
membrana
In anatomia, nome generico di molte formazioni macroscopiche e microscopiche, di struttura varia, caratterizzate dal prevalere della lunghezza e della larghezza sullo spessore.
Nel linguaggio della biologia, della fisica e della chimica, struttura permeabile o semipermeabile che regola il passaggio selettivo di soluti, solventi o particelle tra componenti cellulari o tra la cellula vivente e l’ambiente a essa esterno, o tra due soluzioni di caratteristiche diverse.
Anatomia e medicina
La struttura delle m., che fanno parte dei più diversi organi e apparati, è quanto mai varia: così, la membrana otturatoria, che chiude il forame otturato dell’osso iliaco, è formata da connettivo fibroso, la membrana del timpano è formata da connettivo rivestito da epidermide sulla faccia esterna e da mucosa sulla faccia mediale, le membrane sierose (pleura, peritoneo, pericardio) sono costituite da mesotelio, mentre la membrana basale (o ialina) la m. sottile e trasparente situata tra l’epitelio e lo strato connettivale sottostante (nella cute, nelle mucose, nelle ghiandole ecc.), è costituita da un sottile intreccio di fibrille reticolari. In patologia, la sindrome della membrana ialina del neonato comporta l’assenza della componente fosfolipidica (lining) che tappezza gli alveoli polmonari e ne riduce la tensione superficiale; come conseguenza, si verifica un’atelectasia polmonare.
Le membrane fetali sono quelle che avvolgono il feto, cioè l’amnio e il corion; la loro rottura, che determina la fuoriuscita del liquido amniotico, costituisce uno dei fenomeni del parto. L’osso da membrana è un osso il cui abbozzo embrionale è rappresentato da una m. di tessuto connettivo, anziché da una formazione cartilaginea.
Biologia
Membrane biologiche animali
Con il nome di membrana biologica s’intende non soltanto la m. plasmatica, considerata generalmente come una barriera che separa la cellula dall’ambiente esterno, ma anche una serie di m. delimitanti i diversi organelli subcellulari presenti nel citoplasma. La m. plasmatica avvolge ogni cellula e ne definisce l’entità mantenendo le differenze essenziali fra il suo contenuto e quello dell’ambiente esterno. È un filtro altamente selettivo che regola il flusso delle molecole verso l’interno della cellula e viceversa; mantiene le concentrazioni ioniche necessarie per lo svolgimento di molte funzioni cellulari; provvede al riconoscimento e alla comunicazione fra le cellule tramite molecole localizzate sulla sua superficie esterna. Le m. degli organelli intracellulari sono caratterizzate da funzioni specifiche dipendenti dal tipo di organello e quindi dalla presenza di particolari molecole aggregate a esse, quali vari sistemi multienzimatici, come la catena respiratoria mitocondriale, o proteine specifiche come le ribonucleoproteine legate al reticolo endoplasmatico rugoso.

Struttura
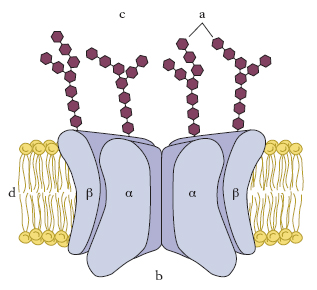
La m. appaiono costituite da tre strati, due scuri all’esterno e uno chiaro all’interno. Il modello strutturale detto modello a mosaico fluido, proposto da S.J. Singer e G.L. Nicolson nel 1972, definisce le m. biologiche come soluzioni bidimensionali di proteine globulari e di lipidi. Secondo questo modello, la m. è costituita da un doppio strato lipidico polare al quale sono associate numerose molecole proteiche (fig. 1). Alcune proteine, completamente incluse nel doppio strato lipidico, stabiliscono un legame di van der Waals con la porzione idrofobica della m. (proteine integrali di membrana); altre, sono associate alla superficie interna o esterna del doppio strato lipidico, per mezzo di interazioni ioniche o legami idrogeno tra le teste polari dei lipidi di m. e le proteine stesse (proteine periferiche di membrana). Il modello a mosaico fluido ha ipotizzato che le proteine di m. possano muoversi liberamente in senso laterale nella matrice lipidica, ma non ruotare tra una superficie e l’altra per non alterare l’asimmetria tra i due lati della m. stessa. Per i lipidi del doppio strato un tale movimento, detto anche diffusione laterale o flip-flop, è possibile, anche se è stato dimostrato essere molto lento. Secondo tale modello le proteine sono rappresentanti fondamentali della struttura delle m. mentre i lipidi riducono le possibilità di combinazione delle molecole proteiche, il che è importante per le loro funzioni biologiche. Anche i microfilamenti e i microtubuli costituenti il citoscheletro della cellula, entrano in contatto con la porzione intracellulare delle proteine di m. e ancorandole in posizioni fisse nel doppio strato lipidico impediscono loro la possibilità di combinazione e di movimento laterale.
