prima, all'epoca in cui i microscopi composti furono costruiti e usati da Leeuwenhoek, Hartsoeker, Malpighi, ecc.: gli ‛animalculi' (Protozoi unicellulari, spermatozoi) che essi descrivevano erano infatti cellule, provviste di nucleo e di citoplasma. La più grande cellula vivente (o piuttosto organismo unicellulare) è l'alga gigante Acetabularia, che può essere lunga più di cinque centimetri e che deve essere stata vista nel Mediterraneo già dai pescatori della preistoria; ma soltanto in questo secolo il microscopio ha mostrato che questa grande alga possiede un unico nucleo e un citoplasma continuo. Essa è, quindi, un'autentica cellula.
Un enorme progresso nella comprensione del significato del concetto di cellula fu fatto quando R. Virchow, nel 1858-1859, dimostrò che tutte le cellule necessariamente derivano, per scissione, da cellule preesistenti: il concetto omnis cellula e cellula esclude la possibilità della formazione di cellule per generazione spontanea. I moderni progressi della tecnica (conservazione delle strutture cellulari per mezzo di ‛fissazione' chimica, colorazione dei costituenti della cellula con coloranti vari) rendono visibili le diverse fasi della mitosi, cioè della scissione cellulare: i lavori di E. van Beneden, Th. Boveri, W. Flemming, O. Hertwig, E. Strasburger e altri portarono alla scoperta dei cromosomi e dimostrarono che questi corpi filamentosi, che sono presenti nei nuclei di tutte le cellule, vengono divisi in parti eguali nelle cellule figlie durante la mitosi. Questa importante scoperta fu all'origine del principio di continuità genetica, che venne decisamente stabilito quando O. Hertwig (1875) descrisse la fusione (‛anfimissi') dei nuclei dell'uovo (pronucleo femminile) e dello spermatozoo (pronucleo maschile) al momento della fecondazione, e quando van Beneden (1885) dimostrò la divisione dei cromosomi durante la scissione dell'uovo fecondato.
Progrediti metodi di fissazione e di colorazione citologici e istologici (rivolti allo studio rispettivamente delle cellule e dei tessuti formati da cellule della stessa ‛famiglia') portarono alla scoperta di molti organuli della cellula, dei quali ci occuperemo diffusamente in questo stesso articolo: la cr0matina e il nucleolo nei nuclei a riposo, non in fase di divisione; i mitocondri, i lisosomi, i corpi di Golgi, i centrosomi, i cinetosomi, ecc., nel citoplasma. Nonostante ciò, durante il primo trentennio di questo secolo si ebbero forti contrasti circa la presunta esistenza di questi organuli: il lavoro di J. Loeb sui colloidi ha dimostrato che, quando le proteine sono precipitate da acidi, sublimato, ecc., formano filamenti e granuli che hanno una qualche rassomiglianza con le strutture osservate nelle sezioni fissate e colorate di cellule in riposo e in divisione. La tendenza a considerare tutti gli organuli della cellula come artefatti prodotti dalla fissazione portò un eminente citologo come E. B. Wilson a elaborare, nel 1922, lo schema estremamente semplificato di una tipica cellula, rappresentato nella fig. 1.
La citologia ha subito una nuova rivoluzione con l'av- vento del microscopio elettronico, specialmente per l'influenza di A. Claude e dei suoi colleghi K. Porter e G. Palade: sono stati introdotti metodi di fissazione più raffinati, che permettono una conservazione decisamente migliore delle delicate strutture della cellula (v. fig. 2).
Con le enormemente accresciute possibilità di ingrandimento (più di 100.000 diametri) e di risoluzione (5 Å o anche meno) dei moderni microscopi elettronici, l'analisi ora raggiunge un livello che permette anche l'osservazione delle macromolecole (acidi nucleici, proteine, glicogeno, ecc.). La fig. 3 rappresenta un moderno schema, molto semplificato, di una cellula tipica: come vedremo, i mitocondri sono i siti delle reazioni ossidative che forniscono energia alla cellula; i lisosomi sono vacuoli digestivi intracellulari che frammentano le grosse molecole; il reticolo endoplasmatico (un intreccio intracellulare di canali) è coperto da minuscoli granuli, i ribosomi, ove avvengono le sintesi proteiche. La struttura del nucleo, d'altra parte, non ha subito cambiamenti così profondi dall'epoca di E. B. Wilson: l'unica differenza è che il reticolo di cromatina dei vecchi citologi è ora considerato un artefatto, poiché oggi riteniamo che la cromatina riempia l'intero nucleo in modo pressoché omogeneo. Nei nuclei della maggior parte delle cellule sono osservabili soltanto locali addensa- menti della cromatina; il resto è riempito con fibrille di cromatina lassa.
Come siamo riusciti a sapere, per esempio, che i mitocondri svolgono un ruolo importante nella respirazione e nella produzione di energia? Lo sappiamo perché si è sviluppata una nuova e potente tecnica, che consente analisi biochimiche di organuli cellulari isolati: è la centrifugazione differenziale di cellule omogeneizzate (A. Claude). Le cellule vengono frantumate meccanicamente in un mezzo appropriato (di solito una soluzione di saccarosio) e l'omogenato risultante viene centrifugato a velocità gradatamente crescenti: in questo modo è possibile riunire, in forme purificate, nuclei cellulari, nucleoli, mitocondri, lisosomi, frammenti di reticolo endoplasmatico, ribosomi liberi, corpi di Golgi, frammenti della membrana cellulare. Queste frazioni possono essere analizzate con metodi biochimici, che sono in grado di dare precise informazioni circa la composizione chimica (contenuto in acidi nucleici, proteine - inclusi tutti i tipi di enzimi - lipidi, ecc.) degli organuli cellulari isolati.
Anche metodi meno drastici possono dare informazioni estremamente importanti sulla composizione chimica dei costituenti della cellula: sono i metodi citochimici, in cui ben note e specifiche reazioni chimiche sono applicate su cellule viventi o loro sezioni microscopiche. Essi si rivelano di grande utilità per individuare la localizzazione intracellulare di macromolecole o di enzimi; in alcuni casi, è possibile perfino effettuare misurazioni quantitative, purché si disponga di apparecchiature adeguate. Il grande vantaggio della citochimica è di dare informazioni su singole cellule; i metodi biochimici sono invece di solito applicati a una grande ed eterogenea popolazione cellulare. I metodi citochimici si sono dimostrati particolarmente utili per la individuazione degli acidi nucleici: la reazione di Feulgen, basata sulle proprietà chimiche dello zucchero desossiribosio, è ideale per rivelare la presenza degli acidi desossiribonucleici (DNA). Gli acidi nucleici possono essere anche individuati con semplici metodi di colorazione, sia con coloranti basici (J. Brachet), sia per mezzo del loro assorbimento nell'ultravioletto (T. O. Caspersson); i due tipi di acidi nucleici (DNA, e acidi ribonucleici: RNA) possono facilmente distinguersi l'uno dall'altro quando le sezioni vengano trattate prima della colorazione o della fotografia all'ultravioletto - con enzimi che specificamente digeriscano l'uno o l'altro tipo di acido nucleico (desossiribonucleasi o ribonucleasi, rispettivamente).
È altresì possibile svelare, mediante semplici reazioni coloranti, proteine acide e basiche e perfino visualizzare al microscopio proteine specifiche con un metodo più elaborato: anticorpi fluorescenti sono preparati contro una proteina pura specifica e le cellule contenenti questa proteina vengono colorate con questi anticorpi; la localizzazione intracellulare della proteina antigenica può poi essere osservata con un microscopio a fluorescenza. Molti enzimi, sia ossidativi che idrolitici, possono essere individuati e localizzati nelle cellule per mezzo di opportune reazioni (v. enzimi). La determinazione citochimica del glicogeno, dell'amido e dei lipidi non offre particolari difficoltà.
Le classiche tecniche citochimiche hanno lo svantaggio di fornire molto spesso soltanto informazioni di natura statica, essendo di solito applicate a cellule fissate, uccise. Ma un'altra tecnica citochimica, l'autoradiografia, ci consente di ottenere informazioni su processi dinamici, come il sito delle sintesi delle macromolecole, o il movimento nella cellula di sostanze appena sintetizzate (per es., dal nucleo al citoplasma). Il principio è semplice: si fanno incorporare a cellule viventi specifici precursori radioattivi in macromolecole di neoformazione (nel caso di proteine sarà usato un amminoacido, nel caso dell'RNA e del DNA, invece, sarà usato un nucleoside, come uridina o timidina, rispettivamente). La localizzazione intracellulare della proteina o dell'acido nucleico appena sintetizzati sarà poi determinata per mezzo di un semplice procedimento fotografico.
Negli ultimi anni, vari metodi citochimici (compresa l'autoradiografia) sono stati adattati alla microscopia elettronica: è divenuta in tal modo possibile un'analisi della localizzazione delle macromolecole e della loro sintesi a livello ultrastrutturale. Molti enzimi possono ora essere individuati al microscopio elettronico ed è molto probabile che, fra pochi anni, i singoli ‛geni' (cioè, segmenti specifici di enormi molecole di DNA) potranno essere osservati in fotografie ottenute mediante il microscopio elettronico: in realtà, ci si è già riusciti nel caso dei geni che controllano la formazione dei ribosomi. Lo studio della cellula, la citologia, in poco più di cento anni dall'epoca di M. J. Schleiden e Th. Schwann, ha ora raggiunto lo stadio ‛molecolare' del suo sviluppo.
2. Il citoplasma
Gli schemi delle figg. 1 e 2 furono ideati per rappresentare cellule ‛tipiche'. Occorre subito precisare che non esiste nulla che si possa considerare come una cellula ‛tipica': cellule provenienti da differenti organi o tessuti (per es., una cellula nervosa e una cellula muscolare, uno spermatozoo e un uovo) hanno tutte una forma molto differente, così come differenti sono le strutture, le funzioni e la composizione biochimica. Le cellule si sono adattate alle loro funzioni e questa è l'essenza del differenziamento cellulare, che sarà brevemente discusso in seguito. Un'altra osservazione che va subito fatta è che queste differenze da cellula a cellula sono di solito più evidenti se si prende in considerazione il citoplasma piuttosto che il nucleo.
a) Membrana cellulare
Le proprietà della membrana cellulare sono oggetto di un interesse sempre crescente; ciò in quanto la complessità della sua struttura e della sua funzione diventa ogni giorno più evidente: il suo ruolo è essenziale, dal momento che la membrana cellulare rende possibili e controlla gli scambi tra la cellula e il mezzo esterno (v. elettrofisiologia; v. trasporto attraverso membrane biologiche). La membrana cellulare è dotata di una proprietà detta ‛permeabilità selettiva': consente cioè la penetrazione nella cellula di sostanze necessarie alle sintesi e alla produzione di energia. Grazie a tale proprietà, la cellula è protetta dalle sostanze tossiche e può eliminare i prodotti finali del proprio metabolismo.
La membrana cellulare è costituita da lipidi e da proteine. La fig. 4 rappresenta il modello proposto da Danielli e Davson nel 1943 per spiegare la struttura e le proprietà fisiologiche della membrana cellulare: un doppio strato di lipidi separa due foglietti di proteine. La microscopia elettronica ha mostrato che la membrana cellulare è veramente formata da un doppio strato; ma indica anche che la sua struttura (nella quale si possono vedere subunità esagonali di proteine) è molto più complessa di quanto non si ritenesse prima. Tra i due strati di lipidi (che sono chimicamente differenti nelle superfici interna ed esterna della membrana cellulare) sono intercalate delle proteine, che sono visibili, al microscopio elettronico, come ‛particelle intermembraniche'; nella superficie esterna della membrana queste proteine contengono residui di zuccheri, che possono combinarsi con proteine di origine vegetale, chiamate lectine. Se queste lectine (in particolare la concavalina A) vengono aggiunte a cellule isolate, si ha aggregazione cellulare nel caso di cellule embrionali o neoplastiche, ma non in quello di cellule differenziate. La membrana cellulare può essere rappresentata come un mare di lipidi, nel quale le proteine si muovono come icebergs. Questa complessità non è sorprendente se si considera l'alta selettività della permeabilità della membrana. Per esempio, le cellule vivono in mezzi che, come il plasma sanguigno, sono ricchi di Na+ e poveri di K+: la membrana riconosce questi due ioni, respingendo l'Na+ e concentrando il K+.
Il funzionamento di questa ‛pompa del sodio' richiede energia, che (come sempre accade nel caso di cellule viventi) trae origine dalla rottura dei legami fosforici ricchi di energia dell'acido adenosin trifosforico (ATP). Tra le proteine che costituiscono la membrana cellulare vi sono enzimi che rompono l'ATP e liberano energia. La membrana stessa contiene anche un gruppo di enzimi che sono necessari per captare il fosfato e il glucosio del mezzo esterno.
Nei Batteri e nei Miceti, specifici enzimi regolano la captazione degli amminoacidi e degli zuccheri: sono le permeasi, che vengono sintetizzate, sotto il controllo di geni specifici, quando il substrato (amminoacido, zucchero) viene aggiunto al mezzo esterno.
Non è noto se le permeasi siano presenti nelle cellule degli organismi superiori. Ma è chiaro che, in cellule non circondate da una spessa parete cellulare protettiva, come i Batteri e i Miceti, sostanze di alto peso molecolare presenti nel mezzo esterno possono giungere nell'interno della cellula per mezzo di un meccanismo (‛endocitosi') che non ha niente in comune con la classica permeabilità: particelle (Batteri, cloroplasti, ecc.) possono essere inglobate, per mezzo di invaginazioni della membrana cellulare, nella cellula ed eventualmente digerite (‛fagocitosi'). Un processo simile (‛pinocitosi'; v. fig. 5) permette la captazione nella cellula di grandi molecole solubili, come proteine o acidi nucleici: gocce del mezzo esterno, contenenti le macromolecole, penetrano nella cellula mediante invaginazioni della membrana cellulare; queste invaginazioni si separano dalla membrana in modo da formare vacuoli intracellulari di pinocitosi. Quando questo processo è molto attivo (come nel caso delle cellule dell'intestino e del rene) la membrana diventa altamente differenziata: al microscopio elettronico si vedono microvilli (v. fig. 6); alla loro estremità basale vi sono numerosi vacuoli di pinocitosi (spesso chiamati ‛fagosomi') pieni di materiali ingeriti.
Alcune cellule vivono liberamente nel mezzo esterno (per es., i globuli rossi e bianchi del sangue), ma la maggior parte si aggrega in maniera da formare una massa solida. Ma come si aggregano? Per la presenza, al di fuori della membrana cellulare, di un rivestimento o matrice che le tiene insieme: questo ‛cemento intercellulare' è costituito da uno speciale tipo di proteine associate con vari zuccheri. Tali glicoproteine, che facilmente fissano il calcio, possono essere distrutte con la digestione per mezzo di enzimi proteolitici (per es., tripsina). Se un tessuto solido è trattato con tripsina o con agenti che rimuovono il calcio, le cellule si liberano. Le cellule dissociate (che si possono eventualmente riaggregare se si rimuove la tripsina o se viene di nuovo aggiunto il calcio) sono ampiamente usate per studi fisiologici e biologici. Per esempio, è stato dimostrato da A. Moscona che cellule dissociate spesso si riaggregano in una maniera tessuto-specifica: se cellule isolate di rene e di retina, provenienti da embrioni di pollo e di topo, vengono messe insieme, le cellule di rene delle due specie si aggregheranno insieme; allo stesso modo si può formare una retina composta da una popolazione mista di cellule di pollo e di topo. Un altro interessante fenomeno rilevato su cellule isolate è quello cosiddetto di ‛inibizione da contatto': quando le cellule sono separate l'una dall'altra, esse possono essere coltivate in un mezzo adatto in cui presentano locomozione e divisione cellulare; ma se due cellule entrano in contatto, sia la locomozione sia l'attività mitotica si arrestano. Questa inibizione da contatto non avviene nelle cellule cancerose, fatto questo che dimostra come la malignità modifichi le proprietà superficiali delle cellule.
Le cellule si mantengono unite non soltanto per la presenza del cemento intercellulare, ma anche perché possono aderire l'una all'altra mediante rinforzi localizzati della loro membrana, i cosiddetti ‛desmosomi' (v. fig. 7). Si ritiene che questi permettano lo stabilirsi di comunicazioni elettriche fra cellule adiacenti.
Infine, il fatto che la membrana cellulare può essere lievemente danneggiata da brevi trattamenti con certi virus (specialmente il virus emolitico Sendai) o con particolari fosfolipidi ha grande importanza: tali trattamenti consentono la ‛fusione' di cellule di diversa origine (per es., una cellula cancerosa umana può fondersi con un globulo rosso del sangue di un uccello) e nuclei di due specie differenti sono presenti contemporaneamente in un citoplasma comune (v. Harris, 1970). Tali eterocarionti (v. fig. 8) sono di considerevole interesse, come vedremo in seguito, per le analisi delle interazioni nucleocitoplasmatiche.
b) Reticolo endoplasmatico
Questo elaborato intreccio di doppie membrane, che fu descritto per la prima volta da Porter e Palade, unisce la membrana cellulare alla membrana nucleare e costituisce una specie di scheletro (un ‛citoscheletro') per la cellula. Il concetto che tutte le membrane della cellula, comprese quelle dei mitocondri, dei lisosomi, ecc., siano continue è stato proposto da J. D. Robertson. Sebbene questa ipotesi sia stata combattuta, essa certamente contiene un buon fondo di verità: infatti dobbiamo considerare le membrane cellulari come sottoposte a continui cambiamenti e trasformazioni; esse sono in una condizione dinamica e non statica.
Come mostra la fig. 9, si possono distinguere due differenti tipi di reticolo endoplasmatico: quello ‛ruvido' in cui le doppie membrane sono ricoperte di piccoli granuli (i ribosomi), e quello ‛liscio' in cui i ribosomi mancano. Sia il reticolo endoplasmatico ruvido sia quello liscio possono essere recuperati, in forma frammentaria, da cellule omogeneizzate mediante ultracentrifugazione; il primo costituisce i ‛microsomi' (A. Claude), che possono essere dissociati in ribosomi liberi e membrane per mezzo del trattamento con detergenti. Le due frazioni separate possono poi essere sottoposte alle analisi chimiche. Le membrane del reticolo endoplasmatico sono costituite di fosfolipidi e proteine. Queste ultime, nel caso del fegato, comprendono un certo numero di enzimi (in particolare, la glucosio-6-fosfatasi) che intervengono nel metabolismo dei carboidrati e dei lipidi (specialmente degli steroli e dei carotenoidi). Contengono anche, nel fegato, un certo numero di enzimi (ossidativi o no) che hanno una parte fondamentale nella funzione di disintossicazione, distruggendo le sostanze tossiche aromatiche (derivati dell'anilina, per esempio). Poiché le membrane lisce sono molto abbondanti nelle cellule che contengono grandi quantità di glicogeno (fegato), ormoni steroidi (testicoli) o vitamina A (retina), è probabile che esse intervengano nella sintesi o nell'immagazzinamento di carboidrati e di alcune specie di sostanze grasse.
L'abbondanza di ribosomi, d'altra parte, è direttamente correlata, come per primi dimostrarono T. O. Caspersson e J. Brachet nel 1940-1941, con l'intensità delle sintesi proteiche. I ribosomi (v. fig. 10) sono costituiti da quantità approssimativamente uguali di proteine e RNA; contengono un gran numero (più di 50) di differenti proteine e tre tipi di RNA ribosomiale (rRNA). Tutti i ribosomi sono composti di due ‛subunità', che possono venire separate sperimentalmente e che prendono il loro nome dalla rispettiva costante di sedimentazione (valore S) nell'ultracentrifuga: le costanti di sedimentazione per i ribosomi animali, le loro subunità grandi e piccole sono rispettivamente di 80 S, 60 S e 40 S. Valori più piccoli (70 S, 50 S e 30 S) si trovano per i ribosomi dei Batteri e le loro subunità. La subunità ribosomiale più grande contiene un rRNA 28 S (peso molecolare: 1.200.000 dalton) e la più piccola un rRNA 18 S (peso molecolare: 700.000 dalton). Inoltre, un RNA 5 S, molto più piccolo, si trova in tutti i ribosomi (peso molecolare 35.000 dalton).
I ribosomi intervengono direttamente nelle sintesi proteiche, ma la funzione dell'rRNA (che rappresenta l'80% dell'RNA totale) non è ancora conosciuta. Tutto quel che sappiamo è che l'integrità dei ribosomi è assolutamente necessaria per il successo delle sintesi proteiche. Andrebbe senza dubbio oltre lo scopo della presente trattazione discutere il complesso meccanismo delle sintesi proteiche (v. biologia; v. proteine), ma un punto essenziale deve essere menzionato: il sito vero e proprio dove avviene la sintesi di una proteina specifica è il ‛polisoma' (v. fig. 11), un aggregato di ribosomi uniti insieme da un filamento di RNA messaggero (mRNA). Questo mRNA è una esatta copia (‛trascrizione'), fatta nel nucleo, del gene (cioè di una definita sequenza di basi in una macromolecola di DNA); esso dirige, nel citoplasma, la sintesi di una particolare proteina (‛traduzione'). L'mRNA si muove dal nucleo al citoplasma trasportando l'informazione genetica che era codificata nel DNA e si lega alle più piccole subunità dei ribosomi. Nelle subunità più grandi, c'è un sito che lega specificamente le molecole di RNA solubile o di trasferimento (comunemente designato in inglese come RNA transfer, tRNA); questi tRNA, che sono di piccola misura (4 S, peso molecolare circa 13.000), portano i vari amminoacidi che costituiranno la catena proteica. Tali combinazioni tra ribosomi, tRNA e mRNA, richiedono l'intervento di parecchi fattori di natura proteica, che sono necessari per l'avvio, l'allungamento e la conclusione di una nuova catena proteica.
Tornando al reticolo endoplasmatico, ci sarebbe da aggiungere che la cellula contiene polisomi sia liberi, sia inseriti sulla membrana: questi ultimi sono spesso specializzati nella sintesi delle proteine che saranno portate fuori dalla cellula (per es., enzimi pancreatici, proteine del sangue sintetizzate dal fegato). L'autoradiografia associata con la microscopia elettronica mette in chiara evidenza che, nel pancreas, gli enzimi (tripsina e altri) sono sintetizzati dai polisomi che stanno attaccati alle membrane del reticolo endoplasmatico ruvido; queste proteine si muovono nel lume del reticolo endoplasmatico e si accumulano dentro le dilatazioni di esso, note come ‛cisterne'. Esse infine formano granuli di secrezione (vedi sotto, È g), che sono versati nel lume dei dotti pancreatici.
c) Mitocondri
Queste particelle a forma di bastoncello si possono vedere con il microscopio ottico nel citoplasma di tutte le cellule, tranne che nei Batteri. Possono essere facilmente osservati, nella cellula vivente, col microscopio a contrasto di fase, o per mezzo della colorazione vitale con il colorante verde Janus; test citochimici per gli enzimi ossidativi dimostrano chiaramente la loro presenza nel citoplasma.
Il microscopio elettronico mostra che i mitocondri hanno sempre la medesima ultrastruttura (v. fig. 12): essi sono circondati da una doppia membrana (esterna e interna); quest'ultima accresce la sua superficie mediante invaginazioni e forma le ‛creste': dentro questa membrana interna ripiegata, c'è la cosiddetta ‛matrice'.
I mitocondri possono essere facilmente isolati mediante la centrifugazione differenziale di omogenati di cellule. La purezza delle frazioni mitocondriali può essere stabilita per mezzo dell'esame chimico di enzimi rivelatori (citocromossidasi, succinicodeidrogenasi, che sono enzimi strettamente mitocondriali) e per mezzo del microscopio elettronico.
I mitocondri isolati contengono fosfolipidi, molte specie di proteine e piccole quantità di acidi nucleici (sia RNA che DNA). Negli ultimi anni, è stato anche possibile analizzare separatamente le membrane interne ed esterne, lo spazio intermembrana e la matrice interna dei mitocondri. La composizione enzimatica di questi quattro ‛compartimenti' dei mitocondri è differente, così che enzimi ‛rivelatori' possono essere usati per studiarli.
Le differenze nella composizione chimica sono importanti ove si confrontino le membrane mitocondriali esterna e interna: mentre la prima presenta alcune somiglianze (ma non una identità) con le membrane lisce del reticolo endoplasmatico, le ‛creste' della membrana interna contengono tutti gli enzimi che sono necessari per la produzione di energia (ossidazione e fosforilazione). L'integrità della struttura mitocondriale e, in particolare, la presenza dei lipidi sono necessarie per l'accoppiamento tra ossidazione e fosforilazione (cioè per la sintesi dell'ATP). Secondo alcuni sperimentatori, la fosforilazione ossidativa avverrebbe in alcune particelle elementari molto piccole (le ‛particelle trasportatrici di elettroni' di D. Green). Questa opinione, però, non è accettata da tutti, perché procedimenti che provocano la scomparsa morfologica di quelle particelle elementari non impediscono necessariamente la fosforilazione ossidativa (v. fosforilazione ossidativa).
Una gran mole di lavoro è stata recentemente dedicata agli acidi nucleici mitocondriali. Mentre per lungo tempo il DNA è stato ritenuto uno specifico costituente del nucleo cellulare, gli studi di questi ultimi anni hanno chiaramente dimostrato la presenza costante nel citoplasma di DNA mitocondriale. Questo DNA si differenzia sotto molti aspetti da quello nucleare, che sarà descritto in seguito (v. sotto, cap. 3, § a): mentre quest'ultimo è costituito da filamenti estremamente lunghi, il DNA mitocondriale (v. fig. 13) è molto spesso costituito da molecole circolari, di un tipo molto più piccolo (5 μ di lunghezza; peso molecolare attorno a 10.106 dalton). Anche il DNA batterico è circolare, ma il suo peso molecolare è molto più alto, dell'ordine di 2,5•109 dalton. Questo significa che un mitocondrio contiene circa 2.000 volte meno ‛informazioni' che un batterio; ne consegue che il DNA mitocondriale non contiene abbastanza informazioni per codificare tutte le proteine presenti nel mitocondrio: alcune di esse devono essere formate dai polisomi e gli mRNA corrispondenti devono essere di origine nucleare. Lavori di genetica e di biochimica hanno confermato queste deduzioni: noi sappiamo, per esempio, che il citocromo c (che svolge una funzione essenziale nel sistema di trasferimento degli elettroni e che è accumulato nei mitocondri) è sintetizzato nel reticolo endoplasmatico sotto il controllo di molti geni nucleari. La stessa cosa è probabilmente vera per tutti gli enzimi della catena respiratoria e attualmente si ritiene che il ruolo principale del DNA mitocondriale sia quello di dirigere la sintesi delle proteine ‛strutturali', non enzimatiche, dei mitocondri. Una piccola parte dell'enzima respiratorio citocromossidasi, comunque, è sintetizzata sotto il controllo diretto del DNA mitocondriale nei lieviti.
Comunque, i mitocondri contengono tutti gli enzimi necessari per la sintesi del DNA, dell'RNA e delle proteine: essi possono duplicare il proprio DNA, e questo avviene quando i mitocondri si dividono. Il DNA mitocondriale può anche essere ‛trascritto', cioè può dare origine a tutti e tre i principali tipi di RNA (mRNA, rRNA e tRNA). In effetti, i mitocondri contengono un piccolo numero di ribosomi che, come i ribosomi batterici, sono più piccoli di quelli citoplasmatici. Essi contengono anche tRNA speciali, che non esistono nei polisomi citoplasmatici e che probabilmente sono una copia del DNA mitocondriale. Gli rRNA mitocondriali sono anche sintetizzati dal DNA mitocondriale, che non ha niente in comune col DNA nucleare. Comunque, è ora chiaro che la maggior parte del DNA mitocondriale è costituita da segmenti che hanno significato genetico: essi non controllano la sintesi di RNA, in altri termini non ‛trascrivono'. Questi segmenti inattivi, che sono stati chiamati ‛spaziatori', hanno subito nel corso dell'evoluzione enormi cambiamenti: per questa ragione il DNA mitocondriale può essere molto differente, nella sua composizione chimica, quando si esaminino due specie di rane tanto correlate tassonomicamente che i loro ibridi siano vitali e fertili.
Mitocondri isolati, come abbiamo detto, sono capaci di sintesi proteica. Questa sintesi differisce, sotto molti aspetti, dalla sintesi proteica polisomale ed è molto simile alla sintesi proteica batterica. Per esempio, la sintesi proteica mitocondriale, come quella batterica, è molto sensibile all'antibiotico cloramfenicolo, che invece non ha effetto sulla sintesi proteica polisomale; l'inverso accade per la puromicina, che è un inibitore molto attivo della sintesi proteica nei polisomi, ma non ha alcun effetto sulla sintesi proteica mitocondriale.
È opportuno ricordare che certi farmaci, usati per uccidere i tripanosomi (i quali posseggono un solo grande mitocondrio), possono specificamente distruggere il DNA mitocondriale (v. fig. 14). Per esempio, cellule di lievito, trattate con acriflavina o con bromuro di etidio, formano colonie più piccole delle cellule normali nello stesso mezzo ambientale; le dimensioni ridotte di queste ‛piccole' colonie sono dovute alla totale assenza di enzimi respiratori (B. Ephrussi). Il DNA mitocondriale, in queste colonie ‛piccole', diventa sempre più piccolo e la sua composizione chimica diventa sempre più anormale quanto più la dimensione delle colonie diminuisce. Al microscopio elettronico, nei casi estremi della mutazione citoplasmatica delle ‛piccole' colonie, si possono vedere soltanto le ombre dei mitocondri, formate da poche membrane.
Si può concludere che il DNA mitocondriale, anche se contiene soltanto poche informazioni genetiche, si comporta in molti casi come il DNA nucleare. Questa conclusione è resa più valida dalle recenti osservazioni di Slonimski: la sua analisi genetica indica che le cellule di lievito contengono due distinte popolazioni di mitocondri, che hanno due ‛sessi' differenti (+ e −). I mitocondri, come i Batteri, sarebbero capaci di accoppiamento e ricombinazione genetica. È chiaro che i mitocondri e i Batteri hanno molto in comune: il concetto che i mitocondri sono batteri simbiotici, modificati nel corso dell'evoluzione, non è da considerare fantastico o assurdo.
d) Lisosomi
Particelle lievemente più leggere dei mitocondri sono state isolate da omogenati di fegato da C. De Duve nel 1949. Egli scoprì che esse sono assolutamente differenti dai mitocondri per quello che riguarda il loro contenuto enzimatico; non contengono enzimi respiratori, ma idrolitici (proteasi, fosfatasi acida, desossiribonucleasi, ribonucleasi, ecc.) capaci di frammentare grandi molecole come proteine e acidi nucleici. Questi enzimi idrolitici (che hanno sempre un optimum di attività in ambiente acido, a pH 5) sono presenti in forma ‛latente': essi vengono liberati e diventano attivi quando la membrana che li circonda viene distrutta dai detergenti. A questo ‛sacchetto' di enzimi, racchiusi in una membrana, De Duve diede il nome di ‛lisosoma'. Egli per primo considerò i lisosomi come dei ‛sacchetti suicidi', intendendo che se gli enzimi lisosomiali vengono liberati dalla loro membrana, la morte della cellula (citolisi) sarà inevitabile. Lavori più recenti portarono De Duve e i suoi collaboratori a una differente interpretazione: ai lisosomi si deve la digestione intracellulare del materiale penetrato nella cellula mediante pinocitosi; essi possono anche digerire organuli cellulari (mitocondri, per esempio) in fase di degenerazione. Essi quindi hanno una funzione importante e universale nell'economia della cellula, con la distruzione, per digestione, di sostanze di rifiuto, seguita dalla eliminazione di residui solidi (nei dotti biliari, per esempio): grazie a loro la cellula è mantenuta ‛pulita'.
I lisosomi sono visibili, al microscopio elettronico, come ‛corpi densi', pieni dei residui della parziale digestione (v. fig. 15). Tutti i lisosomi possono facilmente essere individuati, sia al microscopio ottico sia a quello elettronico, per la loro attività fosfatasica acida. Questo enzima è anche un rivelatore molto utile della frazione lisosomiale negli esperimenti biochimici fatti su omogenati.
I lisosomi sono presenti in tutti i tipi di cellule; sono particolarmente abbondanti nel fegato e nelle cellule che hanno forti attività fagocitiche (globuli bianchi del sangue, macrofagi).
L'origine dei lisosomi e le loro relazioni con gli altri sistemi di membrane intracellulari (reticolo endoplasmatico, apparato reticolare di Golgi) sono troppo complesse per essere discusse qui particolareggiatamente (per una recente rassegna sintetica, v. Jacques, 1969). Le tappe principali sono la formazione dei lisosomi primari, derivanti probabilmente dal reticolo endoplasmatico, e la loro fusione con gocce di pinocitosi (‛fagosomi', contenenti materiale che dovrà essere digerito), in maniera da formare i lisosomi secondari, dove avviene la digestione. Se, dopo la digestione, i lisosomi restano pieni di residui solidi, essi diventano ‛corpi residui', densi al microscopio elettronico. Quando digeriscono organuli cellulari in degenerazione, vengono chiamati ‛autofagosomi' o ‛citolisosomi' (v. fig. 16).
Alcune cellule contengono quelli che chiamiamo lisosomi modificati o specializzati; per esempio, quando proteine eterogenee sono iniettate nel sangue, esse vengono riassorbite dal rene, dove formano delle gocce (fagosomi), in cui gli enzimi lisosomiali si accumulano e digeriscono la proteina anomala (W. Straus). Le uova contengono vacuoli che assumono la colorazione vitale con coloranti basici e contengono fosfatasi acida: questi ‛corpi multivescicolari' (v. fig. 17) intervengono nella formazione delle ‛placchette vitelline', che trattengono alcuni degli enzimi lisosomiali. Le teste di molti spermatozoi contengono uno speciale granulo, l'‛acrosoma', pieno di enzimi idrolitici che, liberati al momento della fecondazione, servono alla digestione delle membrane che proteggono l'uovo. I vacuoli digestivi dei Protozoi, le granulazioni neutrofile dei leucociti che si rompono durante la fagocitosi, sono anche lisosomi modificati e specializzati.
e) Perossisomi
De Duve riuscì anche a isolare e a identificare ‛microcorpi' (vacuoli che contengono al centro una inclusione di aspetto cristallino) osservati nel fegato con la microscopia elettronica. Essi contengono un'altra classe di enzimi, che ossidano substrati come amminoacidi o acido urico in maniera ‛diretta', con formazione di perossido di idrogeno (H2O2). Essi contengono anche catalasi, l'enzima che decompone il perossido d'idrogeno e per questo motivo De Duve li chiamò ‛perossisomi'. Sono stati riscontrati in molte cellule animali e vegetali (ma non in tutte), e sembra che in queste ultime essi abbiano un'importante funzione biochimica.
f) Corpi di Golgi
La presenza di un reticolo nelle cellule nervose fu descritta, per la prima volta, da C. Golgi (1898) che usò, per la sua individuazione, una delicata tecnica citologica. In seguito si scoprì che qualcosa di simile si può osservare, in alcune cellule viventi, mediante il microscopio a contrasto di fase, sotto forma di granuli rifrangenti che furono chiamati ‛dittosomi' o ‛corpi di Golgi'.
La struttura di un corpo di Golgi (v. fig. 18) è molto complessa: esso è formato da una certa quantità di membrane (‛lamelle') spesse e lisce, organizzate in ‛tubuli', che terminano poi in ‛sacculi'. Da questi ultimi si distaccano piccole ‛vescicole' di Golgi che rimangono libere nel citoplasma.
I rapporti tra i corpi di Golgi e le altre membrane intracellulari (lisosomi, reticolo endoplasmatico) sono attualmente oscuri e i pareri espressi dai diversi cultori di microscopia elettronica sono ancora troppo contraddittori per essere qui discussi. Una cosa è, comunque, certa: i corpi di Golgi svolgono un'importante funzione nell'accumulo e probabilmente nella sintesi delle glicoproteine, le quali formano la matrice che tiene insieme le cellule (v. sopra, § a). Questo è stato provato sia dalle analisi citochimiche, sia da esperienze biochimiche effettuate su corpi di Golgi isolati: essi contengono un enzima rivelatore che interviene nella sintesi delle glicoproteine (la cosiddetta ‛galattosiltransferasi').
Nelle cellule delle piante, i corpi di Golgi si accumulano, alla fine della divisione cellulare, nella regione dove si formerà la nuova parete cellulare (v. fig. 19). I corpi di Golgi sono anche siti di accumulo (deposito) di sostanze (proteine, lipidi e glicogeno) che sono state prodotte in eccesso dalla cellula.
g) Granuli di secrezione (zimogeno)
Grossi granuli si vedono spesso nelle ghiandole secretive all'apice terminale della cellula: contengono il prodotto (finito o no) della secrezione. Nel pancreas, per esempio, si può dimostrare con metodi immunologici che questi granuli di secrezione contengono i principali enzimi pancreatici (tripsina, chimotripsina ecc.). Abbiamo già visto che questi enzimi sono sintetizzati dai polisomi aderenti al reticolo endoplasmatico; si dirigono nelle cisterne del reticolo endoplasmatico, quindi vanno ai corpi di Golgi e infine si fondono insieme per formare i granuli di zimogeno, senza una ulteriore sintesi proteica. I granuli di secrezione giungono infine alla membrana cellulare e il loro contenuto è espulso nei dotti secretori: questo processo di esocitosi - che è probabilmente mediato da enzimi proteolitici - è l'inverso di quello già menzionato della pinocitosi.
h) Centrosomi (centrioli) e cinetosomi (corpi basali)
Questi due tipi di granuli esercitano nella cellula funzioni molto differenti, ma sono morfologicamente molto simili e hanno in comune una proprietà molto importante: essi possono duplicarsi così che da un granulo ne deriva un altro assolutamente identico al primo. I centrosomi (o centrioli), come vedremo in seguito, svolgono un ruolo molto importante nella divisione cellulare; i cinetosomi (o corpi basali) si trovano alla base delle ciglia o flagelli responsabili della motilità propria di certe cellule (v. fig. 20).
