
batteriofago
Particella virale submicroscopica (detta anche fago), endoparassita obbligata della cellula batterica, di cui provoca la lisi. I b. sono stati raggruppati in sei tipi differenti (A, B, C, D, E, F).
Come i virus, i b. sono costituiti da DNA o da RNA e sono circondati da un rivestimento proteico, il capside. Il tipo A, il più complesso, presenta una morfologia allungata, con una testa a struttura icosaedrica, un collare, una coda costituita da una struttura tubulare cava avvolta da una guaina contrattile e, infine, da una piastra esagonale connessa con sei lunghe fibre. Le dimensioni dei b. sono variabili; alcuni possono essere molto piccoli e contenere 5 o 6 geni, altri molto grandi, e contenerne più di 100.
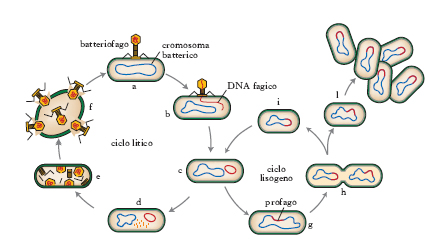
In base al diverso ciclo biologico si distinguono b. virulenti e b. temperati. In entrambi i casi il ciclo vitale (v. fig.) inizia con l’adesione del b. alle cellule batteriche ospiti, mediante proteine virali che riconoscono recettori specifici delle superfici batteriche. Successivamente l’acido nucleico viene iniettato all’interno della cellula batterica. La moltiplicazione dei b. virulenti dà luogo a un ciclo litico; l’acido nucleico si replica subito dopo il suo ingresso e se ne formano centinaia di copie. Alla sintesi delle proteine capsidiche segue la fase di assemblaggio tra i genomi e le strutture proteiche virali, per formare particelle fagiche complete infettanti che sono rilasciate dopo la lisi della cellula batterica. I b. temperati, contenenti sempre DNA a doppio filamento, possono passare attraverso un ciclo litico, oppure attraverso un ciclo alternativo definito ciclo vitale lisogeno. In questo caso i geni del b. sono repressi e l’acido nucleico virale viene integrato nel cromosoma del batterio ospite. Il genoma fagico integrato viene definito profago, i batteri che contengono il profago si chiamano batteri lisogeni e la situazione di integrazione viene definita lisogenia.
Il batterio lisogeno può trasportare uno o più geni che influenzano la virulenza del batterio ospite: per es. Corynebacterium diphteriae causa la difterite nell’uomo solo quando è presente come profago il fago β che codifica la produzione dell’esotossina difterica. Anche la produzione di tossine da parte di Clostridium botulinum e di Streptococcus pyogenes e le modificazioni antigeniche riscontrabili in alcune salmonelle e shigelle sono riconducibili a geni del b. i quali non risentono dei processi che determinano la loro repressione e sono stati incorporati nel DNA dell’ospite per ricombinazione. Nella seconda metà degli anni 1990, nell’ambito del Progetto genoma (➔ genoma), sono stati sequenziati numerosi genomi batterici e virali. Uno dei risultati più sorprendenti è stata proprio la dimostrazione della grande capacità di trasferimento di geni in microrganismi differenti spesso segnalata dalla presenza di tipiche sequenze di DNA. Molti di questi tratti contengono sequenze simili a noti geni di batteriofagi.
I batteriofagi presentano uno spettro di specificità d’ospite molto limitato, anche nell’ambito di una stessa specie batterica. Tale proprietà ( tipizzazione fagica), riconosciuta in laboratorio osservando la presenza della lisi dei batteri, viene utilizzata come strumento diagnostico. Una sua applicazione pratica riguarda, per es., le indagini epidemiologiche delle infezioni ospedaliere da stafilococchi. I b. sono anche in grado di trasferire l’informazione genetica da una cellula batterica a un’altra (➔ trasduzione).