
Ominidi
Famiglia zoologica dell’ordine dei Primati che comprende la specie umana attuale, le specie umane estinte, note attraverso resti fossili, e le grandi scimmie. Il complesso di processi evolutivi che per serie successive di modificazioni, morfologiche e strutturali, ha condotto all’uomo attuale è detto ominazione.
Sistematica
Gli Ominidi costituiscono una famiglia (Hominidae) che comprende oltre all’uomo (Homo sapiens) le scimmie antropomorfe africane Gorilla gorilla e Pan con due specie, Pan troglodytes, il comune scimpanzé e Pan paniscus, il bonobo o scimpanzé pigmeo. La famiglia degli O. fa parte della superfamiglia degli Hominoidea – insieme alla famiglia dei Pongidi, comprendente il Pongo pygmaeus (l’orango asiatico) –, e a quella degli Ilobatidi, con il genere Hylobates, con più specie riferibili ai gibboni e al siamango, confinati nell’Asia insulare di Sud-est.
Nel Miocene inferiore gli Ominoidea si sono separati per divergenza evolutiva dai Cercopithecoidea, l’altra superfamiglia delle scimmie catarrine del Vecchio Mondo caratterizzate dalla coda. Esse, con le scimmie platirrine del Nuovo Mondo, appartengono al subordine degli Anthropoidea che insieme a quello dei Prosimii costituiscono l’ordine dei Primati.
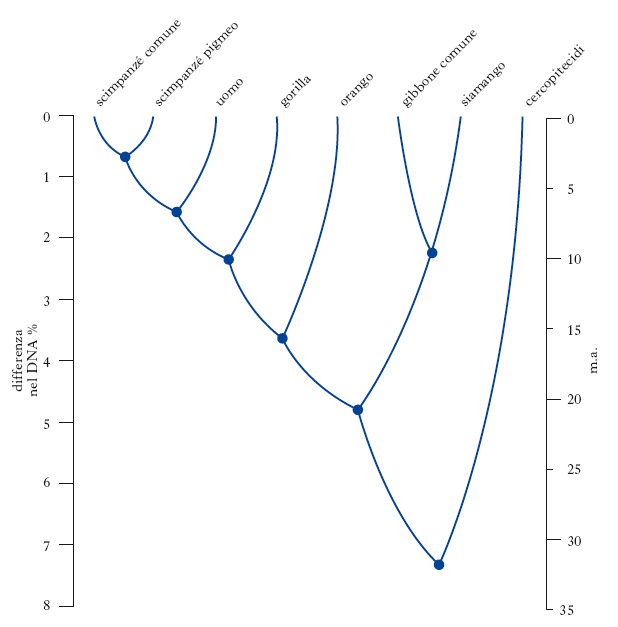
In passato, in base alle omologie strutturali (presenza in 2 o più specie di caratteri derivati da un antenato comune) gorilla e scimpanzé erano inclusi nella famiglia dei Pongidi insieme all’orango; l’uso del termine O. era limitato all’uomo e a quegli antenati fossili nei quali la testimonianza anatomica dell’acquisizione della postura eretta e del bipedismo marcava gli inizi della linea evolutiva umana. Gli studi di genetica sviluppati dall’antropologia molecolare a partire dagli anni 1960 hanno dimostrato l’affinità genetica di scimpanzé e gorilla con l’uomo, i quali sono quindi entrati di diritto nella famiglia (fig. 1). Pertanto, se la morfologia adattativa legata all’ambiente di foresta accomuna il gorilla e lo scimpanzé all’orango, dal punto di vista della parentela filetica le due antropomorfe sono più affini all’uomo che non all’orango. In particolare, lo scimpanzé presenta più del 98% delle sequenze nucleotidiche uguali a quelle dell’uomo a dimostrazione che la divergenza evolutiva delle due specie si è verificata tra 7 e 5 milioni di anni. Al momento comunque è ancora impossibile stabilire quale tra le tre linee gorilla-scimpanzé-uomo si è separata per prima, poiché gli eventi di speciazione che hanno portato alla loro separazione in termini evolutivi sono talmente ravvicinati (2-3 milioni di anni) che non si è in grado di identificare le tre specie in modo univoco in base alle mutazioni derivate dall’ultimo antenato comune. Inoltre il genoma umano ha una struttura a mosaico nella quale ciascuna regione cromosomica ha la sua specifica relazione con quella di altre specie a confronto. In base al confronto delle sequenze di una regione cromosomica del genoma umano si verifica, per es., che gorilla e uomo risultano identici, mentre rispetto alle sequenze presenti in un’altra regione è lo scimpanzé che risulta più simile all’uomo. Nel complesso tuttavia la maggiore similitudine genetica dell’uomo allo scimpanzé indica che nel gruppo monofiletico il gorilla si sarebbe separato un po’ prima degli altri. Oggi c’è un generale accordo nel sistemare l’uomo e i suoi antenati fossili bipedi nella sottofamiglia Ominini separata da quella che si riferisce a gorilla e scimpanzé. Secondo alcuni studiosi, alla famiglia degli O. andrebbe aggregato anche il Pongo che in questo caso andrebbe raggruppato con gorilla e scimpanzé nella sottofamiglia dei Pongini.