Il microscopio elettronico ha dimostrato una notevole somiglianza strutturale, se non identità, tra i centrosomi e i cinetosomi. Entrambi hanno la forma di un cilindro, rinforzato da un gruppo di nove fibrille; ognuna di queste fibrille è costituita da una ‛tripletta' di subfibrille (v. fig. 21).
Quando i centrosomi si raddoppiano durante la mitosi, il centrosoma figlio sta sempre ad angolo retto rispetto a quello preesistente.
Molto poco si sa circa la composizione chimica dei centrosomi e dei cinetosomi: sono probabilmente costituiti di subunità proteiche attaccate linearmente l'una all'altra. La presenza del DNA (che ci si aspetterebbe data la capacità di questi organuli cellulari di duplicarsi) è possibile ma non è stata accertata. Infatti, è più probabile che le cellule contengano molte molecole di precursori dei centrosomi (precentrosomi) sparse nel loro citoplasma e che queste molecole siano mantenute insieme da un processo simile alla cristallizzazione.
i) Ciglia e flagelli
Le ciglia e i flagelli determinano la motilità di molte cellule: si differenziano essenzialmente per le dimensioni, essendo le ciglia più corte dei flagelli. L'ultrastruttura dei due organuli è identica, anche se più complessa, a quella del cinetosoma a cui sono attaccati e da cui si originano (v. fig. 22): essi dispongono di un gruppo di ‛9+2' fibrille. Le 9 fibrille esterne sono costituite da ‛doppiette' di subfibrille, che di solito presentano delle protuberanze laterali (arms). Abbiamo una buona quantità di informazioni circa la loro composizione chimica, poiché è possibile separare i flagelli dagli spermatozoi e le ciglia dai Protozoi: il costituente principale è una (proteina fibrosa (tubulina), che si contrae in presenza di ATP. Quest'ultimo fornisce l'energia per la contrazione (che è causa della ‛vibrazione' delle ciglia e dei flagelli): durante la contrazione, i legami fosforici ricchi di energia vengono spezzati da un enzima (ATPasi), che è associato, negli arms dei filamenti esterni delle ciglia, alla proteina contrattile. È interessante notare che proteine simili, dotate anch'esse di attività ATP-asica, sono state rinvenute sempre dove ci sono cellule capaci di contrarsi: il caso estremo è, naturalmente, il muscolo, il quale contiene una proteina (actomiosina) che si contrae quando si aggiunge ATP. In relazione a questa somiglianza, le ciglia sono state chiamate ‛muscoli monomolecolari'.
l) Microtubuli
Sono questi delle proteine della stessa famiglia cui appartengono le proteine contrattili delle ciglia, dei flagelli, dei muscoli ecc.; probabilmente esistono in tutte le cellule: sono presenti nella membrana cellulare e il citoplasma spesso li contiene (v. fig. 23), orientati più o meno parallelamente. Questi microtubuli, come vedremo, svolgono una funzione importante nella divisione cellulare. Anche essi sono costituiti da una proteina (tubulina), che si contrae in presenza di ATP. Si ritiene che abbiano una certa importanza nei cambiamenti reversibili di forma che alcune cellule possono subire durante la differenziazione embrionale.
m) Materiali di riserva
Quando le cellule animali sono ben nutrite, esse accumulano riserve di grassi e glicogeno, che saranno utilizzati durante i periodi di digiuno. I lipidi non sono goccioline libere di grasso, ma sono circondate da una membrana: l'insieme forma i cosiddetti ‛lipocondri'. Il glicogeno si trova sotto forma di granuli, che qualche volta danno origine a ‛rosette' visibili al microscopio elettronico (v. fig. 24); queste particelle, come abbiamo detto, sono spesso associate alle membrane lisce del reticolo endoplasmatico.
Di regola le cellule non immagazzinano riserve di proteine; questo è invece il caso dei semi e delle uova: l'embrione in sviluppo costruisce le proprie proteine a spese delle preesistenti proteine accumulate. Queste ultime, nel caso delle uova, costituiscono il tuorlo o vitello (la parte gialla dell'uovo di gallina), che è formato da placchette vitelline visibili al microscopio. Al microscopio elettronico, le placchette vitelline appaiono come costituite di due parti: un nucleo centrale che contiene la proteina (più precisamente una fosfoproteina) in una forma disidratata, cristallina, e uno strato esterno (v. fig. 25) di corpi multivescicolari.
n) Cloroplasti
I granuli che contengono la clorofilla, presenti in tutte le piante verdi, i ‛cloroplasti', possono facilmente essere visti al microscopio ottico. Sono costituiti da lamelle parallele, chiamate ‛tilacoidi', di natura proteica (v. fig. 26). Nei cloroplasti delle piante superiori, ma non in quelli delle Alghe, zone dense (‛grani') interrompono i tilacoidi; la clorofilla e molti enzimi richiesti per la fotosintesi sono accumulati in questi grani, che probabilmente esercitano una funzione simile a quella delle creste nei mitocondri (onde assicurare uno stretto contatto fra gli enzimi attivi nel trasporto di elettroni). L'amido sintetizzato per fotosintesi rimane accumulato nei cloroplasti stessi.
Cloroplasti isolati sono capaci di fotosintesi indipendente. Tali cloroplasti isolati si contraggono in presenza di ATP e di luce: proteine contrattili devono perciò esistere nei tilacoidi e nella membrana cloroplastica.
I cloroplasti hanno in comune con i mitocondri una importante proprietà: contengono DNA e sono capaci di moltiplicarsi. Il DNA cloroplastico è fibrillare, come il DNA cromatinico; come nei mitocondri, la quantità di DNA presente in un singolo cloroplasto è insufficiente ad assicurare una completa autonomia, sebbene sia più abbondante che in un mitocondrio. Si ritiene che il DNA cloroplastico diriga la sintesi delle proteine strutturali dei tilacoidi, ma non quella degli enzimi fotosintetici.
Il DNA cloroplastico è capace di duplicarsi; può anche essere trascritto, cioè copiato nella forma di molecole di RNA (mRNA, rRNA e tRNA). Come i ribosomi batterici e mitocondriali, i ribosomi cloroplastici sono più piccoli (70 S invece di 80 S) e hanno un rRNA più piccolo dei ribosomi citoplasmatici. Ancora, come nei Batteri e nei mitocondri, la sintesi delle proteine cloroplastiche non risponde alla stessa maniera dei ribosomi citoplasmatici a inibitori quali il cloramfenicolo o la puromicina. La stessa questione di prima si può sollevare per i cloroplasti come per i mitocondri: sono essi organismi (alghe unicellulari nel caso dei cloroplasti, Batteri in quello dei mitocondri) che una volta vivevano liberamente e sono poi entrati in simbiosi nel corso dell'evoluzione? Le attuali acquisizioni biochimiche favoriscono una risposta affermativa: forse la scoperta, su un altro pianeta, di forme primitive di vita darà, prima o poi, una risposta definitiva a questo difficile problema.
3. Il nucleo
Tranne i Batteri, che hanno un nucleo molto semplificato e che sono chiamati per questo motivo ‛Procarioti', i nuclei delle cellule più complesse (quelli degli ‛Eucarioti') contengono sempre i seguenti costituenti visibili (v. fig. 2): una membrana nucleare che li circonda o li avviluppa; cromatina densa e lassa e i nucleoli, immersi insieme nel succo nucleare. Sebbene i nuclei siano quasi sempre di forma sferica od ovoidale, possono occasionalmente essere molto grandi e irregolari, come nelle ghiandole secretrici del baco da seta. Anche gli spermatozoi e i globuli rossi dei Vertebrati (tranne i Mammiferi) hanno un nucleo insolito, costituito da una cromatina estremamente addensata, quasi cristallina, nel quale, non è possibile vedere né nucleoli né succo nucleare.
Considereremo prima il nucleo come un tutto unico; poi, separatamente, daremo maggiori particolari sulla membrana nucleare, la cromatina, i nucleoli e il succo nucleare.
a) Composizione chimica del nucleo
Svariati metodi consentono l'isolamento di nuclei con buone rese e con una soddisfacente conservazione della loro struttura.
L'analisi chimica mostra che il nucleo è essenzialmente costituito di acidi nucleici (DNA e RNA) e proteine: fra queste ultime, si dovrebbe fare una distinzione tra proteine basiche e acide. Per prima cosa, esamineremo brevemente le principali proprietà di questi costituenti chimici di tutti i nuclei eucariotici.
DNA. - Il DNA (v. acidi nucleici) è associato con certe proteine basiche (istoni), in forma di un complesso nucleoproteico (nucleoistone). Quei siti che nella doppia elica del DNA non sono occupati dagli istoni possono legare proteine acide.
Metodi citochimici (per esempio la già menzionata reazione di Feulgen) consentono la individuazione del DNA e mostrano che esso è un componente fondamentale della cromatina e dei cromosomi delle cellule in divisione. Questi metodi non sono generalmente abbastanza sensibili da individuare il DNA mitocondriale e cloroplastico. Comunque, se una cellula, come l'alga unicellulare Acetabularia, contiene un singolo nucleo e molte migliaia di cloroplasti e mitocondri, i metodi biochimici mostrano che la quantità di DNA citoplasmatico può essere molto più elevata di quella del DNA nucleare.
Il contenuto di DNA varia considerevolmente nei nuclei di specie differenti, ma rimane piuttosto costante in tutti gli organi della stessa specie. Anche specie non molto lontane, da un punto di vista zoologico, possono contenere quantità molto differenti di DNA nei loro nuclei (c'è almeno 4 volte più DNA nel nucleo di una cellula di salamandra, per esempio, che in quello di una cellula di rana). Ma tutti i nuclei, siano essi grandi o piccoli (il rapporto tra volume nucleare e volume citoplasmatico tende a rimanere costante, così che cellule grandi hanno grossi nuclei), hanno approssimativamente lo stesso contenuto di DNA, nell'ambito della stessa specie: non c'è, in questo senso, un'apprezzabile differenza tra nuclei di cellule cardiache, nervose, renali, ecc.
Questa regola della costanza del DNA per nucleo non può, comunque, essere accettata in maniera dogmatica: in alcuni casi sicuramente autentici, come in quello dei cromosomi ‛giganti' di alcuni insetti (v. sotto), solo il contenuto di DNA di una certa parte della cromatina aumenta, in risposta a un trattamento ormonale.
Naturalmente, il contenuto in DNA del nucleo cambia nelle cellule prossime alla divisione: come vedremo, il DNA è duplicato prima che avvenga la mitosi; ne consegue che il contenuto in DNA delle varie cellule è differente nei tessuti dove le mitosi sono frequenti. Le cellule che stanno sintetizzando DNA possono facilmente distinguersi dalle altre: diversamente dai nuclei ‛quiescenti', quelli che sintetizzano DNA e che si accingono alla mitosi incorporano timidina radioattiva (un precursore specifico del DNA). I nuclei delle cellule ‛quiescenti' non incorporano timidina, fatto questo che mostra come il DNA sia una molecola metabolicamente molto stabile.
Ricapitolando, nei tessuti in cui la divisione cellulare non avviene (per es. nel cervello), il contenuto in DNA dei singoli nuclei è essenzialmente lo stesso e le molecole di DNA rimangono notevolmente stabili.
Se il DNA è il materiale genetico, come è possibile che il contenuto in DNA vari tanto da una specie all'altra? Perché il nucleo di una rana contiene 10.000 volte più DNA di un batterio e perché un nucleo di salamandra contiene 4 volte più DNA di un nucleo di rana? Ci si aspetta naturalmente che un batterio contenga meno ‛informazioni' genetiche di una rana, ma non in tale misura. La spiegazione di questo paradosso è venuta recentemente, quando fu scoperto, con l'aiuto della tecnica di ‛ibridazione molecolare', che fornisce precise informazioni sul grado di omologia fra due preparati di DNA (Britten e Kohn; Mc Carthy), che il DNA, nella cromatina degli Eucarioti, è altamente ‛ripetitivo': sequenze abbastanza lunghe di molecole di DNA (lunghe più di 300 nucleotidi) sono ripetute, moltiplicate molte volte. Queste ‛extracopie' di sequenze di DNA possono dare ragione dell'80- 90% del DNA nei nuclei ricchi di DNA della salamandra. È probabile che questa frequente ripetizione di certi segmenti di DNA abbia avuto una funzione importante nel corso dell'evoluzione.
Uno dei metodi fisici usati per la caratterizzazione dei DNA è la misura della loro ‛densità', che dà informazioni circa la loro composizione chimica, cioè sul contenuto relativo in guanina e citosina (G e C) in confronto all'adenina e alla timina (A e T). L'uso di questo metodo dimostra che molte cellule contengono, oltre al ‛picco' principale del DNA cromosomico, picchi minori di DNA satelliti. Uno di questi, il DNA satellite del topo (che naturalmente non ha niente a che fare col DNA mitocondriale), è un esempio di DNA estremamente ripetitivo, essendo costituito da un milione di copie della stessa (o molto simile) sequenza di DNA, che è lunga soltanto 300 nucleotidi. È probabile che esso svolga un ruolo nella mitosi, come vedremo più avanti.
Un altro DNA satellite ripetitivo è presente nella maggior parte dei nuclei: si tratta del DNA degli ‛organizzatori nucleolari', i quali, come vedremo, dirigono la formazione dei nucleoli nel nucleo e la sintesi dei ribosomi nel citoplasma. Questo DNA è di solito un satellite ‛pesante', con un contenuto in G+C più alto di quello della massa del DNA cromosomico. Esiste in circa 130 copie nell'insetto Drosophila, 600 copie nei polli e qualcosa come 1.000.000 di copie negli ovociti del rospo Xenopus laevis. Poiché la sua composizione base è simile a quella dei ribosomi, è normalmente chiamato ‛DNA ribosomiale' (rDNA).
Istoni. - Queste proteine basiche, a basso peso molecolare, sono strettamente legate col DNA nella cromatina e nei cromosomi. La loro basicità è dovuta a un eccesso di amminoacidi basici (arginina, lisina). In tutti i tessuti sono state trovate le stesse 5 frazioni fondamentali di istoni; sono state registrate soltanto differenze quantitative nella proporzione di queste 5 frazioni. Tuttavia, i globuli rossi nucleati degli Uccelli contengono uno speciale tipo di istoni; gli spermatozoi contengono sempre o istoni molto ricchi in arginina o protammine (nei Pesci), che hanno un peso molecolare più basso e un contenuto in arginina ancora più alto degli istoni. Abbiamo già visto che la cromatina dei globuli rossi degli Uccelli, come pure quella degli spermatozoi, è insolitamente addensata; è anche metabolicamente inattiva, nel senso che queste cellule non sintetiz- zano né RNA, né proteine. È molto probabile che questa inattività sia dovuta al forte legame del loro DNA con le proteine ricche di arginina.
Alcuni tipi di istoni possono essere modificati chimicamente (fosforilazione, acetilazione) a opera di enzimi presenti nei nuclei: tale modificazione chimica potrebbe avere una funzione nel controllo dell'attività cromatinica.
RNA. - Tutti i tipi di RNA, come diremo in seguito, sono sintetizzati nei nuclei: infatti questi ultimi contengono molte specie di RNA (di grandezza molecolare molto diversa), alcune delle quali non vengono mai trovate nel citoplasma.
Una parte dell'RNA è sempre strettamente associata alla cromatina e presenta una composizione in basi simile a quella del DNA cromosomico, per cui è spesso chiamata RNA DNA-simile (o D-RNA). Il resto dell'RNA è presente nei nucleoli: è di alto peso molecolare e, come l'rRNA dei ribosomi e il DNA degli organizzatori nucleolari, è normalmente ricco in G+C.
In contrasto con la stabilità del DNA, gli RNA nucleari sono metabolicamente molto labili: sono rapidamente sintetizzati, ma sono anche rapidamente degradati dagli enzimi intranucleari. La sintesi degli RNA nucleari, in particolare quella degli RNA nucleolari, è specificamente inibita da due farmaci: l'actinomicina D e l'α-amanitina. La prima agisce legandosi al DNA (specialmente all'rDNA), che non può più essere raggiunto dagli enzimi che sintetizzano l'RNA (le RNA-polimerasi, che sono inattive in assenza del DNA e che sono perciò DNA-dipendenti). L'α-amanitina inibisce l'attività di una di queste RNA-polimerasi, agendo sull'enzima stesso e non sul DNA. Incidentalmente, possiamo dire che anche i mitocondri e i cloroplasti contengono una RNA-polimerasi DNA-dipendente, ma diversa dagli enzimi presenti nel nucleo: come l'RNA-polimerasi batterica, non è attaccata dall'α-amanitina, ma da un altro farmaco, la rifamicina, che ha scarso effetto sulle RNA-polimerasi del nucleo.
Mentre, come già sappiamo, il contenuto di DNA per nucleo è approssimativamente costante, si riscontrano grandi variazioni nel contenuto di RNA nei nuclei delle cellule dei vari organi dello stesso organismo: questo contenuto è massimo in quelle cellule che sintetizzano più attivamente proteine e hanno il maggior contenuto in ribosomi.
Proteine acide. - Sono anche chiamate proteine ‛residue', perché rimangono insolubili quando i nucleoistoni sono stati rimossi dai nuclei (per es., per mezzo di soluzioni saline concentrate). Chimicamente, la frazione proteica acida è estremamente complessa; oltre ad albumine, globuline e fosfoproteine, nei nuclei sono presenti molti enzimi: alcuni di questi catalizzano la produzione di energia (la glicolisi, e probabilmente anche la fosforilazione ossidativa); altri hanno un ruolo essenziale nella sintesi degli acidi nucleici (DNA-polimerasi e RNA-polimerasi) e dei loro precursori, i nucleotidi. Nei nuclei isolati sono anche presenti enzimi idrolitici (ribonucleasi, per esempio), che scindono gli acidi nucleici. Mentre gli istoni sono molto simili in tutte le specie e in tutti i tessuti, le proteine non istoniche (acide) mostrano specificità di specie e di tessuto.
Sintesi di macromolecole da parte di nuclei isolati. - Se si aggiungono precursori adatti, i nuclei isolati possono sintetizzare sia proteine sia RNA. In entrambi i casi, la sintesi richiede l'integrità della struttura nucleare: la digestione, anche parziale, del DNA con l'enzima desossiribonucleasi interrompe rapidamente la sintesi; l'aggiunta di DNA (anche di differente origine, o polimeri ottenuti da sintesi artificiale) ripristina in gran parte la sintesi proteica.
Il sito e i meccanismi biochimici della sintesi proteica nucleare rimangono sconosciuti: l'autoradiografia fa pensare che la sintesi proteica avvenga principalmente nel nucleolo, ma la probabile assenza di veri ribosomi e polisomi nei nuclei rimane enigmatica per i biochimici. La sintesi proteica nucleare è bloccata dai veleni che inibiscono la produzione di energia (cianuri, dinitrofenolo), ma è molto meno influenzata dalla ribonucleasi di quanto non lo sia la sintesi proteica operata dai polisomi. Comunque, la sintesi proteica nei nuclei è quantitativamente soltanto di minore importanza, e non v'è dubbio che la maggior parte delle proteine nucleari abbia origine dal citoplasma: le proteine sintetizzate nei ribosomi citoplasmatici migrano nel nucleo ove vengono accumulate (in particolare nel nucleolo).
La sintesi nucleare dell'RNA in vitro presenta minori problemi: è sensibile all'actinomicina e all'α-amanitina, e richiede anch'essa produzione di energia. Di particolare interesse è il fatto che l'aggiunta di istoni blocca la sintesi nucleare dell'RNA, mentre l'eliminazione di queste proteine basiche per mezzo di una moderata digestione triptica, la stimola. Gli effetti inibitori degli istoni possono essere contrastati dalle proteine acide della cromatina e da certe frazioni di RNA. Nuclei isolati da cellule in rapida divisione possono anche sintetizzare piccole quantità di DNA: essi probabilmente completano la sintesi di catene incomplete di DNA, senza iniziare la sintesi di nuove.
b) Membrana nucleare
Come mostra la fig. 27, la membrana nucleare è a doppio strato e interrotta da ‛pori'. Questi pori, come dimostrato da recenti lavori basati sull'impiego della microscopia elettronica ad alta risoluzione, non sono semplici fori nella membrana. Essi vengono ora chiamati ‛complessi porosi nucleari' e certamente meritano questo nome: sono parzialmente occlusi da un granulo centrale e rinforzati lateralmente da 8 granuli laterali (che danno loro una simmetria ottagonale). Questi granuli laterali formano l'anello che circonda e rinforza i pori. La fig. 28 è un diagramma schematico di un complesso poroso in cellule vegetali.
La membrana nucleare si origina dai reticolo endoplasmatico e rimane in diretto contatto con queat'ultimo. Essa può essere isolata e analizzata chimicamcnte: la sua composizione complessiva è simile a quella delle membrane del reticolo endoplasmatico, ma ci sono notevoli differenze che indicano, ancora una volta, che le membrane intracellulari sono in uno stato dinamico, sempre mutevole. Questa opinione è ulteriormente confermata dall'esistenza nel citoplasma (e qualche volta anche dentro il nucleo) di strutture chiamate ‛lamelle annulate' (v. fig. 29): esse sono molto simili, ma non identiche, alla membrana nucleare. Probabilmente sono copie della membrana nucleare che vengono versate nel citoplasma, dove subiscono nuove disposizioni strutturali. Sono frequenti nelle cellule che sintetizzano attivamente proteine.
La permeabilità della membrana nucleare presenta interessanti problemi: esperimenti fisiologici (misurazione del rigonfiamento del nucleo dopo l'iniezione di varie sostanze nel citoplasma) indicano che molecole (proteine incluse) con un peso molecolare fino a 70.000 si muovono facilmente dal citoplasma al nucleo. Ma questo non spiega come enormi molecole di RNA, con peso molecolare superiore a i milione, possano passare dal nucleo al citoplasma: come nel caso della captazione di macromolecole mediante pinocitosi nella membrana cellulare, si deve ritenere che debbano esistere meccanismi più delicati della semplice permeabilità. La chiave di questi meccanismi probabilmente risiede nel modo (ancora sconosciuto) in cui funzionano i complessi porosi.
c) Cromatina
È il più importante costituente della cellula, perché contiene la maggior parte del DNA, che è il materiale genetico.
Il microscopio elettronico (v. fig. 30) mostra che la cr0- matina può essere suddivisa in due parti: una disciolta e una addensata, equivalenti rispettivamente alla eucromatina e alla eterocromatina dei citogenetisti. La eucromatina è geneticamente e biochimicamente più attiva della eterocromatina: per esempio, la cromatina disciolta sintetizza più RNA di quella addensata; quest'ultima è più lenta a duplicare il suo DNA, prima della divisione cellulare, di quanto non faccia la prima.
È probabile che due diverse cose siano confuse sotto lo stesso nome di eterocromatina: in alcuni casi, la cromatina addensata si identifica con la cromatina lassa, ma ha una conformazione differente, più addensata, dovuta all'aumentato aggomitolamento dei filamenti di cromatina. La cromatina addensata, in altri casi, è costituita di un DNA chimicamente differente da quello del resto della cromatina. Questo è il caso, per esempio, degli organizzatori nucleolari (che sono particolarmente ricchi in guanina e citosina) e del ‛DNA satellite' del topo, ricco di adenina e timina, che ha un'importante funzione nella divisione cellulare. Si può quindi concludere che ci sono due diversi tipi di eterocromatine: la facoltativa e la costitutiva. In quest'ultima intervengono differenze non soltanto nella composizione chimica del DNA, ma anche nella quantità e nella natura delle proteine associate agli acidi nucleici.
d) Nucleoli
Generalmente i nuclei contengono due nucleoli, tuttavia esistono delle eccezioni a questa regola: per esempio, la maggior parte delle uova non hanno nucleoli durante la segmentazione; d'altra parte gli ovociti in formazione degli Anfibi contengono da 1.000 a 2.000 nucleoli nel loro grande nucleo (spesso chiamato vescicola germinale).
L'ultrastruttura dei nucleoli è molto costante: non sono mai circondati da una membrana e sono costituiti di due parti: un ‛nucleo centrale' (core) fibrillare, circondato da una ‛corteccia' granulare (v. fig. 31). Nelle uova in segmentazione i nucleoli sono sostituiti da ‛corpi nucleolari', che possiedono soltanto la componente fibrillare.
Sia la corteccia granulare sia il ‛centro' fibrillare contengono RNA e proteine. Il contenuto in RNA della corteccia è molto più elevato di quello del centro: i granuli che formano la corteccia sono paragonabili ai ribosomi, ma sono più grandi. La sintesi dell'RNA inizia, comunque, nel centro fibrillare: le molecole di RNA appena sintetizzate sono trasportate da questo centro ai granuli della corteccia.
È possibile analizzare nucleoli isolati da ovociti o da cellule epatiche: essi contengono molte proteine, di cui qualcuna simile alle proteine ribosomiali, in aggiunta all'RNA. Quest'ultimo rassomiglia all'RNA ribosomiale nella sua composizione in basi (alto contenuto in guanina e citosina); ma l'RNA nucleolare è molto più grande degli rRNA: la sua costante di sedimentazione è 45 S (peso molecolare di 4.200.000 dalton) invece di 28 S (peso molecolare di 1.600.000 dalton) e 18 S (peso molecolare di 700.000 dalton).
È ora accertato che l'RNA nucleolare 45 S è un ‛precursore' degli rRNA citoplasmatici 28 S e 18 S, e che la sua sintesi è diretta dall'‛organizzatore nucleolare': abbiamo già visto che questo segmento di eterocromatina è costituito da DNA, che ha una composizione chimica simile a quella dei ribosomi e che è chiamato, per questa ragione, rDNA. Il suo ruolo diretto nella sintesi dell'rRNA citoplasmatico è stato chiaramente dimostrato da Gurdon e Brown (v., 1965), che lavorarono su un ‛mutante' del rospo Xenopus in cui gli organizzatori nucleolari mancano per una delezione. Questi organizzatori nucleolari mutanti non formano nucleoli e sono completamente incapaci di sintetizzare rRNA e ribosomi. Essi sono, naturalmente, letali e muoiono non appena fatti nascere in condizioni di omozigosi. Oli eterozigoti, che hanno un singolo organizzatore nucleolare, formano soltanto un nucleolo; possono sintetizzare ribosomi e raggiungere la forma adulta.
Abbiamo buone conoscenze circa l'origine degli organizzatori nucleolari negli ovociti di Xenopus. Negli animali giovani, che hanno appena subito la metamorfosi, gli ovociti sono allo stadio pachitene della meiosi. I loro nuclei hanno una struttura molto peculiare (v. fig. 32): vicino alle coppie di cromosomi, si può vedere una capsula contenente DNA addensato. L'ibridazione molecolare, a livello citologico, mostra che queste capsule sono costituite di rDNA, cioè un DNA che facilmente si ibrida con gli rRNA, perché entrambi hanno molte sequenze di basi in comune. Le capsule sono i siti di estesa sintesi di DNA, ma non (in contrasto con i cromosomi in pachitene) di sintesi di RNA: sono, infatti, i siti di una enorme moltiplicazione degli organizzatori nucleolari, i quali costruiscono migliaia di copie di se stessi. Il risultato di questa estesa amplificazione dei geni che dirigono la sintesi dell'rRNA è la formazione di 1.000 o più nucleoli nel nucleo dell'ovocito: quando gli organizzatori nucleolari si sono moltiplicati, lasciano la capsula e formano i 1.500 nucleoli che si trovano in questi nuclei.
La struttura dell'organizzatore nucleolare, in ciascun nucleolo, è ora ben conosciuta (perlomeno negli ovociti degli Anfibi) grazie a un intenso lavoro biochimico e di microscopia elettronica. La fig. 33 rappresenta il ‛ritratto' dei geni che dirigono la sintesi degli rRNA. L'organizzatore nucleolare è una molecola gigante circolare di DNA, in cui i geni che codificano gli rRNA 18 S e 28 S sono associati in coppie e si ripetono molte volte (amplificazione del gene). Le serie ripetute di geni sono separate da zone ‛intercalari' che non sono trascritte nell'interno del RNA. Nella fig. 33, le catene di RNA in accrescimento (probabilmente associate a proteine) appaiono come filamenti che si distaccano dall'asse del DNA; questi filamenti sono assenti negli intercalari, dove non c'è sintesi di RNA.
e) Succo nucleare
Sorprendentemente scarse sono le conoscenze sulla natura chimica e sulla funzione biologica del succo nucleare, anche nel caso delle vescicole germinali degli ovociti di Anfibi, che contengono abbastanza succo nucleare per l'analisi biochimica con i moderni micrometodi. Le proteine sono certamente i costituenti principali e comprendono numerosi enzimi idrolitici. Alcune di queste proteine sono di origine citoplasmatica: dopo la sintesi su polisomi, esse penetrano nell'interno del nucleo. Le proteine del succo nucleare si mescolano con il citoplasma circostante quando la membrana nucleare si frammenta durante la maturazione dell'ovocito sotto la stimolazione ormonale; questo mescolamento delle proteine nucleari e citoplasmatiche è un prerequisito indispensabile per la divisione mitotica nell'uovo.
Anche il succo nucleare contiene RNA, presumibilmente una miscela complessa di mRNA e dei loro precursori giganti associati con proteine, di precursori degli rRNA (anch'essi probabilmente associati a proteine in modo da formare particelle nucleoproteiniche) e di vari tipi di tRNA. La presenza di DNA nel succo nucleare è stata suggerita da vari ricercatori, ma per adesso la questione rimane aperta.
f) Il nucleo batterico
I Procarioti posseggono un nucleo grandemente semplificato, costituito da una fibra circolare di DNA, che probabilmente non è combinata con proteine. Nei Batteri non ci sono né membrana nucleare, né nucleoli, e nemmeno succo nucleare. La fibra di DNA qualche volta aderisce alla membrana cellulare per mezzo di speciali membrane chiamate ‛mesosomi'. La membrana cellulare, nei Batteri, contiene gli enzimi respiratori necessari alla produzione di energia.
Da un punto di vista morfologico, i Batteri sono semplici (v. fig. 34): una membrana, che esplica anche la funzione dei mitocondri degli Eucarioti, circonda il citoplasma, che è pieno di ribosomi e contiene il materiale genetico in forma di una singola fibra di DNA: il batterio è una macchina per la sintesi, controllata dai geni, di proteine specifiche. Questa semplicità è la ragione per cui la biologia molecolare ebbe origine da studi fatti su Batteri e su Virus.
4. Divisione cellulare (mitosi)
Le cellule spesso si dividono quando hanno raggiunto la loro massima grandezza; ma la ragione per cui le cellule nervose, per esempio, non si dividono mai dopo la nascita, mentre continuamente avvengono mitosi nella pelle, nell'intestino o nella milza durante tutta la vita, rimane sconosciuta.
Lo scopo principale della divisione cellulare è la uguale distribuzione del materiale genetico (DNA) nelle due cellule figlie. Questo fatto avviene con un meccanismo piuttosto complicato, che è rappresentato nella fig. 35 e, in modo schematico, nella fig. 36. La mitosi, benché in realtà sia un processo continuo, è, secondo un modello classico, divisa in 5 fasi successive: interfase, profase, metafase, anafase e telofase. L'intero processo dura da pochi minuti a molte ore nei diversi tipi di cellule.
L'interfase (v. fig. 35A) corrisponde al nucleo ‛quiescente' che è stato diffusamente descritto nel precedente capitolo. Quando la cellula è pronta per la mitosi, il centrosoma, che è già appaiato con un centrosoma ‛figlio', si divide e attorno a esso si sviluppa una irradiazione fibrillare, l'aster.
Durante la profase (v. fig. 35B), filamenti allungati, i cromosomi, diventano visibili nella cromatina, in precedenza omogenea; contemporaneamente, i nucleoli scompaiono e la membrana nucleare comincia a frammentarsi. I centrosomi si distaccano, raggiungendo infine i poli opposti del nucleo; ognuno di essi è adesso circondato da un aster; i due aster sono uniti insieme da un sistema di fibre costituente il fuso mitotico. Alla fine della profase, spesso denominata pro-metafase, i cromosomi sono attaccati ad alcune fibre dell'aster per mezzo di una regione speciale, il centromero o cinetocoro. In materiali favorevoli, si può vedere che i cromosomi sono già divisi in due cromosomi-figli, i cosiddetti ‛cromatidi', tranne nella regione del centromero, che rimane indivisa.
Nella metafase (v. fig. 35C), i cromosomi, che per un processo di spiralizzazione sono diventati più corti e più spessi fin dalla pro-metafase, sono situati nel centro del fuso, dove formano la ‛piastra equatoriale'. Questo stadio è particolarmente adatto al conteggio del numero dei cromosomi di una determinata specie. Gli aster hanno accresciuto le loro dimensioni attorno ai centrosomi.
Nell'anafase (v. fig. 35D), i centromeri finalmente si dividono e i cromosomi-figli, che sono di solito disposti a V, si muovono verso i due poli dell'‛apparato mitotico': essi vengono spostati in senso opposto, verso i due centrosomi.
Finalmente, nella telofase (v. fig. 35E), i cromosomi si srotolano e diventano meno distinti: essi si avvicinano sempre più ai centrosomi, mentre gli aster gradualmente si dissolvono. Una membrana nucleare si forma, a spese del reticolo endoplasmatico, attorno a ciascuna delle due serie di cromosomi, e i nucleoli diventano di nuovo visibili. Nello stesso tempo si forma un ‛solco' che divide la cellula in due cellule figlie: queste ultime posseggono metà del citoplasma, ma lo stesso numero di cromosomi contenuto nella cellula madre.
Ci sarebbe da aggiungere che, al contrario di quanto precedentemente descritto, le cellule dei vegetali superiori non hanno aster e probabilmente nemmeno centrosomi. Tratteremo ora più dettagliatamente i cromosomi, l'apparato mitotico (aster e fusi) e le variazioni biochimiche caratteristiche della cellula in divisione.
a) Cromosomi
Cromosomi metafasici. - Sono i portatori dei geni, le unità dell'ereditarietà, contenuti in codice nelle molecole del DNA cromosomico. In una normale cellula diploide (2n), metà dei cromosomi sono di origine materna; e l'altra metà di origine paterna, sin dalla fecondazione dell'uovo: ogni gamete porta un complemento cromosomico aploide (n), caratteristico della specie. Nel diagramma semplificato della fig. 36, dove i cromosomi si dividono, il numero aploide di cromosomi sarebbe 2, ma di solito è molto più alto (nell'uomo è 23).
I cromosomi metafasici possono essere isolati da cellule artificialmente arrestate a questo stadio di mitosi da trattamenti che bloccano la formazione dell'apparato mitotico (colchicina, demecolcina) o interferiscono reversibilmente con la sintesi del DNA. Trattamenti ripetuti delle cellule con alte e basse temperature (shock termici) provocano pure la ‛sincronizzazione' delle cellule. Tale sincronizzazione indotta della divisione cellulare di una popolazione di cellule è, naturalmente, di grandissima importanza per l'analisi biochimica della mitosi.
I cromosomi metafasici isolati non differiscono molto chimicamente, almeno per quanto ne sappiamo, dalla cromatina delle cellule in interfase: sono costituiti da DNA associato a istoni, proteine acide e piccole quantità di RNA. Sono state trovate, nei cromosomi metafasici, alcune proteine che non sono presenti nella cromatina del nucleo in interfase; esse potrebbero forse determinare la forte condensazione dei cromosomi nella metafase.
Visti con il microscopio ottico, i cromosomi metafasici di solito appaiono come formati da due cromatidi separati da una sottile fenditura; i cromatidi, specialmente nelle cellule vegetali lasciate per un certo tempo in ambiente freddo, sono avvolti in ‛cromonemi'. Si può anche distinguere il punto dell'attacco al fuso, il centromero e, in casi favorevoli, la presenza, all'estremità dei due cromosomi della stessa coppia, degli organizzatori nucleolari (sfere o bastoncini fortemente condensati).
I cromosomi metafasici, naturalmente, contengono gli stessi tipi di DNA contenuti nella cromatina: una massa di DNA cromosomico con le sue sequenze ripetitive e uniche, DNA-satelliti (di solito più leggeri, per la minore proporzione di sequenze di guanina+citosina) e un pesante rDNA degli organizzatori nucleolari. Un recente lavoro ha mostrato, con un metodo molto elegante e fine di ibridazione molecolare su sezioni di tessuto, che il DNA satellite del topo è accumulato nella regione del centromero. Ma non sappiamo ancora perché sequenze più volte ripetute di DNA satellite del topo sono accumulate nella regione dove le fibre del fuso aderiscono al cromosoma. È chiaro che la replicazione del centromero avviene più tardi di quella dei cromatidi: abbiamo visto che i cromosomi non sono spostati in senso opposto, proprio alla fine della metafase, finché il centromero non si è diviso. Infatti, la principale differenza tra la meiosi e la mitosi è che, nella prima, i centromeri non si dividono. È molto probabile che l'accumulo di DNA-satellite nella regione del centromero svolga un ruolo essenziale nella separazione dei cromosomi, ma nulla sappiamo circa i meccanismi molecolari di questa separazione.
L'analisi ultrastrutturale dei cromosomi metafasici è resa difficile dalla loro lunghezza: molecole intere di DNA lunghe 100 μ (0,1 mm) e anche 1 mm sono state viste, rispettivamente, nello spermatozoo del riccio di mare e nei cromosomi di un roditore (il criceto cinese).
Si può calcolare che il DNA di una singola cellula umana aploide, se fosse dato di poterlo estendere interamente, formerebbe un filo lungo un metro. Questo significa che le catene di DNA-istone, nei cromosomi metafasici, debbono essere fortemente ammassate come i fili di lana in un gomitolo. Il microscopio elettronico mostra che i cromosomi sono costituiti di filamenti molto lunghi e sottili, spessi circa 250 Å, che subiscono un esteso aggomitolamento. Di solito, soltanto gli anelli fatti da tali filamenti, che si dipartono dalla massa del cromosoma, possono esser visti (v. fig. 37). La stessa struttura si può osservare nella cromatina dei nuclei scissi in interfase. L'esatta natura delle fibre di cromatina spesse 250 Å rimane tuttora in discussione: esse potrebbero essere costituite da una singola fibra aggomitolata di 100 Å, o da due di tali fibre strettamente aderenti.
