
riproduzione
Biologia
Il processo o l’insieme dei processi mediante i quali gli esseri viventi perpetuano la propria specie, producendo nuovi individui che ripetono ciclicamente le medesime fasi di sviluppo fino al raggiungimento della forma definitiva propria di quelli da cui sono derivati. La r. può essere asessuata o sessuata.
R. asessuata
La r. asessuata è prevalentemente (ma non esclusivamente) adottata da organismi unicellulari (Batteri ecc.) ma anche da alcuni organismi pluricellulari, come per es. l’idra. Essa avviene con modalità simili per cellule aploidi. In ogni caso, dà origine a una progenie di individui geneticamente identici (a meno di errori o mutazioni) tra loro e all’organismo parentale.
R. sessuata
Caratteri generali
La r. sessuata, che è stata adottata dalla stragrande maggioranza degli organismi pluricellulari ma anche occasionalmente da alcuni unicellulari (Protozoi, lieviti), comporta la messa in comune e la ripartizione riassortita dei genomi di due individui diversi della stessa specie. Produce una progenie di individui che potranno essere geneticamente diversi l’uno dall’altro nonché da entrambi i genitori. Essa implica un meccanismo, la meiosi, per due casi tipici. Nel caso di organismi diploidi la meiosi porta alla formazione di cellule aploidi specializzate (i gameti), che sono capaci di fondersi (fecondazione), ridando origine a una cellula diploide (lo zigote): da questa, per r. mitotica, si formerà il nuovo individuo, che, alla stregua dei due organismi genitori, conterrà in ogni sua cellula due copie di ciascun cromosoma, una di origine paterna e una di origine materna.
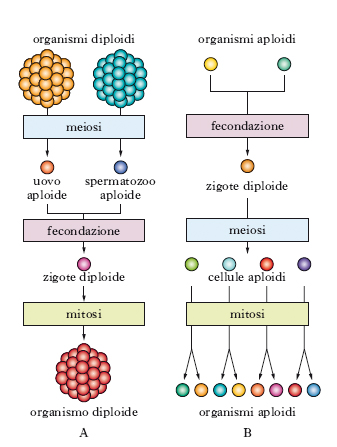
Nel caso di organismi aploidi, la meiosi segue direttamente la fecondazione e sono le cellule aploidi a riprodursi mitoticamente (fig. 1). Nel corso del processo di aploidizzazione, si ha il riassortimento casuale dei cromosomi provenienti dall’uno o dall’altro dei genitori; può altresì avvenire uno scambio fisico (ricombinazione genetica) di porzioni di cromosomi corrispondenti (omologhi) di provenienza paterna e materna (➔ meiosi). Il meccanismo descritto fa sì che ogni cellula della nuova generazione aploide porti un nuovo assortimento di geni, derivanti in parte dall’una e in parte dall’altra delle due cellule parentali della precedente generazione aploide. Così, tramite cicli di aploidia, fusione, diploidia e meiosi vengono smembrate e riassortite combinazioni di geni.
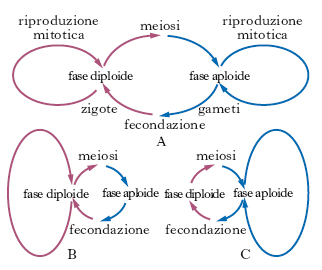
Il ciclo di r. sessuata, implicante, come si è visto, un’alternanza di generazioni mitotiche, può essere rappresentato con assoluta generalità nel diagramma della fig. 2. Ogni organismo dotato di r. sessuata passa infatti nel suo ciclo vitale attraverso fasi di vita aploide e diploide. La predominanza dell’una o dell’altra fase può essere schematizzata dall’estensione dei cicli laterali, che possono anche mancare. La predominanza della fase diploide ha importanti implicazioni dal punto di vista dell’evoluzione. Le cellule specializzate per la fusione sessuale, cioè i gameti, sono morfologicamente ben distinte in due tipi. Di norma, un tipo di gamete (gamete femminile o ovocellula) è di notevoli dimensioni, mentre l’altro (gamete maschile) è normalmente di piccole dimensioni e capace di movimento attivo.
Nella maggior parte degli organismi superiori è ben netta la distinzione tra le cellule della linea germinale, da cui deriverà, per meiosi, la prossima generazione di gameti aploidi, e le cellule della linea somatica, che, in attiva r. vegetativa per mitosi, costituiscono tutto il resto dell’organismo e non hanno continuità con la progenie. Da un certo punto di vista è utile affermare che il ruolo delle cellule della linea somatica è solo quello di permettere alle cellule della linea germinale di sopravvivere e di propagarsi. È in queste ultime che è scritta la storia evolutiva della specie, in quanto la selezione naturale, operando selettivamente sugli organismi, elimina combinazioni di geni non adatte.
