
Embriologia
Embriologia
SOMMARIO: 1. Lo sviluppo e l'embriologia: a) embriologia descrittiva; b) embriologia sperimentale. 2. La triade crescita-differenziamento-morfogenesi: a) crescita; b) differenziamento cellulare; c) morfogenesi. 3. Eventi cellulari: a) divisione cellulare; b) differenziamento cellulare; c) interazioni fra le cellule e movimenti cellulari. 4. Strumenti concettuali sovracellulari: a) mappatura dei territori presuntivi e mappe del destino embriologico; b) campi morfogenetici; c) gradienti e sistemi di coordinate; d) induzione. 5. Cenni conclusivi. □ Bibliografia.
1. Lo sviluppo e l'embriologia
Lo ‛sviluppo' di un organismo pluricellulare è quella serie di eventi che portano dallo ‛zigote', cioè la cellula uovo fecondata, all'individuo adulto capace di sostentarsi, interagire e riprodursi. Lo studio di questi eventi definisce il campo classico della ‛embriologia' con la sua doppia anima, descrittiva e sperimentale.
a) Embriologia descrittiva.
La ‛embriologia descrittiva' registra la serie di fenomeni biologici, per lo più macroscopici, che costituiscono lo sviluppo embrionale - e talvolta postembrionale - di un sistema biologico o di più sistemi biologici affini. I primi eventi susseguenti la fecondazione della cellula uovo sono più o meno gli stessi per tutti gli animali (v. Gilbert, 19944). Immediatamente dopo la fecondazione della cellula uovo da parte di uno spermatozoo ha inizio una fase detta di ‛segmentazione', consistente nel succedersi di un certo numero di rapide divisioni cellulari, le quali hanno l'effetto di produrre alcune decine di cellule che costituiscono il primo abbozzo dell'embrione e che sono dette ‛blastomeri'. I blastomeri sono in genere di dimensioni abbastanza ridotte rispetto allo zigote, perché le divisioni cellulari della fase di segmentazione hanno l'effetto di suddividere nei vari blastomeri l'enorme massa di citoplasma dello zigote. Il complesso dei primi blastomeri prende talvolta il nome di ‛morula', e alla fine della fase di segmentazione le poche cellule presenti si dispongono in modo da formare una sferetta cava detta ‛blastula'. La cavità centrale della blastula è detta ‛blastocele'.
A un certo momento le cellule della blastula entrano in agitazione e la sferetta nel suo complesso è sottoposta a complessi sommovimenti. Molte cellule che si trovavano all'esterno della blastula passano all'interno e alcuni straterelli di cellule si dilaminano per dar luogo a più foglietti. Questo processo fondamentale, detto ‛gastrulazione', costituisce lo stadio nel quale si delineano le basi dell'architettura del futuro individuo. È in questo stadio che negli animali più evoluti, detti triblastici, si individuano i tre foglietti embrionali fondamentali, detti ectoderma, mesoderma ed endoderma: il foglietto più esterno, l'ectoderma, darà luogo all'epidermide e al sistema nervoso; il più interno, l'endoderma, darà luogo all'epitelio di rivestimento interno del tubo digerente e delle vie respiratorie e a quello degli organi interni che sono associati all'apparato digerente, quali il pancreas e il fegato; infine, il foglietto intermedio, il mesoderma, darà luogo al tessuto connettivo, ai muscoli, al cuore e ai vasi, ai reni, alle gonadi, ai tessuti emopoietici (v. embriologia, vol. II; v. sviluppo embrionale, vol. VII).
In alcune specie sono poi molto evidenti le trasformazioni all'interno dell'ectoderma dorsale (nei Cordati) o ventrale (negli Artropodi), che culminano nella formazione del tubo neurale. Per gli embrioni di queste specie, e in particolare per i Vertebrati, si parla allora di un processo di ‛neurulazione' e della formazione di una ‛neurula'. Contemporaneamente alla formazione del tubo neurale si registra l'inizio della strutturazione degli organi principali del futuro organismo. Questa lunga fase di eventi morfogenetici più o meno cospicui prende di solito il nome di ‛organogenesi'.
L'organogenesi ha termine con la formazione di un organismo capace di vivere e di riprodursi. Le cellule che ne compongono il corpo e ne permettono il funzionamento sono dette ‛cellule somatiche' o ‛cellule della linea somatica', mentre le cellule che daranno luogo ai futuri gameti, permettendo così la sopravvivenza della specie, sono dette ‛cellule germinali' o ‛cellule della linea germinale'. La separazione fra le due linee è uno degli eventi più precoci dello sviluppo in tutte le specie, anche se il processo più manifesto, cioè la maturazione finale dei gameti, avviene di solito in età matura (v. embriologia, vol. II).
b) Embriologia sperimentale.
Al di là di questi primi stadi relativamente uniformi, le variazioni che lo sviluppo biologico può presentare nelle diverse specie sono praticamente infinite, così che di fatto non esiste un'unica embriologia descrittiva generale, ma vi sono piuttosto diverse embriologie speciali. La ‛embriologia sperimentale', invece, essendo centrata sullo studio dei meccanismi che sono alla base dei vari processi ontogenetici, presenta un aspetto più unitario e consiste in pratica nella descrizione degli effetti prodotti da manipolazioni più o meno vistose cui vengono appositamente sottoposti embrioni di uno o più sistemi biologici. Queste manipolazioni - eseguite con varie tecniche, dalle più dirompenti, come legature, cauterizzazioni o amputazioni, alle più innocue, come la colorazione con traccianti vitali - hanno lo scopo comune di turbare il corso naturale degli eventi in un sistema in via di sviluppo e di osservarne gli effetti.
Con il passare degli anni la componente sperimentale dell'embriologia ha preso sempre maggior vigore e sono notevolmente aumentati il numero e la varietà delle tecniche di intervento. In alcuni sistemi, ad esempio, lo strumento principe di questa investigazione sistematica si è rivelata l'analisi genetica, che può essere considerata una forma tutta particolare di intervento mirato sull'embrione. A seguito di questi sviluppi si è pensato che si potesse ormai parlare della nascita di una nuova disciplina, la ‛biologia dello sviluppo' (v. Slack, 19912; v. Boncinelli, 1994; v. Gilbert, 19944). Avvalendosi di una trattazione descrittiva ridotta al minimo, questa branca della biologia dovrebbe riassumere gli elementi e i principî della meccanica dello sviluppo nei vari sistemi, giungendo a enunciare, nei limiti del possibile, principî generali riguardo a come e perché gli organismi si sviluppano. Il compito è certamente immane, ma la sfida è entusiasmante: si tratta infatti dell'obiettivo più importante che la biologia di questi anni si è posta, in attesa di affrontare le problematiche poste dal cervello e dal suo rapporto con la mente.
2. La triade crescita-differenziamento-morfogenesi
È consuetudine considerare la catena di eventi indicati col termine ‛sviluppo' come potenzialmente suddivisa in tre processi considerati più elementari, e cioè la crescita, il differenziamento cellulare e la morfogenesi, o sviluppo in senso stretto (v. Boncinelli, 1994).
a) Crescita.
Il parametro forse più evidente in un organismo che si sta sviluppando è l'aumento delle sue dimensioni: questo fenomeno è spesso indicato con il termine ‛crescita'. La crescita dell'embrione consiste in gran parte nell'aumento del numero delle cellule che lo compongono. Condizione necessaria per la crescita e quindi per tutto lo sviluppo embrionale è la proliferazione cellulare, la quale deve essere controllata con precisione in ogni fase dello sviluppo. Tradizionalmente, però, questo aumento del numero delle cellule dell'embrione non viene considerato come un fattore essenziale e specifico dello sviluppo.
b) Differenziamento cellulare.
Più vicino al cuore del problema si trova invece il fenomeno del differenziamento cellulare. Con questa espressione si intende il complesso di eventi che portano le cellule di un embrione a differenziarsi fra di loro a partire da un'iniziale omogeneità. Questo processo è a buon diritto considerato della massima importanza, talché spesso si adopera la locuzione ‛differenziamento e sviluppo' come se si trattasse di una cosa sola. Tutte le cellule di un organismo adulto derivano, come abbiamo visto, da un'unica cellula progenitrice, lo zigote. I primi blastomeri derivanti dalle divisioni cellulari della fase di segmentazione sono generalmente molto simili tra di loro, mentre non c'è dubbio che nell'organismo adulto vi sono centinaia di tipi cellulari diversi. Durante tutto lo sviluppo si devono quindi essere diversificati tra di loro, cioè differenziati, vari gruppi di cellule. Il differenziamento cellulare è appunto il complesso di queste diversificazioni e i meccanismi molecolari e cellulari che ne stanno alla base costituiscono l'oggetto di studio primario della moderna embriologia molecolare e cellulare.
Il differenziamento si può idealmente scomporre in una serie di differenziamenti di linee cellulari specifiche, quali quello della linea delle cellule del sangue, quello della linea delle cellule nervose, oppure quello della linea delle cellule muscolari, o delle cartilaginee, e così via. Alcuni di questi processi differenziativi specifici possono essere studiati in laboratorio, anzi si può dire che la maggior parte delle conoscenze che si è andata accumulando negli ultimi decenni deriva da questo tipo di analisi: sarebbe quindi assurdo disconoscere l'enorme contributo che lo studio del differenziamento ha dato alla comprensione della biologia dello sviluppo e alla biologia in generale.
Questo approccio così lineare e proficuo nasconde però un'insidia: quello che si finisce per studiare spesso non è tanto il differenziamento delle cellule nei vari tipi quanto il ‛differenziamento terminale' dei singoli tipi cellulari, vale a dire l'ultimo passo che una data cellula deve compiere lungo la sua linea differenziativa. Per esempio, il passaggio a globulo rosso, o eritrocita, è l'ultima tappa lungo la linea di differenziamento eritroide delle cellule del sangue, così come il passaggio da mioblasto a miotubulo è l'atto conclusivo lungo la linea di differenziamento della cellula del muscolo scheletrico. Questi eventi differenziativi si possono seguire abbastanza facilmente, in vivo o in vitro, perché gli eventi morfologici che li accompagnano sono spesso vistosi e perché sono disponibili molti marcatori cellulari o molecolari in grado di evidenziare il trapasso da un tipo cellulare all'altro.
L'indubbio successo di tali studi non deve però far pensare che in questa maniera si stia studiando il differenziamento in generale. Esiste tutta una catena di eventi che porta la cellula, o per meglio dire una linea differenziativa di cellule, dallo stato relativamente indifferenziato di blastomero a quello quasi terminalmente differenziato di mioblasto o di eritroblasto o di linfocita pre-B. Questa catena di eventi resta ancora tutta da studiare. A tutt'oggi non c'è tuttavia motivo di pensare che i principî e i meccanismi implicati in questi eventi precoci siano diversi da quelli che stanno alla base del differenziamento terminale che porta rispettivamente l'eritroblasto a eritrocita, il mioblasto a miotubulo e il linfocita pre-B a linfocita B maturo.
Possiamo affermare che il problema dello sviluppo degli organismi pluricellulari si identifica con il problema del differenziamento cellulare? Come succede spesso per questo tipo di domande, la risposta è prima di tutto una questione di semantica. In questo caso specifico tutto dipende da cosa si intende col termine differenziamento. Alcuni biologi tendono a darne una definizione onnicomprensiva e non avrebbero quindi alcuna esitazione a rispondere affermativamente al quesito che ci siamo posti; noi preferiamo adottare una definizione meno ampia e pensare che lo sviluppo sia qualcosa di più del differenziamento cellulare, anche se quest'ultimo ne costituisce il fenomeno più cospicuo.
c) Morfogenesi.
Dalla definizione generale di differenziamento cellulare rimane escluso il concetto di disposizione spaziale delle cellule appartenenti alle varie linee cellulari, che viene spesso compreso in quello più tecnico di ‛morfogenesi', nel senso di creazione di forme, cioè di strutture biologiche complesse organizzate nello spazio e nel tempo. La disposizione spaziale delle cellule - spesso indicata con il termine inglese di pattern formation (v. Slack, 19912) - può riguardare la formazione di strutture parziali come l'ala di una farfalla, la mano di un primate o l'occhio di un cefalopode, oppure anche di strutture globali, quali il piano corporeo complessivo di un organismo come un moscerino o un anfibio.
Ci piace pensare che in una mano ci sia qualcosa di più di un complesso di tessuti che si sono differenziati nel momento giusto e nella maniera giusta. Si ha l'impressione che in una struttura complessa sia presente un altro tipo di informazione biologica, distinto da quella differenziativa o istologica. Si parla a questo proposito di ‛informazione posizionale', un concetto che, introdotto da poco più di venti anni (v. Wolpert, 1969 e 1989) per rendere meno imprecisa e sfocata la sensazione che una struttura ordinata contenga qualcosa di più di un complesso di tessuti, si è rivelato tutt'altro che evanescente; inoltre, in tempi recenti, è stato anche possibile chiarire molti aspetti del controllo genetico dell'informazione posizionale (v. sviluppo embrionale, vol. VII).
3. Eventi cellulari
Lo sviluppo nel suo complesso può essere pensato come articolato in una serie di eventi di cui è facile individuare il livello cellulare e in un'altra serie di eventi che, pur avendo la loro base in fenomeni cellulari, non sono a questi facilmente riconducibili e che noi attribuiremo a un livello sovracellulare. Possiamo parlare quindi di ‛eventi cellulari' e di ‛strumenti concettuali sovracellulari'. I primi possono essere schematizzati in vari modi, uno dei quali è riportato nella tab. I; i secondi (v. cap. 4) comprendono una serie di concetti scaturiti dagli studi condotti in anni relativamente più recenti.

Gli eventi cellulari comprendono la divisione cellulare, il differenziamento, le interazioni fra le cellule e i movimenti cellulari.
a) Divisione cellulare.