Mentre i cromosomi metafasici sono lunghi soltanto pochi micron, cromosomi molto più grandi esistono in natura: sono i ‛cromosomi giganti' di alcuni Insetti (Ditteri) e i ‛cromosomi a spazzola' degli ovociti degli Anfibi. Il loro studio ha ampliato sensibilmente le nostre conoscenze sulla struttura e la funzione dei cromosomi.
I cromosomi giganti e il puffing. - Come mostra la fig. 38, i cromosomi giganti (i quali sono 10 volte più lunghi e 100 volte più spessi dei normali cromosomi metafasici) sono costituiti da una successione di bande scure (ricche di DNA) e di interbande chiare (povere di DNA). Sono stati trovati nei nuclei in interfase delle ghiandole salivari presenti nelle larve di Ditteri (Drosophila, Chironomus, Rhynchosciara, tre specie che sono state particolarmente studiate). La loro presenza in questi nuclei dimostra chiaramente la persistenza dei cromosomi nel nucleo in interfase. Lo spessore dei cromosomi giganti è dovuto al loro modo di formarsi: essi risultano da una ‛endomitosi', cioè da un ripetuto moltiplicarsi delle fibre cromosomiche, senza divisione cellulare o nucleare. Di conseguenza, i cromosomi giganti sono composti da molte fibre: per questa ragione vengono anche chiamati cromosomi ‛politenici'. Che le bande scure in realtà corrispondano ai geni, fu scoperto grazie a un accurato confronto tra la mappa genetica e quella citologica della Drosophila.
A livello molecolare, è probabile che ciascuna fibra di cromosoma gigante sia costituita di singole molecole di DNA, che si piegano a formare le bande scure, e che sono circondate da istoni e proteine acide. Si dispiegano poi nelle interbande, che contengono poco DNA rispetto al grande contenuto proteico.
Quando lo sviluppo della larva si avvicina allo stadio della metamorfosi, viene prodotto un ormone chiamato ‛ecdisone'; i cromosomi giganti reagiscono all'ecdisone con rigonfiamenti locali detti puffs. Le strutture fibrillari dei cromosomi diventano meno distinte nei puffs e ciò è dovuto al dispiegarsi o allo srotolarsi delle fibre del cromosoma. I puffs appaiono in determinati momenti e in determinate bande soltanto nei cromosomi giganti: si possono considerare come una manifestazione microscopicamente visibile dell'attività genetica. Mentre i puffs possono essere indotti mediante trattamento delle ghiandole con ecdisone, altri agenti (riscaldamento, ribonucleasi, aggiunta di Zn2+, ecc.) possono anche produrre puffs in modo molto meno specifico. È stato suggerito che il puffing è la risposta ai mutamenti nel rapporto Na+/K+ nel nucleo, che sarebbero indotti da molti e svariati agenti fisici o chimici.
Il puffing non può avvenire in assenza di sintesi di RNA, poiché viene abolito dall'aggiunta di actinomicina o di istoni. Infatti, le sintesi del DNA, dell'RNA e delle proteine sono tutte indotte dall'ecdisone alla banda specifica dove avviene il puffing. Ma sembra attualmente che la prima reazione dei cromosomi giganti all'ormone sia un accumulo locale - nel sito dove in seguito si formerà il puff - di una proteina acida presente nel succo nucleare; questa proteina ‛de-reprimerebbe' il DNA presente in questa particolare banda, cioè permetterebbe la sua trascrizione dall'RNA-polimerasi e la sintesi dell'mRNA che causa la formazione del puff. L'ultramicroanalisi della composizione in basi ha inoltre dimostrato che gli RNA presenti nei diversi cromosomi, nei puffs e nei nucleoli sono tutti differenti.
Di particolare interesse è il fatto che, nelle Rhynchosciara, certe bande possono reagire all'ecdisone con sintesi individuali di DNA: la risposta all'ormone in tali casi è una localizzata amplificazione genica, che interessa uno o pochi geni, e non avviene in altri.
Cromosomi a spazzola. - Anche le vescicole germinali (nuclei) di ovociti in accrescimento (uova ovariche) dei Vertebrati (tranne quelle dei Mammiferi) contengono cromosomi molto grandi: questi hanno un curioso aspetto di spazzole e per tale motivo sono stati chiamati ‛cromosomi a spazzola'.
Abbiamo già discusso il ruolo svolto dalle ‛capsule' contenenti rDNA, nella formazione di centinaia di nucleoli a uno stadio molto iniziale (pachitene) di questi ovociti. I cromosomi a spazzola diventano visibili in uno stadio più avanzato, quando l'ovocito sta attivamente accrescendosi e può già essere visto a occhio nudo. Se tale ovocito (che ha un diametro di circa 0,5 mm) è aperto delicatamente con una pinza, la sua grande vescicola germinale salta fuori e la si può osservare al microscopio a contrasto di fase; si possono facilmente vedere, oltre ai nucleoli, anche i cromosomi a spazzola (v. fig. 39): essi sono costituiti da un asse, che presenta ispessimenti locali (i ‛cromomeri'), da cui si dipartono ‛barbe' laterali. I cromomeri, in cui la presenza di DNA può essere dimostrata con la reazione di Feulgen, sono gli equivalenti delle bande dei cromosomi giganti. Come queste bande, essi probabilmente rappresentano dei geni individuali. Lo sviluppo delle barbe varia dappertutto nel cromosoma: si possono considerare, come i puffs, un'espressione visibile dell'attività di singoli geni.
Esperimenti basati sulla digestione enzimatica hanno mostrato che, come nei cromosomi giganti, l'intera lunghezza del cromosoma a spazzola (incluso l'asse delle barbe) è costituita da un filo continuo di DNA: questo filo è ripetutamente arrotolato nel cromosoma e srotolato altrove (cioè tra i cromomeri e nelle barbe).
La fig. 40 rappresenta in modo schematico la probabile struttura di un cromosoma a spazzola; essa mostra anche come l'RNA venga continuamente sintetizzato nell'asse di DNA delle barbe e come questa sintesi cominci a una estremità della barba e termini alla estremità opposta. Sembra quasi che le molecole di RNA-polimerasi DNA-dipendenti si siano spostate lungo l'asse della barba in maniera unidirezionale.
Quest'abbondante sintesi di RNA da parte dei cromosomi a spazzola è ovviamente correlata con la copiosa sintesi proteica che avviene durante l'accrescimento dell'ovocito. Tuttavia, la massa di proteine accumulate nell'ovocito che si sta sviluppando è costituita dalle placchette vitelline, ed è ora ben accertato che le fosfoproteine che costituiscono il vitello non sono fabbricate dall'ovocito stesso: il loro precursore, la vitellogenina, è sintetizzata, sotto la stimolazione ormonale, dal fegato delle femmine e riversata nel sangue; essa allora penetra nell'interno dell'ovocito in accrescimento per intensa pinocitosi e viene trasformata nella fosfoproteina cristallina delle placchette vitelline.
Due altre osservazioni dovrebbero essere fatte riguardo ai cromosomi a spazzola: in primo luogo sta diventando sempre più chiaro che molti cromosomi (se non tutti) hanno barbe laterali come quelli a spazzola; ma queste barbe sono naturalmente molto più piccole e meno visibili. In secondo luogo, i cromosomi a spazzola sono cromosomi meiotici e non mitotici: come si vede nella fig. 39, essi sono tenuti assieme da nodi, chiamati ‛chiasmi'. Il microscopio elettronico ha mostrato che i chiasmi sono strutture complesse, in cui i cromosomi omologhi sono tenuti assieme dai cosiddetti ‛complessi sinattinemali' (v. fig. 41) di natura proteica. (Sui cromosomi, v. anche genetica: Citogenetica, ed embriologia).
b) Apparato mitotico
Metodi citochimici hanno dimostrato la presenza di proteine e RNA come costituenti maggiori delle fibre dell'aster e del fuso. Tra queste ultime alcune sono continue, da un centrosoma all'altro; le altre collegano uno dei centrosomi al centromero di un cromosoma. Il microscopio elettronico mostra che le fibre dell'aster e del fuso sono in realtà dei microtubuli, e sono perciò cavi. Molti ribosomi possono vedersi nel fascio dei microtubuli.
Un enorme progresso fu realizzato quando D. Mazia riuscì a isolare l'apparato mitotico da uova fecondate di riccio di mare (v. fig. 42). L'analisi chimica divenne allora possibile e dimostrò che l'apparato mitotico è costituito principalmente da una proteina, la tubulina. Questa proteina preesiste, in forma di piccole subunità, nel citoplasma dell'uovo non fecondato: la formazione dei lunghi microtubuli è il risultato di una polimerizzazione lineare (che implica ossidazione di gruppi −SH− e formazione di legami −SS−) al momento della mitosi. Alcuni agenti che inibiscono la mitosi (colchicina, vinblastina) sono attivi perché si legano alla tubulina: essi impediscono la sua polimerizzazione e, come conseguenza, la formazione di un fuso organizzato. Sostanze che, come il mercaptoetanolo, riducono i ponti −SS− inibiscono la costruzione dell'apparato mitotico: gli asters e i fusi non si formano, ma sia i centrosomi sia i cromosomi possono replicarsi.
Più approfondite analisi hanno dimostrato che l'apparato mitotico contiene, in effetti, tre distinte proteine. Una di loro appartiene alla classe delle proteine contrattili: si contrae in presenza di ATP e possiede attività ATP-asica.
c) Anormalità mitotiche
La cellula in divisione è più vulnerabile da parte degli agenti esterni (fisici e chimici) della cellula in interfase. Di conseguenza, le anormalità mitotiche sono frequenti e di vario tipo, specialmente nelle uova che si sviluppano in condizioni avverse. Le più frequenti di queste anormalità possono essere qui ricordate: sono le apparizioni dei ‛citoaster' (centinaia di asters nello stesso citoplasma), mitosi ‛pluricentriche' (i cromosomi sono distribuiti fra tre o più apparati mitotici), ‛amitosi' (il nucleo si divide per germinazione in assenza di apparato mitotico), ‛c-mitosi' (assenza di un fuso organizzato dopo trattamento con colchicina, calore, ecc.; se i cromosomi possono replicarsi, la cellula risultante sarà poliploide, con 4n, 8n, ecc. cromosomi), rottura e viscosità dei cromosomi (quando il DNA è parzialmente degradato, per esempio dopo trattamento con raggi X o con onde ultraviolette), picnosi (completa degenerazione della cromatina, che forma una sfera amorfa), ecc.
La maggior parte di queste aberrazioni mitotiche, che sono particolarmente frequenti nelle cellule cancerose, portano a una anormale distribuzione dei cromosomi nelle cellule figlie: si realizzano così condizioni di ‛aneuploidia', le quali sono per lo più letali, perché l'organismo muore rapidamente quando i nuclei delle sue cellule contengono un numero molto variabile di cromosomi.
d) Variazioni biochimiche durante la mitosi.
Produzione di energia. - La presenza di ossigeno e glucosio, come fonte di energia, è normalmente necessaria per la divisione della cellula; le uova di rana, tuttavia, possono dividersi molte volte in assenza totale di ossigeno o in presenza di cianuri, che bloccano quasi completamente la respirazione. Ma le uova di rana, come tutte le altre cellule, non si dividono in presenza di dinitrofenolo, che blocca la sintesi dell'ATP: l'energia prodotta dai mitocondri sotto forma di ATP è perciò assolutamente indispensabile per la divisione cellulare.
Nelle uova, sono state dimostrate piccole fiuttuazioni della respirazione durante il ciclo mitotico: c'è un massimo durante l'interfase, un minimo durante la metafase e l'anafase.
Sintesi del DNA. - Il DNA non è sintetizzato durante la mitosi, ma durante la precedente interfase. Questo è stato dimostrato con l'uso di tecniche citochimiche (misurazione del contenuto in DNA di ciascun nucleo con la reazione di Feulgen, studi autoradiografici sull'incorporazione della timidina nel DNA) e con analisi biochimiche di cellule sincronizzate. Da tutto questo lavoro è emerso il concetto di ‛ciclo cellulare' (v. fig. 43): l'interfase, per una cellula che sta per subire la mitosi, si può dividere in tre periodi: G1, S e G2. S è lo stadio dell'effettiva sintesi del DNA e dura poche ore: il contenuto di DNA del nucleo si raddoppia esattamente, passando da un valore di 2n a 4n. Questa fase è preceduta dalla fase preparatoria Gi ed è seguita dal periodo G2 (entrambi durano poche ore, nel caso di cellule che si dividono attivamente). L'esistenza della fase G2 prova che, se la sintesi del DNA è necessaria per la divisione della cellula, non è tuttavia sufficiente a indurre la mitosi: altri eventi biochimici, di natura ancora sconosciuta, sono necessari prima che la cellula che ha replicato il suo DNA possa entrare nella profase. Talvolta si dice che le cellule che non si dividono mai rimangono in G0 (G è l'abbreviazione di gap, intervallo).
La sintesi del DNA è probabilmente il più fondamentale di tutti i processi biologici: il DNA preesistente deve essere copiato fedelmente prima di essere distribuito nelle cellule figlie. Qualsiasi errore fatto in quel momento provocherebbe alterazioni nel materiale genetico e il risultato sarebbe una mutazione, generalmente dannosa per la discendenza se avviene durante la formazione del gamete.
Noi sappiamo molto di più sul meccanismo di duplicazione del DNA nei Batteri e nei batteriofagi che non nelle cellule eucariotiche; ma non c'è ragione, fino a ora, per credere che i meccanismi di duplicazione del DNA siano sostanzialmente differenti nei Procarioti e negli Eucarioti. In entrambi i casi la duplicazione del DNA è di tipo ‛semiconservativo', come è rappresentata in maniera schematica nella fig. 44; le due fibre delle molecole di DNA composte da una doppia elica si separano, formando una ‛forchetta', visibile nei Batteri col microscopio elettronico. Ognuna delle due fibre è replicata a opera dell'enzima DNA-polimerasi (il quale, come l'RNA-polimerasi, è DNA-dipendente e perciò richiede la presenza di DNA ‛stampo'), che agisce sui precursori, i desossiribonucleosidi trifosfati. Tale meccanismo implica che, in una cellula pronta per la mitosi e già nel periodo G2 del ciclo cellulare, metà del DNA presente è ‛vecchio' e l'altra metà è appena sintetizzato durante il periodo S. Come si può facilmente vedere dalla fig. 44, il modello a doppia elica di Watson e Crick è in grado di dare una perfetta spiegazione di una corretta duplicazione del DNA. Esperimenti condotti prima su Batteri e poi su cellule eucariotiche non lasciano dubbio che la duplicazione semi-conservativa del DNA sia un processo universale nelle cellule viventi.
Tuttavia, le cose stanno diventando, come normalmente accade, sempre più complicate, e ancora oggi il meccanismo della duplicazione del DNA, perfino nei Batteri, è oggetto di accalorate discussioni: è molto dubbio, per esempio, che la classica DNA-polimerasi sia il ‛vero' enzima, perché mutanti batterici che mancano di questo enzima possono ciononostante duplicare il loro DNA. Un'altra DNA-polimerasi, più specifica e probabilmente legata alla membrana batterica, è forse il vero enzima. È anche abbastanza evidente che, nei Batteri, solamente brevi tratti di molecola di DNA sono copiati da un enzima che ‛cammina' lungo una delle eliche del DNA e ‛torna indietro' lungo l'altra.
Una situazione simile probabilmente esiste nelle cellule degli Eucarioti: studi effettuati mediante autoradiografia sulla sintesi del DNA nei cromosomi giganti indicano che la sintesi avviene, nel cromosoma, in ‛punti di inizio' multipli (detti anche ‛repliconi'): la sintesi del DNA può perfino procedere in due opposte direzioni, partendo da due differenti repliconi sullo stesso cromosoma. Sebbene le molecole di DNA nei cromosomi giganti siano continue, la loro sintesi è discontinua: essa non procede da un'estremità all'altra del cromosoma; il DNA appena sintetizzato forma soltanto dei corti tronconi, non più lunghi di 1.000 nucleotidi, e contiene un breve segmento di RNA. Questo DNA neosintetizzato, transitorio, è meno stabile del ‛vecchio' DNA. La velocità di duplicazione del DNA è stata calcolata in certe cellule ed è risultata dell'ordine da 0,5 a 1,5 μ al minuto. I metodi autoradiografici ci consentono ora di seguire al microscopio la duplicazione del DNA nella cromatina durante la fase S: la velocità di sintesi del DNA dipende sia dal numero e dall'accostamento dei punti di inizio sia dalla velocità di movimento delle molecole di DNA-polimerasi lungo le fibre del DNA. Durante lo sviluppo embrionale, il numero dei punti di inzio è dapprima molto alto, così che questi punti sono strettamente accostati l'uno all'altro e la duplicazione del DNA è molto rapida (la duplicazione del DNA avviene in meno di 10 minuti durante la segmentazione delle uova di Drosophila); più tardi, il numero dei punti di inizio progressivamente diminuisce e la sintesi del DNA diviene notevolmente lenta. Questo numero è ancora più basso durante la meiosi, nel corso della quale la duplicazione del DNA ha luogo abitualmente in diversi giorni (H. G. Callan).
È stato ripetutamente sostenuto che la sintesi del DNA inizia sempre a contatto della membrana nucleare; tuttavia, le più recenti osservazioni suggeriscono piuttosto che la duplicazione del DNA inizia in molti punti differenti e in modo casuale nell'intero nucleo. L'eterocromatina, che è spesso adesa alla membrana nucleare, si replica di solito più tardi (nella seconda metà) nella fase S: ciò è vero sia per gli eterocromosomi sessuali X e Y, sia per le ripetute sequenze ricche di AT dell'eterocromatina centromerica.
In ragione della sua estrema delicatezza, la sintesi del DNA (la fase S del ciclo cellulare) è particolarmente sensibile agli agenti fisici e chimici: è bloccata, per esempio, dai raggi X e dall'aggiunta in grande eccesso di timidina (un normale precursore nella sintesi del DNA) o di analoghi della timidina (fluoro-desossiuridina, per esempio), di idrossiurea, ecc. Poiché queste sostanze, così come gli shock termici, esplicano effetti reversibili, è relativamente facile ottenere una popolazione di cellule bloccate in fase S e permetterne il recupero: si otterrà una sincronia di divisioni cellulari, che durerà almeno per 2-3 cicli mitotici.
Il caso delle uova merita una breve menzione: subito dopo la fecondazione, esse subiscono ripetute e rapide divisioni. L'analisi del loro ciclo cellulare durante questo periodo iniziale di sviluppo, che dura 10-20 ore, mostra che esse non hanno né il periodo G1 né il G2: come i Batteri, esse sintetizzano ininterrottamente DNA (tranne, naturalmente, durante l'effettiva fase di mitosi). Questo è possibile perché l'uovo non fecondato già contiene una buona scorta di precursori (desossiribonucleotidi trifosfati) per la sintesi del DNA. Il loro citoplasma contiene anche grandi quantità di enzimi che sintetizzano DNA, del tipo DNApolimerasi. Durante la divisione, questa DNA-polimerasi citoplasmatica migra progressivamente nei nuclei. Infine, molte uova di dimensioni notevoli (quelle degli Anfibi, in particolare) contengono, oltre a DNA mitocondriale, una riserva di DNA citoplasmatico localizzato nelle placchette vitelline: i nuclei in divisione possono probabilmente attingere da questa riserva di DNA del tuorlo in maniera da replicare il proprio DNA il più rapidamente possibile.
Infine, potrebbe aggiungersi che vi è qualche indizio, per il momento ancora legato a particolari circostanze, che le suddette variazioni cicliche della respirazione durante la divisione dell'uovo e la sintesi del DNA possano in qualche modo essere in correlazione con questi fenomeni: la respirazione è al punto massimo durante la fase S, un fatto questo per nulla sorprendente, dal momento che la sintesi sia del DNA che dei suoi precursori richiede energia.
RNA e sintesi proteica. - Mentre la sintesi del DNA è limitata al periodo S, la sintesi dell'RNA è continua durante l'intera interfase. Ma diminuisce durante la profase e scende bruscamente quasi a zero durante la metafase e l'anafase. A questo proposito, la cellula in mitosi è paragonabile, come vedremo in seguito, a una cellula che non ha affatto un nucleo. È ovviamente un errore chiamare il nucleo in interfase un nucleo ‛in riposo': esso è, al contrario, molto attivo, impegnato nella sintesi sia del DNA che dell'RNA; in confronto, la cellula in divisione, con i suoi cromosomi addensati e inattivi, potrebbe definirsi una cellula ‛pigra' e in riposo.
Nel periodo G2, alla fine dell'interfase, il nucleo contiene notevoli quantità di RNA, che sono diffuse nel citoplasma durante la profase; una quantità esigua (se pure ve ne è) di questo RNA viene recuperata dai nuclei quando essi si riformano durante la telofase.
Nelle uova di riccio di mare e di Anfibi, la divisione di cellule, fino a molte centinaia, può avvenire in presenza di actinomicina, che blocca completamente la sintesi dell'RNA: ciò dimostra che queste uova già contengono, prima dell'inizio della loro divisione, tutte le informazioni necessarie per la formazione ripetuta dell'apparato mitotico. Questa informazione è contenuta in codice in molecole di mRNA stabili e preesistenti. D'altra parte, la divisione è quasi immediatamente arrestata dalla presenza della puromicina, fatto questo che dimostra che la sintesi proteica è assolutamente necessaria per la divisione della cellula.
La sintesi proteica globale, come la sintesi dell'RNA, aumenta durante l'interfase e la profase: ma, a differenza della sintesi dell'RNA, non si blocca mai durante la metafase e l'anafase; essa risulta solamente ridotta e può continuare perché molecole stabili di mRNA sono ancora presenti nel citoplasma.
Cambiamenti in un certo numero di proteine specifiche durante la mitosi sono stati già studiati: per esempio, gli istoni, quantunque siano così strettamente associati al DNA nei cromosomi, non si replicano in maniera semiconservativa. È probabile che queste proteine basiche siano sintetizzate nel citoplasma su piccoli polisomi e migrino poi da questi nell'interno del nucleo; la sintesi degli istoni è spesso in aumento durante la fase S, ma è ancora rilevabile durante i periodi G1 e G2.
Le proteine dei microtubuli sono di continuo sintetizzate durante l'intera interfase e servono durante la seguente fase di mitosi. Nelle uova, come abbiamo visto, esse sono già presenti prima della fecondazione; il citoplasma dell'uovo non fecondato contiene anche una riserva di molecole di mRNA tubulina, che sono tradotte dopo la fecondazione e durante la segmentazione. La sintesi della tubulina, e la formazione degli asters, è così possibile quando frammenti anucleati di uova non fecondate di riccio di mare subiscono la partenogenesi (v. sotto, cap. 5).
Molti enzimi sono sintetizzati in maniera ciclica durante la divisione di cellule sincronizzate. Alcuni di essi, come la timidinachinasi, intervengono nella sintesi dei precursori del DNA e si accumulano, come ci si poteva aspettare, poco prima del periodo S. Ma anche altri, che manifestamente non hanno niente a che fare con la mitosi (glucosio- e lattato-deidrogenasi, per esempio) mostrano un comportamento ciclico durante il ciclo cellulare. Uno degli enzimi che entra nella sintesi del DNA, la ribonucleotidereduttasi (che catalizza la riduzione dei ribonucleotidi nei corrispondenti desossiribonucleotidi), è interessante perché la sua attività aumenta considerevolmente (fino a 100 volte) allorché viene stimolata la divisione di cellule in riposo aggiungendo loro un mezzo di coltura fresco o quando cellule di fegato sono indotte a entrare in mitosi mediante asportazione chirurgica di una parte dell'organo: ma, sebbene questo enzima possa avere diritto a un ruolo decisivo nella regolazione della sintesi del DNA, esso non mostra sempre un particolare incremento di attività durante la fase S.
e) Inibizione e stimolazione della mitosi
Molti veleni, alcuni dei quali sono usati regolarmente nella chemioterapia dei tumori maligni, bloccano efficacemente la mitosi. Essi sono o veleni del fuso, o veleni dei cromosomi.
Tra i veleni del fuso, la colchicina, la demecolcina e la vinblastina agiscono, come abbiamo visto, legandosi alla proteina principale dell'apparato mitotico. Anche gli inibitori dei gruppi −SH e gli agenti riducenti interferiscono con la formazione dell'apparato mitotico.
I principali veleni dei cromosomi sono le radiazioni (raggi X e ultravioletti) e i veleni radiomimetici (‛mostarde azotate') che degradano direttamente o modificano le molecole di DNA. Altre sostanze inibiscono la duplicazione del DNA durante la fase S: mitomicina, idrossiurea, desossiadenosina, fluorodesossiuridina, ametopterina, ecc. Anche sostanze semplici come la caffeina e l'acido nitroso minano l'integrità del DNA. Ad alte dosi, questi agenti producono la degenerazione picnotica dei cromosomi; a basse concentrazioni, sono mutageni.
Più interessanti sono i ‛caloni', ancora pochissimo conosciuti: sembra che queste sostanze producano un'inibizione organo-specifica della mitosi. Per esempio, un estratto di rene adulto inibirebbe specificamente la mitosi nel rene, ma non in altri organi dell'embrione; la crescita in vitro di cellule umane maligne provenienti da un tumore umano della mucosa uterina (le cosiddette ‛cellule HeLa') sarebbe arrestata da un estratto di mucosa uterina. I caloni sono attualmente oggetto di una gran mole di lavoro, e non c'è dubbio che nelle cellule differenziate esistano sostanze che inibiscono specificamente le mitosi nelle cellule dello stesso tessuto. Queste sostanze sono state isolate e purificate in molti casi: esse sono o piccoli peptidi (peso molecolare 2.000-3.000 dalton) o grandi molecole glicoproteiche.
Non si hanno sufficienti conoscenze circa la stimolazione della mitosi; nelle cellule vegetali, l'attività mitotica è stimolata dalle ‛citochinine', che sono derivati poco comuni delle basi degli acidi nucleici: molte di loro sono state isolate da preparazioni di RNA solubile. Si suppone che esse agiscano accorciando il periodo G2 dell'interfase.
Nelle cellule animali, i linfociti (globuli bianchi del sangue) non si dividono quando circolano nel sangue; tuttavia, quando sono trattati con sostanze di origine vegetale chiamate ‛fitoemoagglutinine', essi realizzano prontamente la sintesi di DNA e la mitosi. Si sa anche che la prima risposta delle cellule che producono anticorpi a un antigene è la divisione cellulare. Anche certi ormoni (per es., la eritropoietina, che stimola la formazione di globuli rossi del sangue) provocano la sintesi di DNA e la divisione cellulare. In tutti questi casi, la sintesi del DNA è preceduta da quella dell'RNA e delle proteine; ma il primo effetto dei ‛mitogeni' si esplica certamente sulla membrana cellulare, poiché essi non penetrano nella cellula. Gli esperimenti di induzione della duplicazione del DNA e della mitosi con l'aggiunta di lectine (che legano le glicoproteine alla superficie cellulare) o di fitoemoagglutinine, dimostrano chiaramente che la membrana cellulare esercita effetti regolatori sui processi che si svolgono nel nucleo.
f) Meccanismi operanti nella mitosi
Non c'è niente da aggiungere a quanto è stato già detto circa il meccanismo di duplicazione del DNA e la divisione dei cromosomi.
La migrazione dei cromosomi verso i poli nell'anafase e nella telofase non è, probabilmente, un processo attivo, ma una conseguenza delle proprietà delle fibre del fuso: i cromosomi non strisciano nella direzione del centrosoma, essi sono ‛trascinati' dalle fibre dell'aster come se i loro centromeri fossero attaccati a corde.
Poiché noi sappiamo che l'apparato mitotico contiene proteine contrattili, è molto facile immaginare che un accumulo locale di ATP potrebbe produrre un accorciamento delle fibre del fuso attaccate ai cromosomi. È anche possibile che, come nel muscolo, i filamenti del fuso scivolino l'uno sull'altro, accorciando così il fuso.
C'è infine il problema della formazione del solco: la divisione del citoplasma (plasmodieresi, citocinesi) è essenzialmente dovuta alle proprietà contrattili della membrana cellulare e del sottostante strato corticale. Tutto ciò è provato dal fatto che, se si asporta per micromanipolazione l'apparato mitotico di un uovo di riccio di mare durante la metafase o l'anafase, la divisione della cellula ora senza nucleo avviene in un tempo normale. Che la membrana cellulare e la corteccia abbiano proprietà contrattili è stato dimostrato aggiungendo ATP alle cellule che erano state trattate con glicerolo in maniera da estrarre la maggior quantità possibile del citoplasma interno. Inoltre, una proteina corticale contrattile è stata isolata dalle uova di riccio di mare in divisione: essa reagisce all'aggiunta di ATP e subisce gli stessi cambiamenti nella proporzione di residui solforati ridotti e ossidati (tiolo ⇆ disolfuro: 2 −SH ⇆ −SS−) come l'intero uovo durante il ciclo mitotico.
Per poter spiegare la formazione del solco è stata avanzata l'ipotesi di una contrazione da parte dei microtubuli; ma questa ipotesi sembra improbabile ora che la presenza di microfilamenti alla base del solco in formazione è stata dimostrata in molti tipi di cellule: questi microfilamenti sono molto piccoli (soltanto 40-50 Å) e sono stati scoperti recentemente mediante il microscopio elettronico. Un farmaco, la citocalasina B, impedisce la loro formazione, senza danneggiare i microtubuli; questa sostanza inibisce la formazione del solco nell'uovo di riccio di mare: fatto, questo, che rafforza notevolmente l'ipotesi che tali sottili microfilamenti (che sono presumibilmente dotati di contrattilità) rivestano un ruolo essenziale nella divisione del citoplasma nella telofase.
5. Interazioni nucleocitoplasmatiche
Nei capitoli precedenti, il citoplasma e il nucleo sono stati analizzati separatamente: ma la cellula dovrebbe essere considerata un tutto unico, come una unità di vita in cui nucleo e citoplasma continuamente interagiscono. È un fatto che il citoplasma anucleato mantiene per un certo tempo molte delle sue attività, compresa la vibrazione delle ciglia e anche la capacità di rigenerazione, come presto vedremo; cionondimeno, il citoplasma anucleato, anche se è ancora ‛vivente', ha perso la più fondamentale proprietà della vita: la riproduzione. In questo capitolo, descriveremo la funzione biochimica del nucleo cellulare preso come un tutto unico, senza dimenticare che gli scambi nucleocitoplasmatici possono aver luogo in entrambe le direzioni.
Tre vecchie teorie che riguardano il ruolo biochimico del nucleo cellulare possono essere menzionate, perché ci danno l'occasione di ricordare alcuni fatti importanti.
Per J. Loeb, il nucleo era il centro principale della respirazione cellulare. Noi ora sappiamo che la respirazione e la produzione di energia sono le funzioni più importanti dei mitocondri; nondimeno, il nucleo isolato è, in qualche misura, capace di produrre energia per mezzo della glicolisi e probabilmente anche della fosforilazione ossidativa.
Per E. B. Wilson, il nucleo sarebbe un sito di accumulo di enzimi e coenzimi. Questo non è vero per la maggioranza degli enzimi; ma alcuni enzimi, specialmente quelli operanti nella sintesi degli acidi nucleici e dei nucleotidi, vengono in effetti accumulati nei nuclei. Uno dei coenzimi che esercita un ruolo importante nella glicolisi e nella respirazione, il nicotinammide-adenin-dinucleotide (NAD), è sintetizzato dal nucleo, per lo meno nel fegato.
Per M. Verworn, il nucleo controllerebbe i processi di sintesi; secondo T. Caspersson, esso avrebbe un ruolo primario nella sintesi proteica. Come vedremo, la funzione principale del nucleo è proprio la sintesi: ma piuttosto sintesi di acidi nucleici, che non di proteine. Già sappiamo che la massa delle proteine è sintetizzata nel citoplasma dai polisomi, anche nel caso della sintesi di proteine nucleari, come gli istoni.
Come possiamo allora studiare il ruolo biochimico del nucleo? Ci sono tre sistemi principali: a) autoradiografia e studi biochimici sulla incorporazione di precursori specifici nelle macromolecole di cellule intatte; b) confronti di frammenti nucleati e anucleati di uova o di grandi organismi unicellulari; c) studi biochimici su nuclei isolati e su organuli citoplasmatici.
I risultati ottenuti con quest'ultimo sistema sono già stati ampiamente discussi nei due capitoli precedenti e non è necessario riprenderli in considerazione. Prenderemo quindi in esame i risultati ottenuti su cellule integre e su frammenti nucleati e anucleati.
a) Lavori su cellule integre
L'autoradiografia ha presto dimostrato che l'incorporazione dei precursori dell'RNA, per esempio di uridina radioattiva, è particolarmente rapida nel nucleo: infatti, il precursore attraversa l'intero citoplasma senza essere usato per la sintesi dell'RNA ed è incorporato per la prima volta nell'RNA nucleolare. Se la cellula è sottoposta a un breve ‛trattamento' (per pochi minuti) con uridina marcata, ed è poi rimessa in un mezzo normale non radioattivo, si può vedere che l'RNA marcato abbandona progressivamente il nucleo e si trasferisce nel citoplasma: questo esperimento ci dice chiaramente che il nucleo è più attivo del citoplasma nella sintesi dell'RNA e che una parte dell'RNA nucleare è trasferito nel citoplasma. Comunque, non è possibile dire, sulla base di questi esperimenti, se il citoplasma sia capace o no di eseguire sintesi indipendenti di RNA.
I risultati non sono così chiari nel caso della sintesi proteica: l'autoradiografia mostra una marcatura, dopo l'aggiunta di un amminoacido radioattivo, sia del citoplasma sia del nucleo. Relativamente alla cellula che viene studiata e alla lunghezza del trattamento con l'amminoacido, i nuclei possono essere più o meno fortemente marcati di quanto non lo sia il circostante citoplasma. Per poter trarre conclusioni meno generiche, sarebbe necessario un lavoro più preciso, comprendente un'analisi cinetica dettagliata del tempo necessario alla sintesi proteica e l'utilizzazione di potenti inibitori di essa, come la puromicina o la cicloesimmide: i dati autoradiografici convalidano l'ipotesi che il nucleo e il citoplasma siano entrambi capaci di sintesi proteica, ma non v'è dubbio che la maggior parte delle proteine nucleari sono di origine citoplasmatica.
L'autoradiografia, allo stato attuale, non ci può dire molto sulla identità chimica degli acidi nucleici e delle proteine che sono sintetizzate in un dato sito della cellula. Per l'identificazione delle molecole di RNA che si spostano dal nucleo al citoplasma è necessaria un'analisi biochimica: essa ha solo chiaramente dimostrato che tutti i tipi di RNA (mRNA, rRNA e tRNA) sono sintetizzati nel nucleo e si muovono da questo verso il citoplasma.
Nel caso degli RNA messaggeri, che vengono sintetizzati in siti specifici dei cromosomi, si hanno sufficienti indicazioni che essi non attraversano la membrana nucleare come tali, ma come complessi nucleoproteici: gli ‛informoferi' di Georgiev sono presenti soltanto nel nucleo e corrispondono a un complesso tra i grandi precursori dell'mRNA e le proteine; essi consentono il trasporto dei precursori dell'mRNA dalla cromatina alla membrana nucleare. Gli ‛informosomi' di Spirin sono localizzati nel citoplasma; essi sono un complesso tra mRNA e proteine, e consentono il trasporto dell'mRNA dalla membrana nucleare ai ribosomi, quando l'mRNA vi si lega per formare i poliribosomi. Il legarsi dell'RNA messaggero a una proteina protegge probabilmente il messaggero contro una degradazione causata dagli enzimi idrolitici come la ribonucleasi. Come è già stato accennato prima, il nucleo contiene molti tipi di RNA DNA-simile, di tutte le grandezze possibili che vanno da un peso molecolare di molti milioni fino a 10.000-20.000. Le molecole giganti di RNA messaggero devono essere divise in molecole più piccole e questo solleva un interessante interrogativo: come può questo spezzettamento essere fatto correttamente da un enzima come la ribonucleasi? Per poter ottenere molecole ‛significative' di RNA messaggero, gli enzimi dovrebbero essere in grado di ‛riconoscere', nel tagliare a pezzi la grande molecola di mRNA, i punti in cui le predette molecole dovrebbero terminare. Poiché le ribonucleasi conosciute non hanno lo stesso alto potere di specificità come le RNA-polimerasi, è probabile che la risposta a questo quesito sia altrove; è più probabile che la molecola gigante di mRNA contenga alcune sequenze di basi che non possono essere attaccate dalle ribonucleasi intranucleari, di modo che frammenti significativi di molecola sfuggono alla degradazione.
Sembra che soltanto il 50% delle molecole di RNA DNA-simile sfuggano alla degradazione nel nucleo, si muovano nel citoplasma (sotto forma di informosomi) e si leghino ai ribosomi: soltanto a questo 50% spetta il nome di RNA messaggero, poiché soltanto le informazioni genetiche contenute in esso saranno poi trasportate nelle proteine. Deve perciò esistere un meccanismo di selezione che consenta alle sole molecole ‛significative' di mRNA di trasferirsi nel citoplasma, mentre l'altra metà dell'mRNA - che dovrebbe essere chiamato mRNA ‛senza significato' - viene degradata nel nucleo stesso.
È probabile che la membrana nucleare, svolgendo quasi il ruolo dei doganieri o degli ufficiali di polizia ai confini tra due Stati, riconosca, con i suoi complessi porosi, l'mRNA ‛buono' da quello ‛cattivo' e passi attentamente al vaglio le due categorie. Recenti esperimenti (v. Harris, 1970), di cui diremo in seguito, indicano che anche il nucleolo potrebbe avere una parte in questa selezione tra gli mRNA che entrano nel citoplasma e quelli che rimango- no nel nucleo.