Ogni individuo sessuato diploide contiene in ogni sua cellula un doppio corredo cromosomico. A livello di una particolare coppia di cromosomi esistono tuttavia differenze che determinano il sesso. Nei Mammiferi e in diversi altri animali le differenze riguardano i cromosomi detti X e Y. La femmina, produttrice di cellule uovo, possiede due copie del cromosoma X in ogni sua cellula somatica, mentre il maschio, produttore di spermatozoi, possiede due cromosomi solo parzialmente omologhi, X e Y, che nella meiosi segregano normalmente. Di conseguenza, mentre una cellula uovo contiene necessariamente nel suo corredo aploide un cromosoma X, uno spermatozoo può contenere alternativamente un cromosoma X (e dar luogo, dopo fecondazione, a un embrione femminile diploide XX) o un cromosoma Y: in questo caso, dall’uovo fecondato si svilupperà un individuo di sesso maschile, caratterizzato da una coppia di cromosomi XY. Mentre lo spermatozoo contribuisce alla formazione dell’embrione con il solo apporto del proprio corredo cromosomico, l’embrione utilizza l’intero citoplasma materno per il suo sviluppo; avviene così che caratteri portati da DNA extracromosomici (per es., DNA mitocondriale) vengono ereditati per via materna.
R. sessuata ed evoluzione
Numerosi argomenti sono stati proposti per giustificare la straordinaria prevalenza della r. sessuata nel mondo vivente. I più importanti possono essere riassunti nel modo seguente: a) la varietà di nuove combinazioni di geni, resa possibile a ogni evento generativo dal meccanismo della ricombinazione meiotica, allargando lo spettro dei genotipi possibili, sarebbe vantaggiosa per il mantenimento di una specie in un ambiente variabile; b) la riunificazione in un unico individuo della progenie di mutazioni vantaggiose casualmente avvenute su geni diversi nell’uno e nell’altro dei genitori, resa possibile dalla r. sessuata, permette la rapida diffusione nella popolazione della combinazione vantaggiosa. Nel caso di r. asessuata, viceversa, ciascuno dei due organismi inizialmente mutati darebbe origine a un clone che tenderebbe a eliminare l’altro dalla popolazione. Per avere la doppia mutazione vantaggiosa occorrerebbe attendere che nel clone mutato vincente si verificasse casualmente, tra le tante possibili, l’altra mutazione vantaggiosa. Calcoli quantitativi mostrano come il meccanismo della r. sessuata, invocato per spiegare una riduzione nel tempo necessario all’evoluzione rispetto a quello valutato nell’ipotesi di successive mutazioni indipendenti, porti un effettivo vantaggio nel caso di popolazioni numericamente superiori a 106 individui; c) in assenza di r. sessuata, nel genoma di un organismo diploide si accumulerebbero mutazioni letali (o comunque svantaggiose) recessive, verificatesi nel corso di generazioni successive. Tali mutazioni, infatti, non sarebbero in alcun modo controselezionate, data la presenza nel genoma di una copia (allele) corretta per ogni gene mutato. Solo individui in cui si verificassero indipendentemente mutazioni letali (o svantaggiose) a carico delle due copie dello stesso gene sarebbero rimossi dalla popolazione. A lungo andare l’organismo si comporterebbe di fatto come aploide, non avendo più la disponibilità di un secondo allele per ogni gene. Il meccanismo di r. sessuata, che implica il riassortimento di due copie mutate nel 25% della progenie in individui incapaci di sopravvivere o comunque svantaggiati, favorisce la rimozione della mutazione dannosa dalla popolazione, che rimane a tutti gli effetti diploide. Il grande vantaggio della diploidia risulta evidente se si considera che, in organismi diploidi, una copia di ciascun gene è libera di mutare senza danneggiare la funzione cui presiede il gene interessato. Duplicazioni geniche derivanti con una certa frequenza da errori nella replicazione del DNA, seguite da ricombinazione meiotica con alleli mutati, possono portare (e hanno in effetti portato in molti casi) a un arricchimento genetico che non è possibile per organismi aploidi e, comunque, in assenza di r. sessuata. Si può pertanto affermare che quest’ultima ha creato le condizioni per una più rapida evoluzione in piante e animali superiori.
Stabilità genomica e alternanza delle fasi riproduttive
La scoperta, relativa ai geni delle immunoglobuline, di un riarrangiamento genico che interviene nel corso dello sviluppo tra la fase embrionale e la fase di completo differenziamento (riarrangiamento che porta a congiungere tra loro porzioni di geni distanti lungo il genoma embrionale) ha messo in evidenza come nella r. mitotica delle cellule somatiche possano aversi eccezioni alla regola dell’identità genetica tra cellule figlie di una cellula di riproduzione mitotica e la cellula madre. Meccanismi di amplificazione di determinati geni o di porzioni non codificanti e meccanismi di trasposizione e riarrangiamento hanno gettato nuova luce sul problema dello sviluppo differenziato dei diversi tessuti e organi negli organismi superiori (sicuramente legato, peraltro, alla regolazione dell’espressione genica; ➔ regolazione). Il problema proposto da queste e altre recenti osservazioni riguarda il modo di conciliare questa instabilità, o plasticità, del genoma con la conservazione dei caratteri ereditari della specie. Una delle ipotesi che si possono avanzare è che il programma contenga un sottoprogramma della propria instabilità. Mentre non si è in grado di rispondere compiutamente a domande di carattere così generale, possono essere dati per chiariti i due seguenti punti fondamentali: a) i riarrangiamenti genomici irreversibili che hanno luogo nelle cellule della linea somatica negli animali superiori, e che possono essere connessi con il programma di sviluppo somatico, non interessano la stabilità e la conservazione dei caratteri ereditari, proprio in quanto cessano fisicamente di esistere con la morte dell’organismo; la conservazione dei caratteri è affidata viceversa alle cellule della linea germinale. Nel caso delle piante e degli organismi unicellulari, in cui le cellule vegetative sono o possono essere i progenitori di cellule germinali, i riarrangiamenti genomici, se avvengono, devono essere reversibili (per es., copia e trasposizione di un gene con conservazione della copia originale); b) la possibilità di variazioni discontinue accelera i tempi dell’evoluzione. La possibilità di amplificazione ed eventuale trasposizione di intere sequenze permette divergenze rapide, che possono portare a incompatibilità riproduttiva e quindi a eventi di speciazione.