Le scimmie antropomorfe
In generale nelle sequenze del DNA i geni che ne costituiscono le unità fisiche funzionali presentano regioni codificanti le proteine, i cosiddetti mattoni che formano l’impalcatura di base dell’organismo, affiancate da regioni regolatrici (‘sequenze promotrici’) che ne controllano l’espressione fenotipica. Queste impartiscono le istruzioni sui tempi e modi con cui le sequenze strutturali devono assemblare il materiale di base durante il ciclo ontogenetico. Nel gorilla e nello scimpanzé le strutture anatomiche di base sono uguali a quelle dell’uomo, ma i tempi di regolazione per lo sviluppo dell’impalcatura anatomica sono accelerati perché le strutture rispondano fin dalla nascita con assetti specializzati che adattino l’individuo a sopravvivere nella specifica nicchia ecologica della specie, la foresta equatoriale. Pertanto la diversità fisica rispetto all’uomo riguarda le specializzazioni anatomiche legate alla vita arboricola, la locomozione, la dieta e i comportamenti sociali.
Per arrampicarsi e volteggiare tra i rami vengono utilizzati i quattro arti con l’arto toracico molto più lungo dell’arto pelvico e l’articolazione scapolo-omerale molto mobile; mano e piede sono prensili con pollice e alluce opponibili (l’alluce nel feto è allineato con le altre dita ma alla nascita esso è divergente); nel piede manca l’arco plantare e il calcagno è ridotto; nel cranio il foro occipitale è posteriore rispetto alla linea mediana; la colonna vertebrale è priva di curve, le ali dell’ileo sono alte e strette; la postura a terra è clinograda e per gli spostamenti viene adottata la cosiddetta knuckle-walking, un tipo di locomozione basato sull’appoggio a terra del dorso della mano ripiegata; nel cranio la faccia (splancnocranio) è molto più sviluppata del neurocranio in relazione al grande sviluppo dell’apparato masticatorio con molari e premolari grandi, specializzati per una dieta essenzialmente frugivora, foglivora e granivora; i canini sono molto sviluppati, più nel maschio che nella femmina, e richiedono uno spazio (diastema) nell’arcata superiore e inferiore per l’alloggio di ogni canino. La capacità cranica è pari a circa 380 ml nello scimpanzé maschio e 530 ml nel gorilla maschio, in rapporto alla sua maggior mole corporea.
Per la comunicazione sociale le antropomorfe sfruttano principalmente la vista e il tatto: la gestualità e la mimica facciale sono ben sviluppate e il contatto fisico (grooming) cementa i legami interindividuali secondo regole stabilite. Le antropomorfe non hanno la capacità di comunicare attraverso la parola poiché nella laringe il loro organo della fonazione non è adatto all’emissione di suoni articolati e nel cervello manca un centro nervoso (centro di Broca) necessario per la motilità dell’organo. Il ciclo vitale si esaurisce tra 40 e 50 anni e il periodo delle cure parentali si completa tra gli 8 e i 10 anni.
L’uomo
Tra gli O. l’uomo è una specie generalista che ha acquisito la postura ortograda e per spostarsi utilizza solo gli arti pelvici (deambulazione bipede e andatura ‘a passi alterni’). L’apparato masticatorio è ridotto con denti iugali (molari e premolari) che non sono specializzati (la dieta è onnivora), il canino piccolo e senza diastema non presenta dimorfismo sessuale e, a differenza delle antropomorfe africane, lo smalto dei denti è spesso. Il volume cerebrale valutato rispetto alla mole corporea raggiunge in media 1350 ml ma è soprattutto la superficie del cervello (neopallio) che si è straordinariamente sviluppata. L’estensione della rete interneuronica esalta le attività intellettive ed è questa l’unica vera specializzazione dell’uomo che gli ha permesso di diffondersi su tutta la terra.