Poco si sa sulla sorte degli mRNA che restano racchiusi nel nucleo: la maggior parte viene probabilmente degradata, ma non è impossibile che alcune molecole di mRNA nucleare svolgano un ruolo di regolazione: esse potrebbero legarsi a specifici segmenti di DNA, sottraendo gli istoni che le bloccavano e impedendo l'accesso dell'RNA-polimerasi alla molecola del DNA. In altre parole, mentre gli istoni esercitano il ruolo di repressori aspecifici generici, gli RNA nucleari, come pure le proteine acide aspecifiche, ‛de-reprimerebbero', cioè attiverebbero, i geni: questi ultimi, divenendo accessibili all'RNA-polimerasi, potrebbero essere trascritti dentro molecole di mRNA ‛significative' e perciò diventare ‛attivi' (v. Paul, 1970; v. Georgiev e Samarina, 1969). Ci sarebbe da aggiungere che questa spiegazione dell'‛attivazione dei geni' e del ruolo regolatore dell'RNA nucleare e delle proteine acide della cromatina resta per ora largamente ipotetica.
Molto di più si sa circa la sintesi, da parte del nucleo, dell'RNA ribosomiale. Questo è in gran parte dovuto al fatto che, come già è stato detto nella parte riguardante il nucleolo, una considerevole amplificazione dell'rDNA, che dirige la sintesi dell'rRNA, avviene negli ovociti degli Anfibi: mentre nelle cellule normali l'rDNA corrisponde allo 0,2% del DNA totale, nel nucleo dell'ovocito avviene una amplificazione di 1.000 volte, correlata alla presenza di più di 1.000 nucleoli. Abbiamo anche visto che gli organizzatori nucleolari, che sono costituiti da rDNA, sono responsabili della sintesi della totalità dell'rRNA (il che corrisponde poi a più dell'80% dell'RNA totale della cellula).
Infine, già sappiamo che l'rDNA degli organizzatori nucleolari è circolare e che i cerchi di rDNA sono costituiti da una uguale quantità di geni che codificano per la sintesi dei due rRNA pesanti (28 S e 18 S) e da una zona intercalare molto ricca in G e C, e che non è trascritta. È il caso di aggiungere che sia i geni dell'RNA 28 S e 18 S, sia le zone intercalari hanno un peso molecolare di circa 9 milioni di dalton e si ripetono intorno a 450 volte in ogni singolo organizzatore nucleolare: queste cifre dimostrano con molta evidenza l'amplificazione dei geni dell'rRNA.
Prima di passare dall'uovo di Xenopus alle cellule somatiche, occorre sottolineare che la sintesi di almeno 10 proteine ribosomiali è coordinata con quella degli rRNA: la sintesi di queste proteine è simultanea, durante lo sviluppo, a quella degli rRNA 28 S e 18 S. Tale coordinazione non avviene nel caso del terzo tipo di rRNA, la piccola molecola di rRNA 5 S: il gene che possiede il codice per l'rRNA 5 S è ridondante (cioè esiste in molti esemplari), come quelli che codificano per gli rRNA 18 S e 28 S, ma è localizzato in altri cromosomi e non è amplificato negli ovociti di anfibio. È stato recentemente dimostrato che ovociti di Xenopus molto giovani, a uno stadio in cui possiedono solamente pochi nucleoli ma hanno già aumentato il loro rDNA, sintetizzano attivamente l'rRNA 5 S, ma non lo rRNA ad alto peso molecolare 18 S e 28 S. Il citoplasma di questi ovociti contiene pochissimi ribosomi e i loro nucleoli mancano della normale corteccia ricca di RNA. È chiaro che, a questo stadio, l'rDNA è già aumentato, ma che la sua trascrizione negli rRNA 28 S e 18 S è completamente repressa; nello stesso tempo l'rRNA 5 S viene attivamente sintetizzato.
L'rDNA fortemente amplificato degli ovociti dello Xenopus è chimicamente assai poco differente dall'rDNA delle comuni cellule somatiche (fegato, rene, eritrociti) della stessa specie. In queste cellule, i due organizzatori nucleolari sintetizzano un precursore degli rRNA 28 S e 18 S solo poco più grande di queste molecole: il suo peso molecolare è 2,5×106 dalton (invece di 2,2×106 nell'ovocito) e un tratto di DNA con peso molecolare di soli 300.000 dalton (invece di 2,5×106 nell'ovocito) ha il ruolo di ‛spaziatore', cioè di zona intercalare.
Valori leggermente più alti (2,9×106 dalton, costante di sedimentazione di 45 S) sono stati a più riprese riscontrati nel precursore degli rRNA nelle cellule dei Mammiferi. Questo RNA 45 S viene sintetizzato molto rapidamente quando si aggiunge alle cellule di Mammiferi uridina marcata ed è localizzato nei nucleoli. La complicata ‛maturazione' di questo precursore, in modo da produrre poi gli rRNA 18 S e 28 S citoplasmatici, avviene interamente nel nucleo: soltanto gli rRNA 18 S e 28 S ‛finiti' si spostano dal nucleo al citoplasma. Sui dettagli di questa ‛formazione' non esiste ancora accordo completo: secondo un recente lavoro di Penman (1970), circa il 50% del precursore 45 S è degradato nel nucleo, dove può essere recuperato sotto forma di molecole anormali di RNA 36 S e 24 S. I normali intermediari tra il precursore 45 S e gli rRNA 28 S e 18 S compiuti avrebbero, rispettivamente, i valori 32 S e 20 S. Lo schema riportato nella fig. 45, leggermente diverso, è stato proposto da Quagliarotti e altri (v., 1970): esso contiene indicazioni sia sui valori di S sia sui pesi molecolari. Secondo questo schema, tre differenti parti (P1, P2 e P3) di RNA sarebbero degradate durante la formazione del precursore nucleolare dell'rRNA 45 S.
Conosciamo invece molto meno circa la sintesi dell'RNA solubile (tRNA): appare certo che, come gli altri RNA, questi RNA dal piccolo peso molecolare (4 S) siano sintetizzati nel nucleo. Molti sono i geni attivi nella sintesi di differenti tipi di tRNA, che si trovano in diversi cromosomi (a differenza dei geni 18 S e 28 S, che sono sempre associati in coppia). È stata accertata una certa sovrabbondanza di questi geni tRNA, che esistono in circa un centinaio di copie negli ovociti di Xenopus. Comunque, le molecole di tRNA, quando escono dal nucleo, non sono ancora in grado di funzionare: per poter esplicare il loro compito, che è quello di accogliere l'amminoacido corrispondente, è assolutamente necessaria l'aggiunta di una piccola sequenza di nucleotidi. Questa sequenza, che viene aggiunta alla ‛coda' della molecola a forma di trifoglio, è costituita di soli tre nucleotidi, aventi come basi citidina, citidina e adenina (C−C−A).
L'aggiunta o la rimozione di questa sequenza finale C−C−A è una funzione del citoplasma ed è uno dei molti meccanismi in grado di controllare la sintesi proteica a livello della traduzione.
Non c'è altro da aggiungere a quanto è stato già detto circa la sintesi del DNA: già sappiamo che la sintesi del DNA cromosomico avviene solamente durante la fase S del ciclo cellulare e che, nel citoplasma, i mitocondri (e i cloroplasti, quando esistono) duplicano il loro DNA, sia continuamente sia in sincronia con la fase S, a seconda della specie.
Il citoplasma, fino a ora, sembra avere un ruolo abbastanza passivo, tranne che nel caso della sintesi proteica. Ma recenti esperimenti hanno definitivamente dimostrato che esso esercita, al contrario, una funzione molto attiva nel controllo della sintesi del DNA e dell'RNA. Questi sono gli esperimenti di J. B. Gurdon (1970) sulle uova di Xenopus e di H. Harris su ibridi di cellule somatiche. Gurdon fece degli esperimenti di trapianto nucleare iniettando, in uova non fecondate anucleate, nuclei presi da tessuti adulti (per es., dal cervello). Nel cervello questi nuclei non si dividono mai; essi hanno grandi nucleoli ricchi di RNA e sintetizzano l'rRNA molto attivamente. Quando sono innestati nel giovane citoplasma dell'uovo non fecondato e anucleato, subiscono un marcato rigonfiamento e perdono i loro nucleoli nel giro di pochi minuti. Se della timidina radioattiva viene iniettata nell'uovo assieme ai nuclei, le autoradiografie dimostrano che questi ultimi sintetizzano DNA molto velocemente. Talvolta si possono osservare cromosomi e mitosi anormali. Dopo circa 24 ore accade che i nuclei dell'embrione (stadio di blastula avanzata) cominciano a formare nucleoli e a sintetizzare rRNA. I nuclei innestati fanno esattamente lo stesso: in essi ricompaiono i nucleoli. È chiaro che la decisione tra la sintesi di DNA o di RNA è stata dettata ai nuclei dal citoplasma: un citoplasma molto giovane favorisce la sintesi di DNA, un citoplasma più vecchio favorisce la sintesi di rRNA. Non sappiamo ancora niente circa la natura dei fattori citoplasmatici che spostano la sintesi verso il DNA o l'RNA. Si sa soltanto che il rigonfiamento iniziale dei nuclei innestati non è semplicemente dovuto alla imbibizione d'acqua, ma alla assunzione di proteine citoplasmatiche da parte dei nuclei. Sappiamo anche che i fattori regolatori, qualunque sia la loro natura, non sono specie- specifici: nuclei adulti di tutte le specie finora esaminate reagiscono nella stessa maniera al giovane citoplasma circostante. Abbiamo già visto che gli ovociti adulti degli Anfibi che non sintetizzano più rRNA, contengono una proteina che si lega fortemente all'rDNA e inibisce la sintesi dell'rRNA. È probabile che questo repressore sia già presente nelle uova non fecondate e che esso impedisca la sintesi dell'rRNA nei nuclei adulti innestati. È stato anche osservato che il citoplasma di cellule in attiva divisione (tumori, fegato in rigenerazione) contiene un fattore che stimola la sintesi di DNA nei nuclei isolati dal fegato normale. L'identificazione di tali sostanze regolatrici citoplasmatiche, le quali stimolano la sintesi del DNA e reprimono la sintesi dell'RNA, sarebbe un grande passo avanti nelle nostre conoscenze circa il controllo (positivo o negativo) che il citoplasma esercita sul nucleo.
Gli esperimenti di Harris sono di tipo molto diverso, ma conducono alle medesime conclusioni. Come abbiamo già detto, è possibile fondere due differenti cellule, trattandole entrambe con virus Sendai inattivato. Il risultato di tale fusione è la formazione di un ibrido somatico, in cui due differenti nuclei (‛eterocarionti') sono disposti in un citoplasma comune (v. fig. 8). Un tipico esperimento di Harris è il seguente: un eritrocito di pollo è fuso con una cellula umana cancerosa HeLa. Il nucleo nell'eritrocito è completamente ‛represso' e non sintetizza né DNA né RNA. La cellula HeLa, al contrario, è molto attiva ed entrambe le sintesi, sia del DNA che dell'RNA, sono rilevanti. Qual è il risultato della fusione tra le due cellule? Il nucleo dell'eritrocito si gonfia e comincia a sintetizzare DNA; in seguito diventa visibile un nucleolo, segno che anche la sintesi dell'rRNA è stata riattivata.
Come negli esperimenti di Gurdon, il citoplasma ha esercitato sul nucleo represso un controllo positivo, non specie-specifico. Ulteriori esperimenti con lo stesso sistema biologico hanno dimostrato che non appaiono proteine specifiche di pollo nel citoplasma dell'ibrido prima che il nucleo della cellula di pollo abbia formato un nucleolo: questo è il motivo per cui Harris pensa che il nucleolo sia importante nella selezione tra gli mRNA che passano dal nucleo al citoplasma e quelli che non raggiungono mai il citoplasma e sono degradati nel nucleo.
Esaminiamo ora i risultati ottenuti con il lavoro effettuato su frammenti nucleati e anucleati di cellule, uova od organismi unicellulari.
b) Lavori su frammenti nucleati e anucleati
I principali materiali biologici usati fino a oggi sono l'ameba Amoeba proteus, l'alga Acetabularia, le uova di riccio di mare e i reticolociti (eritrociti immaturi) di coniglio.
Amoeba proteus. - Questa grande ameba (v. fig. 46) è visibile a occhio nudo e può essere facilmente divisa in due metà. Molto rapidamente, le metà anucleate diventano sferiche e interrompono la locomozione e la nutrizione (le amebe ingeriscono prede vive mediante fagocitosi).
Le metà nucleate, al contrario, si muovono e fagocitano normalmente; esse si dividono in due dopo due o tre giorni di nutrimento. Per poterli confrontare, i due tipi di frammenti devono perciò essere tenuti a digiuno. Quando sono digiune, le metà anucleate mostrano soltanto una leggera diminuzione della ‛vitalità': esse sopravvivono per circa 12-14 giorni, mentre le metà nucleate muoiono di inanizione dopo 14-16 giorni.
L'enucleazione produce, dopo pochi giorni, un certo numero di alterazioni morfologiche: il numero dei ribosomi diminuisce, i mitocondri si gonfiano, i corpi di Golgi scompaiono. Queste alterazioni scompaiono se nel frammento anucleato viene micro-iniettato un nucleo di ameba.
Comparazioni biochimiche tra metà anucleate e nucleate hanno, sfortunatamente, solo un limitato valore, perché il citoplasma dell'ameba contiene di regola un gran numero di microrganismi parassiti. Qualche conclusione, comunque, può sicuramente trarsi dagli esperimenti di enucleazione.
Per esempio, la respirazione e il contenuto di ATP delle metà nucleate e anucleate rimangono praticamente uguali per parecchi giorni: questo prova che, contrariamente alla vecchia concezione di J. Loeb, il nucleo non è il centro di produzione dell'energia della cellula; questo mostra inoltre che il nucleo non esercita uno stretto controllo sul funzionamento dei mitocondri. Le metà anucleate, comunque, differiscono sotto certi aspetti dalle loro controparti nucleate: non utilizzano le loro riserve alimentari (glicogeno, grassi) e bruciano invece le proprie proteine. Il risultato di questa ‛autofagia' è che, con il digiuno, il contenuto proteico diminuisce più rapidamente nelle amebe anucleate che in quelle nucleate. Non tutte le proteine sono attaccate nella stessa misura: mentre alcuni enzimi non diminuiscono affatto di attività, anche dopo 10 giorni, nelle metà anucleate, altri invece mostrano un forte e rapido declino di attività.
Le metà anucleate incorporano ancora amminoacidi nelle loro proteine e sono perciò capaci di sintesi proteiche indipendenti. Questa sintesi proteica è stata analizzata per mezzo di fini e interessanti esperimenti da Prescott e Goldstein (v., 1969): essi hanno marcato, per mezzo di un amminoacido radioattivo, le proteine di una ameba, poi hanno isolato il nucleo ‛caldo', cioè radioattivo, e lo hanno iniettato in una ameba ‛fredda' non radioattiva. Hanno così trovato, per mezzo dell'autoradiografia, che un certo numero di proteine radioattive lascia il nucleo ‛caldo', penetra nel citoplasma e infine invade il nucleo ‛freddo' dell'ameba ricevente. L'analisi di questo esperimento ha dimostrato che esistono due classi (una che si muove lentamente, l'altra velocemente) di proteine ‛migranti'. Esse sono state chiamate ‛citonucleo-proteine' dal momento che vanno e vengono dal nucleo al citoplasma: esse potrebbero avere un ruolo importante nel controllo dell'attività genetica, ma tutto ciò resta ancora un'ipotesi.
Confrontiamo ora gli effetti della rimozione del nucleo sul metabolismo dell'RNA con ciò che è stato appena detto circa il metabolismo delle proteine. Il contenuto in RNA delle metà anucleate diminuisce molto più rapidamente del loro contenuto proteico: in 5 giorni scende a circa 1/3 del suo valore iniziale, e si può con sicurezza concludere che il nucleo esercita un controllo molto più diretto sulla sintesi dell'RNA che non su quella delle proteine. La composizione in basi dell'RNA non cambia considerevolmente nel citoplasma anucleato e gli esperimenti hanno mostrato che l'rRNA è particolarmente sensibile alla rimozione del nucleo. Quando il nucleo di un'ameba che ha incorporato precursori radioattivi nell'RNA è innestato in un'ameba normale, la radioattività si sposta molto velocemente dal nucleo al citoplasma. Ma c'è una leggera radioattività nel nucleo del ricevente, cioè l'ameba ‛fredda': se le citonucleo-proteine si muovono liberamente dal nucleo al citoplasma e viceversa, il movimento dell'RNA è quasi unidirezionale (dal nucleo al citoplasma).
C'è una certa incorporazione di uridina nell'RNA e di timidina nel DNA quando questi precursori vengono aggiunti a frammenti anucleati: ma la già menzionata presenza di microrganismi contaminanti nel citoplasma delle amebe non permette di trarre delle conclusioni, tranne che la sintesi dell'RNA nel citoplasma anucleato deve essere di scarsa importanza. In realtà, una tale incorporazione potrebbe rappresentare non una vera e propria sintesi, ma l'aggiunta della sequenza terminale −CCA a molecole già esistenti di tRNA.
Acetabularia. - L'interesse per quest'alga gigante unicellulare (lunga 5 cm o più) nacque quando J. Hämmerling dimostrò che essa è capace di un'estesa rigenerazione in assenza del nucleo. La fig. 47 rappresenta l'alga completamente sviluppata: il nucleo si trova all'estremità inferiore, basale dell'alga. All'estremità apicale c'è un ombrello, normalmente chiamato ‛cappello', che serve per la formazione dei gameti durante la riproduzione sessuale. Questo cappello ha una morfologia specie-specifica: è facile riconoscere, dal suo aspetto, le differenti specie di Acetabularia.
È facile tagliare l'alga, subito prima della formazione del cappello, in due metà: come Hämmerling (v., 1953) scoprì, le metà anucleate non solo sopravvivono per lungo tempo (fino a tre mesi), ma possono perfino rigenerare un cappello perfettamente normale (v. fig. 48). Infatti, i cappelli si formano più rapidamente in assenza che in presenza del nucleo, ma cessano di crescere dopo poche settimane. Naturalmente, esse rimangono sempre sterili, dal momento che non possiedono il nucleo. Hämmerling dai suoi esperimenti trasse la conclusione che la formazione del cappello è dovuta a ‛sostanze morfogenetiche' che vengono formate dal nucleo e distribuite secondo un gradiente di rigenerazione decrescente dall'apice alla base (v. fig. 49): queste sostanze rimangono stabili, nel citoplasma anucleato, per parecchie settimane Che le sostanze morfogenetiche siano realmente formate dal nucleo è stato dimostrato tramite eleganti esperimenti di innesti interspecifici compiuti da Hämmerling: egli combinò la metà nucleata di una specie con la metà anucleata di un'altra e scoprì che, anche se qualche volta si può formare per primo un cappello ‛ibrido', il cappello finale è sempre del tipo nucleare. Sembra che ci sia una competizione tra le sostanze morfogenetiche preesistenti immagazzinate nella metà anucleata, e le sostanze appena sintetizzate formate dal nucleo dell'altra specie; questa competizione, alla fine, è vinta dal nucleo.
Un buon numero di lavori biochimici sono stati fatti sull'Acetabularia, sia nel laboratorio di Hämmerling sia in quello di Brachet. Un primo punto accertato è che la rimozione del nucleo non esercita importanti effetti sulla produzione dell'energia, tranne il fatto che il contenuto di ATP dei cloroplasti diminuisce dopo periodi prolungati di enucleazione. Ma né la respirazione, né la fotosintesi sono notevolmente colpite durante i primi giorni successivi alla rimozione del nucleo. Questo risultato indica che il nucleo dell'alga non esercita uno stretto controllo sull'attività dei cloroplasti e dei mitocondri.
Un'osservazione dovrebbe farsi ancora circa la fotosintesi: se si danno alle alghe periodi successivi di luce e di buio (per es., 12 ore di luce e 12 ore di buio), esse acquistano un ‛ritmo circadiano' di capacità fotosintetica. Questo ritmo è conservato per molte settimane in frammenti anucleati; ma, se si combina una metà nucleata di un'alga avente un dato ritmo con una metà anucleata che ha un ritmo differente, il ritmo nucleare ha il sopravvento su quello citoplasmatico. Nello stesso modo, frammenti anucleati di alghe che hanno perso il loro ritmo circadiano di capacità fotosintetica per prolungato trattamento con antibiotici, ne acquistano uno se sono innestati su un frammento nucleato di un'alga che ha un ritmo normale. Pertanto i ritmi circadiani, che sono importanti eventi biologici, possono persistere in assenza del nucleo; ma la regolazione dell'‛orologio circadiano' è effettuata dal nucleo, se esso è presente.
Molto importante è il fatto che la sintesi proteica, compresa quella di un certo numero di enzimi specifici, va avanti per molte settimane in assenza del nucleo. Per esempio, l'attività delle fosfatasi aumenta fortemente quando si forma il cappello, sia che il nucleo sia presente, sia che esso manchi. Poiché la sintesi proteica è controllata dai geni, la spiegazione logica dei fatti è la seguente: nell'Acetabularia certi RNA messaggeri sarebbero estremamente stabili e capaci di una lunga sopravvivenza in assenza di nucleo. Le sostanze morfogenetiche di Hämmerling apparterrebbero a questa classe di mRNA molto stabili. Il fatto che gli inibitori della sintesi proteica citoplasmatica (puromicina, cicloesimmide) impediscano completamente la formazione del cappello suggerisce inoltre l'ipotesi che le proteine specifiche richieste per la formazione del cappello siano costruite sotto il controllo di questi mRNA citoplasmatici, che erano stati sintetizzati molto prima nell'interno del nucleo.
Comunque, un'approfondita analisi della sintesi degli enzimi nell'Acetabularia mostra che non sarebbe da trascurare un ruolo ‛specifico' del nucleo. Per esempio, la sintesi delle fosfatasi può essere regolata, in molti organismi, con la modificazione del contenuto di fosfato inorganico del mezzo: se quest'ultimo non contiene fosfato, o ne contiene poco, la sintesi delle fosfatasi viene aumentata. Nell'Acetabularia, soltanto i frammenti nucleati reagiscono in questa classica maniera all'acqua di mare priva di fosfato: la regolazione della sintesi degli enzimi tramite cambiamenti nella composizione del mezzo esterno si perde in assenza del nucleo. È stato anche dimostrato che, negli innesti interspecifici tra Acetabularia mediterranea e Acetabularia crenulata, un enzima (malicodeidrogenasi) presente nel frammento anucleato viene progressivamente sostituito da un differente enzima, che è sintetizzato continuamente sotto il controllo dei geni nucleari.
Tutte queste osservazioni concordano con l'ipotesi che esistano molecole stabili di mRNA nel citoplasma dell'Acetabularia, ma una prova vera e propria potrebbe essere ottenuta soltanto per mezzo di studi sul metabolismo dell'RNA nell'alga. Tali studi hanno finora rafforzato l'attendibilità dell'ipotesi suddetta; tuttavia la prova decisiva ancora manca. Per esempio, l'ipotesi fa prevedere che, se l'mRNA è distrutto (per mezzo, supponiamo, di un trattamento in vivo dell'alga con ribonucleasi), le metà anucleate diventeranno irreversibilmente incapaci di rigenerare, mentre nel caso dei frammenti nucleati c'è da aspettarsi una reversibilità. D'altra parte, se la sintesi dell'mRNA fosse inibita dall'actinomicina, i frammenti anucleati rigenererebbero ancora i cappelli, mentre le metà nucleate non li formerebbero fintanto che fossero tenute in presenza dell'inibitore. La fig. 50 mostra, in maniera schematica, che queste due ‛previsioni' possono essere confermate dagli esperimenti.
L'autoradiografia, in accordo con l'ipotesi precedente, mostra che l'RNA prima viene sintetizzato nel nucleo, poi passa nell'interno del citoplasma e si distribuisce secondo un gradiente di morfogenesi apico-basale. Esperimenti biochimici hanno inoltre dimostrato che certi tipi di molecole di RNA possono davvero ‛sopravvivere' per lunghissimo tempo nel citoplasma: dopo aver marcato le alghe con uridina radioattiva per un'ora e averle poi lasciate immerse in normale acqua di mare, si può ancora reperire, dopo tre mesi, un tipo di RNA radioattivo molto stabile.
Comunque, non è stato finora possibile provare la sintesi dell'mRNA da parte del nucleo, per motivi inaspettati: se si confronta la sintesi di RNA in frammenti nucleati e anucleati, non è assolutamente possibile trovare alcuna differenza in un periodo di poche settimane. In due settimane, il contenuto in RNA si raddoppia in entrambi i tipi di frammenti e non è possibile individuare la sintesi di uno speciale tipo di RNA nelle metà nucleate. Questo risultato è dovuto alla sintesi molto intensa di RNA che avviene nei cloroplasti, sia che il nucleo sia presente sia che esso manchi.
Il grande nucleo dell'Acetabularia contiene un enorme nucleolo, ma così poco DNA che esso non può essere svelato per mezzo di metodi citochimici. La sua morfologia e il suo contenuto in RNA sono fortemente influenzati dal citoplasma circostante: per esempio, se l'alga è tenuta al buio per qualche giorno, in maniera da arrestare la produzione di energia da parte dei cloroplasti, il nucleo si restringe; il suo nucleolo, a forma di cordone, diventa sferico e perde molto del suo RNA (v. fig. 51). Questi cambiamenti sono invertiti se l'alga viene di nuovo illuminata. Tali esperimenti mostrano che il nucleo è largamente influenzato dal citoplasma per quanto riguarda la produzione di energia. Altri esperimenti hanno mostrato che, come nelle uova di Xenopus, è il citoplasma che ‛decide' se il nucleo sintetizzerà RNA o DNA: se si innesta lo stelo dell'alga che sta già formando un cappello su di un frammento nucleato di alga giovane, in quest'ultima la mitosi sarà accelerata. Al contrario, innestando uno stelo giovane su un frammento nucleato vecchio, si impedisce al nucleo di entrare in mitosi (v. Hämmerling, 1953).
Come è stato mostrato in modo schematico nella fig. 50, i cappelli formati dai frammenti anucleati in presenza di actinomicina sono più piccoli che nei controlli (confrontare A con C): questo potrebbe essere dovuto all'unione della actinomicina con il DNA cloroplastico e si potrebbe veramente provare che i cloroplasti dell'Acetabularia contengono DNA, che si combina in vivo con l'actinomicina. Questa osservazione suggerisce l'idea che il DNA cloroplastico potrebbe duplicarsi in assenza del nucleo: gli esperimenti hanno provato che questo è quanto accade, e che il contenuto di DNA delle metà anucleate può raddoppiarsi nel giro di una settimana.
Anche il conteggio dei cloroplasti dimostra che essi possono aumentare di numero in assenza del nucleo. La loro moltiplicazione è, comunque, più lenta nelle metà anucleate che in quelle nucleate; nelle prime, molti cloroplasti sono grandi, a forma di manubrio, come se fossero stati capaci di accrescersi, ma non di dividersi, in assenza di nucleo.
Questi esperimenti dimostrano chiaramente che i cloroplasti possiedono un grado molto elevato di autonomia nei riguardi del nucleo: essi possono duplicare il proprio DNA per parecchi giorni e anche dividersi. Il DNA cloroplastico è trascritto e tutti i tipi di RNA cloroplastici sono sintetizzati in assenza del nucleo. Infine, anche in cloroplasti isolati di Acetabularia può aversi traduzione di mRNA cloroplastici e sintesi di proteine cloroplastiche. Ma l'informazione genetica del DNA cloroplastico non è sufficiente per una piena autonomia: mRNA di origine nucleare sono indispensabili per la sintesi di alcuni degli enzimi contenuti nei cloroplasti, per la continua divisione di questi organuli cellulari e infine, cosa non tra le meno importanti, per la morfogenesi. Recenti lavori sperimentali hanno dimostrato che molti costituenti dei cloroplasti sono in realtà sintetizzati sotto il controllo nucleare, come appare evidente nelle esperienze in cui un frammento anucleato di specie A è innestato in un frammento nucleato di specie B: dopo appena una settimana, molti enzimi cloroplastici del frammento anucleato A presentano variazioni e divengono di tipo B. Lo stesso accade per le proteine che formano le membrane dei cloroplasti e anche per le proteine dei ribosomi cloroplastici; ma, in questi casi, le variazioni non si rendono evidenti prima di molte settimane: ciò dimostra che il controllo esercitato sulla sintesi delle proteine dei cloroplasti varia a seconda della proteina che è sintetizzata. Sebbene i cloroplasti, nell'Acetabularia, mostrino un elevato grado di autonomia - in accordo con l'ipotesi che i cloroplasti siano alghe unicellulari divenute simbiotiche nel corso dell'evoluzione - è chiaro che in definitiva essi dipendono dall'attività dei geni nucleari.
Uova di riccio di mare. - Un paragone tra le uova di riccio di mare e l'Acetabularia è opportuno, dal momento che esso dimostra che il citoplasma anucleato di una cellula animale è di molto inferiore a quello di una cellula di pianta verde sia per quel che riguarda la morfogenesi sia per la sintesi delle macromolecole. Le uova di riccio di mare non fecondate possono facilmente essere divise in due metà mediante centrifugazione (v. fig. 52): la metà leggera e trasparente contiene il nucleo, gocce di grasso, ribosomi, poche placchette vitelline e molti mitocondri; la metà pesante, rossa, contiene gli stessi costituenti di quella leggera, ma in proporzioni differenti, non possiede il nucleo e in più è presente in essa un pesante pigmento rosso scuro. Se vengono fecondati, entrambi i frammenti possono svilupparsi in larve normali. Se vengono attivati con un agente partenogenetico, per esempio con acqua di mare ipertonica, soltanto le metà nucleate si sviluppano normalmente: i frammenti anucleati possono, al massimo, subire una anormale segmentazione, che ha molto poco in comune con la tipica segmentazione delle uova di riccio di mare (v. fig. 53).
Questo è invero uno sviluppo molto povero se paragonato alla formazione degli eleganti cappelli dell'Acetabularia.
La sintesi proteica è eccezionalmente scarsa nelle uova di riccio di mare non fecondate; ma essa è rapidamente (anche nel giro di pochi minuti) e assai fortemente accresciuta dalla fecondazione (A. Monroy). Dapprima si ritenne che questo aumento della sintesi proteica potesse essere attribuito alla produzione, da parte dei nuclei dell'uovo e dello spermatozoo, di nuove molecole di mRNA in grado di attaccarsi a ribosomi preesistenti e di formare polisomi attivi. Ma gli esperimenti compiuti su frammenti di uova nucleati e anucleati hanno reso impossibile questa spiegazione: frammenti nucleati e anucleati di uova non fecondate sintetizzano molto poco, se non affatto, le proteine. Ma, se vengono attivati dall'acqua di mare ipertonica, entrambi sintetizzano proteine rapidamente e molto attivamente. In effetti, la sintesi proteica è anche più abbondante nelle metà anucleate che in quelle nucleate, dopo l'attivazione artificiale.
Questi e altri esperimenti hanno portato alla conclusione che l'uovo non fecondato di riccio di mare contiene una riserva di molecole di mRNA stabili, le quali sono ‛mascherate' in modo tale che non possono reagire con i ribosomi. Questi ultimi sono anormali nell'uovo non fecondato (v. Monroy, 1965): essi contengono un inibitore che impedisce loro di legarsi all'mRNA preesistente. Ma molti altri meccanismi di controllo negativo della sintesi proteica esistono nell'uovo non fecondato di riccio di mare: parziale deficienza nei fattori proteici che promuovono il legame dell'mRNA ai ribosomi e l'allungamento della catena proteica; mancanza degli enzimi attivanti gli amminoacidi; presenza di tRNA incompleti; presenza di un inibitore della sintesi proteica nei ribosomi di uova non fecondate, ecc. Il blocco della sintesi proteica, nelle uova di riccio di mare non fecondate, è quindi dovuto a una repressione multipla a livello della traduzione.
Metà nucleate e anucleate contengono quantità quasi uguali di RNA e di DNA. Questi acidi nucleici vengono sintetizzati, come le proteine, anche in assenza del nucleo? La risposta è positiva per l'RNA, probabilmente negativa per il DNA. Non c'è sintesi di RNA nei frammenti anucleati prima dell'attivazione con acqua di mare ipertonica; dopo pochi minuti, la sintesi dell'RNA comincia e diventa facilmente misurabile. Questa sintesi non è limitata alla semplice aggiunta della sequenza −CCA alle molecole di tRNA incomplete: tre differenti tipi di RNA sono sintetizzati nelle metà anucleate, con costanti di sedimentazione di 15S, 13S e 11S. Le specie 13S e 11S corrispondono agli rRNA mitocondriali; la 15S può essere un precursore di questi rRNA mitocondriali o forse un mRNA. È stato dimostrato che gli RNA mitocondriali - al pari degli RNA cloroplastici nell'Acetabularia - non possono legarsi ai ribosomi citoplasmatici e quindi non possono formare poliribosomi. La intensa sintesi proteica che ha luogo nei frammenti anucleati delle uova di riccio di mare, dopo l'attivazione partenogenetica, non è quindi dovuta alle molecole di mRNA di origine materna. Tra questi mRNA in ‛maschera' materna, quelli che dirigono la sintesi della tubulina e degli istoni sono già stati identificati nei frammenti anucleati delle uova di riccio di mare.
Nelle uova di riccio di mare, il DNA citoplasmatico è localizzato nei mitocondri: finora non è stato possibile determinare una sintesi di DNA mitocondriale nelle metà nucleate attivate di uova di riccio di mare. Sembra quasi che, in questo materiale, il DNA mitocondriale possa essere trascritto nell'RNA, ma non duplicato. Perciò non è probabile che i mitocondri possano aumentare di numero nei frammenti anucleati di uova di riccio di mare, anche dopo attivazione partenogenetica: questo è in forte contrasto con quanto abbiamo visto per i cloroplasti nell'Acetabularia. Una tale differenza tra mitocondri e cloroplasti non sorprende dal momento che è noto che un cloroplasto contiene come minimo 100 volte più DNA (cioè informazione genetica, a meno che il DNA cloroplastico sia sovrabbondante) di un mitocondrio.
Reticolociti. - I reticolociti sono il risultato di un esperimento di enucleazione fatto dalla natura stessa durante la formazione degli eritrociti: questi ultimi, nei Mammiferi, non hanno nucleo e non contengono quasi per niente RNA. Sono pieni di emoglobina e non sintetizzano più proteine. I reticolociti sono eritrociti immaturi: essi hanno già perso il loro nucleo, ma contengono ancora ribosomi e sintetizzano attivamente emoglobina. Durante la maturazione dei reticolociti, il nucleo e i mitocondri vengono espulsi: i polisomi si disgregano e i ribosomi liberi infine degenerano.
Sono stati isolati i polisomi dei giovani reticolociti: essi sono specializzati nella sintesi di una sola proteina, che è l'emoglobina. È stato possibile isolare, nel laboratorio di H. Chantrenne (v. Chantrenne e altri, 1967), l'mRNA che lega insieme i ribosomi dei reticolociti: questo è l'RNA messaggero dell'emoglobina. Come è da aspettarsi dalla grandezza delle catene proteiche che formano l'emoglobina, si tratta di una molecola piuttosto piccola di RNA (9 S); è associato con una proteina, come avviene anche nel caso degli ‛informosomi'. Molto recentemente, Lockard e Lingrel (v., 1969), hanno ottenuto la sintesi dell'emoglobina in vitro con l'mRNA 9 S: questa è la prima volta che un mRNA isolato da una cellula animale ha diretto, in un sistema acellulare, la sintesi di una proteina specifica. Più recentemente Gurdon (v. Gurdon e altri, 1971) iniettando mRNA dell'emoglobina estratto dai reticolociti di coniglio negli ovociti del rospo Xenopus, ha potuto ottenere una notevole sintesi di emoglobina di coniglio nelle uova di questo anfibio. Questi esperimenti dimostrano chiaramente che gli ovociti di rospo contengono in eccesso tutti gli apparati necessari per la sintesi proteica; ma l'identità della proteina sintetizzata è specificata dall'mRNA che si lega ai ribosomi.
c) Conclusioni
Ricapitolando, gli esperimenti eseguiti sia su cellule integre sia su loro frammenti anucleati conducono alle stesse conclusioni generali, e precisamente alle seguenti: 1) il nucleo non è un centro di produzione dell'energia: al contrario, esso è in gran parte dipendente dai mitocondri (e dai cloroplasti nelle piante verdi) per il mantenimento delle sue attività sintetiche; 2) il nucleo è il centro principale della sintesi del DNA e dell'RNA. Esso controlla la sintesi proteica, che è una funzione del citoplasma, mediante la produzione e la diffusione di tutti i tipi di RNA (mRNA, rRNA e tRNA); 3) gli organuli cellulari contenenti DNA, cioè i mitocondri e i cloroplasti, mostrano un certo grado di ‛autonomia' nei confronti del nucleo. Questa autonomia, specialmente la capacità di duplicarsi in assenza del nucleo, è più grande per i cloroplasti (che hanno un più alto contenuto di DNA) che non per i mitocondri; 4) il citoplasma ha una parte importante, ma probabilmente non altamente specifica, nell'orientare il nucleo sia verso la duplicazione del DNA (di solito seguita dalla mitosi), sia verso la trascrizione del DNA (seguita dalla sintesi dell'RNA del nucleo e delle proteine citoplasmatiche).
6. Differenziamento della cellula
Come è già stato detto, le cellule che compongono i vari tessuti sono morfologicamente, fisiologicamente e biochimicamente differenti: esse hanno subito il differenziamento cellulare durante lo sviluppo embrionale. La fig. 54 mostra un esempio di differenziamento cellulare: le cellule della cartilagine (sopra) e del muscolo (sotto) hanno la medesima origine embrionale. Esse sono già molto differenti in un embrione di anfibio di 7 giorni: le cellule cartilaginee sono circondate da una matrice omogenea, costituita di glicoproteine, mentre le cellule muscolari allungate stanno fondendosi in un ‛sincizio' e formando le fibrille delle proteine contrattili, la miosina e l'actina. Queste cellule hanno già struttura, funzione e composizione chimica differenti.
Come cellule embrionali identiche subiscano il differenziamento, rimane uno dei grandi misteri della biologia dei nostri giorni e sarà il maggiore problema dei biologi molecolari di domani. La questione ha più che un interesse accademico e la sua soluzione getterebbe luce sul problema del cancro. Le cellule neoplastiche, in generale, non si dividono più rapidamente delle cellule normali: ma invece di differenziarsi, continuano a dividersi e infine invadono l'intero organismo.