La r. delle piante
Nel mondo vegetale la conservazione della specie è assicurata in molti modi che riflettono la più lunga storia naturale delle piante, e la loro maggiore flessibilità adattativa rispetto agli animali, in relazione sia alla minore differenziazione sia al regime più vario e rigoroso di rapporti con l’ambiente.
R. sessuata
Negli organismi vegetali a più semplice organizzazione manca o è rarissima la riproduzione sessuata. Essa può identificarsi con la divisione cellulare (talora amitotica, o diretta, per scissione, gemmazione, politomia ecc.) come nei procarioti (Batteri, Cianobatteri); oppure può avvenire per mezzo di cellule differenziate (esoconidi, endoconidi) o per frammentazione di colonie o di talli, da cui si staccano pezzi come ormogoni, soredi, coremi, oidi ecc. (procarioti e parecchi esempi in alghe, funghi, licheni).
R. asessuata
Processi di moltiplicazione agamica per frammentazione e formazione di organi di moltiplicazione ricorrono naturalmente anche nelle piante superiori, ove la riproduzione agamica è fatta anche artificialmente (talee, margotte, propaggini ecc.) e i frammenti e organi che servono alla produzione asessuata di nuovi individui (geneticamente identici alla pianta madre, con la quale costituiscono cloni), possono provenire dal fusto (bulbilli, gemme quiescenti, rizomi ecc.), dalla foglia (intera o in parte, bulbilli ecc.), da foglie e fusto (bulbi), dalla radice (apice, gemme avventizie, bulbilli o tuberi radicali), dal fiore (le cui parti talora si modificano espressamente); infine, si possono ottenere nuove piante, agamicamente, da tessuti coltivati in vitro.
Nelle piante inferiori (Alghe e Funghi) è molto diffuso un fenomeno, che non ha riscontro nel mondo animale, consistente nella produzione di spore e propaguli asessuali unicellulari, sia durante la fase diploide sia durante quella aploide del ciclo biologico.
R. e ciclo biologico
Nelle piante si alternano due generazioni: una aploide, l’altra diploide. La prima a maturità produce i gameti in ricettacoli detti gametangi; la seconda inizia con lo zigote. I gametangi, talora semplicissimi, possono essere di varia fattura e complessità e, in qualche caso, possono contenere gameti femminili e maschili (Fucali). Il fatto che durante la fase vegetativa dei gametofiti (aploidi) o dello sporofito (diploide) s’inseriscano modi collaterali di riproduzione (agamica) aumenta enormemente la possibilità di propagazione di certe specie (si pensi a tanti funghi patogeni) in ambienti favorevoli; in questi casi, il ricorso alla r. sessuata coincide per lo più con condizioni sfavorevoli per deficienza di alimenti o di acqua, per eccessi termici, per il sopraggiungere della stagione sfavorevole. In quest’ultimo caso vengono a volte prodotti organi speciali adatti a superare tali periodi avversi (spore durature, sclerozi ecc.).
La durata e lo sviluppo relativo delle due generazioni dipendono dal momento del ciclo biologico in cui avviene la meiosi, che può essere iniziale o zigotica; intermedia; terminale o gametica. Il ciclo con meiosi terminale o gametica è quello proprio degli animali e ricorre nelle piante soltanto nelle Diatomee e nelle Alghe brune Fucali, che hanno, come la maggior parte degli animali, ciclo diplonte e nelle quali, quindi, le cellule aploidi prodotte per meiosi funzionano direttamente come gameti. Se la meiosi è iniziale, e quindi precoce, verificandosi alla germinazione dello zigote, resta eliminata tutta la generazione diploide (Alghe verdi Coniugate, molti funghi ecc.). Se la riduzione meiotica è intermedia, cioè cade in qualche momento del ciclo biologico e separa le due generazioni, queste sono entrambe rappresentate e il loro sviluppo può essere relativamente equilibrato (Alghe brune isogenerate, Muschi), oppure una delle due generazioni può nettamente prevalere, per durata e sviluppo, sull’altra. Un tratto caratteristico del regno vegetale è che, man mano che si sale nella scala evolutiva, la meiosi diventa sempre più ritardata; ciò comporta che progressivamente diviene preponderante la fase diploide del ciclo biologico. Per es., nelle piante superiori essa è nettamente preminente e occupa gran parte del ciclo, mentre la fase aploide è ridottissima sia per durata (da qualche giorno a poche settimane, in genere) sia per lo sviluppo, essendo accolta nelle strutture fiorali (che, a loro volta, sono effimere).