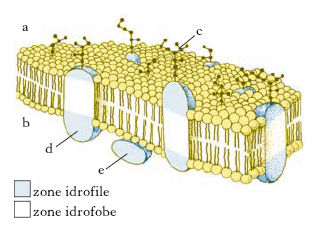
Al microscopio elettronico la linea nera esterna ha uno spessore maggiore di quella interna; ciò è dovuto a una particolare struttura associata esternamente alla m. cellulare denominata glicocalice; essa è costituita dai residui glucidici delle glicoproteine e dei glicolipidi legati alla m., e spesso comprende anche glicoproteine e proteoglicani secreti e poi adsorbiti dalla superficie cellulare (fig. 2). Le catene oligosaccaridiche svolgono un ruolo di ancoraggio e orientamento delle proteine di m., ma potrebbero anche rivestire un ruolo fondamentale nella interazione e nel riconoscimento reciproco cellulare.
Costituzione chimica
Nella maggioranza delle m. biologiche delle cellule animali, i lipidi costituiscono circa il 50% in massa; sono molecole anfipatiche, che contengono cioè sia una regione idrofilica (la testa polare) sia una idrofobica (code idrocarburiche degli acidi grassi).
I tre principali lipidi di m. sono i fosfolipidi, i glicolipidi e il colesterolo. I fosfolipidi sono presenti in grandi quantità nelle m. eucariotiche nelle 4 forme principali: la fosfatidilcolina, la fosfatidiletanolammina, la fosfatidilserina e la sfingomielina. I primi tre sono fosfogliceridi mentre la sfingomielina presenta una molecola di sfingosina e non contiene glicerolo. I fosfolipidi sono caratterizzati da gruppi polari differenti per dimensione, forma e carica elettrica che, oltre a influenzare direttamente la funzionalità di alcune proteine di m., sono responsabili dell’asimmetria e della diversa distribuzione della carica elettrica del doppio strato lipidico. I gruppi glucidici dei glicolipidi esposti sulla superficie cellulare esterna hanno, come funzione principale, quella di recettori di superficie cellulare, sia verso particolari antigeni, sia verso molecole fisiologiche utilizzate nella normale comunicazione intercellulare. I glicolipidi si differenziano in glicolipidi neutri e in gangliosidi; i primi si trovano sia nelle cellule eucariotiche sia in quelle procariotiche, ma alcuni di essi si rinvengono solo in certi Mammiferi, per es., il galattocerebroside nella mielina. I gangliosidi contengono uno o più residui di acido sialico, che gli conferiscono una carica elettrica negativa; essi abbondano soprattutto nella m. plasmatica dei neuroni. Il colesterolo ha una funzione regolatrice della fluidità della m., esso aiuta a mantenere la stabilità meccanica del doppio strato lipidico. Doppi strati fosfolipidici possono essere prodotti in laboratorio, dando origine ai cosiddetti liposomi.
