
diversità
Biologia
Diversità anticorpale In immunologia, la varietà di anticorpi che il sistema linfatico è in grado di produrre verso gli antigeni del mondo esterno all’organismo.
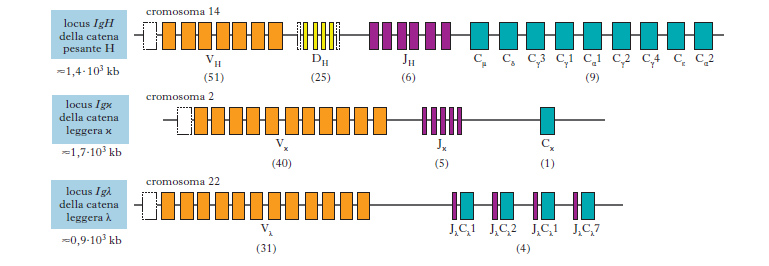
Le molecole di anticorpo vengono sintetizzate da linfociti B attivati. La struttura di un anticorpo viene definita da informazioni geniche localizzate su tre loci distinti presenti in cromosomi diversi: 14 (per le catene pesanti H, heavy), 2 (per le catene leggere χ) e 22 (per le catene leggere λ). La struttura base dei tre loci è simile; essi infatti codificano per le regioni variabili (V) e costanti (C) delle rispettive catene che costituiranno l’anticorpo (fig. 1). Gli aspetti caratteristici dei geni riguardano i processi di riarrangiamento che si realizzano nel tempo della differenziazione della cellula linfocitaria B, con ridistribuzione attraverso meccanismi casuali dei segmenti genici stessi. In tal modo è possibile interpretare l’ampia variabilità degli anticorpi. In particolare, nel corso degli studi sulle fasi del riarrangiamento genico, si è potuto interpretare il complesso dei fenomeni che consentono agli anticorpi di combinarsi con le varianti antigeniche possibili.
L’analisi della sequenza primaria della regione anticorpale che si combina con l’antigene (sito combinatorio) e gli studi sulle altre componenti della molecola hanno descritto alcuni aspetti strutturali, tra i quali risulta fondamentale l’identificazione delle regioni variabili V codificate da segmenti genici diversi (processo di riarrangiamento che si verifica nelle prime fasi della differenziazione linfocitaria). Per le catene pesanti (H) sono noti tre segmenti V: VH (variabilità), D (d.), JH (giunzione). Per le catene leggere (L, light) i geni per le regioni V (χ e λ) sono simili a quanto descritto per la catena H, mentre è assente il segmento D. Gli anticorpi sono formati da due catene che consentono almeno 10 specificità differenti. Poiché ogni catena è costituita da una regione variabile e una costante, la combinazione casuale delle due catene H e L consente un notevole risparmio di DNA. Del resto il vantaggio si consolida nella natura stessa della risposta all’antigene, perché quest’ultimo è costituito da più determinanti che possono essere identificati da anticorpi a diversa affinità. Si deve ricordare come la regione variabile delle catene H e L sia composta da sequenze ipervariabili e da strutture costituite, nell’ambito del sito combinatorio, da sequenze costanti.
Nella struttura di un’immunoglobulina anticorpale confluiscono le informazioni derivate dall’assemblaggio modulare dei geni che codificano le catene pesanti e le catene leggere. Prescindendo dalle componenti costanti delle catene leggere (χ e λ) e pesanti (μ, α, γ, δ, ε), le quali identificano la struttura che non include il sito combinatorio, quest’ultimo deriva da una serie di eventi che si realizzano durante il processo di differenziazione del linfocita B secondo le tappe che seguono: un frammento genetico V si associa al frammento J e il binomio VJ si associa alla frazione costante (strutturazione della catena leggera), mentre nella catena pesante si inserisce, tra V e J, il frammento D.
In linea teorica, il riarrangiamento può avvenire su ogni cromosoma dotato dei geni per la struttura dell’anticorpo e ogni linfocita B potrebbe dar luogo a geni attivati per codificare due catene pesanti H, due leggere χ e due leggere λ. In realtà solo uno dei due alleli (quello di origine materna o paterna) viene riarrangiato e trascritto, mentre l’altro evolve verso un riarrangiamento abortivo (che non è trascrivibile) oppure non viene riarrangiato. Si verifica così per le cellule B quel fenomeno relativamente raro definito di esclusione allelica, caratteristico delle cellule della femmina, in cui viene espresso uno dei due alleli dei geni presenti sul cromosoma X. Si deve inoltre aggiungere come in ogni cellula B si osservi un arrangiamento efficace di un gene χ o di un gene λ, ma in nessun caso di entrambi. Quindi una molecola anticorpale, a completamento della sua sintesi, presenta due catene pesanti (H) con definito isotipo, mentre le due catene leggere sono entrambe χ o λ.
Il repertorio clonale dal quale ha luogo la generazione della d. anticorpale è così il risultato di vari meccanismi che, sommandosi, sono in grado di giustificare la diversità stessa:
a) carattere proprio della linea germinale con le varie combinazioni delle catene pesanti e leggere;
b) opzioni combinatorie (ricombinazione genetica) in segmenti genici che codificano le regioni variabili;
c) mutazioni somatiche (circa il 5% del totale).
