
vita
Biologia
La condizione degli organismi dotati di una forma specifica, di una costituzione chimica determinata, capaci di mantenersi in una situazione di equilibrio dinamico, cioè di avere un ambiente interno costante nonostante gli scambi con l’ambiente esterno (omeostasi) e di riprodurre queste proprietà in altri organismi simili.
Elementi chimici della vita

La composizione chimica della materia vivente è molto differente da quella della crosta terrestre e dell’atmosfera. I quattro elementi più abbondanti nella biosfera sono idrogeno (H), ossigeno (O), carbonio (C), azoto (N), mentre i quattro elementi più abbondanti nella crosta terrestre sono ossigeno, silicio (Si), alluminio (Al) e ferro (Fe). H e O rappresentano l’88,5% degli atomi del corpo umano; C rappresenta il 9,5% e N l’1,4% (tab. 1).
H, O, N e C sono gli elementi a numero atomico più basso che possano formare legami covalenti, mettendo in comune gli elettroni dello strato più esterno. Hanno infatti bisogno, rispettivamente, di 1, 2, 3 e 4 elettroni per completare il loro strato esterno e formare legami covalenti stabili. Inoltre, C, O e N possono mettere in comune sia 1 sia 2 elettroni, permettendo così la formazione di legami singoli o doppi. Questa proprietà permette una considerevole varietà di legami chimici. Il carbonio può anche formare legami tripli, legandosi sia con un altro atomo di C sia con un atomo di N. C, N, O e H non sono gli unici elementi ad avere tali proprietà, ma sono i più leggeri fra quelli capaci di formare legami forti (covalenti). Degli altri elementi che sono stati dimostrati essenziali per la v. di un animale, soltanto 3 hanno un numero atomico superiore a 34: molibdeno (numero atomico 42), stagno (50) e iodio (53), che sono necessari soltanto in tracce. Particolarmente importante è la capacità degli atomi di C di legarsi tra loro e con O, H, N e lo zolfo (S) per formare un’immensa varietà di composti con diversi gruppi funzionali. I composti chimici del C hanno, inoltre, un’altra importante caratteristica. A causa della struttura a tetraedro degli orbitali degli elettroni che sono messi in comune, composti con la stessa formula chimica possono assumere strutture tridimensionali differenti (isomeri ottici). Oltre al C solo il Si possiede la capacità di combinarsi con sé stesso, mettendo in comune quattro elettroni. Tuttavia, sebbene sia molto più abbondante nella litosfera, il Si non è adatto a realizzare le caratteristiche funzionali e strutturali tipiche degli organismi viventi. Infatti, mentre la maggior parte dei composti ossidati del Si sono insolubili, il C, sotto forma di anidride carbonica, non solo ha un’alta solubilità in acqua ma è anche liposolubile. Proprietà quest’ultima che rende possibile il passaggio di questa molecola attraverso le membrane biologiche. Inoltre il Si, combinandosi con sé stesso, forma catene molto corte e i legami Si-Si sono estremamente instabili in presenza di acqua, ammoniaca od ossigeno.
C, O, H, N, S e fosforo (P), legandosi fra loro, formano le molecole essenziali della v.: amminoacidi, carboidrati, acidi grassi, purine, pirimidine e nucleotidi. Queste ultime molecole, oltre ad avere un proprio ruolo biochimico indipendente, si polimerizzano e danno luogo a macromolecole complesse fondamentali, come le proteine, il glicogeno, i polisaccaridi e gli acidi nucleici. P svolge inoltre un ruolo primario nel bilancio energetico di una cellula, attraverso i due legami pirofosfato presenti nella molecola di ATP.
Origine della vita
Il problema dell’origine della v. è affrontato da scienziati di varie discipline. Consiste nel capire come, e in che condizioni, un sistema fisico complesso possa assumere l’ordine spazio-temporale caratteristico di una cellula che si accresce e si riproduce. La cellula è la sede di due specie di processi interni, che si svolgono contemporaneamente, condizionandosi a vicenda. Da un lato c’è una complessa rete di vie biosintetiche e degradative, costituite da una successione di trasformazioni ciascuna delle quali coinvolge contemporaneamente un elevato numero di molecole: si tratta di reazioni chimiche, per lo studio delle quali l’approccio termodinamico rappresenta una tecnica di elezione. Dall’altro lato c’è però un gran numero di processi che si svolgono per singoli atti reattivi, coinvolgendo una sola o pochissime molecole per volta, in una successione rigorosamente ordinata nello spazio e nel tempo. Tali processi, quali, per es., la duplicazione del DNA o la catena di trasporto degli elettroni dall’acqua al NADP+ nella fotosintesi, non possono che essere studiati a livello di meccanica molecolare.
