
nervoso, tessuto
Tessuto organico proprio dei Metazoi destinato alla ricezione degli stimoli e alla trasmissione degli eccitamenti ad altri tessuti i quali reagiscono in vario modo: con il movimento o con la secrezione. Per queste sue funzioni il tessuto n. mette l’organismo in relazione con l’esterno e stabilisce rapporti fra le varie parti dell’organismo, esplicando una funzione di correlazione (correlazione n.) che avviene in modo del tutto diverso dal meccanismo d’azione ormonale. Il tessuto nervoso è costituito da due popolazioni cellulari: i neuroni, che sono cellule specializzate nella conduzione degli impulsi ad alta velocità, e dalle cellule della nevroglia, che hanno funzioni di sostegno e trofiche.
Neurone
Il neurone è formato da un corpo, il pirenoforo, e da una serie di prolungamenti, i dendriti e il cilindrasse (detto anche assone o neurite).

Forma e volume del pirenoforo sono molto variabili: può essere sferico, ovale, piriforme, fusiforme o poliedrico; il diametro può essere di 4 come di 150 μm e, analogamente a quel che capita per l’estensione dell’arborizzazione dendritica, il volume è grossolanamente proporzionato all’estensione della zona con cui è in rapporto il neurone. Nel pirenoforo oltre alle strutture comuni alle altre cellule (membrana, nucleo con nucleoli, apparato reticolare di Golgi, mitocondri) sono presenti la sostanza cromofila di Nissl e le neurofibrille. La sostanza cromofila nelle colorazioni con il metodo di Nissl, che l’ha scoperta, appare sotto forma di masserelle distribuite nel citoplasma e nel tratto iniziale dei dendriti: i particolari ultrastrutturali rivelati dal microscopio elettronico permettono di identificarla nell’ergastoplasma, descritto in altre varietà cellulari. Le neurofibrille si evidenziano con l’impregnazione argentica: appaiono come sottili filamenti intrecciati distribuiti nel citoplasma e che si continuano nei dendriti e nel neurite, fino alle più esili ramificazioni terminali: risultano dall’aggregazione di sottili neurofilamenti di circa 100 Å di spessore (fig. 1).
I dendriti sono espansioni del corpo cellulare e rappresentano la maggior parte della superficie recettiva del neurone. Sono in generale multipli. Appaiono come ramificazioni tozze e grossolane; si evidenziano senza limite netto dal pirenoforo e terminano a breve distanza dal corpo cellulare. Hanno spesso superficie rugosa per la presenza di numerose spine o gemmule; si suddividono ripetutamente come i rami di un albero (da ciò il loro nome). I rami terminano liberamente in seno alla sostanza grigia, di rado ne oltrepassano i limiti decorrendo nella sostanza bianca. I dendriti contengono quasi tutti gli organuli presenti nel pericarion: mitocondri, corpi di Nissl (ergastoplasma), neurofibrille; e inoltre lunghi tubuli paralleli di circa 200 Å di diametro detti neurotubuli.
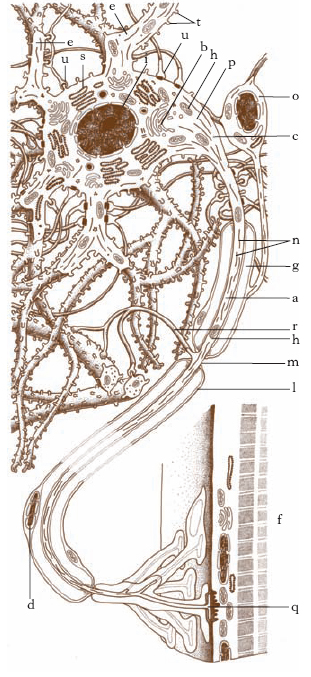
L’assone (o cilindrasse o neurite) è di solito unico. Si origina da un’elevazione conica del corpo cellulare, detta cono di emergenza. È più sottile dei dendriti, liscio, e non vi si riscontrano spine o rugosità. Nell’assone sono assenti i corpi di Nissl. Esso contiene mitocondri, tubuli orientati longitudinalmente di reticolo endoplasmatico liscio, neurotubuli simili a quelli dei dendriti e neurofilamenti (fig. 2). L’assone abbandona il luogo d’origine nella sostanza grigia, attraversa la sostanza bianca diventando una fibra nervosa e termina in un’altra parte della sostanza grigia oppure entra a far parte di un nervo periferico terminando alla periferia. Questi neuroni provvisti di un assone lungo sono chiamati cellule del I tipo di Golgi. In altri neuroni l’assone è corto, non entra nella sostanza bianca ma si ramifica ripetutamente esaurendosi nella sostanza grigia, a breve distanza dal corpo cellulare. Tali neuroni prendono il nome di cellule del II tipo di Golgi.
