
tessuto
Biologia
In istologia, aggregato di cellule (e di sostanze da esse prodotte) che hanno forma, struttura e funzioni simili, e, per lo più, origine embriologica comune; i t., di cui si hanno diversi tipi, sono i materiali costitutivi degli organi animali e vegetali. Un organo è costituito per lo più da t. di tipo diverso, più o meno intimamente connessi. I t. dell’uomo si raggruppano in quattro grandi categorie: epiteliali, connettivi (che comprendono anche il sangue e la linfa), muscolari e nervosi.
Caratteristiche
L’evoluzione degli organismi pluricellulari ha permesso la formazione di cellule e t. specializzati. Dato che i t., sia negli organismi animali sia nei vegetali, sono strutture organizzate costituite da cellule specializzate nelle quali è assolta una specifica funzione, le cellule dei vari t. presentano strutture e caratteristiche chimiche diverse. In alcuni t., quali, per es., quelli epiteliali, la maggior parte delle cellule stabilisce stretti contatti con le cellule circostanti, interagendo con esse in maniera specifica. In altri t. le cellule secernono una serie di macromolecole che formano una complessa rete di proteine e carboidrati che riempie gli spazi extracellulari e costituisce la cosiddetta matrice extracellulare. La composizione della matrice extracellulare differisce notevolmente da un t. all’altro. Per es., il t. osseo è essenzialmente costituito da una matrice cellulare rigida nella quale è disperso un numero molto limitato di cellule, mentre il t. cartilagineo presenta una matrice con caratteristiche di maggiore flessibilità. Il t. connettivo che circonda le ghiandole e i vasi sanguigni ha invece una matrice gelatinosa, nella quale sono disperse numerose cellule.
Questi esempi illustrano il ruolo fondamentale della matrice nel determinare la dimensione e le proprietà meccaniche dei t. e degli organi. Le molecole che costituiscono la matrice appartengono a tre classi: i glicosamminoglicani e i proteoglicani, tipici della cartilagine, che formano una sostanza idratata gelatinosa; le proteine strutturali, come i collageni e l’elastina, che forniscono ai t. resistenza e flessibilità; le proteine adesive, quali la fibronectina e la laminina, che favoriscono l’unione delle cellule alla matrice. Per la formazione di un t. è necessario che più cellule specializzate, originatesi in quella stessa sede o migrate da sedi lontane, si riconoscano e interagiscano fra loro in modo ordinato.
I processi che sono alla base del riconoscimento fra cellule negli organismi pluricellulari sono complessi; per studiarli è necessario dissociare il t. nelle singole cellule che lo compongono e osservare come queste si riaggregano in vitro.
Formazione dei tessuti
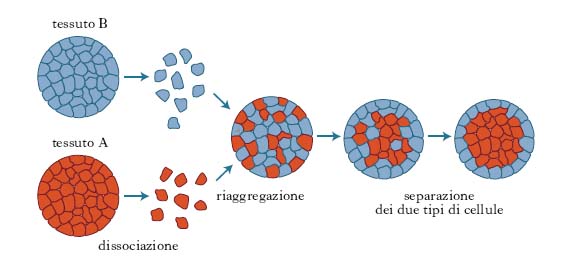
Si è visto che una caratteristica peculiare delle cellule nei vari t. consiste nella loro tendenza ad aderire in maniera specifica a cellule dello stesso tipo o di un tipo simile: se si mescolano insieme cellule dissociate ottenute da t. diversi di un embrione di vertebrato, esse inizialmente si associano in un aggregato misto e successivamente si riuniscono in due gruppi diversi, in base alla loro diversa origine tessutale (v. fig.). Se si ripete l’esperimento con sospensioni cellulari ottenute da due embrioni di specie diverse, le cellule si riuniscono per formare aggregati t.-specifici e non specie-specifici. Per es., disaggregando cellule epatiche, esse tendono a riassociarsi fra loro piuttosto che con altri tipi di cellule dissociate, quali quelle della retina o di un muscolo. Queste interazioni, dette omotipiche, possono avvenire grazie alla presenza sulla superficie cellulare di molecole di adesione. Sono state identificate due famiglie di molecole di adesione intercellulare: quelle che richiedono ioni Ca2+ per esplicare il loro ruolo adesivo, le caderine (➔ caderina), e quelle che non li richiedono. Tra le molecole di adesione intercellulari calcio-indipendenti le più diffuse sono le N-CAM (➔ ).