Sviluppo delle due generazioni
La crescita vegetativa può esaurirsi in qualche mese, o in uno o due anni (piante annuali, con forme invernali e primaverili, e biennali); o continuare per un periodo indefinito, molto variabile, distinto in una fase giovanile, dopo la quale la pianta può fiorire una sola volta e morire (Agave americana, dopo 10-70 anni); o fiorire e fruttificare ogni anno. In certe piante, annualmente vengono prodotti prima i fiori (Forsythia, Calicanthus, Caesalpinia ecc.) e poi le foglie. In certe piante bulbose (tulipano, giacinto, zafferano ecc.), l’accrescimento vegetativo si conclude con la formazione degli organi di r. vegetativa (bulbi, cormi) e sessuata (fiori). In altri casi (piante annuali e perenni), dopo un periodo di accrescimento vegetativo l’accrescimento di organi vegetativi e riproduttivi procede contemporaneamente.
Momento della riproduzione
Alcune piante (orzo) hanno particolari adattamenti fotoperiodici, con la presenza, nell’ambito della stessa specie, di varietà regionali brevidiurne e longidiurne, secondo l’area geografica di distribuzione. Il fenomeno della fioritura è largamente dipendente dalla luce e dalla durata relativa del giorno e della notte (fotoperiodo) e, accanto ad altri tipi, si distinguono sotto questo riguardo anzitutto le piante brevidiurne e neutrodiurne, prevalenti alle basse latitudini, da un lato e dall’altro dell’equatore; e le piante longidiurne, proprie delle medie e alte latitudini, talora con forme invernali e primaverili e con esigenze di un periodo a bassa temperatura (➔ vernalizzazione).
La r. degli animali
Nella r. sessuata o gamica il nuovo individuo deriva da particolari cellule riproduttive differenziate, i gameti, che nella maggioranza dei casi si uniscono due a due nel processo di fecondazione (anfigonia), o anche da una sola cellula sessualmente differenziata (partenogenesi). Nella anfigonia i gameti si fondono in un’unica cellula fecondata, lo zigote (anfionte), da cui trae origine il nuovo individuo.
Nel ciclo riproduttivo degli organismi, la r. agamica può avvicendarsi con quella sessuata, oppure l’anfigonia con la partenogenesi; in un caso e nell’altro si parla di alternanza di generazioni: metagenesi nel primo caso; eterogonia nel secondo.
R. agamica
Negli organismi unicellulari la r. avviene previa divisione diretta o indiretta del nucleo, e cioè per scissione o emitomia, per gemmazione o divisione ineguale o per politomia o divisione multipla. La divisione nucleare può essere complicata per la presenza di due tipi di nucleo, il macro- e il micronucleo, come nei Ciliati. Anche alcuni Metazoi, Celenterati, Platelminti, Anellidi ecc., possono riprodursi agamicamente per divisione e per gemmazione. La divisione talora consiste nella frammentazione del corpo del genitore in due parti uguali che si completano successivamente, oppure la divisione può ripetersi a diversi livelli del corpo e le parti che ne risultano danno origine a tanti individui figli disposti in serie lineare che poi si separeranno (per es., Turbellari, Anellidi); o ancora, come nella strobilazione, caratteristica degli Scifozoi, la divisione dell’individuo originario (scifistoma) è multipla e le porzioni in cui esso si scinde sono tanto meno giovani quanto più si allontanano da quello.
Nella gemmazione, sull’individuo genitore si forma una protuberanza o gemma che poi, separatasi, si accresce dando origine al nuovo individuo. Nei Protozoi corrisponde a una divisione ineguale del corpo citoplasmatico, ma uguale del nucleo, e può essere esogena ed endogena. Le gemme possono talora differenziarsi in gameti. Nei Metazoi la gemmazione può dar luogo a colonie temporanee di individui, oppure, come nei Poriferi, Celenterati, Briozoi, Tunicati, la colonia è permanente e composta da un insieme di individui derivati asessualmente dall’individuo originario, fondatore della colonia, che possono essere tutti uguali (colonie omeomorfe) o differenziati morfologicamente e fisiologicamente (colonie eteromorfe). La gemmazione endogena o endogenesi può avvenire anche nei Metazoi: così, le gemme interne o gemmule delle Spugne e gli statoblasti dei Briozoi, che sono capaci di resistere a condizioni ambientali sfavorevoli.
