
embrione
In biologia animale e vegetale, organismo in via di sviluppo, derivato dall’uovo fecondato, dopo che dalla condizione unicellulare è passato a quella pluricellulare.
In particolare, nell’uomo è detto e. il prodotto del concepimento nella fase prenatale in cui si formano gli abbozzi dei diversi organi, senza che nell’insieme esso abbia ancora assunto le caratteristiche proprie della specie umana. L’e. è contenuto nella cavità (o vescicola) amniotica ed è avvolto dal corion. Si chiamano annessi embrionali le formazioni che non fanno parte dell’e., ma hanno funzioni protettive, respiratorie e nutritive: l’amnio, l’allantoide, la sierosa e il sacco vitellino.
Biologia
Embriogenesi
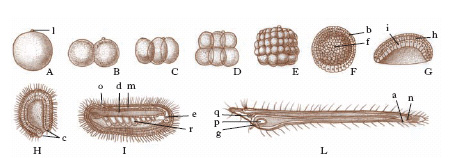
Lo sviluppo dell’embrione dall’uovo fecondato o attivato (embriogenesi o ontogenesi o sviluppo embrionale) inizia subito dopo la fecondazione (un’eccezione è data dalle uova a sviluppo partenogenetico) con il processo di segmentazione, consistente in una serie di divisioni mitotiche in conseguenza delle quali l’uovo fecondato si trasforma in una massa costituita da numerose cellule (blastomeri), che nel caso più comune delimitano una cavità centrale (blastocele). Allo stadio caratterizzato da pochi blastomeri sferici e da una cavità centrale si dà il nome di morula. Si chiama blastula lo stadio definitivo raggiunto dalla segmentazione, con blastomeri disposti intorno a una cavità di segmentazione più ampia. Segue la gastrulazione o formazione della gastrula, stadio embrionale didermico in cui i blastomeri si ripartiscono e si differenziano in due strati, l’ectoderma o ectoblasto (esterno), l’entoderma o entoblasto (interno). L’esempio più dimostrativo di questo processo, nei Cordati, è dato dalla gastrulazione dell’uovo di anfiosso (fig. 1): una metà della blastula si invagina nell’altra metà e si formano così i due foglietti e una cavità d’invaginazione, l’archenteron o intestino primitivo.
Nella maggior parte dei Metazoi, all’individuazione dell’ectoderma e dell’entoderma si accompagna o segue immediatamente quella del terzo foglietto, il mesoderma, situato tra ectoderma ed entoderma. Dai 3 foglietti embrionali derivano tutti i tessuti che formeranno i vari organi. Nei Vertebrati la regione dorsale dell’ectoderma si ispessisce e forma la piastra neurale, che poi per sollevamento e saldatura dei margini si trasforma in un organo assile, il tubo neurale, il quale darà origine al sistema nervoso centrale: encefalo e midollo spinale con tutti i loro derivati. Dal rimanente ectoderma avranno origine l’epidermide con i suoi derivati e particolari strutture, i placodi, che entrano nella formazione di taluni organi di senso. L’entoderma dà origine agli elementi epiteliali di gran parte del canale digerente, delle ghiandole annesse (fegato e pancreas) e degli organi respiratori. Il terzo foglietto forma un cordone massiccio di grosse cellule: la corda dorsale o notocorda. Ai lati della corda si originano, sempre dal terzo foglietto, i somiti. Nell’anfiosso questi s’individuano dapprincipio come due masserelle, le quali successivamente si segmentano in senso antero-posteriore, e formano tante tasche (sacchetti celomatici). La zona caudale del mesoderma, presso la regione blastoporale, rimane insegmentata e si chiama teloblastema. Da esso si originano nuovi somiti man mano che la larva si allunga. La parte dorsale dei somiti rimane segmentata, le parti laterali s’insinuano fra entoderma ed ectoderma verso la regione ventrale e, fondendosi con quelle dei segmenti contigui, danno origine alle piastre laterali, costituite da due foglietti, splancnopleura e somatopleura, separati dalla cavità celomatica. Nei Vertebrati la parte dorsale dei somiti (epimero) dà origine alla muscolatura dorsale, alle vertebre e al derma. Nella zona intermedia (mesomero) fra il somite e la piastra laterale si origineranno gli organi escretori e le gonadi. La piastra laterale (ipomero) viene a formare il peritoneo e il mesentere, in corrispondenza rispettivamente della somatopleura e della splancnopleura.
