
evoluzione
Biologia
E. biologica
Il processo di cambiamento adattivo per il quale tutte le specie viventi – e solo queste – mutano di generazione in generazione per adattarsi all’ambiente in continua trasformazione.
Teorie dell’evoluzione
L’idea che le specie viventi mutino nel tempo è relativamente recente. Il famoso aforisma di Linneo, il fondatore della moderna sistematica, «species tot numeramus, quot ab initio numeravit Infinitum Ens» mostra come ancora nel Settecento si sostenesse nella maniera più categorica la fissità delle specie viventi: fissità che si rifaceva alla lettera del dettato biblico. Teorie evolutive furono accennate, sempre nel 18° sec., da P.-L. Moreau de Maupertuis e G.-L. Leclerc de Buffon, il quale fu tuttavia costretto a ritrattare le sue idee quando furono criticate dalla facoltà di Teologia della Sorbonne di Parigi. Teorie organiche dell’evoluzione furono formulate soltanto a partirte dagli inizi del 19° sec. da J.-B. de Lamarck in Francia e, quasi contemporaneamente in Inghilterra, da Erasmus Darwin, nonno di Charles, e dominarono il campo della biologia dopo l’uscita, nel 1859, dell’Origine delle specie di quest’ultimo. Una teoria identica a quella di Darwin fu formulata dal naturalista inglese A.R. Wallace.
La visione lamarckiana
La teoria evolutiva di Lamarck sostiene che l’organismo è continuamente modificato in maniera adattiva dall’ambiente in cui vive e che i cambiamenti a lungo andare diventano ereditabili: ciò provocherebbe nell’organismo, con il passare delle generazioni, cambiamenti profondi e il sorgere di nuove specie. I cambiamenti fisiologici provocati dall’uso e dal disuso degli organi vengono ereditati in tempi molto lunghi: la teoria prevede quindi l’ereditarietà dei caratteri acquisiti. Lamarck si spinge ancora oltre e sostiene che la necessità, quindi l’utilità, per un organismo di possedere un nuovo organo conferisce a esso la capacità di formarlo.
La dimostrazione dell’inaccettabilità della teoria lamarckiana risale alla seconda metà del 20° sec.: l’ereditarietà dei caratteri acquisiti necessita il passaggio dell’informazione genetica dal soma, che è modificato dall’ambiente, al gene, che è il determinante ereditario dalla cui struttura dipende quella del soma; una modificazione somatica non è trasmissibile al gene e quindi il carattere acquisito non può essere ereditabile.
La teoria darwiniana
La teoria darwiniana si basa su un principio molto semplice: prendendo in considerazione tempi relativamente brevi, la consistenza demografica delle popolazioni animali e vegetali è abbastanza costante. Come aveva notato R. Malthus, il potenziale riproduttivo di ogni specie è talmente alto che, ove non vi fosse un’altissima mortalità, il numero degli individui andrebbe continuamente e rapidamente crescendo; questo non avviene in quanto lo spazio e le risorse a disposizione sono limitate. Un’altra osservazione fondamentale è che in tutte le popolazioni esiste una notevolissima variabilità. L’idea che ha portato alla formulazione della teoria della selezione naturale è che la morte degli individui che nascono in eccesso (la stragrande maggioranza dei nati di ogni specie) non avvenga a caso, ma che in ogni momento sopravvivano quelli più adatti alle condizioni date. Poiché le condizioni in ogni nicchia ecologica non sono costanti, se si ammette che la variabilità della popolazione abbia una base ereditaria, questo processo porterà gradualmente a mutare le caratteristiche della specie e, a lungo andare, a formarne di nuove.
La teoria neodarwiniana o sintetica
Darwin stesso era conscio di una difficoltà nella sua teoria: è vero che ogni popolazione animale o vegetale è altamente variabile, ma certamente l’e. non può avvenire se non esiste negli organismi una fonte continua di variabilità, la cui origine era sconosciuta ai tempi di Darwin. Le ricerche successive al 1900, con la nascita della genetica come scienza, hanno risolto questo problema. La causa della variazione che permette l’e. degli organismi è rappresentata dalla mutazione o meglio da vari tipi di mutazione, intendendo per mutazione una variazione improvvisa del gene, o comunque del materiale ereditario, che avviene spontaneamente per cause diverse. La rielaborazione della teoria darwiniana sulla base delle nuove acquisizioni della genetica, operata nei primi decenni del 20° secolo, prende il nome di teoria neodarwiniana dell’evoluzione, o teoria sintetica dell’evoluzione.