R. sessuata
Nella r. sessuata i gameti possono essere morfologicamente identici (isogameti) anche se fisiologicamente diversi e si ha allora la isogamia propria di vari Protozoi e vegetali inferiori. Quando i gameti sono diversi, talora molto diversi, si ha l’anisogamia e i due anisogameti sono detti rispettivamente microgamete o spermatozoo, il gamete maschile più piccolo e più attivo; macrogamete o uovo, quello femminile più grande e meno mobile, o immobile. La singamia, cioè la fusione dei due gameti, consta della cariogamia o fusione dei due nuclei e della plasmogamia o fusione dei due citoplasmi. In questo processo ha una importanza fondamentale la meiosi che determina la condizione aploide nel nucleo dei due gameti, i quali sono destinati a fondersi, con la fecondazione, in una unica cellula, lo zigote, da cui ha inizio il nuovo individuo. Nello zigote la cariogamia, raddoppiando il numero dei cromosomi, ripristina quello caratteristico della specie (condizione diploide) e che compare in tutte le cellule del corpo (somatiche) e nelle cellule riproduttive (germinali) prima che sia avvenuta la riduzione del numero dei cromosomi. Questa si compie, in genere, durante la gametogenesi, immediatamente prima della formazione dei gameti; in tal caso si parla di meiosi terminale o gametica. Nella meiosi iniziale (Sporozoi, molti vegetali inferiori) e in quella cosiddetta intermedia (Foraminiferi, vegetali inferiori e tutte le piante superiori) la riduzione avviene invece, rispettivamente, subito dopo la formazione dello zigote oppure con la formazione delle spore dall’organismo a cellule diploidi.
Nei Protozoi i gameti derivano da trasformazione di individui ordinari, oppure da individui gametogeni, gamonti o gametociti, macro- e microgametociti che producono rispettivamente macro- e microgameti, cioè anisogameti. Lo zigote in certi casi (Sporozoi) si moltiplica per sporulazione che dà luogo alla formazione di stadi duraturi, le spore. La fecondazione, sempre nei Protozoi, si chiama anfimissi, se i due individui che si originano sono di diversa origine; automissi, se sono derivati dalla medesima cellula madre; autogamia, se l’unione interessa soltanto i due nuclei derivati dalla divisione dell’individuo iniziale, senza che questo si sia diviso. Caratteristica dei Ciliati è la coniugazione, cioè l’unione temporanea di due individui che hanno il valore di gamonti in determinati periodi e in date condizioni. La separazione dei due coniuganti avviene dopo complicate trasformazioni del duplice apparato nucleare (macro- e micronucleo) di ciascun coniugante, che fondamentalmente consistono nella distruzione del macronucleo e nella fusione di parti dei micronuclei che si scambiano fra i due coniuganti, previe successive divisioni maturative di ciascun micronucleo originario. Si ha perciò dopo la fecondazione un rinnovamento dell’apparato nucleare in ciascun gamonte, che, separatosi dall’altro, inizia una nuova serie di r. agamiche.
Nei Metazoi soltanto alcune cellule si differenziano per la funzione riproduttiva: sono appunto le cellule germinali, che generalmente si trovano nelle gonadi. Le gonadi femminili si chiamano ovari, quelle maschili testicoli; nelle gonadi si compiono la ovogenesi e la spermatogenesi che conducono rispettivamente alla formazione delle ovocellule o uova e delle cellule spermatiche o spermatozoi. Non sempre le gonadi sono portate da individui diversi, come nel dioicismo o diecia o gonocorismo. Se sono portate invece dallo stesso individuo che produce gameti sia maschili sia femminili si ha la monoecia o monodieciaismo o ermafroditismo. In molti animali ermafroditi una stessa gonade (ovotestis) può produrre i due tipi di gameti. Le gonadi, più o meno varie e complesse, sono in genere associate ad apparati per l’emissione dei prodotti sessuali, per assicurare l’incontro e l’unione dei gameti, e a un complesso di ghiandole e organi vari che contribuiscono alla funzione riproduttiva e vengono indicati come organi sessuali accessori.
Ma negli animali a sessi separati, talora, i sessi non si distinguono soltanto per la presenza di testicoli o di ovari, cioè per questi caratteri sessuali primari, cui corrispondono come s’è detto strutture sessuali accessorie, ma anche per un complesso di caratteri differenziali (di aspetto, di comportamento ecc.) che non riguardano direttamente l’apparato riproduttore. Sono questi i caratteri sessuali secondari che stabiliscono il dimorfismo sessuale, la cui determinazione può dipendere o dalla attività delle gonadi (caratteri eusessuali) o dagli stessi fattori determinanti il sesso delle cellule germinali (caratteri somatosessuali).
Importanza fondamentale ha il problema della determinazione del sesso, che può essere genotipica, cioè legata a un determinato corredo cromosomico, o fenotipica, dipendente da fattori estrinseci ai gameti. La fecondazione può essere interna quando lo spermio raggiunge l’uovo all’interno del corpo della femmina, già nell’ovario o nell’ovidutto; o esterna, come accade nella maggioranza degli animali acquatici, specialmente Invertebrati; in questo caso l’incontro e l’unione dei due gameti avviene nell’acqua e, poiché le probabilità d’incontro dei gameti sono più scarse che nella fecondazione interna, il numero dei gameti prodotti è grandissimo. Alla fecondazione esterna segue lo sviluppo dell’uovo nell’ambiente esterno, mentre nella fecondazione interna lo sviluppo si compie nella maggioranza dei casi all’interno del corpo materno, a volte in un particolare organo che protegge l’embrione (utero).