Molte cellule derivate dal foglietto mesodermico non hanno una tipica disposizione epiteliale, e sono distribuite come un tessuto lasso fra gli abbozzi dei vari organi. Costituiscono il mesenchima, da cui si originano il sistema vascolare, molte parti del sistema scheletrico e diversi tipi di tessuti connettivi.
Embriologia
L’embriologia è la disciplina biologica che studia l’ontogenesi.
Branche dell’embriologia
L’embriologia chimica, nata come disciplina autonoma con l’opera di J. Needham, Chemical embryology (1931), tratta della chimica dell’uovo e dell’embrione senza alcun riferimento biochimico ai problemi della morfogenesi; suoi problemi fondamentali sono: la chimica della fecondazione; lo studio di metabolismi particolari legati alla gastrulazione, al differenziamento e accrescimento; la struttura chimica dell’uovo, ma soprattutto lo studio dei meccanismi del differenziamento cellulare. L’embriologia comparata (o comparativa) è lo studio fatto paragonando i vari stadi di sviluppo di specie più o meno vicine sistematicamente; ha avuto grande incremento nell’ultimo quarto del 19° sec. sotto l’influenza del darwinismo, con il tentativo di armonizzare i fatti dello sviluppo embrionale con i concetti evolutivi, allo scopo di ricostruire, sulla base di tali fatti, le genealogie dei vari gruppi animali; il primo trattato di embriologia comparata è quello di F.M. Balfour (Comparative embryology, 1880-81). L’embriologia sperimentale (o causale) utilizza il metodo sperimentale per determinare le cause dei processi attraverso i quali originano le forme individuali: suo principale compito è la ricerca dei fattori che governano lo sviluppo. La realizzazione della forma specifica dell’organismo, che si modifica e si complica con il procedere dello sviluppo, è intesa, sotto questo aspetto dinamico, come una delle peculiari funzioni dell’essere vivo.
Cenni storici
L’embriologia animale, come scienza autonoma, si può dire abbia inizio con i lavori di Ch.I. Pander (1817) e K.E. von Baer (1827), ma ha, naturalmente, origini più remote. Già presso gli antichi Greci, in Aristotele e Galeno, si trovano notizie frammentarie sullo sviluppo degli animali, desunte principalmente da osservazioni dirette sull’uovo di pollo in incubazione. I primi tentativi di interpretazione dei fenomeni dello sviluppo e le prime descrizioni di e. risalgono al 17° sec. con G. Fabrici d’Acquapendente, W. Harvey e M. Malpighi. Fu Harvey a formulare per primo l’importante principio che tutti gli animali provengono dall’uovo per una graduale comparsa delle varie parti dell’essere futuro. I primi a sperimentare sull’uovo furono, fra gli altri, E. Haeckel (1859) e C. Chun (1880).
