
regolazione
Intervento su fatti, fenomeni, processi naturali (anche fisici o fisiologici) o meccanici, per modificarne o disciplinarne le condizioni o lo svolgimento, in base a determinate esigenze.
Biologia
Gli organismi viventi costituiscono una complessa macchina biochimica in cui le varie reazioni, in ciascun particolare stadio del ciclo di una cellula o dello sviluppo di un organismo, devono svolgersi in maniera ordinata ed essere compatibili fra loro. Inoltre, un organismo è in grado di reagire ai segnali dell’ambiente attivando e disattivando specifiche reazioni; variazioni di concentrazione di protoni (pH) all’interno della cellula o variazioni di temperatura possono cambiare l’attività di un enzima di vari ordini di grandezza. In termini di attività metabolica cellulare, questo significa che la quantità e l’attività degli enzimi all’interno della cellula devono essere accuratamente regolate. In alcuni casi il bersaglio del meccanismo regolativo è l’attività di un enzima o di un ormone e non la sua produzione (r. enzimatica o metabolica); in altri casi, più numerosi, i processi di r. agiscono a livello della sintesi degli enzimi attivando o reprimendo l’espressione dei geni che li codificano (r. genica).
R. enzimatica o metabolica
La r. delle vie metaboliche è operata da una serie di fattori che si distinguono in base al loro meccanismo d’azione, alla loro localizzazione intracellulare e all’ordine temporale della loro azione. La r. metabolica avviene a due diversi livelli: il primo è dato dagli enzimi regolatori allosterici, la cui azione si manifesta con fenomeni di inibizione, di allosterismo e di retroinibizione (➔ regolatore); il secondo, soprattutto negli organismi superiori, è esercitato dalla r. ormonale (➔ ormone), efficace sia a livello cellulare sia subcellulare.
R. genica
Le conoscenze sulla r. dei geni che si sono accumulate nel corso degli anni 1990 riguardano sia la struttura e la funzione della cromatina (➔ epigenesi) sia, soprattutto, la funzione degli elementi regolativi della trascrizione. Risulta ormai chiaro che, in tutte le cellule, la fase più importante della r. dell’espressione genica riguarda l’inizio della trascrizione, il processo nel quale l’enzima RNA-polimerasi si lega a un sito specifico sul DNA e inizia a trascrivere una molecola di RNA utilizzando il DNA come stampo. La r. della trascrizione è un processo straordinariamente complesso, basato sull’interazione di proteine dette fattori che agiscono in trans, o fattori transattivi, con specifiche sequenze di DNA dette elementi che agiscono in cis. Le interazioni delle proteine fra di loro e con il DNA permettono la r. della trascrizione dei geni adatti nel momento adatto e quindi la produzione della giusta quantità di una determinata proteina.
Elementi che agiscono in cis
Sono brevi sequenze di nucleotidi che possono essere adiacenti a un gene, nel suo interno o distanti da esso migliaia di paia di basi. L’elemento in cis, comune a tutti i geni, è il promotore, sito nel quale il complesso di proteine necessarie alla trascrizione comprendente l’RNA-polimerasi viene legato al DNA. Nel promotore sono presenti due sequenze con una localizzazione più o meno fissa: l’elemento iniziatore (Inr), che determina il sito di inizio della trascrizione, e una regione ricca di adenine-timine, localizzata 25-30 paia di basi a monte di Inr.
Altri elementi di r. che agiscono in cis sono gli elementi promotori prossimali e quelli distali. Gli elementi promotori prossimali sono caratterizzati da sequenze, lunghe 50-100 paia di basi, localizzate a monte del sito di partenza e non sono elementi regolativi di tutti i geni. Un primo tipo di sequenze consiste in citosina-adenina-adenina-timina (CAAT box) e guanina-citosina (GC box), che fungono da siti di legame per proteine denominate fattori di trascrizione a monte. Questo tipo di sequenze non è presente nei geni housekeeping. Un secondo tipo di elemento promotore prossimale è costituito dall’elemento di risposta (RE). Gli elementi promotori distali possono presentarsi a una distanza di migliaia di paia di basi dal sito di inizio della trascrizione. Tali siti distali sono in grado di intensificare o annullare la trascrizione e sono chiamati rispettivamente intensificatori (enhancer) o silenziatori (silencer): esercitano la loro funzione a grande distanza e possono localizzarsi a monte o a valle del gene da regolare.