Si può ritenere che la precoce comparsa, in momenti diversi dello sviluppo, di molecole di adesione con diversa specificità abbia un ruolo importante nei processi dello sviluppo e della morfogenesi. L’adesione fra cellule e matrice nei diversi t. è determinata dalla fibronectina e da altre proteine che si è riusciti a isolare, quali la laminina, la vitronectina, la trombospondina, la tenascina. Molecole chiave nelle interazioni fra cellula e matrice sono inoltre le integrine, proteine transmembrana che hanno la funzione di recettori di varie molecole di adesione. Anche le interazioni fra cellule presenti in t. diversi giocano un ruolo particolarmente importante durante il differenziamento dei tessuti. Un esempio è costituito dal differenziamento del cristallino dalle cellule dell’epidermide, indotto da particolari cellule nervose dell’ectoderma nel calice ottico. Utilizzando modelli sperimentali di cellule in coltura è stato possibile inoltre dimostrare, per es., che le cellule epiteliali, come quelle della ghiandola mammaria, possono assumere la loro peculiare architettura e sintetizzare proteine t.-specifiche grazie alla presenza di una struttura denominata lamina basale (➔ lamina). Inoltre è noto che i particolari contatti fra cellule di diverso tipo che caratterizzano il sistema nervoso sono il risultato della capacità degli assoni delle cellule nervose di accrescersi, seguendo percorsi specifici che li conducono verso i rispettivi bersagli cellulari.
Terapia tessutale
Espressione coniata da V.P. Filatov (1937) per designare un metodo terapeutico da lui proposto, detto anche tessutoterapia, che utilizza, di solito per innesti sottocutanei o per iniezioni, le proprietà biologiche acquisite da alcuni t. di provenienza animale o vegetale, isolati dall’organismo.
Botanica
Nelle piante inferiori le cellule che ne costituiscono il corpo (tallo) restano sempre molto autonome e sono relativamente poco differenziate. I loro aggregati formano pseudotessuti, la cui origine può avvenire: a) per associazione dovuta all’incontrarsi fortuito di cellule; b) per il permanere da una generazione all’altra dell’involucro gelatinoso che le racchiude; c) per divisione cellulare non seguita dalla separazione delle cellule figlie. Le cellule degli pseudotessuti si dividono secondo una, due o tre direzioni dello spazio: ne derivano nel primo caso cellule filamentose, nel secondo cellule laminari, nel terzo cellule massicce. Le Alghe e i Funghi sono esempi di piante costituite di pseudotessuti.
Nelle piante vascolari (Cormofite) si possono identificare diversi tipi di cellule che si distinguono in base alla loro origine da determinate linee embrionali, alle caratteristiche morfologiche e alle funzioni. Gli aggregati cellulari che esse formano sono veri t., che hanno una localizzazione e sono associati negli organi (radice, fusto, foglia) in modo sensibilmente tipico e costante. I t. detti definitivi, o adulti, provengono da altri t., detti meristematici, o embrionali, le cui cellule hanno la capacità di moltiplicarsi attivamente e talora indefinitamente per formare poi, attraverso i processi di differenziamento, le cellule dei t. definitivi. Nelle piante, a differenza degli animali, l’accrescimento è indefinito: lo dimostra, per es., la periodica formazione di nuovi rami e foglie, che ha luogo per tutta la durata della vita della pianta. Questa crescita illimitata è assicurata dai meristemi apicali (degli apici vegetativi e radicali) che derivano direttamente, come il cambio intrafasciale, dall’embrione (fanno eccezione le radici e le gemme avventizie). Man mano che, partendo dall’embrione, la pianta si sviluppa e cresce longitudinalmente (in lunghezza) e radialmente (in grossezza) si formano continuamente nuove cellule destinate a differenziarsi in elementi di t. definitivi; questa produzione di cellule avviene in ben determinate regioni della pianta che restano costantemente allo stato meristematico. Le cellule vegetali hanno un’elevata capacità di rigenerazione e perciò, tranne rari casi, poste in adatte condizioni, manifestano la loro totipotenza o riacquistano la capacità di dividersi attivamente (come le cellule dei meristemi), e possono produrre nuovi organismi interi. Tale capacità è, peraltro, manifestata in parte anche dalla pianta intera con la frequente produzione di nuovi germogli, la rimarginazione di ferite, la produzione di radici secondarie e di gemme avventizie e latenti.
I t., oltre alla loro specializzazione funzionale e morfologica, presentano alcuni caratteri costanti, come la continuità protoplasmatica fra le cellule, dovuta al fatto che le pareti cellulari sono perforate da piccoli canali capillari, i porocanali, che si corrispondono fra cellule attigue e sono attraversati da filamenti, o ponti, protoplasmatici (plasmodesmi); perciò spesso non vi è una netta separazione tra le cellule del t., ma una continuità morfologica e una collaborazione funzionale assai strette, che fanno del t. un insieme organizzato.