Le funzioni specifiche delle m. sono svolte in larga misura dalle proteine a esso associate. Le proteine sono generalmente di natura acida e povere di gruppi sulfidrilici; si distinguono in proteine transmembranali (o integrali) e proteine periferiche, in base al loro modo di associazione con il doppio foglietto lipidico. Alcune proteine possono attraversare il doppio strato sporgendo ai due lati di questo con le loro regioni idrofile, altre sono ancorate a proteine integrali differenti da legami covalenti, altre ancora sono appena appoggiate a uno dei due lati delle m. e, probabilmente, ne esistono anche alcune che penetrano nella m. solo con una porzione della propria catena polipeptidica. La gran parte delle informazioni sulle proteine di m. è stata ottenuta grazie a particolari metodiche di purificazione e di indagine quali l’elettroforesi su gel di poliacrilammide con sodiododecilsolfato o la microscopia elettronica a criodecapaggio. Le proteine di m., possono fungere da recettori (nelle m. plasmatiche) o da enzimi (nelle m. biologiche in genere). A volte, alcune proteine di m. possono svolgere la loro funzione solo se associate a particolari componenti lipidici. Ne è un esempio la cooperazione funzionale tra la guanilatociclasi della m. delle cellule miocardiche e il fosfolipide cardiolipina; questo è presente anche nella m. interna mitocondriale, dove svolgerebbe un ruolo fondamentale per assicurare la particolare permeabilità agli ioni da parte dei mitocondri, il che indica come la differente composizione lipidica e proteica di m. diverse garantisca una diversa funzione.
Esempi di specializzazione di m. molto particolari, sono quella del reticolo endoplasmatico liscio e rugoso (➔ reticolo), la m. nucleare (➔ nucleo) e la m. interna mitocondriale (➔ mitocondrio).
Meccanismi generali di trasporto
Si possono definire 3 gruppi fondamentali di trasporto.
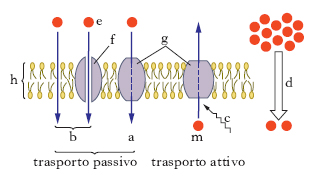
Diffusione semplice o passiva
Si verifica quando una sostanza penetra nella cellula per movimento spontaneo secondo un gradiente di concentrazione (dalla concentrazione più elevata a quella minore). Le molecole diffondono attraverso la fase lipidica del doppio strato come conseguenza della solubilità delle molecole nella fase lipidica stessa (per es., i narcotici) oppure attraverso canali idrofili transmembranali costituiti da proteine di m., dette proteine canale o pori acquosi (fig. 3). La diffusione passiva è il tipo di trasporto utilizzato per le piccole molecole prive di carica come acqua, urea, glicerolo, che si spostano attraverso la m. con velocità inversamente proporzionale alle loro dimensioni.
Quando le molecole diffusibili sono dotate di carica elettrica, come i piccoli ioni inorganici, il loro passaggio non dipenderà solo dal gradiente di concentrazione ma anche, nel caso delle m. plasmatica e mitocondriale, dal gradiente di potenziale elettrico totale. Nella m. plasmatica questi gradienti costituiscono il gradiente elettrochimico che genera il potenziale di membrana necessario per il funzionamento e la vita stessa delle cellule. Il potenziale di m., che consiste in una differenza di potenziale elettrico esistente tra i due lati della m. (−70 mV negli organismi eucariotici) è mantenuto dall’equilibrio di Gibbs-Donnan che determina la diversa distribuzione degli ioni Cl− e K+ tra i compartimenti extra- e intracellulare. Questo equilibrio si verifica nelle cellule grazie alle caratteristiche di permeabilità selettiva della m. plasmatica. All’interno delle cellule, sono presenti gli anioni proteici incapaci di permeare verso il liquido extracellulare: l’ambiente intracellulare ha un elevato numero di cariche negative appartenenti alle proteine. L’origine del potenziale di m. è determinato anche da processi di trasporto attivo di ioni.
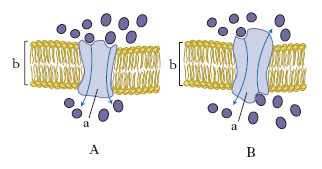
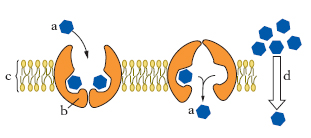
Diffusione mediata da un trasportatore (o proteina vettrice)
Si svolge lungo il gradiente di concentrazione della molecola da trasportare e dipende dalla presenza nella struttura della m. di proteine adibite al trasporto di sostanze attraverso il doppio strato lipidico. Ogni proteina di trasporto è specifica per una particolare molecola, o gruppo di molecole molto simili fra loro. Alcune proteine vettrici si limitano a trasferire un solo composto da un lato all’altro della m. (uniporto), mentre altre funzionano come sistemi di cotrasporto, nei quali il trasferimento di una data molecola dipende da quello, simultaneo o successivo, di una seconda molecola, sia nella medesima direzione (simporto o cotrasporto), sia nella direzione opposta (antiporto o controtrasporto; fig. 4). È probabile che quando la molecola da trasportare si lega a uno dei due siti di legame della proteina trasportatrice induca in essa una modificazione conformazionale che permette il trasferimento e il rilascio della molecola stessa sul lato opposto della m., consentendo il ritorno della proteina al primitivo stato conformazionale (fig. 5). Il processo è reversibile e la direzione del trasporto dipende solo dalla differenza tra le concentrazioni della molecola da trasportare ai due lati della membrana. Questo tipo di trasporto presenta caratteristiche cinetiche simili a quelle delle reazioni enzima-substrato (➔ enzima).