Fattori di trascrizione
Esperimenti di biologia molecolare hanno portato all’identificazione di una serie di fattori di trascrizione che agiscono in trans, necessari alle RNA-polimerasi per il corretto inizio della trascrizione, che si legano a specifici siti sul DNA. Questi fattori possono essere classificati in 5 gruppi. Al primo appartengono i fattori generali di trascrizione (GTF, General Transcription Factor), che sono assemblati a livello del sito promotore e richiamano sul sito la RNA-polimerasi II affinché possa iniziare la trascrizione dell’RNA messaggero. Vi sono poi fattori di trascrizione comuni, detti anche fattori di trascrizione a monte, che sono prossimali a molti svariati promotori quali le sequenze CAAT e GC, le proteine attivatrici, che si legano agli intensificatori, e i fattori proteici repressori, che si legano agli elementi silenziatori e inibiscono l’avvio della trascrizione. La quinta categoria è costituita dai fattori di trascrizione inducibili, proteine non sempre attive in una cellula che, in seguito a stimoli esterni, potenziano il loro legame con le sequenze RE, dando luogo ad aumentata trascrizione di particolari geni. Per la maggior parte, le proteine che fungono da fattori di trascrizione hanno in comune due proprietà fondamentali: si legano sia a sequenze specifiche di DNA sia ad altri fattori di trascrizione. Molte proteine che attivano i geni presentano, infatti, un motivo strutturale (il motivo a dita di zinco, il motivo a cerniera di leucine, il motivo elica-ansa-elica) che corrisponde alle sequenze specifiche del DNA, con interazioni così perfette da far supporre che le unità strutturali di base degli acidi nucleici e delle proteine si siano evolute insieme. Oltre a questi motivi le proteine presentano un dominio, detto di transattivazione, costituito da una sequenza amminoacidica ricca di serina, treonina e glutammina, che interagisce con gli altri fattori proteici che formano il complesso trascrizionale di base.
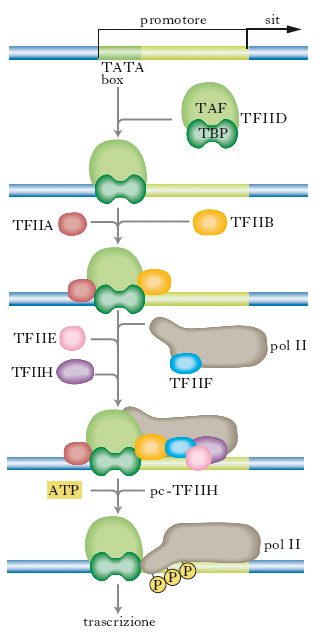
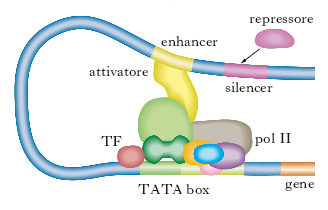
La fig. 1 mostra un esempio di come alcuni GTF si assemblano alla RNA-polimerasi II per dare inizio alla trascrizione. Il nome dei diversi fattori è formato dalla sigla TF (transcription factor), dal numero romano II (controllo della RNA-polimerasi II) e dalle lettere maiuscole (A, B ecc.) che definiscono i differenti fattori. Come mostrato nella fig. 1, inizialmente TFIID riconosce la sequenza TATA del promotore mediante una sua subunità proteica (TBP); in successione si assemblano sul promotore, prima la polimerasi II insieme al fattore di trascrizione F e poi i fattori di trascrizione E e H. La polimerasi II è fosforilata dalla subunità che funge da chinasi della TFIIH e la fosforilazione determina il suo distacco dal complesso dei fattori e l’inizio della trascrizione. La fig. 2 fornisce una plausibile spiegazione di come una proteina attivatrice possa regolare un gene lontano: in questo caso si deve ipotizzare che il DNA fra la sequenza enhancer e il promotore si ripieghi, formando un’ansa, per permettere alle proteine legate all’enhancer di interagire direttamente con uno dei TF o con l’enzima RNA-polimerasi.
Dato che i geni devono rispondere nella maniera più flessibile ai segnali che vengono dall’esterno, essi devono essere attivati ma anche rapidamente disattivati. Questo può avvenire o attaccando o togliendo gruppi fosforici ai TF, oppure mediante la funzione dei silenziatori, gli appositi fattori repressori a influenza negativa che si legano, come gli intensificatori, a sequenze regolative molto lontane dal gene.