Molte teorie sono state proposte per cercare di spiegare come un numero limitato di cellule, localizzate in una parte ben definita dell'embrione che si sta sviluppando, subiscano un differenziamento a un dato stadio del proprio sviluppo; ma così pochi fatti sono accertati che è ancora molto difficile pensare di provarle sperimentalmente.
La teoria oggi più accettata è quella dell'attivazione differenziale del gene: sotto l'influenza di circostanze ancora sconosciute, un dato gene o gruppo di geni diventerebbe attivo e dirigerebbe la sintesi delle proteine specifiche caratteristiche dei vari tipi di cellule (per es., l'emoglobina negli eritrociti, le proteine contrattili actina e miosina nelle cellule muscolari ecc.). Questa teoria postula che tutte le cellule abbiano gli stessi geni, ma che i meccanismi regolatori agiscano in maniera tale da attivare specialmente alcuni di loro, mentre il resto del genoma rimane in uno stato represso. Purtroppo si sa così poco della regolazione dei geni negli Eucarioti che, al massimo, si può supporre che le proteine cromosomiche (gli istoni e specialmente le proteine acide) abbiano una parte in questa funzione regolatrice.
Altri autori pensano che il DNA stesso possa essere modificato durante il differenziamento, forse come risultato di una specifica metilazione delle basi (E. Scarano). Un'altra possibilità sarebbe che certi geni (per es., quelli per la sintesi dell'emoglobina) subiscano una duplicazione individuale al tempo del differenziamento cellulare: questi geni sarebbero specificamente duplicati (e conseguentemente amplificati), mentre il resto del genoma non lo sarebbe. Questa possibilità non dovrebbe essere scartata in modo superficiale: abbiamo visto che i geni per la sintesi dell'rRNA subiscono una considerevole duplicazione durante l'ovogenesi degli Anfibi; il risultato di questo eccezionale moltiplicarsi di geni è la sintesi, con l'accrescimento dell'ovocito, di un enorme numero di ribosomi. La duplicazione selettiva dei geni è stata anche dimostrata nei cromosomi giganti di certi Insetti: abbiamo visto che, come risposta a una stimolazione ormonica, una singola banda subisce la duplicazione del DNA, mentre il resto del cromosoma rimane inattivo. Ma recenti esperimenti hanno reso questa attraente ipotesi molto inverosimile: è stato definitivamente dimostrato che le cellule che producono emoglobina nei conigli o seta nei bachi da seta contengono lo stesso numero di geni per l'emoglobina o per la seta contenuto in tutte le altre cellule; inoltre, questi geni esistono in una singola copia nel genoma aploide.
Si sono anche fatti ripetuti tentativi per spiegare il differenziamento sulla base dell'esistenza di particelle citoplasmatiche autoduplicantesi, i ‛plasmageni': questa possibilità non può essere esclusa, ma non esiste per ora una prova sicura dell'esistenza di tali ipotetici plasmageni. Non c'è alcuna prova diretta che gli organuli citoplasmatici contenenti DNA (mitocondri e cloroplasti) esercitino una parte di maggior rilievo nel differenziamento cellulare; un ruolo accessorio non può, comunque, essere escluso.
Lasciamo adesso le teorie per i fatti: il più importante di questi viene dagli esperimenti di J. B. Gurdon (v. Gurdon e Woodland, 1970), il quale ha chiaramente dimostrato che i nuclei di cellule già differenziate (presi da vecchie larve o da cellule adulte provenienti da colture di tessuti) mantengono le stesse capacità di sviluppo caratteristiche del nucleo molto giovane dell'uovo appena fecondato. Se questi nuclei sono trapiantati in uova anucleate non fecondate, è occasionalmente possibile, uno sviluppo normale e completo: alcuni dei nuclei dell'adulto sono quindi totipotenti, cioè, capaci di dirigere tutti i tipi possibili di differenziamento cellulare. Per esempio, dopo il trapianto di nuclei dal rene, dal polmone o dalla pelle in uova anucleate non fecondate, si possono ottenere girini vitali e mobili, che contengono tutti i tipi possibili di cellule differenziate (nervi, muscoli, cartilagine, intestino ecc.): questo fatto dimostra chiaramente che le cellule adulte hanno conservato tutti i geni necessari per un completo differenziamento embrionale. Tali risultanze contrastano con le teorie che implicano modificazioni chimiche irreversibili delle molecole stesse di DNA.
Il differenziamento embrionale è, in generale, preceduto dalla sintesi di mRNA di vita assai breve. Questa sintesi iniziale è seguita da quella di mRNA più stabili, che vengono prodotti quando diventano visibili il differenziamento istologico e la sintesi di proteine specifiche. Questo spiega, per esempio, come l'actinomicina può impedire il differenziamento delle cellule che formeranno il cuore (inibendo la sintesi degli mRNA di breve vita), mentre non impedisce lo sviluppo e la contrattilità delle giovani fibre muscolari del cuore (come già sappiamo l'actinomicina inibisce la sintesi dell'RNA, ma non ha nessun effetto sugli mRNA già sintetizzati). Sia questi esperimenti, sia quelli già menzionati per l'Acetabularia, dimostrano chiaramente che molti meccanismi di controllo, tanto a livello della trascrizione quanto a livello della traduzione, devono operare nell'embrione per assicurare il differenziamento cellulare.
Ma gli embrioni non sono i soli materiali utilizzabili per lo studio del differenziamento cellulare: una grande quantità di lavoro si sta svolgendo attualmente su cellule coltivate in vitro. Esse costituiscono un facile sistema per lo studio sia del differenziamento cellulare sia dello sdifferenziamento (perdita di differenziamento): cellule embrionali indifferenziate possono essere coltivate in un mezzo artificiale in cui esse subiranno un differenziamento sia morfologico che biochimico. D'altra parte, cellule differenziate (cartilaginee, muscolari, nervose, cellule pigmentifere, ecc.) possono anche essere coltivate in vitro: è allora facile vedere se esse trattengono o perdono la loro condizione differenziata sotto varie condizioni sperimentali. In tali studi, i criteri biochimici danno spesso più precise informazioni che non la semplice osservazione morfologica delle cellule coltivate: è possibile misurare gli enzimi che sono specifici di certi tipi di cellule, o seguire la sintesi o la perdita di proteine tipiche come l'emoglobina o l'actomiosina. I metodi immunologici sono particolarmente utili: in virtù di essi è possibile dimostrare se alcuni specifici antigeni (in particolare gli antigeni superficiali della membrana cellulare) sono presenti o si sono perduti durante la coltivazione in vitro delle cellule.
La quantità di lavoro attualmente fatta su tali colture di tessuti è così estesa che tutto ciò che possiamo fare è di indicare, in un modo generico e incompleto, un buon numero di promettenti approcci al problema del differenziamento cellulare. Per esempio, certi ormoni (i corticosteroidi, in particolare) provocano, in cellule coltivate in vitro, la sintesi di enzimi specifici: l'analisi degli scambi metabolici provocati dall'ormone in tali cellule ha portato a un marcato progresso nelle nostre conoscenze dei meccanismi molecolari dell'azione ormonale, in particolare a livello della trascrizione e della traduzione.
Le cellule coltivate in vitro costituiscono anche un materiale ideale per lo studio dei problemi immunologici: è possibile determinare, in una grande popolazione di cellule, quelle che reagiscono a un antigene con la produzione di anticorpi. Lo stesso sistema è anche ben utilizzabile negli studi sull'infezione e la moltiplicazione virale: di grandissima importanza per il nostro attuale problema del differenziamento cellulare è il fatto che alcuni virus, i cosiddetti virus oncogeni, provocano la trasformazione maligna di cellule sane; le proprietà fisiologiche e biochimiche di tali cellule trasformate sono naturalmente ampiamente modificate, in particolare la composizione chimica della loro membrana cellulare.
Particolarmente interessante è il lavoro attualmente fatto, in parecchi laboratori, sugli effetti della 5-bromo-2′-desossiuridina (BUDR) sul differenziamento cellulare in vitro. Questa sostanza è un ‛analogo' della timidina, che è un normale precursore del DNA durante la sua sintesi: poiché le due sostanze sono chimicamente molto simili tra loro, le cellule facilmente fanno errori durante la duplicazione del DNA e sintetizzano molecole di DNA in cui una parte dei residui di timidina sono sostituiti dalla BUDR. Se si aggiunge BUDR a cellule in via di differenziamento, accade qualche cosa di molto importante: avendo la precauzione che la BUDR sia aggiunta in concentrazioni così basse da non interferire con la divisione della cellula, si assiste a un arresto del differenziamento. Si tratta, come è stato dimostrato, del caso delle cellule pigmentifere (le quali diventano incolori), delle cellule muscolari (che non formano fibrille contrattili), delle cellule cartilaginee (che non formano quella matrice omogenea mostrata nella fig. 43), delle cellule nervose (che non formano più terminazioni nervose), ecc. In tutti questi casi è inibita la sintesi della proteina specifica della cellula: non c'è sintesi di melanina nelle cellule pigmentifere, di actina e miosina nelle cellule muscolari, di glicoproteine nelle cellule cartilaginee, ecc.
Un aspetto molto importante è che questi particolari effetti della BUDR possono essere eliminati quando la sostanza è rimossa dalle cellule: l'inibizione del differenziamento cellulare, in questo caso, è quindi reversibile. La spiegazione che è stata proposta è la seguente: mentre tutte le cellule debbono produrre un gran numero di proteine, in particolare enzimi, per sopravvivere, le cellule differenziate producono, in più, un tipo specifico di proteine. Tali proteine specifiche sono state chiamate proteine ‛di lusso' in contrapposizione alle universali proteine ‛economiche'. La BUDR contrasta la sintesi delle proteine ‛di lusso' senza intervenire in quella delle proteine ‛economiche'. In considerazione del fatto che la BUDR può alterare la sintesi del DNA, è stata avanzata l'ipotesi che i geni codificanti la sintesi delle proteine ‛di lusso' siano amplificati: essi esisterebbero in un più ampio numero di copie che non gli altri geni e potrebbero essere duplicati fuori del periodo S del ciclo della cellula. La BUDR bloccherebbe selettivamente la duplicazione di questi geni. Per il momento questa è nulla più che un'ipotesi, come abbiamo visto non molto probabile: infatti, sappiamo che la BUDR non agisce soltanto sulla sintesi del DNA, ma che essa può anche modificare le proprietà della membrana cellulare. Una completa spiegazione degli effetti della BUDR verrà certamente, presto o tardi; nel frattempo, questa sostanza manterrà un eccezionale interesse per lo studio del differenziamento cellulare.
Un altro potente mezzo per questo studio è l'ibridazione cellulare: abbiamo già visto che le cellule somatiche si possono fondere e possono quindi formare eterocarionti quando vengono trattate con virus emolitici inattivati. Questa tecnica, che è stata particolarmente usata da H Harris (v., 1970), è l'equivalente, per le cellule somatiche, dei trapianti nucleari fatti su uova non fecondate da J. B,. Gurdon. Abbiamo già visto che l'ibridazione cellulare ha dato informazioni estremamente importanti circa le interazioni nucleocitoplasmatiche; lo stesso metodo darà indubbiamente informazioni non meno importanti circa il differenziamento cellulare. È naturalmente possibile fondere insieme cellule differenziate e indifferenziate: provocherà una tale fusione il differenziamento delle cellule indifferenziate? O, al contrario, si arresterà il differenziamento nelle cellule che già si stanno differenziando? La regola generale, in questi esperimenti, è che il carattere differenziato è ‛estinto' nell'ibrido: una cellula giovane non differenziata annulla il differenziamento dopo la fusione. Le cellule pigmentifere cessano di produrre pigmento (melanina) dopo la fusione con una cellula embrionale; lo stesso avviene per le cellule cerebrali o cartilaginee. Se una cellula carcinomatosa, maligna, è fusa con una cellula normale, essa perde di solito i caratteri di malignità. Sfortunatamente, quando queste cellule ibride sono coltivate per molte generazioni, esse perdono progressivamente un certo numero dei loro cromosomi: in particolare, quando una cellula umana è fusa con una cellula di topo, i cromosomi umani sono progressivamenti perduti (una situazione che ha consentito notevoli progressi nel campo della genetica umana). Ma quando è stato perso un certo numero di cromosomi, il carattere soppresso (una proteina ‛di lusso' nelle cellule differenziate, la malignità nelle cellule cancerose) abitualmente riappare.
Gli effetti di altre sostanze chimiche sulla crescita e sul differenziamento sono stati ora ampiamente studiati: queste sostanze sono i nucleotidi ciclici (cAMP, cGMP), presenti in tutte le cellule. Un aumento nel contenuto di cAMP in una cellula determina l'arresto della divisione cellulare e induce il differenziamento. Il cGMP esercita effetti opposti; è quindi possibile che il bilancio tra le concentrazioni del cAMP e del cGMP sia responsabile dell'orientamento della cellula verso la divisione o, al contrario, verso il differenziamento.
Questi esperimenti sugli ibridi cellulari e sugli effetti dei nucleotidi ciclici dimostrano chiaramente che nella cellula svolgono un ruolo importante opposti meccanismi regolatori: i meccanismi positivi inducono il differenziamento, mentre i controlli negativi lo sopprimono e favoriscono la divisione cellulare.
Uno dei compiti più importanti per il futuro sarà quello di analizzare i meccanismi genetici e biochimici che operano in tali casi di regolazione positiva e negativa: abbiamo già riferito della probabile esistenza dei ‛caloni', vale a dire di sostanze presenti nei tessuti adulti le quali specificamente inibiscono l'attività mitotica negli organi embrionali omologhi (per es., estratti di rene adulto inibiscono la mitosi nel rene embrionale).
Abbiamo già visto che il citoplasma di cellule in rapida divisione contiene dei fattori chimici che stimolano la sintesi del DNA nei nuclei isolati da fegato adulto, dove la divisione cellulare è eccezionale. Naturalmente, la divisione cellulare e il differenziamento cellulare non sono la stessa cosa: infatti, abbiamo visto che essi si escludono a vicenda e che una cellula che si divide è incapace di differenziamento. Tuttavia, è stato dimostrato recentemente che la formazione di cartilagine può procedere anche durante la mitosi; e inoltre, il differenziamento è, quasi sempre, preceduto da un periodo di divisione cellulare. I caloni e le sostanze che, al contrario, stimolano la divisione cellulare, potrebbero perciò avere una parte nel differenziamento cellulare, così come nella mitosi. In questo campo, l'ultima parola sarà detta dai biochimici, i quali stanno cercando di isolare, purificare e identificare le sostanze chimiche che controllano il differenziamento cellulare.
Si possono prevedere altri approcci al problema del differenziamento cellulare. Per esempio, K. W. Jeong (v. Jeong e altri, 1970) è recentemente arrivato a separare, per mezzo della micromanipolazione, il nucleo, il citoplasma (con tutti i suoi organuli) e la membrana di un'ameba, e a riunire successivamente questi tre componenti: l'ameba ricostituita era perfettamente vivente, poiché era capace di moltiplicarsi indefinitamente. È perciò possibile estrarre da una cellula il nucleo e il citoplasma, lasciando soltanto la membrana; in un secondo tempo, si può iniettare nella membrana vuota il citoplasma e il nucleo di un'altra cellula, e ottenere una cellula perfettamente vitale. Tali delicati esperimenti di microchirurgia potrebbero, teoricamente, essere eseguiti su grandi cellule differenziate, come le cellule nervose. Chi sa cosa succederebbe, per esempio, se si iniettasse dentro la membrana di una cellula nervosa il citoplasma di una cellula epatica e il nucleo di una cellula muscolare? Un tale esperimento potrebbe apparire pseudoscientifico, ma non c'è ragione di dubitare che questo diventerà possibile presto o tardi.
Un'altra possibilità futura sarà quella di introdurre nuovi geni, sia per iniezione, sia per pinocitosi, dentro le cellule: il DNA di un gene (il cosiddetto ‛lac operone') è già stato isolato in forma pura dai Batteri, mentre il DNA del gene che dirige la sintesi di uno degli RNA solubili è stato sintetizzato in laboratorio. Non c'è dubbio che un numero sempre maggiore di geni sarà isolato e sintetizzato in futuro. Il giorno in cui essi saranno usati per lo studio del differenziamento cellulare non è forse così lontano come si potrebbe pensare: per esempio, l'mRNA per l'emoglobina è stato isolato in forma pura, ed è stato possibile con questo mRNA sintetizzare enzimaticamente il gene dell'emoglobina. Se si iniettasse questo gene nel citoplasma di un uovo appena fecondato, dirigerebbe esso la sintesi dell'emoglobina e porterebbe al differenziamento di eritrociti? Ancora una volta, ciò potrebbe apparire pseudoscientifico: ma non è così, poiché abbiamo visto che Gurdon ha dimostrato che l'iniezione di mRNA dell'emoglobina in ovociti di Anfibi è seguita dalla sintesi di emoglobina.
Il problema del differenziamento cellulare rimane uno degli enigmi principali della moderna biologia: ma esso può essere e sarà risolto da esperimenti condotti con coraggio e immaginazione. (Su tutto ciò, v. embriologia).
bibliografia
Afzelius, B. A., Ultrastructure of cilia and flagella, in Handbook of molecular cytology (a cura di A. Lima-de-Faria), Amsterdam 1969, pp. 1219-1242.
Baudhuin, P., Peroxisomes (microbodies, glyxysomes), in Handbook of molecular cytology (a cura di A. Lima-de-Faria), Amsterdam 1969, pp.1179-1195.
Beermann, W., Clever, C., Chromosome puffs, in The living cell (a cura di D. Kennedy), San Francisco-London 1965, pp. 232-240.
Berendes, H. D., Beermann, W., Biochemical activity of interphase chromosomes (polytene chromosomes), in Handbook of molecular cytology (a cura di A. Lima-de-Faria), Amsterdam 1969, pp. 500-519.
Borst, P., Biochemistry and function of mitochondria, in Handbook of molecular cytology (a cura di A. Lima-de-Faria), Amsterdam 1969, pp. 914-942.
Brachet, J., Biochemical cytology, New York 1957.
Brachet, J., Nucleocytoplasmic interactions in unicellular organisms, in The cell (a cura di J. Brachet e A. E. Mirsky), vol. III, New York 1961, pp. 771-841.
Brachet, J., Embriologia molecolare, Milano 1973.
Brachet, J., Bonotto, S., Biology of Acetabularia, New York 1970.
Brachet, J., Malpoix, P., Macromolecular synthesis and nucleocytoplasmic interactions in early development, in ‟Advances in morphogenesis", 1971, X, pp. 263-326.
Brachet, J., Mirsky, A. E. (a cura di), The cell, 6 voll., New York 1959-1964.
Briggs, R., Genetic control of early development in the Mexican axolotl, in ‟Annales d'embryologie et de morphogenèse", 1969, suppl. 1, pp. 105-114.
Callan, H. G., Biochemical activities of chromosomes during the prophase of meiosis, in Handbook of molecular cytology (a cura di A. Lima-de-Faria), Amsterdam 1969, pp. 540-552.
Callan, H. G., Replication of DNA in the chromosomes of Eukaryotes, in ‟Proceedings of the Royal Society" (Series B), 1972, CLXXI, pp. 19-41.
Chantrenne, H., Isolement du RNA messager de l'hémoglobine, in ‟Biochimica et biophysica acta", 1965, CIII, pp. 409-417.
Chantrenne, H., Burny, A., Marbaix, G., The search for the messenger RNA of hemoglobin, in ‟Progress in nucleic acid research and molecular biology", 1967, VII, pp. 173-194.
Curtis, A. S. G. The cell surface. Its molecular role in morphogenesis, London 1967.
Du Praw, E. J., Cell and molecular biology, New York 1968.
Duve, C. de, The lysosome, in The living cell (a cura di D. Kennedy), San Francisco-London 1965, pp. 72-80.
Favard, P., The Golgi apparatus, in Handbook of molecular cytology (a cura di A. Lima-de-Faria), Amsterdam 1969, pp. 1130-1155.
Fawcett, D., Cilia and flagella, in The cell (a cura di J. Brachet e A. E. Mirsky), vol. II, New York 1960, pp. 217-298.
Frenster, J. H., Biochemistry and molecular biophysics of heterochromatin and euchromatin, in Handbook of molecular cytology (a cura di A. Lima-de-Faria), Amsterdam 1969, pp. 251-276.
Gall, J. G. The genetic ribosomal RNA during oogenesis, in ‟Genetics", 1969, LXI, suppl. 1, pp. 123-132.
Georgiev, G. P., Samarina, O. P., Nuclear complexes containing informational RNA, in ‟Annales d'embryologie et de morphogenèse", 1969, suppl. 1, pp. 81-87.
Goldblatt, P. J., The endoplasmic reticulum, in Handbook of molecular cytology (a cura di A. Lima-de-Faria), Amsterdam 1969, pp. 1101-1129.
Granick, S., The chloroplast. Inheritance, structure and function, in The cell (a cura di J. Brachet e A. E. Mirsky), vol. II, New York 1960, pp. 489-602.
Green, D. E., The mitochondrion, in The living cell (a cura di D. Kennedy), San Francisco-London 1965, pp. 63-71.
Gurdon, J. B., Biochemical aspects of nuclear transplantation in multicellular organisms, in ‟Annales d'embryologie et de morphogènese", 1969, suppl. 1, pp. 127-132.
Gurdon, J. B., Brown, D. D., Cytoplasmic regulation of RNA synthesis and nucleolar formation in developing embryo of Xenopus laevis, in ‟Journal of molecular biology", 1965, XII, pp. 27-35.
Gurdon, J. B., Lane, C. D., Woodland, H. R., Use of frog eggs and oocytes for the study of messenger RNA and its translation in living cells, in ‟Nature", 1971, CCXXXIII, p. 177.
Gurdon, J. B., Woodland, H. R., On the long-term control of nuclear activity during cell differentiation, in ‟Current topics on developmental biology", 1970, V, pp. 39-70.
Hämmerling, J., Nucleocytoplasmic relationships in the development of Acetabularia, in ‟International review of cytology", 1953, II, pp. 475-498.
Harris, H., Cell fusion, in The Dunham lectures, Oxford 1970.
Jacques, P. J., Lysosomes and homeostatic regulation, in Ciba foundation symposium on homeostatic regulators, London 1969, pp. 180-192.
Jeong, K. W., Lorch, I. J., Danielli, J. F., Reassembly of living cells from dissociated components, in ‟Science", 1970, CLXVII, pp. 1626-1627.
‟Journal of cell biology" (a cura di R. B. Griffiths), New York, anni 1955 ss.
‟Journal of cell science" (a cura di A. V. Grimstone e H. Harris), London, anni 1966 ss.
Kennedyt, D. (a cura di), The living cell, San Francisco-London 1965.
Kroon, A. M., DNA and RNA from mitochondria and chloroplasts (biochemistry), in Handbook of molecular cytology (a cura di A. Lima-de-Faria), Amsterdam 1969, pp. 943-971.
Lebleu, B., Marbaix, G., Huez, G., Burny, A., Temmerman, J., Chantrenne, H., Characterization of the messenger ribonucleoprotein released from reticulocyte polyribosomes by EDTA treatment, in ‟European journal of biochemistry", 1971, XIX, pp. 264-269.
Lefort-Tran, M., Cytochemistry and ultrastructure of chloroplasts, in Handbook of molecular cytology (a cura di A. Lima-de-Faria), Amsterdam 1969, pp. 1047-1076.
Lima-de-Faria, A. (a cura di), Handbook of molecular cytology, Amsterdam 1969.
Lockard, R. E., Lingrel, J. B., The synthesis of mouse hemoglobin β-chains in a rabbit reticulocyte cell-free system programmed with mouse reticulocyte 9 S RNA, in ‟Biochemical and biophysical research communications", 1969, XXXVII, pp. 204-212.
Mazia, D., Mitosis and the physiology of cell division, in The cell (a cura di J. Brachet e A. E. Mirsky), vol. III, New York 1961, pp. 77-412.
Miller, O. L., Beatty, B. R., Nucleolar structure and function, in Handbook of molecular cytology (a cura di A. Lima-de-Faria), Amsterdam 1969, pp. 605-619.
Mirsky, A. E., Osawa, S., The interphase nucleus, in The cell (a cura di J. Brachet e A. E. Mirsky), vol. II, New York 1960, pp. 677-770.
Monesi, V., DNA, RNA and protein synthesis during the mitotic cycle, in Handbook of molecular cytology (a cura di A. Lima-de-Faria), Amsterdam 1969, pp. 472-499.
Monroy, A., Chemistry and physiology of fertilization, New York 1965.
Munn, E. A., Ultrastructure of mitochondria, in Handbook of molecular cytology (a cura di A. Lima-de-Faria), Amsterdam 1969, pp. 875-913.
Novikoff, A. B., Mitochondria (chondriosomes), in The cell (a cura di J. Brachet e A. E. Mirsky), vol. II, New York 1960, pp. 299-421.
Paul, J., DNA masking in mammalian chromatin. A molecular mechanism for determination of cell type, in ‟Current topics in developmental biology", 1970, V, pp. 317-352.
Perry, R. P., Nucleoli. The cellular sites of ribosome production, in Handbook of molecular cytology (a cura di A. Lima-de-Faria), Amsterdam 1969, pp. 620-636.
Pitelka, D. R., Centriole replication, in Handbook of molecular cytology (a cura di A. Lima-de-Faria), Amsterdam 1969, pp. 1199-1218.
Porter, K. R., The ground substance. Observations from electron microscopy, in The cell (a cura di J. Brachet e A. E. Mirsky), vol. II, New York 1960, pp. 621-675.
Prescott, D. M., Goldstein, L., Protein in nuclear-cytoplasmic interactions in Amoeba proteus, in ‟Annales d'embryologie et de morphogenèse", 1969, suppl. 1, pp. 181-188.
Puiseux-Dao, S., Acetabularia and cell biology, London 1970.
Quagliarotti, G., Hedwigi, E., Wikman, J., Busch, H., Structural analysis of nucleolar precursors of ribosomal ribonucleic acid, in ‟Journal of biological chemistry", 1970, CCXLV, pp. 1962-1969.
Ris, H., The molecular organization of chromosomes, in Handbook of molecular cytology (a cura di A. Lima-de-Faria), Amsterdam 1969, pp. 221-250.
Robertson, J. D., The membrane of the living cell, in The living cell (a cura di D. Kennedy), San Francisco-London 1965, pp. 45-52.
Robertson, J. D., Molecular structure of biological membranes, in Handbook of molecular cytology (a cura di A. Lima-de-Faria), Amsterdam 1969, pp. 1403-1443.
Scarano, E., Enzymatic modifications of DNA and embryonic differentiation, in ‟Annales d'embryologie et de morphogenèse", 1969, suppl. 1, pp. 51-62.
Stevens, B. J., André, J., The nuclear envelope, in Handbook of molecular cytology (a cura di A. Lima-de-Faria), Amsterdam 1969, pp. 837-871.
Wattiaux, R., Biochemistry and function of lysosomes, in Handbook of molecular cytology (a cura di A. Lima-de-Faria), Amsterdam 1969, pp. 1159-1178.
Wilson, E. B., The cell in development and heredity, New York 1925.
Patologia della cellula
di Massimo Aloisi
SOMMARIO: 1. Patologia della cellula e ‛patologia cellulare'. 2. Le apparenze strutturali elementari e le interpretazioni biochimiche della patologia spontanea: a) considerazioni generali; b) eziologia e patogenesi generali dei vari aspetti della patologia cellulare; c) i quadri relativamente specifici della patologia cellulare spontanea. 3. La riproduzione sperimentale analitica di vari aspetti della patologia della cellula. 4. La rigenerazione. □ Bibliografia.
1. Patologia della cellula e ‛patologia cellulare'
Nell'articolo di J. Brachet (v. cellula: Fisiologia della cellula) sono prese in esame la struttura, la fisiologia e la biochimica della ‛cellula' secondo l'accezione generale e fondamentale del termine e, pur dovendo necessariamente portare esempi concreti di determinate cellule, l'argomento è svolto in modo da considerare quelle strutture-funzioni che sono più generalmente comuni a tutte le cellule, i Batteri o Procarioti compresi.
Una trattazione della ‛patologia della cellula' rigidamente compresa entro gli stessi limiti, tuttavia, risulterebbe, almeno oggi, estremamente semplice, monotona: essa non andrebbe oltre le osservazioni e i dati piuttosto superficiali che un qualunque esperto di colture di tessuti o, per esempio, di morfologia cancerologica, ha continuamente a portata di mano.
Del resto, nel descrivere la natura e l'operatività delle strutture cellulari diviene inevitabile - e talora illuminante per la futura sperimentazione - considerare i casi, come già fa Brachet, in cui quella operatività venga a cessare o a essere deviata, trasformata: se lo credessimo opportuno, potrebbe convenirsi di chiamare quella una patologia elementare e sperimentale della cellula, integrando la descrizione delle suddette modificazioni primitive con quella di tutte le loro conseguenze finali, coincidano esse con il completo recupero della normale operatività della cellula (o della sua parte in oggetto), oppure con un permanente difetto o con la morte della cellula.
Ma all'infuori della osservazione, che è canonica per ogni studioso di patologia generale, secondo cui l'evento patologico non è che l'altro versante dialettico del corrispondente evento fisiologico o comunque questo stesso evento in condizioni soltanto quantitativamente diverse dalla norma statistica, quella siffatta patologia generale e sperimentale della cellula è in realtà ancora quasi tutta da indagare, o, nella misura in cui è nota, coincide in buona parte con l'intervento sperimentale stesso rivolto allo studio delle caratteristiche fisiologiche. È questa un'epoca in cui si possono trovare o preparare cellule isolate prive di mitocondri, o si possono asportare e trapiantare nuclei, organuli cellulari, fondere citoplasmi, inibire o comunque influenzare specificamente, e in modo precisamente noto, le attività DNA- e RNA-sintetica in tutte le loro specificazioni, o l'operatività dei mitocondri o le funzioni di membrana; e persino trasportare le membrane, eccitare o inibire la formazione dei lisosomi, l'autofagia, ecc. È evidente che questi studi di fisiologia cellulare rappresentano classicamente la descrizione positiva della patologia provocata (intesa come alterazione della struttura-funzione). Sembra lecito ritenere che appunto questa potrà essere la fisiopatologia generale della cellula nel prossimo futuro, l'approccio e il canone interpretativo più rigorosi per comprendere anche gli eventi patologici spontanei e non solo sperimentali a livello cellulare, e appare ovvia l'ineluttabilità di tale conclusione.
Tuttavia, rebus sic stantibus, occorre anche fare altre considerazioni. La patologia spontanea o sperimentale dei viventi, anche se indirizzata secondo uno studio cellulare e molecolare, non può limitarsi a modelli relativamente semplici come organismi monocellulari (o non cellulari), per i quali del resto si ripetono le stesse monotone prospettive di decadimento morfo-funzionale già prima lamentate; qui appare altrettanto ovvio che nei riguardi di questo tipo di viventi prevarrà di gran lunga l'interesse per l'intervento sperimentale o genetico al fine di comprendere il significato positivo, cioè funzionale e generale, di determinate strutture più o meno ubiquitarie, ossia presenti in quasi tutti i viventi. Con le dovute eccezioni, naturalmente: il parassitismo ne offre una vistosa, quale è quella, per esempio, della patologia batterica in rapporto al suo parassita, il fago.
Ma l'altra considerazione è che quando si rivolga l'attenzione ai viventi multicellulari (o cellulari), per l'inerente fenomeno della differenziazione, non è più tanto lecito (se non in sede paradigmatica e di base) parlare di cellule generiche, poiché in essi esistono solo cellule specifiche, e questa loro specificità (differenziazione) largamente influenza il loro modo di essere patologico. Per una patologia concreta, che cioè finisca in qualche modo, pur rimanendo generale, col dare informazioni integrabili nei processi morbosi manifestati dall'organismo, le cellule non possono concepirsi isolate l'una dall'altra, bensì, come esse in realtà sono nei vari tessuti, associate in unità sovracellulari, immerse in un ‛mezzo interno' che le condiziona e che esse stesse condizionano, sottoposte a molteplici sistemi di correlazione.
Infine, l'ultima considerazione è quella che le ‛cause' degli eventi patologici (fattori eziologici) e il loro modo di agire sono, nella patologia spontanea degli organismi, complesse o divengono spesso, anche durante quella sperimentale, molteplici e intricate, onde al patologo rimane spesso l'obbligo del registrarne gli effetti (che appariranno come epifenomeni ricchi e svariati) senza poter sempre risalire con sicurezza e con rigore lungo tutta la loro catena patogenetica. Si dica subito che i patologi, tuttavia, non hanno aspettato (e ben hanno fatto) né aspettano che la biologia molecolare completi (se possibile) il suo iter conoscitivo per fondare e perfezionare la loro ricerca e le loro dottrine: l'integrabilità tra i diversi approcci del conoscere scientifico è fattibile periodicamente, dalla biologia molecolare alla psicologia, volta a volta che determinati ponti conoscitivi vengano consolidati.
Alcuni degli ‛aspetti' patologici riscontrabili nelle cellule degli organismi complessi possono osservarsi nelle cellule isolate, in coltura, o negli organismi non cellulari: per esempio, un eccesso di contenuto lipidico, o le più note alterazioni nucleari, o la gelificazione o fluidificazione del citoplasma o la sua vacuolizzazione; ma è assai dubbio che qui essi abbiano lo stesso significato eziopatogenetico. Ed è plausibile e vera anche la situazione reciproca. Né va dimenticato che la patologia generale è nata come disciplina medica.
Con ciò abbiamo fornito la ragione per cui qui, per tutto un settore, si parlerà, sia pur criticamente, di ‛patologia cellulare', piuttosto che di ‛patologia della cellula'; non è questo un giuoco di parole, bensì una precisazione dipendente dalla necessità che ancora oggi abbiamo di rimanere nel termine storico del corrispondente capitolo della ‛patologia generale' delle malattie spontanee. Tale richiamo storico è d'altronde interessante anche per una considerazione della evoluzione dei concetti di organismo e di affezione morbosa.
Con la ‛teoria cellulare' di Theodor Schwann (1839) si affermò l'importante acquisizione che gli organismi viventi sono costituiti da una o più unità discrete che furono chiamate cellule, nella quale acquisizione era implicito che tutte le manifestazioni fenomeniche degli organismi non fossero che la somma (per dirla con un termine che anche per allora poteva sembrare semplicistico, a parte gli esempi che già si avevano delle ‛colonie' di cellule) delle attività fisiologiche delle singole cellule. Era quindi inevitabile che tale concezione investisse il campo della patologia, tanto più che già dal passato veniva un solenne imperativo da G. B. Morgagni, quello di ricercare localisticamente gli effetti e quindi le cause dei morbi.
Fu Rudolf Virchow che, circa venti anni dopo la formulazione della teoria cellulare, trasse la conclusione che sembrò più importante per la patologia e la medicina, quella cioè che la malattia organismica poteva risolversi nella patologia delle sue cellule, volta a volta variamente distribuita e variamente estesa. Così egli fondò Die Cellularpathologie in ihrer Begründung auf physiologische und pathologische Gewebelehre (Berlino, 1858).
Naturalmente, come oggi non riteniamo più che un organismo sia meramente la somma delle sue cellule e delle sue sostanze intercellulari, ma piuttosto una complessa unità derivante dal reciproco coordinamento dell'attività di tutte le cellule attraverso i sistemi di correlazione umorali e nervosi (alla cui base stanno pure, naturalmente, delle cellule), così anche i fenomeni patologici non possono più essere considerati come il semplice giustapporsi di alterazioni patologiche elementari delle singole cellule. La patologia cellulare analitica virchowiana è stata convenientemente e felicemente filtrata attraverso la trama ‛sintetizzante' di Claude Bernard.
È anche vero che, con la scoperta delle funzioni specifiche degli organuli endocellulari, straordinariamente favorita dall'avvento della microscopia elettronica, si è di nuovo affacciata la possibilità di spiegare alcuni aspetti della patologia organismica in termini di patologia di queste parti costitutive della cellula (le quali peraltro non sono affatto, o non sempre, eguali nelle varie cellule); così procedendo, seguendo l'analisi strutturale-biochimica, tuttora in corso, di queste parti costitutive, si è passati a ipotizzare l'esaustivo passaggio della patologia cellulare (o mitocondriale, o nucleare, o delle membrane, ecc.) a una più basilare e fondamentale ‛patologia molecolare'. Un tale studio, molto fecondo e istruttivo per molte o tutte le malattie dette ereditarie, perché appunto condizionate da un'alterazione molecolare del genoma traducentesi in un'alterazione molecolare del soma (per es., difetti enzimatici ubiquitari o cellulospecifici), non è altrettanto facilmente fattibile o risolvibile, almeno per ora, per una gran parte della patologia acquisita, ove le determinanti in gioco sono moltissime, quantitativamente graduate, e ove per ciò stesso prevale sull'alterazione fissa e molecolare (la molecola sbagliata) piuttosto quella dei sistemi di controllo autoregolativi, ossia della cibernetica cellulare e organismica (quando poi non si tratti, diametralmente, dell'ingresso endocellulare di un parassita, batterio, protozoo, virus, ecc.).
Ma per tutta questa dialettica circa i vari livelli di studio della patologia cellulare e per l'evolversi storico di essa, dal Virchow in poi, si rimanda all'articolo patologia sperimentale.
Qui occorrerà passare in rassegna le classiche categorie della patologia cellulare, con l'aggiornamento dovuto in senso interpretativo e sperimentale allo sviluppo della ricerca, ivi compresa, là dove è possibile, la biologia cellulare molecolare.
2. Le apparenze strutturali elementari e le interpretazioni biochimiche della patologia spontanea
a) Considerazioni generali
È tuttora ricca di significato interpretativo la persuasione che non esistono malattie senza corrispondenti alterazioni cellulari, siano o non siano queste facilmente riconoscibili dalla struttura, a seconda dei mezzi di indagine che abbiamo a disposizione. Parlare di alterazioni puramente funzionali in realtà è oggi quasi divenuto un non senso, poiché a un livello fondamentale non esistono funzioni che si sovrappongono alle strutture, né strutture che non abbiano funzione: si può dire anzi che la fisiologia è la struttura nel tempo.