Tecnica
Manufatto costituito da un insieme di fili, ricavati dalle fibre tessili e intrecciati fra loro, mediante l’opera di tessitura, secondo un determinato ordine. Tipi particolari di t., ottenuti con materiali non tessili, sono i t. metallici e i t. di vetro. Con il nome di non t., o anche di t. non t. (TNT), si indicano infine varie strutture tessili piane prodotte con mezzi diversi dalla tessitura e dalla maglieria (con esclusione dei feltri e strutture analoghe) e, con senso più ristretto, una struttura tessile consistente in un velo di fibre (fondo) tenute insieme con vari procedimenti di collegamento: con leganti chimici (per es., latice di gomma naturale o artificiale), mediante trattamenti termici (per le fibre termoplastiche), o con lavorazioni meccaniche (per es., cuciture); i non t., che hanno bassi costi di produzione, vengono usati per imbottiture e rinforzi di ogni tipo, oggetti da gettare dopo l’uso, articoli sportivi, cuoi artificiali, filtri.
Tipi di tessuto
Il t., nella sua accezione più ampia, si può ottenere in tre modi: a) per intreccio di una serie di fili disposti longitudinalmente (ordito) con uno o più fili disposti trasversalmente (trame); si realizza sul telaio o sulla macchina per tessere; b) per concatenazione, secondo anse o boccole, di uno o più fili disposti secondo la stessa direzione; si realizza manualmente per mezzo di bastoncini metallici (ferri) o meccanicamente grazie a telai e macchine di maglieria; c) per intreccio di una serie di fili longitudinali con un filo trasversale, che all’inizio del lavoro è anche esso longitudinale e che si pone nell’assetto trasversale solo al momento dell’intreccio; con questo sistema si realizzano pizzi, tende, merletti ecc. Economicamente di gran lunga più importanti sono i t. per intreccio ortogonale di ordito e trama (commercialmente indicati semplicemente come t.) e i t. per evoluzione di una sola serie di fili (commercialmente indicati come maglie).
T. per intreccio di ordito e trama
I t. veri e propri sono realizzati su telaio o su macchina per tessere e sono caratterizzati da tre dimensioni: altezza, lunghezza, spessore. La prima è vincolante e caratterizza l’uso del t. stesso; la seconda serve solo per quantificare il t. a livello commerciale; la terza, indirettamente, determina la massa e l’impiego del tessuto. Nella terminologia della tessitura, i t. veri e propri danno luogo a due fasi ben distinte: a) preparazione dell’ordito (orditura, eventuale imbozzimatura, passatura o rimettaggio nei licci e nel pettine, attrezzaggio della macchina da tessere); b) formazione dell’intreccio sulla macchina da tessere.
Le possibilità d’intreccio o di legatura sono praticamente infinite, comunque riconducibili a tre armature base (tela, saia, raso). Le varie armature (base e/o derivate) si ottengono a telaio, programmando in maniera diversa l’alzata e l’abbassata contemporanea dei fili d’ordito, in modo da promuovere il cosiddetto passo d’ordito attraverso cui lanciare la navetta (telaio) o il dispositivo porta-trama (macchina da tessere). Tutti i t. sopra descritti, ottenuti con movimento dei fili d’ordito (in alzata o in abbassata) determinato da licci (➔ telaio) cui i fili sono solidali, sono detti lisci; per contrapposizione, si dicono operati i t. in cui i fili d’ordito sono suscettibili di movimento indipendente, grazie alla macchina jacquard. Chiaramente, per quanto riguarda l’aspetto finale del t., concorrono non solo il tipo d’armatura, il colore dei filati adoperati, la presenza di orditi o trame supplementari, la tecnica di tessitura, la varietà del disegno, a seconda si tratti di t. lisci od operati, ma anche il pregio della materia prima con cui si sono realizzati i filati, le caratteristiche costruttive (titolo, torsione ecc.) dei medesimi, il tipo di rifinizione cui il t. viene sottoposto.
T. a maglia
Le maglie, o i t. a maglia, vanno innanzitutto suddivisi in due categorie: a maglia in trama e a maglia in catena. Nella maglia in trama, l’alimentazione del filo singolo o di pochi fili si svolge orizzontalmente ed effettua la formazione delle maglie in successione, sino ad avere un rango completo; si dice anche a maglie raccolte, in quanto è l’ago che prende il filo. Nella maglia in catena, l’alimentazione di una serie di fili paralleli è simultanea per tutti gli aghi in lavoro che formano così, cumulativamente, un rango di maglie. In ambedue i casi il manufatto è caratterizzato da grande estensibilità o elasticità, dal momento che le boccole o anse possono distendersi in un senso o nell’altro. La compattezza di una maglia, oltre che dal titolo o grossezza dei filati adoperati, sarà determinata anche dalla densità delle maglie (numero di ranghi e di fili al centimetro); l’aspetto finale del manufatto, oltre che dal tipo di maglia, sarà invece determinato dalla natura e dalle caratteristiche costruttive dei filati, da alcune varianti di esecuzione sulle macchine, dal tipo di rifinizione.