Sono stati isolati e studiati in vitro molti diversi fattori di trascrizione, ma non sono ancora noti gli esatti processi con i quali tali fattori regolano l’avvio della trascrizione. Dato che la crescita, lo sviluppo e l’omeostasi sono controllati in molti casi da ormoni o da fattori di crescita, sono noti processi di r. della trascrizione mediati da recettori cellulari. In questo contesto sono importanti alcuni fattori di trascrizione, denominati STAT, coinvolti sia nell’attivazione della trascrizione, sia nei processi di trasduzione del segnale (➔ trasduzione).
R. della morfogenesi
I geni coinvolti nella r. dei processi di sviluppo in quasi tutte le specie eucariotiche presentano una caratteristica sequenza di 180 bp all’interno della regione codificante, definita omeobox perché scoperta per la prima volta nei geni omeotici di Drosophila melanogaster (➔ omeotici, geni). Dai geni omeotici dipende il differenziamento dei vari segmenti che danno origine alle regioni cefaliche, toraciche e addominali. Nell’embrione in via di sviluppo essi si esprimono dopo i geni materni, zigotici e di segmentazione, ma prima dei geni che determinano il differenziamento dei singoli tipi cellulari all’interno di ciascun segmento. Alcuni geni dotati di omeobox codificano proteine che regolano la trascrizione, le quali influenzano a loro volta l’espressione sia di altri geni con omeobox sia di geni per proteine strutturali non regolative. Omeobox sono state scoperte in geni che regolano lo sviluppo in numerosi organismi quali l’uomo, i topi, le rane, i vermi. Così l’embrione, nello stadio iniziale, in virtù dei geni omeotici, risulta suddiviso in una regione anteriore e una posteriore, già dotate di un potenziale sviluppo differente, prima ancora che avvenga la formazione dei singoli organi. L’omeobox, in qualsiasi gene venga riscontrata, codifica una sequenza di 60 amminoacidi con il motivo elica-ansa-elica sopradescritto. La comprensione dei meccanismi che regolano l’attività degli enzimi o l’espressione dei geni durante la formazione dell’embrione non è sufficiente per spiegare il complesso problema dello sviluppo.
Un punto fondamentale è la comprensione dei meccanismi che determinano come in certi membri di una popolazione cellulare, apparentemente omogenea, un certo gruppo di geni in una cellula e un altro in cellule apparentemente identiche diventi attivo o inattivo, indirizzando conseguentemente i due gruppi in differenti programmi di sviluppo. I processi sono tutti basati su differenze preesistenti, sia chimiche sia topografiche (➔ sviluppo). Un citoplasma non uniforme può essere identificato già nell’uovo non ancora fecondato. Durante la divisione cellulare, il citoplasma, non omogeneo, può essere distribuito non uniformemente e conseguentemente creare un differente ambiente citoplasmatico per i nuclei delle cellule figlie. Sostanze regolative citoplasmatiche potranno influenzare in maniera differenziale l’espressione dei geni, che a loro volta cambieranno la composizione del citoplasma, e così via. In un tale sistema piccole differenze biochimiche iniziali possono essere amplificate in grandi differenze morfologiche e fisiologiche.
R. del fago
Il batteriofago λ presenta due cicli vitali alternativi: la crescita litica e quella lisogenica. Nel ciclo litico tutti i geni trovano espressione secondo una sequenza temporale, che culmina con la produzione di circa 100 nuovi fagi e la lisi della cellula batterica infettata. In alternativa, il virus infettante può intraprendere uno sviluppo lisogenico, nel quale il suo DNA è inserito nel DNA della cellula ospite a livello di un sito specifico del cromosoma. La forma integrata viene chiamata profago. Nelle cellule lisogeniche i meccanismi della cellula stessa replicano il DNA del profago per molte generazioni, come se si trattasse di un normale segmento del genoma, e tutti i geni del fago tranne uno sono disattivati.
