
respirazione
Processo fisiologico, essenziale per gli esseri viventi aerobici (Uomo, animali e vegetali), che consiste nell’assunzione dell’ossigeno atmosferico e nell’eliminazione di anidride carbonica e acqua, per mezzo di apparati e organi diversi (polmoni, branchie, trachee e inoltre diffusione dei gas attraverso la superficie corporea).
La r. consiste essenzialmente in uno scambio gassoso tra gli organismi viventi e l’ambiente. In tale processo si possono distinguere due momenti fondamentali: la r. esterna, in cui l’ossigeno ambientale viene assunto secondo diversi meccanismi, in rapporto con il grado di complessità dell’apparato respiratorio, e la r. cellulare, consistente in un processo di ossidazione aerobica di materiali nutritizi, che si svolge all’interno delle strutture cellulari in presenza di ossigeno, con liberazione di energia e produzione di anidride carbonica.
Cenni storici
Anticamente la r. era identificata con la r. esterna, cioè con la ritmica immissione ed emissione di aria (ventilazione polmonare), cui le varie scuole mediche attribuivano diversi significati fisiologici. L’interpretazione della r. come fenomeno chimico in certo senso analogo alla combustione, con assunzione di ossigeno (O2) e liberazione di anidride carbonica (CO2), è dovuta alle ricerche di J. Priestley e di A. Lavoisier nella seconda metà del 18° secolo. Il matematico G.L. Lagrange (1791) fece osservare che il processo di combustione respiratoria non può avvenire nei polmoni – che non hanno una temperatura particolarmente elevata – ma nei vari organi. L. Spallanzani (1803) verificò sperimentalmente l’ipotesi di Lagrange. E.F. Hoppe-Seyler (1864) dimostrò che negli organi respiratori dei Vertebrati (branchie, polmoni) l’ossigeno si combina in modo reversibile con l’emoglobina dei globuli rossi del sangue. L’anidride carbonica emessa dai tessuti, invece, si combina con varie sostanze contenute nel sangue, da cui poi, a livello dei polmoni, viene emessa all’esterno. Nella maggior parte degli Invertebrati, l’ossigeno si lega ad altri pigmenti respiratori come l’emocianina, l’emeritrina e l’eritrocruorina.
Nelle piante il riconoscimento dei fenomeni respiratori è più difficile, perché mascherato dalla fotosintesi clorofilliana, in cui viene assorbita anidride carbonica ed emesso ossigeno. J. Ingenhousz (1779) osservò che le parti verdi delle piante emettono ossigeno in presenza di luce e anidride carbonica nell’oscurità. T. de Saussure (1804) riconobbe il significato dei due processi della fotosintesi e della r., la quale avviene anche, costantemente, nelle piante. Si stabilì così che la r., intesa come processo chimico, avviene in tutti gli esseri viventi. L. Pasteur scoprì poi che alcuni microrganismi possono vivere in condizioni anaerobie. Si poté in seguito dimostrare che anche in questi avvengono fenomeni di ossidazione analoghi alla r., senza l’intervento dell’ossigeno atmosferico (r. anaerobica ➔ fermentazione). Fenomeni di r. anaerobica si osservano anche in alcuni tessuti degli organismi pluricellulari (per es., nel tessuto muscolare).
R. esterna
Negli organismi monocellulari (Batteri, Protozoi) e in alcuni Metazoi di piccole dimensioni lo scambio dei gas respiratori con l’esterno avviene per diffusione e non presenta particolari difficoltà, dato che la superficie di rivestimento è relativamente grande rispetto alla piccola mole di questi organismi.