Classificazione dei tessuti
A prescindere da alcune classificazioni commerciali e merceologiche, i t. possono normalmente essere classificati: a) in relazione alla tecnologia di fabbricazione: t. veri e propri e t. a maglia; b) in relazione all’altezza e all’assetto con cui si presentano: stretti, bassi, alti; a semplice o doppia altezza; avvolti in rullo o piegati a barchetta; in tutta altezza (t. veri e propri e maglie), tubici (t. e maglie), circolari (maglie) ecc.; a maglia diminuita o aumentata ecc.; c) in base alla materia prima: a seconda del tipo di fibra con cui si sono realizzati i filati (da fibre discontinue) o i fili continui impiegati, si hanno t. di lana, di cotone, di fibre liberiane (lino, canapa), di seta, di fibre minerali, di fibre chimiche, misti (nel caso siano presenti più fibre). I t. di lana a loro volta si distinguono in cardati o pettinati, a seconda del ciclo di lavorazione utilizzato per l’allestimento dei filati; i t. di cotone, oltre che in cardati e pettinati, si suddividono anche in radi e fitti, a seconda della massa e del tipo di armatura o di intreccio adottati; i t. di fibre chimiche, in rapidissimo sviluppo nel corso della seconda metà del 20° sec., in t. di fibre artificiali (da polimeri naturali) e di fibre sintetiche (da polimeri sintetici); d) in base alla destinazione: t. per abbigliamento, arredamento, per impieghi tecnici.
Caratteristiche dei tessuti
Innumerevoli possono essere le caratteristiche proprie di ciascun t., sia esso un t. vero e proprio o una maglia, che variano a seconda dell’impiego specifico a cui il manufatto è destinato. Le caratteristiche possono riguardare la struttura del t. (rado, fitto, compatto, leggero, pesante), la mano o sensazione al tatto e l’aspetto superficiale (mano cascante, sostenuta, morbida, ruvida ecc.; aspetto superficiale liscio, rasato, crespato, coperto, a pelo ecc.), la vestibilità e il drappeggio, l’attitudine alla confezione, la coibenza termica, le doti di minima o normale manutenzione, la resistenza a determinati agenti aggressivi (alle tarme, alle muffe, alla fiamma, alle macchie, alla luce, agli sfregamenti ripetuti, alle gualciture, allo strappo ecc.) e così via. I principali fattori che riescono a influenzare le caratteristiche di un t. sono: a) qualità e proprietà delle materie prime con cui si sono realizzati i filati (finezza e lunghezza per la lana; finezza, tiglio e pulizia per il cotone ecc.); b) vincoli e caratteristiche costruttive dei filati utilizzati (per es., filati di lana e cotone cardati o pettinati); c) vincoli e caratteristiche costruttive del t.; d) tipo di rifinizione cui viene sottoposto il t., inteso a modificare l’aspetto superficiale del t. e i legami intermolecolari delle fibre, allo scopo di conferire determinati effetti di mano, stabilità dimensionale, particolari proprietà antistatiche, di resistenza alla fiamma ecc. Rientrano nella varietà dei possibili cicli di rifinizione anche la scelta di una determinata tecnica di tintura o di una classe di coloranti compatibile con la fibra da tingere.
Sui t. è possibile inoltre compiere una serie di prove di laboratorio, intese ad accertare la corrispondenza della fornitura al campione, il rispetto di determinati capitolati, la presenza di particolari proprietà richieste dall’uso ecc. Le principali analisi eseguibili su t. sono: la determinazione delle caratteristiche dei filati impiegati, le prove di resistenza alle sollecitazioni fisiche (perforazione, lacerazione, pilling ecc.), le prove di resistenza al degrado e alle sollecitazioni chimiche (attacco di tarme e muffe, acqua, sfregamento ecc.), la verifica di determinate proprietà come l’idrorepellenza, l’oleorepellenza ecc.
È obbligatoria l’etichettatura dei t. comunque prodotti e commercializzati. L’etichettatura riguarda la composizione delle fibre impiegate, con le percentuali elencate in ordine decrescente.