
mitocondrio
Organello intracellulare di forma granulare o filamentosa, di dimensioni comprese tra 0,5 e 3 μm, presente in tutte le cellule eucariotiche con metabolismo aerobio. La sua attività è respiratoria e avviene solo in presenza di ossigeno. All’interno dei m. sono contenuti gli enzimi dei principali cicli metabolici deputati alla produzione di energia, quali il ciclo dell’acido citrico, della β-ossidazione degli acidi grassi e della fosforilazione ossidativa, processi che producono la maggior parte dell’energia sotto forma di ATP utilizzata dalla cellula.
Struttura
Il numero di m. presente in ogni cellula varia nei diversi tipi cellulari, secondo le dimensioni della cellula e le richieste energetiche. In condizioni metaboliche diverse i m. modificano la loro forma, diventando più o meno allungati fino ad assumere una forma quasi sferica; ciò è evidente quando il m. passa dallo stato di riposo a quello attivo. In alcuni tipi di cellule, i m. sono localizzati dove avvengono processi metabolici che richiedono ATP. La struttura del m. è identica nelle cellule animali e vegetali, così come la sua funzione.
Il m. è delimitato da due membrane concentriche, con proprietà e funzioni biologiche distinte, separate da uno spazio intermembrana (o intermembranoso), di circa 50-150 Å; il doppio sistema membranoso delimita il compartimento interno che contiene una fase il cui stato fisico è simile a un gel, detta matrice mitocondriale.
La membrana mitocondriale esterna (spessore 50-60 Å) consiste per circa il 50% di fosfolipidi e per il restante 50% di proteine; contiene una concentrazione considerevole di colesterolo. In essa è presente la proteina porina che, formando canali nel doppio strato lipidico, permette a sostanze di peso molecolare inferiore a 10.000 di diffondere liberamente nello spazio intermembrana.
La membrana mitocondriale interna, molto più impermeabile, si ripiega in un grande numero di pliche, dette creste, che ne aumentano notevolmente la superficie; è costituita per il 20% di lipidi e per l’80% di proteine; i lipidi sono essenzialmente fosfogliceridi contenenti colina ed etanolammina, oltre a fosfolipidi contenenti serina e inositolo. La membrana mitocondriale interna del muscolo cardiaco è ricca di fosfatidilcolina, fosfatidiletanolammina e cardiolipina.
Funzioni
Alla membrana mitocondriale esterna sono associati numerosi enzimi che partecipano a diverse reazioni della biosintesi degli acidi grassi e dei fosfolipidi e sono responsabili di alcune reazioni di ossidazione; un enzima caratteristico di questa membrana è la monoamminossidasi .
Anche lo spazio intermembrana contiene molti enzimi, come l’adenilato chinasi, che utilizzano l’ATP prodotto dai sistemi enzimatici presenti nella matrice e sulla membrana mitocondriale interna per fosforilare altri nucleotidi necessari al metabolismo cellulare.
La membrana mitocondriale interna contiene numerose proteine che possono essere riunite in tre gruppi principali: gli enzimi della catena respiratoria, coinvolti nella fosforilazione ossidativa; le proteine che costituiscono il complesso enzimatico detto ATP-sintetasi; le specifiche proteine vettrici (dette anche sistemi navetta) che regolano il passaggio di vari metaboliti (fosfato inorganico, ADP, ATP, acidi di- e tricarbossilici, Ca2+ e alcuni amminoacidi) verso e dalla matrice mitocondriale. Il complesso dell’ATP-sintetasi forma piccole protrusioni, chiamate corpi elementari o particelle della membrana, ed è costituito da due componenti principali denominati F0 e F1; F1 sporge verso la matrice ed è attaccato tramite un peduncolo a F0 che è inserito nello spessore della membrana interna, attraversandola. Alla membrana mitocondriale interna si trovano associati anche due enzimi del ciclo dell’acido citrico, l’aconitasi e la succinatodeidrogenasi.
La matrice mitocondriale contiene una miscela concentrata di enzimi diversi, tra cui gli enzimi del ciclo dell’acido citrico, della β-ossidazione degli acidi grassi, il sistema della piruvatodeidrogenasi; contiene inoltre: ATP, ADP, AMP, fosfato inorganico (Pi), NAD, NADP, il coenzima A, vari ioni come K+, Mg2+ e Ca2+. Qui ha luogo il ciclo dell’acido citrico, o degli acidi tricarbossilici, i cui principali prodotti finali sono CO2, NADH e FADH2; questi ultimi due costituiscono l’elemento di raccordo tra il ciclo dell’acido citrico e la fosforilazione ossidativa; essi cedono la propria carica elettronica agli enzimi della catena respiratoria, al termine della quale si ha la formazione di ATP e acqua.