Nella maggior parte dei Metazoi invece, nei quali la mole del corpo è relativamente grande rispetto alla superficie di rivestimento (che, per di più, spesso è costituita in modo da non permettere la diffusione dei gas), e nei quali i processi metabolici sono molto più attivi, la semplice diffusione non basta ad assicurare l’apporto di una quantità sufficiente di ossigeno né la rapida eliminazione dell’anidride carbonica. Né questi organismi dispongono di depositi di ossigeno analoghi ai depositi di glicidi e di lipidi: si pensi, per es., che nei liquidi organici di un uomo sono presenti, in soluzione, circa 1000 cm3 di ossigeno, mentre il consumo di questo, che a riposo è di circa 300-400 cm3 al minuto, raggiunge facilmente i 3400 cm3 durante il lavoro muscolare. L’Uomo (e parecchi altri Metazoi) ha pertanto bisogno di attingere continuamente alla riserva di ossigeno – praticamente illimitata – costituita dall’atmosfera o (se si tratta di animali acquatici) all’ossigeno in soluzione nell’acqua. In questi casi lo scambio gassoso avviene nell’apparato respiratorio variamente differenziato nei diversi animali, che consiste fondamentalmente in una vasta superficie in corrispondenza della quale l’ambiente interno è in contatto con l’ambiente da cui viene attinto l’ossigeno: acqua per gli animali acquatici, aria atmosferica per gli animali terrestri.
Con meccanismi particolari (movimento della faringe e delle branchie nei Pesci, movimenti inspiratori ed espiratori nell’Uomo e negli altri Vertebrati terrestri ecc.) viene assicurato il continuo rinnovamento dell’aria (o dell’acqua) in contatto con la superficie respiratoria, mentre l’ossigeno assorbito viene rapidamente trasportato in tutti i tessuti dal sangue, che assicura contemporaneamente il trasporto dell’anidride carbonica dai tessuti all’apparato respiratorio.
La r. nell’Uomo
Fisiologia. La r. esterna nell’Uomo comprende essenzialmente fenomeni di natura meccanica che assicurano il rinnovo dell’aria nell’interno dei polmoni, e gli scambi respiratori che avvengono a livello degli alveoli polmonari (➔ respiratorio, apparato). Nelle persone di sesso maschile è prevalente l’allungamento del diametro verticale della gabbia toracica (r. diaframmatica), nella donna, in condizioni abituali, prevale l’allungamento dei due diametri orizzontali (r. costale). La frequenza del respiro – ovvero l’alternarsi ritmico e uniforme dei movimenti respiratori (eupnea) – in condizioni di riposo, nell’uomo adulto è di 15-20 atti respiratori al minuto; circa 45 nel neonato, 35 nel lattante, 25 nei bambini durante la prima infanzia. La sospensione degli atti respiratori è detta apnea, l’accelerazione polipnea, il rallentamento bradipnea; per designare genericamente una r. anormale si usa il termine di dispnea. In alcuni tipi di questa il ritmo della r. ha delle particolari evoluzioni cicliche (respiro periodico di Cheyne-Stokes, respiro di Kussmaul, respiro di Biot).
Gli scambi respiratori si giovano della straordinariamente grande superficie complessiva delle pareti degli alveoli polmonari, che è pari a 30 volte o, a seconda dei calcoli, a 45 volte quella corporea, essendo stata stimata rispettivamente in 60 o 90 m2. La parete degli alveoli è ricoperta da un esile velo liquido, nel quale i gas atmosferici si disciolgono; poi, superando la parete alveolare e l’endotelio dei suoi capillari, si diffondono verso il sangue legandosi rapidamente e reversibilmente all’emoglobina degli eritrociti: l’istituzione di tale legame permette al sangue di trasportare nei tessuti una quantità di ossigeno pari al 19% del suo volume e non semplicemente allo 0,36% come avverrebbe se si trattasse di una semplice soluzione; la rapidità con cui avviene la fissazione permette all’ossigeno di fissarsi all’emoblogina nel breve tempo (0,7 s) impiegato dalle emazie a percorrere i capillari alveolari; la reversibilità del legame consente all’ossigeno di staccarsi dall’emoglobina, quando la sua tensione nel plasma diminuisce per effetto del rilascio a livello dei tessuti. Questa reazione è facilitata dalla presenza in molti tessuti di sostanze analoghe all’emoglobina, ma dotate di un’affinità per l’ossigeno maggiore di questa, come la mioglobina del muscolo e l’emoglobina fetale.