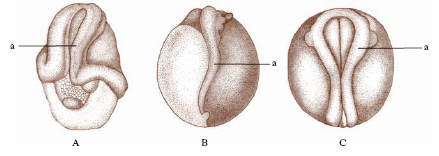
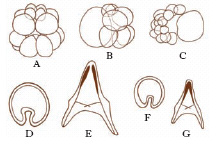

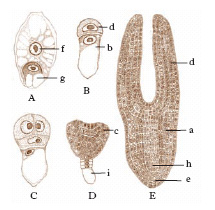
Con lo studio delle potenze dell’uovo l’embriologia sperimentale esordisce con i famosi esperimenti, quasi contemporanei (1885) ma indipendenti, di W. Roux e di L. Chabry che dimostrarono, l’uno sull’uovo di rana (fig. 2), l’altro di ascidia, come alla distruzione di uno o di alcuni dei primi blastomeri seguisse la formazione di un e. difettoso di quegli organi e di quelle strutture che normalmente si sviluppano dal blastomero o dai blastomeri eliminati. Sulle uova di ricci di mare H. Driesch (1892-1900) ottenne invece risultati sorprendenti (fig. 3), che apparentemente contraddicevano quelli di Roux e di Chabry: un blastomero isolato allo stadio di 2, 4 e perfino 8 cellule, pur segmentandosi come una parte dell’intero uovo, si sviluppava in un e. nano, ma normale e completo; il blastomero cioè si manifestava totipotente o equipotenziale. Driesch ne dedusse che la segmentazione si doveva intendere come una separazione di blastomeri indifferenti, il cui destino era fissato dalla posizione che essi venivano ad assumere nel tutto. I risultati ottenuti sulle uova dei Celenterati, Nemertini, Ctenofori, Molluschi Gasteropodi, Srafopodi, Nematodi, Anellidi, Ascidie, Teleostei, Anfibi ecc. condussero a istituire due categorie di uova, regolative e a mosaico: le prime capaci di modificare le loro parti sì da assumere la configurazione e la potenza dell’uovo intero; le uova ‘a mosaico’, al contrario, incapaci di questa regolazione, in quanto ciascuna parte dell’uovo avrebbe un proprio destino determinato. Superato così il periodo in cui insistentemente si affermò che sostanze organo-formative contenute nell’uovo rappresentassero i materiali grezzi per la formazione degli organi dell’e., fu precisato da esperimenti ulteriori il ruolo della divisione citoplasmatica nei confronti delle localizzazioni e fu appurata la generale esistenza di fenomeni regolativi nello sviluppo, che per primo Driesch aveva messo in luce. Si stabilì che nell’uovo, che è apparentemente omogeneo all’inizio nella sua costituzione e struttura, si fissano precocemente una polarità e spesso una bilateralità. Il successivo progredire di questi studi portò via via ad attenuare il contrasto fra sviluppo indeterminato (uova regolative) e sviluppo determinato (uova a mosaico), escludendo una rigida determinazione e conseguente stereotipica organizzazione iniziale dell’uovo. Si era impostato così il problema di una determinazione iniziale nello sviluppo, che sotto un nuovo aspetto si richiamava alle concezioni preformistiche o evolutive (preformazione), in contrapposto a una determinazione progressiva, come nella teoria della epigenesi. Le indagini si rivolsero allora allo studio delle potenze o capacità formative dell’uovo e soprattutto alla ricerca di un substrato materiale di tali capacità, che si credette di potere identificare nelle cosiddette localizzazioni germinali da E.B. Wilson descritte per primo nell’uovo di Dentalium (fig. 4).
La ricerca moderna
Con l’applicazione fatta più di recente dei metodi biochimici e citochimici all’embriologia, alla base dell’organizzazione iniziale dell’uovo si è posta la complessa composizione chimica del suo citoplasma, alcuni componenti del quale vanno gradualmente raggruppandosi e concentrandosi probabilmente sotto l’influenza di fattori che risiedono nella parte corticale dell’uovo (cortex), che è forse la prima a manifestarsi diversa dal resto dell’ooplasma; e così da una condizione omogenea si instaura una prima eterogeneità chimica dell’uovo, da cui derivano proprietà differenti e non più equivalenti del suo citoplasma: a questo insieme di processi è stato dato il nome di chemiodifferenziamento (J.S. Huxley) dell’uovo, che interviene ora precocemente, ora più o meno tardivamente nello sviluppo. In questo indirizzo l’e. sperimentale poggia su una vastissima sperimentazione. Sulle uova di Echinodermi a opera degli embriologi scandinavi (J. Runnström, S. Hörstadius, P.E. Lindhal ecc.) si è pervenuti a dimostrare nell’uovo l’esistenza di gradienti che interreagiscono fra loro e cui corrispondono proprietà morfogenetiche fondamentali diverse, legate a differenti proprietà fisico-chimiche e a metabolismi specifici. Questo e altri dati dimostranti l’insorgere di variazioni progressive di proprietà regionali nell’uovo secondo un asse (ciò che corrisponde all’originario concetto di ‘gradiente’ introdotto per l’uovo di Ascaris da T. Boveri), o secondo due o tre dimensioni (il concetto di ‘campo’ di A. Gurwitsch e di P. Weiss), hanno portato l’embriologia sperimentale a modificare profondamente la nozione fondamentale di localizzazioni germinali, intese come mosaico di territori organo-formativi. Si preferisce oggi considerare lo sviluppo come derivato dalla graduale interreazione tra i vari componenti chimici dell’uovo, le cosiddette ‘sostanze determinanti’; non esclusa l’azione dei geni del nucleo che, distribuiti dalla segmentazione ai vari distretti ooplasmatici, possono essere alcuni attivati, altri lasciati inerti, venendo ad avere così un’efficacia determinante sul destino delle cellule in cui l’uovo si segmenta.