L’ipotesi della sintesi prebiotica
Basandosi sull’osservazione che tutte le forme di vita sono costituite dalle stesse macromolecole A.I. Oparin (1923), J.B.S. Haldane (1929), J.D. Bernal (1951) e M. Calvin (1959) suggerirono che le condizioni geologiche della Terra primitiva avrebbero potuto rendere possibile una sintesi prebiotica, vale a dire la sintesi di una sufficiente quantità e varietà di molecole biologicamente importanti partendo da composti inorganici. Appena la Terra si raffreddò formando una crosta solida, i gas, formatisi al di sotto di essa, uscendo dalle fessure della superficie terrestre, crearono un’atmosfera primitiva. Molto probabilmente, l’acqua arrivò alla superficie della Terra sotto forma di gas e l’oceano (più piccolo dell’attuale di circa un fattore 10) si formò come risultato di milioni di anni di piogge torrenziali. Era attivo il ciclo dell’acqua e il pH dell’oceano non doveva essere molto diverso dall’attuale in quanto, anche allora, tamponato dai silicati derivati dall’erosione delle rocce superficiali. L’atmosfera primitiva era sostanzialmente priva di ossigeno; questo gas, da cui dipende la maggior parte della v. attuale, sembra si sia formato dall’acqua attraverso gli organismi fotosintetici che si svilupparono più tardi. Seguendo le ipotesi di Oparin e S.L. Miller, l’opinione prevalente fino agli anni 1980 era che l’atmosfera primitiva fosse fortemente riducente, in quanto avrebbe contenuto soprattutto idrogeno, metano, ammoniaca e monossido di carbonio, qualora l’uscita dei gas dalle fessure della crosta terrestre fosse avvenuta prima che la maggior parte del ferro metallico affondasse nel cuore della Terra. Presumibilmente, invece, una certa quantità di ferro metallico rimase in superficie, reagendo con i gas uscenti e formando soprattutto anidride carbonica, poco monossido di carbonio e scarsissime quantità di ammoniaca e metano.
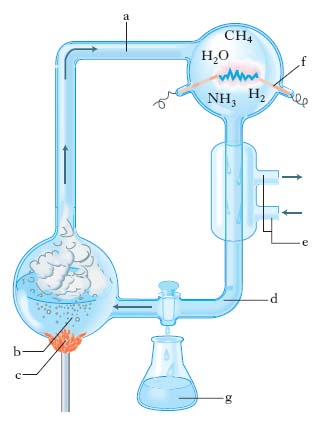
L’opinione prevalente negli anni più recenti è a favore di un’atmosfera primitiva con caratteristiche pressoché neutre. I quattro elementi fondamentali della v. (H, O, N, C), comunque presenti nell’atmosfera primitiva insieme all’energia, derivata soprattutto dai raggi ultravioletti solari e dalle scariche elettriche dei temporali, avrebbero portato alla sintesi di una quantità di molecole organiche che, dilavate dalle piogge, dovevano poi accumularsi nell’oceano. C’è la possibilità che le fratture profonde sotto gli oceani siano state la sede di sintesi di molecole organiche. In queste regioni, ricche di zolfo, fosforo e catalizzatori minerali, sarebbe avvenuta la formazione di NH3 e CH4 in quanto vi era disponibilità sia degli atomi necessari alla sintesi di composti organici sia di sorgenti di energia capaci di promuovere reazioni chimiche. Gli esperimenti di Miller (1953-57), basati su alcune premesse teoriche di H.C. Urey (1952), hanno dimostrato la possibilità di ottenere vari composti organici in una miscela di acqua, ammoniaca, metano e idrogeno sottoposta per lungo tempo a elevata temperatura e a scariche elettriche ad alto potenziale e posta in un ambiente chiuso (fig. 1). Molte differenti condizioni di reazione che simulano plausibili condizioni prebiotiche possono condurre a sintesi di molecole organiche. Di particolare interesse sono gli esperimenti effettuati da C. Ponnaperuma (1963) che, utilizzando luce ultravioletta come sorgente di energia e sostanze inorganiche e organiche semplici (carbonio, composti dell’azoto, acido cianidrico), ottenne la sintesi di amminoacidi e nucleotidi.