I geni del fago codificano proteine con una precisa sequenza temporale e vengono chiamati precoci e tardivi. I geni precoci codificano proteine necessarie alla duplicazione del DNA e all’integrazione del DNA fagico all’interno del cromosoma batterico, oltre che proteine regolatrici che controllano l’espressione dei geni tardivi. I geni tardivi codificano proteine necessarie alla formazione di nuovi capsidi, all’impacchettamento del DNA fagico appena duplicato in nuovi fagi e alla lisi della cellula ospite. Tutti i geni tendono a essere raggruppati in operoni. La scelta fra il ciclo lisogenico e quello litico è dovuta alla r. dell’espressione di molti geni. Se viene seguita la via lisogenica, i geni responsabili per la riproduzione fagica e la lisi sono inattivati da un repressore prodotto dal gene virale (cI). Se la cellula intraprende il ciclo litico, il gene cI viene represso dalla proteina Cro codificata da un altro gene fagico. Le cellule lisogeniche possono intraprendere lo sviluppo litico spontaneamente, con bassa frequenza, o in seguito a esposizioni a vari agenti fisici o chimici. Per es., le radiazioni ultraviolette inducono il ciclo litico in quanto causano un cambiamento nella funzione di una proteina batterica, la proteina RecA, normalmente coinvolta in eventi di ricombinazione. Quando il DNA viene danneggiato, essa assume una funzione proteasica e taglia il repressore codificato da cI. La RNA polimerasi in tal modo può liberamente trascrivere Cro. Le prime proteine Cro sintetizzate bloccano l’attacco della RNA polimerasi al promotore di cI, impedendo l’ulteriore sintesi di repressore. A questo punto la svolta è stata effettuata e, a tutto ciò, segue il ciclo litico. La via litica e quella lisogena si escludono a vicenda e l’ingresso nella seconda via può avvenire solo in un momento precoce e molto specifico dell’infezione. Una volta che il fago ha avviato un’infezione litica produttiva, il batterio non potrà più sopravvivere, anche nel caso in cui l’infezione abortisca.
Per quanto riguarda la r. naturale della fertilità ➔ contraccezione.
Tecnica
Operazione che nel funzionamento di un sistema mantiene il valore di determinate grandezze costante, o variabile entro limiti stabiliti, oppure, nell’ambito dei servosistemi ne garantisce uno stabilito andamento in funzione di grandezze variabili di entrata. In genere, la r. si compie mediante strumenti di misura che rilevano il valore delle grandezze da controllare e, ove si superino i livelli di tolleranza, ne danno avviso agli addetti affinché intervengano (r. manuale), o comandano direttamente i dispositivi automatici (r. automatica o autoregolazione). Si ha r. diretta o indiretta se si agisce direttamente sull’organo che modifica il funzionamento nel senso dovuto o se si agisce con servomotori, ad aria compressa (r. pneumatica), a liquido (r. idraulica), elettrici (r. elettrica). La r. è di norma continua, altrimenti, se interviene quando il valore della grandezza controllata supera massimo o minimo stabiliti (anche a fini di sicurezza, arrestando temporaneamente la macchina o l’impianto), si dice di massimo o di minimo.
La stabilità della r. rappresenta la sua capacità di riportare la grandezza al valore stabilito, con oscillazioni smorzate a sufficienza o con rapido moto aperiodico, l’insensibilità è la variazione del valore della grandezza da regolare entro cui il regolatore non interviene.
R. nei controlli automatici
Indica un problema di sintesi, cioè di progetto di un dispositivo R che, connesso al sistema o processo da controllare P, mantenga il valore di regime della variabile controllata y (in condizioni operative del sistema alterate per es. da disturbi z) costante (e pari al valore di riferimento r) o, nei problemi di asservimento, mantenga l’uscita y a distanza stabilita dal riferimento r(t) variabile nel tempo. In genere la r. si basa su una struttura a retroazione in cui l’azione di controllo si sviluppa in funzione dell’errore, con lo scopo di ridurlo; il sistema risultante deve essere asintoticamente stabile e, in corrispondenza di determinate classi di riferimenti e disturbi, rispondere con errore nullo in regime permanente. Per sistemi lineari a un ingresso e una uscita, si usano tecniche per tentativi nel dominio della frequenza (➔ regolatore), assegnando il tipo di sistema (grazie a un numero sufficiente di poli nell’origine nella funzione di trasferimento ad anello aperto) per soddisfare la specifica di progetto sul regime permanente, e successivamente usando reti di correzione elementari (anticipatrice e attenuatrice) per rendere asintoticamente stabile il sistema ad anello chiuso.