Negli studi di e. sperimentale ha fatto epoca, per il completo rinnovamento che ne seguì, la scoperta dovuta a H. Spemann dell’induzione embrionale, cioè di quell’influenza determinante il destino dei vari aggruppamenti di cellule componenti l’uovo in segmentazione che talora, come Spemann riuscì magistralmente a dimostrare negli Anfibi, parte da particolari territori della blastula e della gastrula, detti centri organizzatori.
Embriologia molecolare
La scoperta delle caratteristiche strutturali delle molecole degli acidi nucleici che permettono a questi, attraverso l’alternarsi di quattro basi azotate, la codificazione delle informazioni necessarie a costruire una cellula, con tutte le sue peculiarità, ha portato a importantissime acquisizioni in campo embriologico e all’affermazione di una nuova scienza sintetica, l’embriologia molecolare. Le ricerche in cui è stata particolarmente feconda di risultati sono quelle condotte sul differenziamento cellulare e quelle sulle interazioni cellulari nella morfogenesi.
Per quanto riguarda il differenziamento cellulare, il problema principale che ci si pone è come da un’unica cellula, lo zigote, si formino cellule che, con un solo tipo di informazione genetica, possono assumere caratteristiche e funzioni diverse (cellule ossee, cellule epatiche, cellule muscolari ecc.). La diversità di queste cellule è rappresentata da una diversità delle loro proteine (emoglobina nelle cellule del sangue, miosina nelle cellule muscolari ecc.). La sequenza degli amminoacidi nelle proteine è dettata dalla sequenza dei nucleotidi nel DNA; il problema quindi è come lo stesso DNA, contenuto in tutte le cellule di un e., possa far sintetizzare proteine diverse quando queste cellule si differenziano. Sono state formulate varie ipotesi, tra cui una è che il DNA stesso divenga diverso nelle diverse cellule, ma non vi sono prove sperimentali che suffraghino questa ipotesi. Un’altra è che alcuni tratti del DNA siano funzionanti in certe cellule e altri in altre cellule e in realtà, attraverso studi di ibridazione DNA-RNA, si è osservato che ogni tipo di cellula trascrive in effetti solo il 10% del proprio DNA. Esperimenti di J. Paul hanno dimostrato che la cromatina isolata produce RNA specifici del tipo di cellula da cui proviene. Numerosi esperimenti hanno inoltre messo in luce che: il DNA isolato è capace, in presenza degli enzimi adatti, di produrre tutti gli RNA che una cellula qualsiasi è in grado di produrre; il DNA isolato, cui si aggiungano gli istoni, diviene silente, cioè incapace di sintetizzare alcun tipo di RNA; se però si aggiungono le proteine acide specifiche della cromatina di cellule differenziate, il DNA dirige la sintesi di proteine dello stesso tipo di quello delle cellule differenziate. È pertanto verosimile che siano le proteine acide cromatiniche, liberando alcuni settori del DNA dagli istoni, a consentire la trascrizione di esso in RNA specifico del tipo cellulare. Infine l’ultima ipotesi è che esistano meccanismi nello stesso citoplasma che consentono a certe cellule di uno stesso organismo di usare certi messaggi e ad altre di usarne altri; quest’ipotesi è in parte confortata da esperimenti su uova di riccio di mare in cui gli embrioni sono in grado di usare messaggi diversi nei diversi momenti dello sviluppo. In effetti, il citoplasma ha un’importanza fondamentale nel mantenimento dello stato differenziato della cellula. Verosimilmente, quando determinati geni entrano in funzione, sono sintetizzate determinate proteine, parte delle quali agisce sul nucleo per sostenere la trascrizione di particolari geni e parte agisce sul citoplasma per promuovere la stabilità di determinati mRNA o per incrementare la sintesi di alcune proteine strutturali; in tal modo il citoplasma di una cellula differenziata acquista una serie di molecole regolatrici che rinforzano la condizione differenziata. L’embriologia molecolare ha così stabilito alcuni punti fermi nella conoscenza del meccanismo del differenziamento cellulare, e cioè: il differenziamento non comporta cambiamenti nel DNA nucleare, ma una trascrizione limitata del DNA e implica inoltre cambiamenti auto-consolidantisi nel citoplasma.

