
glicolisi
Processo di degradazione anaerobica del glucosio ad acido lattico (detto anche ciclo di Embden-Meyerhof), caratteristicamente legato alla produzione di energia sotto forma di ATP. Questo tipo di demolizione del glucosio si realizza in molti microrganismi e nelle cellule della maggior parte degli animali superiori. Nella g. si ha un’ossidoriduzione interna, per cui lo stato di ossidazione del prodotto è lo stesso di quello del substrato. L’equazione generale bilanciata della g. è data da: C6H12O6+2Pi+2ADP→
→2CH3CHOHCOOH+2ATP+2H2O
avendo indicato con Pi i gruppi fosfato inorganico che entrano nella sequenza di reazioni. L’analisi del bilancio energetico della g. comprende l’esame di due distinti processi che avvengono contemporaneamente: quello esoergonico di degradazione del glucosio ad acido lattico e quello endoergonico di accumulo di energia sotto forma di ATP. Le variazioni di energia libera standard indicano che la demolizione del carboidrato fornisce una quantità di energia superiore a quella richiesta dalla fosforilazione di due molecole di ADP ad ATP; in particolare, questo eccesso ammonta al 31% circa dell’energia totale posta in gioco e che viene quindi immagazzinata sotto forma di legame fosforico. L’intera catena della g. procede perciò con una diminuzione netta molto elevata di energia libera, pari a −116 kJ/mol: si tratta quindi di un fenomeno altamente irreversibile anche se alcune delle sue tappe a minor decremento energetico vengono spesso adoperate dai sistemi cellulari nella biosintesi del glucosio a partire da acido lattico o altri precursori. La g. si svolge attraverso l’azione sequenziale di un gruppo di 11 enzimi localizzati nel citoplasma e non associati tra loro a formare un complesso multienzimatico stabile. Alcuni risultati sperimentali hanno mostrato però che esistono enzimi della catena che tendono, sia pur debolmente, ad associarsi alla membrana plasmatica, alle miofibrille, ai mitocondri. I composti intermedi tra glucosio e acido piruvico sono fosforilati; i gruppi fosforici assumono tre funzioni: quella di fornire a ciascun intermedio un gruppo polare caricato negativamente che impedisce l’evasione dalla cellula attraverso la membrana, quella di fungere da gruppo di legame o riconoscimento nella formazione di complessi enzima-substrato e, infine, quella legata al processo di accumulo di energia. I gruppi fosforici sono infatti destinati a diventare i gruppi terminali dell’ATP neoformato.
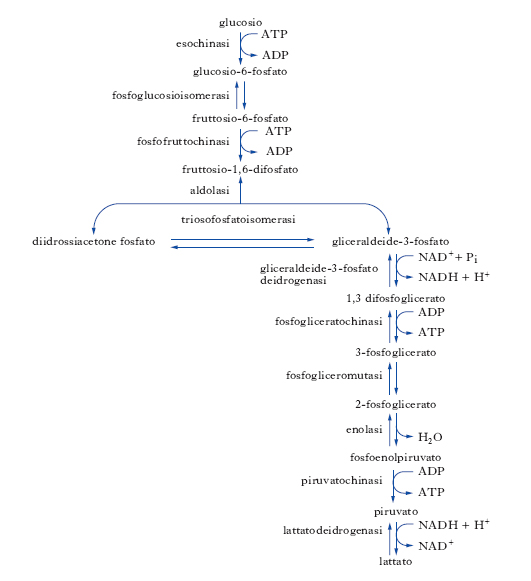
La g. anaerobica si svolge in due fasi, nella prima delle quali il glucosio viene preparato alla sua degradazione da reazioni fosforilanti e quindi scisso per formare la gliceraldeide-3-fosfato. In questa fase preparatoria, inoltre, alcuni esosi diversi dal glucosio possono, sia pure con diversa (minore) affinità, entrare nella catena glicolitica ed essere convertiti nel prodotto comune; in ordine a queste trasformazioni vengono consumate due molecole di ATP per produrre la fosforilazione delle posizioni 1 e 6 dell’esoso trasformato (fruttosio-1,6-difosfato), costituendo l’innesco alla fase successiva. Nella seconda fase della g. avviene la conversione della gliceraldeide-3-fosfato in acido lattico attraverso la serie di ossido-riduzioni che consentono al sistema di immagazzinare energia sotto forma di ATP; in questa porzione del processo, comune a tutti gli zuccheri ossidati, si formano infatti quattro molecole di ATP che provocano una resa netta pari a due molecole di ATP ottenute per molecola di glucosio degradata ad acido lattico. Durante la g. avvengono pertanto tre diversi tipi di trasformazioni chimiche: una sequenza di reazioni degradative della molecola dell’esoso con formazione di acido lattico, la trasformazione di ADP in ATP e una sequenza di reazioni ossidoriduttive (v. fig.). I passaggi più rilevanti ai fini energetici sono la fosforilazione del glucosio a glucosio-6-fosfato, la fosforilazione del fruttosio-6-fosfato a fruttosio-1,6difosfato e la defosforilazione dell’acido fosfoenolpiruvico ad acido piruvico. Nelle cellule aerobie, in condizioni di aerobiosi, la degradazione del glucosio attraverso la g. si arresta ad acido piruvico. Questo composto, che successivamente è completamente degradato a CO2 e H2O attraverso il ciclo di Krebs e la fosforilazione ossidativa (per aumentare la resa energetica della degradazione del glucosio), rappresenta il punto di unione tra metabolismo anaerobio e aerobio dei carboidrati semplici. Nell’uomo, principalmente a carico degli eritrociti, è nota una serie di enzimopatie ereditarie a carico degli enzimi della g. le quali provocano, genericamente, emolisi, cambiamento della forma dei globuli rossi, splenomegalia ecc.
La glicoregolazione è il controllo agente sulle vie del metabolismo glicidico nel passaggio da glucosio ad acido piruvico e viceversa. La g. è regolata dalla fosfofruttochinasi, un enzima allosterico stimolato dalla presenza di fruttosio-1,6-difosfato, fruttosio-2,6-difosfato, AMP e ADP, ma inibito dall’ATP, da acido citrico e dal NADPH; il secondo livello di regolazione avviene durante il passaggio catalizzato dall’esochinasi, controllato dalla concentrazione di glucosio-6-fosfato, dall’acetilcoenzima A e dal fosfoenolpiruvato. Anche la piruvatochinasi regola la g. poiché inibita da ATP, NADH e alanina e stimolata dal fruttosio-1,6-difosfato e glucosio-6-fosfato. Il ciclo anabolico è realizzato dalla cellula per la sintesi di glucosio (➔ gluconeogenesi). In questo come negli altri casi analoghi è la prima reazione della via biosintetica a essere sottoposta a regolazione.