Abbiamo visto che la divisione o proliferazione cellulare è una condizione necessaria, anche se non sufficiente, per lo sviluppo di un organismo superiore. Abbiamo anche visto che subito dopo la fecondazione lo zigote comincia a dividersi, prima in due cellule, poi in quattro, poi in otto e così via, e abbiamo chiamato segmentazione questa prima fase dello sviluppo. In parecchi organismi queste prime divisioni cellulari sono molto veloci, più veloci di una divisione cellulare ordinaria. Dopo questa prima fase si osserva, in genere, un rallentamento del susseguirsi delle divisioni cellulari.
È appena il caso di ricordare che la proliferazione cellulare, con modalità differenti per le varie cellule, accompagna l'organismo per tutta la sua vita: ogni cellula, se deve dividersi, si divide secondo un determinato ritmo. Esistono infatti cellule che da un certo momento in poi non si dividono più, come ad esempio i neuroni; cellule che ordinariamente non si dividono, ma che all'occorrenza possono dividersi, come quelle del fegato; e cellule che si dividono continuamente anche nella vita adulta, come quelle dell'epidermide o i precursori delle cellule del sangue. Questo schema proliferativo deve essere rigidamente controllato. La catena di eventi che regola la ‛decisione' di dividersi o non dividersi da parte di una singola cellula è stata studiata in maniera molto approfondita negli ultimi anni. Quando qualche elemento di questa catena perde la sua capacità di intervenire nel controllo della proliferazione, le cellule che ne dipendono possono dividersi disordinatamente e dare origine a un tumore.
Va da sé che la divisione cellulare può anche assumere, durante lo sviluppo embrionale, un ruolo marcatamente morfogenetico. Nella generazione di forme complesse, infatti, può influire in modo fondamentale anche una differenza nella velocità di crescita di parti diverse di un organo o dell'intero embrione: le regioni a più alta velocità di replicazione si espanderanno a spese di altre dotate di un ridotto ritmo proliferativo, e queste ultime finiranno per costituire barriere o strozzature; e rigonfiamenti e strozzature costituiscono gli elementi sui quali si basa il riconoscimento delle forme stesse.
Consideriamo adesso i vari tipi di divisione cellulare nello sviluppo (v. fig. 2; v. Boncinelli, 1994), tenendo comunque presente il fatto che non esiste praticamente divisione cellulare che non getti il seme, palese o recondito, di un differenziamento delle cellule figlie. L'uno o l'altro dei vari tipi di divisione cellulare può prevalere in fasi diverse dello sviluppo di sistemi diversi. Come abbiamo visto, la divisione cellulare di tipo I prevale spesso nelle primissime fasi embrionali successive alla fecondazione: essa può essere definita ‛divisione cellulare per segmentazione'. Gli altri tipi di divisione subentrano in fasi successive. Ad esempio, le modalità II, III e IV sono collettivamente rappresentative di un tipo di proliferazione cellulare, la ‛proliferazione cellulare esponenziale', che caratterizza molta parte dei processi di crescita vera e propria nell'embrione e nell'organismo. Un'opzione a sé è costituita dalla modalità V, caratteristica della crescita delle cosiddette cellule staminali (stem cells): con questo termine, spesso utilizzato con significati alquanto diversi, noi intendiamo indicare cellule che possono replicarsi per un certo numero di generazioni restando essenzialmente uguali a se stesse, ma che in taluni casi possono dare origine a cellule figlie diverse che imboccano la via del differenziamento, originando a loro volta un certo numero di cellule mature diverse. L'esempio tipico è dato dai precursori delle cellule del sangue - ma lo stesso discorso vale per l'epidermide, la mucosa intestinale o i gameti maschili - le quali si rinnovano continuamente e costituiscono un fondo comune, o pool cellulare, da cui ogni tanto si distacca una cellula che inizia a specializzarsi per dar luogo a cellule differenziate di tipo linfoide, eritroide o mieloide.
Esaminiamo ora la divisione cellulare ordinaria di tipo esponenziale, la più comune e la più interessante. Se si prendono in considerazione le caratteristiche morfologiche delle due cellule figlie derivate da una divisione cellulare di questo tipo, si possono distinguere almeno tre possibilità, rappresentate nella fig. 2 come tipi II, III e IV. Nella divisione del tipo II, le due cellule figlie sono morfologicamente indistinguibili tra di loro e dalla cellula madre; nel tipo III, una è indistinguibile dalla madre mentre l'altra è diversa; infine, nel tipo IV le due cellule figlie sono entrambe diverse dalla cellula madre e diverse tra di loro. Se però si analizza accuratamente il fenotipo complessivo o il destino embriologico delle due cellule figlie, si deve concludere che la divisione di tipo II, quella apparentemente più semplice in cui si generano due cellule identiche alla cellula madre, rappresenta di fatto niente di più che un'astrazione teorica: non a caso le tre cellule in questione sono state indicate nella figura come A, A′ e A″.
Consideriamo in dettaglio questo punto. Nella fig. 2 le due cellule figlie originate da una divisione del tipo II sono rappresentate come indistinguibili tra di loro e dalla cellula madre. Quando diciamo ‛indistinguibili' intendiamo ‛indistinguibili dal punto di vista fenotipico', cioè esteriore, il che spesso significa anche dal punto di vista della biochimica cellulare. Ma non dobbiamo lasciarci ingannare. L'esperienza accumulata negli ultimi decenni ci dice che certamente le due cellule non saranno indistinguibili nel loro intimo e che prima o poi si individuerà un marcatore capace di distinguerle, sia esso macroscopico o molecolare.
A questo proposito si possono portare molti esempi. Uno dei più istruttivi è rappresentato dalle cellule dell'embrione precoce del moscerino Drosophila melanogaster, uno degli organismi più interessanti per la moderna biologia dello sviluppo (v. Lawrence, 1992). Allo stadio di blastoderma, l'embrione ha la struttura di una palla cava un po' allungata nel cui interno si trova praticamente soltanto del materiale nutritizio e la cui superficie esterna è costituita da circa 6.000 piccole cellule. Queste appaiono tutte identiche, sia all'osservazione macroscopica che a quella microscopica, eppure soltanto in alcune di esse è espresso un determinato gene, per esempio il gene fushi tarazu (ftz) (v. Gilbert, 19944). Questa espressione è evidenziabile solamente nelle cellule appartenenti a sette strisce, sette cinture trasversali non più larghe di 3-4 cellule e separate fra di loro da un intervallo (interstriscia) di 4-5 cellule: si usa parlare in questo caso di un profilo d'espressione ‛zebrato', o ‛a zebra'.
Una cellula appartenente a una delle sette strisce apparentemente non differisce dalla cellula contigua appartenente a una interstriscia, e fino a una decina di anni fa non esisteva alcuno strumento in grado di consentire la distinzione tra le due cellule. Tuttavia, il fatto che una sola delle due cellule produca la proteina di ftz è indicativo del fatto che tra esse una differenza, per quanto sottile, deve esistere. Le due cellule figlie derivanti da una divisione cellulare possono quindi essere diverse fra di loro, ma lo strumento, molecolare o macroscopico, necessario per distinguerle può non essere stato ancora scoperto e quindi non essere ancora disponibile.
Per distinguere tra di loro le due cellule figlie derivanti da una particolare divisione cellulare si può in alcuni casi cercare di identificarne il ‛destino embriologico'. Ciò può essere realizzato nella maniera più semplice mediante una ricognizione del tipo di cellule a cui queste avranno dato origine, in altre parole operando l'analisi della loro ‛discendenza cellulare' (cell lineage). In alcuni sistemi, come nel nematode Caenorhabditis elegans, un altro organismo protagonista della più recente biologia dello sviluppo, questo tipo di indagine è relativamente semplice, in quanto è sufficiente condurre per un certo tempo un'osservazione microscopica accurata: il numero totale di cellule di cui è costituito ciascun animale di questa specie, infatti, è costante e non supera il migliaio. Alcuni autori hanno così potuto fare l'inventario completo di tali cellule e del loro grado di parentela (v. Sulston e Horvitz, 1977; v. Sulston e altri, 1983). Anche se non è sempre possibile utilizzare metodologie così semplici e dirette, in molti casi si può ugualmente seguire il destino della discendenza di una determinata cellula mediante traccianti biologici, particelle materiali adsorbite o marcatori genetici.
Ritorniamo alle modalità di divisione cellulare nello sviluppo, illustrate nella fig. 2. Se è possibile seguire in una maniera o nell'altra le differenze nel fenotipo o nel destino embriologico delle varie cellule, raramente si vedrà qualcosa di simile a una divisione del tipo II, mentre più facilmente si osserveranno divisioni del tipo III o del tipo IV. Un sottotipo della modalità III (e IV) è rappresentato dalla situazione indicata come IIIa (e IVa) in cui il destino di una delle due cellule figlie è rappresentato da una ‛morte programmata', che in questo contesto si può considerare come una forma tutta particolare di fenotipo: in condizioni di sviluppo normali quella data cellula è condannata a una morte programmata, determinata geneticamente.
In chiusura di questo paragrafo sulla divisione cellulare, dobbiamo introdurre una distinzione. Non c'è dubbio che una cellula imbocchi la strada del proprio destino embriologico prevalentemente sulla base della sua ‛ascendenza cellulare', cioè sulla base della natura della cellula madre e della sua storia precedente. Se il suo destino è determinato esclusivamente o quasi esclusivamente su questa base si parla di ‛sviluppo a mosaico', mentre se l'ascendenza cellulare è solamente una delle componenti e in questa determinazione entrano in gioco anche fattori diversi si parla di ‛sviluppo di tipo regolativo'. Fra questi fattori diversi vanno annoverati in primo luogo la vicinanza fisica ad altre cellule e l'esposizione a sostanze induttrici diffusibili (v. anche sviluppo embrionale: Analisi del differenziamento, vol. VII).
b) Differenziamento cellulare.
I. Paradigmi molecolari. - Abbiamo già parlato del differenziamento come fenomeno generale; ne riparliamo adesso come fenomeno cellulare specifico. Ci stiamo riferendo in questo caso a specifici eventi differenziativi subiti da uno specifico tipo di cellule, ad esempio il differenziamento delle cellule dell'ovidotto del pollo o quello della linea differenziativa dei linfociti B. Si registra oggi una certa tendenza a utilizzare in questo caso il termine ‛differenziazione' cellulare, ma qui preferiamo continuare a utilizzare il termine ‛differenziamento'.
In ciascun caso di transizione differenziativa controllata da un tipo cellulare a un altro, analizzato sia in vivo che in vitro, è stato possibile evidenziare l'accensione e/o lo spegnimento specifico di almeno una unità genica. Ci sono d'altra parte ottime ragioni per ritenere che, salvo poche eccezioni, il patrimonio genetico sia lo stesso per tutte le cellule di uno stesso organismo. Ciò suggerisce che il meccanismo molecolare alla base del differenziamento cellulare lungo linee differenziative specifiche sia rappresentato dall'espressione differenziale di gruppi di geni. Sembra a questo punto ragionevole estendere questa conclusione e utilizzarla anche per spiegare le differenze macroscopiche fra cellule chiaramente diverse, come le cellule nervose e le cellule del fegato. Secondo questa visione, nella cellula epatica sarà attivo un certo gruppo di geni, diciamo epato-specifici, mentre tutti gli altri saranno spenti; nella cellula muscolare, viceversa, sarà attivo un altro gruppo di geni, diciamo muscolo-specifici, mentre saranno spenti tutti gli altri, compresi naturalmente quelli epato-specifici.
L'affermazione di carattere generale che il meccanismo molecolare alla base del differenziamento cellulare è rappresentato dall'espressione differenziale di gruppi di geni costituisce un po' il ‛paradigma molecolare' della moderna biologia dello sviluppo. È molto difficile apprezzare oggi tutta la portata storica di questo paradigma: occorrerebbe ricordare di quali e quante fumosità e vaghezze ha fatto giustizia e da quanti presunti dogmi ha sgombrato il campo della biologia dello sviluppo.
Un secondo paradigma della biologia dello sviluppo si è andato col tempo affiancando al primo. Lo possiamo definire come il paradigma della ‛progressività degli eventi determinativi nel differenziamento'. L'analisi istologica dimostra che già i precursori delle cellule dei diversi tessuti sono caratterizzati da differenze più o meno marcate. Precursori di linee differenziative diverse imboccano infatti abbastanza presto la via di un destino istologico specifico e tutti gli eventi successivi non fanno che spingerli più avanti su quella via. Cellule destinate a divenire tessuto muscolare sono diverse da quelle destinate a divenire epitelio intestinale, come queste sono a loro volta diverse da quelle che daranno luogo agli eritrociti, e le loro differenze aumentano con il procedere dello sviluppo. L'indagine molecolare conforta in tutto e per tutto questa conclusione. Ci possiamo raffigurare questa successione di eventi come un'accensione sequenziale di gruppi di geni specifici di quel tessuto e un concomitante spegnimento di altri. L'accensione del gene epato-specifico n + 1 non produrrebbe gli stessi effetti in una cellula in cui non fossero già stati precedentemente accesi gli altri n geni epato-specifici. Possiamo dire che le cellule dei cloni destinati a diventare epatociti vengono progressivamente ‛determinate' in direzione epato-specifica. Il termine ‛determinazione' è utilizzato per indicare una serie di processi determinativi parziali che, succedendosi nel tempo, preparano il differenziamento vero e proprio della cellula verso un fenotipo istologico definitivo. Il concetto stesso di differenziamento è legato alla possibilità di osservare una chiara trasformazione morfologica della cellula o anche soltanto una trasformazione del suo fenotipo biochimico o molecolare. Tutto ciò è preceduto però da una serie di eventi determinativi il cui effetto morfologico può essere molto difficile da mettere in risalto. Spesso, infatti, che una data cellula sia in un certo stadio già determinata in una precisa direzione istologica lo si può dedurre soltanto a posteriori, considerando i suoi successivi cambiamenti istologici o la specificità delle cellule discendenti. Si può supporre inoltre che da un certo punto in poi il processo di determinazione divenga irreversibile, anche se non è sempre facile determinare il momento esatto di questo cambiamento.