Nuove teorie
Un secondo problema della teoria darwiniana non è stato risolto: se il processo evolutivo utilizza la selezione naturale, la quale in ogni momento favorisce la sopravvivenza del più adatto, è difficile capire come si siano formati gradualmente gli organi e le funzioni complesse. La formazione improvvisa di tali funzioni è anche impossibile in quanto necessita l’improbabile comparsa contemporanea di numerosissime mutazioni. A queste difficoltà concettuali, notevoli ai tempi di Darwin quando le basi dell’ereditarietà erano ignote e ancor più grandi adesso che sono note, si aggiunge il fatto che la teoria darwiniana dell’evoluzione prevede che i fossili forniscano una serie ragionevolmente continua e graduale dell’evolversi della vita: in realtà la serie dei fossili è tutt’altro che continua. Mentre ai tempi di Darwin era facile pensare che le serie fossero lacunose semplicemente per la difficoltà di reperire i fossili, adesso è molto più difficile sostenere questa semplice spiegazione: o l’evoluzione non è stata continua o alcune forme sono durate così poco e sono state così poco numerose che non riusciamo a reperirne i fossili.
Sulla base dell’e. darwiniana sono state sviluppate altre teorie, come quella dell’e. neutrale, secondo la quale i cambiamenti evolutivi sono dovuti a una deriva genetica casuale e non sempre sono adattivi (potrebbero quindi essere anche sfavorevoli). Secondo la teoria dell’e. a salti (o degli equilibri intermittenti; ➔ equilibrio), si sarebbero alternati lunghi periodi di stabilità a brevi periodi di cambiamenti; questa teoria spiegherebbe la mancanza, in molti casi, di forme di transizione fossili tra i diversi gruppi zoologici. Infine, la teoria dell’e. a mosaico sostiene che, nelle diverse strutture (anche a livello molecolare) di un individuo, i cambiamenti evolutivi possono avvenire in tempi diversi.
Le prove dell’evoluzione
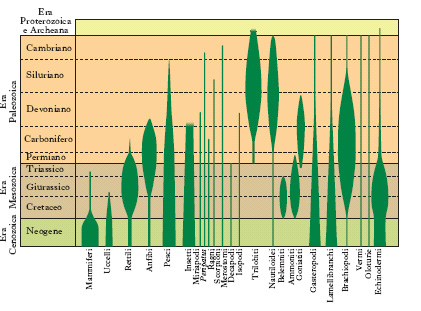
Prove basate sui reperti fossili
Anche se incomplete, le serie fossili non sono spiegabili se non accettando l’insieme di un processo evolutivo. I fossili più antichi che sono stati trovati finora sono residui di organismi microscopici, simili agli odierni batteri, reperiti in rocce che risalgono a 3,5 miliardi di anni fa. Fossili simili agli attuali cianobatteri, presumibilmente fotosintetici, compaiono in rocce di circa 2 miliardi di anni, mentre cellule con strutture simili a quelle degli eucarioti risalgono a circa 1,7 miliardi di anni fa. In un periodo molto più recente, e cioè nel Cambriano (570 milioni di anni fa), si assiste a un’incredibile diversificazione degli esseri viventi: quasi all’improvviso compaiono i rappresentanti di tutti i principali gruppi animali attualmente esistenti. Ancora oggi non esiste una spiegazione convincente di come nel giro di pochi milioni di anni (un tempo evolutivamente brevissimo) vi sia stata una simile esplosione delle forme viventi. Nelle ere successive si trovano evidenze delle prime forme di vita terrestre, sia vegetale sia animale, secondo la successione indicata nella fig. 1, fino a che in tempi evolutivamente recentissimi, circa un milione di anni fa, si assiste alla comparsa dei primi uomini. È importante notare che moltissimi degli organismi che troviamo allo stato fossile sono estinti perché gradualmente sostituiti da altri, più adatti all’ambiente in continuo cambiamento.
