
cellula
Biologia
Unità morfologica e fisiologica elementare di tutti gli organismi animali e vegetali; questi possono essere costituiti da una o più cellule. Il termine c. è stato introdotto in biologia nel 1665 da R. Hooke, che per primo osservò le c. al microscopio; furono poi M. Schleiden e T. Schwann, nel 1838-39, a definire la c. come unità fondamentale dei viventi (teoria cellulare).
I metodi di studio della struttura e della fisiologia cellulare sono ormai molto raffinati; al classico microscopio ottico, con un potere di risoluzione di 0,2 mm, si è affiancato il microscopio elettronico, con potere di risoluzione di 10 Å; per lo studio della chimica e della fisiologia cellulare ci si avvale di metodi come la spettrofotometria, la diffrazione ai raggi X, la centrifugazione frazionata e l’autoradiografia.
Struttura e fisiologia della cellula
L’organizzazione interna e il movimento delle c. eucariotiche dipendono da una complicata rete di filamenti proteici presenti nel citoplasma che hanno funzioni statiche e dinamiche e che costituiscono, nel loro complesso, il citoscheletro: filamenti di actina, microtubuli, filamenti intermedi. Ogni c. conserva, sviluppa e riproduce la sua organizzazione in condizioni fisiche rigorosamente definite; infatti tutte le sue reazioni chimiche sono sottoposte a catalisi e regolazione da parte di molecole che sono sostanzialmente le stesse nell’uomo come nel batterio.
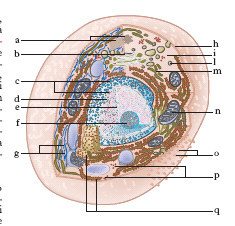
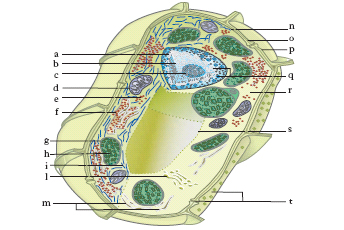
Organizzazione
Ogni c. è delimitata dalla membrana plasmatica o cellulare, al cui interno si trova il citoplasma, in cui sono presenti numerosi organelli cellulari: il reticolo endoplasmatico granulare o ergatoplasma e quello liscio o agranulare, i ribosomi, l’apparato di Golgi, i mitocondri, i lisosomi, i centrioli e, limitatamente alle cellule vegetali, i plastidi (fig. 1 e 2). Tutte le c. si possono suddividere in procariotiche ed eucariotiche.
Le c. procariotiche (tipiche di Batteri, Cianofite) non hanno un vero nucleo delimitato da membrana e il DNA è libero nel citoplasma.
Le c. eucariotiche possiedono invece una regione nucleare differenziata, separata dal citoplasma mediante una membrana nucleare. Inoltre i procarioti non vanno incontro al caratteristico processo di divisione cellulare, chiamato mitosi, che è proprio delle c. eucariotiche.
Specializzazione
Alcuni organismi viventi sono costituiti da un’unica c. (protozoi e protofiti); gli organismi pluricellulari (metazoi e metafiti) vanno invece dalle poche decine di c. degli organismi inferiori ai miliardi di c. di quelli superiori. Nei metazoi e nei metafiti le c., variamente differenziate in rapporto al lavoro cui sono adibite, si associano formando i tessuti, che compongono i diversi organi. Le c. di un tessuto differiscono da quelle di un altro tessuto in quanto, mediante un processo chiamato differenziamento, assumono caratteristiche morfologiche e funzionali specifiche che le rendono adatte a compiere una funzione specializzata, diversa da tessuto a tessuto. Le dimensioni delle c. sono molto varie; si va dai 20 μm di lunghezza di Escherichia coli, fino ai parecchi centimetri della c. uovo di struzzo (7,5 cm). L’alga verde Acetabularia è formata da un’unica c. che può raggiungere i 10 cm di diametro. Alcune c. sono dotate di movimento: il movimento ameboide, tipico dei protozoi del genere Amoeba, avviene per mezzo di deformazioni della superficie cellulare che permettono alla cellula di strisciare sul substrato, altre c. per muoversi si servono, invece, di ciglia o di flagelli, come nel caso degli spermatozoi.