L’ipotesi dell’origine dallo spazio
La Terra non sarebbe tuttavia l’unico luogo del Sistema solare dove si possono originare molecole organiche. Già S.A. Arrhenius nel 1900 aveva postulato che la v. fosse arrivata nel nostro pianeta dagli spazi extraterrestri. Analisi di meteoriti, specialmente di quella che precipitò a Murchison, in Australia, nel 1969, hanno mostrato che esse contengono anche gli amminoacidi presenti nei sistemi biologici. Poiché questi amminoacidi sono stati trovati all’interno della meteorite non possono essere considerati come contaminazioni di superficie. Analisi, condotte mediante sonde spaziali e spettrofotometri, sul nucleo della cometa Halley hanno dimostrato la presenza di polimeri di formaldeide e di poliossimetano (HCO). Gli studi effettuati utilizzando la sonda Giotto hanno rivelato che nel nucleo della cometa sono presenti particelle silicato-simili, carbonio, ossigeno, azoto.
La presenza di sostanze organiche in oggetti extraterrestri suggerisce che i componenti essenziali delle macromolecole biologicamente importanti si formano facilmente in natura e potrebbero essersi formate circa 4 miliardi di anni fa attraverso reazioni non biologiche. Non essendovi allora né ossigeno libero né metabolismo cellulare, queste molecole organiche potrebbero essere rimaste molto stabili. In conseguenza di ciò negli oceani si sarebbero accumulate concentrazioni abbastanza alte di reagenti inorganici che hanno poi subito ulteriori trasformazioni. Il ‘brodo primordiale’ nel quale si è formata la v., ipotizzato da Haldane e Oparin, è quasi certamente esistito. Rimane comunque aperto il problema di come si sia passati dai primi composti organici molto semplici alle proteine funzionali e al sistema informativo a base di acidi nucleici che garantisce riproduzione e conservazione di funzioni.
Il ruolo dell’RNA
Anche se non vi sono prove sufficienti, si ritiene che i catalizzatori primitivi dei primi composti organici siano stati le argille e gli ammassi di pirite. Alla fine degli anni 1960 L. Vogel, F. Crick e C. Woese, indipendentemente, considerato il triplice ruolo dell’RNA nella sintesi proteica (➔ proteine), ipotizzarono che questa molecola doveva essere il primo polimero formatosi all’origine della vita. Nel 1970 H.M. Temin ha dimostrato che da molecole di RNA è possibile ottenere molecole di DNA, utilizzando la trascrittasi inversa. Questo enzima è molto diffuso in natura ed è presente anche in un batterio molto antico, Myxococcus xanthus. È pertanto ragionevole pensare che la conversione del mondo dell’RNA a quello del DNA abbia potuto utilizzare un’attività enzimatica del tipo di quella della trascrittasi inversa. Il ruolo primario dell’RNA nell’evoluzione precellulare è ulteriormente confermato dalle numerose scoperte derivate dall’uso di nuove tecnologie molecolari. È noto infatti che l’RNA serve da innesco nella sintesi di DNA, è parte necessaria dei telomeri permettendo l’aggiunta del DNA terminale ai cromosomi eucariotici ed è presente nell’apparato secretorio delle proteine. Ma la scoperta veramente significativa è stata quella della sua attività catalitica che include un’attività autonoma di taglio, polimerizzazione e saldatura (➔ ribozima).
Un altro punto cruciale e non ancora compreso dell’evoluzione prebiotica è l’insorgenza di una relazione ordinata tra l’RNA (o tra i suoi precursori) e le proteine; furono infatti queste interazioni che portarono alla nascita del codice genetico e ai principi della sua traduzione. L’universalità del codice genetico, che è uniforme in tutti gli organismi, dai virus ai batteri all’uomo, è uno dei più forti argomenti a favore dell’unicità dell’origine di tutti gli esseri viventi. Si concorda infatti sul fatto che, una volta scelto un determinato codice, questo si sia mantenuto in quanto qualsiasi cambiamento sarebbe stato fonte di gravi perturbazioni ed è molto probabile che un codice a tre lettere fosse presente al momento in cui si formarono le prime cellule.