Prove fornite dalla biochimica e dalla biologia molecolare
L’analisi dei costituenti biochimici e delle vie metaboliche degli organismi, dai batteri all’uomo, mettono in evidenza alcune caratteristiche fondamentali della vita. Tutti gli organismi hanno come mattoni fondamentali due macromolecole, le proteine e gli acidi nucleici, la cui struttura fondamentale è identica in tutti gli esseri viventi. Le proteine sono sempre composte dagli stessi 20 amminoacidi (il numero possibile di questi è molto superiore) in forma L (la forma L e la forma D sono funzionalmente identiche). Anche gli acidi nucleici, nei quali risiedono i determinanti ereditari, hanno la stessa identica struttura in tutti gli esseri viventi, così come è identico (salvo trascurabili eccezioni) il codice genetico, cioè l’insieme delle regole con cui l’informazione contenuta negli acidi nucleici viene utilizzata per costruire le proteine. Ciò si spiega solo ammettendo che tutti gli organismi derivino da un organismo primordiale che già possedeva le caratteristiche fondamentali di quelli attuali. Inoltre, prendendo in esame le vie metaboliche, vale a dire quelle serie di reazioni biochimiche con cui gli organismi costruiscono i propri componenti e utilizzano i substrati, è possibile osservare che esse sono quasi identiche in tutti gli esseri viventi, osservazione che avvalora ulteriormente le conclusioni precedentemente esposte.
Prove derivate dall’anatomia comparata, dalla fisiologia e dall’embriologia
L’esame comparato degli organismi indica chiaramente che esistono fra di essi notevoli somiglianze e che possono essere ordinati sulla base della loro struttura, secondo una serie in cui la complessità aumenta progressivamente. Gli animali pluricellulari più semplici sono le Spugne e i Celenterati, il cui corpo è formato da due soli foglietti cellulari (l’ectoderma e l’entoderma) e presentano una sola apertura che è insieme anale e buccale. Seguono i Platelminti, o vermi piatti, nei quali compare il terzo foglietto cellulare (il mesoderma) e poi i Nematelminti, o vermi tondi, nei quali compaiono una seconda apertura, quella buccale, organi complessi, che più o meno modificati troveremo in tutti gli animali, e un accenno di formazione del celoma, ovvero la cavità che separa l’intestino dal tegumento. Il celoma si perfeziona negli Anellidi dove la cavità risulta circondata completamente di una sua propria membrana. Esaminando un gruppo di animali più ristretto, per es. i Mammiferi placentati, si può osservare che tutti, pur così differenti tra di loro come una giraffa da un topo o un pipistrello da una foca, sono costituiti esattamente dai medesimi organi, i quali hanno la stessa struttura fondamentale: gli animali sono diversi fra loro solo perché lo sviluppo di ogni singolo organo è differente nei diversi animali. Prendendo in considerazione un gruppo vasto quale quello dei Vertebrati, che comprende i Pesci (cartilaginei e ossei), gli Anfibi, i Rettili, gli Uccelli e i Mammiferi, è possibile in alcuni casi vedere come molti organi che compaiono nel gruppo più primitivo (quello dei Pesci cartilaginei, che sono comparsi per primi nella storia evolutiva, come si desume dallo studio dei fossili) si conservano, seppure adattati alle nuove situazioni, nei gruppi più recenti. Un caso tipico è rappresentato dal sistema circolatorio, e in particolare dal cuore, che presenta nei Pesci un singolo atrio e un singolo ventricolo, due atri e un solo ventricolo negli Anfibi, due atri e due ventricoli non completamente separati nei Rettili, due atri e due ventricoli completamente divisi nei Mammiferi e negli Uccelli. Questi cambiamenti progressivi sono correlati a esigenze funzionali del sistema circolatorio; in ogni caso, la condizione più semplice è quella presente nell’organismo che è comparso per primo sulla scena dell’evoluzione. Queste osservazioni sono interpretabili nella maniera più logica supponendo la formazione di piani strutturali che sono andati lentamente evolvendo, a seconda delle specifiche necessità dei diversi organismi: per es., il sistema circolatorio è drasticamente cambiato nei Vertebrati con il passaggio dalla vita acquatica a quella terrestre.