Composizione
L’analisi chimica dei componenti cellulari rivela la presenza di un’alta percentuale di acqua, che varia da poco più del 50% fino al 98%, come nel caso delle meduse. Tra i costituenti chimici principali delle cellule vi sono: a) le proteine, divise in proteine strutturali, che formano l’ossatura dell’edificio cellulare, e gli enzimi, che svolgono la funzione di catalizzatori biologici, regolando e accelerando le reazioni biologiche; b) gli zuccheri polisaccaridi, dalla cui degradazione viene ricavata l’energia necessaria alla vita della cellula; c) i grassi, che costituiscono una riserva di energia; d) gli acidi nucleici, infine, che controllano i meccanismi ereditari della cellula. Nella c. si trovano, inoltre, molecole organiche più semplici e ioni inorganici.
Il nucleo
Il nucleo è la regione differenziata del citoplasma, delimitata da una propria membrana. Questa membrana è formata da un differenziamento locale del reticolo endoplasmatico ed è regolarmente attraversata da pori di diametro variabile fra 250 e 1000 Å; questi pori non consentono un passaggio indiscriminato di sostanze, ma rappresentano un vero e proprio filtro. In genere le c. sono mononucleate, ma esistono c. bi- e polinucleate; alcune forme viventi sono costituite da una massa citoplasmatica indivisa, in cui sono immersi numerosi nuclei: una struttura di questo genere viene chiamata sincizio. Nel nucleo delle c. eucariotiche, il messaggio ereditario, contenuto nel DNA, viene trascritto su molecole di RNA messaggero (RNAm) che migra nel citoplasma e viene tradotto in catena polipeptidica a livello dei ribosomi; a quest’ultimo processo partecipano altri due tipi di RNA: l’RNA ribosomale (RNAr) e l’RNA di trasporto (RNAt), che trasporta ai ribosomi gli amminoacidi attivati.
Membrane
Gli ultimi due decenni del 20° sec. sono stati testimoni di un incremento esplosivo nella comprensione delle proprietà fondamentali delle c. eucariotiche e soprattutto delle loro interazioni negli organismi pluricellulari. Le c. utilizzano una varietà di organuli e di componenti cellulari per portare a termine le loro principali funzioni, ma sempre di più emerge che le più importanti attività cellulari coinvolgono eventi che si verificano all’interno o sulla superficie di un complesso sistema di membrane. Le membrane, oltre a circondare la superficie della c. e il nucleo determinano una fondamentale compartimentazione del citoplasma. Tutti gli eventi che si verificano a livello di questo complesso sistema di membrane sono mediati da proteine con ruoli e funzioni diversi: complessi multienzimatici, nei quali la posizione spaziale dei diversi componenti è determinante per la loro funzione di catalizzatori dei processi del metabolismo; recettori e trasduttori di segnali; proteine coinvolte nella secrezione cellulare e nell’endocitosi (➔ clatrina); proteine integrali di membrana, necessarie al trasporto di molecole e ioni all’interno e all’esterno della c.; proteine di adesione, necessarie per il riconoscimento e l’interazione fra più c. specializzate per la formazione dei tessuti durante lo sviluppo (➔ adesione). È inoltre da notare che, mentre in alcuni tessuti, per es. negli epiteli, la maggior parte delle c. stabilisce stretti contatti con le c. che la circondano, mediante diversi tipi di giunzioni (➔ connessina), in altri tessuti le c. secernono una serie di macromolecole che formano una complessa rete di proteine e carboidrati che costituisce la matrice extracellulare. Tale matrice, oltre ad avere una funzione di supporto strutturale, influenza le dimensioni, la motilità e la crescita delle c. e, talvolta, soprattutto nelle prime fasi dello sviluppo embrionale, costituisce un substrato attraverso il quale le c. si muovono (➔ matrice).