La miglior illustrazione del concetto di determinazione e del suo rapporto con il differenziamento è probabilmente fornita dallo studio dei ‛dischi imaginali' della drosofila (v. fig. 5; v. Gilbert, 19944). Una volta completata l'embriogenesi, quest'insetto passa attraverso due stili di vita, prima da larva, poi da insetto adulto. La larva rappresenta una forma di vita autonoma, pressoché svincolata da quella dell'adulto per ciò che riguarda sia l'anatomia che la fisiologia. Affondate nel corpo della larva si trovano una ventina di masserelle compatte costituite da piccole cellule tondeggianti: queste masserelle sono dette dischi imaginali - da imago, il nome latino della forma adulta - e rappresentano i primordi di molti organi e di tutte le appendici cuticolari dell'animale adulto.
Quando la larva è giunta al termine del suo ciclo, smette di nutrirsi, si ferma e si trasforma in pupa. Questo processo di ‛metamorfosi' è scatenato da un ormone specifico, detto ecdisone. All'interno dell'involucro della pupa ha allora luogo una serie di processi morfogenetici che rimodellano completamente il corpo del futuro moscerino. I tessuti larvali vengono praticamente distrutti, mentre i dischi imaginali si evaginano e le strutture da essi derivate prendono il sopravvento. Dalla coppia di dischi imaginali per l'ala si originano le ali, dalla coppia di dischi imaginali per la zampa protoracica si originano le zampe protoraciche e così via. Durante la vita larvale i dischi imaginali restano quiescenti e compiono soltanto qualche ciclo di divisione mitotica, perché nella larva è alto il livello di un secondo ormone, detto neotenina od ormone giovanile, il cui ruolo è appunto quello di prevenire la transizione dell'animale dallo stadio di larva a quello di pupa. Al momento programmato per questa transizione si verifica contemporaneamente un abbassamento del tasso di neotenina e un picco di concentrazione di ecdisone (v. ormoni negli invertebrati, vol. VI).
La morfologia complessiva dei dischi dei diversi tipi è abbastanza riconoscibile, ma le cellule che compongono un determinato disco non sono distinguibili tra di loro né da quelle di un disco di un altro tipo. In altre parole, i vari dischi sono composti di tante piccole cellule tutte uguali. Ciò nonostante ogni gruppo di cellule di ogni disco sa quello che deve fare durante la metamorfosi pupale (v. fig. 5B). Utilizzando questo sistema sono stati compiuti molti studi in un'epoca pionieristica dell'embriologia sperimentale della drosofila. Singoli dischi imaginali possono essere isolati da una larva e trapiantati in un'altra larva, così che quando quest'ultima subirà la sua metamorfosi mostrerà un'appendice soprannumeraria esattamente corrispondente al disco trapiantato: il disco dell'ala darà luogo a un'ala, il disco dell'antenna darà sempre origine a un'antenna. Se viene trapiantata solo una porzione di un disco, si osserverà la comparsa della corrispondente porzione dell'organo adulto. Possiamo quindi dire che le cellule dei vari dischi sono determinate per un loro destino embriologico, anche se questo diventerà manifesto solo al momento della metamorfosi.
Tale stato di determinazione è molto stabile: i vari dischi imaginali, infatti, possono essere subcoltivati per lunghi periodi nell'addome di moscerini adulti senza che perdano la loro specificità; in questo ambiente abnorme sono in grado di mantenersi e di crescere, ma non possono differenziarsi per l'assenza di ecdisone. Un dato disco può poi essere recuperato da questo primo ospite e trapiantato in un secondo ospite adulto, e così via per decine e decine di volte. Di tanto in tanto si può valutare lo stato di determinazione delle cellule di quel disco trapiantandone una porzione in una larva e osservando ciò che accade al momento della sua metamorfosi. Il disco originariamente destinato a formare un'ala darà un'ala anche dopo numerosi cicli di trapianto in ospiti adulti, e così per tutti gli altri dischi.
Di tanto in tanto, con il passare delle generazioni e dei cicli di trapianto, qualche disco perde la sua determinazione, o per meglio dire ne acquista una diversa. Il disco destinato a generare una zampa può produrre un'antennina, mentre il disco per l'antenna può produrre un'ala e da allora in poi produrrà un'ala. A questo fenomeno di acquisizione di una nuova determinazione da parte di un disco imaginale subcoltivato per un lungo periodo è stato dato il nome di ‛transdeterminazione' (v. fig. 5C; v. Hadorn, 1965). Anche in questo fenomeno aberrante si può tuttavia individuare un'indicazione di progressività. Gli eventi transdeterminativi non sono infatti completamente casuali: lo schema della fig. 5C mostra che esistono delle transizioni preferenziali, ad esempio quasi nessun disco transdetermina nella direzione dei dischi genitali, mentre i dischi labiali, quando transdeterminano, si trasformano quasi invariabilmente in dischi per le antenne.
Dall'analisi di questi esperimenti si possono trarre almeno tre conclusioni. In primo luogo, il patrimonio genetico delle cellule componenti i vari dischi rimane evidentemente sempre lo stesso attraverso i vari stati di determinazione, altrimenti non si può spiegare il fenomeno della transdeterminazione. Come conseguenza, la determinazione può consistere solamente nell'accensione o nello spegnimento di un gruppo di geni piuttosto che di un altro, cioè in una espressione genica differenziale. In secondo luogo, questa determinazione è molto stabile ma non totalmente irreversibile. Infine, esiste una preferenzialità nella successione degli eventi transdeterminativi. Poiché è ragionevole supporre che la transdeterminazione derivi in alcuni casi da una perdita parziale di elementi determinativi, possiamo raffigurarci questo fenomeno come una progressiva ‛sdeterminazione'. Ne possiamo concludere che la determinazione consiste in una progressione di eventi decisionali che comportano l'accensione e lo spegnimento di gruppi specifici di geni.
Consideriamo un secondo esempio concreto di determinazione, frutto di indagini molecolari più recenti. Il sogno di ogni biologo molecolare che studia lo sviluppo è quello di poter individuare un singolo gene responsabile del differenziamento lungo una specifica linea differenziativa. Ebbene, esiste un gene il cui comportamento sembra conformarsi in parte a questo sogno. Si tratta del gene myoD (v. Weintraub e altri, 1991), la cui espressione sembra necessaria e sufficiente per il differenziamento delle cellule del muscolo scheletrico. Se si forza la sua espressione in mioblasti proliferanti in coltura, si osserva che questi smettono immediatamente di replicare e cominciano a differenziarsi in cellule muscolari mature. Ma non basta: sotto la spinta della sua azione anche alcune linee di fibroblasti in coltura sono in grado di impegnarsi nel programma differenziativo che conduce alla cellula del muscolo scheletrico; non solo, ma l'espressione forzata di questo gene riesce a riconvertire a un programma di differenziamento muscolare anche cellule coltivate in vitro di linee istologicamente molto distanti, come quelle di adipociti, melanomi, epatomi, neuroblastomi, osteosarcomi e teratocarcinomi nonché colture primarie di condrociti, muscolo liscio, tappeto retinico, fibroblasti e perfino neuroni.
Per esplicare questa funzione durante lo sviluppo effettivo di un vertebrato, myoD ha bisogno dell'ausilio di altri geni, alcuni dei quali ugualmente attivi nel differenziamento della cellula muscolare: si tratta di myogenin e di altri due geni, chiamati myf-5 e mrf-4, che sembrano in grado di compiere anch'essi molte delle funzioni di myoD. I quattro geni sembrano così costituire una famiglia genica, la famiglia myoD, appunto. Il prodotto di myoD è una proteina nucleare capace di agire come fattore trascrizionale e di promuovere il differenziamento cellulare stimolando l'accensione di geni specifici della cellula del muscolo scheletrico. Una volta acceso, myoD si mantiene attivo perché il suo prodotto proteico ne stimola l'espressione promuovendo inoltre anche quella degli altri geni della famiglia myoD, così come ciascuno di essi è in grado di promuovere l'espressione di tutti gli altri e di myoD stesso. In questo modo una rete di reciproche attivazioni assicura il mantenimento di un alto livello d'espressione di proteine regolatrici della miogenesi del muscolo scheletrico. È l'esistenza di questo circuito regolativo chiuso su se stesso e capace di autorinforzo che sta alla base del comportamento straordinario del gene myoD.
Nei mioblasti proliferanti in coltura è presente il prodotto proteico di myoD, eppure queste cellule non differenziano, almeno fintantoché vengono fatte crescere in abbondanza di siero, come si fa normalmente. In queste cellule ci deve essere quindi qualcosa che esercita un controllo negativo sull'attività della proteina di myoD. Si è trovato che le cellule a questo stadio contengono alti livelli di un'altra proteina, la proteina codificata dal gene Id, così chiamato perché inibisce il differenziamento, ovviamente dei mioblasti. Questa proteina è in grado di combinarsi con quella di myoD e di inattivarla. In presenza di alte concentrazioni della proteina di Id, quindi, myoD non può svolgere il suo ruolo, a meno che non venga sovraespresso, come negli esperimenti che hanno portato alla sua identificazione. I mioblasti proliferanti in coltura possono essere indotti a differenziare togliendo loro il siero, perché la riduzione della percentuale di siero nel mezzo di coltura ha come effetto immediato un abbassamento della concentrazione del prodotto di Id in queste cellule.
Siamo evidentemente in presenza di un meccanismo regolativo che sembra fatto apposta per illustrare il concetto di determinazione progressiva. Se nelle cellule di un dato tessuto non c'è il prodotto di altri geni della famiglia o se c'è una sovrabbondanza della proteina codificata da Id, myoD, pur così potente, non può agire. Questo ovviamente per parlare soltanto dei meccanismi molecolari noti. D'altra parte, se si fa esprimere myoD in un embrione di rospo allo stadio di due cellule si può osservare l'accensione transitoria di alcuni geni specifici del muscolo scheletrico, ma lo sviluppo dell'embrione non risulterà in alcun modo influenzato da questo evento precoce. Il fatto che in altri tipi di cellule l'espressione di myoD sia sufficiente a dirigere il programma differenziativo che porta alla miogenesi suggerisce che in queste cellule siano presenti alcuni fattori necessari per la realizzazione di quella determinazione istologica, mentre in altre cellule questi fattori non esistono.
Consideriamo adesso un altro esperimento. Nel nematode C. elegans, la proteina codificata da myoD può esser messa in evidenza in un embrione allo stadio di 100 cellule. La proteina è localizzata precisamente nei precursori delle cellule del muscolo della parete corporea. Queste si differenzieranno apertamente solo dopo altre tre o quattro generazioni cellulari. D'altra parte, la trascrizione di myoD in questo organismo comincia molto prima, già allo stadio di 28 cellule. Tutte le cellule da cui più tardi deriverà il tessuto muscolare sono in grado di trascrivere questo gene, ma il dominio cellulare di questa espressione genica è molto più ampio: in questa fase anche alcune cellule che non daranno luogo a tessuto muscolare sono in grado di esprimere il gene. Le discendenti di alcune delle cellule embrionali di C. elegans che sintetizzano l'RNA messaggero di myoD arriveranno successivamente a produrre effettivamente la proteina e quindi si differenzieranno in tessuto muscolare, mentre altre si avvieranno verso un destino diverso, divenendo ad esempio neuroni o cellule epiteliali.
Come abbiamo visto, ci troviamo in presenza di un chiaro esempio di determinazione progressiva per quanto riguarda i precursori del muscolo scheletrico. Ma si osserva allo stesso tempo anche una riduzione progressiva della capacità di differenziarsi in più di un tipo istologico, cioè una riduzione progressiva della ‛potenzialità differenziativa' o ‛istologica' a carico delle cellule degli altri tipi. A uno stadio molto precoce alcune cellule sono infatti in grado di esprimere myoD e quindi di differenziarsi in direzione miogenica, ma più tardi perdono questa capacità. Vale la pena di notare che la ‛riduzione progressiva della potenzialità differenziativa' può essere vista come l'altra faccia del processo di determinazione progressiva. Il primo termine coglie il progressivo allontanarsi delle cellule dal loro stato iniziale indifferenziato, mentre il secondo si riferisce alla marcia di avvicinamento della singola cellula verso il suo obiettivo finale, il differenziamento terminale.
L'idea di progressività risale in verità a tempi lontani, cioè ai numerosi esperimenti di trapianto eseguiti a più riprese su embrioni di un certo numero di specie diverse verso la fine del secolo scorso e nella prima metà di questo. In questi esperimenti venivano trapiantate da un embrione all'altro intere regioni di tessuto embrionale, cellule singole o, più recentemente, anche singoli nuclei.
Lo zigote risultante dalla fecondazione di una cellula uovo è ovviamente in grado di dar luogo a un intero organismo, il quale è costituito per definizione da cellule di tutti i tipi. Per indicare questa sua proprietà si dice che lo zigote è una cellula ‛totipotente'. Il suo nucleo deve quindi possedere tutta l'informazione genetica necessaria per dirigere lo sviluppo di un intero organismo e anch'esso può essere considerato totipotente. Ma che cosa succede se si introduce in una cellula uovo privata del proprio nucleo il nucleo di una cellula diversa?
Nel 1952 Robert Briggs e Thomas King (v., 1952) riuscirono nell'impresa di introdurre in un uovo anucleato di rana (Rana pipiens) un nucleo prelevato da un'altra cellula. La cellula così ottenuta può essere ancora considerata alla stregua di uno zigote, diciamo uno pseudozigote. Alcuni degli pseudozigoti così ottenuti non si dividono affatto e degenerano; altri si dividono per un certo numero di generazioni cellulari; altri infine vanno molto più avanti nel loro sviluppo. Abbiamo visto che un nucleo capace di sostenere lo sviluppo di un individuo adulto completo è detto totipotente, cioè è in grado di dar luogo a cellule di ogni tipo. Un nucleo capace di sostenere uno sviluppo molto avanzato anche se non completo può essere definito ‛pluripotente', mentre nuclei che sostengono uno sviluppo via via più ridotto sono da considerarsi nuclei di ridotta potenzialità embriologica. Il concetto di ‛potenzialità embriologica' così definito dovrebbe ragionevolmente comprendere quello di potenzialità istologica di cui abbiamo parlato precedemente.