Per quanto riguarda il problema della morfogenesi, l’embriologia molecolare non è riuscita ancora a raggiungere risultati paragonabili a quelli acquisiti nel differenziamento cellulare; il problema consiste nel chiarire i meccanismi che consentono alle cellule di interagire per formare tessuti e organi. Notevoli sono i contributi apportati dalle ricerche sulla morfogenesi in larve di Drosophila: il nucleo dello zigote subisce una serie di 9 mitosi sequenziali, di modo che nel citoplasma sono presenti circa 500 nuclei che non si mescolano tra loro ma conservano nel citoplasma il loro orientamento reciproco; ogni nucleo migra poi in una posizione ben definita alla periferia della cellula e si divide tre volte formando in tutto ca. 4000 nuclei, ognuno dei quali viene racchiuso in membrane citoplasmatiche; il singolo sincizio multinucleato si trasforma così in una struttura multicellulare, detta blastoderma. Le cellule da cui derivano tutte le strutture esterne dell’adulto (antenne, ali ecc.) sono contenute in regioni ben definite del blastoderma, dette dischi imaginali presunti: durante il 3° stadio larvale le cellule discendenti dalle prime sono collocate in 17 dischi imaginali separati tra loro (fig. 5). Trapiantando un disco imaginale in un adulto, le cellule del disco si moltiplicano senza differenziarsi, ma se il trapianto è eseguito in una larva, le cellule del disco si differenziano di nuovo e danno origine, dopo la metamorfosi, alla struttura (antenna, proboscide, ala ecc.) cui erano originariamente destinate. Appare quindi verosimile che le cellule embrionali di questi insetti possano trasmettere ai loro discendenti una potenzialità a dare origine a una particolare struttura e solo a quella (determinazione). In alcuni casi possono aversi degli errori occasionali; per es., le cellule del disco imaginale delle antenne, dopo trapianti successivi, possono dare origine a cellule delle zampe dell’adulto; il fenomeno è detto di transdeterminazione e fornisce l’evidenza che la determinazione, pur essendo un evento molto stabile, produce cambiamenti non irreversibili nel patrimonio genetico della cellula. Questi dati relativi ai dischi imaginali indicano l’esistenza di geni di controllo che regolano le loro caratteristiche.
L’embrione umano
Il fondatore della moderna embriologia umana è W. His, che nel 1880 pubblicò un trattato sull’anatomia dell’e. umano.
I primi stadi dello sviluppo dell’e. umano, dall’annidamento della blastocisti nel mesenchima spugnoso della mucosa uterina, risalgono approssimativamente al 7° giorno dalla fecondazione, cioè alla gastrulazione (stadi 7-8). All’11°-13° giorno è lo stadio 10, in cui ha inizio la linea primitiva che si completa nello stadio 12 di 19 giorni. La lunghezza del disco embrionale è di 0,7 mm. Alla fine della 5a settimana l’e. è completamente formato e misura 6-7 mm: è lo stadio 25 di 38 somiti. Successivamente subisce una metamorfosi con i necessari adattamenti alle condizioni di vita fetale, e gradualmente assume la forma e l’organizzazione umana. A 10 settimane si è accresciuto fino a 40-50 mm, a 20 settimane ha raggiunto i 160-200 mm. In generale si può concludere che le curve dell’accrescimento e del peso hanno aspetto sigmoide.