Parallelamente alla saturazione di ossigeno, nel sangue si verifica un aumento di anidride carbonica che si è prodotta nel corso della r. interna. Il sangue può trasportare una quantità considerevole di anidride carbonica, in quanto solo una piccola parte di questo gas rimane in soluzione (sotto forma di acido carbonico) mentre il rimanente si combina con basi monovalenti (sodio e potassio) formando i rispettivi bicarbonati. A livello della parete degli alveoli polmonari avviene il processo inverso, in quanto l’acido carbonico (trasformandosi in anidride carbonica) si diffonde nell’aria alveolare, mentre dai bicarbonati si riforma l’acido carbonico. Il rapporto tra la quantità di acido carbonico e di bicarbonati presenti in soluzione nel plasma è influenzato in modo determinante dalla r., ed è di fondamentale importanza, in quanto costituisce uno dei principali sistemi tampone del sangue.
Controllo chimico della respirazione. Numerosi stimoli (fisiologici e patologici) sono in grado di modificare la ventilazione per adeguarla alle necessità dell’organismo nelle più diverse condizioni. Nella risposta ventilatoria all’alterato biochimismo dell’ambiente interno (ipossia, ipercapnia, acidosi) sono coinvolti due sistemi sensoriali: i chemiocettori periferici (glomi carotidei in particolare) capaci di rispondere rapidamente a modificazioni (soprattutto) della pressione parziale di ossigeno, PaO2 (ma anche della PaCO2 e della [H+]a, ossia concentrazione idrogenionica del sangue arterioso), e i chemiocettori centrali (situati sulla superficie ventro-laterale del bulbo), che rispondono più lentamente alle modificazioni della PaCO2 e della [H+] del liquor.
Regolazione della r. durante il sonno. La conoscenza dell’influenza che le diverse fasi del sonno (sonno lento non-REM e REM) esercitano sulla regolazione della r. è importante sia in condizioni normali sia patologiche. Il sonno deprime la ventilazione, la veglia la stimola. Le principali conseguenze clinico-funzionali sono rappresentate da: a) ipoventilazione alveolare, con conseguente sviluppo (in soggetti predisposti: pazienti pneumopatici cronici) di ipossiemia e ipercapnia; b) disritmie respiratorie (respiro periodico) indotte da ‘instabilità’ chemiorecettoriale; c) apnea ostruttiva quale conseguenza della perdita di tono della muscolatura (e conseguente ostruzione) delle vie aeree superiori, tipica dei soggetti forti russatori.
Regolazione del ritmo del respiro. La r. è sottoposta a un duplice sistema di controllo, neurochimico e volontario. Il primo, che è automatico e involontario, ha come principali obiettivi: l’omeostasi dei gas respiratori, vale a dire la possibilità di minimizzare le oscillazioni dei gas nel sangue arterioso, anche in presenza di notevoli variazioni di attività, altitudine, gravità; l’omeostasi dell’equilibrio acido-base, mediante gli effetti della ventilazione sulla PaCO2; la regolazione della frequenza respiratoria e del volume corrente, in modo tale da minimizzare lo sforzo e l’energia richiesti per la respirazione. Il controllo volontario (comportamentale), attuato attraverso componenti nervose situate in strutture sopramidollari e corticali, serve fondamentalmente funzioni diverse dallo scambio gassoso, per es., la fonazione.