Ecologia
La d. di una comunità è la valutazione del numero di specie che sono presenti nella comunità stessa ( ricchezza in specie) e della loro rappresentatività relativa, in termini di numero di individui per specie ( equipartizione, in inglese equitability, o uniformità, in ingl. evenness) e viene misurata tramite indici. In ciascuna comunità, solo una piccola percentuale delle specie presenti è rappresentata da numerosi individui, mentre la gran parte delle altre è rappresentata da pochi individui. Le poche specie rappresentate da molti individui, dette dominanti, contribuiscono in maniera determinante alla struttura e alla funzionalità della comunità (per es., in esse passa la gran parte del flusso di energia); le altre, dette specie rare, sono invece determinanti per la d. della comunità. La d. può essere determinata in un’area ristretta, e allora si parla di d. locale o d. alfa, oppure può essere determinata in un’area geografica di maggiori dimensioni, e si parla di d. regionale o d. gamma, o ancora può essere determinata nelle zone di transizione (➔ ecotone) tra ecosistemi diversi (per es., tra un ecosistema terrestre, un prato, e uno acquatico, un fiume), e si ha la d. beta. La d. viene valutata quantitativamente tramite gli indici di d., in grado di dare informazioni sui rapporti funzionali tra le varie popolazioni e tra queste e il loro ambiente, e di indicare se un dato sistema si trova in uno stadio giovane o maturo, di equilibrio o di stress. Lo studio della d., con le sue regolarità nelle relazioni tra il numero di individui per specie e il numero delle specie presenti in un determinato contesto ambientale, permette quindi di caratterizzare l’ecosistema nei vari momenti della sua esistenza. Il concetto di d. è dunque di grande interesse e utilità sia per l’ecologia teorica sia per quella applicata.
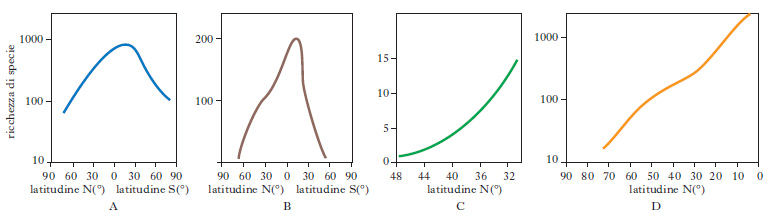
La d. è necessaria per il mantenimento della struttura complessa degli ecosistemi: infatti l’organizzazione in reti alimentari e tutte le altre relazioni interspecifiche della componente biotica di un ecosistema dipendono da una certa regolarità numerica del sistema, che viene mantenuta tramite i meccanismi di autoregolazione dell’ecosistema stesso. La d. varia durante l’evoluzione delle comunità dagli stadi pionieri a quelli più maturi: negli ecosistemi giovani la d. è più bassa, con predominanza di una o poche specie; man mano che il sistema matura la d. aumenta, fino a una distribuzione regolare delle specie. La fase pioniera può corrispondere a una nuova fase di colonizzazione, all’effetto di una perturbazione oppure a una variazione rapida dell’ambiente. La d. varia anche lungo gradienti geografici: nella maggior parte dei casi essa cresce andando dai poli verso l’equatore (fig. 2); sono rari i gradienti in direzione opposta. Al diminuire della latitudine aumenta la disponibilità di microambienti diversi in un sito, che permette l’insediamento di un maggior numero di organismi diversi. La progressiva alterazione antropica della biosfera in futuro comporterà per le comunità terrestri da una parte l’aumento della d. beta, dovuto, per es., a un maggiore tasso di mortalità delle singole popolazioni (che provoca un aumento della struttura a mosaico del territorio il quale, a sua volta, facilita la colonizzazione da parte di nuove specie); dall’altra, il decremento della d. gamma, se l’incrementato disturbo permetterà, su scala globale, solo la colonizzazione da parte di specie caratteristiche delle prime fasi delle successioni.
L’aumento di d. dai poli verso i tropici può essere riconosciuto in numerosi gruppi di organismi. Si riscontra sia negli ecosistemi terrestri, sia in quelli marini e d’acqua dolce; è evidente non solo a livello di ampie regioni geografiche, ma anche nelle comunità di piccole dimensioni. Le spiegazioni date a questo fenomeno sono numerose: è stato correlato all’aumento di produttività che si osserva andando dai poli verso l’equatore; alla maggiore intensità di predazione nelle zone tropicali rispetto a quelle più polari; al clima, cioè la minore variazione stagionale di molti fattori ambientali andando dai poli verso i tropici; alla maggiore età evolutiva delle regioni tropicali rispetto a quelle polari. Esistono importanti eccezioni all’andamento latitudinale, non giustificate dalle precedenti spiegazioni: per es., le isole o i deserti, poco produttivi e poco diversificati, sebbene vicini ai tropici; le sorgenti calde e le lagune, poco diversificate sebbene molto produttive.