Il primo dato emerso da questa analisi è che i nuclei prelevati dalle cellule dell'embrione di rana allo stadio di blastula sono da considerarsi praticamente totipotenti. Questi si dimostrano infatti in grado di promuovere lo sviluppo di girini vitali in una quota oscillante fra il 60 e l'80% degli esperimenti. Questo risultato non giunse totalmente inatteso nemmeno per i primi sperimentatori. Ma che cosa succede se si trapiantano nuclei di cellule appartenenti a stadi di sviluppo via via più avanzati e che sono quindi verosimilmente via via più determinate? La conclusione che se ne trae è che la percentuale di pseudozigoti che producono con successo un girino capace di nuotare e di nutrirsi decresce con continuità se si trapiantano nuclei di stadi via via più avanzati e diviene nulla per i nuclei prelevati dalle cellule somatiche di un girino allo stadio del cosiddetto bottone caudale.
Si possono fare a questo punto alcune considerazioni. Una prima considerazione si riferisce alla ‛costanza del patrimonio genetico' durante lo sviluppo. Se il nucleo delle cellule più determinate avesse perso in maniera irreversibile informazioni genetiche essenziali, non potrebbe essere in grado di dirigere alcun tipo di sviluppo. È chiaro che non è così, come non è così nel caso della determinazione e transdeterminazione dei dischi imaginali della drosofila. Di conseguenza, le alterazioni nucleari associate al procedere dello sviluppo devono essere alterazioni funzionali dotate di un certo livello di reversibilità. Ce le possiamo raffigurare come ‛alterazioni del grado di esprimibilità' di certi gruppi di geni. È chiaro che il fenomeno a cui si assiste in questo tipo di esperimenti è una ‛riprogrammazione' della gestione dell'informazione genetica contenuta nel nucleo. Questa riprogrammazione sarà possibile soltanto nella misura in cui risulterà reversibile il controllo del livello di esprimibilità dei geni implicati.
D'altra parte, anche se i nuclei delle cellule in via di sviluppo sono in grado di dirigere un certo grado di maturazione dell'embrione, la loro potenzialità appare sempre più ridotta. Queste cellule perdono progressivamente la loro potenzialità con il procedere dello sviluppo e questa perdita è registrata dal nucleo. Alla luce delle precedenti considerazioni possiamo allora concludere che, se è vero che la regolazione dell'esprimibilità dei geni è in genere potenzialmente reversibile, la riprogrammazione di questa regolazione risulterà tanto più ardua quanto più irreversibili saranno, con il procedere dello sviluppo, le alterazioni del grado di esprimibilità dei gruppi di geni in questione.
I risultati degli esperimenti di Briggs e King con le uova di rana escludevano la possibilità che il nucleo di una cellula somatica matura fosse totipotente. Operando con cellule del rospo africano Xenopus laevis, l'inglese John Gurdon (v., 1962) ottenne risultati molto simili a quelli ottenuti con la rana, ma ancora più sorprendenti. Anche in questo caso egli osservò una perdita progressiva di potenzialità da parte dei nuclei prelevati dalle cellule somatiche, ma la progressività di questa perdita apparve molto più lenta. Un risultato particolarmente interessante si osserva trapiantando nuclei prelevati da cellule di endoderma intestinale di girino: gli pseudozigoti originati dal trapianto di questi nuclei si sviluppano fino allo stadio di girino in una quota che può arrivare al 7%, e alcuni di questi girini subiscono una regolare metamorfosi fino a dare animali adulti in grado di riprodursi. In linea di principio questi nuclei possono quindi essere considerati totipotenti!
In conclusione, i nuclei di cellule adulte non possono essere considerati totipotenti ma sono certamente pluripotenti. Si registra una progressiva perdita di potenzialità embriologica, ma il livello di riprogrammabilità del patrimonio genetico resta sempre incredibilmente alto, almeno in questi animali. Basti pensare che un nucleo prelevato da un globulo rosso, una cellula che ordinariamente non si divide più e non produce neppure più RNA messaggero, è ancora in grado di generare un girino vitale.
Accanto ai due paradigmi già ricordati ne è stato enunciato un terzo, il paradigma della ‛specificazione posizionale' (v. Wolpert, 1989; v. Boncinelli, 1994): parallelamente alla determinazione del destino istologico di una cellula o di un gruppo di cellule si ha la loro specificazione posizionale. Perché un organismo pluricellulare si sviluppi correttamente occorre che ogni sua cellula riceva al tempo giusto una doppia serie di informazioni: una prima riguardante il suo destino istologico e una seconda riguardante il suo destino posizionale. La cellula deve sapere a quale tipo di tessuto apparterrà e in quale posizione andrà a finire nell'organismo adulto. Nel primo caso si parla, abbiamo visto, di ‛determinazione istologica', nel secondo caso di ‛specificazione posizionale'. Il primo concetto, di cui abbiamo parlato abbondantemente, permea molta della letteratura degli ultimi trent'anni; del secondo concetto, invece, si parla ancora abbastanza poco e perfino nella terminologia non c'è un accordo universale (v. Wolpert, 1969, 1989 e 1991).
Dilungarsi a chiarire l'importanza dell'informazione posizionale nell'embrione prima e nell'organismo adulto poi potrebbe apparire superfluo. Basta considerare il fatto che un corpo che contenesse tutti i tessuti normalmente presenti in un organismo adulto ma disposti senza un ordine spaziale non sarebbe un organismo vivente. Eppure la problematica posizionale è emersa solo di recente, e molto faticosamente, attraverso il susseguirsi di varie fasi. In un primo tempo hanno predominato i problemi inerenti all'organizzazione spaziale locale necessaria per spiegare la generazione di forme o di strutture complesse. Si tratta, come abbiamo visto, di spiegare la genesi di strutture biologiche superiori di vario tipo, come una mano, un'ala, una penna, un tubulo renale, l'occhio composto di una mosca o il disegno sull'ala di una farfalla. Al di là e al di sopra dell'organizzazione locale c'è poi il problema della ‛specificazione regionale', intesa come la strutturazione di intere parti del corpo composte di vari organi contenenti tipi diversi di tessuti disposti ordinatamente. E al di sopra di tutto ciò esiste il problema dell'organizzazione spaziale generale del corpo: la testa deve stare al suo posto, così come il torace e gli arti, e l'arto superiore deve essere distinto da quello inferiore e così via.
Questo tipo di problemi è stato ovviamente sempre presente sullo sfondo di qualsiasi indagine embriologica, ma l'aspetto posizionale restava, per così dire, nell'ombra. Questo stato di cose è andato avanti finché qualcuno non ha ritenuto di dover puntualizzare che forse una cellula muscolare del braccio è simile ma non del tutto equivalente a una cellula muscolare della coscia, o che una cartilagine della testa non è equivalente a una del ginocchio. Da notare che queste osservazioni così elementari sono assurte al rango di principio e hanno preso il nome di ‛principio di non equivalenza' (v. Lewis e Wolpert, 1976). A partire dagli anni settanta la problematica che ruota intorno alla gestione dell'informazione posizionale e al conseguente ‛indirizzamento posizionale' delle varie cellule ha ricevuto sempre maggiore attenzione. Tuttavia, non esiste ancora un corpus adeguato di riflessioni teoriche comparabile a quello accumulatosi intorno al problema della determinazione istologica e del differenziamento cellulare.
Una delle conclusioni concettualmente più interessanti tratte da questo tipo di analisi è rappresentata dalla constatazione che la specificazione posizionale trascende spesso la discendenza cellulare e opera su più cloni cellulari in una volta. A questo proposito è stato coniato anche un termine specifico: un gruppo di cloni cellulari che hanno ricevuto uno stesso indirizzo posizionale viene definito ‛policlone' (v. Crick e Lawrence, 1975; v. Ingham e Martinez Arias, 1992). Consideriamo, ad esempio, l'ala della drosofila rappresentata nella fig. 5B. I cloni di cellule che costituiscono la superficie dorsale dell'ala costituiscono un policlone, così come quelli che ne costituiscono la superficie ventrale. All'interno della superficie dorsale anche i cloni appartenenti al compartimento anteriore costituiscono un policlone, e così quelli che costituiscono il compartimento posteriore. Già nella fase di disco imaginale le cellule dei vari policloni si sono assortite e appaiono separate da bordi netti (linee 1 e 2 della fig. 5B). Nella figura dell'ala si nota un clone appartenente al compartimento anteriore che, pur espandendosi disordinatamente, non attraversa mai il bordo di confine tra il suo compartimento anteriore e quello posteriore. Si tratta di una barriera invisibile ma insuperabile. Questa barriera è controllata geneticamente: è il prodotto del gene engrailed (en) che sovrintende a questa segregazione (v. García-Bellido e altri, 1973). In moscerini mutanti per questo gene, i cloni cellulari dell'ala sono liberi di traversare il confine tra un compartimento e l'altro e lo fanno in maniera casuale.
Specificazione posizionale e determinazione istologica sono al momento due concetti astratti particolarmente utili ai fini di un'esposizione degli elementi fondamentali della biologia dello sviluppo. Spetterà alla ricerca futura chiarire il loro rispettivo ruolo e rispondere a domande del tipo: specificazione posizionale e determinazione istologica vengono generate e veicolate indipendentemente o si avvalgono di un supporto materiale comune? E, in particolare, è possibile che l'informazione richiesta per la specificazione posizionale sia riconducibile, in tutto o in parte, a quella riguardante la determinazione istologica? Quello che è certo è che a un determinato momento dello sviluppo ogni cellula riceve entrambi i tipi di informazione biologica, se non qualche altra cosa ancora. Notiamo per concludere che è stato il riconoscimento della necessità di una specificazione posizionale, in aggiunta alla determinazione istologica, che ci ha spinto a tenere distinte tra loro potenzialità embriologica (od ontogenetica) e potenzialità istologica, anche se i due termini vengono spesso considerati sinonimi.
II. Segregazione di determinanti citoplasmatici. - Abbiamo visto che quasi tutti gli eventi differenziativi dello sviluppo possono essere spiegati con l'ausilio del concetto di espressione genica differenziale di gruppi specifici di geni. Molto si è imparato in questi decenni sui meccanismi dell'azione genica e sulla sua regolazione nelle cellule di un organismo che sta completando il suo processo di sviluppo. Molto meno si sa invece sui fattori che regolano l'espressione differenziale dei vari geni nelle cellule di un organismo che sta cominciando a svilupparsi.
Si conoscono a tutt'oggi solamente due meccanismi generali capaci di controllare l'espressione genica nel primo periodo dello sviluppo: la localizzazione, e conseguente segregazione di determinanti citoplasmatici, e l'induzione (v. Horvitz e Herskowitz, 1992; v. Boncinelli, 1994; v. biologia, vol. I; v. embriologia, vol. II; v. sviluppo embrionale, vol. VII). Partiamo dall'ipotesi che un gene di una data cellula continui a esprimersi come sta facendo se non sopravvengono elementi nuovi e diversi. Questi elementi non possono che essere costituiti da segnali biologici e questi segnali possono venire dall'interno della stessa cellula o dal suo esterno. Nel primo caso la cellula si comporta come se fosse un'entità sorda e cieca, la cui evoluzione nello sviluppo è semplicemente la conseguenza di un piano prestabilito scritto da qualche parte nel suo interno. Nel secondo caso lo stato della cellula evolve sulla base di informazioni biologiche provenienti da altre cellule o dal mondo circostante in generale. In ogni cellula, in ogni momento della sua vita, sono presenti entrambi i tipi di segnale. A seconda degli organismi, dei tessuti e degli stadi può talvolta predominare una segnalazione dall'interno o una dall'esterno.
Non è difficile comprendere come possa arrivare alla cellula un segnale dall'esterno. Queste segnalazioni dall'esterno devono confluire prima o poi in una specifica molecola o complesso di molecole che entrano nel nucleo, si vanno a piazzare su una o più specifiche regioni del genoma e dettano le istruzioni a uno o più geni. Per ogni gene queste istruzioni sono relative al fatto di stare spento o acceso e, se deve stare acceso e quindi esprimersi, con quale intensità lo deve fare.
Un po' più complesso appare comprendere il meccanismo di regolazione dall'interno. Possiamo pensare a un'informazione contenuta in maniera più o meno nascosta nel citoplasma di certe cellule e assente in quello di altre. All'atto della divisione mitotica della cellula che lo contiene, questo principio o ‛determinante citoplasmatico' può passare in una sola delle due cellule figlie e da questa a una parte della progenie. Questo principio può agire immediatamente o può venire conservato in uno stato di latenza per generazioni e generazioni cellulari. La maniera più semplice di raffigurarsi questo principio è ipotizzare la presenza di un prodotto proteico sequestrato in un determinato compartimento cellulare o quella di un RNA messaggero che venga tradotto solo al momento opportuno. L'uscita dallo stato di latenza deve essere a sua volta regolata da altri determinanti endocellulari, o da un cambiamento delle condizioni al di fuori della cellula, oppure semplicemente dal tempo. Possiamo immaginare, ad esempio, che la molecola che blocca l'estrinsecazione del principio si trasmetta attraverso le generazioni cellulari in modo da diluirsi progressivamente. All'inizio la quantità presente è sufficiente per tenere a bada la manifestazione del principio latente, ma a ogni divisione cellulare la sua concentrazione diminuisce fino a che, a un certo momento, non ce ne sarà più abbastanza per far fronte al suo compito.