Bioetica
Le conoscenze acquisite con la biologia testimoniano che la vita umana esiste fin dal concepimento. L’e. umano, a partire dalla fusione dello spermatozoo e dell’ovocita, è un nuovo individuo, distinto dai suoi genitori, nella fase iniziale dello sviluppo vitale, che lo condurrà – attraverso un processo coordinato, continuo e graduale – a divenire adulto. L’e. manifesta, fin dall’inizio, una caratteristica essenziale: è una cellula intrinsecamente orientata e determinata a una ben definita e precisa evoluzione grazie alla presenza del genoma, l’informazione genetica di cui è dotato. È, infatti, il genoma a conferire all’e.: a) un’identità specifica umana, che ne stabilisce l’appartenenza alla specie umana; b) un’identità individuale, che ne marca la singolarità e unicità biologica, che lo distingue da ogni altro zigote; c) un piano-programma che ne dirige l’ulteriore sviluppo, dalla cui attuazione verrà realizzato il corpo, con una ben definita figura umana. Lo sviluppo delle potenzialità differenziative e morfogenetiche dell’e., dunque, dipende dal genoma. Potenzialità che si esprimono gradualmente e che non dicono pura possibilità di essere, ma realizzazione graduale di un essere già esistente che, per legge intrinseca, si deve autocostruire e crescere sulla base di un piano-programma che porta in sé. L’e., quindi, fin dalla fusione dei gameti, non è un potenziale soggetto umano – come taluni hanno ipotizzato – ma è un reale soggetto umano, unico e irripetibile. Dalla continuità dello sviluppo embrionale discende l’impossibilità di supporre qualsiasi salto qualitativo rispetto al momento della fecondazione. Ciò significa che l’individuazione di stadi distinti (principio di gradualità) ha valore descrittivo e che tale gradualità suppone il mantenimento da parte dell’e. della sua identità e individualità attraverso tutto il processo di sviluppo e crescita. In altre parole: l’e. umano è un individuo che, alla fusione dei gameti, inizia la sua esistenza e che da quel momento cresce secondo un naturale processo di sviluppo, che empiricamente consente di riconoscerne l’identità umana. In base ai dati scientifici premessi, è perciò innegabile l’appartenenza dell’e. alla specie umana, ossia la sua natura sostanzialmente umana. Il riferimento a un concetto sostanziale di natura è necessario per non limitare il discorso a un mero riduzionismo biologico ed esprimere, piuttosto, la coerenza che la natura umana possiede in termini sia empirici sia metafisici: una natura, cioè, che si manifesta in una struttura genetica tipicamente umana, riconoscibile in quanto tale in un momento preciso, la fecondazione, che al tempo stesso eccede la dimensione biologica, esprimendo un’identità individuale antropologicamente significativa nella propria unicità e irripetibilità esistenziale. La persona umana non ha un corpo, ma è un corpo, e lo è necessariamente dal momento iniziale del suo sviluppo, la fecondazione. La tutela della vita umana va fatta coincidere con quel momento, nel quale per la prima volta appare l’entità individuale, ossia l’embrione.
Alcune teorie gradualiste riconoscono all’e. lo statuto di individuo umano solo dopo il raggiungimento di determinati stadi di sviluppo biologico (la cariogamia, l’impianto in utero, la comparsa della stria primitiva, la presenza di un aspetto umano, la capacità di provare dolore), e per esse la tutela della vita umana sarebbe da farsi risalire a un momento distinto e successivo rispetto a quello iniziale. Sulla base di tali teorie, si sono anche coniati termini come embrione preimpianto, pre-embrione. (C. Grobstein, External human fertilization, in Scientific American, 1979) e ootide (ossia ovulo fecondato nelle prime 36-72 ore dal concepimento), per dimostrare il minor valore di un e. umano precoce e poterlo utilizzare per finalità sperimentali; si sono altresì invocati i fenomeni della gemellanza e delle chimere per negarne l’individualità. Accanto alle teorie riduzionistiche di matrice biologico-genetica, si sono anche diffuse teorie centrate sullo sviluppo di caratteristiche psicosociali, come la capacità di dimostrare razionalità umana, autocontrollo, curiosità, capacità di entrare in relazione con gli altri o di essere riconosciuti dalla propria madre come individui meritevoli di cura e tutela, o l’essere dotati di senso della comunità. Tali teorie stabiliscono che solo al raggiungimento di determinate fasi di sviluppo e crescita l’individuo umano possa essere considerato meritevole di tutela e che in taluni casi questa tutela sia riconoscibile solo nella vita adulta, o solo nell’ipotesi in cui altri soggetti decidano di riconoscerla.