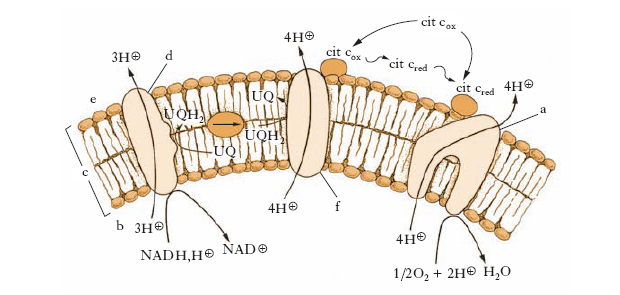

Gli elettroni provenienti dall’idrogeno di NADH e FADH2, formatisi nel ciclo dell’acido citrico, vengono trasportati lungo la catena del trasporto elettronico nella membrana mitocondriale interna (la catena respiratoria) e l’energia che si libera al loro passaggio serve a pompare protoni, attraverso la membrana mitocondriale interna, dalla matrice allo spazio intermembrana. Si instaura così un gradiente elettrochimico protonico tra i due lati della membrana mitocondriale interna e il flusso inverso di protoni lungo il gradiente serve a pilotare l’attività dell’enzima ATP-sintetasi, che catalizza la trasformazione ADP + Pi → ATP e conclude il processo della fosforilazione ossidativa. I gruppi che si susseguono lungo la catena respiratoria presentano affinità progressivamente crescente per gli elettroni: gli elettroni si muovono in cascata dal NADH a livelli energetici via via più bassi, andando da un complesso enzimatico al successivo fino a raggiungere l’ossigeno molecolare, che ha nei loro confronti la massima affinità. Gli enzimi della catena respiratoria sono in grado di trasportare un solo elettrone alla volta; dato che ogni NADH cede due elettroni e una molecola di ossigeno, per trasformarsi in acqua, deve riceverne il doppio, esistono lungo questa catena enzimatica dei punti di raccolta e dei punti di dispersione degli elettroni. Gran parte dell’energia liberata durante il trasporto degli elettroni è utilizzata per sospingere i protoni idrogeno (H+) dalla matrice verso lo spazio intermembrana e poi al di fuori del m. (fig. 1). Si genera così un gradiente di pH attraverso la membrana mitocondriale interna caratterizzata da una concentrazione di H+ nella matrice mitocondriale molto inferiore rispetto a quella presente nel resto della cellula. Il gradiente elettrochimico protonico prodotto ha l’effetto di risospingere i protoni all’interno del m. attraverso il complesso proteico transmembranale dell’ATP-sintetasi che utilizza l’energia del flusso protonico per sintetizzare ATP, a partire da ADP e Pi, all’interno della matrice mitocondriale (fig. 2). Grazie al gradiente elettrochimico protonico e all’azione di specifiche proteine vettrici, sia gli enzimi della matrice sia la stessa ATP-sintetasi ricevono i propri substrati in concentrazioni elevate mediante trasporto attivo.
Altra funzione vitale del gradiente elettrochimico della membrana mitocondriale interna consiste nel pompare ioni Ca2+ con un meccanismo attivo dal citoplasma cellulare alla matrice mitocondriale. Il meccanismo di accumulo del calcio nella matrice mitocondriale è di notevole importanza nella regolazione della concentrazione del Ca2+ nel citoplasma cellulare, in genere molto bassa.
Esistono molti altri sistemi trasportatori (➔ navetta) attraverso la membrana mitocondriale interna, specifici per altre sostanze necessarie ai vari cicli metabolici del mitocondrio: per es., il piruvato per il ciclo dell’acido citrico. Il sistema navetta per l’acido grasso, ha l’effetto di tenere separati i pool extramitocondriale e intramitocondriale degli acidi grassi e del coenzima A (CoA). Gli acidi grassi sono attivati sulla membrana mitocondriale esterna, mentre la loro ossidazione ha luogo nella matrice mitocondriale; dato che le molecole di acil-CoA a lunga catena carboniosa non possono attraversare facilmente la membrana mitocondriale interna, vengono trasportate dalla carnitina.
Riguardo l’origine e l’evoluzione dei m., esistono prove convincenti secondo le quali la presente struttura mitocondriale deriva da una simbiosi tra una cellula eucariotica, in grado di aver ben sviluppato il processo glicolitico anaerobico, e una cellula procariotica (per es., un batterio) che aveva sviluppato un sistema di una catena trasportatrice di elettroni transmembrana.