Trasporto attivo
Avviene contro un gradiente di concentrazione, richiede un dispendio di energia e proteine vettrici integrate nel doppio strato lipidico; se ne considerano tre tipi diversi: il trasporto attivo primario, il trasporto attivo secondario e il trasferimento di gruppo. Il trasporto attivo primario dipende da fonti primarie di energia, come l’idrolisi dell’ATP, il trasporto di elettroni e la luce. Uno degli esempi più importanti di questo tipo di trasporto nelle cellule animali è il passaggio (del tipo antiporto) degli ioni sodio (Na+) e potassio (K+) a opera dell’enzima Na+-K+-ATPasi. Nel trasporto attivo secondario l’energia è fornita da gradienti ionici e da differenze di potenziale che si stabiliscono tra le opposte facce delle membrane. Le molecole da trasportare vengono trascinate all’interno della cellula, contro il proprio gradiente di concentrazione, da ioni che irrompono nel compartimento intracellulare a causa del repentino abbassamento del loro gradiente di concentrazione. Un esempio è quello presente a livello delle cellule epiteliali dei villi intestinali e dei tubuli renali, dove il trasporto della sostanza è accompagnato al movimento di Na+, al di sotto di un gradiente di concentrazione. Zucchero e Na+ sono legati a recettori specifici e differenti del trasportatore e si muovono simultaneamente nell’interno della cellula. Il Na+ viene estruso per opera della pompa sodica e lo zucchero diffonde dalla cellula. In tal meccanismo l’energia è richiesta non per il trasporto per sé stesso, ma per il funzionamento della pompa sodica. Nel processo di trasferimento di gruppo il trasporto di molecole attraverso la m. comporta la trasformazione della molecola trasportata. Questo meccanismo è utilizzato da molte cellule batteriche, per es. per il passaggio del glucosio attraverso la loro m. plasmatica; in questo caso il meccanismo del trasporto avviene simultaneamente a un processo di fosforilazione del glucosio a glucosio-6-fosfato.
Trasporto degli amminoacidi
Numerosi tessuti accumulano amminoacidi contro un gradiente di concentrazione, per mezzo di un processo di trasporto attivo. Nelle m. plasmatiche di numerose cellule animali esistono almeno tre differenti trasportatori: uno per gli amminoacidi acidi, un secondo per quelli basici e un terzo per quelli neutri. È probabile che esistano interazioni fra trasporto degli zuccheri e trasporto di amminoacidi.
Trasporto degli ioni e potenziale di membrana
Il potenziale di m. è generato e mantenuto sia dall’equilibrio di Gibbs-Donnan che dalla pompa Na+-K+-ATPasi, un processo di trasporto attivo degli ioni Na+ verso l’esterno della m. accoppiato a quello degli ioni K+ verso l’interno, contro il gradiente elettrochimico. La pompa Na+-K+-ATPasi (➔ pompa), un sistema antiporto ATP-dipendente, introduce 2 ioni K+ nella cellula, estraendo 3 ioni Na+ per ogni molecola di ATP idrolizzata. Ogni molecola di Na+-K+-ATPasi, in condizioni ottimali, può idrolizzare un centinaio di molecole di ATP al secondo. La Na+-K+-ATPasi è una proteina strutturale di m. composta da 2 tipi di subunità, dotate di siti specifici di legame per gli ioni Na+ sul lato citosolico e per il K+ sul lato extracellulare della m. plasmatica (fig. 6). In questa proteina, sia il ciclo catalitico sia le attività di pompa ionica dipendono da reazioni di trasferimento di gruppi fosforici e da modificazioni conformazionali della molecola. L’importanza della Na+-K+-ATPasi è fondamentale nel mantenimento delle distribuzioni ioniche ai due lati della m.; ogni variazione delle concentrazioni degli ioni può sia provocare fenomeni di depolarizzazione della m., sia generare variazioni nella pressione osmotica della cellula. Questi fenomeni sono stati dimostrati mediante l’uso di inibitori specifici della Na+-K+-ATPasi, quali l’ouabaina che blocca contemporaneamente l’idrolisi dell’ATP e il trasferimento degli ioni. In alcuni sistemi di m. specializzati esiste la pompa Ca2+-ATPasi, il cui scopo è quello di trasportare attivamente gli ioni Ca2+ tra i due lati della membrana. Si trova per es., nella m. interna mitocondriale (➔ mitocondrio) e nella m. del reticolo sarcoplasmatico (➔ reticolo). Essa interviene, quindi, per reinserire gli ioni Ca2+ nel reticolo sarcoplasmatico determinando il termine della contrazione e ponendo la fibra muscolare in grado di contrarsi nuovamente in risposta a un altro impulso nervoso (➔ muscolo).