Nei Mammiferi l’embrione per accrescersi trae il proprio nutrimento dalla madre, contraendo stretti rapporti funzionali con essa attraverso un complesso organo vascolare, la placenta. In tutti i Vertebrati e in alcuni Invertebrati, particolari organi di diversa struttura e origine, gli annessi embrionali, provvedono alla nutrizione, respirazione e protezione dell’embrione.
I fenomeni della r. sessuata si verificano di norma negli animali col raggiungimento dello stato adulto, ossia di completo sviluppo, e della maturità sessuale, che in molte forme si accompagna a profonde trasformazioni somatiche. Le forme larvali e quelle giovanili non sono in genere in grado di riprodursi, ma nella pedogenesi, come accade in alcuni Insetti, sia le larve sia le ninfe possono diventare sessualmente mature e produrre uova ed embrioni sviluppantisi per partenogenesi, o per anfigonia. Quando un animale è capace di riprodursi tanto durante la vita larvale quanto allo stato adulto si ha la dissogonia. La capacità di r. di un organismo ancora in abito larvale o giovanile è detta neotenia: ne è un esempio classico l’Urodelo Ambystoma mexicanum, che nella sua forma branchiata di axolotl si riproduce senza metamorfosi. Alla maturità sessuale, l’attività riproduttiva degli animali assume per lo più una periodicità: a periodi di inerzia delle gonadi si alternano periodi di intensa attività funzionale. Ciò è dipendente dai fattori esterni ambientali e dei fattori interni fisiologici, attività ormoniche principalmente, che sono intimamente collegate alla situazione endocrina dell’animale e dalle quali derivano cambiamenti del metabolismo. In questa attività periodica in relazione alla stagione riproduttiva rientra l’estro periodico, o calore, nelle femmine dei Mammiferi, che coincide con la preparazione dell’uovo alla fecondazione. La periodicità dei fenomeni di r. negli animali può essere, infine, connessa ai cambiamenti di fase lunare o addirittura a certe ore del giorno.
Economia
Costo di r.
È il costo che si dovrebbe sopportare per riprodurre direttamente una merce (r. fisica) o per procurarsela mediante lo scambio (r. economica) o per produrre o ottenere attraverso lo scambio un surrogato (r. economica per via di surrogati), al quale si adeguerebbe il valore della merce suddetta, indipendentemente dal costo effettivamente sopportato per produrla. La teoria del costo di r., legata al nome di Francesco Ferrara (➔), benché elaborata anche da H.C. Carey e anticipata da N.W. Senior, ha incontrato numerose critiche specie per ciò che concerne la r. fisica (che in realtà si traduce in possibilità di produzione degli stessi beni da parte di altri a costi diversi) e la r. economica per surrogati (dato che i beni succedanei arrecano una soddisfazione soltanto approssimativamente simile a quella del bene principale e quindi possono influenzarne ma non determinarne il prezzo). Il caso della r. economica per via di scambio si può invece considerare il ponte di passaggio tra la teoria del valore fondato sul costo di produzione e quella del valore determinato dalla utilità marginale.
Scienze sociali
I problemi relativi alla r. delle popolazioni umane si presentano piuttosto complessi e variamente intrecciati fra loro. Ai fattori biologici che regolano i meccanismi della fecondazione si aggiungono i fattori sociali che influenzano in diversa misura l’andamento della procreazione naturale.
I demografi hanno introdotto varie misure della riproduttività, partendo da un livello neutro di fecondità della donna in grado di assicurare il semplice ricambio generazionale: il numero delle nuove nascite compensa il numero delle morti annuali. Valori superiori a tale livello di stazionarietà comportano accrescimento della popolazione; viceversa valori inferiori portano a contrazione della popolazione e rapido invecchiamento. Il livello di stazionarietà non è fisso, ma dipende dall’influenza della mortalità sulla sopravvivenza delle donne in età feconda: nei paesi in via di sviluppo, che sono più indietro nel processo di evoluzione demografica, il numero dei figli che ogni donna in età feconda deve mettere al mondo è più elevato, in quanto si deve tenere conto che alto è il numero di donne morte di parto, di malattie infettive, di deperimento organico. Viene definito tasso di fecondità totale (TFT) il numero complessivo di figli che in media ogni donna di una certa generazione mette al mondo nel corso di tutta la sua vita riproduttiva. Tecnicamente il TFT si ottiene sommando tutti i tassi specifici per età delle donne di una certa generazione, tra i 15 e i 50 anni (periodo in cui si manifesta la r. umana). Per il calcolo, occorrerebbe seguire una generazione di donne per 35 anni (da 15 a 50 anni di età) e rilevare il numero di figli avuti anno per anno. Solo quando tale generazione di donne si è estinta in termini di r., tenendo conto della mortalità che può aver colpito alcune di esse, è possibile calcolare il rapporto tra il numero di figli avuti nel corso di tutta la vita riproduttiva e il numero delle donne della generazione. Un valore neutro di TFT accettabile per le popolazioni dei paesi demograficamente più avanzati, quali sono i paesi industrializzati, è di 2,05 figli per donna di età feconda. Tutti i paesi europei si trovano attualmente al di sotto di tale livello (v. tab.). L’Italia si attesta sul valore di 1,32 figli per donna in età feconda, dato che manifesta il trend di moderata ripresa della fecondità dopo il minimo storico registrato nel 1995 con il valore di 1,19 figli per donna. I livelli di r. tuttavia rimangono prossimi alla metà di quanto sarebbe necessario per il semplice avvicendamento numerico delle generazioni. Le conseguenze sono molteplici sia sul piano demografico (rapido invecchiamento della popolazione) sia su quello economico (contrazione della componente giovanile in età scolastica, contrazione della forza-lavoro, invecchiamento della classe produttiva, modificazione nei comportamenti di consumo), sia su quello sociale (aggravio della spesa sanitaria e pensionistica).
