
encefalo
Regione anteriore del sistema nervoso centrale dei Vertebrati racchiusa nella scatola cranica, sede dei centri della sensibilità olfattiva, visiva, acustica, statica, nonché dei centri associativi, integrativi, e dei centri superiori della corteccia cerebrale.
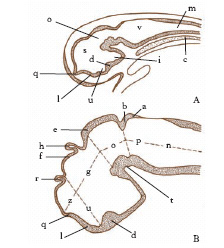

L’e. origina dall’ispessimento della piastra neurale, ectodermica, i cui bordi si ispessiscono (pieghe neurali) e si sollevano formando la doccia neurale; questa, per un processo di rottura e saldatura, si trasforma in un tubo, aperto anteriormente con il neuroporo e comunicante posteriormente con l’archenteron attraverso il canale neuroenterico. La piastra neurale si differenzia in due regioni, una anteriore dilatata che darà origine all’ archiencefalo o prosencefalo e una posteriore più stretta che darà origine al deuterencefalo e al midollo spinale. Successivamente il deuterencefalo si suddivide in due vescicole, il mesencefalo e il rombencefalo. È questo lo stadio a 3 vescicole (fig. 1A), cui corrispondono le tre funzioni fondamentali dell’e., apparse agli stadi più precoci della filogenesi: la funzione olfattiva propria del prosencefalo, quella visiva propria del mesencefalo e quella acustico-statica propria del rombencefalo. Segue lo stadio a 5 vescicole (fig. 1B), comune a tutti i Vertebrati. Esse sono, in senso cefalo-caudale: telencefalo e diencefalo, derivati dalla divisione del prosencefalo; mesencefalo, che rimane indiviso; metencefalo e mielencefalo, derivati dalla divisione del rombencefalo. A questo stadio l’e. presenta 3 curvature (fig. 2): la curvatura cefalica, a concavità ventrale, tra mesencefalo e metencefalo; la curvatura del ponte, a concavità dorsale, nella regione del metencefalo; la curvatura nucale, a concavità ventrale, per cui l’e. risulta piegato ventralmente rispetto al midollo spinale. Queste curvature, evidenti negli Uccelli e nei Mammiferi e accennate negli altri Vertebrati, sono dovute all’ineguale accrescimento delle pareti dorsale e ventrale dell’encefalo. Nel midollo spinale, allo stadio di tre vescicole, si ispessiscono le pareti laterali per attiva moltiplicazione dei neuroblasti, mentre le pareti dorsale e ventrale rimangono sottili: queste ultime danno rispettivamente la placca del tetto (lamina dorsale), e la placca basale (lamina ventrale). Gli ispessimenti delle pareti laterali sono suddivisi internamente in due parti: latero-dorsale, detta lamina alare, con funzione sensitiva, e lateroventrale, detta lamina basale, con funzione motoria.
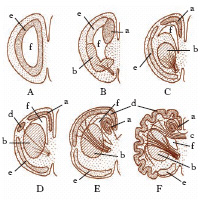
Dal telencefalo si sviluppano tratti, bulbi e lobi olfattori (formazioni pari in relazione con i corrispondenti organi di senso), i corpi striati e i gangli della base. Gli emisferi cerebrali, che si formano come evaginazioni del telencefalo, sono presenti solo dagli Anfibi in su, nelle classi più recenti dei Vertebrati. La parte latero-dorsale degli emisferi costituirà il pallio. Nell’evoluzione filogenetica si ha una successione di 3 palli (fig. 3): dapprima il paleopallio, che è strettamente connesso e spesso s’identifica con i lobi olfattivi; quindi l’archipallio, che diverrà l’ippocampo, e infine il neopallio, che forma la corteccia cerebrale. Anteriormente agli emisferi possono estroflettersi i lobi olfattivi con i tratti e i bulbi. Formazioni telencefaliche impari sono: la commissura anteriore o paleopalliale e la lamina anteriore col recesso neuroporico; la commissura archipalliale o di ippocampo, che compare negli Anfibi; il corpo calloso, neopalliale, che compare nei Mammiferi placentati.
Nel diencefalo dalle lamine laterali si differenziano i talami ottici, centri di correlazione visiva, e dalle lamine dorsali e ventrali rispettivamente l’epitalamo e l’ipotalamo, che hanno frequentemente funzione ghiandolare.
Dal mesencefalo si sviluppa dorsalmente il tetto ottico a funzione visiva (lobi ottici o quadrigemini anteriori) e, nei Mammiferi, anche acustica (quadrigemini posteriori); ventralmente si ha il corpo distinto in tegumento e peduncoli cerebrali contenenti vie ascendenti e discendenti.
Il metencefalo è costituito dorsalmente dal cervelletto, organo di correlazione statica e motoria (presente come semplice banderella nei Ciclostomi), e ventralmente da fasci di fibre, che nei Mammiferi collegano la corteccia cerebrale con quella cerebellare (ponte di Varolio) e, nei Vertebrati inferiori, il cervelletto con il midollo allungato.
Il mielencefalo, che a differenziamento avvenuto si chiama midollo allungato, rappresenta la regione di transizione tra e. e midollo spinale. La parte dorsale resta sottile a formare un tetto epiteliale fortemente vascolarizzato (tela coroidea); le pareti laterali, ispessite, che limitano la cosiddetta fossa romboidale, sono sede di importanti centri nervosi somatici e vegetativi.
Nel progressivo sviluppo delle vescicole cerebrali la cavità originaria del prosencefalo, con la formazione del telencefalo, si suddivide a formare i due ventricoli laterali situati in ciascun emisfero; la cavità del diencefalo forma il III ventricolo comunicante attraverso i forami interventricoli (di Monro) coi ventricoli cerebrali; la cavità del mesencefalo (mesocele), ampia nell’encefalo delle classi più antiche dei Vertebrati, si riduce a uno stretto canale, l’iter (acquedotto), che fa comunicare il III con il IV ventricolo al di sotto del cervelletto e che si estende nel mielencefalo restringendosi gradualmente fino a continuarsi con il canale centrale del midollo spinale. Nei Vertebrati di origine più antica la parte anteriore del IV ventricolo può estendersi, come cavità a sé, nel cervelletto e costituire un metacele o epicele. I derivati delle vescicole subiscono ancora fondamentali cambiamenti morfologici in seguito allo sviluppo di tratti trasversali di fibre (commissure) che connettono le metà opposte, destra e sinistra dell’e., contribuendo alla sua unità funzionale.
Per la descrizione complessiva dell’e. nell’uomo ➔ nervoso, sistema.