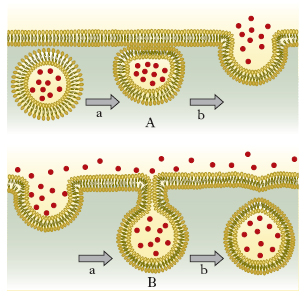
Trasporto di macromolecole e particelle attraverso la membrana plasmatica
La maggior parte delle cellule secerne e ingerisce macromolecole rispettivamente mediante i processi di esocitosi ed endocitosi. Nell’esocitosi il contenuto di particolari vescicole intracellulari si libera all’esterno della cellula quando esse si fondono con la m. plasmatica. Nell’endocitosi si ha il fenomeno inverso: alcune zone della m. plasmatica si invaginano e si staccano formando piccole vescicole di trasporto (vescicole di pinocitosi) o vescicole più grandi (vescicole o vacuoli di fagocitosi). Alla fine del processo la maggior parte delle vescicole di endocitosi dà origine a strutture intracellulari dette lisosomi (fig. 7). Sia l’endocitosi sia l’esocitosi sono risposte localizzate su alcune zone specifiche della m. plasmatica e del sottostante citoplasma.
Regolazione dei sistemi omeostatici
In conseguenza dei suoi incessanti scambi di materiali con l’ambiente, la cellula è in una condizione particolare di equilibrio, denominata equilibrio dinamico o stato dinamico stazionario. La m. plasmatica contribuisce a mantenere costanti le proprietà chimico-fisiche del mezzo interno, ed è pertanto il principale regolatore dell’omeostasi. Tutte le condizioni che determinano alterazioni degli scambi dei metaboliti attraverso la m. plasmatica possono determinare profonde alterazioni delle attività biochimiche della cellula, che portano a una serie di disturbi dell’omeostasi.
Membrana e parete delle cellule vegetali
Parete cellulare e tonoplasto
Nella cellula vegetale il citoplasma è, come in quella animale, delimitato dalla m. cellulare citoplasmatica (plasmalemma), bistratificata, semipermeabile, lipoproteica. Per essa vale, perciò, nelle linee generali, quanto è stato detto sopra. La cellula vegetale è, peraltro, più complessa di quella animale per la presenza, all’esterno del plasmalemma, di una seconda m. denominata correntemente parete cellulare, che racchiudendo la cellula come in un astuccio rigido e in parte estensibile, determina la forma della cellula e la protegge da danni meccanici, da lisi osmotica ecc. Questa parete, a differenza della m. citoplasmatica, che ha permeabilità selettiva, è permeabile all’acqua e a varie altre sostanze; essa manca nelle forme mobili di molte Alghe flagellate, nei Funghi Archimiceti e Mixomiceti e nelle forme apocitarie, cioè negli organismi dotati, almeno in certi stadi del loro ciclo biologico, di movimento ameboide.
La cellula vegetale contiene, nel citoplasma, ampi spazi, i vacuoli, delimitati da una esile m. monostratificata, il tonoplasto. Nella cellula matura un solo grande vacuolo occupa gran parte del volume cellulare. Il citoplasma è allora ridotto a un sottile strato periferico e talora attraversa il vacuolo mediante sottili filamenti (briglie citoplasmatiche). Nel vacuolo è contenuto il succo cellulare, costituito di acqua e di varie sostanze (prodotti di riserva, oli essenziali, acidi organici, glucosidi, alcaloidi, prodotti secondari terminali del metabolismo, sostanze di rifiuto precipitate in forma amorfa o cristallina).