Per sistemi lineari multivariabili o non lineari, le tecniche si basano sulla rappresentazione di P con lo spazio di stato (➔ stato), esplicitando le equazioni differenziali che legano l’evoluzione di stato x e uscita y del sistema alle variabili di ingresso u e ai disturbi. La struttura del controllore dipende dalle informazioni disponibili sul sistema. Quando tutte le componenti (stato x del processo controllato e w del sistema che genera il disturbo) sono misurabili, l’ingresso di controllo si ottiene dalla combinazione delle variabili di stato come u=α(x, w); si parla di retroazione dallo stato e il controllore R è un sistema senza memoria. Se sono misurabili solo le componenti del vettore di errore e, si usa la retroazione dall’errore e il controllore è un sistema dinamico.
Nel primo caso, se l’approssimazione lineare di P intorno all’origine dello spazio di stato è stabilizzabile, condizione necessaria e sufficiente perché esista una soluzione è che sia risolubile un sistema misto di equazioni algebriche e differenziali alle derivate parziali, dette equazioni del regolatore; la condizione è costruttiva, poiché la soluzione fornisce la struttura del regolatore.
Nel secondo caso vale la stessa condizione, ma l’approssimazione lineare di P deve essere anche rilevabile, si deve cioè poter realizzare un osservatore asintotico dello stato che fornisca una stima dello stato (x, w) del sistema composito asintoticamente esatta a partire da misure dell’errore e. Si è nell’ambito della r. robusta se un medesimo controllore (con valori fissati dei parametri) risolve la r. per l’intera classe di processi ottenuta dal processo nominale variandone i parametri all’interno di un dato insieme.
R. delle macchine
Il problema della r. assume aspetti diversi secondo il tipo della macchina regolata. Nelle macchine operatrici a fluido la r. si riduce essenzialmente alla r. della portata di acqua nelle pompe, di aria nei compressori ecc.; nelle macchine motrici (per es., quelle che sono destinate a comandare generatori elettrici) si riduce essenzialmente alla r. automatica della velocità. In quest’ultimo caso si tratta d’impedire che con il variare del lavoro resistente la velocità angolare dell’albero motore possa allontanarsi troppo, in più o in meno, dalla velocità di regime normale, dando luogo a scarti non compatibili con il grado di regolarità desiderato. Tale scopo può essere ottenuto variando il lavoro motore in modo da riportarlo nel minor tempo possibile a un valore uguale al lavoro resistente. È in questo senso specifico che nella tecnica è usato il termine regolazione.
In senso più generale, come accade nelle macchine motrici da trazione, la r. si propone invece di variare la potenza del motore a seconda delle esigenze di servizio, consentendo una variazione del numero di giri entro limiti più ampi (variabili da tipo a tipo di motore), tali comunque da mantenere in ogni caso un buon rendimento. Tale r. è per lo più a volontà del conducente, ma può anche essere automatica, come nel caso della r. di sicurezza, destinata a impedire che in determinate circostanze, annullandosi improvvisamente il carico, la motrice possa assumere velocità pericolose (velocità di fuga).
Nelle macchine motrici a fluido, qualunque sistema di r., automatico o a mano, e in entrambi i sensi sopra specificati, agisce direttamente o indirettamente sull’organo di ammissione del fluido motore. La r. della potenza può essere ottenuta: a) variando la quantità di fluido (acqua, vapore o gas) introdotta nell’unità di tempo, lasciandone però inalterate le caratteristiche fisiche (r. per quantità di fluido); b) mantenendo sensibilmente costante la quantità di fluido introdotto nell’unità di tempo variandone però le caratteristiche (r. per qualità di fluido).