Ma, mentre in alcuni casi l'amplificazione fisiologica di una alterazione primitiva, quale si può verificare al livello organismico (si pensi, per es., a un'alterazione neuronica), è più facilmente descrivibile e interpretabile appunto in termini di fisiopatologia organismica che non in termini di patologia cellulare o subcellulare o molecolare (ciò però dipende naturalmente ‛anche', ma non in tutto, dalle contingenti possibilità di analisi), all'opposto si hanno lesioni cellulari senza che si possa parlare di stato morboso o di malattia dell'organismo, senza cioè che l'economia fisiologica integrata dell'organismo ne sia affetta per superamento dei meccanismi autoregolativi di compenso.
Una lesione o noxa non è descrivibile di per sé come patologia: una ‛fissazione' idealmente perfetta di un tessuto e delle sue cellule (con cui sono immediatamente uccise) non è patologia; patologia è la sequenza nel tempo delle reazioni cellulari, tessutali e organismiche a quella lesione o noxa, se essa è di natura tale da permetterle.
Possiamo anzi dire che lesioni cellulari (nel semplice e comune senso di spostamento reversibile degli equilibri dinamici morfo-funzionali) si determinano continuamente nel nostro organismo per i necessari contatti con l'ambiente e per tutta la fisiologia organismica, senza che esse arrivino quantitativamente a manifestarsi come riconoscibile fatto morboso, anzi senza essere un evento morboso. Come si vede, la definizione di questo concetto richiederebbe molto senso di relatività.
Del resto, molte cellule e i fluidi e le sostanze intercellulari sono, in un organismo, in un variabile equilibrio dinamico anche riguardo al loro numero e alla loro entità totale, per cui si ricambiano continuamente e in modo vario a seconda della loro diversa natura e posizione; e alcune cellule vanno continuamente incontro alla regressione e alla morte per essere sostituite da nuovi elementi della stessa specie, mentre altre subiscono una senescenza che, più o meno, coincide con la senescenza dell'intero organismo.
Queste considerazioni rendono plausibile che in uno studio della patologia cellulare si incontreranno molto spesso fenomeni che in sé non differiscono da quelli osservabili già nel corso della vita fisiologica di vari tipi di cellule o tessuti. Difficilmente infatti la fenomenologia patologica - persino nel parassitismo - sarà qualcosa di completamente nuovo, non avente cioè alcun riscontro nella fenomenologia morfo-funzionale (e cioè anche biochimica) che regola la vita fisiologica. Spesso, anzi, i fatti patologici elementari, pur in apparenza qualitativamente diversi, non sono che deviazioni quantitative di alcuni settori della fisiologia cellulare, oppure manifestazioni che si chiamano patologiche solo perché intervengono fuori tempo o fuori luogo rispetto allo svolgersi normale della vita di un organismo. Naturalmente esistono anche quadri apparentemente del tutto inusitati, per esempio quelli che dipendono da cause esterne per lo più violente e specifiche, la cui azione può essere essenzialmente nuova e comunque tale da soverchiare ogni possibilità di adattamento e di reazione biologico-omeostatica da parte dell'organismo: azioni fisiche e chimiche massimali e letali per le cellule o per l'organismo (ove esiste allora ‛lesione', ma non ‛malattia'), invasione delle cellule da parte di un parassita (ove la presenza di questo può provocare la comparsa di reazioni e di strutture apparentemente nuove).
Ma anche in questi casi, in parte per la grossolanità e superficialità tuttora esistenti nel nostro armamentario fisico-biochimico di indagine, in parte per il fondamentale limite nella qualità di risposta che una cellula animale o vegetale può dare, inerente alla sua stessa natura generica, si assisterà a una certa monotonia nelle risposte nei confronti della molteplicità delle cause di disturbo: non del tutto simile, però, a quella cui si è accennato, riscontrabile negli organismi unicellulari o nelle cellule isolate.
Gli spostamenti degli equilibri dinamici cellulari, che oltre una certa misura (statisticamente valutabile) rientrano nell'ambito delle deviazioni patologiche, sono molto spesso reversibili. Ma vi è una reversibilità al livello cellulare e un'altra al livello tessutale, nel senso che cellule irreversibilmente colpite e quindi necessariamente morenti possono esser sostituite da altre nel tessuto (come avviene del resto di norma nell'equilibrio numerico-cellulare di un tessuto). Altre volte non vi è irreversibilità e morte, ma nemmeno recupero completo (o restitutio ad integrum), bensì raggiungimento di un diverso e spesso meno efficiente punto di equilibrio dinamico (compenso parziale). Come si vedrà, nel caso di una singola cellula isolata e indifesa è molto facile che una deviazione morfo-funzionale raggiunga il punto della irreversibilità, anzi il processo patologico può assumere l'andamento della retroazione positiva (‛circolo vizioso') con risultati cataliticamente ingravescenti fino alla morte.
I fenomeni patologici elementari, visti come espressione di uno spostamento quantitativo abnorme di uno o più aspetti dei complessi equilibri dinamici della fisiologia cellulare, vengono schematicamente suddivisi in due categone: fenomeni a carattere regressivo, che più direttamente rappresentano il danno subito dalle cellule o dal mezzo intercellulare nel quale vivono, onde ne deriva una minorazione specifica o generica dell'attività fisiologica; e fenomeni a carattere progressivo, che rappresentano il compenso, la riparazione, il ritorno all'equilibrio dinamico primitivo, o a un nuovo livello di equilibrio, molto spesso raggiungibile, espressione della ‛adattabilità' della cellula. Come ben si comprende, non è tuttavia sempre facile distinguere questi due momenti in un complesso fenomeno patologico, ove possono essere strettamente correlati e commisti.
La limitazione alla varietà delle alterazioni morfologiche quali furono dapprima elencate dai patologi di discendenza virchowiana, nonostante la notevole varietà delle noxae che le provocavano (e le provocano), poteva essere (ed è) legata alla relativa inespressività del dato puramente morfologico: anche la microscopia elettronica, che oggi impone una revisione (non sempre remunerativa) di tutta la classica patologia cellulare, trova difficoltà a chiarire differenze che pur sussistono notevolissime nell'ambito delle cause (della eziologia). Anzi, molto spesso la microscopia ottica, che permette di cogliere in sintesi un insieme differenziato di deviazioni strutturali e anche i rapporti di gruppi di cellule fra loro, è assai più informativa e discriminante della microscopia elettronica. Di qui tutto l'indirizzo moderno della patologia cellulare ricostruita in termini biochimici, che sembra l'unico modo per uscire dalle semplici classificazioni utili soltanto ai fini pratici, diagnostici.
Ma va subito detto che anche al livello biochimico la patologia può apparire relativamente monotona se non si riesce a descrivere altro che gli eventi terminali di un processo, i quali d'altronde sono i più facili da osservare e i più appariscenti per un'avvenuta amplificazione consequenziale, come è nel caso della irreversibilità e del feedback positivo. In effetti è constatazione fondamentale che a un certo momento della disfunzionalità della cellula, specie quando si arrivi - e con relativa facilità vi si arriva - a menomare i meccanismi della produzione e della conservazione della energia (vedi produzione, riserva e dispendio di ATP), non solo la fisiologia della cellula è per ciò stesso gravemente compromessa, ma ne sono compromessi le stesse compagini strutturali, la membrana, i mitocondri, il reticolo endoplasmatico e gli altri sistemi canalicolari-membranosi, la cui integrità dipende dal rifornimento energetico, i lisosomi e infine anche il nucleo.
La lettura dell'articolo di J. Brachet (v. cellula: Fisiologia della cellula) confermerà chiaramente questa dipendenza costante del modo di essere della cellula e delle sue strutture dalla esigenza fondamentale della disponibilità energetica nelle forme dovute, di cui quella che passa per la sintesi o scissione dell'ATP è una delle principali.
È così che i risultati finali ‛leggibili' di un processo patologico saranno vari a seconda che la noxa colpisca un processo metabolico nelle sue determinazioni a monte, oppure intervenga direttamente nelle parti terminali dei gradienti metabolici cellulari e specie là dove i tre metabolismi principali (glicidico, lipidico e protidico) largamente confluiscono. Nel primo caso, e nonostante la naturale circolarità del metabolismo, avremo una varietà maggiore di situazioni patologiche: il caso paradigmatico è quello del difetto genico di determinati settori del metabolismo, ove la malattia è inizialmente definibile come monomolecolare e come tale può presentare in linea di principio altrettanta e altrettale variabilità quanto la costituzione genomica lo permette, in relazione anche alla differenziazione delle cellule nei tessuti e organi. Nel secondo caso, invece, per esempio quando si tratti di azioni nocive massimali e senza dialettica reattività da parte delle cellule (azioni direttamente mortifere, necrotizzanti), o quando l'evento deviante o patologico si inserisca comunque nelle fasi confluenti ed energeticamente condizionanti del metabolismo, avremo un risultato che, tradotto in termini metabolico-morfologici, non differirà molto da caso a caso, se non per quel tanto per cui i tessuti (cioè le varie cellule differenziate e specializzate) differiscono nella intensità della richiesta energetica. Naturalmente alcune differenze, cospicue sul piano descrittivo, ma talora secondarie in termini patogenetici e generali, dipenderanno dalla presenza, dalla qualità e dalla quantità nelle cellule e nelle sostanze intercellulari dei prodotti della differenziazione: una patologia cellulare generale delle fibre muscolari non può prescindere dal fatto che in esse qualunque primaria deviazione del metabolismo influisce sulla produzione, sulla conservazione e sul modo di essere e di reagire fisico-chimico della notevolissima massa di proteine specifiche, miosina, actina e altre ivi presenti, senza dire che la particolare architettura di queste cellule condizionerà anche l'aspetto puramente morfologico dell'avvenuta o intervenente deviazione patologica. Così si può immaginare sarà per una cellula nervosa (neurorete, sostanza cromofila) o per una cellula ematica (capacità di trattenere l'emoglobina) o per una cellula ghiandolare (difficoltà di formazione o di espulsione del secreto), ecc. Ma, reciprocamente, potranno darsi situazioni patologiche che primariamente consistono in deviazioni chimico-strutturali o della velocità del ricambio proprio di questi prodotti della differenziazione (in genere sono comprensibilmente alterazioni genetiche), onde avremo modificazioni primarie delle proteine e dell'apparato contrattile, del metabolismo specifico della cellula nervosa, emogiobine patologiche, inadeguatezza di determinate secrezioni, con riflesso secondario su tutta la vita cellulare.
b) Eziologia e patogenesi generali dei vari aspetti della patologia cellulare
I fenomeni che rientrano classicamente nello studio della patologia cellulare, progressivi e regressivi, necessariamente riguardano i vari aspetti della vita fisiologica delle cellule e quindi tutti gli equilibri dinamici relativi, endocellulari e dei rapporti tra le cellule e il loro ambiente, il che vuol dire anche tra cellule e cellule o fra le cellule e le sostanze intercellulari (quali ad esempio la sostanza fibrillare collagene e la sostanza fondamentale, mucopolisaccaridica, del connettivo nei Vertebrati; la cuticola degli Insetti: la cellulosa dei Vegetali, ecc.).
Per aver presente, in linea schematica e del tutto provvisoria, l'interdipendenza delle varie funzioni cellulari e quindi le varie risultanze delle deviazioni patologiche a seconda del punto iniziale di attacco della noxa ci serviremo del diagramma della fig. 1, che schematizza le operazioni comuni a quasi tutte le cellule di un organismo. Lo schema è dunque necessariamente generico e la sua lettura quindi introduttiva. Vi sono anche accennati i rapporti correlazionali della cellula con il resto dell'organismo: sistema di correlazione chimico, a breve (per contatto, vedi chaloni e simili) o a lungo raggio di azione (ormoni); sistema di correlazione nervoso (con i relativi mediatori chimici). I blocchi non colorati rappresentano le operazioni biochimiche e biofisiche generali; i blocchi colorati in rosso le strutture a cui quelle operazioni sono associate. Le connessioni rappresentano circuiti semplificati di interdipendenza funzionale che presuppongono tutte le relative retroazioni (o feedbacks); in rosso sono le connessioni provenienti dall'esterno (ormoni e impulso nervoso).
I tre settori dello schema, A, B e C, operano ovviamente in stretta connessione sinergica, ma si può schematicamente dire che in A (di cui va considerata tutta l'estensione endoplasmatica delle cavità del reticolo) si svolgono i processi fondamentali dei rapporti della cellula col mondo esterno (e quindi anche quelli ‛nutritivi') e di quelli dell'interno di molte substrutture intracellulari col restante citoplasma (succo cellulare) in cui sono incluse: sono i processi per cui si creano e si mantengono i gradienti energetici, fonte prima di tutto il funzionamento cellulare (e in ciò la massima importanza spetta ai mitocondri), si controllano la segregazione e il flusso ionico e acquoso, quali espressioni della integrità funzionale della cellula e del suo potenziale di membrana, quindi della sua eccitabilità o attivabilità; e si realizzano alcune attività periodiche quali la secrezione, l'impulso nervoso e la contrazione (a seconda dei tipi cellulari).
Il settore B è quello incentrato nel metabolismo più o meno specifico della cellula, che in buona misura diversifica a seconda della differenziazione cellulare; è il settore che include, oltre al succo cellulare vero e proprio, buona parte dell'apparato canalicolare (reticolo endoplasmatico, apparato di Golgi, ecc., i microcorpi, e altri peroxisomi) e ove compaiono gli adattamenti enzimafici della cellula in dipendenza di situazioni variate, anche esterne; è anche il settore del catabolismo cellulare, della demolizione delle strutture invecchiate o alterate (autofagia). Il settore C è quello dei meccanismi riproduttori, cioè sia della completa moltiplicazione cellulare, quando perduri come fenomeno fisiologico, sia del rinnovo delle strutture e in genere della continua resintesi proteica (e quindi enzimatica) attraverso l'apparato nucleare e ribosomiale.
Tutto ciò ricordato, possiamo comprendere la seguente schematica e provvisoria classificazione dei fenomeni elementari della patologia cellulare: alterazioni dei processi del traffico con l'esterno, comprendenti processi disnutritivi come ipo- e ipertrofia, inclusioni o infiltrazioni esogene, fagocitosi, endocitosi e anomalie della secrezione; alterazioni dei processi moltiplicativi, come ipo- e iperplasia, rigenerazione, deviazione e trasformazione tumorale; alterazioni dell'attività nucleo-citoplasmatica (DNA-RNA) non necessariamente in fase moltiplicativa, come alterazioni della differenziazione, suo arresto o ipersviluppo e specificazione, attivazione o depressione della competenza immunitaria (che però è legata anche ai fenomeni moltiplicativi cellulari); alterazioni delle condizioni di lavoro della macchina metabolica e della produzione e conservazione dell'energia, ossia di tutto il lavoro del sistema enzimatico variamente specifico, consistenti in degenerazioni o distrofie, inclusioni o infiltrazioni endogene, tesaurismosi, pigmentazioni, ecc., necrobiosi e autolisi. Aggiungiamo l'invasione parassitica e, ancora, un fenomeno che qui sarà trattato solo per accenni in quanto è realizzato da un concorso di vari fattori cellulari e non cellulari, cioè quello della mobilitazione difensiva, quale si verifica nella flogosi o infiammazione (processo chiaramente sovra- e intercellulare). Ovviamente tutti questi settori di attività, e pertanto di deviazione patologica, sono o possono essere tra loro strettamente interdipendenti e comparire insieme o sequenzialmente: così possiamo avere atrofia semplice e atrofia degenerativa o distrofia; ipoplasia, come ora vedremo, associata a ipotrofia, ecc. Tali processi devianti dal suo livello normale l'omeostasi cellulare, consistenti anzi essenzialmente in questa deviazione, possono essere reversibili - e sarà una questione di grado - ma possono essere anche irreversibili: cioè la deviazione può giungere a un punto tale da non poter più consentire un ritorno alle condizioni iniziali, in quanto a quel punto si inaugura una serie di processi consequenziali in cui sono largamente implicate la formazione, la funzione e la conservazione dei lisosomi, che terminano con la denaturazione globale del citoplasma e delle sue strutture (necrobiosi, necrosi). Naturalmente, a seconda della natura del tessuto, alla irreversibilità dei processi che conducono alla necrobiosi cellulare può contrapporsi il compenso sostitutivo per rigenerazione.
I termini della patologia cellulare or ora introdotti, per quanto detto sopra, non indicano in realtà soltanto fenomeni patologici intesi come inerenti solamente alle situazioni morbose: atrofie e ipertrofie, ipo- e iperplasie, deviazioni unilaterali o pauci-laterali del metabolismo cellulare fino alla scomparsa dalla cellula del prodotto stesso del suo metabolismo, fenomeni necrobiotici, attività rigenerativa, sono tutti aspetti che vengono esemplificati nella vita normale dei tessuti, e di ciò non mancheremo di dare sufficienti esempi, poiché l'orientamento concettuale che ne deriva è fondamentale in patologia.
c) I quadri relativamente specifici della patologia cellulare spontanea
1. Atrofie, ipotrofie, ipertrofie, ipo- e iperplasie. - Se l'insieme dell'apparato cellulare (v. fig. 1, A, B e C) è generalmente e genericamente ipofunzionante, per esempio per carenza non grave di apporto nutritivo, o per deficiente stimolazione funzionale ormonica o nervosa, o per abbassamento del regime termico nei poichilotermi entro certi limiti di compatibilità, si avrà un caso di ipotrofia con diminuzione volumetrica delle cellule (e quindi del tessuto) senza una deviazione se non quantitativa del metabolismo; al massimo grado si potrà parlare di atrofia, ma allora è difficile che non siano compromessi in senso alterativo determinati settori della vita cellulare e non vi siano quindi note degenerative, regressive, per attivazione lisosomiale, culminanti nella scomparsa autolitica della cellula (vedi necrosi e necrobiosi).
Se però si tratta di una cellula che normalmente mantiene attivo il suo apparato di duplicazione (cellule labili, secondo Bizzozero), uno stato di ipotrofia non può non tradursi, al livello tessutale, in una diminuzione anche numerica delle cellule (atrofia numerica o ipoplasia). All'opposto, un'attivazione particolarmente o abnormemente accentuata del complesso apparato (eccesso di ormoni o di stimolazione funzionale nervosa, per le correlazioni a lungo raggio, attivazione o inibizione dei sistemi chalonici, per le correlazioni a breve raggio, apporto cospicuo di nutrienti) porterà a un aumento volumetrico della cellula e del tessuto per ipertrofia e si tradurrà invece, addirittura, in iperplasia se tra le funzioni delle cellule nel tessuto considerato vi è normalmente anche quella di una moltiplicazione periodica.
Naturalmente in questi casi di ipo- come di iperpiasia la noxa agisce in modo da raggiungere direttamente o indirettamente il settore C dello schema della nostra cellula tipo: nel corso dell'organogenesi può essere pressoché del tutto annullata la responsività di questo settore, e si ha allora il caso della aplasia (mancanza congenita di un tessuto od organo).
Senza arrivare a questi estremi è possibile una serie di deviazioni quantitativo-qualitative dell'apparato proteicocostruttore della cellula, indipendentemente dalla sua capacità moltiplicativa, e quindi il realizzarsi di difetti o anomalie della differenziazione cellulare. Ma alterazioni della differenziazione possono aversi anche nel corso delle atrofie semplici, in quanto un abbassamento di tutti i processi cellulari nella identica misura quale è postulato a base di questo tipo di atrofie è in definitiva un'astrazione, sia perché è facile il trapasso col tempo in una fenomenologia qualitativamente differente (alterazioni quantitative e qualitative del metabolismo, atrofie degenerative), sia, e soprattutto, perché può avvenire che le zone più precocemente e più palesemente colpite in un simile stato di cose, per una cellula altamente differenziata, siano appunto quelle onde nasce e si conserva la differenziazione e si organizzano i suoi prodotti: per esempio le miofibrille delle fibre muscolari, le fibrille collagene, i secreti. Si verifica allora un vero stato di ipoplasia differenziativa (talora ingravescente) al livello subcellulare. All'opposto, una semplice ipertrofia al livello cellulare può essere l'espressione di una iperplasia delle strutture specifiche al livello subcellulare (per es. nella ipertrofia delle fibre muscolari aumentano numericamente le miofibrille e i miofilamenti, ecc.) (v. fig. 2).
Ma si deve anche dire, per tener presente quanto più è possibile l'omeostasi strutturale-funzionale sempre in atto entro la cellula, la quale anche si realizza per spostamenti alternativi dei processi sintetici e differenziativi, che in tutti questi processi sono coinvolte anche le strutture più generiche delle cellule, quali i mitocondri, le membrane del reticolo endoplasmatico liscio e rugoso, l'apparato di Golgi, i lisosomi, i microcorpi, i microtubuli, ecc.; tutte queste strutture endocellulari variano numericamente nelle circostanze suddette del ‛trofismo' cellulare, e anche in senso inverso all'accadimento generale al livello cellulare. Così in una fibra muscolare atrofica possono riscontrarsi, accanto alla regressione ipoplastica del prodotto differenziato, il materiale contrattile, iperplasie anche cospicue (documentate dall'andamento della sintesi) del sistema delle membrane del reticolo endoplasmatico, o dei mitocondri, ecc. (v. Muscatello e altri, 1965; v. figg. 3 e 4). Esempi fisiologici di ipotrofie, ipoplasie e aplasia possono essere quelli inerenti allo sviluppo ontologico per cui sia durante la vita embrionale sia successivamente alcuni organi o abbozzi di organi, divenendo funzionalmente via via meno importanti per i cangiamenti della correlazione fisiologica, subiscono una progressiva e più o meno rapida involuzione, che è naturalmente insieme ipotrofica e ipoplastica, fino alla loro definitiva scomparsa o, come più spesso accade, alla loro sostituzione con tessuto adiposo, ecc. A tale proposito, pur prescindendo dai processi molto cospicui e drammatici che coinvolgono anche fenomeni estesi di degenerazione e di necrobiosi quali quelli che stanno alla base delle metamorfosi degli Anfibi, degli Insetti e di altri animali o delle involuzioni periodiche in molte cellule dei Vegetali, si consideri ad esempio l'atrofia degli abbozzi branchiali e della corda dorsale, di alcune parti dell'albero circolatorio nell'ontogenesi intrauterina dei Mammiferi, degli organi della sfera sessuale (mammella compresa) a una certa età della vita extrauterina, del timo nell'adolescenza, l'ipotrofia del midollo osseo emopoietico, dei muscoli e di alcune strutture ossee e cutanee nella senescenza, ecc.
Alla base di questi fenomeni di involuzione atrofica stanno non solo fatti di riduzione nutritiva per contemporanea involuzione circolatoria, bensì anche modificazioni nelle correlazioni ormoniche o dell'attività nervosa. Spesso si tratta anche di una sorta di circolo vizioso, nel senso che ogni causa che diminuisca l'uso, l'esercizio funzionale di un organo, anche indipendentemente dal deficit nutritivo (o in aggiunta a questo), finisce per provocare atrofia, così come è naturalmente vero anche l'opposto. Nella donna, anche durante il periodo della maturità sessuale, le cellule della mucosa uterina, del miometrio, della mammella subiscono periodici processi di ipertrofia-iperplasia alternati ad altri di ritorno all'equilibrio dinamico preesistente, in coincidenza dei cicli ipofisari.
Si è visto anzi di recente che l'involuzione postpuerperale delle cellule degli adenomeri della mammella non è classificabile come semplice riduzione del trofismo, bensì è sostenuta anche (ma, come si è detto, ciò è constatabile anche altrove, quando l'entità del processo superi certi limiti) da fenomeni complessi di degenerazione citoplasmatica con attivazione di lisosomi, da comparsa di citosegresomi (autofagia) e da fagocitosi di residui esterni di altri citoplasmi (eterofagia) (v. Helminen e Ericsson, 1968).
Nel settore delle atrofie cellulari patologiche e distrettuali riconosciamo molti fattori eziopatogenetici: a) una diminuzione o alterazione della microcircolazione (ischemia non grave o iperemia passiva); b) la compressione (che però spesso anche si traduce in diminuzione della circolazione); c) certe avitaminosi; d) il non uso o comunque l'assenza dello stimolo specifico all'uso (atrofia muscolare da immobilizzazione, da tenotomia, da denervazione); e) talora, all'opposto, l'eccessivo impegno funzionale di un organo (per es. di una ghiandola, del tessuto emopoietico, del sistema cellulare immunopoietico). Si hanno atrofie sistemiche di certe mucose per un'autoaggressione immunitaria blanda ma continuata (vedi, per la mucosa del sistema digestivo e gastrica in particolare, la conseguente anemia perniciosa). Per quanto riguarda l'atrofia per mancata o inefficiente azione neurotrofica per molte cellule, ma in particolare per quelle muscolari, si discute molto ancora se e quanto essa si differenzi da una semplice atrofia da non uso: tutta la discussione è imperniata sul dubbio se, oltre allo stimolo attivante la specifica funzione (per es. le scariche acetilcoliniche dalle terminazioni nervose colinergiche), provengano dal nervo altri fattori necessari per l'omeostasi della cellula innervata.
È un'osservazione comune che le fibre muscolari possono svilupparsi, crescere e differenziarsi in coltura (cioè produrre tutto l'apparato delle proteine contrattili, morfologicamente e fisiologicamente normale), ma solo fino a un certo momento non ben precisabile morfo-fisiologicamente, oltre il quale quelle fibre vanno incontro a un progressivo processo di atrofia: la meccanica distensione e distensibilità appaiono del resto, per le cellule muscolari e per il tessuto fibrillare collageno, un elemento molto importante per il mantenimento della omeostasi.
Nelle fibre muscolari cardiache l'ipotrofia non è mai, ovviamente, una ipotrofia ‛da non uso', ma interviene di regola nella avanzata senescenza; in questo caso si trovano anche note degenerative, nel senso che rimangono nelle fibrocellule del miocardio molti cosiddetti ‛pigmenti da usura', incarcerati dentro vacuoli autofagici o dentro usosomi secondari: essi sono relitti, spesso perossidati, di strutture cellulari, di pigmeriti mioglobinici e granuli di origine lipidica autossidati e genericamente denominati come ‛lipofuscina'. Tali materiali si trovano particolarmente accumulati nel citoplasma paranucleare (ai due poli del nucleo della fibrocellula) e impartiscono al miocardio atrofico un colorito bruno, onde il nome di ‛atrofia bruna' (v. fig. 5). A una atrofia da compressione può paragonarsi invece quella che nel miocardio si verifica per una cronica ipervolemia (dei bevitori di birra).
L'atrofia delle fibre muscolari scheletriche, che interviene dopo la denervazione, non interessa naturalmente solo il bilancio relativo fra le due fasi, mioplasmatica (prodotto differenziativo contrattile) e sarcoplasmatica (il resto del citoplasma muscolare), ma investe di necessità anche l'attività elettrica della membrana, come è documentato dall'insorgere, a più o meno breve distanza di tempo dalla denervazione, di una attivazione detta ‛spontanea' delle singole fibre muscolari, e pertanto elettrofisiologicamente rappresentata da potenziali assai piccoli per 1 o 2 millisecondi, i quali costituiscono il quadro elettrofisiologico della fibrillazione (v. Denny-Brown e Pennybacker, 1938). Essa è dovuta a una diffusa (e forse maggiorata) sensibilità della fibra all'acetilcolina, cioè a una perdita della localizzazione e del controllo dello stimolo, operato a livello della placca motrice. La fibrillazione è in genere anche l'inizio di processi endocellulari che portano a una regressione non più semplicemente atrofica della fibra.
Gli esempi fisiologici di ipertrofia e di iperplasia sono egualmente numerosi: durante la gravidanza e il puerperio la ghiandola mammaria, sotto lo stimolo ormonico della prolattina, acquista uno stato trofico e funzionale ben diverso da quello della mammella quiescente (v. sopra, invece, per il ritorno ‛involutivo' a questo stato). Così, sempre nella gravidanza, le cellule muscolari liscie del miometrio sono grandemente ipertrofiche, oltre che aumentate di numero; le cellule stromali della mucosa dell'utero, che si trasformano in cellule deciduali, possono essere buoni esempi di ipertrofia-iperplasia e di modulazione funzionale ormonodipendente. Anche i muscoli scheletrici, specie quelli legati alla determinazione dei caratteri sessuali secondari, possono subire ipertrofie e ipotrofie a seconda del sesso, dell'età, del periodo ciclo-ormonico.
Ma per i muscoli scheletrici il fattore più generalmente determinante l'ipertrofia, così come all'opposto per l'ipotrofia, è l'esercizio funzionale: sottoposto a continuo e regolare lavoro, quale quello dell'allenamento, un muscolo si ipertrofizza; in realtà, l'espressione ‛esercizio funzionale' racchiude tutta una serie di fattori cooperativi: non solo il susseguirsi di impulsi nervosi attraverso i quali arrivano alle fibre muscolari messaggi biochimici (forse non solo di univoca natura, acetilcolinica), ma anche lo stesso effetto meccanico della distensione e della contrazione ripetuta delle fibre stesse. Né va dimenticato il consensuale aumento della microcircolazione, quando sia possibile.
Almeno per il muscolo scheletrico, come per il muscolo liscio, non si può escludere un certo grado di iperplasia (cioè persino al livello cellulare, poiché è ovvia l'iperplasia del sistema contrattile negli elementi muscolari ipertrofici); essa può essere realizzata sia da nuovi contributi cellulari contrattili da parte di cellule staminali commiste alle già differenziate (‛cellule satelliti' che si moltiplicano e poi incrementano l'entità globale della differenziazione), sia, probabilmente, per scissione longitudinale di precedenti fibre (v. Aloisi e altri, 1972). Tali dati modificano l'affermazione comunemente accettata del Morpurgo (v., 1897) secondo cui l'ipertrofia muscolare da esercizio non è mai accompagnata da aumento numerico delle fibre.
Una comune e ben nota iperplasia fisiologica è quella della epidermide nelle zone di particolare attrito con l'esterno (al limite, la callosità); essa è correlata ad un eccesso di risposta del meccanismo autoregolativo, per cui le cellule basali dello strato epidermico vengono informate in qualche modo dell'entità degli strati sovrastanti cheratinizzati e vi reagiscono conseguentemente. Messaggeri sono le sostanze già citate come chaloni ad azione repressiva (a breve raggio) sulla attività mitotica dello strato basale dell'epidermide, e l'eccesso di risposta che provoca l'iperpiasia è dovuto alla prolungata e, quantitativamente, congrua stimolazione in superficie tendente a rimuovere continuamente materiale produttore di chaloni.
Analogamente, nell'adattamento di un organismo a condizioni di relativamente bassa pressione di O2 si ha iperplasia del tessuto emopoietico con policitemia adattativa.
Patologiche si considerano convenzionalmente le ipertrofie e le iperplasie conseguenti a un evento in qualche modo morboso; ma esse rispondono alle stesse correlazioni morfo-funzionali delle variazioni fisiologiche, e in tal senso deve essere interpretata l'ipertrofia miocardica che si verifica in quei settori del cuore che per un vizio dell'apparato valvolare sono sottoposti a maggior distensione e a maggior lavoro (compenso funzionale). Qui il meccanismo determinante l'aumento del materiale contrattile sembra sia esclusivamente o prevalentemente endocellulare (legge di Maestrini-Starling), anche se questo tipo di ipertrofia miocardica, come in misura di poco minore avviene per il muscolo scheletrico, è sempre accompagnato anche da sensibile mobilitazione (con proliferazione cellulare) del mesenchima circostante.
Più complessa appare la genesi dell'ipertrofia vicariante (che è anch'essa in qualche misura) compensatoria degli organi pari per asportazione di uno dei due (per es., rene). Anche qui vi è un momento iperplastico (minore o posteriore nel tempo) che si aggiunge a quello puramente ipertrofico. Forse il meccanismo è da ricercarsi nelle modificazioni fisiologiche indotte dal sovraggiunto maggior lavoro; ma in altri casi si è spinti a pensare (e a cercare) un meccanismo più lato, che coinvolga messaggi percorrenti il circolo sanguigno e informanti della quantità di tessuto in azione (quindi agenti in modo autoregolativo, verso il ripristino dell'omeostasi). È comunque accertato che anche nella ipertrofia vicaria del rene esiste una dipendenza dall'ormone somatotropo ipofisario.
Tra le ghiandole endocrine esistono stretti collegamenti autoregolativi ed è pertanto molto facile trovare in esse esempi fisiologici e patologici di ipertrofie e iperpiasie: basti pensare alle cellule cortico-surrenali e all'ormone ipofisario corticotrofico (ACTH), o alle cellule dei follicoli tiroidei e all'ormone tireotrofico, e considerare le relazioni speculari, cioè l'influsso sulle cellule ipofisarie degli ormoni delle altre ghiandole. Una ipocalcemia (per es., da deficiente assorbimento di calcio dall'intestino) provoca una iperplasia delle paratiroidi e una conseguente ‛atrofia' della parte calcificata dell'osso, come tentativo di ripristino della omeostasi calcio-ionica del ‛mezzo interno' (plasma sanguigno, ecc.).
2. Alterazioni della differenziazione. - Abbiamo già visto che ipotrofia e ipertrofia significano spesso, al livello sub-cellulare, anche una diminuzione o un aumento del prodotto differenziato della cellula. Il tessuto muscolare è esemplare a questo proposito (modificazioni della fibrillogenesi). Analogamente, una iperplasia epidermica può essere accompagnata da aumento della cheratinizzazione; una produzione di emergenza di globuli rossi può essere accompagnata da imperfetta loro differenziazione (residui nucleari, non completo arricchimento di emoglobina, ecc.).
Alterazioni qualitative della differenziazione si presentano come produzione di prodotti differenziativi abnormi e quindi in certo senso anche come ‛mostruosità'. Tale possibilità, non disgiunta dalle variazioni puramente quantitative, si verifica in particolare in alcuni cloni di una popolazione di cellule che abbia subito la trasformazione neoplastica (v. sotto) ed è una dimostrazione della possibilità - seppure non frequente e peculiare - di una utilizzazione del tutto diversa della informazione genetica, che rimane tutta conservata, e persino disponibile in circostanze abbastanza eccezionali per le cellule animali (mentre è assai facile presso i vegetali, ove cellule già altamente differenziate del soma possono riprodurre l'intera pianta, gameti compresi).
Molto più frequentemente è possibile osservare nelle cellule animali modulazioni reversibili di uno stesso processo differenziativo, e in questo caso si deve ammettere che le variazioni riscontrabili, e con particolare incidenza anche in sede patologica (per es., sotto l'influenza di stimolatori ormonici o per fattori esterni), siano dovute a oscillazioni più o meno ampie dell'equilibrio tra repressori e derepressori del sistema genetico della cellula (settore C della fig. 1) (v. Harris, 1964). Un epitelio pavimentoso composto non cheratinizzante può produrre uno strato corneo, come se fosse un ulteriore proseguimento della differenziazione (prosoplasia), in circostanze varie, quali l'abnorme contatto col mondo esterno, come avviene nei prolassi uterini e rettali, o in alcune avitaminosi; ad esempio, l'epitelio bronchiale, che è cilindrico, può divenire (reversibilmente) pavimentoso composto (metaplasia malpighiana) e cheratinizzare per stimoli esterni come il fumo o nella avitaminosi A; ancora, stimoli ormonici producono pure metaplasia malpighiana (a tipo epidermico) negli epiteli degli sbocchi ghiandolari cervico-uterini. Un esempio che può apparire del tutto abnorme e, come si diceva, quasi ‛mostruoso', è la presenza di ciglia vibratili fuori sede, come nei tubuli renali o nei canalicoli biliari.
Nelle cellule tumorali maligne si assiste spesso alla tendenza opposta a quella differenziativa, specie nei gradi avanzati di malignità, cioè a perdere molti caratteri distintivi della differenziazione (anaplasia) cui erano pervenute le cellule prima della loro trasformazione neoplastica.
Ma bisogna anche tener presente che quando si parla di differenziazione, dedifferenziazione (o sdifferenziazione o anaplasia) o di differenziazione abnorme, ecc., occorrerebbe precisare se ci si riferisce a eventi implicanti singole, individuali cellule (come per le fibre muscolari scheletriche) o in genere a una popolazione cellulare. Quest'ultima, in quanto tale, può cambiare la sua omeostasi differenziativa per la prevalenza numerica di elementi cellulari più o meno differenziati rispetto ad altri, di cellule in continua sollecitazione moltiplicativa rispetto ad altre nelle quali subentra il processo differenziativo specifico (v. sotto, punto N, Trasformazione neoplastica).
Anomalie della differenziazione possono investire anche le sostanze intercellulari, peraltro prodotte dalle cellule: formazione ectopica di osso o, all'opposto, difetti della ossificazione; incapacità delle fibre collagene a maturare, come nella avitaminosi C o nella intossicazione da amminoacetonitrile (v. sotto, cap. 3, punto H), ecc.
3. Degenerazioni o distrofie. - Quando un evento patologico investe in modo più o meno specifico i settori A e B dell'apparato cellulare (v. fig. 1) si possono verificare gravi disordini della compagine citoplasmatica e del suo metabolismo (esistono anche, come vedremo, alterazioni degenerative dell'apparato nucleare, le quali però coincidono a scadenza più o meno breve con la morte della cellula o, come avviene fisiologicamente per i globuli rossi, con la comparsa di irreversibili modificazioni cellulari). Tali disordini con facilità arrivano a un livello sufficientemente elevato da produrre conseguenze strutturali, morfologiche, quelle che classicamente vanno sotto il nome di degenerazioni cellulari e di distrofie tessutali. La gravità di queste evenienze può anzi essere o divenire tale da innescare una serie di ulteriori processi irreversibili e largamente comuni, indipendentemente dalla specifica azione della noxa originaria: cioè la degenerazione può anche concludersi con la necrobiosi.
Uno sguardo alle componenti di questi due settori, e soprattutto la considerazione del fatto che in A compaiono i meccanismi rifornitori dell'energia, fa comprendere come questo esito comune sia sempre immanente. Le stesse strutture subcellulari quali i mitocondri, il reticolo endoplasmatico, l'apparato di Golgi, i lisosomi, la membrana esterna sono costituiti da membrane la cui integrità morfo-funzionale, che è condizione della reversibilità delle oscillazioni fisiologiche della cellula, è a sua volta condizionata dal rifornimento energetico.