Botanica
Nelle Fanerogame l’e. è la piantina abbozzata, derivata per divisione dello zigote e delle cellule successivamente formatesi, racchiusa nel seme. A un certo stadio di sviluppo l’e. arresta il suo accrescimento per riprenderlo all’epoca della germinazione, che normalmente avviene dopo qualche mese ma, se mancano le condizioni propizie, anche dopo molto tempo, talvolta parecchi anni. Durante questo periodo di quiescenza l’e. è in vita latente. Generalmente l’e. è differenziato nei 3 costituenti fondamentali del cormo: radice, fusto (piumetta) e foglie. La radice, detta radichetta, ha forma di cilindro e si continua nell’ipocotile, porzione più o meno sviluppata nelle diverse piante, chiamata fusticino o caulicino e recante al suo estremo superiore i cotiledoni o embriofilli (uno nelle Monocotiledoni; due nelle Dicotiledoni, parecchi nelle Gimnosperme). Al di sopra dell’ipocotile si trova l’epicotile, che rappresenta il primo internodo del fusto e finisce alla prima foglia di questo; le foglie o i loro abbozzi più o meno evoluti sono raccolti all’apice dell’epicotile in forma di corpicciolo di norma ovoide, che è la gemma apicale, detta piumetta, la quale nell’ulteriore sviluppo formerà l’asse epigeo, ossia il fusto. In alcune piante l’e. è indifferenziato, come nelle Orchidacee, nelle quali consta di una masserella di cellule meristematiche che non mostra neppure un accenno di differenziazione in fusto, radice e foglie. L’e. è accompagnato in certe piante dall’albume o anche dal perisperma. Nelle Pteridofite, è detto e. il primo stadio di sviluppo dello sporofito, e si distingue da quello delle Fanerogame perché lo sviluppo dell’e. nella pianta adulta avviene senza interruzione, manca cioè un periodo di quiescenza e non si sviluppa un seme.
L’embriogenesi ha inizio dopo la fecondazione dell’oosfera (formazione dello zigote), con la comparsa della membrana alla periferia di questa e procede con ripetute divisioni; tra la fecondazione e l’inizio delle segmentazioni trascorre un tempo variabile, da qualche ora a qualche giorno e perfino a vari mesi, come nel colchico, la cui oosfera, fecondata in autunno, comincia a segmentarsi nella primavera successiva. Di regola (fig. 6), lo zigote si divide trasversalmente in due cellule: l’una più piccola terminale, l’altra più grande basale e attaccata alla volta del sacco embrionale. Le due cellule possono ancora dividersi trasversalmente, sicché si forma un corpo bi- o pluricellulare, detto proembrione, dal quale si forma, per successive divisioni, l’embrione definitivo, e precisamente dalla sua cellula terminale deriva la maggior parte del corpo dell’embrione stesso, e dalle cellule sottostanti l’apice della radichetta embrionale e l’embrioforo (detto anche sospensore). Questo è un corpo pluricellulare di vario aspetto, che con un’estremità tiene sospeso l’embrione delle Fanerogame nella cavità del sacco embrionale e con l’altra aderisce alla parete di questo; ha la funzione di nutrire l’embrione. Nelle Angiosperme deriva dalle divisioni successive della cellula più esterna delle due nelle quali si divide lo zigote; nelle Gimnosperme l’origine è diversa e più complicata. In certe Pteridofite è detta embrioforo un’appendice pluricellulare dell’embrione, la quale unisce questo al protallo, sottraendovi materiali nutritizi. In altre piante, oltre alla cellula terminale, concorrono alla formazione del corpo dell’embrione una o più cellule sottostanti.