Movimento degli ioni attraverso la membrana e la parete cellulare
La presenza della parete cellulare complica notevolmente lo studio delle relazioni ioniche, ma esistono nel mondo vegetale organismi con cellule grandi (Valonia, Bryopsis, Derbesia, Hydrodictyon, Halicistis, Chara, Nitella ecc.) che permettono tale tipo di ricerche. Nelle piante il trasporto di ioni ha grande importanza perché esse assorbono direttamente dal terreno soluzioni diluite di sali inorganici, che assimilano e incorporano in composti organici presenti nelle cellule, sintetizzando copiose riserve nutritive. Mentre la cellula animale può considerarsi come un unico compartimento, in quella vegetale vi sono 3 compartimenti: la parete, il plasmalemma, il vacuolo. Un aspetto rilevante dello spostamento di ioni consiste nel movimento netto (trasporto) attraverso le m. cellulari, fino a che si stabilisce per accumulo una concentrazione interna degli ioni stessi maggiore che all’esterno. Il potere di accumulo rappresenta un caso in cui il movimento di un soluto avviene in direzione opposta e a velocità diverse da come ci si aspetterebbe, e quindi con un aumento della energia potenziale del sistema. Il movimento di ioni richiede in tal caso consumo di energia cellulare e si ha un trasporto attivo, che presuppone una fonte energetica (ATP), e un sistema di trasportatori. Gli studi con cellule giganti di alghe hanno messo in evidenza, al livello del plasmalemma, differenze di potenziale che determinano il funzionamento di pompe ioniche (per es., per l’espulsione di Na+ e l’accumulo di K+ e Pi).
La parete cellulare, permeabile, rigida ma abbastanza estensibile, fa sì che la cellula non possa rigonfiarsi illimitatamente. Ne consegue una limitazione delle relazioni osmotiche, perché l’assunzione di acqua trova un limite insuperabile nel grado di estensibilità della parete cellulare. Quando la pressione osmotica eguaglia la pressione idrostatica esercitata dalla parete stessa tesa al massimo, cessa l’assunzione di acqua dall’esterno verso il vacuolo e la cellula è in stato di turgore. La deturgescenza, ripetutamente reversibile, e la capacità di resistere a forti oscillazioni della concentrazione del succo cellulare, sono proprietà peculiari della cellula vegetale e ne caratterizzano la adattabilità a condizioni estreme. In piante esposte a venti marini si possono raggiungere concentrazioni del succo cellulare corrispondenti a potenziali osmotici dell’ordine di centinaia (fino a 700-800) di bar. La perdita di acqua determina il progressivo svuotamento del vacuolo fino a raggiungere la plasmolisi, con contrazione e distacco del citoplasma dalla parete cellulare. La plasmolisi, anch’essa entro ampi limiti reversibili, produce l’afflosciamento delle parti erbacee della pianta e può raggiungere valori elevati, che sarebbero letali per le cellule animali.
Formazione della parete cellulare
La parete cellulare ha origine in corrispondenza della divisione cellulare. Essa si genera o come proliferazione verso l’interno della parete preesistente, la quale in forma di cercine avanza dalla periferia verso il centro e finisce per separare in due la massa citoplasmatica (per es., Spirogyra e altre Tallofite); oppure come derivato del fragmoplasto, che si forma in seno alle fibrille del fuso mitotico, in seguito alla divisione nucleare. I dittiosomi dell’apparato di Golgi secernono piccole vescicole contenenti pectina ed emicellulose. Queste vescicolette, venendo a contatto, costituiscono un primo setto divisorio, comune alle due cellule originatesi per mitosi, detto lamella mediana, costituito prevalentemente di pectine. Sulla lamella mediana si deposita poi la parete primaria, formata di un sottile strato di microfibrille di cellulosa, orientata e del diametro di 4-20 nm; esse sono inglobate in una sostanza non fibrillare, la matrice, di natura non cellulosica.