R. automatica dei motori elettrici
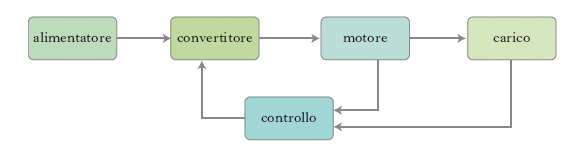
I progressi più significativi nelle prestazioni dei motori elettrici sono legati alla possibilità, che si è concretizzata su larga scala nel corso della seconda metà del 20° sec., di controllarne la velocità. Quest’obiettivo, nato con i primi motori elettrici, è stato perseguito a lungo, ma con evidenti limiti d’impostazione teorica e con scarso rilievo applicativo. L’unico motore di cui si era in grado di variare la velocità, e anche in questo caso a gradini e con rendimenti non elevati, era il motore a corrente continua, che così rappresentava la più valida soluzione offerta sia alle applicazioni industriali sia alla trazione elettrica. Lo sviluppo dei primi componenti elettronici di potenza allo stato solido ha consentito la realizzazione di raddrizzatori e convertitori controllati che permettevano al motore a corrente continua di soddisfare in maniera quasi esaustiva le esigenze di numerosi settori industriali, come quello delle industrie tessili, cartarie, siderurgiche, delle macchine utensili, e di tutto il settore della trazione elettrica. Inoltre, i componenti anzidetti, e ancor più tutti gli altri componenti elettronici successivamente introdotti sul mercato, hanno consentito lo sviluppo di convertitori statici di potenza in grado di produrre tensione alternata con ampiezza e frequenza variabili, che rappresenta la maniera più efficiente e affidabile di regolare le grandezze meccaniche di un motore a corrente alternata. Il motore elettrico è quindi diventato uno dei componenti di un sistema complesso che prende il nome di azionamento elettrico, il cui schema è mostrato in fig. 3. Con riferimento allo schema, un azionamento elettrico è costituito da un convertitore controllato di potenza e da un motore elettrico che aziona un carico meccanico con velocità variabile. Il convertitore di potenza è alimentato con tensione continua o alternata dalla sorgente di energia e alimenta a sua volta il motore con valori appropriati di tensione e corrente definiti dal sistema di controllo. A seconda del tipo di r. elettrica utilizzata, gli azionamenti si dividono in due categorie: in corrente continua (c.c.) e in corrente alternata (c.a.).
L’invenzione dei tiristori e il conseguente sviluppo dei convertitori c.a./c.c. ha dato inizio a una nuova era per gli azionamenti elettrici, che ha significato in quel momento la diffusione degli azionamenti in corrente continua. Questo è avvenuto poiché alla possibilità d’interrompere elettronicamente correnti elevate e, di conseguenza, variare opportunamente le forme d’onda di tensione e corrente, si accoppiava la prerogativa intrinseca del motore a c.c. di poter essere controllato rapidamente e con buon rendimento.
Il controllo della velocità dei motori a corrente alternata risulta in genere più complesso, poiché si tratta di sistemi non lineari, a molte variabili, nei quali è più complessa l’individuazione delle variabili di controllo capaci di agire direttamente su coppia e flusso della macchina, come avviene per l’eccitazione e la tensione di armatura nel caso dei motori a c.c.; si aggiunga che in talune macchine i flussi rotorici non sono accessibili e quindi non sono controllabili direttamente. In conseguenza di questi fatti, mentre un azionamento con motori a c.c. ha una struttura di controllo standardizzata, nel caso degli azionamenti a c.a. sono emerse innumerevoli soluzioni per gli altrettanto innumerevoli problemi di controllo che li riguardano, con riferimento alle varie applicazioni possibili e alle prestazioni diverse che in tali applicazioni sono richieste. Essenzialmente si distinguono applicazioni con basse e con alte prestazioni dinamiche, includendo tra le prime le macchine per il trattamento dei fluidi (pompe, ventilatori), per la trazione e per l’industria manifatturiera; mentre le seconde applicazioni comprendono la robotica e le macchine utensili.
Riguarda soprattutto gli impianti di propulsione di elevata potenza, per i quali l’esigenza dell’economicità di servizio richiede che il funzionamento delle diverse apparecchiature sia mantenuto a un livello di rendimento termico sempre vicino al massimo, anche nelle variazioni di carico. Alla r. automatica è normalmente abbinato il telecomando, effettuato da apposite stazioni, che consente sia la commutazione dalla condotta automatica a quella manuale e viceversa delle apparecchiature sia la variazione a distanza della velocità angolare delle motrici (quindi anche dell’elica).
Nelle Marine militari il telecomando dell’apparato di propulsione si esegue da un locale separato, detto centrale di propulsione, per consentire alla nave l’attraversamento di zone con aria (necessariamente introdotta nella sala motori, in quanto indispensabile per la combustione) contaminata dalla ricaduta di sostanze nocive, prodotte dallo scoppio di ordigni nucleari, biologici o chimici.