Progressi della biologia della cellula
Molte informazioni si hanno oggi sul ruolo di DNA e RNA nel flusso dell’informazione genetica: sono stati meglio chiariti soprattutto i processi di regolazione dell’espressione dei geni (➔ regolazione, epigenesi, silenziamento). Le scoperte più importanti della biologia della c. sono state spesso realizzate in seguito all’introduzione di nuove tecniche o alla modificazione di tecniche già sperimentate. La microscopia rimane un punto di partenza essenziale ed è ancora uno strumento insostituibile in questo campo insieme con le tecniche più recenti, basate sull’impiego della radiazione di sincrotrone. Le c. si possono isolare dai diversi tipi di tessuti di appartenenza e far crescere in vitro: da queste c. si possono isolare gli organelli in forma pura e quantificare e isolare i singoli tipi di molecole e di ioni presenti. La tecnica più sofisticata di separazione cellulare, che permette di ottenere popolazioni omogenee, prevede la marcatura di c. specifiche con anticorpi accoppiati a una sostanza fluorescente. Le c. marcate vengono separate da quelle non marcate in un separatore cellulare attivato dalla fluorescenza (➔ citometria, FACS). Il frazionamento delle c. permette inoltre di ottenere filtrati acellulari, contenenti molecole funzionanti. Tali filtrati sono stati e continuano a essere utilizzati per studi sulla sintesi proteica, la replicazione del DNA, lo splicing dell’RNA, il ciclo cellulare, la mitosi, il movimento delle vescicole dell’apparato di Golgi e vari tipi di trasporto intracellulare.
Sono disponibili numerose tecniche per rivelare e localizzare qualunque molecola in una cellula. Ogni molecola può essere infatti marcata mediante incorporazione di uno o più atomi radioattivi o di sostanze fluorescenti. L’emissione di radiazioni ionizzanti o di luce fluorescente permette di rivelare la molecola, di seguirne i movimenti o di risalire alla concentrazione. Importante è l’uso di microelettrodi di vetro sia per studiare i potenziali elettrici e le correnti ioniche attraverso la membrana plasmatica, sia per misurare la concentrazione di specifici ioni intracellulari e per iniettare in c. vive molecole, quali proteine marcate con sostanze fluorescenti e anticorpi. Il comportamento dinamico di molti tipi di molecole all’interno delle c. può essere anche seguito costruendo un precursore che, dopo essere stato introdotto, viene attivato in una regione selezionata della c. da una reazione stimolata dalla luce.
Una rilevante innovazione per la comprensione delle funzioni cellulari è derivata sicuramente dalla creazione degli anticorpi monoclonali che, teoricamente, possono essere usati per localizzare e purificare qualunque macromolecola e, più in generale, dalle tecnologie del DNA ricombinante che hanno permesso di manipolare l’informazione genetica della cellula.
Ciclo cellulare
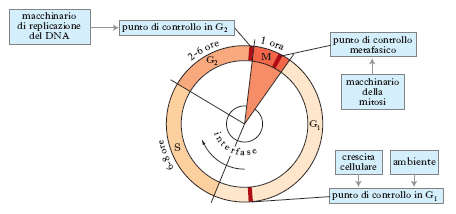
Nella cellula eucariotica la serie di eventi che avvengono tra una mitosi e quella successiva costituisce il ciclo cellulare. Durante l’interfase, che precede la mitosi (detta fase M), si susseguono (fig. 3) tre fasi rispettivamente denominate G1, S, G2 (dove G sta per gap «intervallo» e S sta per sintesi). La fase S è la fase di sintesi del DNA e di duplicazione dei cromosomi. La fase G1 è generalmente più lunga e variabile mentre la G2 è la più breve; in alcuni casi le cellule passano direttamente dalla fase S alla profase mitotica, saltando la G2. Il ciclo cellulare nei Procarioti consiste nella replicazione del DNA, seguita immediatamente dalla divisione cellulare. Per es., Escherichia coli si divide in circa 30 minuti e tutto il tempo, eccetto pochi minuti, è utilizzato per completare la sintesi del DNA.
Nei Mammiferi, le differenze nel tasso di divisione cellulare sono in gran parte dovute a diversità nella durata del periodo che intercorre fra l’entrata e l’uscita dalla fase G1. Questa variabilità è determinata da una occasionale fase di quiescenza, G0, nella quale le cellule della fase G1 possono entrare o rimanere per periodi che variano a seconda del tipo di cellula e delle condizioni ambientali; le altre fasi del ciclo (S, G2 e M) hanno invece una durata fissa.