Nel corso dell’evoluzione l’acquisizione di tutti questi caratteri ha comportato una serie di modificazioni anatomiche preannunciatesi al momento della divergenza dalle antropomorfe. La verticalizzazione del tronco ha richiesto in primo luogo il perfetto bilanciamento del cranio sulla colonna vertebrale ottenuto con lo scivolamento in avanti della base cranica e l’espansione della squama dell’occipitale; conseguentemente il foro occipitale si è allineato con la colonna vertebrale, nella quale si alternano curvature a concavità ventrale (cifosi) e dorsale (lordosi) che con il gioco muscolare permettono di bilanciare il peso corporeo mantenendo il busto eretto. Il bacino osseo è basso e largo con le ali dell’ileo espanse lateralmente per contenere i visceri; il peso del corpo si scarica attraverso l’articolazione ileo-femorale (acetabolo) lungo l’arto pelvico di destra e di sinistra. Il femore è lungo e molto robusto e i muscoli glutei sono ben sviluppati per sostenere il busto e per imprimere la spinta in avanti della gamba durante il movimento. Nel piede l’alluce non è opponibile, le dita si sono raccorciate e allineate e si è formato un arco longitudinale per cui l’appoggio plantare è su tre punti (tripode plantare): calcagno (che è grande e molto robusto), metatarso, estremità dell’alluce. L’arto toracico è più corto dell’arto pelvico e nella mano l’acquisizione della piena opponibilità del pollice permette la presa di precisione, cioè la possibilità di assemblare piccolissimi oggetti, di scrivere, cucire, afferrando lo strumento di lavoro tra il pollice, che si è allungato, e l’estremità dell’indice o di più dita. Il ciclo vitale si esaurisce intorno a 70-80 anni.
Il periodo delle cure parentali si protrae fino all’età giovanile rendendo possibile un lungo apprendimento grazie al progressivo e straordinario sviluppo dell’arborizzazione interneuronica.
Le teorie evoluzionistiche
Si riteneva un tempo che l’evoluzione dell’uomo avesse proceduto per così dire ‘dalla testa ai piedi’: un fossile, per poter essere considerato come antenato nella linea dell’uomo, doveva essere, per prima cosa, dotato di una grande capacità cranica a garanzia di un cervello ‘intelligente’; gli altri caratteri anatomici erano di secondaria importanza. Questa visione era legata alla concezione antropocentrica secondo la quale l’emergenza di ‘qualità’ intellettive specificamente umane ponevano l’uomo di diritto in una posizione dominante la natura. Già le prime indagini paleoantropologiche della seconda metà dell’Ottocento – ma soprattutto quelle numerosissime del Novecento, lette alla luce della teoria darwiniana – hanno evidenziato che il primo segnale nel processo di ominazione è stato invece l’acquisizione della postura eretta e della locomozione bipede, mentre il processo di encefalizzazione ha comportato tempi assai più lunghi per completarsi. La suddivisione geografica e cronologica dei primi Ominini africani mette in evidenza la grande variabilità di forme che talvolta si sono succedute le une alle altre, ma che in gran parte hanno convissuto anche nello stesso territorio in un arco di tempo che va da 5 a circa 1,5 milioni di anni (Plio-Pleistocene) quando si sono definitivamente estinte. La ricchezza di generi (Orrorin, Ardipithecus, Kenianthropus, Australopithecus, Paranthropus) con più specie testimonia che l’evoluzione della linea degli Ominini fin dalla sua origine non è stata frutto di una progressione evolutiva continua di una forma nell’altra. Più che di un albero, si parla infatti di un cespuglio di specie in ciascuna delle quali è manifesto un mosaico di caratteri: la sperimentazione di ‘novità evolutive’ si accompagna spesso a caratteri arcaici.