Si stabilisce così facilmente un circolo vizioso che porta a spostamenti eccessivi del pH verso l'acidità (abbassamento dell'ambito normale del pH intracellulare), fughe ioniche e conseguenti inibizioni o esaltazioni abnormi di determinati processi biochimici (tra queste l'abnorme consumo di O2 per disaccoppiamento delle ossidazioni dalle fosforilazioni e perdita del controllo respiratorio da parte dei mitocondri alterati), ulteriore diminuzione del rifornimento energetico, attivazione e aumento dei lisosomi e crollo dell'attività di membrana esterna (vedi necrobiosi e necrosi).
Naturalmente, così inteso, un simile processo al suo inizio specifico può apparire come solo degenerativo, sarà differenziato e anche si dimostrerà reversibile; anzi vero- similmente l'attività lisosomiale deve essere associata ai meccanismi di rinnovo e quindi di autoconservazione della cellula, ed è solo quando subentra il circolo vizioso e la irreversibilità che inizia il processo necrobiotico.
Esaminiamo ora i vari tipi di degenerazioni, comprese quelle delle sostanze intercellulari.
A. Rigonfiamento torbido, degenerazione vacuolare e idropica. Sono aspetti degenerativi che riguardano principalmente il settore A dello schema della cellula, la membrana esterna, il sistema canalicolare e vacuolare topologicamente continui con lo spazio extracellulare, i mitocondri anch'essi inclusi (per la loro membrana esterna) nello stesso sistema. Come tali, si tratta o di strutture molto esposte agli agenti esterni fisici, tossici, immunologici, o comunque di strutture le cui membrane limitanti in buona misura lipoproteiche, eventualmente rivestite per alcuni tratti della sola membrana esterna dai mucopolisaccaridi della membrana basale (glycocalyx), mantengono la loro integrità e le loro importanti caratteristiche elettriche (potenziali di membrana) solo con continuo dispendio energetico. Tale dispendio si fa a opera di sistemi enzimatici di trasporto o configurazionali della membrana e a spese di composti facilmente liberanti energia come l'ATP; il sistema stesso è in grado, nella sua integrità, di ripristinare tale energia risintetizzando quei composti (a livello del reticolo endoplasmatico e soprattutto a livello dei mitocondri) a spese di processi metabolici di cui fondamentali sono la glicolisi e l'ossidazione (fase endocellulare della respirazione).
Integrità funzionalmente reversibile di queste membrane (così come di quelle interne dei mitocondri: cristae) e gradiente ionico e potenziale elettrico non sono che aspetti di uno stesso stato di cose; la distribuzione ionica da una parte e dall'altra delle membrane è quasi sempre il risultato di un lavoro biologico; ai fenomeni di diffusione fisico-chimica si superimpongono fatti di attiva segregazione delle componenti ioniche la cui distribuzione si allontana per ciò stesso dall'equilibrio (per es. l'equilibrio multi-ionico di Gibbs-Donnan), e invece tende a raggiungere questo equilibrio proprio con la degradazione e la morte cellulare. Si comprende il lavoro che si svolge a livello di queste membrane anche solo pensando che, mentre attraverso esse e possibile la diffusione di ioni come K+ e Na+ e di alcuni anioni endocellulari, la distribuzione reale dei primi è attivamente diversa dall'effetto diffusione e varia a seconda degli stati funzionali della cellula. In tal modo anche l'ingresso dell'acqua, che non è soggetto a trasporto attivo, viene tuttavia a essere controllato: una diminuzione di questo controllo tende a rigonfiare la cellula per l'esistenza in essa, di regola, di una osmolarità e quindi di una relativa pressione maggiore di quella esterna.
Oltre alla regolazione del trasporto attraverso l'attività di membrana, la cellula ha commercio col mondo esterno attraverso alcuni meccanismi di endocitosi, quali la pinocitosi e la fagocitosi (v. fig. 6): si tratta di processi per cui piccole porzioni dell'ambiente esterno, costituite da liquidi, particelle in essi contenute tra cui particelle virali, corpi estranei, batteri, protozoi, penetrano nella cellula per una microinvaginazione della membrana cellulare e, prima di subire varie trasformazioni, rimangono nel citoplasma in forma di microvacuoli. Anche il fenomeno reciproco si verifica: microvacuoli con inclusioni varie si avvicinano alla membrana cellulare (si ricordi che la parete di questi microvacuoli, comunque originati dal sistema canalicolare, è omologa alla membrana cellulare ma con polarità invertita, poiché il loro interno è omologo allo spazio extracellulare), si fondono con essa e poi si aprono verso l'esterno; si realizza in tal caso quel fenomeno, facente parte del più generale processo della esocitosi, definito come ‛eiezione' (v. fig. 7).
Un aspetto topologicamente invertito o speculare è quello di piccole porzioni del gel o succo citoplasmatico, con i loro possibili contenuti (granuli e rosette di glicogeno, ribosomi liberi, microtubuli e microfilamenti, gocce lipidiche), che si estroflettono, rivestite all'esterno dalla membrana (ed eventualmente dal glycocalyx), e si staccano all'esterno: è quanto accade nelle cellule ragiocrine e in quelle secretive (in quest'ultimo caso, nella estroflessione, è contenuto il prodotto secretivo), secondo i processi esocitosici indicati rispettivamente con i termini ‛clasmatosi' e ‛secrezione'. È anche possibile in qualche misura il reciproco evento, per esempio della fusione tra due cellule omologhe. Da alcuni (v. LaVia e Hill jr., 1971) questo processo di estroflessione è chiamato ‛exotropico' (v. fig. 7). L'omologia fra membrane e spazi interni della cellula con la membrana esterna e lo spazio extracellulare autorizza a parlare anche di una esotropia e di una exotropia, con i loro reciproci aspetti di reversibilità, e secondo le modalità or ora accennate, anche tra gel citoplasmatico (con i suoi contenuti) e il sistema canalicolare e vescicolare della cellula (reticolo endoplasmatico, apparato di Golgi, apparato lisosomiale), realizzandosi così un complesso movimento delle membrane relative, una loro notevole plasticità e tramutabilità (vedi rapporti tra reticolo canalicolare del Golgi e quello endoplasmatico, i lisosomi, i vacuoli secretivi, ecc.; vedi anche le vie di comunicazione con la membrana esterna dei mitocondri e quella del nucleo).
Quanto ai mitocondri, il loro volume, che pure è regolato da una serie di eventi legati al trasporto attivo, subisce per ciò stesso variazioni in relazione alla loro specifica attività fisiologica: un modesto rigonfiamento di tali organuli dallo stato di riposo si verifica reversibilmente in presenza di substrati utilizzabili per l'ossidazione e di ossigeno. La reversibilità sussiste anche quando sia disponibile una certa quantità di accettore di fosfati (ADP) per cui il processo ossidativo viene esaltato e buona parte dell'energia derivante viene conservata sotto forma di ATP. Si dice allora che i mitocondri sono in grado di accoppiare le ossidazioni con le fosforilazioni, o che il controllo respiratorio esercitato dall'ADP (come innesco) è in atto (v. fig. 8).
In condizioni patologiche varie, per esempio in anossia (v. fig. 9), i mitocondri possono rigonfiarsi eccessivamente e irreversibilmente (perdita del controllo del trasporto ionico, squilibrio elettrodinamico, entrata di ioni Ca++ e dell'acqua, accumulo di fosfati di calcio tipo idrossiapatite), e la loro membrana esterna può rompersi.
Alterazioni della membrana esterna possono portare a modificazioni configurazionali (v. fig. 10) riconoscibili alla microscopia elettronica e persino ottica, formazione eccessiva e abnorme di gemmazioni exotropiche (analoghe a quelle fisiologiche osservabili dopo la divisione cellulare in cellule libere in coltura), alterazioni delle giunzioni fra cellule, produzione di ‛figure mieliniche' costituite principalmente dalla parte fosfolipidica della membrana che è così profondamente alterata, notevole perdita del controllo volumetrico della cellula stessa (entrata di acqua, o degenerazione idropica, e facile avvio verso la necrobiosi irreversibile) (v. fig. 11). Un eccessivo accumulo di vescicole pinocitosiche per abnorme stimolo all'assunzione di materiale dall'esterno, come nel caso di un eccessivo riassorbimento da parte dell'epitelio tubulare renale dopo somministrazione di quantità notevoli di saccarosio o di mannitolo, realizza il quadro della degenerazione vacuolare (nefrosi osmotica); ma si è visto questo tipo di degenerazione anche in condizioni di ischemia (deficiente irrorazione sanguigna, quindi anossia cellulare), cioè in condizioni in cui non è facile il rifornimento di energia.
Quando i mitocondri subiscono globalmente un sensibile rigonfiamento (anche del tipo funzionale e reversibile), essi possono fare assumere alla cellula, specie se coesiste un certo stato di idratazione del citoplasma, l'aspetto del rigonfiamento torbido (la torbidità essendo dovuta alla granulosità mitocondriale che si fa evidente anche con le comuni colorazioni al microscopio ottico). Dalla sua primitiva descrizione e interpretazione virchowiana si sono avute varie e contrastanti opinioni sul rigonfiamento torbido; si sa che, fino a un certo punto, esso è reversibile e che può esser causato da azioni tossiche o tossiniche sulla cellula (per es. dalla tossina difterica nelle fibre muscolari cardiache o scheletriche) o infettive (tifo addominale, epatite virale) o in condizioni di anossia, o per eccesso di ormone tiroideo (tiroxina). L'epitelio dei tubuli renali va facilmente incontro a rigonfiamento torbido reversibile in condizioni di iperlavoro (anche all'inizio della ipertrofia vicariante), nella febbre, ecc. (significato di maggior impegno funzionale).
Al microscopio elettronico si nota, oltre al rigonfiamento e al distanziamento delle cristae, anche frammentazione di queste, perdita di materiale granulare proveniente dalla matrice. Si comprende bene come, in uno stadio successivo, irreversibile per riduzione massimale dell'apporto energetico alla cellula, il quadro si accompagni a una degenerazione idropica e porti alla necrobiosi cellulare.
B. Degenerazione grassa, steatosi. Molte cellule normali presentano variabili quantità di grasso, che può aumentare in condizioni fisiologiche fino a veri e propri accumuli; a parte il tessuto adiposo e altre cellule che normalmente si caricano di prodotti di secrezione costituiti da sostanze lipidiche o liposolubili, compare grasso nella senescenza delle cellule, per esempio nelle cellule cartilaginee, nervose, ecc. Un abnorme accumulo di grassi entro una cellula, quantitativamente tale da divenire visibile in modo cospicuo anche al microscopio ottico e persino da costituire una massa omogenea occupante quasi tutto il volume cellulare (v. fig. 12), come nel tessuto adiposo, è possibile per le seguenti principali ragioni (che valgono per qualunque cellula particolarmente competente per il metabolismo lipidico, ma soprattutto per quella epatica, ove massimamente si determina e fruttuosamente si può studiare il processo di accumulo): per un arrivo di acidi grassi o grassi neutri eccessivo rispetto alle possibilità metaboliche di utilizzo o di secrezione; per un'abnorme attività neolipogenetica dagli idrati di carbonio o dagli amminoacidi; infine, per lo scompaginamento delle numerose strutture lipoproteiche (membrane) della cellula, che è seguito dalla evidenziazione della parte meno solubile e cioè lipidica (fosfolipidica, ecc.). Quest'ultima evenienza, nota agli antichi osservatori come ‛lipofanerosi', è naturalmente espressione di grave alterazione cellulare (specialmente se essa è già rilevabile al microscopio ottico) e sinonimo di lesione letale in atto (v. fig. 13).
Si ha accumulo e degenerazione grassa nelle cellule del fegato in occasione dei seguenti stati morbosi: iperalimentazione grassa, digiuno, kwashiorkor e altre carenze alimentari (e in special modo quelle di fattori lipotropi come la colina), alcolismo acuto e cronico, anossie e anemie, intossicazioni (vedi steatosi sperimentali), azioni tossiniche e stati infettivi (tubercolosi, infezioni virali del fegato), malattie metaboliche (diabete), encefalopatie, ecc. I grassi accumulati sono in tutti questi casi prevalentemente trigliceridi, ma una così vasta serie di situazioni eziopatogenetiche fa naturalmente pensare alla steatosi come a un epifenomeno terminale facilitato dalle caratteristiche metaboliche della cellula epatica. Non è negabile una conclusione del genere, ma è tuttavia possibile oggi, anche per le informazioni derivanti dalle ricerche della patologia sperimentale, quali poi saranno qui in seguito esposte, semplificare entro certi limiti tale molteplicità eziopatogenetica, cioè trovare per alcune forme di steatosi epatica tratti patogenetici comuni.
Appare cioè fattore importante e centrale in molti casi la disponibilità della cellula epatica a compiere sintesi proteiche in misura adeguata alla regolazione della veicolazione al suo esterno dei grassi, sotto forma di complessi lipoproteici a bassa densità. Si comprende allora come tale tratto della fisiologia della cellula epatica possa venir compromesso in molte delle suesposte circostanze patologiche, secondo una patogenesi anche assai diversa, ma conducente allo stesso risultato, cioè all'incapacità o inadeguatezza della cellula a liberarsi degli acidi grassi e dei trigliceridi che le arrivano o vi si producono: o per una eccessiva alimentazione grassa (complessi chilomicronici dall'intestino); o per una mobilizzazione degli acidi grassi dai trigliceridi delle riserve adipose (digiuno); o per una dieta, eventualmente anche abbondante dal punto di vista quantitativo, ma povera di proteine (kwashiorkor, inanizione ipoproteica, ipoalimentazione o iporiassorbimento dell'alcolista), la quale riduce la riserva di amminoacidi disponibile per le sintesi proteiche; o per una scarsa disponibilità energetica per la sintesi (anossia da stasi circolatoria, anemie gravi, intossicazioni da veleni che blocchino i processi metabolici ossidativi); o per una disorganizzazione della macchina sintetica delle proteine di secrezione della cellula epatica (veleni che alterino il reticolo endoplasmatico rugoso o deviino i metaboliti intermedi); o, infine, per un'eccessiva lipogenesi da altri settori del metabolismo (diabete, ecc.).
Un'altra causa di steatosi della cellula del fegato è rappresentata dalla carenza (che può prodursi sperimentalmente, ma che spesso è associata a una generale carenza proteica) di fattori detti ‛lipotropi', di cui la colina (o i composti metilanti che la realizzano) è uno dei più noti: la carenza di colina si traduce in carenza di produzione di fosfolipidi, i quali possono avere molta importanza sia nella utilizzazione dei grassi (nella cellula epatica o altrove), sia come materiali di rinnovo delle strutture stesse della cellula (membrane), onde si può tornare al più generale processo prima ricordato.
Altri tipi di cellule che possono subire in modo cospicuo la degenerazione grassa sono le cellule miocardiche sotto l'azione della tossina difterica, gli epiteli renali in certi tipi di nefropatie (nefrosi lipoidea) associati a iperlipemia e ipercolesterolemia, gli epiteli intestinali per disturbo della veicolazione dei grassi al livello postassorbimento (anche qui è importante la presenza delle proteine, poiché nella formazione dei chilomicroni, ai trigliceridi si associano fosfolipidi e β-lipoproteine), le cellule del processo infiammatorio, durante l'evoluzione di questo.
C. Degenerazione glicogenica. Abnormi quantità di glicogeno possono trovarsi, in varie condizioni patologiche, nel citoplasma, e persino nel nucleo delle cellule. Nel diabete, sia per l'elevata concentrazione del glucosio ematico nei confronti di cellule, come quelle del miocardio, che appaiono meno dipendenti dall'insulina, sia per neoglucogenesi cellulare (fegato, rene, muscolo scheletrico), si può avere un accumulo di glicogeno. Ma notevolissimi accumuli di glicogeno si trovano nelle ‛tesaurismosi' glicogeniche o glicogenosi sulle quali, essendo alterazioni a carattere genetico, sarà fatto cenno in seguito.
D. Degenerazioni ialine. Depositi di materiale, che appare acidofilo, compatto e trasparente, vitreo (ialino) al microscopio ottico, si possono trovare in cellule che hanno subito lesioni di varia natura. Il termine infatti ha solo un riferimento morfologico. Si tratta quasi sempre di materiali proteici o glicoproteici che si accumulano nel citoplasma come prodotti abnormi, o quantitativamente eccessivi in cellule che normalmente ne contengono: gocce ialine proteiche spesso incluse nelle strutture vacuolari (esocitosi) o lisosomiali degli epiteli dei tubuli renali, quando questi riassorbano proteine abnormemente filtrate dai glomeruli (glomerulonefriti, glomerulonefrosi); corpi ialini (al microscopio elettronico finemente filamentosi) delle cellule epatiche nella cirrosi alcolica (v. fig. 14); corpi paracristallini fortemente acidofili nelle cisterne (reticolo endoplasmatico rugoso) secretive delle plasmacellule iperstimolate nei processi fiogistico-immunitari (v. figg. 15 e 16); masse ialine nelle fibre muscolari distrofiche, ove peraltro esse rappresentano porzioni di mioplasma coagulato e risolto in zolle (zolle di Zenker) per fenomeni contratturali terminali; conglomerati piastrinici che si fondono e assumono in breve tempo aspetto omogeneo e ialino. Dal punto di vista fisiologico possiamo considerare una metamorfosi ialina quella delle cellule epidermiche che si caricano di materiali cheratinici e infine si trasformano in squame lamellari ialine, acidofile (così avviene nei tumori cheratinizzanti: perle cornee ialine).
E. Degenerazioni mucose e mucoidi. Le cellule caliciformi e altri elementi epiteliali delle - o annessi alle - mucose secernono mucina, che è un insieme di proteine coniugate con oligo- e polisaccaridi contenenti anche acetil-glucosammina e mucopolisaccaridi acidi. Una sostanza simile, detta ‛mucoide', si trova come sostanza fondamentale anista del mesenchima; glicoproteica è, almeno in parte, anche la natura delle membrane basali.
Già nella normale secrezione di muco da parte delle cellule caliciformi assistiamo al loro riempimento progressivo di secreto, che si accumula nella parte distale e centrale della cellula respingendo il citoplasma residuo e il nucleo (che diviene spesso picnotico: v. sotto) all'estremità prossimale. Questo è un quadro fisiologico, ma in talune circostanze, come ad esempio nelle stesse mucose sotto lo stimolo infiammatorio o in cellule derivate da epiteli mucipari, si determina un eccesso di secrezione accompagnato da fenomeni regressivi della cellula, fino al suo disfacimento: si parla allora di degenerazione mucosa degli elementi in questione.
Della patologia del mucoide dei connettivi diremo appresso; ma va ricordato fin da ora che prodotti quantitativamente e qualitativamente abnormi di questa sostanza (talora anche la sostanza amiloide; v. sotto) possono penetrare (o anche prodursi) entro le cellule, e allora è possibile osservarli all'interno dei vacuoli o dei fagosomi o direttamente nelle cisterne del reticolo endoplasmatico.
F. Aspetti patologici delle cellule nervose. Già fisiologicamente, si possono osservare nelle cellule nervose gangliari e del nevrasse accumuli di prodotti lipidici perossidati (cromolipidi) che rivelano la senescenza del neurone (sappiamo che in condizioni fisiologiche molti neuroni degenerano o scompaiono ancora prima del raggiungimento del completo sviluppo postnatale). Ma il carattere più specifico dei fenomeni regressivi del tessuto nervoso è legato alla integrità di quella unità chiamata neurone, di cui la cellula, o pirenoforo, è il centro trofico. Il taglio dell'assone, che ne è un prolungamento, o la soppressione delle connessioni e degli stimoli afferenti provoca fenomeni regressivi: nella cellula si ha la cromatolisi (possibilmente reversibile), caratterizzata da una rapida scomparsa della massa ribosomiale e comunque dell'RNA e delle ribonucleoproteine citoplasmatiche; nel moncone periferico del cilindrasse si ha la ‛degenerazione walleriana', consistente in disgregazione e autofagia della mielina e di frammenti dell'assone da parte delle cellule di Schwann; al microscopio elettronico sono visibili numerose figure mieliniche.
G. Attacco di anticorpi sulla membrana basale e superficie cellulare. Anticorpi (etero-, iso- e autoanticorpi) verso determinate cellule possono circolare e raggiungere appunto quelle cellule che posseggono i ricettori antigeni superficiali; anticorpi e complessi solubili antigene-anticorpo possono circolare e aderire a determinate strutture endoteliali o alle loro membrane basali o alle membrane di altre cellule alterandone il trofismo. Il sopraggiungere su tale complesso di frazioni complementari del siero provoca vera e propria lisi cellulare. Il fenomeno è in modo esemplare verificabile nei globuli rossi, la cui membrana ne risulta gravemente danneggiata (v. fig. 17) così da determinare la fuoriuscita della emoglobina (emolisi). Si ricorda però che una lisi delle membrane, in specie dei globuli rossi, si può determinare anche in circostanze che non sono immunitarie, per effetto di tossine batteriche ad azione emolitica (emolisine batteriche); per un difetto biochimico congenito dei globuli rossi (più spesso della emoglobina; v. sotto) da cui risulta una minore resistenza dei globuli rossi stessi, un più precoce invecchiamento e lisi extra-cellulare; infine, ovviamente, per fagocitosi.
H. Patologia cellulare relativa al nucleo e al nucleolo. Nei processi patologici il nucleo può subire diverse alterazioni morfologiche globalmente regressive: rigonfiamento e lisi (cariolisi), all'opposto picnosi; si possono accumulare in esso gocce lipidiche o depositi di glicogeno (vacuolizzazioni nucleari) fino a ridurre le strutture cromatiniche tutte e solo a ridosso della membrana nucleare propria (v. figg. 18 e 19). Naturalmente il nucleo può contenere inclusioni virali (v. fig. 20). Un ingrandimento del volume nucleare e una distribuzione particolare della cromatina intorno al nucleolo (o ai nucleoli), anch'esso sensibilmente ingrandito e ben acidofilo, si riscontrano in alcune cellule, come quelle muscolari, i cui nuclei anche senza potersi suddividere in situ, vengono tuttavia ‛attivati' in situazioni che preludono a una rigenerazione (v. sotto, cap. 4).
Nella divisione nucleare si possono manifestare anormalità del genoma, nel senso di addizioni cromosomiche (per es., trisomia autosomiale, come nella sindrome di Down; trisomia G o addizioni di cromosomi del sesso, come nella sindrome di Klinefelter), oppure nel senso opposto di delezioni (assai rara la delezione completa autosomiale come nella monosomia G, più frequenti le delezioni parziali di cromosomi autosomiali) con conseguenze fenotipiche varie, metaboliche e strutturali; invece è riscontrabile la delezione completa di un cromosoma del sesso (come nella sindrome di Turner). Tali anomalie, come è quasi sempre il caso della patologia genetica descrivibile, hanno naturalmente conseguenze varie nel fenotipo in dipendenza della sezione del genoma alterato, a meno che non si tratti di semplici translocazioni compensate e senza perdita effettiva di materiale genetico. Altrimenti possono aversi variabili risultanze patologiche fenotipiche e certamente non tutte arrivate all'osservazione.
Molte delle aberrazioni genomiche si determinano al momento della meiosi e sono pertanto - se vitali - trasmissibili ai prodotti del concepimento. Esse possono essere anche dipendenti da fattori esterni fisici (radiazioni), o da altri agenti mutageni e comunque lesivi della compagine nucleare, specie nella sua fase mitotica o meiotica, se applicati agli organi portatori delle cellule germinali. Naturalmente sono possibili alterazioni genomiche anche nel corso della mitosi delle cellule somatiche, durante tutta la vita e per diversi stimoli, tra cui, molto importante, l'inclusione di un virus, la formazione di un protovirus o di un provirus, ecc. (v. Temin, 1972). Tali alterazioni sono state studiate soprattutto a proposito della trasformazione tumorale delle cellule (v. sotto), ma fin da ora possiamo dire che esse, anche se molto frequenti (poliploidie, dispersione e alterazioni di cromosomi, cariocinesi atipiche, tripolari o multipolari), non rappresentano il segno essenziale della malignità tumorale né della natura tumorale della cellula; sono piuttosto il risultato sporadico della frequenza abnorme di mitosi e delle condizioni nelle quali queste si verificano nelle cellule già divenute tumorali. Non si può tuttavia escludere la possibilità di reperire neoplasie con costante, regolare alterazione genomica: finora, peraltro, una simile condizione è stata riscontrata soltanto nella leucemia mieloide cronica umana, in cui Nowell e Hungerford (v., 1962) hanno dimostrato in un'alta percentuale di casi un cromosoma abnorme (con molta probabilità il 21) per delezione di una sua parte; tale cromosoma alterato è stato denominato cromosoma Philadelphia (Ph1) (per i rapporti tra natura tumorale e genoma delle cellule, v. Nowell, 1965).
I. Patologia cellulare relativa ai difetti genetici (e quindi più o meno congeniti) del metabolismo. Un difetto nell'assetto genomico di un determinato tipo di cellula che investa la parte di quel genoma che si esprime fenotipicamente (cioè si trascrive e si traduce secondo le normali sequenze e caratterizza appunto il tipo di cellula) non può non tradursi in un difetto enzimatico-strutturale di tutte le cellule di quel tipo differenziativo, con conseguenze ulteriori molto varie, a seconda dei casi, per l'organismo intero. Alcuni di questi difetti possono essere chiaramente ereditabili.
1. Le tesaurismosi glicogeniche. Già nella ontogenesi normale conosciamo numerosi esempi di cellule che, mentre da una parte sintetizzano attivamente glicogeno, dall'altra lo demoliscono assai poco per le vie conosciute nella vita adulta: in questi casi la replezione di glicogeno, coincidendo con la morte della cellula, termina con meccanismi autofagosomiali e lisosomiali. La replezione e il successivo disfacimento delle cellule hanno qui un valore morfogenetico (apertura di cavità, modulazioni delle mucose, canalizzazioni ghiandolari: v. Aloisi, 1933): la demolizione metabolica del glicogeno è, per molte cellule, acquisizione relativamente tardiva durante lo sviluppo.
Vi è una condizione patologica, caratterizzata dalla permanenza di uno stato analogo a quello embrionale, in cui buona parte del glicogeno cellulare trova il suo destino nel sistema lisosomiale; ma in questo manca, per difetto genetico, la maltasi acida o α-1,4-glucosidasi (v. Hers, 1965). Ne consegue una patologica replezione della cellula di lisosomi carichi di glicogeno non scindibile e sottratto alle catene glicogenolitiche ordinarie: è questo il caso della tesaurismosi glicogenica tipo II o glicogenosi tipo II di Cori, o morbo di Pompe, che coinvolge molti tipi di cellule. Altre glicogenosi sono: quella per difetto congenito della glucoso-6-fosfatasi con compromissione delle cellule epatiche e di quelle renali (malattia di von Gierke), o quella per difetto della fosforilasi; in quest'ultimo caso, poiché le fosforilasi del fegato e del muscolo sono molecolarmente diverse, si distinguono di regola due forme di glicogenosi, la epatica o malattia di Hers e la muscolare o malattia di McArdle, come due forme cioè topograficamente dissociate. Infine, possono esservi glicogenosi caratterizzate dalla presenza di glicogeno abnorme, in quanto questo polisaccaride si modifica enzimaticamente dal punto di vista della architettura molecolare prima di completare la sua polimerizzazione. Tutte queste forme patologiche, come quelle che citeremo tra poco, sono evidentemente esempi ormai classici di patologia molecolare.
2. Altre tesaurismosi. Vi sono tesaurismosi lipidiche o lipidosi, distinte in colesteroliche (malattia di Hand-Schüller-Christian), fosfatidiche (malattia di Nieman-Pick), cerebrosidiche (malattia di Gaucher); tesaurismosi glicoproteiche, comprendenti le varie mucopolisaccaridosi (malattia di Hurler, malattia di Morquio, malattia di Sanfilippo) e forse la lipodistrofia intestinale (malattia di Whipple) che interessano vari tipi di cellule e spesso quelle del mesenchima (sistema reticolo-istiocitario); frequentemente il materiale che si accumula è in sede lisosomiale (v. Dingle e Fell, 1969; v. Hers, 1965).
3. Emoglobine patologiche. Una patologia molecolare dei globuli rossi consiste nella presenza in essi, per difetto genetico ed ereditabile, di emoglobine molecolarmente abnormi o di un'abnorme popolazione di emoglobine normali: un esempio di questa condizione è caratterizzato da un'eccedenza di emoglobina fetale (HbF) per carenza di catene β della globina, anomalia che determina una delle forme di ‛talassemia' contrassegnata, anche nello stato eterozigote, dal tratto morfologico della microcitemia (globuli rossi più piccoli del normale) e nello stato omozigote dalla notevole facilità all'emolisi (anemia emolitica per distruzione dei globuli rossi). Analogamente l'emoglobina S (presenza di valina al posto dell'acido glutammico nella catena β della globina), che allo stato ridotto cristallizza e deforma il profilo dei globuli rossi (globuli rossi a falce), nell'omozigote determina egualmente una anemia emolitica.
4. Disordini congeniti enzimatici delle cellule degli epiteli renali possono portare a incapacità varie nell'attività secretiva e riassorbente, con comparsa di prodotti anomali nelle urine; moltissime altre condizioni collegabili con difetti genetici potrebbero essere qui elencate a carico dell'intero organismo e di suoi particolari gruppi di cellule differenziate. Ci limitiamo qui a citare le distrofie muscolari ereditarie, e particolarmente quelle associate a difetto recessivo di un cromosoma X; in questi casi il carattere patologico, trasmissibile ai maschi da femmine portatrici eterozigoti, interessa in modo precipuo le fibre muscolari striate, le quali vanno incontro a distruzione lenta ma progressiva (forma di distrofia muscolare progressiva di Erb).
L. Infiltrazioni corpuscolari esogene, patologia legata alla fagocitosi, al parassitismo endocellulare; la mobilitazione flogistica. Particelle del mondo esterno non parassitanti, quali polveri inerti (carbone) o capaci di produrre sequele nell'apparato respiratorio (silicio), possono venire malate e ritrovarsi nelle cellule fagocitanti anche persistentemente nella misura in cui non siano (come quelle suaccennate) in alcun modo digeribili dagli enzimi lisosomiali e fagosomiali. Nelle virosi e nelle infezioni batteriche, protozoarie, fungine, ecc. si determina tutta una modulazione delle cellule ospiti (non necessariamente le cellule del sistema fagocitante mesenchimale, ma anche cellule epatiche, muscolari, nervose o endoteliali, ecc.) con frequente formazione, come nel caso dei virus, di ‛corpi inclusi' contenenti il parassita, come i corpi del Negri delle cellule nervose degli animali affetti da rabbia. In alcuni casi, la presenza persistente dei microrganismi nell'interno delle cellule, o vera infezione endocellulare, è il prodotto di un ‛parassitismo opportunistico', cioè di un equilibrio dinamico raggiunto tra cellule ospiti e attività aggressiva da parte del microbo; in tal caso la modulazione cellulare può essere morfologicarnente molto vistosa, come la formazione di cellule giganti multinucleate assai simili agli osteoclasti normali dei Mammiferi (v. fig. 21).
Le cellule del sistema mesenchimale che partecipano all'evento patologico complesso e coordinato quale è la flogosi (polimorfonucleati, linfociti, monociti) e quelle di esse che innescano la reazione generale immunitaria (monociti, linfociti) subiscono modificazioni fisiologiche e patologiche (degenerazione grassa, per es.) legate alla fagocitosi, alla digestione e distruzione del materiale fagocitato, cui sono associati notevoli cangiamenti nel metabolismo (per i leucociti, v. Rossi e Zatti, 1966), alla trasformazione e, comunque, veicolazione del materiale antigene (dai monociti attivati) verso le cellule immuno-competenti; in queste ultime poi si determina la trasformazione ‛blastica' che è la condizione della produzione anticorpale (vedi immunità). Plasmacellule in attività immunopoietica esaustiva possono trattenere inclusi globulinici concentrati in forme paracristalline, quali i già ricordati corpi di Russell. In alcune reazioni di ipersensibilità (a base immunologica cellulomediata) monociti, linfociti e altre cellule subiscono fenomeni regressivi simili a quelli cui si è accennato come lesione da incontro antigene-anticorpo-complemento.
M. Infiltrazioni endogene. Abbiamo già parlato (vedi atrofia) dei pigmenti da usura endocellulari che si possono accumulare nell'apparato lisosomiale-autofagosomiale. Pigmenti endogeni dell'organismo possono però depositarsi entro cellule diverse da quelle in cui o per cui si sono formati: pigmenti di origine ematica dopo abbondanti emorragie, come l'ematina e l'emosiderina, possono essere assunti da varie e distanti cellule dell'organismo, ma in particolare da quelle fagocitanti o con attiva pinocitosi; così avviene per le porfirine e per i pigmenti biliari (ittero). Pigmenti emosiderinici (come gli altri) si trovano allogati nel sistema lisosomiale e di qui possono venir rilasciati, con conseguente possibilità di riutilizzazione del ferro. Importante è il fenomeno della infiltrazione calcarea o calcificazione di cellule da parte di ioni calcio e fosfati del mezzo interno, che si verifica di solito in cellule gravemente danneggiate (alterazioni mitocondriali con notevole rigonfiamento e accumulo di fosfati di calcio) o, più spesso, nelle cellule necrotiche (v. sotto, punto P; v. fig. 22).
N. Trasformazione neoplastica. A parte la frequenza già ricordata nelle cellule neoplastiche di gravi alterazioni genomiche e di mitosi abnormi, che sono da considerarsi piuttosto conseguenze che fattori casuali della cancerogenesi, non vi è dubbio che la trasformazione neoplastica rappresenta un evento di squisita patologia cellulare. Intendiamo qui trasformazione in senso lato, comprendente cioè anche quella benigna (ossia relativa ai tumori non maligni), ove peraltro quelle aberrazioni dell'apparato nucleare non sono di regola osservate. Siccome in definitiva non si posseggono appariscenti criteri assolutamente discriminativi, né morfologici né biochimici, fra cellule normali e cellule neoplastiche, al di fuori di quelli legati al fatto e ai modi di crescita e ai rapporti intercellulari, non sembra il caso di insistere sulla patologia della cellula neoplastica, la quale è appunto fenomeno secondario, anche se molto frequente per le condizioni stesse di crescita, e ripete comprensibilmente tutti i quadri già descritti come patologia cellulare delle cellule non neoplastiche.
In definitiva, dunque, l'essere ‛patologico' della cellula neoplastica, almeno per quella a carattere maligno, consiste in una rottura del controllo moltiplicativo, quale normalmente in parte è realizzato all'interno delle cellule stesse, in parte è il risultato di rapporti intercellulari. Tale rottura della omeostasi ontogenetica non è del tutto irreversibile per i vari cloni cellulari che dilagheranno lungo la proliferazione tumorale, ma è tendenzialmente permanente in alcuni di essi, cui è appunto legata la malignità.
L'osservazione morfologica della oncogenesi spontanea e di quella sperimentale, la possibilità di osservare in coltura cellule neoplastiche e, anzi, di trasformare in vitro cellule normali in cellule neoplastiche (mediante subcolture prolungate, immissione di virus, ecc.) ha permesso, almeno, di definire meglio questa trasformazione. Essa consiste: 1) in una graduale o improvvisa perdita dei rapporti epitelio-stromali rispetto all'equilibrio dinamico raggiunto nella ontogenesi normale: tali rapporti rimangono pressoché inalterati e solo deformati nei tumori benigni, vengono rotti nella trasformazione maligna (v. Pinkus, 1972); 2) in una diminuzione della coesione fra cellula e cellula (per le cellule epiteliali), in un allargamento della membrana basale e in una sua maggiore permeabilità, talché micropseudopodi cellulari possono penetrarla e spingersi all'esterno (microinvasività della cellula preneoplastica, v. Sugar, 1972); nelle colture di cellule tumorali questa tendenziale indipendenza cellulare si traduce nella soppressione della inibizione da contatto che invece dimostrano le cellule normali, le quali, quando col crescere della popolazione vengono a contatto, bloccano i loro movimenti migratori; con questi fenomeni riguardanti le membrane cellulari, le quali perdono la capacità di legare ioni Ca++ e divengono meno elettronegative e si disgiungono, la trasformazione neoplastica ci appare come legata o associata a una patologia delle giunzioni; indipendentemente dal problema della primarietà o secondarietà di queste trasformazioni, appare però evidente la loro importanza nel determinismo delle metastasi; 3) in una diminuzione dell'effetto densità sulla proliferazione cellulare: in coltura, il raggiungimento di una certa densità cellulare esercita un'azione inibitrice sul ciclo riproduttivo, e l'inibizione anche qui può essere da contatto e non da fattori diffusibili; 4) nella creazione di stipiti cellulari stabili a capacità moltiplicativa indefinita (v. Pontèn, 1971); 5) nella perdita o nell'acquisto di caratteristiche antigeniche rispetto alle cellule normali, a parte quelle portate da eventuali virioni: è questa una strada molto interessante per comprendere la trasformazione neoplastica (v. Weiss, 1971).
A questo punto occorre però definire brevemente il concetto di ‛atipia', così utile nella distinzione diagnostica dei tumori maligni da quelli benigni e dai tessuti normali; per quanto detto sopra, esiste una ‛atipia cellulare' e una ‛atipia sovracellulare' (delle costruzioni sovracelluiari), ambedue conseguenze della malignità, ma mentre la prima può mancare in cellule veramente maligne (è piuttosto una ‛patologia' di esse), l'altra è più genuina espressione della aberrazione ontogenetica, intercellulare e intertessutale che caratterizza la malignità.
O. Patologia delle sostanze intercellulari. Per quanto è stato detto nel capitolo introduttivo, la patologia tessutale e organismica non è solo descritta né solo prodotta dalle alterazioni delle singole cellule, ma anche da quelle delle sostanze intercellulari (dalle membrane basali alle strutture connettivali) e da quelle del mezzo interno. D'altronde la stessa vita delle cellule dipende da questo interstizio con i fluidi che lo percorrono, che reciprocamente le cellule, anche distrettualmente, concorrono a costruire e a determinare funzionalmente.