Utilizzando vari metodi, è stata effettuata un’analisi accurata della relazione fra la sintesi del DNA e la divisione cellulare nelle cellule eucariotiche e si è osservato che ogni stadio del ciclo è caratterizzato da varie attività di sintesi. La fase M del ciclo delle cellule somatiche diploidi corrisponde alla mitosi, che assicura l’uguaglianza genetica delle cellule figlie, mentre nella produzione delle cellule germinali aploidi (spermatozoo, uovo) la fase M è la meiosi, che assicura la continuità genetica tra le generazioni negli organismi pluricellulari. Durante la fase G1 vengono sintetizzati le proteine, i carboidrati e i lipidi caratteristici della cellula. Anche se l’assemblaggio di tutti i tipi di molecole contenute nella cellula continua in S e G2, la fase S è l’unica in cui avvengono sia la replicazione del DNA, sia la sintesi delle proteine dei cromosomi (istoni e proteine acide). La fase G2 è caratterizzata invece dalla sintesi di un gruppo di proteine necessarie per la progressione attraverso la mitosi. Negli Eucarioti inferiori, quali il lievito, il momento critico della tarda fase G1, in cui avviene l’attivazione della fase S, viene chiamato start; negli Eucarioti superiori, questo momento, nel quale le attività della cellula sono ristrette alla divisione, viene invece chiamato punto di restrizione (R).

A partire dalla seconda metà degli anni 1980 sono stati compiuti progressi consistenti nella ricerca e nell’identificazione dei geni coinvolti nella regolazione del ciclo cellulare, utilizzando i lieviti Saccharomyces cerevisiae e Saccharomyces pombe, in cui sono state identificate circa 150 mutazioni in più di 30 geni diversi, chiamati complessivamente Cdc (Cell division cycle). Alla luce degli studi effettuati negli anni successivi, il sistema di controllo del ciclo nelle c. di mammifero si è rivelato molto più complesso, anche se sono state meglio comprese alcune strategie comuni. La progressione del ciclo cellulare viene sostanzialmente effettuata sotto la spinta di reazioni biochimiche sostenute dall’attivazione di molteplici chinasi chiamate Cdk (Cycline dependent kinase), a opera di proteine note come cicline. Una volta attivato, ciascun complesso Cdk-ciclina fosforila specifiche proteine. La ciclina, infatti, riconosce e si lega a una proteina bersaglio, che può così essere fosforilata dalla Cdk. La proteina bersaglio fosforilata non è più in grado di rimanere legata alla ciclina e viene rilasciata dal complesso (fig. 4). Dato che nelle varie fasi del ciclo sono presenti cicline differenti, ogni fase è caratterizzata dalla fosforilazione di proteine bersaglio differenti.
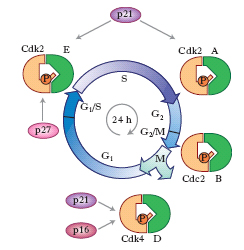
Oltre ai fattori positivi che promuovono la progressione ordinata delle c. attraverso il ciclo, quali le cicline e le specifiche Cdk, gli studi degli anni 1990 hanno portato all’identificazione di fattori negativi che arrestano il ciclo stesso. I fattori negativi, noti come inibitori delle chinasi dipendenti dalla ciclina (CKI), quali p16, p21 e p27, inattivano i complessi Cdk-ciclina (fig. 5). Talvolta possono essere inattivati anche mediante processi diversi quale, per es., la degradazione della ciclina causata dal sistema di proteolisi della ubiquitina. Queste complesse reazioni biochimiche caratterizzano punti cruciali nel ciclo, detti punti di controllo, che sorvegliano la progressione normale delle cellule. Le varie fasi presentano uno o più punti di controllo cellulare, dove il ciclo si può arrestare finché non viene corretto l’eventuale difetto per impedire che sia trasmesso alle c. figlie.
Alcuni tipi cellulari
C. alfa e beta (o cellule A e B)
Nel pancreas c. degli isolotti di Langerhans che secernono, rispettivamente, il glucagone e l’insulina.
C. fisalifore
Grosse c. con numerosi vacuoli, contenenti glucogeno e grasso, caratteristiche del cordoma.