In conclusione, per la realizzazione di un processo di determinazione mediante localizzazione e segregazione di un determinante citoplasmatico occorre il concorso di diversi meccanismi. Innanzitutto si deve avere la ‛localizzazione' di questo determinante all'interno del citoplasma di una cellula embrionale. La corretta localizzazione del determinante citoplasmatico in una data cellula serve a controllare la sua successiva segregazione in una o nell'altra delle due cellule figlie all'atto della divisione cellulare. La ‛segregazione' è il fenomeno più importante e deve avvenire in modo che il determinante in questione finisca nella cellula giusta e non nell'altra. Si richiede infine un ‛meccanismo marcatempo' che scandisca il tempo biologico dello smascheramento dilazionato di quel dato determinante.
In questa ottica, non sorprende che la localizzazione di determinanti citoplasmatici cominci già nell'uovo, la cellula asimmetrica per eccellenza. Anzi, un esempio estremo di influenza dei determinanti citoplasmatici è dato proprio dalla localizzazione e dall'abbondanza del materiale vitellino all'interno dell'uovo. Queste caratteristiche specifiche determinano di fatto l'andamento delle prime divisioni cellulari dopo la fecondazione e tutta l'impostazione della gastrulazione. In base alla quantità e alla localizzazione del vitello, le uova si possono distinguere in ‛isolecitiche' (o ‛oligolecitiche') se ne contengono poco, ‛mesolecitiche' se ne contengono una quantità intermedia e ‛telolecitiche' se ne contengono una quantità notevole (v. tab. II). In quest'ultimo caso, il vitello deve necessariamente essere localizzato in maniera chiaramente asimmetrica a un'estremità della cellula uovo, cioè al ‛polo vegetativo', contrapposto a quello ‛animale'. Per completezza dobbiamo ricordare poi che le uova della maggior parte degli insetti sono dette ‛centrolecitiche' perché hanno tutto il vitello concentrato in posizione centrale.
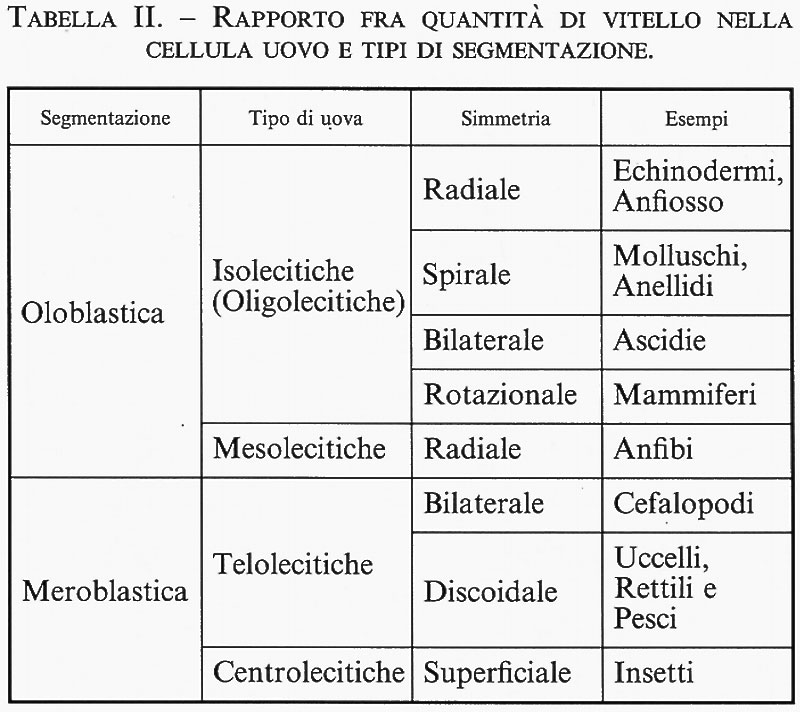
La quantità e la posizione del vitello influenzano direttamente il tipo di segmentazione dello zigote. Il vitello è in genere un ostacolo alla segmentazione, così che questa può essere completa o ‛oloblastica' se il vitello è scarso, oppure parziale o ‛meroblastica' se il vitello è abbondante. Nella segmentazione oloblastica i solchi delle divisioni cellulari attraversano tutta la massa dello zigote. Nella segmentazione meroblastica soltanto una porzione del citoplasma viene suddivisa, perché i solchi delle divisioni cellulari non penetrano a fondo nel vitello. La segmentazione meroblastica può essere discoidale, come nell'uovo di pollo che è particolarmente ricco di vitello, o superficiale, come negli Insetti. Le varie divisioni cellulari nella segmentazione oloblastica possono susseguirsi secondo diverse classi di simmetria, la quale può essere radiale, spirale, rotazionale, o bilaterale. Tutto ciò si riflette a sua volta nelle modalità della gastrulazione, così che la quantità e la posizione del vitello influenzano indirettamente anche il tipo di gastrulazione cui va incontro l'embrione. Queste diverse modalità di gastrulazione in embrioni di specie diverse mascherano la fondamentale identità di questo processo: se si fa astrazione dalle differenze nel vitello, infatti, la gastrulazione del pollo non è molto diversa da quella degli Anfibi né quest'ultima da quella dell'anfiosso. Si può passare dalla prima alla seconda e da questa alla terza se si immagina di rimuovere progressivamente la gigantesca massa di vitello presente nell'uovo degli Uccelli.
Abbiamo già parlato del ruolo della discendenza cellulare diretta nella determinazione cellulare in generale e abbiamo visto che quando il destino di una data cellula è determinato esclusivamente in base alla sua ascendenza cellulare si parla di sviluppo a mosaico, mentre quando l'ascendenza cellulare è solamente uno di più o meno numerosi fattori di varia natura che intervengono in tale determinazione si parla di sviluppo di tipo regolativo. Le varie specie animali possono essere ordinate lungo un asse continuo di preferenzialità che si estende fra due estremi. A un estremo di questa scala stanno alcuni gruppi di molluschi, tunicati e nematodi che mostrano uno sviluppo prevalentemente a mosaico: in questi organismi è quindi privilegiata la determinazione precoce mediante segregazione di determinanti citoplasmatici. All'altro estremo si trovano Mammiferi, Anfibi e ricci di mare, che mostrano uno sviluppo di tipo più regolativo, nel quale le interazioni cellulari hanno un peso maggiore. Vediamo adesso brevemente alcuni esempi noti di segregazione e di induzione nelle prime fasi dello sviluppo.
La segregazione di determinanti citoplasmatici può essere osservata abbastanza chiaramente negli embrioni precoci di Nematodi, Tunicati e Molluschi. In alcune specie di molluschi lo zigote mostra un'estrusione di materiale citoplasmatico, che prende il nome di ‛lobo polare', immediatamente prima della prima divisione cellulare. In certi casi il lobo polare è molto evidente, perché separato dal resto dello zigote da un sottile peduncolo. La prima divisione cellulare è asimmetrica: il lobo polare resta connesso soltanto con uno dei due blastomeri, indicato con CD. Le dimensioni del lobo polare a volte sono tali da farlo scambiare per una terza cellula, cosicché il tutto prende l'aspetto di un trifoglio. A questo punto il blastomero CD riassorbe il lobo polare per riestruderlo immediatamente prima della seconda divisione. Dopo questa seconda divisione cellulare il lobo polare resta connesso esclusivamente con il blastomero D, dal quale viene poi riassorbito per non ricomparire mai più. Se si rimuove il lobo polare allo stadio di due cellule, le successive divisioni cellulari procedono in modo del tutto normale, ma la larva risulterà assolutamente priva di tessuti di derivazione mesodermica. Una larvetta anormale più o meno dello stesso tipo si ottiene anche in seguito a rimozione del blastomero D da un embrione allo stadio di 4 blastomeri (v. Wilson, 1904). Se ne può concludere che il citoplasma normalmente racchiuso nel lobo polare contiene l'informazione necessaria per la determinazione delle cellule in direzione mesodermica e che questa informazione conferisce al blastomero D la sua capacità di formare mesoderma. Si può dimostrare l'esistenza di una specifica regione del citoplasma dell'uovo che contiene tutti quei fattori o principî necessari per la specificazione del destino embriologico del blastomero D e della sua capacità di svilupparsi in direzione mesodermica: il nucleo di questo blastomero riceve quindi dal citoplasma uno specifico complesso di istruzioni che lo condurranno a dare origine a un certo tipo di cellule.
III. I vari tipi di induzione. - Veniamo adesso al fenomeno dell'induzione, che possiamo definire in astratto come un'interazione fra due gruppi di cellule in conseguenza della quale viene a restringersi la potenzialità embriologica delle cellule di almeno uno dei due gruppi. In pratica con questo termine si intende l'azione di un gruppo di cellule (‛induttore') su un secondo gruppo contiguo di cellule (‛bersaglio'), con il risultato che le cellule di questo secondo gruppo cambiano il loro stato di determinazione e talvolta alcuni loro tratti fenotipici.
È usuale distinguere ancora un'interazione induttiva permissiva da una istruttiva. Ci troviamo in presenza di un'induzione ‛permissiva' quando il gruppo di cellule sul quale si esercita l'azione induttiva ha già un suo destino embriologico predeterminato e attende solo un segnale per incamminarsi su questa via. Ci troviamo in presenza di un'induzione ‛istruttiva' quando il gruppo di cellule sul quale si esercita l'azione induttiva acquisisce dal segnale induttivo le informazioni sul suo destino embriologico e sul suo piano di attuazione. Un'interazione istruttiva perfettamente corrispondente a questa definizione non è facile da immaginare; più realistico è invece parlare di induzione istruttiva quando il gruppo di cellule sul quale si esercita l'azione induttiva abbia a disposizione una certa gamma di destini embriologici alternativi e riceva dal segnale induttivo istruzioni relative a quale debba essere quello verso cui dirigersi effettivamente.
Lo stesso segnale induttivo può sortire un certo effetto su un gruppo di cellule e non sortirne affatto su un altro gruppo. La predisposizione a rispondere a un determinato stimolo induttivo è chiamata ‛competenza'. La competenza non è che un aspetto particolare dello stato di determinazione di un gruppo di cellule e come tale è limitata nel tempo e nello spazio. La limitazione più interessante della condizione di competenza è forse quella temporale: alcune cellule acquisiscono a un certo stadio dello sviluppo la competenza specifica a rispondere a un certo stimolo e la mantengono per un lasso di tempo limitato, detto ‛periodo di competenza', senza palesarne alcun segno prima o dopo.
I primi fenomeni induttivi furono osservati in embrioni di anfibio da Hans Spemann e Hilde Mangold. Nel 1924, questi ricercatori, grazie alle possibilità di indagine aperte dagli esperimenti di trapianto, si accorsero che una specifica regione dell'embrione in gastrulazione mostrava un'eccezionale autonomia di comportamento in tutti i tipi di trapianto: si tratta del labbro dorsale del blastoporo, il quale è capace di promuovere in quasi ogni circostanza l'inizio di una nuova gastrulazione e la formazione di un nuovo asse embrionale, completo di notocorda, di corda neurale e di muscolatura parassiale, e conserva tale capacità anche se viene trapiantato nella regione ventrale dell'embrione. Operando con tessuti di salamandre di due colori diversi fu possibile dimostrare che il nuovo asse era indotto dal frammento trapiantato, ma era costituito da cellule dell'embrione ricevente: il labbro dorsale trapiantato era capace quindi di sovvertire l'assetto delle cellule circostanti il punto del trapianto, specificandone nuovamente il destino embriologico. Il potere del frammento trapiantato è tale da riuscire a organizzare queste cellule dell'embrione ricevente in una struttura superiore della complessità di un nuovo asse dorsale: per questa sua capacità la regione corrispondente al labbro dorsale del blastoporo di un anfibio in gastrulazione venne battezzata ‛organizzatore' e ancora oggi viene designata come ‛organizzatore primario' o ‛organizzatore di Spemann'. A questo tipo di induzione è stato dato il nome di ‛induzione primaria' o induzione embrionale primaria allo scopo di distinguerla da altri tipi di induzione.
L'induzione primaria, osservata per la prima volta mediante esperimenti di trapianto, è in realtà un fenomeno naturale attraverso il quale l'organizzatore di Spemann regola la formazione dell'asse dorsale negli Anfibi all'atto della gastrulazione. Anche nei Mammiferi esiste una struttura embrionale capace di compiere funzioni analoghe al momento della gastrulazione: si tratta della parte più anteriore della stria primitiva in formazione, detta ‛nodo', omologa alla struttura dell'embrione di pollo denominata ‛nodo di Hensen'. Questa regione possiede molte interessanti proprietà, fra le quali quella di indurre la formazione di un asse secondario una volta trapiantata in una sede ectopica di un embrione di pollo o di topo all'inizio della gastrulazione.
La formazione di un asse dorsale secondario completo è un fenomeno complesso che comporta l'instaurazione di una nuova polarità dorso-ventrale e il riconoscimento di una polarità rostro-caudale. La formazione dell'asse dorsale primario in un embrione normale è anch'essa un fenomeno molto complesso e interessante. In un caso e nell'altro colpisce l'alta specificità regionale delle strutture formate, soprattutto di quelle che costituiscono il sistema nervoso centrale: l'encefalo viene a trovarsi in posizione rostrale rispetto al midollo spinale e all'interno dell'encefalo il proencefalo precede il mesencefalo e questo il rombencefalo. Il sistema nervoso in formazione acquisisce questa specificità regionale dal sottostante mesoderma assiale dorsale, detto anche ‛cordomesoderma'. Il cordomesoderma quindi non solo induce la formazione del tessuto che costituisce il sistema nervoso centrale, ma ne determina anche la specificità regionale. Questo flusso di informazione posizionale dal cordomesoderma dell'embrione di anfibio al sovrastante tessuto nervoso in formazione può essere messo in evidenza sperimentalmente trapiantando nella cavità blastocelica di un embrione allo stadio di gastrula iniziale porzioni di cordomesoderma già determinato di un secondo embrione: se si trapianta una porzione anteriore si otterrà l'induzione di strutture cefaliche, se si trapianta una regione più centrale si otterrà l'induzione di strutture cervicali, se si trapianta una regione posteriore si otterrà l'induzione di strutture caudali.