Si ripetono quindi i fenomeni di ipo- e ipertrofia o ipo- e iperplasia, che qui sarà collagena e interstiziale, spesso in qualche modo coerente (ipotrofia o ipertrofia generali) o all'opposto sostitutiva, nel senso che alla regressione di cellule parenchimali può far riscontro una iperplasia cellulare e collagena del connettivo interstiziale e adiposo (adiposi sostitutiva, come nella fisiologica senescenza del midollo osseo o del timo, oppure nelle miocardiopatie o miopatie con distruzione di fibre muscolari). Può prevalere, alla fine, l'iperplasia delle fibre collagene rispetto alla popolazione cellulare, come avviene nelle cicatrici (riparative, esito di infiammazioni, ecc.). Può anche aversi un aumento abnorme della sostanza fondamentale mucopolisaccaridica (ground substance) come nelle malattie vascolari, in certe neoplasie, in malattie congenite quali le già ricordate tesaurismosi mucopolisaccaridiche. Il complesso interstiziale, fibre e sostanza fondamentale, può subire facilmente anche fenomeni di metaplasia (oltre che la trasformazione neoplastica), di cui il più frequente è quello della metaplasia ossea (ossificazioni patologiche eterotipiche).
I fenomeni degenerativi o infiltrativi dell'interstizio sono molteplici: ialinosi, degenerazione amiloide, degenerazione mucoide ed elastotica, infiltrazione grassa-colesterolica, infiltrazione calcarea, infiltrazione di pigmenti o materiali inorganici esogeni ed endogeni.
1. Ialinosi. È una evoluzione patologica, molto tipica dei connettivi (e dei muscoli lisci) dei piccoli vasi, riscontrabile nella arteriosclerosi, nelle glomerulopatie ecc., per cui queste strutture risultano omogenee per imbibizione di una sostanza acidofila e trasparente al microscopio ottico, ma elettron-densa al microscopio elettronico. A questo tipo di alterazione corrisponde anche l'ispessimento ialino delle membrane basali dei glomeruli in alcune gravi nefropatie (v. fig. 23).
2. Degenerazione amiloide. Si tratta di una inifitrazione prevalentemente interstiziale di un complesso mucopolisaccaridico-proteico (per pinocitosi o fagocitosi esso può ritrovarsi anche dentro cellule); quando l'amiloide è secondaria ad altri e diversi stati morbosi dell'organismo, è generalmente indice di estrema gravità. Ha una struttura submicroscopica fibrillare e risulta birifrangente alla luce polarizzata dopo alcuni trattamenti; presenta anche una colorabilità istochimica abbastanza caratteristica, legata alla presenza dei mucopolisaccaridi (v. fig. 24). Si verifica negli stati iperimmuni, nelle suppurazioni e altre flogosi croniche e, il che è equivalente per un certo verso, a seguito di riassorbimento (o somministrazione) prolungato di materiali proteici anche autologhi, ma denaturati. È considerata o un aspetto della patologia del sistema globulinopoietico combinato con un'abnorme produzione di mucoproteine nel tessuto interstiziale, oppure come soltanto dovuta a questa deviazione puramente interstiziale, mesenchimale; comunque sia, il materiale amiloide è riassorbito nel sangue e di qui filtra (onde il termine di infiltrazione) attraverso i vasi in diverse altre sedi, amalgamando e distruggendo le cellule parenchimali (fegato, rene, milza, e altri organi). L'amiloide, o una sostanza simil-amiloide, è riproducibile sperimentalmente mediante introduzione parenchimale prolungata di sostanze proteiche (anche di origine batterica) incongrue e stimolanti l'immunopoiesi.
3. Degenerazione mucoide ed elastotica. Consistente nella abnorme presenza di mucopolisaccaridi (normali o meno) della sostanza fondamentale, onde la reattività istochimica del connettivo vira nettamente verso la basofilia (per i vari acidi mucoidinici), si trova nel teatro di alcune flogosi, nell'interstizio di tumori, anche benigni, nelle lesioni precancerose.
4. Infiltrazione grassa-colesterolica. Riscontrabile prevalentemente nel tessuto connettivo dei vasi sanguiferi arteriosi nella arteriosclerosi; all'infiltrazione grassa-colesterolica può seguire quella calcarea.
5. Infiltrazione calcarea. È molto frequente nelle pareti vasali arteriosclerotiche, ma è frequente anche in tutti i tessuti necrotici ove per accumulo di prodotti fosforilati di una glicolisi pre- o post-mortale si liberano (a opera di fosfatasi) ioni PO4-- che fissano i Ca++ circolanti in idrossi-apatite [Ca10(PO4)6(OH)2] (v. sotto, punto P, Necrosi); ma si possono avere anche calcificazioni di tessuti non necrotici né degenerati (calcinosi), come nel caso della iperincrezione paratiroidea per mobilizzazione di calcio dalle ossa.
6. Infiltrazione di pigmenti esogeni ed endogeni. Una fagocitosi abbondante di materiali organici o inorganici (derivati emoglobinici, polveri inerti di carbone, composti calcarei o silicici) può concludersi, specie in questi ultimi casi ove una digestione intra- ed extracellulare è praticamente impossibile, in un riversamento nello spazio extracellulare del materiale fagocitato; questo diviene un corpo estraneo che cronicamente stimola l'iperplasia collagena e la sua degenerazione ialina (noduli silicotici, per es.).
P. Necrosi. Sui meccanismi che si concludono con la morte cellulare (si è già accennato ad alcuni rilievi critici) sarà detto nel capitolo seguente. Qui verranno succintamente ricordate le cause spontanee di necrosi e i quadri che ne risultano.
Vi è una morte della cellula senza necrosi: quando si sottopongono le cellule all'azione di un sistema ‛fissativo' essenziale per la microscopia ottica ed elettronica, si ha (o si dovrebhe avere) tutto l'opposto che la perdita di struttura, anzi i mezzi (soluzioni) fissativi intanto sono pregevoli in quanto non deformano troppo le strutture, ne impediscono il decadere necrobiotico (annullamento di ogni azione enzimatica), permettono, semmai, di mettere meglio in evidenza determinate strutture, conservano i prodotti metabolici (espressioni delle attività enzimatiche) che si siano verificati prima della fissazione (sono, questi ultimi, i metodi citochimici e istochimici).
All'opposto, una cellula necrotica ha come caratteristica quella di aver perduto tutte o quasi le sue strutture e di apparire pressoché omogenea al microscopio ottico e in buona misura anche a quello elettronico. Ciò vale anche per la necrosi che investe le sostanze intercellulari, con la sola osservazione aggiuntiva che in questo caso l'omogeneizzazione può anche essere l'oggetto di una sovrabbondanza di materiale apparentemente anisto (al microscopio ottico) che amalgama e cancella le sottostanti strutture quali sono i fasci di fibrille collagene. È il caso, in sede del tutto normale, della formazione della sostanza fondamentale della cartilagine ialina, ove i normali mucopolisaccaridi amalgamano e apparentemente cancellano, con la loro basofilia, le fibre collagene acidofile che pur sono il fondo strutturale del tessuto cartilagineo.
La perdita delle strutture è un fenomeno che si svolge dunque nel tempo, e nella misura in cui esso è determinato dai meccanismi stessi della fisiologia cellulare, oltre un certo punto di deviazione patologica divenuta irreversibile, è un processo che può chiamarsi ‛necrobiotico', per distinguerlo dalla necrosi subitanea prodotta da cause esogene massimali quali il calore eccessivo (ustione), gli acidi o le basi forti, le scariche elettriche, verso le quali cause non vi è reattività biologica possibile.
Tra le cause di morte cellulare, quale terminalmente si raggiunge attraverso il comune processo necrobiotico, citiamo: a) la mancata irrorazione sanguifera (necrosi ischemica, infarto), cioè la soppressione dell'apporto dell'ossigeno e dei metaboliti (v. fig. 25); b) fattori fisici quali l'agitazione termica eccessiva, ma non fino al livello di ustione, e all'opposto il congelamento che produca cristalli di H2O meccanicamente dirompenti le strutture cellulari e la membrana esterna; lo spostamento ionico da corrente elettrica; l'energia radiante (fotoni UV o ionizzanti); c) fattori chimici, quali sali di metalli pesanti (Hg, Cd, Pb, Ag, W., ecc.) che si legano alle proteine delle membrane e ad altre strutture interne alle cellule; composti organici inibitori più o meno specifici di determinate funzioni enzimatiche o che si pongono come competitori metabolici (falsi metaboliti); d) fattori parassitari, quale l'azione citopatica dei virus, le eso- ed endotossine batteriche; e) fattori infiammatori e immunitari, quale l'arrivo di anticorpi specifici o l'accumulo all'esterno della cellula di complessi antigene-anticorpo comunque prodotti, favorito dal fatale intervento dei fattori complementari del siero circolante, i quali determinano la definitiva lisi e morte cellulare.
3. La riproduzione sperimentale analitica di vari aspetti della patologia della cellula
Con l'introduzione nella pratica sperimentale di sostanze di cui si conosca l'azione specifica su determinate strutture e attività enzimatiche cellulari, e con l'ausilio del riscontro morfologico offerto dalla microscopia elettronica (e da altri mezzi fisici di indagine strutturale), si è potuto creare una patologia sperimentale spesso ‛artificiale' nei confronti dell'interesse interpretativo medico, ma molto utile sia come ‛modelli' patogenetici a sé stanti, sia come ‛simulazioni eziologiche' portanti a processi patogenetici in tutto eguali o comparabili a quelli della patologia spontanea, che è costretta a ricalcare strade obbligate e comuni (v. cap. 2, § a).
A. Concetto di lesione biochimica. Nel 1931 R. A. Peters, in un lavoro in collaborazione con N. Gavrilescu, introdusse l'espressione di ‛lesione biochimica', mentre era intento allo studio della ragione dei sintomi della avitaminosi B1 (tiamina) nel piccione (per es. l'opistotono e i fatti convulsivi): fu dimostrato che questi dipendevano dalla incapacità a utilizzare ulteriormente il piruvato (che deriva in prevalenza dalla glicolisi) in quanto la tiamina, fosforilata, rappresenta il coenzima di una ossido-decarbossilasi piruvica, un enzima che appunto rende possibile l'ingresso dei prodotti conseguenti nel ciclo di Krebs. Così si potevano far risalire i sintomi patologici a una precisa e localizzabile interruzione di una singola operazione metabolica, anello di un ciclo, che poi è naturalmente interconnesso con altri. Una tale interruzione (lesione biochimica) ha come primo effetto, evidentemente, l'accumulo di metaboliti a monte e la carenza operativa a valle del vettore biochimico. Ma il concetto di lesione biochimica si perfezionò ulteriormente dal punto di vista sperimentale quando via via si scoprirono le molte sostanze che riescono a bloccare una determinata operazione biochimica o per competizione sugli enzimi (antimetaboliti), o per effetto deviante o tossico specificamente mirato su un punto del ciclo metabolico. Fu lo stesso Peters (1947-1949) che, per esempio, scoprì l'azione del fluoroacetato, di per sé poco tossico, ma in grado di obbligare le cellule a produrre una sintesi per loro letale (v. Peters, 1963): quella di un fluorocitrato (anziché del normale citrato), il quale è un forte inibitore della aconitasi (o aconitoanidrasi o aconitoidratasi), enzima che converte, nel ciclo degli acidi tricarbossilici, il citrato in iso-citrato. Tale sintesi letale è possibile, dunque, per una relativa aspecificità dell'enzima implicato. Si accumula così nei mitocondri un'abnorme quantità di citrato, con conseguenze sintomatologiche cospicue e in gran parte ancora da delucidare, ma legate al blocco di quel ciclo (quindi con analogie con la deficienza di tiamina).
Come si vede, il concetto di lesione biochimica implicitamente sembra superare e capovolgere quello vecchio virchowiano di patologia cellulare, in quanto si può intendere come lesione ubiquitaria per ogni e qualsiasi cellula la cui fisiologia dipenda dall'operazione biochimica disturbata (e quindi di tutte le cellule qualora si tratti di un ciclo biochimico fondamentale). Tuttavia molti fattori accessori, la permeabilità della membrana, l'assetto enzimatico intramitocondriale, capacità adattative differenziali, rendono diverso il danno secondo il tipo di cellula. Ma, soprattutto oggi, sappiamo che in molti casi la lesione biochimica non può disgiungersi da una lesione strutturale, come è necessariamente vero l'opposto, una volta che il termine di ‛struttura' da un'accezione soltanto morfologica passi a quella veramente reale includente le configurazioni molecolari e intermolecolari.
B. Alterazioni della membrana cellulare e delle giunzioni intercellulari (v. Allison, 1968; v. LaVia e Hill jr., 1971). Sostanze che leghino fortemente il Ca++ (come il chelante acido etilendiammino-tetra-acetico, EDTA) e lo asportino quindi dai legami con la membrana cellulare determinano o favoriscono la separazione delle cellule; lo stesso avviene per lieve tripsinizzazione, in quanto cellule normali tripsinizzate perdono un componente proteico della membrana e divengono disponibili all'agglutinazione da Concanavalina A (sostanza proteica che si lega fortemente ai glucidi), così come lo sono spontaneamente le cellule tumorali. Ma la rimozione di una certa quantità di Ca++ dalla superficie cellulare finisce per produrre danni che investono la permeabilità e la funzione attiva di trasporto (v. sotto), i quali si concludono quasi inevitabilmente con la morte della cellula. Anche le alterazioni o la soppressione dei fosfolipidi (a opera di anestetici, fosfolipasi, detergenti) della membrana conduce agli stessi effetti con notevoli modificazioni configurazionali (figure mieliniche) della membrana stessa (v. Trump e altri, 1970). Analogamente avviene per tutti i mezzi che realizzano una perossidazione dei lipidi, quali la avitaminosi E o l'irradiazione ultravioletta.
Si è già accennato alle lesioni delle membrane cellulari per l'arrivo di anticorpi specifici seguiti dall'azione litica del complemento. Modificazioni della membrana si hanno nelle infezioni da virus, sia al momento dell'ingresso delle particelle virali, sia quando quelle neoformate riabbandonano la cellula.
C. Alterazioni del controllo degli scambi ionico-acquosi e quindi del volume cellulare, degenerazione vacuolare o idropica. Lesioni primitive anche della sola membrana esterna, se menomano la capacità di contrarre legame con gli ioni Na+ o K+ e del loro trasporto attivo, provocano la rottura dell'equilibrio dinamico (anti Gibbs-Donnan) della cellula e l'avvio alla sua distruzione (v. schema della fig. 26).
D. Alterazioni dei mitocondri. Modificazioni fisiologiche volumetriche e strutturali dei mitocondri sono state studiate anche in vitro su questi organelli isolati e posti in condizioni diverse della loro attività metabolico-produttrice di energia (v. anche Buffa e Pasquali Ronchetti, 1972). In generale si è visto che si ha contrazione e decontrazione della matnce a seconda dello stato energetico dell'organello, il quale può essere tradotto nel rapporto ATP/ADP; una diminuzione di tale rapporto si accompagna fisiologicamente a contrazione ed è espressione di un lavoro compiuto (per es., trasporto attivo), ma decontrazione (rigonfiamento di modico grado) può intervenire solo se è presente substrato (metaboliti), O2 e ADP. Pertanto, nelle stesse condizioni, mitocondri in vivo o isolati da cellule sofferenti per anossia ischemica, o avvelenati da sostanze che bloccano od ostacolano il trasporto elettronico lungo la catena respiratoria o disaccoppiano le funzioni ossidative da quelle fosforilative, non riescono a controllare questo loro equilibrio strutturale-funzionale, divengono alla fine molto rigonfi (l'integrità strutturale-funzionale della membrana interna dei mitocondri dipende dalla riproduzione di ATP) e si avviano alla distruzione; spesso presentano anche accumuli di sostanze elettrondense nella matrice. Rigonfiamenti cospicui (vedi rigonfiamento torbido della cellula) si ottengono anche per azione della tiroxina e di ioni Ca++ o per interferenza con i lipidi strutturali dei mitocondri. Negli stadi più avanzati di queste alterazioni si ha abnorme permeabilità al Ca++ e rilascio di materiali lipidici (vedi lipofanerosi).
E. Alterazioni del reticolo endoplasmatico (RE) (v. fig. 27). La natura delle membrane limitanti questo sistema canalicolare della cellula è molto simile a quella della membrana esterna (trilaminare), tuttavia alcuni autori distinguono due classi di membrane limitanti, diverse nello spessore, alle quali si attribuiscono diverse possibilità evolutive o di rapporto nei confronti di altre strutture citoplasmatiche quali i vacuoli pinocitosici, l'apparato di Golgi, i vacuoli secretivi, i lisosomi, i fagosomi, ecc. (v. Arstila e Trump, 1968). In ogni modo, la fondamentale struttura fosfolipoproteica (con associazioni glucidiche) si ripete e si ripetono i danni ottenibili per azione fosfolipasica, di quella dei detergenti, per perossidazione. In queste condizioni si separano i componenti della membrana e, fatto molto importante se si tratta di reticolo rugoso, si distaccano i relativi ribosomi, così come si possono disaggregare i poliribosomi o polisomi (v. sotto). Il reticolo endoplasmatico liscio, così come quello rugoso, risponde a queste noxae in modo abbastanza monotono, che è quello della dilatazione delle cisterne e dell'accumulo in esse di materiale, forse identificabile in glicoproteine.
Una semplice rimozione dei fosfolipidi da parte della fosfolipasi C del Clostridium welchii altera, non solo morfologicamente (al microscopio elettronico), la struttura delle membrane del reticolo endoplasmatico, ma, ovviamente, anche molte delle attività enzimatiche associate; è importante la constatazione che si può fino a un certo punto (il che accade in qualche misura anche con i mitocondri o loro frammenti) ripristinare la configurazione strutturale e quelle attività risospendendo le membrane isolate, e così alterate, insieme ai competenti fosfolipidi.
La compagine del reticolo endoplasmatico può aumentare notevolmente come quantità e variare qualitativamente nelle cellule tutte le volte che si determini sperimentalmente uno stimolo alle attività metaboliche di loro competenza come quelle conducenti, nel fegato in particolare, alla detossificazione con adattamento enzimatico (cioè con incremento adattativo di determinati sistemi enzimatici); ciò si verifica anche verso sostanze naturali per l'organismo come gli ormoni steroidei. Nel muscolo, come si è visto, la denervazione che produce ipoplasia del materiale fibrillare contrattile determina, almeno all'inizio, iperplasia del reticolo endoplasmatico (sarcoplasmatico).
F. Alterazioni dei lisosomi. Questi organelli, circondati dalla loro membrana, rappresentano una raccolta di enzimi, in generale acido-idrolasici (fosfatasi, proteasi, peptidasi, amilasi, ecc.), che sono attivati solo dall'ingresso dei relativi substrati, che dipende dalla permeabilità della membrana e dalla sua integrità. Come si vede nello schema della fig. 28, un danno grave di questa membrana (cui non è estranea la disponibilità energetica o di ATP) ne determina la disgregazione e la fuoriuscita degli enzimi all'esterno con autodigestione (o digestione intracellulare) del citoplasma e delle sue strutture (necrosi, lisi). Questa rottura della barriera lisosomiale può avvenire dentro vacuoli di fagocitosi (fagosomi) e determinare la digestione intracellulare dell'oggetto fagocitato, il quale, come si è già ricordato, può essere anche un insieme di strutture cellulari deteriorate (autofagosomi). Gli enzimi intralisosomiali, come possono essere difettosi o mancare per ragioni genetiche (vedi, per es., le tesaurismosi genetiche), così possono essere indotti (accresciuti) per stimolazione da parte dei particolari relativi substrati.
G. Alterazioni del sistema ribosomiale e dell'RNA citoplasmatico. Sostanze come il tetracloruro di carbonio (CCl4) inducono (in specie nel fegato) una perossidazione delle membrane del reticolo rugoso con distacco dei ribosomi e anche una disgregazione dei poliribosomi: questa è la prima lesione nella steatosi sperimentale da CCl4 (e da suoi derivati). In passato si riteneva che l'azione primitiva di tali veleni si esercitasse a livello dei mitocondri, ma si è poi dimostrato che questi vengono compromessi secondariamente dai perossidi così formati; con ciò si ha una doppia sorgente di menomazione delle sintesi proteiche, quella del meccanismo stesso delle sintesi, e quella del rifornimento di energia (v. schema della fig. 29). Altre sostanze agiscono in modo analogo, come la puromicina (ma non la actinomicina D, che blocca la sintesi di mRNA); altre, come l'etionina, arrivano allo stesso risultato sottraendo adenina per la sintesi di mRNA; altre ancora, come l'acido orotico, possono ostacolare la coniugazione dei trigliceridi con le proteine e il rilascio delle lipoproteine nel plasma; altre infine, come l'acetilnitrosammina, estendono la loro azione fino alla necrosi delle cellule epatiche. Così molti esperimenti offrono varie possibilità interpretative della steatosi epatica (ne manca purtroppo una univoca per la steatosi da alcool), della necrosi e della cirrosi conseguente.
H. Influenze sulla differenziazione e sulle strutture differenziate. Ne citiamo alcune come esempi: l'avitaminosi E produce per meccanismi non ancora molto noti, ma legati quasi certamente a fenomeni perossidativi, gravi alterazioni necrotiche primarie o secondarie del mioplasma delle fibre muscolari striate; in modo analogo agiscono un eccesso di corticosteroidi (cortisone, nonostante la sua azione stabilizzatrice sui lisosomi: ma la necrosi e lisi delle fibre muscolari scheletriche si svolge con meccanismo assai probabilmente, almeno all'inizio, extralisosomiale), oppure sostanze farmacologicamente attive come il plasmocid e altri derivati chinolinici.
Molto studiate di recente sono le alterazioni regressive della cellula nervosa, culminanti in cromatolisi, che però è espressione di lavoro ricostitutivo delle strutture differenziate alterate e distrutte, indotte da composti organofosforici (alchil- o aril-fosfati), alcuni dei quali usati, è noto, come anticrittogamici; si è però osservato che effetti simili sono determinati da varie altre sostanze a struttura molto diversa (v. Cavanagh, 1969). Altre azioni neurotossiche, con fenomeni di demielinizzazione sia sui nervi periferici sia sulle strutture centrali, sono esercitate dall'idrazide dell'acido isonicotinico (isoniazide, usato come chemioterapico contro la tubercolosi) e da un altro antimetabolita della nicotinammide, la 6-amminonicotinammide (6-AN), che è paralizzante: quest'ultima produce gravi lesioni e distruzioni di alcune cellule nervose, e forse anche di nevroglia del midollo spinale, inducendo una ‛sintesi letale' di coenzimi piridin-nucleotidici ‛sbagliati', cioè contenenti 6-AN anziché la normale nicotinammide (v. Herken, 1971).
Vi sono infine alcuni composti, di cui un prototipo si trova nella specie di pisello Latyrus odoratus, i quali ostacolano la maturazione del collagene (ammino-acetonitrile, β-ammino-propionitrile, ad azione inibente sui legami in- tra- e intermolecolari del tropocollagene) con conseguenze varie nei riguardi della formazione, della struttura e della consistenza dello scheletro e in generale dei tessuti fibrosi: in senso opposto, queste sostanze possono contrastare le fibrosi patologiche (sclerosi, silicosi, ecc.) (v. Levene, 1971).
I. Alterazioni del nucleo e del nucleolo. Numerose sostanze cancerogene producono lesioni strutturali del nucleo, ma soprattutto del nucleolo. Egualmente si può dire degli agenti fisici e degli antimetaboliti che interferiscono con la sintesi di RNA da parte del nucleolo. Molto studiata di recente è la lesione nucleolare nel fegato dovuta al polipeptide tossico α-amanitina, prodotto dal fungo Amanita phalloides (v. Fiume e Laschi, 1965); anche tale polipeptide interferisce sulla RNA polimerasi in condizioni di alta forza ionica. (Per questo e per altri danni sperimentali sull'apparato nucleare-nucleolare, v. Stirpe e Novello, 1971).
L. Critica dei meccanismi terminali culminanti nella necrobiosi sperimentale. Abbiamo cercato, durante questa trattazione, di mettere in luce come la patologia della cellula non sia solo l'effetto primario della noxa, bensì consista nella serie delle reazioni e controreazioni dovute allo spostamento dell'omeostasi cellulare. Ciò, si è detto, è la condizione stessa della reversibilità del danno, almeno fino al punto, che spesso è comune a molte cellule, in cui si innescano processi irreversibili di degradazione strutturale-funzionale. Ora, con l'uso dei suddetti agenti lesivi noti si è potuto meglio valutare sperimentalmente quando e per quali motivi un processo alterativo non è più compensabile. Si è potuto così constatare che i fenomeni degenerativi non sono mai un passivo subire da parte della cellula del danno primariamente inflittole, ma rappresentano invece una serie di meccanismi che realizzano livelli diversi di omeostasi. Si afferma anzi da alcuni (v. Farber e altri, 1971) che persino quando la noxa ‛naturale' è delle più gravi, come nell'assoluta ischemia, si possono osservare, all'inizio, recuperi di funzione (per es. mitocondriale), prima dell'innesco della degradazione irreversibile. Questi autori sostengono anche che la stessa attivazione dei lisosomi e il rilascio di enzimi degradativi nel citoplasma o in grossi vacuoli autofagici rappresenterebbe piuttosto una conseguenza della morte cellulare anziché la causa di essa (suicidio cellulare).
Producendo specifiche lesioni biochimiche, per esempio nelle attività connesse con la sintesi delle proteine, si è potuto dissociare l'inibizione della proliferazione dalla morte cellulare. Un antibiotico glutarimidico, la cicloheximide (prodotto dallo Streptomyces griseus), inibisce la sintesi proteica attraverso il sistema ribosomiale del citoplasma fondamentale, mentre lascia indisturbata la sintesi proteica mitocondriale (così come non influisce su quella dei batteri). Si ottengono così cellule, per esempio quelle dei fondi ghiandolari intestinali che normalmente proliferano per il continuo rimpiazzo, le quali interrompono questa loro importante funzione e retroagiscono inibendo la sintesi di DNA (cioè il ciclo mitotico), ma sopravvivono. Tali risultati, paragonati con quelli ottenibili con tossici atti a inibire la sintesi proteica attraverso l'inibizione primaria del DNA, hanno fatto arrivare alla conclusione quasi sorprendente che l'inibizione del DNA associata a quella della produzione di qualche proteina è meno lesiva della sola inibizione del DNA. Provando tutte le ipotesi possibili per una spiegazione di tale effetto, Farber e altri (v., 1971) hanno trovato infatti che l'inibitore delle sintesi proteiche citoplasmatiche, la cicloheximide, è capace anche di proteggere le cellule intestinali dalle lesioni mortifere da azioni dirette sul DNA (inibitori specifici, agenti alchilanti, radiazioni); tale effetto non si ha invece per altri tipi di cellule come quelle del sistema linfatico dello stesso intestino, ove anzi i due interventi si possono sommare. Per converso, l'azione protettiva sulle cellule epiteliali dell'intestino non è esercitata solo dalla cicloheximide, ma anche da altri inibitori delle sintesi proteiche citoplasmatiche, con struttura del tutto diversa dalla cicloheximide (impiegati per escludere ogni effetto diretto o diverso da quello inibitorio). Viene proposto il seguente
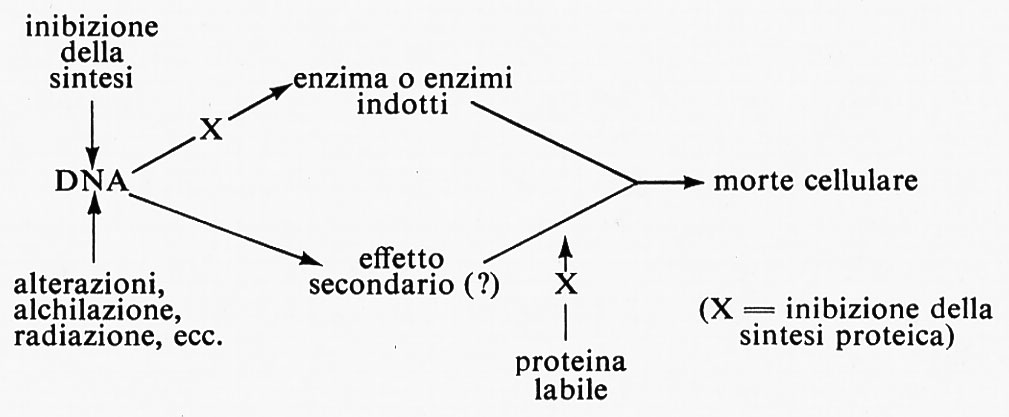
che rappresenta due ipotesi interpretative, una che presuppone che l'interferenza con la duplicazione del DNA provochi l'induzione di uno o più enzimi la cui operatività porti ad alterazioni irreversibili della cellula, l'altra che considera invece la possibile normale presenza di una proteina enzimatica, a vita media molto breve, alterante sub-strati (macromolecolari) essenziali per la vita cellulare. È naturalmente da aggiungere che se - come sembra giusto - consideriamo gli eventi patologici delle cellule come risposte, variabili nel tempo quantitativamente e qualitativamente, della organizzazione cibernetica della cellula, essi dovranno essere necessariamente diversi a seconda del tipo di cellula, almeno all'inizio e fin che non sia raggiunto il punto di irreversibilità, che però sarà anche questo vario da cellula a cellula e da stato funzionale a stato funzionale. La monotonia delle risposte, così facile da rilevare, è verosimilmente dovuta al fatto che fino a oggi abbiamo potuto apprezzare solo i fenomeni terminali di una complessa catena, e probabilmente molti di questi erano e sono mere conseguenze di una morte già avvenuta.
Del resto, sappiamo e in parte è stato anche qui ricordato, che già in condizioni fisiologiche - come quella di alcuni tipi di secrezione cellulare coinvolgenti la distruzione di buona parte della cellula la quale rimane inclusa nel secreto stesso, o quella dei processi di involuzione periodica fisiologica, per esempio il passaggio di una ghiandola mammaria secernente alla sua fase quiescente - si verificano estesi processi distruttivi che, se anche di per sé non sempre reversibili, non coincidono però con la morte cellulare, poiché la reversibilità consiste nella riproduzione delle strutture distrutte a opera di quelle residue.
4. La rigenerazione
In termini puramente di biologia della cellula, la rigenerazione consiste appunto nella capacità omeostatica della riproduzione delle strutture endocellulari deteriorate e anche trasformate nel lavoro complesso della vita cellulare e dei suoi rapporti con l'esterno: per esempio, riproduzione dei mitocondri o dei plastidi alterati o distrutti per autofagia, ricostituzione delle membrane, ecc.
Il termine rigenerazione è, però, storicamente noto per descrivere la capacità di ricostituirsi di una o più unità so- vracellulari variamente coordinate. Alcuni anzi sostengono che la vera rigenerazione sarebbe quella complessa di un intero arto di un animale (quale può avvenire in un anfibio urodelo), ricominciando a imbastire la nuova crescita con la riproduzione del blastema primitivo indifferenziato. Una rigenerazione di singoli tessuti è possibile, o perché si tratta di una popolazione cellulare fisiologicamente in continua crescita dinamicamente equilibrata con una continua distruzione (per es., epiteli di rivestimento o ghiandolari), o perché si fa a spese di una riserva di cellule indifferenziate e quiescenti (cellule satelliti delle fibre muscolari), o, infine, per un ritorno alla moltiplicazione di cellule che risultavano ormai stabili (rigenerazione del fegato dopo epatectomia parziale).
Nel caso del muscolo, alla riattivazione delle cellule satelliti può associarsi una ‛segregazione' di una porzione nucleata di citoplasma sarcoplasmatico, la quale, svincolatasi allora dal resto degenerato o morto della fibra, contribuirà al fenomeno rigenerativo attraverso moltiplicazione o successiva differenziazione.
Nel caso della rigenerazione di unità sovracellulari, di tessuti o parti di organi e anche di elementi complessi (come sono le fibre muscolari, sincizi cellulari, neurodipendenti, o neuromioni), sono assai interessanti da studiare le modalità d'informazione che sono alla base dello stimolo rigenerativo e che controllano e poi limitano il processo rigenerativo stesso. In alcuni casi, come in quello del fegato rigenerante (v. fig. 30), è verosimile che esistano canali informativi locali legati alla discontinuazione del tessuto e però anche canali informativi più generali che dipendano dalla quantità di tessuto presente in atto: si considera possibile, pertanto, l'esistenza di fattori circolanti. Nel caso dei neuromioni, quasi tutta l'attività delle due parti fisiologiche, nervosa e muscolare, è mutuamente interdipendente e anche la rigenerazione dell'una non può completarsi morfologicamente e funzionalmente senza la rigenerazione dell'altra.
bibliografia
Allison, A. C., Some effects of pharmacologically active compounds on membrane, in ‟British medical bulletin", 1968, XXIV, pp. 35-140.
Aloisi, M., Studio sistematico sul glicogeno in ‛Cavia cobaja' durante lo sviluppo, in ‟Archivio italiano di anatomia e di embriologia", 1933, XXXII, pp. 25-109.
Aloisi, M., Pierobon Bormioli, S., Schiaffino, S., Cell multiplication and histochemical changes in compensatory muscle hypertrophy, in Symposium on structure and function of normal and diseased muscle and peripheral nerve, Kazimierz 1972.
Arstila, A. U., Trump, B. F., Studies on cellular autophagocytosis. The formation of autophagic vacuoles in the liver after glucagon administration, in ‟American journal of pathology", 1968, LIII, pp. 687-733.
Buffa, P., Pasquali Ronchetti, I., Energy transduction and mitochondrial configuration, in ‟Journal of submicroscopical cytology", 1972, IV, pp. 106 ss.
Cameron, G. R. (a cura di), Pathology of the cell, Edinburgh 1952.
Cavanagh, J. B., Toxic substances and the nervous system, in ‟British medical bulletin", 1969, XXV, pp. 268-273.
Denny-Brown, D. E., Pennybacker, J. B., Fibrillation and fasciculation in voluntary muscle, in ‟Brain", 1938, LXI, pp. 311-334.
De Reuck, A. V. S., Knight, J. (a cura di), Cellular injury. Ciba foundation symposium, London 1964.
Dingle, J. T., Fell, H. B. (a cura di), Lysosomes in biology and pathology, Amsterdam 1969.
Farber, E., Verbin, R. S., Lieberman, M., Cell suicide and cell death, in Mechanisms of toxicity (a cura di W. N. Aldridge), London 1971, pp. 163-173.
Fiume, L., Laschi, R., Lesioni ultrastrutturali prodotte nelle cellule parenchimali epatiche dalla falloidina e dalla α-amanitina, in ‟Lo sperimentale", 1965, CXV, pp. 288-297).
Florey, H. W. (a cura di), General pathology, London 19704.
Gavrilescu, N., Peters, R. A., Biochemical lesions in vitamin B deficiency, in ‟The biochemical journal", 1931, XXV, pp. 1397-1409.
Gavrilescu, N., Peters, R. A., On the function of torulin. An ‛in vitro' effect of antineuritic vitamin concentrates, in ‟The biochemical journal", 1931, XXV, pp. 2150-2161.
Harris, M., Cell culture and somatic variation, New York 1964.
Helminen, H. J., Ericsson J. L. E., Studies on mammary gland involution, in ‟Journal of ultrastructure research", 1968, XXV, pp. 193-252.
Herken, H., Antimetabolic action of 6-aminonicotinamide on the pentose phosphate pathway in the brain, in Mechanisms of toxicity (a cura di W. N. Aldridge), London 1971, pp. 189-203.
Hers, H. G., Inborn lysosomal diseases, in ‟Gastroenterology", 1965, XLVIII, pp. 625-633.
King, D. W. (a cura di), Ultrastructural aspects of disease, New York 1966.
LaVia, M. F., Hill, R. B. Jr., Principles of pathobiology, New York 1971.
Levene, C. I., Effect of lathyrogenic compounds on the cross-linking of collagen and elastin ‛in vivo', in Mechanisms of toxicity (a cura di W. N. Aldridge), London 1971, pp. 67-85.
Morpurgo, B., Über Activitäts-Hypertrophie der willkürlichen Muskeln, in ‟Virchow's Archiv für pathologische Anatomie", 1897, CL, pp. 522-554.
Muscatello, U., Margreth, A., Aloisi, M., On the differential response of sarcoplasm and myoplasm to denervation in frog muscle, in ‟Journal of cell biology", 1965, XXVII, pp. 1-24.
Nowell, P. C., Chromosome changes in primary tumors, in ‟Progress in experimental tumor research", 1965, VII, pp. 83-103.
Nowell, P. C., Hungerford, D. A., Chromosome studies in human leukemia. IV. Myeloproliferative syndrome and over atypical myeloid disorders, in ‟Journal of the National Cancer Institute", 1962, XXIX, pp. 911-931.
Peters, R. A., Biochemical lesions and lethal synthesis, Oxford 1963.
Pinkus, H., Histological evidence of disturbed tissue interactions in human preneoplastic and neoplastic skin, in Tissue interactions in carcinogenesis (a cura di D. Tarin), London 1972.
Pontèn, J., Spontaneous and virus induced transformation in cell culture, Berlin 1971.
Rossi, F., Zatti, M., The mechanism of the respiratory stimulation during phagocytosis in polymorphonuclear leucocytes, in ‟Biochimica et biophysica acta", 1966, CXIII, pp. 395-397.
Stirpe, F., Novello, F., Ribonucleic acid polymerase in nuclear damage, in Mechanisms of toxicity (a cura di W. N. Aldridge), London 1971, pp. 147-161.
Sugar, J., Ultrastructural and histochemical changes during the development of cancer in various human organs, in Tissue interactions in carcinogenesis (a cura di D. Tarin), London 1972.
Temin, H. M., The protovirus hypothesis and cancer, in RNA viruses and host genome (a cura di P. Emmelot e P. Bentvelzen), Amsterdam 1972, pp. 351-363.
Trump, B. F., Duttera, S. M., Byrne, W. L., Arstila, A. U., Membrane structure. Lipid protein interactions in microsome membranes, in ‟Proceedings of the National Academy of Sciences", 1970, LXVI, pp. 433-440.
Weiss, D. W. (a cura di), Immunological parameters of host tumor relationships, New York 1971.