Struttura delle fibre nervose
Il neurite delle cellule del I tipo di Golgi conserva gli stessi caratteri nel suo lungo decorso, ma nei Vertebrati adulti, a una certa distanza dall’origine, si riveste di guaine proprie diventando una fibra nervosa; a queste si sovrappongono nei nervi periferici cerebrospinali altre guaine dipendenti dal connettivo. La fibra nervosa è quindi composta dall’assone e dalle sue guaine. Le fibre sono indicate come: fibre mieliniche con neurilemma; fibre mieliniche senza neurilemma; fibre amieliniche con neurilemma e senza (neuriti nudi).
In tutti i Vertebrati fuorché nei Ciclostomi, le fibre mieliniche con neurilemma sono la continuazione di neuriti delle cellule motrici del midollo e dell’encefalo e delle cellule dei gangli sensitivi cerebro-spinali. Sono lunghe quanto il nervo di cui fanno parte, così che la loro lunghezza risulta proporzionale alla mole corporea. Vi è inoltre un rapporto quasi costante tra lo spessore della fibra e il volume della cellula da cui essa si origina. La guaina mielinica costituisce un manicotto che avvolge il neurite, il quale nelle fibre fresche risulta quindi mascherato dall’opacità della mielina. Al microscopio elettronico la guaina mielinica risulta costituita da una serie concentrica di lamine scure e chiare che si ripetono con un periodo regolare di 120 Å. Essa si interrompe a intervalli regolari (strozzamenti anulari, o nodi, di Ranvier, che nell’uomo sono disposti a distanza di 0,8-1 mm). Quando la fibra si altera, dalle lecitine della mielina si liberano acidi grassi e si formano esteri grassi che si colorano in nero con l’acido osmico.
Alla superficie esterna della guaina mielinica aderisce il neurilemma o guaina di Schwann, costituita dalle cellule di Schwann e dalle loro propaggini citoplasmatiche. Le cellule di Schwann sono di origine ectodermica come le cellule nervose, e sono considerate come cellule di nevroglia che durante lo sviluppo embrionale hanno migrato seguendo l’assone e formando attorno a questo una guaina completa. Vi è una sola cellula di Schwann per ogni segmento internodale, cioè per ogni tratto di fibra tra due consecutivi nodi di Ranvier. In corrispondenza del nodo di Ranvier, dove la guaina mielinica è interrotta, l’assone è circondato parzialmente soltanto dalle propaggini citoplasmatiche di due cellule di Schwann adiacenti e da una membrana basale.
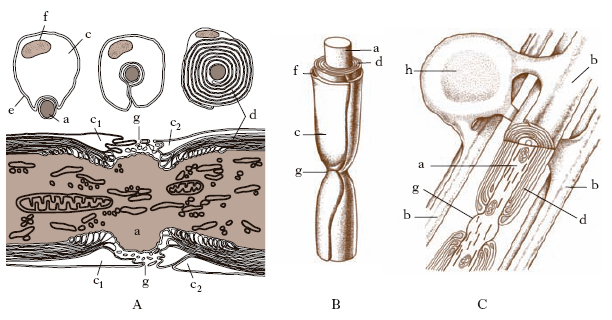
La guaina mielinica è una dipendenza della cellula di Schwann, essendo formata da successivi avvolgimenti concentrici della membrana plasmatica della cellula di Schwann attorno all’assone. Ogni segmento internodale risulta perciò costituito da una cellula di Schwann con le sue lamelle mieliniche (fig. 3A e 3B). Nelle fibre del sistema nervoso centrale la guaina mielinica è formata dalle cellule di oligodendroglia. Si ritiene che la mielinizzazione avvenga, come nei nervi periferici, mediante un avvolgimento dei prolungamenti della cellula di glia intorno agli assoni; in questo caso, però, un singolo oligodendrocito avvolge più di un assone (fig. 3C).
Le fibre mieliniche costituiscono la sostanza bianca dei centri nervosi e dalla presenza della mielina (che non presenta strozzamenti anulari) dipende il loro aspetto bianco splendente; sono sparse in gran numero anche nella sostanza grigia. Non si anastomizzano mai l’una con l’altra, ma in punti determinati emettono rami collaterali diretti verso la sostanza grigia dove si esauriscono con arborizzazioni terminali. Le fibre amieliniche si trovano nell’embrione di tutti i Vertebrati e nel sistema n. centrale dei Ciclostomi. Sono sprovviste di guaina anche un certo numero di fibre delle radici e dei nervi periferici cerebrospinali dei Vertebrati adulti, un tratto del neurite in prossimità immediata della cellula, molte fibre del simpatico (fibre grigie) ecc.