Queste conclusioni sono ulteriormente avvalorate dallo studio dell’embriologia. Gli organismi sono fra loro molto più simili nella vita embrionale di quanto non lo siano nella vita adulta. In base a questa considerazione E. Haeckel fondò la cosiddetta legge biogenetica fondamentale, la quale afferma che l’ontogenesi (sviluppo embriologico) è la ricapitolazione della filogenesi (storia evolutiva). Sebbene la validità di tale legge appaia oggi più limitata di quanto non si pensasse ai tempi di Haeckel, è certo che alcune affinità e omologie si possono stabilire solo in base a ricerche embriologiche (fig. 2). Inoltre, nel corso della vita embrionale si formano, spesso senza mai divenire funzionali, organi che sono presenti e funzionali in organismi più primitivi.
La distribuzione geografica degli animali e delle piante presenta singolarità (per es., l’assenza di Mammiferi nella Nuova Zelanda, dei Placentati nell’Australia, di orsi bianchi nell’Antartide e di pinguini nell’Artide; la presenza dei Marsupiali soltanto in Australia e, con pochissime specie, in America) che si possono interpretare abbastanza bene in termini evoluzionistici. Infine la sistematica zoologica e botanica, che appunto si basa sulle affinità di struttura fra gli esseri viventi, offre argomenti di prima importanza a favore dell’evoluzione.
Anche l’uomo è il prodotto di un’e. a partire da antenati del gruppo delle scimmie, durante il Pliocene e il Pleistocene. Gli antenati più prossimi erano simili alle attuali scimmie antropomorfe. Questa affermazione, già contenuta in Darwin, ha poi trovato conferma nelle numerose scoperte di resti fossili di Preominidi e Ominidi (➔) che dimostrano alcune tappe di questa evoluzione.
Meccanismi dell’evoluzione
La scoperta dei meccanismi che sono alla base dell’e. è dovuta per la massima parte ai grandi progressi compiuti dalla genetica nel Novecento. I processi evolutivi fino a ora ben dimostrati sono: la mutazione, la selezione, la deriva genetica e l’isolamento geografico. Essi determinano sostanzialmente una variazione delle frequenze alleliche nelle popolazioni. Lo studio della variabilità ereditaria e della sua modulazione nel tempo e nello spazio è stato effettuato da G. Hardy e W. Weinberg (➔ equilibrio).
Le mutazioni
Le mutazioni operano a tre livelli: genico, producendo nuovi alleli, cromosomico, producendo nuove strutture cromosomiche, e genomico, dando luogo a cambiamenti del numero dei cromosomi. Esse rappresentano la base dell’e.: le mutazioni geniche possono produrre effetti sull’insieme dei caratteri visibili di un organismo (fenotipo); avvengono spontaneamente (e cioè per cause ignote) con frequenza molto bassa (circa 1/100.000 o 1/1.000.000 di geni, per gene e per generazione); la loro frequenza può essere aumentata da agenti mutageni quali i raggi X o alcune sostanze chimiche. Comunque le mutazioni che si verificano in un determinato ambiente sono casuali, cioè non si verificano con maggiore frequenza le mutazioni che consentono un migliore adattamento degli individui all’ambiente stesso. Le mutazioni cromosomiche non producono modificazioni delle caratteristiche morfologiche ma controllano caratteristiche funzionali; inoltre rivestono un ruolo importante nel determinare la sterilità degli ibridi negli incroci interspecifici. Le mutazioni genomiche, molto diffuse nelle piante e rare negli animali, causano variazioni dell’assetto cromosomico che, invece di essere diploide (2 n), come di norma, può essere triploide (3 n), tetraploide (4 n) ecc.; possono produrre anche variazioni che interessano una sola o poche coppie di cromosomi (aneuploidia).