Lo studio dei fossili
La discussione sulla chimica precellulare potrebbe tuttavia rimanere un fatto squisitamente speculativo malgrado le molte informazioni accumulate e i notevoli progressi della biologia molecolare. Per quello che riguarda invece le informazioni sulle prime cellule, lo studio dei fossili ha dato qualche indicazione. Reperti fossili di forma sferica, ritrovati a Greenland (Australia) e nella Repubblica Sudafricana in formazioni rocciose sedimentarie vecchie di 3,5 miliardi di anni, sono stati interpretati come residui di cellule in quanto le dimensioni e le forme di questi microrganismi sono simili a quelle degli attuali cianobatteri. Strutture molto simili, chiamate stromatoliti, si formano anche oggi nelle rocce sedimentarie, dove i sedimenti oceanici precipitano intorno a colonie di cianobatteri. Mentre da questi microfossili non si possono ricavare molte informazioni in quanto l’approccio è esclusivamente morfologico, più promettente sembra l’approccio molecolare. Per es., la conoscenza delle sequenze complete di RNA ribosomiale di diverse centinaia di organismi ha condotto all’importante e sorprendente conclusione che esistono tre linee cellulari altrettanto antiche, ciascuna delle quali è ben distinta e ugualmente diversa dalle altre due: gli Archeobatteri, gli Eubatteri (comunemente detti batteri) e gli Eucarioti. Non è pertanto più accettata l’ipotesi, formulata precedentemente, che una di queste linee cellulari abbia preceduto le altre (➔ progenote). Gli studi sulla sequenza del DNA hanno invece confermato l’ipotesi endosimbiotica secondo la quale mitocondri e cloroplasti, originariamente cellule simili ai procarioti, siano stati fagocitati da altre cellule per formare gli Eucarioti attuali.
Sulla base del complesso delle informazioni raccolte, i tempi entro cui le prime forme di v. sono apparse e si sono affermate sulla Terra possono essere così riassunti: a 4,5 miliardi di anni fa è datata la formazione del pianeta e della Luna; successivamente vi sono stati intensi bombardamenti di meteoriti e tra 4,4 e 3,8 miliardi di anni fa sono comparse le prime forme di vita; intorno a 3 miliardi di anni fa, si sarebbero formati i primi organismi in grado di condurre processi fotosintetici e di sviluppare ossigeno nell’atmosfera (organismi autotrofi); verso i 2 miliardi di anni fa, si è sviluppata un’atmosfera ricca di ossigeno, capace di sostenere i primi organismi eterotrofi.
Durata della vita negli animali
Sulla durata media della v. degli animali selvatici non si hanno, ovviamente, dati precisi. Pare accertato che le tartarughe giganti delle Galápagos e di altre isole possano vivere fino a 200 anni. Anche gli elefanti, le balene e altri grossi Mammiferi hanno v. lunga (oltre un secolo). Il cammello può vivere 40-50 anni. Molto più breve è la v. dei Mammiferi di dimensioni più piccole: i ratti vivono 5-7 anni, altri topi 3-4. In tab. 2 sono indicati i dati relativi ad alcuni animali domestici. Fra gli Uccelli, pappagalli, gufi, falchi, picchi, sono piuttosto longevi (fino a 60-100 anni), mentre i Passeriformi hanno in genere una durata di v. molto più breve. I pesci vivono a lungo: anche specie di modeste dimensioni, come l’aringa, vivono fino a 20 anni, la platessa (Pleuronettidi) fino a 70. Di altri pesci, come i lucci e le carpe, si ritiene possano raggiungere il secolo. Anche alcuni Molluschi, come Tridacna e Unio, sono assai longevi (70-100 anni). La v. negli Insetti è breve e si conclude nel giro di un anno, o, in alcune specie, di pochi mesi o giorni. Tuttavia non mancano specie con v. larvale lunghissima, come una cicala (Tibicina septemdecim), la cui larva si metamorfosa dopo 17 anni di v. larvale.