L’evoluzione degli O. durante il Miocene
Il primo rappresentante della superfamiglia degli Hominoidea è Proconsul africanus (Kenya, Tanzania, circa 23 milioni di anni), dallo scheletro postcraniale generalista con tronco corto e segmenti degli arti simili a quelli di una scimmia quadrupede mentre il cranio e la dentatura sono simili a quelli degli attuali Ominoidea. Secondo alcuni esso avrebbe costituito il punto di avvio nell’evoluzione della linea dello scimpanzé, ma in generale esso viene piuttosto considerato un esempio di evoluzione a mosaico.
Nel Miocene medio la linea dei gibboni si è separata dal tronco comune ominoideo; al momento appare comunque difficile rintracciare nel materiale fossile i caratteri specializzati dell’attuale genere Hylobates (estrema lunghezza degli arti toracici adatti alla brachiazione, mano con lunghe dita ricurve).
Nel Miocene superiore i caratteri dello splancnocranio dei resti attribuiti ai generi Sivapithecus e Lufenpithecus, rinvenuti in India e Pakistan, fanno di questi i probabili antenati dell’orango. In Eurasia i resti attribuiti ai generi Driopithecus (13-8 milioni di anni) e Graecopithecus sarebbero piuttosto collegabili alla linea più generalista degli Ominini.
Il percorso evolutivo del gorilla e dello scimpanzé è scarsamente ricostruibile a causa dell’acidità del suolo di foresta che non ha permesso la buona fossilizzazione dei resti scheletrici. Tuttavia, nei resti di un mascellare rinvenuti in Kenya e attribuiti a un nuovo genere, Samburupithecus, la dentatura presen;ta caratteri morfologici molto simili a quelli del gorilla. La datazione stimata in 9,5 milioni di anni verrebbe a conferma del dato neontologico basato sul confronto delle sequenze del DNA: il gorilla si sarebbe separato dalla linea dell’uomo poco prima dello scimpanzé. In Kenya, nello stesso sito in cui sono stati rinvenuti resti attribuiti al genere Homo, sono stati individuati denti con smalto sottile appartenenti a due individui del genere Pan; la datazione pari a circa 0,5 milioni di anni indicherebbe che fin dal tempo della loro divergenza le due linee hanno convissuto (speciazione simpatrica).
Alla fine del Miocene in Africa, a causa del progressivo raffreddamento climatico, il manto forestale si ridusse; quando si verificò la frattura tettonica dal Corno d’Africa fino all’Africa meridionale, si formò un lungo corridoio (la Rift Valley) tra le regioni occidentali dominate dalla foresta equatoriale e quelle orientali dominio della savana. Questi mutamenti avrebbero favorito l’evoluzione di forme arboricole che, pur ancora abili ad arrampicarsi e a spostarsi sugli alberi, dovettero parzialmente modificare i loro comportamenti, abituandosi a passare parte della giornata nella savana alla ricerca di nuove fonti di cibo, raccogliendo semi, bacche, radici. È nella foresta che sono vissuti i primi Ominini che hanno sperimentato il bipedismo, ma è nelle savane orientali e meridionali che essi si sono evoluti abbandonando progressivamente il modello di vita imposto dalla foresta.
L’evoluzione degli O. durante il Plio-Pleistocene
I primi Ominini (fig. 2). - Fa discutere il reperto rinvenuto nel 2002 nel bacino del Ciad di un piccolo cranio scimmiesco, ma con canini piccoli e mancanza di diastema (datato 6,5 milioni di anni), attribuito alla specie Sahelanthropus tchadensis e soprannominato Tumai; purtroppo non mostra nessun indizio sulla postura. È il rinvenimento del cosiddetto Millennium Man, nel 2000, a testimoniare che il primo ominino che ha sperimentato il bipedismo grazie a un femore molto lungo e robusto è stato Orrorin tugenensis, già presente in Africa nella foresta delle colline Tugen (Kenya) 6 milioni di anni fa. Come ulteriore testimonianza dell’emergenza della linea umana esso presenta un canino piccolo senza diastema.
Come Orrorin anche Ardipithecus ramidus è vissuto nella foresta. Resti scheletrici e denti datati 4,4 milioni di anni sono stati rinvenuti ad Aramis nella depressione di Afar in Etiopia. Dapprima essi sono stati attribuiti a una nuova specie del genere Australopithecus, ma a un più attento riesame è risultato che Ardipithecus ramidus rappresenta un morfotipo antenato di tutte le specie di O. successive, poiché alcuni caratteri dei denti e dello scheletro appendicolare appaiono intermedi tra quelli dello scimpanzé e quelli dell’Australopiteco.