Terminazioni nervose
Per le proprietà funzionali si distinguono fibre dei nervi recettori, o afferenti, e degli effettori, o efferenti. Le espansioni dei neuriti a conduzione centripeta, o recettori, si trovano in ogni regione del corpo e hanno connessioni morfologicamente diverse con gli elementi specifici dei singoli organi. Nell’epidermide e in molte mucose le fibre si dividono ripetutamente terminando negli intervalli fra le cellule epiteliali in esili rami destinati a trasmettere le sensazioni di dolore. Invece negli apparati di senso specifico (dell’udito, dell’equilibrio, del gusto) le fibre nervose hanno connessioni più intime con cellule epiteliali differenziate in special modo per la recezione degli stimoli. Soltanto negli organi dell’olfatto le cellule di senso sono vere e proprie cellule nervose (cellule di senso primarie) perché, pur essendo dotate di funzione recettrice, esse stesse con il loro neurite emettono la fibra per la conduzione centrale. Le fibre recettrici che si espandono nel connettivo terminano in parte liberamente, in parte con corpuscoli terminali di forma svariatissima ma che hanno qualche carattere comune. Il connettivo forma un involucro intorno al tratto terminale della fibra, la quale percorre la parte assile del corpuscolo, dopo essersi spogliata delle guaine, e spesso ha un tragitto flessuoso e presenta delle varicosità. Fra i corpuscoli terminali del connettivo che hanno maggiore diffusione nell’uomo sono da ricordare i seguenti: i corpuscoli di Pacini, numerosissimi nel connettivo sottocutaneo della palma della mano e della pianta del piede; i corpuscoli di Golgi-Mazzoni, del tessuto sottocutaneo, destinati a ricevere le sensazioni di freddo; i corpuscoli di Meissner, numerosi nelle papille del derma; i corpuscoli di Ruffini, frequenti nella parte superficiale del connettivo sottocutaneo che, come quelli di Pacini, si ritiene siano destinati a ricevere le sensazioni del caldo; gli organi muscolo-tendinei del Golgi esistenti in quasi tutti i tendini dei muscoli striati: hanno forma affusata, contorno netto e intervengono nella regolazione della contrazione muscolare.
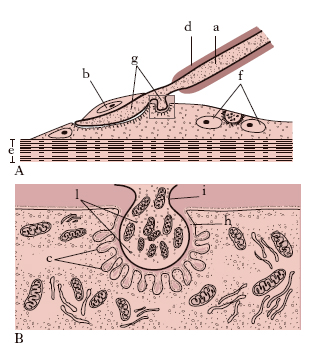
I neuriti a conduzione centrifuga, o effettori, si distinguono in due varietà diverse morfologicamente e funzionalmente: somatici, destinati alla muscolatura volontaria e provenienti da neuroni radicolari del midollo e dell’encefalo, e viscerali, che innervano la muscolatura liscia e le ghiandole e provengono da neuroni dei gangli del simpatico. I primi terminano nella fibra muscolare con le cosiddette placche motrici (fig. 4).
Sinapsi
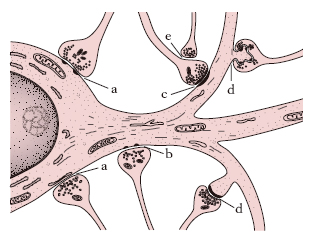
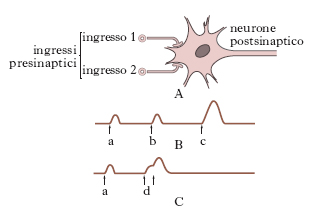
I punti di contatto tra due neuroni prendono il nome di sinapsi. Le sinapsi sono i luoghi di trasmissione dell’impulso da un neurone all’altro; esse possono istituirsi (fig. 5) tra l’assone di un neurone e il corpo cellulare di un altro neurone (sinapsi asso-somatiche), oppure tra assone e dendriti (sinapsi asso-dendritiche) o più raramente tra assone e assone (sinapsi asso-assoniche). Le sinapsi sono costituite dall’affrontamento delle rispettive membrane plasmatiche dell’assone e dei dendriti o del corpo cellulare e non esiste passaggio di neurofibrille da un neurone all’altro attraverso la sinapsi. Nella terminazione dell’assone, a livello della sinapsi, sono assenti le neurofibrille e sono presenti mitocondri e microvescicole, di circa 200-650 Å di diametro, le vescicole presinaptiche o sinaptosomi, che contengono particolari sostanze chimiche, il cosiddetto mediatore dell’impulso nervoso. Le superfici pre- e postsinaptica sono separate da uno spazio largo circa 20 nm (fessura sinaptica). Ogni neurone può contrarre rapporti mediante le sinapsi con un gran numero di altri neuroni. Una cellula del Purkinje del cervelletto può avere migliaia di sinapsi nei suoi dendriti.