C. giganti
Elementi fagocitari forniti di numerosissimi nuclei, di dimensioni relativamente gigantesche, che compaiono quando in un tessuto infiammato vi siano corpi estranei (schegge, fili di seta ecc.) o microrganismi resistenti alle comuni difese organiche (per es. bacilli della tubercolosi, treponemi della sifilide ecc.). Le c. giganti entrano a far parte quindi dei granulomi in genere; c. di Langhans sono dette quelle del granuloma tubercolare; c. giganti di Sternberg quelle del linfogranuloma maligno ecc. Caratteri diversi hanno le c. giganti di tumori (➔ sarcoma).
C. immunocompetenti
C. impegnate nel riconoscimento dell’antigene e nella risposta a esso.
C. di Hargraves
Granulociti neutrofili contenenti abbondanti inclusioni costituite da materiale nucleico e anticorpi antinucleo.
C. mitrali
Le c. piramidali che formano uno degli strati del bulbo olfattorio. C. mitrali (o cardiache) sono anche dette le c. che si rinvengono nell’escreato emorragico dei cardiopatici mitralici, con fenomeni di stasi polmonare.
Cellule RA
Dette anche ragociti, queste c. sono granulociti oppure c. monocitoidi contenenti inclusioni citoplasmatiche e si rinvengono nel liquido articolare nel corso di artropatie infiammatorie.
Fisica
C. fotoelettrica Denominazione generica di dispositivo che, sfruttando uno dei fenomeni della fotoelettricità, converte variazioni di una grandezza luminosa (illuminamento di una superficie) in variazioni di una grandezza elettrica (tensione). A seconda del fenomeno fotoelettrico che è alla base del loro funzionamento, le c. fotoelettriche si distinguono in fotoconduttive, fotoemissive, fotovoltaiche. Molte, e di notevole importanza, le applicazioni: quelle relative ai dispositivi automatici a comando luminoso, alla registrazione ottica dei suoni (cinematografia sonora), alla conversione di energia luminosa in energia elettrica, alla fotometria (in particolare astronomica) e alla spettroscopia.
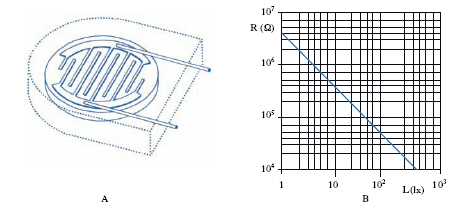
La c. fotoconduttiva (o fotoconduttrice) è basata sul fenomeno della fotoconduzione, cioè sulla variazione della resistività di certe sostanze, in funzione dell’intensità e della lunghezza d’onda della radiazione da cui sono colpite; inserita in un circuito chiuso alimentato e provvisto di uno strumento amperometrico, permette la determinazione dell’intensità della radiazione incidente per via elettrica. È di solito costituita da un sottile strato (0,5-50 μm) di materiale fotoconduttivo (silicio, arseniuro o antimoniuro di gallio, antimoniuro di indio, seleniuro di cadmio) deposto su una lastrina isolante; la resistenza fra i due elettrodi, in genere del tipo interdigitato a pettine (fig. 6A), diminuisce in maniera non lineare all’aumentare dell’illuminamento dello strato fotosensibile (fig. 6B), per radiazioni di lunghezza d’onda inferiori a un determinato valore critico caratteristico del materiale (soglia di fotoconduttività). Le variazioni di resistenza seguono con un certo ritardo (dell’ordine dei millesimi o decimillesimi di secondo) quelle dell’illuminamento, la sensibilità è in genere massima nel rosso o nell’infrarosso vicino.