La specie Ardipithecus kadabba, identificata successivamente, è ancora più antica (5,2-5,8 milioni di anni) e testimonia la fase precoce dell’evoluzione dei denti verso la linea umana.
L’Australopiteco.- Il genere Australopithecus vissuto in Africa presenta più specie che possono essere distinte geograficamente in un gruppo meridionale (Australopithecus africanus) e un gruppo orientale (Australopithecus anamensis, Australopithecus afarensis, Australopithecus ghari) cui vanno aggiunti i resti rinvenuti in Africa centrale (Australopithecus bahrelghazali). Il genere è vissuto tra 4,4 e 1,2 milioni di anni.
Fu con la scoperta nel 1924 a Taung (Sudafrica) di un cranio infantile di tipo scimmiesco ma con dentatura decidua di un bambino che l’anatomista sudafricano R. Dart coniò il termine Australopithecus africanus («scimmia australe africana»). In seguito, reperti appartenenti a individui adulti riferibili alla stessa specie, datati tra 2 e 3 milioni di anni, sono stati rinvenuti a Makapansgat e Sterkfontein: una colonna vertebrale quasi completa e parte di un femore hanno messo in evidenza che l’acquisizione della postura eretta era ormai un fatto compiuto. A Sterkfontein tuttavia, a indicare che l’Australopiteco era ancora un buon arrampicatore, è stato rinvenuto parte dello scheletro di un piede in cui l’alluce diverge alquanto dalle altre dita, mentre il calcagno è tipicamente umano.
La specie più antica del genere è Australopithecus anamensis (4,2 milioni di anni) a cui sono riconducibili i resti di scheletro craniale e postcraniale e i denti appartenenti a 21 individui rinvenuti nei pressi del Lago Turkana, a Kanapoi e Allia Bay (Kenya); i resti della tibia indicano una postura eretta, mentre i denti presentano ancora un certo grado di dimorfismo sessuale.
Di estrema importanza sono i resti di Australopithecus afarensis scoperti ad Hadar nella depressione di Afar; a tale specie sono pure attribuite le cosiddette orme di Laetoli (Tanzania), tipicamente umane per la presenza dell’arco longitudinale del piede, e i resti, sempre rinvenuti a Laetoli, di quella che viene chiamata ‘la prima famiglia’ comprendente più individui di ambo i sessi e di diversa età. Australopithecus afarensis ha vissuto tra 3,2 e 3,7 milioni di anni. Il reperto più completo e il più conosciuto per la sua buona rappresentatività (i resti rappresentano più del 40% dell’intero scheletro) si riferisce a Lucy, il piccolo scheletro di un esemplare femmina (statura e peso stimati pari a 110 cm e 40,6 kg rispettivamente). In Lucy, e più in generale nell’Australopiteco, la postura e la locomozione bipede sono affermate (bacino osseo basso e largo come nell’uomo, riduzione della lunghezza dell’arto toracico, foro occipitale quasi in linea con la colonna vertebrale, femore lungo e robusto, alluce non divergente), mentre il cranio è scimmiesco con una capacità cranica compresa tra 380 e 470 ml. Il ritrovamento in Etiopia, a Dikika, di Australopithecus afarensis, un individuo di sesso femminile di 3 anni di età datato 3,8 milioni di anni, ha messo chiaramente in luce che l’Australopiteco per la morfologia dell’arto superiore, simile per alcuni versi a quella delle antropomorfe africane, era ancora un buon arrampicatore.
Australopithecus ghari è rappresentato da resti di un cranio parziale e da denti rinvenuti a Bouri nel Middle Awash (Etiopia) datati 2,5 milioni di anni: la morfologia dei denti e lo spessore dello smalto pongono questa specie sulla linea degli Ominidi. L’Australopiteco ha vissuto nella savana sviluppando comportamenti adatti a spostarsi in grandi spazi aperti. L’affermazione della postura ortograda, riducendo la superficie del corpo esposta direttamente alle radiazioni solari, sarebbe stata vantaggiosa in un ambiente savanicolo soggetto a forte radiazione, e la maggiore ampiezza del campo visivo avrebbe permesso una migliore esplorazione del territorio; la locomozione bipede a sua volta avrebbe favorito gli spostamenti in tale ambiente con la mano libera che una volta sganciata dall’attività locomotoria poteva essere sfruttata nella raccolta di semi, radici, bacche, tuberi e per trasportare oggetti e i piccoli durante gli spostamenti. Si è discusso molto sul comportamento dell’Australopiteco: in un primo tempo esso fu definito una ‘scimmia assassina’, oggi è considerato piuttosto una preda inerme, priva sia di caratteri fisici sia di strumenti specializzati adatti a fronteggiare i predatori.