Le caratteristiche funzionali delle sinapsi si possono riassumere come segue: l’impulso nervoso è trasmesso unidirezionalmente (a differenza di quanto si verifica nella fibra nervosa, nella quale l’impulso può essere bidirezionale); esiste un ritardo ben definito nella trasmissione dell’impulso, correlato alla liberazione del mediatore chimico; è possibile un’integrazione dei messaggi sinaptici, nel senso che se impulsi subliminali convergono simultaneamente da vie diverse possono risultare efficaci (sommazione presinaptica); non tutte le sinapsi hanno carattere eccitatorio, ma alcune possono svolgere funzioni inibitorie.
Funzionalità della cellula nervosa
Proprietà fondamentali del neurone sono l’eccitabilità, la conduttività e la trasmissibilità dell’eccitamento. L’eccitabilità, che si definisce come la proprietà di reagire a una modificazione dell’ambiente, è specifica; ogni neurone reagisce secondo la sua natura e secondo la natura dello stimolo. Uno stimolo, per eccitare un neurone, deve essere adeguato, avere cioè una determinata intensità e raggiungere un dato valore di soglia; stimoli di intensità insufficiente sono detti subliminali. Una volta eccitato, il neurone risponde con una reazione massima che non aumenta aumentando l’intensità dello stimolo (legge del tutto o niente); lo stimolo può peraltro, aumentando di intensità, eccitare più neuroni, e in questo modo può anche aumentare l’intensità della risposta.
La conduttività, o proprietà di condurre gli eccitamenti, oltre a obbedire alla legge del tutto o niente, presenta altri caratteri: l’impulso decorre isolato come in un cavo elettrico in ogni fibra nervosa (legge della conduzione isolata) e, di fronte a stimoli artificiali, l’eccitamento si diffonde come un’onda indifferentemente in ambedue le direzioni (legge della conduzione indifferente); sarà poi la sinapsi a stabilire la direzione che dovrà seguire. La fibra nervosa inoltre non modifica nella conduzione l’eccitamento. La velocità di propagazione dell’impulso nervoso varia a seconda del tipo di fibra (si distinguono fibre a piccola, media e grande velocità di conduzione, le più lente essendo rappresentate dalle fibre del sistema nervoso vegetativo), ed è compresa fra 0,60-120 m/s.
L’altra importante proprietà del neurone è la trasmissibilità dell’eccitamento che avviene tramite le sinapsi o articolazioni interneuroniche; queste sono dotate di singolari proprietà quali la conduzione lenta e in un sol senso, la facile faticabilità, l’inibizione in determinati casi della propagazione dell’impulso, la sommazione degli impulsi subliminali e la facilitazione della loro trasmissione; la sinapsi è inoltre il luogo su cui agiscono specificamente alcuni farmaci (stricnina). La possibilità di trasmettersi degli impulsi nervosi, oltre a dipendere dall’intensità, è anche legata alla loro durata; per ogni neurone esiste infatti una durata minima di passaggio di una corrente elettrica, sempre di intensità sufficiente, capace di determinare l’eccitamento; essa è detta cronassia: perché l’eccitamento si trasmetta da neurone a neurone è necessario che le rispettive cronassie siano uguali o l’una il doppio dell’altra (legge dell’isocronismo dei neuroni). Neuroni che intervengono in una stessa funzione hanno valori cronassimetrici molto vicini o uguali.