Genetica
Nella matrice mitocondriale sono contenute molte copie identiche del genoma mitocondriale, costituito essenzialmente da DNA; sono presenti anche particolari ribosomi, molecole di tRNA ed enzimi necessari per l’espressione dei geni propri del m., rendendo così il m. in grado di duplicarsi autonomamente anche se la sua replicazione è sempre collegata e regolata dal genoma della cellula a cui appartiene.

DNA mitocondriale. - Il DNA mitocondriale (DNAmt) è una molecola di DNA circolare eccetto che in alcuni Protisti, nei quali è lineare. Le sue dimensioni variano a seconda delle specie (da circa 15.000 paia di basi nei Mammiferi a circa 2 milioni di paia di basi nelle piante) e all’interno della stessa specie. I DNAmt meglio studiati sono quelli del lievito e dell’uomo. Entrambi sono stati sequenziati: il DNAmt del lievito è costituito da circa 70.000 coppie di basi, ma solo un terzo codifica proteine, mentre quello dell’uomo ne contiene 16.569. Nel DNAmt si possono distinguere i due filamenti dalla differente composizione delle basi: un filamento pesante (H, heavy) e un filamento leggero (L, light). Il DNAmt umano codifica 22 tRNA, 2 rRNA e 13 polipeptidi (fig. 3).
La presenza di geni per l’RNA transfer e ribosomale dimostra che i m. possiedono un loro sistema di traduzione anche se le relative componenti proteiche sono codificate da geni nucleari. La notevole simbiosi molecolare fra DNA mitocondriale e nucleare è anche aumentata dal fatto che proteine che agiscono in concerto fra loro, quali quelle della catena respiratoria, vengono codificate in parte dal DNAmt e in parte dal DNA nucleare.
Espressione dei geni dei mitocondri. - Gli schemi di trascrizione dei geni mitocondriali sono piuttosto variabili nelle diverse cellule eucariotiche studiate (per il lievito, ➔ maturasi). Nell’uomo sono trascritti entrambi i filamenti di DNAmt. Le sequenze del genoma mitocondriale umano non contengono introni; molti geni risultano separati da geni che codificano gli RNAt e non da spaziatori non codificanti come nel lievito. Solo dopo la trascrizione gli RNAt sono separati dalle sequenze codificanti proteine. Nel DNAmt è presente l’ansa D, una regione contenente un terzo filamento costituito da un frammento di DNA di circa 700 paia di basi che si lega al filamento L e allontana da questo il tratto corrispondente del filamento H. Nell’ansa D si trovano due promotori, uno per il filamento H e l’altro per il filamento L; sono trascritte due molecole di RNA pari alla lunghezza dell’intero genoma mitocondriale. I due trascritti sono tagliati da nucleasi durante la loro sintesi: dal filamento H si formano i diversi tipi di RNA, mentre la porzione L dell’RNA viene degradata quasi completamente. Gli RNA mitocondriali presentano alcune differenze rispetto a quelli nucleari: all’estremità 5′ della molecola non hanno né il cappuccio di 7-metil guanosina né sequenze non codificanti, ma iniziano direttamente con il codone di inizio AUG.
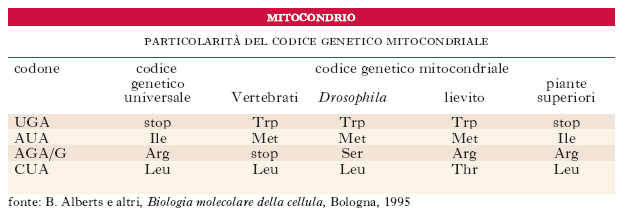
Il codice genetico dei mitocondri. - Le proteine codificate dal DNAmt sono state sequenziate e confrontate con le sequenze geniche: il codice genetico mitocondriale, tranne nelle piante, differisce in piccola parte da quello universale (v. tab.), probabilmente per fenomeni di deriva genetica casuale.
Filogenesi dei mitocondri. - È stato dimostrato che il ritmo di sostituzione di sequenze nucleotidiche nell’evoluzione dei genomi mitocondriali è 10 volte maggiore rispetto a quello dei genomi nucleari. Il ritmo relativamente alto di mutazione dei geni dei m. è pertanto utile per stimare le date di eventi evolutivi relativamente recenti. Inoltre, il genoma mitocondriale è ereditato per via materna; la maggior parte del citoplasma dello zigote è infatti derivato dalla cellula uovo. Supponendo che la velocità di variazione delle sequenze sia costante e dovuta solo a mutazioni, il numero delle differenze riscontrate fra i vari gruppi e fra gli individui dello stesso gruppo geografico può essere utilizzato per dedurre le linee evolutive e il periodo nel quale sia vissuta un’originaria ‘Eva mitocondriale’, cioè l’antenata comune dalla quale tutti gli esseri umani moderni avrebbero ereditato il loro DNAmt.