Durata della vita nelle piante
Nelle piante la durata della v. varia entro limiti molto più estesi che negli animali, da 20 minuti (batteri) ad alcune migliaia di anni (sequoie). Molte piante unicellulari vivono poche ore o pochi giorni, alcune alghe (Macrocystis) molti anni (forse 100), la maggior parte dei funghi poche settimane o qualche mese, ma alcuni sono perenni (Poliporacee) o hanno micelio perennante (oltre a uno o più tipi di spore); nelle Fanerogame si distinguono le annue, le bienni e le perenni e fra queste ultime le erbacee e le legnose: nelle erbacee una parte del corpo, per lo più sotterranea (rizomi ecc.), vive per parecchi anni, mentre i getti sono annuali; fra le legnose si contano i vegetali più longevi, infatti vivono da 1000 a 2000 anni diversi alberi, come Taxodium mexicanum, l’olivo, il cipresso, il castagno ecc., tutti superati dalle sequoie, giacché in Sequoia gigantea, su tronchi morti, sono state contate 2300 cerchie annuali, e per un esemplare è stata calcolata, ma non con sicurezza, l’età di 4000 anni; tuttavia gli esemplari viventi di ambedue le specie di Sequoia non superano i 1500 anni. Meno longevi sono il baobab (400 anni) e Dracaena draco (200 anni). Non esiste proporzione tra età e dimensioni: esistono piante annuali, alte parecchi decimetri quali il mais o il girasole e arbusti nani della flora alpina, che pur essendo alti pochi centimetri, contano molti anni di età. Inoltre in un albero una parte minima della massa è data da cellule vive.
Data la costituzione delle piante superiori così diversa da quella degli animali è opportuno ricordare che la durata dei diversi organi (foglie, fiori, semi) e delle altre parti è diversa anche in uno stesso individuo. Per la durata di singole cellule si può dire che il massimo è raggiunto dagli elementi dei raggi midollari secondari in diversi alberi (decine di anni) e dalle cellule della corteccia e del midollo di alcuni cacti (100 anni).
Vita artificiale
Il settore di ricerca interdisciplinare della v. artificiale esplora la possibilità di nuove forme di v.; esso ha per traguardo la comprensione della v. attraverso la ‘costruzione’ di sistemi viventi invece che attraverso la loro scomposizione sperimentale ed è quindi caratterizzato da un approccio sintetico piuttosto che riduzionista o analitico. Gli studi condotti in quest’ambito riguardano alcuni particolari aspetti della v., come l’intelligenza, l’evoluzione, il linguaggio, lo sviluppo ecc., perciò vertono su sistemi che presentano solo alcune proprietà caratteristiche dei sistemi viventi riguardanti l’area specifica di interesse e non comprendono moltissime altre proprietà vitali. Le ricerche di v. artificiale si articolano in tre aree principali: la sintesi wetware (da wet, umido [come il cervello umano], creazione di modelli teorici puramente speculativi), quella software (realizzazione e studio all’elaboratore di sistemi viventi spesso indicata con la locuzione v. digitale) e quella hardware (realizzazione di sistemi concreti funzionanti). Poiché i sistemi studiati dalla v. artificiale sono creati dal ricercatore, si possono in tal modo esaminare fenomeni biologicamente possibili ma non reali, cioè fenomeni che pur avendo plausibilità biologica non sono presenti in natura; lo studio di tali fenomeni, nella misura in cui sono basati sugli stessi meccanismi e processi di quelli reali, può servire ad allargare il campo dei fenomeni empirici con i quali verificare ipotesi e teorie sul mondo vivente reale. La v. artificiale è interessata a ogni fenomeno e aspetto del mondo vivente; essa studia le diverse entità che esistono ai vari livelli della gerarchia biologica, dalle biomolecole alle cellule, dagli organi agli organismi, dalle popolazioni di organismi alle comunità e agli ecosistemi; inoltre, a ciascun livello di entità biologiche la v. artificiale studia una varietà di fenomeni quali la riproduzione, la crescita e lo sviluppo, l’evoluzione e l’apprendimento, le interazioni delle entità biologiche tra loro e con l’ambiente esterno, e così via. La v. artificiale estende la concezione di v. organica e può essere un utile strumento per alcuni tipi di studi biologici, in particolare perché possiede proprietà che la rendono più semplice da studiare; così, per es., nel caso della v. digitale, è possibile predefinire completamente la sequenza dell’intero genoma di ogni essere nel sistema artificiale realizzato all’elaboratore; tale sistema può essere manipolato a piacimento e la sua evoluzione può essere fatta procedere con una velocità estremamente maggiore rispetto a un sistema vivente naturale.