La selezione naturale
L’orientamento dell’e. è il risultato dei fenomeni di selezione naturale i quali agiscono sulla variabilità genetica, fornita dalle mutazioni casuali, attraverso la riproduzione differenziale; infatti i portatori di combinazioni geniche favorite dalla selezione contribuiscono con un maggior numero di discendenti alla generazione successiva. L’esistenza e l’azione della selezione naturale è suffragata da numerose prove sperimentali, date da esperimenti di laboratorio e da osservazioni in natura. Per es., ceppi di batteri incapaci di sopravvivere in presenza di un certo antibiotico divengono improvvisamente resistenti a esso e si moltiplicano nonostante la presenza dell’antibiotico grazie alla comparsa per mutazione di individui resistenti. Questa mutazione, preesistente nel genotipo dei batteri alla comparsa dell’antibiotico, risulta neutra o addirittura sfavorevole in ambienti privi di antibiotici, ma è invece favorita dalla selezione in presenza di antibiotico; infatti solo gli individui portatori di tale mutazione sopravvivono e si riproducono, mentre tutti gli altri muoiono: ciò spiega perché alcuni antibiotici rivelatisi molto efficaci al momento della loro comparsa perdono in breve tempo la capacità di agire su molti germi patogeni. Analoghi ai fenomeni di resistenza dei batteri agli antibiotici sono quelli di resistenza agli insetticidi da parte di molti artropodi; anche gli insetticidi hanno quindi agito come forze selettive modificando in breve tempo il patrimonio ereditario di numerose popolazioni.
Deriva genetica
Si hanno fenomeni di deriva quando la frequenza di uno o più alleli (o addirittura la presenza o l’assenza di essi) in una popolazione è determinata unicamente dal caso, e ciò avviene quando più alleli hanno valore selettivo simile e quando una popolazione viene drasticamente ridotta di numero (per es., in una popolazione di zanzare che abita una zona trattata con insetticidi, la popolazione che si formerà dopo il trattamento porterà solo gli assetti cromosomici delle zanzare sopravvissute); gli effetti della deriva sono tanto più accentuati quanto minore è il numero degli individui della popolazione.
Isolamento geografico
L’isolamento geografico riveste un ruolo fondamentale nell’e. e in particolare nel processo di formazione delle specie o speciazione, che ne è il punto cruciale. Esso, impedendo il flusso genico e quindi il rimescolamento genetico tra popolazioni adiacenti, permette alle popolazioni isolate di adattarsi indipendentemente ai diversi ambienti, ed è quindi la causa primaria del loro differenziamento in razze, semispecie e specie distinte. Così, se una singola specie è distribuita uniformemente su un certo territorio e in seguito, sia per effetto delle diverse pressioni selettive che si realizzano nelle varie parti dell’areale, sia per la presenza di una barriera geografica (catena montuosa, tratto di mare o più semplicemente un territorio inabitabile per la specie), si stabiliscono differenze genetiche tra gruppi di popolazioni a mano a mano isolati, questi si evolveranno indipendentemente gli uni dagli altri dando origine a razze geografiche o a specie distinte. Se la barriera viene a cadere e i gruppi di popolazioni tornano in contatto, possono o ibridarsi o risultare riproduttivamente isolati, e in tal caso il processo di speciazione è compiuto. È da notare che l’isolamento riproduttivo non viene necessariamente raggiunto nel corso dell’isolamento geografico, perché, finché le popolazioni rimangono isolate, nessuna forza selettiva agisce in favore dell’insorgenza di barriere riproduttive; è quando le popolazioni tornano in contatto che l’isolamento riproduttivo può essere favorito dalla selezione naturale, instaurarsi e rafforzarsi con meccanismi di varia natura (genetica, ecologica, etologica ecc.). Se infatti la prole derivata da due popolazioni precedentemente isolate ha un grado di adattamento (fitness) inferiore a quello della prole derivata da incroci intrapopolazionali, la selezione naturale favorirà le mutazioni e le combinazioni geniche che rendono più probabile l’accoppiamento tra individui della stessa popolazione (➔ speciazione).