Tecnica
R. del suono
Operazione inversa della registrazione, e cioè procedimento per riottenere suoni, immagini, segnali elettrici ecc., che siano stati registrati su un disco, un nastro magnetico, un supporto di memoria digitale ecc. (➔ suono).
Sistemi di riproduzione
Procedimenti volti a realizzare copie di documenti, stampati, disegni, fotografie in forma direttamente leggibile e in numero di copie variabile a piacere anche se per lo più limitato a un massimo di alcune centinaia. I sistemi di r. forniscono sia copie monocrome sia copie a colori o di tipo speciale (per es., i sistemi che aggiungono un leggero effetto rilievo alla fotocopia ottenuta). Si possono raggruppare secondo diversi criteri. Per es., si distinguono sistemi a secco e sistemi a umido, a seconda che forniscano una copia asciutta oppure bagnata, a loro volta classificabili, in base al principio chimico-fisico o fisico adottato, in sistemi fotografici, termici, elettrostatici, digitali (elettronici) ecc.
Sistemi di r. cianografici
Un processo di questo tipo, appartenente alla categoria dei sistemi a umido, può fornire sia negativi, a tratto bianco su fondo azzurro, sia positivi a tratto azzurro su fondo bianco. Il primo tipo, per es., si basa sulla riduzione di sali ferrici a ferrosi, operata per esposizione di un foglio di carta (impregnata, per es., di sali ferrici come il ferricianuro di potassio) a una intensa radiazione UV: luce del sole o, meglio, adatta lampada a vapori di mercurio. Il foglio di carta impregnata, tenuto a contatto con l’originale da riprodurre ed esposto a una intensa radiazione ultravioletta, consente che nelle zone corrispondenti a un fondo trasparente, non stampato, si formi ferricianuro ferroso (azzurro), che si evidenzia immergendo poi il foglio in acqua, ottenendo in tal modo l’immagine negativa di quella originale.
Sistemi di r. diazotipici
Si basano sull’utilizzo di un foglio di carta rivestito di un sottile strato di sali di diazonio: se a esso si appoggia un originale (disegno o tratto) eseguito su foglio traslucido e si espone il tutto alla luce, questa passa liberamente nelle zone non coperte da disegno e trasforma i sali di diazonio. Poiché questi, combinandosi con fenoli e ammine, hanno la proprietà di produrre sostanze colorate, nelle zone non alterate, corrispondenti al disegno, è possibile trasformare il diazocomposto rivelando l’immagine latente: lo sviluppo può essere operato con vapori, oppure a umido, o termicamente, ottenendo sempre l’immagine positiva dell’originale.
Sistemi di r. fotografici
Il sistema di r. fotografica su pellicola, agli alogenuri d’argento, è quello usato da più tempo (➔ fotografia), anche se la diffusione della fotografia digitale, che si è avuta a partire dalla metà degli anni 1990, sta portando rapidamente alla sua sostituzione con sistemi di r. digitale. La r. mediante pellicola consiste nel fotografare l’originale per ottenere un negativo da cui si trarrà poi un positivo. Presenta l’inconveniente di essere un processo a umido, piuttosto lento e costoso, ma conserva vantaggi importanti come la possibilità di riprodurre originali di qualsiasi tipo, l’elevata fedeltà nella riproduzione, specie a colori, la possibilità di preparazione di microfilm. Diverse varianti sono state elaborate per ottenere una semplificazione del procedimento o per potere ricorrere a prodotti di minore costo. Il processo a stabilizzazione dell’alogenuro, per es., prevede l’impiego di carta fotosensibile all’alogenuro d’argento, impressionata e sviluppata come di consueto, che però non viene trattata nel consueto bagno di fissaggio (che asporta per solubilizzazione l’alogenuro d’argento non colpito da radiazioni luminose e perciò non trasformato). Infatti anziché essere asportato, l’alogenuro d’argento viene stabilizzato chimicamente con un bagno che forma un complesso insensibile alla luce. Si ha così un negativo, invertito rispetto all’originale, che può dare un positivo per fotocopiatura e che consente di ottenere direttamente un positivo da un originale pure negativo utilizzando carte particolari (autopositive) e luce di appropriata lunghezza d’onda (blu). Nei processi a trasferimento di gelatina, invece, la carta fotosensibile per formare un negativo ha un’emulsione all’alogenuro d’argento che contiene anche uno sviluppatore e un formatore di colore (tannino). Il negativo passa in un bagno alcalino che provoca la riduzione dell’alogenuro d’argento, che poi si ossida, nelle zone impressionate. I prodotti dell’ossidazione portano all’insolubilizzazione della gelatina e al suo rigonfiamento (nelle zone non impressionate). Comprimendo il negativo così ottenuto su un foglio dotato di spiccate proprietà assorbenti e preventivamente trattato con tiourea, la gelatina rigonfiata passa su di esso trasferendovi l’immagine positiva. L’alogenuro d’argento presente nella gelatina annerisce all’aria e rinforza il colore dell’immagine positiva.