Per il controllo della r. sono presenti tre elementi: a) unità di controllo centrale (centri respiratori localizzati nel tronco encefalico, bulbo e ponte), che integrano le informazioni provenienti dalla periferia e attivano le vie motrici regolando i movimenti respiratori; b) un sistema di sensori composti da: 1) chemiocettori di tipo periferico (glomi carotidei, sensibili soprattutto alla variazione della PaO2) e di tipo centrale (aree chemiosensitive della superficie ventro-laterale del bulbo), sensibili soprattutto alle variazioni della concentrazione [H+] del liquor indotte da variazioni della PaCO2; 2) meccanocettori, costituiti da fibre vagali mielinizzate (recettori a lento adattamento, situati tra le fibre muscolari lisce delle vie aeree, e a rapido adattamento, situati tra le cellule epiteliali delle vie aeree, entrambi sensibili alla espansione polmonare e, i secondi, anche a stimoli irritativi endogeni ed esogeni); 3) fibre vagali amieliniche (terminazioni delle fibre C o recettori J, situati, in prossimità dei vasi, nell’interstizio polmonare); 4) fusi neuro-muscolari della muscolatura respiratoria; c) un sistema di effettori (muscoli respiratori, la sola muscolatura scheletrica che non conosce sosta, dalla nascita alla morte), la cui attività produce ventilazione.
Il sistema può essere altresì immaginato come composto da due sezioni: una sottoposta al controllo (depositi gassosi corporei – sistema circolatorio – mantice meccanico), e una deputata al controllo (chemiocettori – meccanocettori – centri respiratori). Esso opera come sistema di controllo a feedback negativo: cioè, da una parte, aumenta la flessibilità e l’accuratezza della regolazione ma introduce anche, a motivo della complessità dei meccanismi interessati, un possibile conflitto di informazioni (e conseguente instabilità) nella regolazione stessa. Ne sono esempi alcuni tipi di respiro patologico. Resta irrisolto il problema della ritmogenesi respiratoria: se sia generata da gruppi di neuroni dotati di attività pacemaker (attività ritmica propria) oppure da una attività di network (interazione di una rete di neuroni di per sé privi di attività ritmica).
R. e apparato di fonazione. Una delle condizioni necessarie perché l’Uomo possa parlare è la possibilità di modificare lo stato di equilibrio della pressione atmosferica all’interno dell’apparato di fonazione (polmoni, trachea e cavità epiglottidali). La maggior parte dell’aria sottoposta a modificazioni di pressione è aria polmonare, cioè respiratoria (o anche pneumonica).
La fonazione. Gran parte delle lingue utilizza solo aria polmonare per le articolazioni dei propri fonemi. Pochissimi sono i tipi di articolazioni apneumonici , nei quali cioè non interviene aria respiratoria; tra i più importanti: articolazioni avulsive, o clic, in cui l’aria modificata è soltanto quella contenuta in una porzione più o meno grande della cavità orale; articolazioni recursive (o glottidalizzate), che richiedendo una perfetta chiusura della glottide fanno uso dell’aria compressa o rarefatta nella cavità orofaringea.
È invece pneumonica la maggior parte delle altre articolazioni possibili (per es., la totalità delle articolazioni italiane), dove la contrazione o la dilatazione di una cavità dell’apparato di fonazione, quando sia chiusa ogni via di comunicazione dell’interno dell’apparato di fonazione con l’esterno, provocano rispettivamente un aumento o una diminuzione della pressione dell’aria contenuta all’interno dell’apparato; se invece vi è almeno una via di comunicazione aperta, nel primo caso si creerà una corrente d’aria egressiva, nel secondo caso ingressiva. Se questa via di comunicazione si apre dopo che si è creata una compressione o una rarefazione, si avrà un’esplosione o un’implosione. Per es., l’articolazione normale dei fonemi italiani richiede aria polmonare egressiva continua (nelle vocali, semivocali, fricative, vibranti, laterali, nasali) o compressa (nelle occlusive esplosive). L’esame sperimentale della r. fonatoria si esegue con diversi apparecchi: pneumografi, per registrare e misurare i movimenti delle varie parti della cassa toracica; volumografi, per misurare la quantità d’aria inspirata o espirata ecc.
Il volume relativo dell’aria espirata aumenta se aumenta l’intensità articolatoria, diminuisce se aumenta la frequenza delle vibrazioni laringee, o se aumenta la durata dell’articolazione; il volume relativo aumenta progressivamente se si passa dalle vocali aperte alle chiuse, alle consonanti fricative, e alle esplosive; aumenta se si passa dalle articolazioni sonore alle sorde, dalle esplosive alle aspirate ecc.