Da tutto ciò possiamo concludere che il primo effetto dell'organizzatore di Spemann è quello di indurre un cordomesoderma che contenga le strutture appropriate e che, a sua volta, sia capace di indurre la formazione del tessuto nervoso sovrastante con l'appropriata specificità regionale. Dobbiamo notare che organizzatore di Spemann e labbro dorsale del blastoporo non sono definizioni di una struttura materiale, ma di una regione. Questa regione contiene cellule diverse in tempi diversi: in ogni momento della gastrulazione alcune cellule entrano in questa regione mentre altre ne escono. Si può pensare quindi che organizzatori primari prelevati a tempi diversi mostrino proprietà induttive leggermente diverse. Questa ipotesi è stata confermata da una serie di esperimenti. Un labbro dorsale prelevato più precocemente induce strutture assiali anteriori, mentre un labbro dorsale prelevato più tardivamente induce strutture assiali posteriori. Sembra quindi ragionevole supporre che una variazione nel tempo delle proprietà dell'organizzatore primario si traduca durante la gastrulazione in una specificità regionale lungo l'asse maggiore del corpo dell'anfibio. Va detto però che nonostante una ricerca appassionata che dura da decenni ben poco si conosce circa la natura di tali segnali e il meccanismo che è alla base dell'induzione primaria.
Nonostante il nome, l'induzione primaria non costituisce il primo evento di induzione nello sviluppo dell'anfibio: ne esiste infatti un altro, detto ‛induzione del mesoderma', che la precede nel tempo. La blastula dell'anfibio si presenta come una sferetta cava asimmetrica lungo l'asse polo animale-polo vegetativo (v. fig. 18), costituita soltanto dai precursori di due foglietti embrionali. Le cellule dell'emisfero vegetativo sono destinate a dare endoderma, mentre le cellule dell'emisfero animale sono destinate a dare ectoderma. A partire da un certo momento, le cellule di una ristretta fascia equatoriale - detta ‛zona marginale' - sono indotte a cambiare il proprio destino embriologico da quello di cellule ectodermiche a quello di cellule mesodermiche: è il risultato di un'azione induttiva esercitata dalle cellule endodermiche su quelle ectodermiche. Questo fenomeno è stato messo in luce dall'olandese Peter Nieuwkoop (v., 1973), che ha osservato accuratamente il destino embriologico delle varie porzioni isolate della blastula (v. fig. 18A). Se si rimuovono da una blastula le cellule della fascia equatoriale, le due regioni polari isolate non danno luogo ad alcun derivato mesodermico: la calotta polare animale dà luogo a tessuto ectodermico dotato di ciglia, mentre la regione vegetativa dà luogo a formazioni endodermiche. Se però le due regioni vengono combinate, cioè fatte sviluppare giustapposte, si osserva il comparire nella regione animale di strutture mesodermiche come la notocorda, la muscolatura scheletrica e i reni. Per parte sua, la zona marginale isolata da una blastula avanzata è capace di dar luogo a strutture di derivazione mesodermica. Questa sua proprietà è stata acquisita progressivamente attraverso una serie di azioni induttive esercitate sull'ectoderma della regione equatoriale da parte dell'endoderma vegetativo.
Attraverso queste interazioni non si stabilisce soltanto un destino mesodermico per l'intera zona marginale, ma anche una sua suddivisione in regioni ventrali e dorsali. Mentre le cellule dell'emisfero vegetativo dorsale estremo specificano le varie componenti del mesoderma assiale, quali la notocorda e i somiti, le restanti cellule dell'emisfero vegetativo specificano il mesoderma intermedio, essenzialmente le cellule ematiche e il rene, e quello ventrale, cioè sistema muscolare e mesenchima. In questo schema rimane da spiegare come il mesoderma non assiale possa a sua volta essere suddiviso in mesoderma ventrale e mesoderma intermedio. È stato proposto a questo scopo un modello di interazione induttiva del mesoderma implicante tre tipi di segnali e detto per ciò ‛modello a tre segnali' (v. fig. 18B; v. Slack, 19912). Le cellule della regione vegetativa dorsale estrema (DV) inducono direttamente il mesoderma assiale, mentre le cellule della restante regione vegetativa, più ventrale (VV), inducono il mesoderma non assiale. All'interno di questo si osserva un'ulteriore specificazione in mesoderma ventrale e mesoderma intermedio sotto l'azione di un terzo segnale che agisce in senso orizzontale provenendo dal mesoderma assiale presuntivo stesso, che corrisponde alla regione dell'organizzatore primario di Spemann (O). Le cellule del mesoderma più vicino all'organizzatore acquisiranno un destino di mesoderma intermedio, mentre quelle più lontane daranno luogo al mesoderma ventrale. La specificazione in senso dorsale della regione vegetativa dorsale viene acquisita nelle primissime fasi della segmentazione: già a partire dallo stadio a otto blastomeri le cellule del quadrante vegetativo dorsale sviluppano la facoltà di segnalare alle cellule sovrastanti un cambiamento di indirizzo istologico in senso dorsale, o meglio antero-dorsale. Questa regione si comporta quindi come un centro organizzatore precursore dell'organizzatore primario di Spemann ed è stata chiamata ‛centro di Nieuwkoop'.
Ben poco si sa della natura molecolare di questi processi, ma in tempi recentissimi sono stati tentati alcuni approcci che hanno dato risultati molto interessanti. Si è cominciato col mettere a punto una metodica che permettesse di saggiare l'attività mesodermizzante e in grado di dare risultati riproducibili. Abbiamo visto che, fatta crescere in isolamento, la calotta polare animale (animal cap) non è in grado di diventare altro che epitelio ciliato, mentre la sua combinazione con cellule dell'emisfero vegetativo ne trasforma parzialmente l'identità istologica in direzione mesodermica. L'idea vincente è stata allora quella di far crescere in vitro una serie di calotte animali dopo averle trattate con varie sostanze e osservare se in alcuni di questi modelli si notassero segni di mesodermizzazione. Questo è effettivamente avvenuto in presenza di due diverse sostanze già note per la loro capacità di agire come fattori di crescita: l'FGF (Fibroblast Growth Factor) e l'attivina, un membro della famiglia dei fattori di crescita peptidici di tipo TGF-β (Transforming Growth Factor β).
Queste due sostanze non hanno esattamente lo stesso effetto. L'FGF induce nelle cellule trattate caratteristiche più tipiche del mesoderma non assiale e potrebbe quindi rappresentare il principio attivo che deriva dalla regione vegetativa ventrale VV (v. fig. 18B). L'attivina, viceversa, induce caratteristiche tipiche del mesoderma assiale e potrebbe quindi essere un componente del principio attivo che deriva dalla regione vegetativa dorsale DV (v. Green e altri, 1992). L'attivina non può però rappresentare l'unico componente di questo secondo segnale, perché non è in grado di mimarne da sola tutti gli effetti. Si è visto che la sua azione diviene più simile a quella attesa per questo secondo segnale se agisce in combinazione con almeno altri due tipi di sostanze, e precisamente i prodotti dei geni chiamati noggin e Wnt (v. Moon e Christian, 1992). La combinazione dell'attivina con questi prodotti genici conduce a effetti comparabili anche a quelli esercitati dall'ipotetico principio attivo prodotto dalla regione O nel modello a tre segnali della fig. 18B. Questi prodotti genici sono capaci non solo di dorsalizzare il mesoderma, ma anche di indurne la regionalizzazione e il successivo differenziamento in compartimenti istologici diversi rappresentati dalla notocorda, dal muscolo, dall'apparato ematopoietico, dai reni e così via. In seguito all'azione di queste sostanze si osserva anche la formazione dell'asse cordomesodermico e la conseguente induzione del tubo neurale. L'effetto induttivo dell'organizzatore primario di Spemann può essere quindi ricondotto in buona parte all'azione di queste sostanze.
Un terzo tipo di induzione precoce è rappresentato dall'induzione del tessuto nervoso da parte del cordomesoderma a partire dal neuroectoderma dorsale, cioè dall'‛induzione neurale'. Un'enorme mole di esperimenti è stata progettata nel tentativo di identificare la sostanza capace di realizzare questo tipo di induzione, ma a tutt'oggi non è stato ottenuto alcun risultato. A un certo punto parve anzi che quasi ogni sostanza fosse in grado di indurre, in maniera aspecifica, una sorta di placca neurale, e questo condusse all'ipotesi che l'induttore esistesse allo stato latente nelle cellule dell'ectoderma dorsale stesso e venisse liberato ogniqualvolta una qualsiasi sostanza causava uno stress a queste cellule.
Per ‛induzione secondaria' si intende invece ciascuno dei numerosi eventi induttivi che hanno luogo più avanti nell'embriogenesi, quando il piano globale dell'organismo è già stabilito (v. Saxén e Toivonen, 1962). Su meccanismi del genere è basata la formazione del rene, dell'occhio, dell'orecchio, delle piume, delle scaglie, dei denti, del pancreas e così via. Probabilmente la più nota delle induzioni secondarie è quella che conduce alla formazione del cristallino dell'occhio. Questo si forma a partire dalle cellule dell'ectoderma della testa sotto l'influsso della vescicola ottica di origine prosencefalica. All'inizio del secolo Spemann osservò che distruggendo il primordio della vescicola ottica il corrispondente cristallino non si formava. La conclusione non poteva che essere quella che il contatto con questo tessuto neuroectodermico era richiesto per il differenziamento delle cellule ectodermiche e la formazione del cristallino stesso. Se poi la vescicola ottica veniva trapiantata a uno stadio appropriato in una regione cefalica diversa, si osservava in questa nuova sede la formazione ectopica di una struttura molto simile a un cristallino. Sembra chiaro a questo punto che il contatto con la vescicola ottica sia condizione necessaria e sufficiente per l'induzione del cristallino. Le cose non sono ovviamente così semplici, ma questi fenomeni sono certamente alla base dell'induzione del cristallino.
IV. Articolazione del differenziamento cellulare. Differenziamento istologico manifesto, crescita, maturazione e morte. - Consideriamo adesso molto brevemente le varie articolazioni del differenziamento cellulare, quali le abbiamo delineate a suo tempo nella tab. I. Dell'indirizzamento posizionale e della determinazione abbiamo parlato più che esaurientemente. Per ‛differenziamento istologico manifesto' possiamo intendere quel complesso di cambiamenti macroscopici che segnalano in maniera evidente l'acquisizione di nuove proprietà da parte di una cellula che si differenzia. Si tratta quindi semplicemente di un altro modo di designare il differenziamento palese di un determinato gruppo di cellule. Questo evento può essere concomitante ad alcuni passi determinativi, ma anche molto distante nel tempo da alcuni di essi. Ad esempio, le cellule del disco imaginale dell'ala di un moscerino sono determinate già dai primi stadi larvali a divenire parti dell'ala adulta, ma il loro differenziamento manifesto ha luogo soltanto durante il periodo della pupazione. Un caso particolare di differenziamento istologico è rappresentato dalla ‛crescita' e dalla ‛maturazione' di alcune cellule che si avviano allo stadio terminale del loro processo di differenziamento. Quando una cellula perviene a questi stadi, la sua evoluzione è in genere ormai irreversibile, gli eventi determinativi e differenziativi al suo interno volgono al termine. Esempi di questi eventi possono essere osservati nella maturazione dei gameti, delle cellule dell'epidermide o delle mucose intestinali, dei motoneuroni spinali o delle cellule del sangue.
Consideriamo adesso la ‛morte cellulare programmata' come esempio particolare di differenziamento (v. Ellis e altri, 1991). Si tratta di un fenomeno molto diffuso: si è calcolato che un buon 15% di tutti i neuroni che si originano nel sistema nervoso centrale di un mammifero vanno incontro a morte nelle prime fasi del suo sviluppo. È necessario operare una distinzione tra la morte accidentale della cellula, inevitabile e certamente accelerata dall'usura e da fattori esterni, che prende il nome di ‛necrosi cellulare', e la morte attiva e programmata, che prende il nome di ‛apoptosi': il primo tipo di morte cellulare è un evento cellulare passivo, mentre il secondo è un processo attivo.
La cellula partecipa attivamente al proprio suicidio. Si tratta di un vero e proprio processo differenziativo che comporta la ricezione di appropriati segnali esterni, accompagnata da caratteristici eventi nucleari culminanti nell'accensione di geni regolatori specifici e nella produzione di proteine specifiche. Esempi di morte cellulare programmata si possono incontrare nella formazione o nella ‛rifinitura' di diversi organi. Ad esempio, la separazione dell'ulna e del radio o quella delle dita della mano o del piede sono dovute a morte cellulare di specifiche cellule: le dita dell'embrione sono unite tra di loro da un setto dermico, una sorta di membrana palmare, le cui cellule andranno incontro a un processo di apoptosi in seguito al quale le dita risulteranno infine separate. Questo fenomeno è controllato geneticamente: infatti le dita della zampa di un pollo si separano prima della nascita, mentre quelle di un palmipede restano unite per tutta la vita.
Una determinazione estremamente precisa di quali cellule devono morire si può osservare nello sviluppo del nematode Caenorhabditis elegans (v. Ellis e Horvitz, 1986): è questo un organismo con uno sviluppo a mosaico rigidamente determinato, nell'ambito del quale solo certe determinate cellule saranno destinate a morire e non altre. Per uno sviluppo normale devono morire esattamente 131 cellule delle 1.090 che si sono venute formando, ed esattamente 959 devono sopravvivere e differenziarsi. Questa determinazione cellulare è stata studiata nella sua derivazione clonale e nella sua regolazione genetica. Vi sono innanzitutto dei geni che controllano le decisioni che ogni cellula deve prendere a ogni biforcazione dell'albero della discendenza cellulare: individui mutanti per questi geni sono detti ‛mutanti di lineage', perché mostrano turbamenti più o meno estesi nel quadro generale delle identità delle varie cellule. Alcune cellule, in sostanza, scambiano il loro destino embriologico con quello delle sorelle o della cellula madre. Così nei mutanti del gene unc-86 si passa dal normale schema A → B + C allo schema A → B + A, nei mutanti di lin-17 (o di lin-18) dallo schema normale allo schema A → B + B e nei mutanti di lin-44 a quello A → C + B. Ancora più interessante è il caso di lin-12: una mutazione recessiva di questo gene trasforma lo schema normale A → B + C nello schema A → B + B, mentre una mutazione dominante lo trasforma in A → C + C. In questo organismo l'analisi genetica ha quindi permesso di individuare un certo numero di geni regolatori che hanno un ruolo fondamentale nelle decisioni che le cellule devono prendere a ogni divisione cellulare.