Sistemi di r. termografici
Appartengono alla categoria dei processi a secco. Il primo processo di r. che utilizza il calore per la formazione dell’immagine (processo Termofax) si basa su fogli di carta trattati con sostanze che sottoposte all’azione del calore originano prodotti colorati. Un tale foglio di carta va sovrapposto all’originale da riprodurre e irraggiato con una luce infrarossa. Le radiazioni infrarosse risultano maggiormente assorbite nelle zone scure dell’originale e questo provoca un differente riscaldamento con una trasmissione al foglio sensibile di una differente quantità di calore in corrispondenza delle aree al tratto (testo o disegno). Ne deriva che i composti termosensibili si trasformano diversamente, dando origine a un’immagine visibile: esistono carte rivestite con sostanze incolori che si colorano sotto l’azione del calore. Altre carte dispongono di un fondo colorato ma nascosto da uno strato ceroso che lo ricopre e che lo svela, fondendo, solo nelle zone più esposte al calore. Le carte termiche di questo tipo presentano sensibili problemi di conservazione e deterioramento legati appunto alla temperatura (anche ambiente). Il processo di r. Imagic appartiene alla categoria dei processi termografici ma in esso l’originale da riprodurre viene ricoperto da un sottile velo di idrocarburo (non troppo volatile), è posto poi a contatto con un normale foglio di carta ed esposto a radiazioni infrarosse. Il maggiore assorbimento che si verifica nelle zone stampate dell’originale provoca l’evaporazione dell’idrocarburo che si condensa sulla superficie del foglio usato. Si ha un’immagine latente che viene rivelata depositando sul foglio di carta un inchiostro in polvere contenente una resina termoplastica che aderisce solo alle zone impregnate di liquido. Riscaldando la copia si ottiene poi la fusione della resina che accompagna l’inchiostro e quindi la fissazione dell’immagine trasferita.
Sistemi di r. xerografici
Anche questi appartengono alla categoria dei processi a secco. L’immagine da riprodurre, testo, foto o disegno, viene proiettata su di un tamburo (o piastra) ricoperto di un materiale semiconduttore (per es., selenio) che ha la capacità di annullare una carica elettrica positiva se colpito dalla luce. Nelle altre aree conserva quindi la capacità di attirare elettrostaticamente un pigmento (il toner) e formare una matrice ponendo a contatto della quale un foglio di carta si ottiene la copia voluta (➔ xerografia). Un metodo particolare è quello Electrofax, che a differenza della xerografia si serve direttamente di un foglio di carta fotosensibile.
Sistemi di r. digitale
Una particolare forma di r. ricorre all’analisi per punti della superficie di un originale (testo o immagine) e alla digitalizzazione e registrazione di corrispondenti segnali elettrici delle diverse densità rilevate. Immagini trasparenti (per es., diapositive o negativi) e originali opachi (per es., testi o foto su carta) sono così riproducibili e memorizzabili elettronicamente in vario modo tramite dispositivi a scansione (per es., scanner piani o a tamburo, o anche tramite sensori di fotocamere digitali o miniscanner digitalizzatori da tavolo; eventualmente anche tramite videocamere digitali). La r. digitale prevede così che testi e immagini acquisiti mediante digitalizzazione e successiva memorizzazione su un elaboratore, o su memorie portatili (memorie a stato solido, o dischi ottici, o altro), possano essere visualizzati su monitor televisivi, elaborati e successivamente stampati ricorrendo a stampanti a getto d’inchiostro (a volte con coloranti azoici resistenti anche all’acqua o allo sbiadimento provocato da luce UV) oppure adottando carta fotografica tramite sistemi a proiezione (anche laser) o con stampanti termiche (anche a colori). La facilità di ritocco degli originali acquisiti, con adeguati programmi di elaborazione, è un vantaggio del metodo rispetto ad altri sistemi di r., così come la possibilità di trasmissione e condivisione a distanza, via internet, delle immagini digitalizzate.