La trasmissione dell’impulso a livello della sinapsi è dovuto a un meccanismo elettrochimico. La sinapsi effettrice libera dai sinaptosomi una sostanza, il mediatore chimico, la cui natura (acetilcolina, noradrenalina, serotonina, sostanza P) è stata individuata solo per alcune varietà di sinapsi. Il mediatore si diffonde nella fessura intersinaptica e agisce sulla membrana postsinaptica, cioè sulla cellula recettrice, dando avvio a una serie di fenomeni di membrana, globalmente riassumibili in una temporanea alterazione degli equilibri locali, imperniata sull’aumento della conduttanza della membrana per gli ioni Na+ prima, e per gli ioni K+ subito dopo. Gli ioni Na+, la cui concentrazione allo stato di riposo è molto più elevata all’esterno che all’interno della membrana, grazie all’improvviso aumento della permeabilità di questa passano massicciamente all’interno, tendendo a raggiungere il potenziale di equilibrio, che però non toccano perché l’intensa positività indotta da essi stessi con il loro arrivo nell’interno della cellula crea un contrasto tra il gradiente elettrico e il gradiente di concentrazione. La neutralità elettrica così turbata viene ristabilita da un movimento in senso inverso degli ioni K+, che passano all’esterno della membrana. Il potenziale di membrana, in rapporto all’afflusso nel suo interno delle cariche positive rappresentate dagli ioni Na+, dapprima si annulla (depolarizzazione), poi si inverte, e potenziale d’azione viene denominato il potenziale di membrana invertito. L’estrusione delle altre cariche positive rappresentate dagli ioni K+ dà inizio invece alla discesa del potenziale d’azione, al ristabilimento del potenziale di membrana e alla ridistribuzione degli ioni Na+ e K+ secondo le proporzioni che caratterizzano la fase di riposo.
Per ottenere nella cellula nervosa un potenziale d’azione è necessario che gli impulsi sinaptici raggiungano un valore critico: tale condizione si può creare (fig. 6) per un’attivazione contemporanea di più sinapsi (sommazione spaziale) o per una rapida successione degli stimoli (sommazione temporale). Nella fibra neuritica l’eccitamento si propaga con un meccanismo di autorigenerazione, paragonabile alla combustione di una miccia: il potenziale d’azione che insorge in un primo tratto della fibra è stimolo sufficiente all’insorgenza del fenomeno nel tratto prossimale, per cui l’impulso percorre tutta la fibra con pari intensità, cosa che non avverrebbe se la fibra fosse un semplice trasmettitore passivo e non venisse coinvolta attivamente nel processo in tutta la sua estensione; differenze esistono solo nella velocità di trasmissione dello stimolo e dipendono essenzialmente dal diametro delle fibre, dalla presenza o assenza della guaina mielinica e dalla temperatura: la velocità diminuisce con l’abbassarsi della temperatura e la perfrigerazione provoca il blocco della trasmissione dell’impulso.

Nelle fibre mieliniche dei Vertebrati la velocità di conduzione è superiore a quella che dovrebbe essere consentita dalle dimensioni del loro diametro; ciò accade perché la conduzione procede secondo una diversa modalità, cioè a salti (teoria saltatoria): i potenziali d’azione si sviluppano in tutta la loro ampiezza solo a livello dei nodi di Ranvier, dove la fibra è sprovvista di avvolgimento mielinico, e la conduzione nei tratti internodali si propaga passivamente (fig. 7).
Patologia
Le alterazioni delle cellule nervose in condizioni patologiche che non siano malformative o neoplastiche consistono in alterazioni regressive, che si manifestano col rigonfiamento torbido o col raggrinzimento e la sclerosi del corpo cellulare, o anche, nei casi più gravi, con la dissoluzione parziale o totale del corpo cellulare. Altre modalità di alterazione consistono nella scomparsa della struttura fibrillare con omogeneizzazione del corpo cellulare, nella infiltrazione grassosa e nella incrostazione di sali di calcio. Nelle demenze delle età presenile e senile si osserva la conglutinazione delle neurofibrille in fasci compatti a decorso vorticoso e la formazione, a spese della sostanza intercellulare, delle cosiddette placche senili, costituite da conglomerati di filamenti e di granuli argentofili, di detriti cellulari e di elementi neurogliali. Nelle fibre nervose l’alterazione è denunciata dalla frammentazione della guaina mielinica in sferule ovoidi, dalla formazione di prodotti di disfacimento, e dallo spezzettamento dei cilindrassi.
La nevroglia astrocitaria partecipa ai processi patologici del tessuto n. ora con alterazioni regressive che determinano la perdita dei prolungamenti ed eventualmente la trasformazione in senso ameboide dei corpi cellulari, ora con fenomeni di ipertrofia o di proliferazione (gliosi). Nell’oligodendroglia sono ben dimostrate le alterazioni regressive. La microglia subisce alterazioni caratteristiche: la trasformazione in cellule a bastoncello, che sono caratteristiche della paralisi progressiva, in cellule granulo-adipose in tutti quei processi nei quali è stimolata la sua funzione fagocitaria.