Formazione delle specie per poliploidia
Nelle piante, ove è largamente diffusa la riproduzione vegetativa, la formazione delle specie attraverso l’ibridazione di due entità diverse e successiva poliploidizzazione assume un ruolo importante. Infatti, anche se gli ibridi tra due specie sono sterili, a causa di disturbi nella formazione dei gameti, essi possono ugualmente propagarsi per riproduzione vegetativa. Uno dei più noti e importanti organismi poliploidi è il frumento da pane (Triticum aestivum) con 42 cromosomi, che si è originato in seguito alla produzione di un ibrido tra un frumento coltivato, con 28 cromosomi, e una graminacea selvatica dello stesso genere, con 14 cromosomi. Fenomeni simili non sono conosciuti negli animali e ciò è probabilmente in rapporto alla quasi generale assenza in essi della riproduzione vegetativa; sono invece frequenti casi di animali in cui l’ibridazione e la poliploidia hanno sostenuto un ruolo importante nell’e. (platelminti, anellidi, crostacei, insetti, alcuni vertebrati); in questi casi però vi è presenza anche di ermafroditismo o di partenogenesi.
Botanica
Nel mondo vegetale, il processo della e. organica ha prodotto effetti più intensi nelle strutture riproduttive che non su quelle somatiche, a differenza degli animali che, oltre a non essere fissi al suolo, sono sempre eterotrofi e mobili. La spiegazione di tale comportamento è da ricondurre alla peculiare capacità delle piante di accedere direttamente a fonti esterne di energia (chimica o luminosa), alla loro più stretta e continua relazione con il mondo minerale, di cui utilizzano direttamente i materiali organicandoli, alla loro funzione di tramite tra il mondo minerale e quello vivente. La facilità e versatilità dell’alimentazione e, nelle piante acquatiche, i vantaggi dovuti alla vita nell’acqua, hanno reso meno impellente l’esigenza di una forte diversificazione anatomica, tanto che nelle piante la distribuzione del lavoro a livello dei tessuti e degli organi risulta alquanto semplificata: a parte la diversificazione dell’aspetto morfologico complessivo, tutte le piante vascolari, per es., rispondono, per ciò che riguarda la fase vegetativa, a modelli organizzativi poco numerosi e sensibilmente uniformi. Molto più ricca appare invece la filogenesi degli organi riproduttivi. Nei fiori si assiste al passaggio dalle grandi strutture acicliche (per es., nelle Conifere) a quelle verticillate a cinque o quattro elementi per verticillo (per es., Angiosperme Dicotiledoni) o a tre elementi (per es., Angiosperme Monocotiledoni); da fiori a pezzi separati (dialipetale) a fiori a pezzi saldati (gamopetale), da strutture a 5,4,3 piani di simmetria (fiori actinomorfi) a strutture con unico piano di simmetria (fiori zigomorfi) o affatto irregolari; da fiori con ovario libero (supero), da cui derivano veri frutti, a fiori con ovario infossato (infero) e avvolto dal ricettacolo, da cui derivano falsi frutti, e così via. Lo studio comparativo morfologico, citologico e fisiologico degli organi riproduttivi (archegoni, anteridi, oangi ecc.) deve ancora essere approfondito con possibilità di notevoli cambiamenti nella sistematica del Regno vegetale; tale studio, finora effettuato per alcuni gruppi vegetali, si presenta particolarmente complesso nell’ambito delle alghe e dei funghi, ove le relazioni filogenetiche sono tuttora di incerta interpretazione.
Linguistica
La serie delle modificazioni fonetiche, morfologiche, strutturali, semantiche, attraverso le quali passa una lingua nel suo sviluppo storico: l’ e. semantica è il risultato dei successivi cambiamenti di significato che subisce un vocabolo; l’ e. fonetica comprende qualsiasi alterazione fonetica che in una parola, o in una serie analoga di parole, appare come progressiva, in opposizione ai mutamenti improvvisi e immediati.
Medicina
E. del feto
L’insieme dei movimenti compiuti dal corpo fetale durante il parto, culminanti con la sua espulsione ed estrazione.
Scienza militare
Nel linguaggio militare, insieme di movimenti di truppe, di navi, di aerei, che cambiano ordinatamente la loro posizione. In particolare, con riferimento a truppe terrestri, ogni movimento compiuto da reparti per disporsi al combattimento.
In marina, il percorso che, seguendo direzioni varie, viene effettuato da una nave per un determinato fine. Le e. sono i vari movimenti d’insieme che un complesso di navi compie per cambiamento di rotta o di formazione a scopo tattico o di parata.