La c. fotoemissiva è basata sull’effetto fotoelettronico di superficie, cioè sull’emissione di elettroni dalla superficie di alcuni materiali colpiti da radiazioni di appropriata lunghezza d’onda. È costituita da un’ampolla di vetro, a vuoto spinto oppure riempita da un gas inerte a bassa pressione, entro la quale sono due elettrodi metallici a differente potenziale elettrico. Inviando raggi luminosi sull’elettrodo a potenziale minore (fotocatodo), se la lunghezza d’onda della luce è minore della soglia fotoelettrica del fotocatodo (➔ fotoelettrone) da quest’ultimo si distaccano elettroni (fotoelettroni) che, accelerati dal campo elettrico esistente tra gli elettrodi, si dirigono verso l’elettrodo a potenziale maggiore (anodo) e sono da questo raccolti. Si dà il nome di caratteristica di accelerazione della c. al diagramma dell’intensità, ia, della corrente di elettroni raccolti dall’anodo (corrente fotoelettrica) in funzione della tensione, Va, tra anodo e fotocatodo (fig. 7), tenendo costante l’illuminamento di quest’ultimo. Tale caratteristica, che naturalmente è asimmetrica (ia è nulla per Va ⟨ 0), è non lineare. Per valori relativamente bassi di Va vi è un effetto di carica spaziale a opera dei fotoelettroni, tale che ia cresce al crescere di Va; a partire da un certo valore della tensione, l’effetto di carica spaziale viene meno, tutti i fotoelettroni pervengono all’anodo, e ia si mantiene costante: si dice che la c. è allora in regime di saturazione. Ulteriori aumenti di ia potranno ottenersi aumentando l’illuminamento o, che è lo stesso, il flusso luminoso L sul fotocatodo. Al diagramma di ia in funzione di L, rilevato in regime di saturazione, si dà il nome di caratteristica di illuminazione della c.; con riferimento a tale caratteristica, si definisce sensibilità della c. il rapporto ia/L.
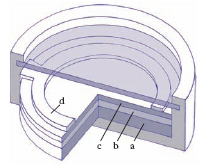
La c. fotovoltaica è basata sull’effetto fotovoltaico (➔ fotovoltaico, effetto) tra materiali diversi, cioè sul fatto che sotto l’azione della luce si genera una forza elettromotrice nella zona di contatto (giunzione) tra un metallo e un semiconduttore (barriera Schottley) oppure tra un semiconduttore di tipo p e uno di tipo n o anche tra un liquido e un semiconduttore (c. fotoelettrochimica). È costituita da una lamina b (fig. 8) di semiconduttore, sorretta da un elettrodo a, sulla quale è depositato uno strato sottilissimo, c, trasparente, di metallo oppure di semiconduttore diverso dal primo, su cui appoggia un secondo elettrodo, d, costituito da un anello opportunamente foggiato: b può essere selenio, che è un semiconduttore di tipo p, e c argento o alluminio (c. a selenio), ovvero di silicio p e di silicio n, rispettivamente (c. a silicio). La forza elettromotrice dipende dalla natura delle due sostanze che danno luogo all’effetto fotovoltaico, dall’illuminamento della giunzione e dalla lunghezza d’onda della radiazione illuminante: alla piena luce solare la forza elettromotrice di una c. a selenio è di circa 0,3 V, quella di una c. a silicio intorno a 0,6 V. L’intensità della corrente che una c. può erogare quando sia chiusa su un circuito dipende anche dall’area della superficie illuminata e dalla resistenza del circuito: quest’ultima, inoltre, influenza fortemente la caratteristica di illuminazione, cioè il diagramma dell’intensità della corrente in funzione dell’illuminamento della giunzione, nel senso che tale caratteristica risulta lineare soltanto per resistenze relativamente basse. Per la conversione dell’energia solare, ottenendo tensioni relativamente alte e cospicue intensità di corrente, più c. identiche vengono collegate in serie-parallelo tra loro e montate su appositi pannelli, anche di notevoli dimensioni superficiali; il costoso silicio monocristallino è spesso sostituito con strati sottili di silicio amorfo o policristallino, arseniuro di gallio, solfuro di cadmio o anche altri composti. Per quel che riguarda il rendimento di tali dispositivi, cioè il rapporto tra l’energia elettrica generata e l’energia della radiazione incidente (circa 1,36 kW/m2 fuori dall’atmosfera, intorno a 1 kW/m2 sulla superficie terrestre a mezzogiorno in una giornata serena), si è intorno al 12-14% per quelli commerciali policristallini e al 4-8% per le c. commerciali a silicio amorfo. Ricorrendo a opportuni metodi ottici (specchi concentratori o lenti di Fresnel) di intensificazione della radiazione incidente, si sono ottenuti rendimenti del 25% con c. all’arseniuro di gallio; grazie a prototipi di laboratorio con particolari strutture multigiunzione sono stati raggiunti rendimenti fino al 40%.