Geni di questa categoria controllano la morte programmata delle singole cellule così come controllano un qualsiasi altro evento differenziativo e non sono quindi specifici per questo evento. Esistono invece geni specifici implicati nella morte programmata, detti ceh (da cell death, morte cellulare): una mutazione in uno di questi geni impedisce la morte di tutte o di una vasta categoria di cellule programmate per morire. Si tratta evidentemente di geni regolatori all'apice della gerarchia della morte cellulare, dei quali i più studiati sono ceh-3 e ceh-4, il cui prodotto è necessario all'interno delle singole cellule perché queste possano morire al momento opportuno. Anche questi geni devono rispondere a determinati segnali, interni o esterni: esiste ad esempio un altro gene, ceh-9, che controlla negativamente l'azione di questi geni, per cui la morte cellulare programmata può avvenire solo in assenza del prodotto di ceh-9. Le cellule nelle quali tale gene è attivo non possono andare incontro a questo tipo di morte; viceversa, embrioni mutanti per una lesione in questo gene mostrano segni di morte cellulare generalizzata. Esempi di morte cellulare programmata osservabili in altre specie sono stati molto meno studiati dal punto di vista genetico, ma non c'è motivo di ritenere che non siano controllati attraverso meccanismi simili. Molto recentemente sono stati trovati geni di mammifero strettamente imparentati al gene ceh-9. Sempre più prezioso appare quindi l'apporto dello studio delle specie inferiori, e soprattutto dei cosiddetti sistemi modello quali C. elegans e Drosophila, per la comprensione della nostra biologia e in definitiva della nostra stessa natura (v. sviluppo embrionale, vol. VII).
c) Interazioni fra le cellule e movimenti cellulari.
Anche se stiamo considerando eventi differenziativi a livello della singola cellula, non è possibile ignorare che con la sua superficie la cellula si trova a contatto con altre cellule e spesso con una matrice extracellulare, con le quali interagisce continuamente mediante fenomeni di interazione cellula-cellula o cellula-matrice extracellulare; talvolta, in base a queste interazioni, cambia forma e si muove. Le molecole interessate in questi fenomeni sono innumerevoli, presenti a volte in quantità minime e soprattutto difficilissime da ‛cogliere sul fatto'. La natura stessa di queste interazioni, deboli e fugaci a livello delle singole molecole anche se forti e vincolanti a livello dell'intera superficie cellulare, rende quasi impossibile il compito di analizzarle in vivo e molto difficile anche quello di studiarle in vitro.
Le cellule interagiscono innanzitutto per formare i tessuti e, da questi, i vari organi e sistemi, ciascuno dei quali passa attraverso una serie di strutture transitorie intermedie che spesso poco o nulla hanno a che fare con la sua configurazione finale o quasi finale. Pertanto, la maggior parte delle interazioni cellulari deve necessariamente essere determinata ma metastabile, e questo introduce un elemento di complicazione ulteriore.
Per interagire, all'interno dei tessuti e tra di essi, le cellule devono prima di tutto riconoscersi. In base a questo riconoscimento le cellule si aggregano o si disaggregano per un fenomeno di affinità differenziale e talvolta si spostano per distanze apprezzabili su scala macroscopica. La cellula confina e comunica col mondo tramite la sua membrana plasmatica. All'interno e alla periferia di tale membrana si possono incontrare molti tipi di proteine, alcune in associazione transitoria, altre invece come parte integrante della membrana stessa: è con alcune di queste proteine che la cellula aderisce ad altre cellule o a un substrato.
Esistono almeno tre classi generali di molecole di adesione (v. Gilbert, 19944): le molecole che mediano l'adesione alle altre cellule, talvolta dette collettivamente CAM (Cell Adhesion Molecules); le molecole che mediano l'adesione al substrato, talvolta dette SAM (Substrate Adhesion Molecules); e le molecole delle giunzioni cellulari, dette CJM (Cell Junctional Molecules) (v. tab. III). Le prime tengono unite le cellule nei foglietti epiteliali e promuovono la condensazione delle cellule del mesenchima. Le seconde sono implicate nell'adesione delle cellule alla matrice extracellulare e comprendono le componenti della matrice extracellulare stessa e i loro recettori presenti sulla superficie delle cellule: molecole di questa classe permettono il movimento delle cellule del mesenchima e mediano la separazione dei foglietti epiteliali. Infine, le molecole delle giunzioni cellulari assicurano la comunicazione fra il citoplasma di cellule contigue e sono alla base delle proprietà di permeabilità selettiva e di resistenza degli strati cellulari degli epiteli.
Per cambiare forma e per spostarsi la cellula ha bisogno di applicare delle forze, deve cioè poter contare su molecole capaci di esercitare una forza, sia per tirare che per spingere. La cellula trova queste forze innanzitutto nella struttura del suo citoscheletro, formato essenzialmente di tre tipi di microstrutture: ‛microfilamenti', ‛microtubuli' e ‛filamenti intermedi'. I primi, costituiti prevalentemente di actina, formano una rete interconnessa per tutto il citoplasma ma sono particolarmente addensati alla periferia della cellula, nella cosiddetta cortex cellulare; i secondi, di calibro maggiore e di maggior consistenza e rigidità dei microfilamenti, sono costituiti prevalentemente da tubulina e si irraggiano dal centro della cellula verso la periferia; infine i filamenti intermedi, più vari nella composizione e nella consistenza, si distribuiscono per tutto il citoscheletro.
La cellula può cambiare forma, cioè muoversi tenendo essenzialmente fermo il suo baricentro, o spostarsi, cioè muoversi con una traslazione del suo baricentro. Per l'uno come per l'altro movimento la cellula ha bisogno di punti d'appoggio e di connessioni fra questi e le molecole del citoscheletro: i punti d'appoggio, su altre cellule o sulla matrice extracellulare, sono forniti dalle molecole d'adesione, mentre il collegamento è assicurato da una miriade di proteine ad azione ancillare quali la talina, la vinculina, l'actinina. Queste proteine - insieme ad altre quali la spettrina, l'ancorina, l'adducina e così via - tengono insieme i microfilamenti corticali e connettono questi alla membrana plasmatica, mentre altre proteine, chiamate collettivamente MAP (Microtubule-Associated Proteins), assistono i microtubuli nella loro azione.
I motori veri e propri sono proteine che trasformano energia chimica in energia meccanica. Ad esempio, la chinesina e la dineina sono in grado di spostarsi lungo i microtubuli trascinando con sé i microorganelli e le altre microstrutture da trasportare, mentre la miosina esercita un analogo ruolo rispetto ai microfilamenti. Questi si possono contrarre quando due molecole di miosina tendono a spostarsi in direzione opposta trascinando con sé in una specie di tiro alla fune due filamenti di actina diversi: vale forse la pena di notare che questo è lo stesso meccanismo molecolare che genera la contrazione macroscopica dei muscoli. Tutte queste proteine utilizzano l'energia libera immagazzinata nelle molecole di ATP per produrre energia meccanica (v. biochimica, vol. I e biochimica, vol. X; v. bioenergetica, vol. I; v. cellula, fisiologia e patologia, vol. X; v. fosforilazione ossidativa, vol. III).
4. Strumenti concettuali sovracellulari
Lo sviluppo è un fenomeno essenzialmente sovracellulare, che si realizza attraverso una costellazione di fenomeni cellulari. Abbiamo parlato abbastanza estesamente di fenomeni cellulari e non ci resta a questo punto moltissimo da dire. Questo non significa che i processi a livello sovracellulare siano del tutto chiariti, né che non vi siano stati progressi in questo campo; anzi, i primissimi studi sono stati condotti tutti a questo livello e si è andata accumulando negli anni una mole impressionante di conoscenze su questo terreno. Da questa analisi sono emersi dei concetti che non possono essere ignorati, come quello di campo morfogenetico, di morfogeno, di gradiente, di coordinate, di mappa, di induzione e di competenza (v. Boncinelli, 1994).
a) Mappatura dei territori presuntivi e mappe del destino embriologico.
I tessuti, gli organi e i sistemi non sorgono dal nulla, bensì si originano attraverso una precisa concatenazione di eventi che possono essere fatti risalire molto indietro nello sviluppo, in alcuni casi fino allo zigote o addirittura alla cellula uovo. In ogni momento dello sviluppo, quindi, si può - almeno in linea di principio - tracciare una mappa di che cosa diventeranno i singoli territori presenti in quel momento. L'aspetto di tale mappa sarà poi tanto più simile alla mappa finale quanto minore sarà stata la migrazione cellulare durante lo sviluppo. È quindi logico che si sia cercato il modo di disegnare almeno qualcuna di queste mappe che prendono il nome, appunto, di ‛mappe dei territori presuntivi' o ‛mappe del destino embriologico'.
Per procedere a tale operazione esistono numerose metodiche che possono essere adattate ai vari sistemi e ai loro vari stadi. La procedura fondamentale consiste nell'identificare una cellula o un gruppo di cellule e osservarne quindi l'evoluzione attraverso un contrassegno di varia natura e più o meno persistente. Oggi, a seguito dell'evoluzione progressivamente registratasi nelle metodiche, si utilizzano essenzialmente due gruppi di marcatori: quello dei coloranti o traccianti vitali non tossici e quello dei marcatori cellulari naturali.
Per colorante vitale, come ad esempio il rosso neutro, si intende una sostanza che viene captata solamente dalle cellule immediatamente prossime al punto di applicazione e può essere rintracciata per qualche tempo nella sua localizzazione. In anni più recenti sono state messe a punto metodiche sempre più sofisticate, basate sull'impiego di sostanze che, microiniettate nelle singole cellule, possono poi essere individuate nelle discendenti di queste durante lo sviluppo: possono essere impiegati coloranti fluorescenti, come il DiI, o enzimi, come la perossidasi di rafano, la cui presenza viene rilevata in uno stadio più avanzato dello sviluppo mediante reazioni chimiche. Dopo alcune divisioni cellulari, però, tutti questi marcatori sono così diluiti che non è più possibile rilevarne la presenza all'interno delle cellule.
I marcatori cellulari naturali possono essere preesistenti o indotti. Le cellule di quaglia, ad esempio, possiedono un nucleo dalla morfologia affatto caratteristica per la singolare distribuzione dei nucleoli, e possono essere facilmente distinte da quelle di pollo: se si trapiantano cellule di quaglia nell'embrione di pollo se ne può seguire il destino istologico e posizionale. Moltissime delle informazioni finora raccolte sulla migrazione e sull'evoluzione delle cellule della cresta neurale negli Uccelli sono state acquisite proprio con questa metodica. Altri marcatori dello stesso tipo possono essere individuati in particolari condizioni genetiche delle cellule delle varie specie: ad esempio, approfittando dell'esistenza di un mutante che non produce un determinato enzima cellulare, si possono trapiantare cellule di questo mutante in un embrione normale e seguirne la discendenza con metodi chimici. Mediante la combinazione di questi metodi si possono ottenere le mappe dei territori presuntivi degli embrioni delle varie specie.
b) Campi morfogenetici.
Possiamo definire un ‛campo morfogenetico' come un determinato territorio dell'embrione in via di sviluppo nel quale ha luogo una serie di interazioni cellulari a lungo, a medio e a corto raggio, che portano alla formazione di una struttura organizzata, ad esempio un arto. L'esigenza di dare un nome specifico a tale territorio nasce dall'impressione che esso goda di una certa autonomia nell'ambito dello sviluppo dell'intero organismo. Questa impressione è derivata da una serie di osservazioni condotte prevalentemente sul campo morfogenetico dell'arto; lo studio dello sviluppo degli arti costituisce di per sé un interessantissimo argomento.
Trapiantando in sedi diverse frammenti prelevati da embrioni di anfibio si osservò che esistevano gruppi di cellule capaci di dare luogo a strutture specifiche come l'arto anteriore, quello posteriore, la coda, i tentacoli e le branchie. Le cellule appartenenti a questi gruppi, ribattezzati appunto campi morfogenetici, mostravano una straordinaria capacità regolativa, erano in grado, cioè, di dare origine alla struttura in questione anche se venivano sottoposte a un certo numero di manipolazioni microchirurgiche. Così, se una parte del campo veniva rimossa o se veniva introdotto in esso del materiale estraneo, l'organo si formava lo stesso. Se infine il campo veniva suddiviso in un certo numero di frammenti, si formavano più copie complete di quell'organo.
Non c'è dubbio che il concetto di campo appaia abbastanza vago e carico di connotati metafisici. Per tale motivo ha perso in tempi recenti molta della sua importanza; molti di questi fenomeni, infatti, possono essere ugualmente spiegati in termini di informazione posizionale, di gradienti e di sistemi di coordinate, concetti ritenuti, più o meno a ragione, più precisi e meglio definiti operativamente.
c) Gradienti e sistemi di coordinate.
Soffermiamoci adesso a considerare un concetto cardine della moderna biologia dello sviluppo, quello di gradiente di concentrazione di una sostanza capace di agire come morfogeno. Per ‛gradiente' si intende ovviamente una distribuzione degradante di una data sostanza in una regione specifica dell'organismo in via di sviluppo, con un'alta concentrazione a un'estremità e una pressoché nulla all'altra. Con il termine ‛morfogeno' si intende designare invece una sostanza dotata di potenzialità organizzatrici della morfogenesi, che sia distribuita secondo un gradiente e che agisca sulle singole cellule bersaglio con una modalità differenziale in qualche modo correlata con la propria concentrazione locale.