R. artificiale. Tecnica con cui si sollecita la ripresa dei movimenti respiratori, che si pratica come intervento d'urgenza per ripristinare una sufficiente ventilazione polmonare in caso di arresto o di grave indebolimento degli atti respiratori.
R. cellulare o dei tessuti
Consiste nel complesso di reazioni che si svolgono nell’interno delle cellule, mediante le quali l’ossigeno viene utilizzato per l’ossidazione dei metaboliti cellulari con conseguente produzione di CO2 e formazione di composti ad alta energia.
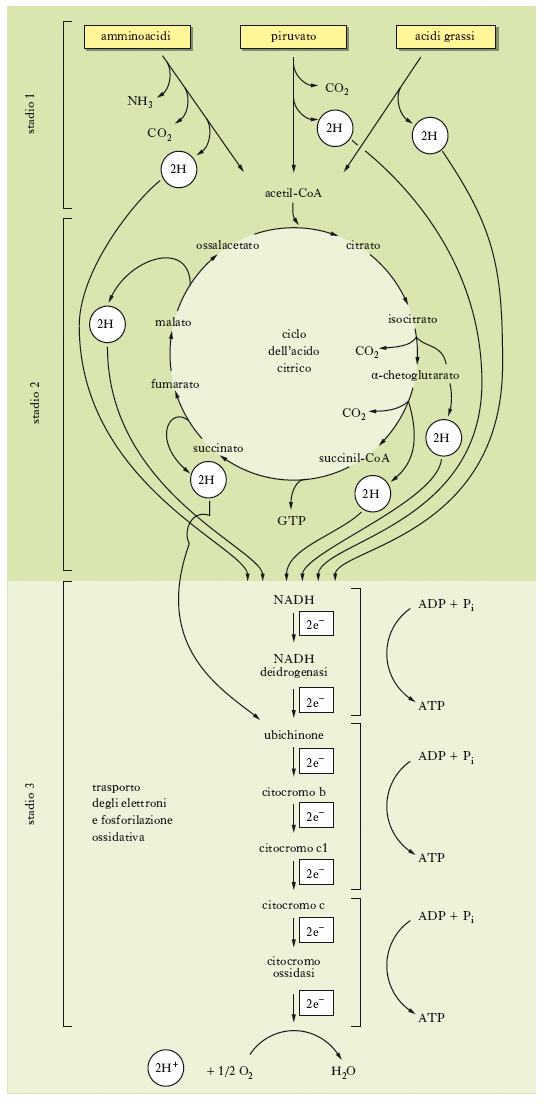
La r. cellulare è un processo articolato in tre fasi principali (v. fig.). Nella prima, le molecole organiche da utilizzare come substrati ‘combustibili’, cioè carboidrati, acidi grassi e alcuni amminoacidi, subiscono processi enzimatici di ossidazione che danno origine ad atomi di idrogeno e a composti a due atomi di carbonio, rappresentati dai gruppi acetilici dell’acetilCoA. I cicli metabolici coinvolti nella prima fase sono la glicolisi e la β-ossidazione dei grassi. Nella seconda tappa, i gruppi acetilici, trasportati dall’acetilCoA, sono degradati enzimaticamente per generare atomi di idrogeno e CO2, che è il prodotto finale di scarto dell’ossidazione dei substrati metabolici organici. In questo caso, il ciclo metabolico coinvolto è il ciclo dell’acido citrico. Nella terza fase della r. cellulare, gli atomi di idrogeno prodotti nella tappa precedente sono trasportati dai coenzimi ridotti, sia nicotinici (NADH) sia flavinici (FADH2), all’interno dei mitocondri. Qui, attraverso una sequenza ordinata di reazioni enzimatiche di ossidoriduzione, viene effettuato un trasferimento di elettroni grazie all’intervento di una serie di molecole accettrici, l’ultima delle quali è rappresentata dall’ossigeno molecolare che viene ridotto ad acqua. Questa sequenza di reazioni, nota come catena respiratoria o catena del trasporto degli elettroni, è accoppiata alla produzione di energia nel processo noto come fosforilazione ossidativa (➔ mitocondrio).