Gli studi sulle reti neurali (➔ neurale, rete), le quali nell’ambito della v. artificiale possono fornire modelli del sistema nervoso e del suo comportamento, sono pervenuti a risultati estremamente interessanti, particolarmente per quanto riguarda il loro impiego come sistemi di controllo di organi sensoriali e motori di un organismo: si è, per es., potuto stabilire che vi è una stretta correlazione (coadattamento) tra il grado di sofisticazione del sistema sensoriale e il grado di sofisticazione del sistema motorio e che, inoltre, se lasciati liberi di evolvere, sia il sistema sensoriale sia quello motorio si modificano, ma si mantengono al tempo stesso coadattati a ogni stadio del processo evolutivo (vi è quindi coadattamento sensomotorio in quanto vi è coevoluzione tra i due sistemi).
Diritto
La v. umana è un bene indisponibile che riceve una particolare tutela dall’ordinamento. Ad es., possono ricordarsi l’art. 1 l. 22 maggio 1978, n. 194 (in base al quale lo Stato tutela la v. umana dal suo inizio), e, più di recente, l’art. 1 l. 19 febbraio 2004, n. 40 (che annovera il concepito tra i soggetti di diritto) e l’art. 10 della Convenzione delle Nazioni Unite sui diritti delle persone con disabilità del 13 dicembre 2006 (ratificata dall’Italia con l. 3 marzo 2009, n. 18): il diritto alla v. è connaturato alla persona umana. Il diritto alla v. è protetto dalla possibile violazione ad opera non solo di terzi (in capo ai quali potrà quindi sorgere l’obbligo di risarcire i danni provocati cagionando la morte di un soggetto), ma anche di sé stessi (i trattamenti terapeutici sono infatti ritenuti legittimi, pure nell’eventualità di rifiuto del paziente, nel caso di un pregiudizio imminente ed irreparabile che integri gli estremi dello stato di necessità ex art. 2045 c.c.). La Corte di cassazione ha affermato che anche il concepito ha diritto alla vita.
Per la tutela penale della v. umana ➔ omicidio.
Economia
Costo della vita
La somma occorrente in un determinato periodo di tempo per il mantenimento di un individuo o, meglio, di un’unità familiare. Le variazioni di questa somma nel tempo, in conseguenza delle variazioni dei prezzi dei beni e dei servizi, sono evidenziate mediante appositi numeri indici di solito elaborati da uffici pubblici. Gli indici del costo della v. permettono infatti di individuare l’andamento dei salari e stipendi reali, in periodi di variazione del valore della moneta, e di rivedere di conseguenza i salari e gli stipendi nominali automaticamente (sistema della scala mobile; ➔ scala) o attraverso nuovi accordi. Per quanto riguarda la costruzione degli indici del costo della v. occorrono: a) una scelta razionale dei beni e servizi ritenuti necessari all’esistenza; b) un’accurata determinazione dell’importanza relativa del consumo dei beni stessi nei bilanci individuali e familiari; c) una rilevazione periodica e attendibile dei relativi prezzi al minuto. I numeri indici possono essere calcolati a consumi immutati (quando si vogliono mettere in evidenza le variazioni dei prezzi di un numero abbastanza grande di beni e servizi, le cui qualità e quantità consumate si immaginano costanti) o a consumi modificati (quando, oltre che dalle variazioni dei prezzi, tali indici dipendono dalle variazioni del tenore di vita determinate dalle condizioni del mercato e dal mutare delle abitudini).
Filosofia
Filosofia della vita
Espressione (ted. Lebens;philosophie) usata per denominare un orientamento filosofico affermatosi all’inizio del 20° sec. con rappresentanti di varia provenienza speculativa: W. Dilthey, G. Simmel, M. Scheler, L. Klages, e specialmente M. de Unamuno, J. Ortega y Gasset, E. d’Ors ecc. Gli autori di questo orientamento, accentuando o sviluppando motivi romantici, schopenhaueriani, nietzschiani, pongono talora un’antinomia tra v. e ragione, oppure riconducono le esigenze della ragione e i valori dello spirito al processo della v. e alle sue leggi immanenti, di cui sarebbero manifestazione. Il risultato di questa «vitalizzazione della ragione» (Ortega) è spesso l’irrazionalismo, il misticismo, l’amoralismo.