Modificazione della parete cellulare
Nelle cellule più mature, internamente alla parete primaria si può depositare un secondo strato, più o meno spesso, ben visibile al microscopio, con contenuto di cellulosa molto alto. La parete cellulare non si ispessisce in corrispondenza dei campi di punteggiature, i meati attraverso i quali passano i plasmodesmi, che connettono il citoplasma di cellule attigue. Nella parete secondaria si possono distinguere 3 o più strati a fibrille diversamente orientate. La parete cellulare può subire nei tessuti definitivi varie modificazioni, venendo incrostata e compenetrata da sostanze di natura non cellulosica: così si originano pareti lignificate, imbevute di lignine, nei tessuti meccanici, di sostegno e di conduzione ascendente; suberificate e cutinizzate per depositi di sostanze di natura grassa nei tessuti tegumentari ed epidermici. In certi casi, l’impregnazione è dovuta al deposito di sostanze inorganiche od organiche varie e si hanno allora pareti silicizzate (Diatomee, alcune Felci), calcarizzate (Alghe calcaree), tannificate, per presenza, nel legno di certi alberi, di tannini e di sostanze coloranti. La parete cellulare può, in qualche caso, depositarsi tanto abbondantemente da occupare quasi tutto il lume cellulare e ciò si verifica in alcuni tessuti meccanici (sclereidi, idioblasti meccanici, cellule pietrose), come quelli dei noccioli di alcuni frutti.
Nelle piante inferiori la parete è costituita di sostanze che presentano variazioni da gruppo a gruppo: peptidoglicani, chitina e derivati insieme a cellulosa. Sono presenti inoltre polimeri di matrice variabili da caso a caso, quali agar, alginati, carrageen, fucoidina.
Chimica
Le m. utilizzate per le operazioni di separazione sono costituite da materiali con elevato rapporto superficie/spessore, che hanno la proprietà caratteristica di rallentare o impedire il passaggio di alcune specie chimiche e di consentire il passaggio di altre (membrane permselettive). Le m. possono essere liquide o solide, naturali o sintetiche, organiche (essenzialmente di tipo polimerico) o inorganiche (metalliche, di materiale ceramico, di vetro).
Le operazioni di separazione con m., sia in fase liquida sia in fase gassosa, hanno assunto importanza sempre maggiore in molti settori industriali. L’interesse per le operazioni a m. è legato al fatto che esse avvengono a basse temperature, non coinvolgono cambiamenti di fase né l’impiego di additivi chimici, sono caratterizzate da elevata flessibilità, da bassi consumi energetici e da alto grado di automatizzazione, sono modulari e richiedono modesti spazi d’ingombro. Inoltre, sono semplici nella loro conduzione e nella loro impiantistica. Oltre alle operazioni a m. già da tempo affermate su scala industriale (microfiltrazione, ultrafiltrazione, osmosi inversa, dialisi, elettrodialisi, separazione di gas), prospettive di sviluppo sempre più promettenti stanno avendo la distillazione a m., la nanofiltrazione, la per vaporazione, la separazione con m. liquide supportate. Quest’ultima utilizza un sottile supporto polimerico contenente nei suoi pori un agente estraente liquido, detto carrier, costituito in genere da un composto complessante; tale supporto tiene distinta la soluzione acquosa, contenente la specie chimica da separare (soluzione di feed), dalla fase acquosa in cui la specie medesima viene ricuperata (soluzione di strip).
Sono stati proposti vari tipi di trasporto in m. liquide supportate. Nel trasporto semplice con carrier il complessante C reagisce all’interfaccia membrana-soluzione di feed con la specie A (secondo la reazione reversibile: A+C⇄AC) e, dopo averla trasportata sotto forma di complesso AC attraverso la m., la rilascia all’interfaccia membrana-soluzione di strip; riacquistata la forma iniziale, il carrier diffonde nuovamente attraverso il supporto polimerico rinnovando il ciclo di trasporto. Il tipo più interessante di operazione attuabile con una m. liquida supportata è il trasporto accoppiato, che può essere esemplificato facendo riferimento alla reazione Cu2++2HR⇄R2Cu+2H+, dove HR rappresenta l’agente complessante in forma idrogenata; tale reazione consente di estrarre il rame da un’acqua di scarico (proveniente, per es., da una lavorazione galvanica). La reazione è spostata verso destra in corrispondenza dell’interfaccia membrana-soluzione di feed, che è mantenuta a un pH più elevato rispetto all’interfaccia membrana-soluzione di strip ove, invece, la reazione è spostata verso sinistra. Il gradiente di pH esistente fra le due facce della m. consente, a differenza di quanto accade non solo nella diffusione passiva ma anche nel trasporto semplice con carrier, di concentrare una specie chimica (il rame nel caso in esame) vincendo un controgradiente di concentrazione.