Tutte le reazioni coinvolte nelle varie tappe della r. cellulare sono catalizzate da enzimi specifici, alcuni dei quali, attraverso la modulazione della loro attività catalitica da parte di piccole molecole effettrici (➔ effettore), regolano la velocità della fase in cui agiscono. Questo significa che la r. cellulare, poiché è accoppiata alla produzione di substrati ad alta energia sotto forma di ATP, è regolata dalle richieste energetiche della cellula stessa, del tessuto cui la cellula appartiene o, in ultimo, dell’intero organismo. Stimoli ormonali, concentrazione cellulare di ATP, richiesta di lavoro muscolare, concentrazione di substrati metabolizzabili, sono tra i principali fattori responsabili della regolazione della r. cellulare.
Nella r. cellulare l’ossigeno non agisce direttamente nell’ossidazione dei substrati che si verifica nelle varie fasi: quest’ultimi sono, infatti, ossidati in tappe enzimatiche successive, nelle quali spesso vengono coinvolti enzimi del gruppo delle deidrogenasi. Un ruolo fondamentale è anche svolto dalle molecole trasportatrici di elettroni della catena respiratoria, il cui componente finale, l’emoproteina enzimatica citocromoossidasi, è responsabile della reazione terminale della r. cellulare che provoca la riduzione dell’ossigeno molecolare ad acqua.
R. nelle piante
Aerobiosi e anaerobiosi. Il processo respiratorio si svolge nelle piante con gli stessi processi chimici descritti per gli animali. Poiché gli organismi vegetali sono comparsi prima degli organismi animali, l’affermarsi del processo respiratorio si può considerare come un adattamento degli organismi alla disponibilità di ossigeno libero prodotto dalle piante con la fotosintesi. L’atmosfera primitiva era costituita essenzialmente di metano, ammoniaca e vapore acqueo e non conteneva ossigeno libero. Le prime cellule e i più antichi organismi avevano forme di anaerobiosi (fermentazioni) proprie anche di parecchi organismi attuali, che vivono su substrati in cui la degradazione di zuccheri, lipidi e proteine si svolge in deficienza o in assenza di ossigeno. Ciò accade in molti Batteri e Funghi; è da considerare che le cellule a vita aerobia di tutti gli altri organismi, oltre alla fase glicolitica anaerobica, possono ricorrere a forme di anaerobiosi, quando condizioni particolari di superlavoro ostacolino l’adeguato rifornimento di ossigeno e rendano impossibile la degradazione completa del materiale respiratorio a CO2 e H2O, e la liberazione di tutta l’energia di legame che esso contiene in origine.
Il processo respiratorio. Mentre la r. aerobica, nel suo insieme (glicolisi, formazione di acetilcoenzima A, ciclo di Krebs e catena respiratoria) può considerarsi come la reazione inversa di quella globale della fotosintesi, cioè
(di questa variazione di energia ΔG circa il 40% viene recuperato in forma chimica utile alla cellula sotto forma di circa 38 molecole di ATP), le reazioni globali delle fermentazioni hanno un rendimento più basso; per es., per la fermentazione alcolica
e per la fermentazione lattica
Accanto agli organismi aerobi, nei quali si svolge la r. completa, diversi organismi vegetali possono respirare o fermentare, a seconda delle condizioni ambientali in cui si trovano (organismi aerobi facoltativi); altri sono, invece, anaerobi obbligati. Molti organismi, specialmente vegetali, hanno una via anaerobica alternativa per l’ossidazione del glucosio, chiamata via dei pentosofosfati (➔ pentoso). Dal punto di vista della r., la via dei pentosofosfati ha circa la stessa efficienza della glicolisi associata al ciclo di Krebs. Questa via alternativa è presente in molte piante, nelle quali si ha un’abbondante formazione di glucosio-6-fosfato; essa è collegata anche al ciclo di Calvin, che serve come fonte di pentosi per molte attività cellulari e per la continua formazione del ribulosio-1,5-difosfato, necessario nella fotosintesi, in quanto accettore della CO2.