Nelle Fanerogame, all’interno della nocella con la meiosi si formano 4 macrospore, di cui 3 degenerano e una diviene il sacco embrionale, entro il quale si origina prima il gametofito femminile con una o più oosfere e, dopo fecondazione, l’embrione. Nelle Angiosperme il gametofito femminile contiene di solito 8 nuclei, 2 dei quali si fondono insieme costituendo il nucleo secondario del sacco, dal quale, in seguito alla doppia fecondazione, si forma il tessuto nutritivo (albume). In parecchie specie vegetali, alla formazione del sacco embrionale possono partecipare due, o tutte e quattro, le macrospore.
In generale, dall’oosfera fecondata proviene un solo e.; però nei pini, nei cedri e in altre Conifere la massa cellulare che deriva dalle prime divisioni dello zigote si scinde in 4 serie longitudinali di elementi, ciascuna capace di sviluppare un distinto e., sicché, in queste piante, il seme a completo sviluppo può contenere più embrioni. Alla segmentazione dell’oosfera, nelle Angiosperme, di solito precede quella del nucleo secondario da cui deriva l’albume. Durante la formazione dell’e. e dell’albume il sacco embrionale si ingrandisce a spese dei tessuti circostanti dell’ovulo, ai quali sottrae materiali nutritizi sia direttamente attraverso la sua superficie sia producendo austori di struttura diversa, generalmente ai due estremi opposti del sacco, cioè in corrispondenza della calaza e del canale micropilare. Tali austori possono prendere origine dalle sinergidi, dalle antipodi, più frequentemente dal sospensore dell’e. o da cellule dell’albume, eccezionalmente (zucca) dal tubo pollinico. L’e. oltre che dall’oosfera fecondata si può originare in vari modi: senza fecondazione per sviluppo di un’oosfera diploide per aposporia, o di un’oosfera apolide per apogamia goniale (➔ partenogenesi); da una o più cellule somatiche del gametofito, quali le sinergidi o le antipodi (apogamia somatica); infine l’e. può avere origine fuori dal sacco, da cellule della nocella o dei tegumenti dell’ovulo, quindi da cellule diploidi dello sporofito, le quali, iniziando le loro divisioni, si spingono nel sacco, dove gli e. possono raggiungere il complesso sviluppo. Quest’ultimo modo chiamato embrionia avventizia, è una forma di apomissia, è spesso associata alla poliembrionia ed è stata riscontrata in poche Angiosperme (Funkia ovata, Opuntia vulgaris ecc.).
Un capitolo interessante dell’embriogenesi si è aperto con lo sviluppo delle tecniche di coltura in vitro di cellule e tessuti. Esse possono venir utilizzate per la propagazione massiva e la produzione di un grandissimo numero di piante: data la flessibilità del materiale vegetale, è possibile ottenere, in adatte condizioni di coltura degli espianti, interi nuovi individui, quasi da qualunque tipo di cellule parenchimatiche, oltre che da quelle meristematiche. Negli espianti di tessuti, in genere, dapprima si ha la crescita indifferenziata di callo, dentro il quale, in un secondo momento, gruppi di cellule si differenziano dando origine a veri e propri e. da cui si sviluppano nuove piante. Composizione del mezzo, dosaggio di ormoni e vitamine, regime di illuminazione e qualità (colore) della luce sono fattori che influiscono sui processi di morfogenesi e costituiscono altrettante possibilità di controllo. La coltura di granelli di polline che si forma dopo la meiosi offre la possibilità di ottenere piantine aploidi, dalle quali, per poliploidizzazione spontanea (o indotta sperimentalmente) si ottengono piante diploidi omozigoti. Questa procedura offre il grande vantaggio di mettere a disposizione materiale geneticamente puro in breve tempo, con risparmio di anni di lavoro nella preparazione di materiale vegetale, per la selezione e il miglioramento delle piante.