È possibile realizzare m. stabili ai solventi organici e capaci di operare ad alte temperature in ambienti aggressivi. Ciò ha creato le premesse per l’impiego, da parte dell’industria chimica, di reattori a membrana, in cui vengono accoppiate operazioni di separazione e reazioni chimiche o biologiche, sia in processi a bassa temperatura (utilizzando catalizzatori enzimatici) sia in processi di ossidazione, idrogenazione, deidrogenazione ecc., fatti avvenire a temperature più elevate.
Fisica
Nella fisica delle particelle elementari, possibili strutture immerse in varietà multidimensionali postulate da alcuni modelli di natura speculativa proposti nel corso degli anni 1990; tali modelli modificano i concetti di base della teoria dei campi per ottenere una teoria unificante che comprenda la gravità, la fenomenologia osservata, descritta dal Modello Standard delle particelle elementari, nonché le proprietà più interessanti della supersimmetria e del modello a stringhe.
Tecnica
Nella scienza delle costruzioni, ogni sistema materiale a due dimensioni (lastre sottili, piane o curve) per il quale sia possibile trascurare la rigidezza a flessione e a torsione. Nelle m., per la loro stessa struttura, il regime degli sforzi interni si riduce ai soli sforzi contenuti nel piano tangente alla superficie media della struttura; questi sforzi di membrana comportano tensioni uniformemente ripartite nello spessore della lastra per l’assenza in ogni punto del momento flettente e del momento torcente.
La m. può concepirsi come l’insieme di due sistemi di fili o archi puri disposti secondo le linee isostatiche, ognuno dei quali sostiene una parte delle forze esterne: precisamente, una parte per la quale esso risulta funicolare. Poiché in ogni punto passano due elementi lineari – fili o archi – incrociantisi ortogonalmente, la forza esterna può suddividersi in ogni punto fra i due, in maniera che tutti gli elementi risultino funicolari dei carichi da essi portati. Gli elementi di un sistema esercitano cioè sugli elementi dell’altro una funzione correttiva delle forze esterne, alterandole con sistemi di forze (di cui essi sono funicolari) tali che le forze risultanti agenti sugli elementi dell’altro sistema ammettano questi come loro curve funicolari. Quanto si è detto diviene particolarmente semplice nel caso che la m. sia una superficie di rivoluzione e sia soggetta a forze esterne simmetriche rispetto al suo asse. In questo caso le linee isostatiche sono i paralleli e i meridiani; i primi sono circonferenze, i secondi possono avere una forma qualsiasi ma comune a tutti. Le strisce disposte lungo i paralleli possono funzionare come funicolari di forze radiali uniformi ma di intensità qualsiasi, e ciascuna di esse assorbe perciò una componente radiale delle forze esterne tale da lasciare alle strisce disposte secondo i meridiani una distribuzione di forze da esse sostenibile come archi puri (➔ cupola). La formulazione matematica delle condizioni di equilibrio statico di una m., nel caso più generale che la sua superficie media sia di forma qualsiasi, è piuttosto complessa. Le equazioni di equilibrio risultano tuttavia molto semplici in alcuni casi particolari notevoli.
Le strutture a membrana sono sempre più attuali in molti campi della tecnica. Nel campo delle costruzioni civili si possono citare le volte sottili in cemento armato e i serbatoi a involucro sottile sia in cemento armato sia metallici. Già da tempo inoltre strutture a m. trovano applicazioni nelle costruzioni navali e aeronautiche (strutture a guscio).
Zoologia
Membrana alare
La ripiegatura cutanea che nei Pipistrelli, negli Pterodattili e nei Ranforincoidei (Rettili fossili) è sostenuta dallo scheletro dell’arto anteriore e costituisce la superficie portante dell’ala. Negli Insetti la m. alare è costituita da una m. chitinosa sostenuta dalle nervature.