Fotorespirazione. Da tempo si era sospettato che l’accelerazione della r. normale (processo oscuro) potesse essere influenzata dalla luce (fotorespirazione). La fotorespirazione è un processo diverso dalla r. oscura, e dal punto di vista energetico essa appare un processo dissipativo. Con luce molto intensa, la fotosintesi è limitata dalla velocità di diffusione di CO2 e in certi casi dall’attività degli enzimi carbossilanti. Nel mais, nel sorgo, nella canna da zucchero, la velocità della fotosintesi, con luce intensa, è 2-3 volte maggiore che nella barbabietola da zucchero, nel tabacco e nel grano. Le piante del primo gruppo sono capaci di abbassare il livello di CO2 esterna a un punto di compensazione prossimo a zero, pari a meno di 5 ppm (parti per milione); quelle del secondo gruppo invece fino a circa 50 ppm. Ciò significa che le piante del primo gruppo riescono a mantenere un’alta velocità di diffusione di CO2 e perciò hanno una fotosintesi molto intensa; in quelle del secondo gruppo, che non possono raggiungere un basso punto di compensazione, vi è la fotorespirazione attivata dalla luce che è biochimicamente distinta dalla r. normale. Queste reazioni tendono anche ad abbassare la concentrazione di ossigeno in prossimità dei cloroplasti e forse a proteggerli da processi fotossidativi. Con α-idrossisolfonato è possibile inibire la fotorespirazione, e allora la velocità della fotosintesi in alta intensità luminosa si approssima a quella delle piante del primo gruppo.
Fattori influenzanti la respirazione. Si devono annoverare la temperatura, la disponibilità di ossigeno e di sali minerali, gli stimoli meccanici e, infine, le ferite. Le rotture di tessuti superficiali stimolano fortemente la r. e la formazione di tessuti meristematici cicatriziali nell’area della ferita, per determinare la formazione del callo da ferita. Probabilmente, l’aumentata intensità respiratoria sulle superfici di ferita dipende dalla mobilizzazione di alte quantità di substrati respiratori. Le radici delle piante possono essere asfissiate nel suolo in cui crescono se troppo fitte, o se sommerse nell’acqua poco ossigenata, come nelle zone paludose delle regioni calde, ove si ha la vegetazione di miriadi di organismi acquatici, che impoveriscono le acque di ossigeno.
Strutture. Nelle piante la r. esterna avviene per diffusione dei gas attraverso la superficie del corpo nelle piante tallofite, attraverso gli stomi e le lenticelle nelle cormofite; l’aria (e ossigeno) da queste aperture si diffonde negli spazi intercellulari e di qui passa nelle singole cellule; il cammino inverso è compiuto dall’anidride carbonica; in certi organi esposti ad asfissia (radici immerse nella fanghiglia di acque ferme ecc.) si sviluppano canali aeriferi più o meno estesi, che servono da serbatoi di aria, in altri casi le radici formano pneumatofori.
Anche nelle piante, in seguito alla r. si ha produzione di calore, la cui entità è però molto meno vistosa che negli animali superiori, tanto più che gran parte del calore prodotto si disperde celermente attraverso la grande superficie delle lamine fogliari; tuttavia in certi casi si può constatare un aumento notevole della temperatura (➔ termogenesi). In generale una pianta superiore verde emette nell’atmosfera, in 24 ore, da 5 a 10 volte il proprio volume di CO2, equivalente a 1/5-1/3 della quantità di CO2 che essa ha fissato per fotosintesi durante la giornata. Il quoziente respiratorio nelle piante è di norma eguale a 1, però può anche essere inferiore, se il materiale respiratorio è un lipide o una proteina.