L'esistenza di un morfogeno distribuito secondo un gradiente di concentrazione è stata postulata diverse volte per spiegare la generazione della disomogeneità prima e dell'aperta diversità poi all'interno di una regione inizialmente omogenea. Dal punto di vista teorico questo problema è stato affrontato nell'ambito del cosiddetto ‛problema della bandiera francese' (French flag in inglese; v. Wolpert, 1969 e 1991). Supponiamo di voler disegnare una striscia di cellule in una posizione definita fra la testa e la coda di un animale ipotetico, indipendentemente dalle dimensioni effettive dell'animale stesso, o fra la spalla e la mano, sempre indipendentemente dalle dimensioni effettive dell'arto. È chiaro che questo equivale concettualmente a disegnare una bandiera francese, o italiana, su un drappo incolore di dimensione qualsiasi. Un modo di procedere può essere quello di tracciare un gradiente lineare di una sostanza, un ipotetico morfogeno, per tutta la lunghezza del drappo e imporre che il campo blu si estenda dall'origine al punto corrispondente, ad esempio, al 20% del valore del gradiente, il campo bianco da questo punto a quello corrispondente al 70% e il campo rosso per il resto del drappo. Questo obiettivo può essere raggiunto se la concentrazione locale del morfogeno viene letta dalle singole cellule di quel territorio con un ‛meccanismo di risposta a soglia' (v. Lawrence, 1992): laddove la concentrazione del morfogeno è inferiore a un primo valore di soglia, i vari punti saranno blu; laddove questa concentrazione è superiore al primo dei due valori di soglia, ma non al secondo, i punti saranno bianchi; infine, nella regione in cui la concentrazione è superiore anche al secondo valore di soglia, i punti saranno rossi. In questo modello la delimitazione dei tre campi dipende dall'interpretazione di un gradiente sulla base di due valori di soglia. Da notare che questa delimitazione è indipendente dalle dimensioni effettive del territorio e non implica nessuna informazione sull'effettiva distanza di ogni singolo punto, cioè di ogni singola cellula, dai due estremi del territorio stesso.
A questo schema teorico, proposto indipendentemente da ogni realizzazione materiale, si può pensare di fornire una base molecolare in termini di regolazione genica delle singole cellule esposte al morfogeno. Supponiamo che il colore dei vari campi del territorio sia dato dall'azione di due geni, che possiamo chiamare a e b. Si può allora pensare che il colore blu si ottenga per quelle cellule nelle quali entrambi i geni sono spenti, che il colore bianco caratterizzi le cellule nelle quali il gene a è acceso mentre il gene b è spento e che il colore rosso, infine, sia proprio di quelle cellule nelle quali entrambi i geni sono accesi. Con questi presupposti è facile spiegare l'origine della diversità ordinata di questa particolare bandiera francese: basta supporre che il gene a si accenda solo laddove il morfogeno è presente in una concentrazione uguale ad almeno il 20% della sua concentrazione massima e il gene b si accenda solo laddove il morfogeno è presente in una concentrazione uguale ad almeno il 70% della sua concentrazione massima. Nella prima regione entrambi i geni saranno allora spenti e si avrà il colore blu, nella seconda regione sarà acceso solamente il gene a e si avrà il colore bianco e infine nella terza regione entrambi i geni saranno accesi e si avrà il colore rosso. In virtù di un meccanismo del genere, la distribuzione graduale di un morfogeno può essere trasformata in una risposta tutto-o-nulla dei prodotti genici e quindi dei fenotipi da esso controllati. A dire il vero resta da spiegare come si possa formare un gradiente del genere. Una soluzione abbastanza semplice può essere quella secondo la quale la fonte della sostanza che agisce da morfogeno è localizzata a un'estremità del territorio, mentre per tutta la lunghezza del territorio stesso opera un meccanismo di degradazione uniforme di questa sostanza. Perché questo sia possibile occorre che il morfogeno sia capace di diffondere abbastanza liberamente all'interno del territorio.
Normalmente si rappresenta un gradiente sopra un sistema di assi cartesiani e si assume che la sua concentrazione locale specifichi l'informazione posizionale in termini di coordinate cartesiane ortogonali. Si è visto però che in alcuni casi è più comodo pensare l'informazione posizionale come specificata in termini di coordinate polari. Questo vale, per esempio, per l'arto, di pollo come di insetto. I valori posizionali possono essere specificati in termini di distanza da un centro e in termini di posizione relativa su una circonferenza: la prima dimensione viene chiamata dimensione radiale, mentre la seconda viene chiamata dimensione angolare. Questo schema concettuale si è rivelato molto utile, ad esempio nella formulazione delle regole della rigenerazione dell'arto.
d) Induzione.
Abbiamo già parlato dell'induzione come di uno dei meccanismi in base ai quali la cellula di un embrione prende le sue decisioni. Resta ancora qualche altra cosa da dire per quanto riguarda gli aspetti sovracellulari di questo fenomeno, sovracellulare per eccellenza. Infatti, anche se il suo effetto immediato è certamente esercitato sulla singola cellula, il risultato di un'interazione induttiva potrà essere apprezzato solo al livello dei tessuti o nel complesso di una o più popolazioni cellulari. Per un gioco di induzioni si determinano, si formano o si completano quasi tutti gli organi. Alcuni dei casi di induzione meglio studiati sono quelli che implicano diversi foglietti epiteliali e le cellule mesenchimatiche adiacenti: si tratta quindi di ‛interazioni epitelio-mesenchimatiche'. In questo tipo di interazioni l'epitelio può derivare da uno qualsiasi dei foglietti embrionali, mentre il mesenchima di solito deriva dal mesoderma lasso. Su questa base si forma la maggior parte delle strutture cutanee, quali peli, piume, unghie e unghielli, ghiandole e denti; i vari organi annessi all'apparato digestivo, quali il fegato, il pancreas e le ghiandole salivari; gli organi faringei, quali i polmoni, il timo e la tiroide; e infine il rene (v. Saxén e Toivonen, 1962).
Nello sviluppo dei denti, ad esempio, lo smalto viene prodotto dalle cellule del tessuto di rivestimento sotto il controllo del mesenchima della gemma dentaria. Se si trapianta quest'ultimo sotto la cute della zampetta di un embrione di topo, si osserverà in quel punto la formazione di un dente. In questo caso le cellule che avrebbero dovuto sintetizzare le proteine della cute si mettono a sintetizzare lo smalto dei denti. D'altra parte, lo strato epiteliale è capace di interpretare a modo suo alcuni dei segnali induttivi provenienti dal tessuto mesenchimatico sottostante. Si tratta appunto di un'interazione reciproca tra epitelio e mesenchima.
Consideriamo più in generale il problema della determinazione della specificità regionale nell'induzione epitelio-mesenchimatica, prendendo ad esempio la pelle. Questa è composta di due tessuti principali, un'epidermide esterna di origine ectodermica e un derma di origine mesodermica. Nel pollo si osservano almeno tre strutture cutanee diverse, originate quasi interamente dall'ectoderma di rivestimento: nelle ali ci sono le penne, nelle cosce le piume e nelle zampe squame e unghie. Il mesenchima del derma orienta la natura di queste strutture a seconda della loro posizione nel corpo: ciò si può verificare facilmente giustapponendo in varie combinazioni il derma derivato da una regione con l'ectoderma derivato da un'altra. Così l'ectoderma dell'ala dà luogo a penne, piume o squame se viene combinato con derma derivante dall'ala, dalla coscia o dalla zampa. Si tratta naturalmente di un'induzione di tipo istruttivo implicante segnali diversi provenienti dalle cellule del derma delle varie regioni del corpo, che suscitano una risposta di volta in volta diversa da parte delle cellule dell'ectoderma in termini di un'espressione differenziale di geni codificanti proteine cutanee.
Un esempio particolarmente interessante è dato dallo sviluppo del rene, argomento che da solo potrebbe meritare gli onori di un trattato. Qui ci dobbiamo limitare ad accennare all'induzione reciproca fra l'epitelio dell'abbozzo dell'uretere e il mesenchima metanefrogeno (v. Grobstein, 1967). Quest'ultimo induce l'epitelio ureterico a penetrare al suo interno, allungandosi e ramificandosi; all'estremità di queste ramificazioni, l'epitelio induce a sua volta le cellule del mesenchima circostante ad aggregarsi e a formare il tubulo renale che darà successivamente luogo al nefrone, l'unità funzionale dell'apparato filtrante renale. In assenza dell'abbozzo dell'uretere il mesenchima non si condensa e non forma tubuli; in assenza del mesenchima l'abbozzo dell'uretere non si ramifica.
5. Cenni conclusivi
Dopo la grande messe di informazioni fornite dall'embriologia descrittiva, i progressi registrati in anni a noi più vicini dall'embriologia sperimentale hanno recato un valido contributo alle nostre conoscenze sulle modalità dello sviluppo embrionale. Al punto in cui siamo sembra aprirsi la strada all'esplorazione degli intimi meccanismi che regolano le varie fasi dello sviluppo e alla comprensione della biologia dello sviluppo embrionale.
L'eccezionale prosperità che attualmente caratterizza la biologia molecolare, con il quotidiano isolamento di nuovi geni, ha già prodotto un'ampia rivisitazione in chiave molecolare della maggior parte degli eventi embriologici descritti. Tutto lascia pensare che nei prossimi anni si assisterà a una vera e propria ‛esplosione' di una embriologia molecolare.
BIBLIOGRAFIA
Boncinelli, E., Biologia dello sviluppo, Roma 1994.
Briggs, R., King, T. J., Transplantation of living nuclei from blastula cells into enucleated frogs' eggs, in ‟Proceedings of the National Academy of Sciences", 1952, XXXVIII, pp. 455-463.
Crick, F., Lawrence, P., Compartments and polyclones in insect development, in ‟Science", 1975, CLXXXIX, pp. 340-347.
Ellis, R. E., Horvitz, H. R., Genetic control of programmed cell death in the nematode C. elegans, in ‟Cell", 1986, XLIV, pp. 817-829.
Ellis, R. E., Yuan, J., Horvitz, H. R., Mechanisms and functions of cell death, in ‟Annual review of cell biology", 1991, VII, pp. 663-698.
García Bellido, A., Ripoll, P., Morata, G., Developmental compartmentalization of the wing disc of Drosophila, in ‟Nature: new biology", 1973, CCXLV, pp. 251-253.
Gilbert, S. F., Developmental biology, Sunderland, Mass., 1985, 19913, 19944 (tr. it.: Biologia dello sviluppo, Bologna 1988).
Green, J. B. A., New, H. V., Smith, J. C., Responses of embryonic Xenopus cells to activin and FGF are separated by multiple dose thresholds and correspond to distinct axes of the mesoderm, in ‟Cell", 1992, LXXI, pp. 731-739.
Grobstein, C., Mechanisms of organogenetic tissue interaction, in ‟National Cancer Institute monographs", 1967, XXVI, pp. 279-299.
Gurdon, J. B., The developmental capacity of nuclei taken from intestinal epithelial cells of feeding tadpoles, in ‟Journal of embryology and experimental morphology", 1962, X, pp. 622-640.
Gurdon, J. B., Transplanted nuclei and cell differentiation, in ‟Scientific American", 1968, CCXIX, pp. 24-35 (tr. it.: Trapianti nucleari e differenziazione cellulare, in ‟Le scienze", 1969, III, 13, pp. 26-37).
Hadorn, E., Problems of determination and transdetermination, in ‟Brookhaven symposia in biology", 1965, XVIII, pp. 148-161.
Horvitz, H. R., Herskowitz, I., Mechanisms of asymmetric cell division, in ‟Cell", 1992, LXVIII, pp. 237-255.
Ingham, P. W., Martinez Arias, A., Boundaries and fields in early embryos, in ‟Cell", 1992, LXVIII, pp. 221-235.
Lawrence, P. A., The making of a fly: the genetics of animal design, Oxford 1992.
Lewis, J. H., Wolpert, L., The principle of non-equivalence in development, in ‟Journal of theoretical biology", 1976, LXII, pp. 479-490.
Moon, R. T., Christian, J. L., Competence modifiers synergize with growth factors during mesoderm induction and patterning in Xenopus, in ‟Cell", 1992, LXXI, pp. 709-712.
Nieuwkoop, P. D., The ‟organization center" of the amphibian embryo: its spatial organization and morphogenic action, in ‟Advances in morphogenesis", 1973, X, pp. 1-39.
Saxén, L., Toivonen, S., Primary embryonic induction, London 1962.
Slack, J. M. W., From egg to embryo. Regional specification in early development, Cambridge 19912.
Spemann, H., Mangold, H., Über Induktion von Embryonalanlagen durch Transplantation artfremder Organisatoren, in ‟Wilhelm Roux Archiv für Entwicklungsmechanik der Organismen", 1924, C, pp. 599-638.
Sulston, J., Horvitz, H. R., Postembryonic cell lineages of the nematode Caenorhabditis elegans, in ‟Developmental biology", 1977, LVI, pp. 110-156.
Sulston, J., Schierenberg, J., White, J., Thomson, N., The embryonic cell lineage of the nematode Caenorhabditis elegans, in ‟Developmental biology", 1983, C, pp. 64-119.
Weintraub, H. e altri, The myoD gene family: nodal point during specification of the muscle cell lineage, in ‟Science", 1991, CCLI, pp. 761-766.
Wilson, E. B., Experimental studies on germinal localization, in ‟Journal of experimental zoology", 1904, I, pp. 1-72.
Wolpert, L., Positional information and the spatial pattern of cellular differentiation, in ‟Journal of theoretical biology", 1969, XXV, pp. 1-47.
Wolpert, L., Positional information revisited, in ‟Development", 1989, CVII, suppl., pp. 3-12.
Wolpert, L., The triumph of the embryo, Oxford 1991 (tr. it.: Il trionfo dell'embrione, Milano 1993).