Fisica

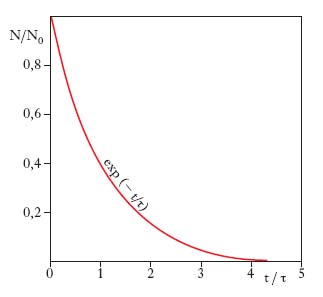
La v. media di un oggetto microscopico in uno stato instabile è l’intervallo di tempo che in media intercorre tra quando ha inizio l’osservazione del sistema e quando ha luogo una transizione spontanea verso uno stato più stabile. Gli oggetti microscopici isolati (molecole, atomi, nuclei, particelle elementari) possono compiere spontaneamente transizioni o decadimenti verso stati a energia (o massa, stante la loro equivalenza) minore con emissione della differenza di energia sotto forma (oltre che di energia cinetica dei prodotti di decadimento) di fotoni nei decadimenti radiativi di molecole o atomi o nuclei inizialmente in stati eccitati, elettroni (e neutrini) o particelle alfa nei decadimenti radioattivi dei nuclei, altre particelle nei decadimenti delle particelle elementari. Questi decadimenti sono caratterizzati dal fatto che se al tempo t=0 sono presenti N0 sistemi microscopici nello stato iniziale, si osserva sperimentalmente che entro il tempo t decade una frazione pari a 1−exp(−t/τ), dove τ è la v. media dello stato (pari al valore medio dell’intervallo di tempo che intercorre tra quando ha inizio l’osservazione e quando ha luogo il decadimento), ovvero al tempo t non sarà decaduta una frazione N/N0=exp(−t/τ) (legge esponenziale del decadimento, fig. 2). Ciò sta a indicare che la probabilità di decadimento in un intervallo di tempo dt molto piccolo rispetto a τ è costante e pari a dt/τ; infatti, per un gran numero di oggetti microscopici tale probabilità può essere espressa mediante la frequenza dei decadimenti −dN/N, dove −dN è il numero di decadimenti nell’intervallo dt, e quindi si ha dN/N=−dt/τ, espressione che, integrata tra 0 e t, si traduce nella legge esponenziale N=N0exp(−t/τ). La v. media è soggetta alla dilatazione relativistica del tempo e va quindi misurata nel sistema di riferimento di quiete dell’oggetto microscopico; se questo si muove con velocità v rispetto all’osservatore, la v. media risulterà pari a τ/√‾‾‾‾‾‾‾‾‾‾‾‾‾‾ 1 − (v/c)2 con c velocità della luce nel vuoto. Ogni sistema microscopico instabile ha una v. media caratteristica, che per i diversi sistemi può assumere valori molto diversi: gli stati eccitati atomici hanno v. media dell’ordine di 10−8 s; i decadimenti radioattivi, v. comprese tra frazioni di secondo e miliardi di anni; i decadimenti delle particelle elementari quasi stabili hanno v. medie comprese tra 10−6 e 10−13 s, quelle che danno luogo a decadimenti radiativi tra 10−16 e 10−20 s e infine, quelle dette instabili (o anche risonanze), hanno τ dell’ordine di 10−23 s (➔ particelle elementari).
La misura della v. media fornisce informazioni dirette sulle interazioni che causano il decadimento (➔ deboli, interazioni; forti, interazioni). Il carattere probabilistico insito nella definizione della v. media deriva direttamente dalla meccanica quantistica: non è possibile prevedere quando una data particella subirà il decadimento ma solo calcolare la probabilità di decadimento. Un’altra conseguenza della natura quantistica degli oggetti microscopici è il collegamento, stabilito dal principio di indeterminazione tra la v. media e l’indeterminazione in energia ΔE dello stato iniziale: ΔE∙τ∿ℏ dove ℏ è la costante di Planck ridotta. Quest’ultima relazione permette di misurare la v. media, quando questa sia estremamente breve (∿10−23 s) e quindi non determinabile direttamente dall’osservazione del decadimento esponenziale, misurando la larghezza ΔE della distribuzione di energia dei prodotti di decadimento.
Matematica
In statistica demografica e in matematica attuariale, v. media (o v. media alla nascita, o speranza media di v.), il numero medio di anni che un individuo di una certa popolazione, supposta in equilibrio demografico, deve aspettarsi di vivere (tale numero è dedotto, con opportuni calcoli, da tavole statistiche di sopravvivenza e mortalità); v. media residua, per un individuo di età x, il numero medio di anni che l’individuo di tale età deve aspettarsi di vivere ancora; v. mediana (o v. probabile), il numero di anni che un individuo di età x potrà oltrepassare con il 50% di probabilità, equivalente al periodo di tempo che deve trascorrere perché i sopravvissuti di una data classe di età si riducano alla metà.