Il Parantropo. - Rispetto all’apparato masticatorio, le specie del genere Australopithecus sono definite ‘gracili’, per distinguerle dalle specie del genere Paranthropus in cui l’apparato masticatorio, e più in generale il cranio, è molto più robusto, tanto che questo presenta lo sviluppo di una cresta sagittale mediana (più nel maschio che nella femmina) per massimizzare l’area di inserzione dei potenti muscoli temporali. Un tempo le tre specie del genere Paranthropus erano attribuite al genere Australopithecus di cui erano considerate le forme ‘robuste’. In vista dell’estrema specializzazione dell’apparato masticatorio adatto a una dieta esclusivamente vegetariana ricca di fibre e materiale coriaceo, che richiedeva uno sforzo notevole nella masticazione per la ‘molitura’ del materiale fibroso, oggi viene generalmente accettata la loro sistemazione nel genere Paranthropus che nell’evoluzione degli Ominini rappresenta un ramo terminale senza sbocchi evolutivi.
Di Paranthropus robustus rimangono resti cranici e postcraniali in Africa meridionale a Kromdraai, Swartkrans e Drimolen, datati 1,8-1,2 milioni di anni.
Di Paranthropus boisei rimangono un cranio quasi completo, definito ‘Schiaccianoci’ per l’estrema robustezza dell’apparato masticatorio rinforzato da una robusta cresta sagittale, rinvenuto nella gola di Olduvai (Tanzania), datato 1,9 milioni di anni; e un altro cranio sulle sponde del Lago Turkana (Kenya) datato 2,8 milioni di anni.
Di Paranthropus aethiopicus rimane un cranio rinvenuto a Shungura (Etiopia) e un altro cranio rinvenuto a Koobi Fora (Kenya), soprannominato ‘The black skull’ per la colorazione dovuta al minerale presente nel sito di rinvenimento. In media la capacità cranica del Parantropo è pari a circa 500 ml.
Keniantropo. - Al genere Kenianthropus è attribuita la specie Kenianthropus platyops; nel cranio rinvenuto a Lomekwi (Kenya), deformato post mortem per deposizione nel terreno e datato 3,5 milioni di anni, la faccia è molto larga e piatta.
Comparsa e affermazione del genere Homo
Estintasi la linea dei Parantropi, l’evoluzione del genere Homo si ricollega a quella dell’Australopiteco, rispetto al quale, nei primi rappresentanti del nuovo genere, si verifica la riduzione dei denti iugali con uno splancnocranio più ridotto e retratto rispetto al neurocranio, il perfezionamento della postura ortograda con il completo allineamento del foro occipitale con la colonna, l’incremento della statura e dell’encefalizzazione, la manifattura dei primi strumenti litici. Al genere Homo fanno capo molte specie oggi tutte estinte: solo la specie Homo sapiens è rimasta a popolare tutta la terra.
Homo habilis. - La prima specie Homo habilis è comparsa in Africa nella gola di Olduvai (Tanzania) nello stesso ‘bed’ del Paranthropus boisei datato 1,9 milioni di anni. I resti sono rappresentati da una mandibola di tipo moderno molto meno robusta di quella del Parantropo, e da un parietale che, per misurazione indiretta, ha fornito la stima di 650 ml di capacità cranica; i resti scheletrici della mano e del piede presentano caratteri ‘moderni’. L’area di distribuzione di Homo habilis si estende fino al Kenya dove, sulle rive orientali del Lago Turkana, è stato rinvenuto un cranio ben conservato (etichettato KNM ER 1470) a grande capacità cranica (750 ml), ma con una faccia ‘arcaica’, più simile cioè a quella dell’Australopiteco per il grande sviluppo dell’apparato masticatorio. Nello stesso sito è stato rinvenuto anche un cranio (KNM ER 1813) nel quale viceversa la capacità è ridotta ma la faccia risulta molto più ‘umana’, con una mandibola poco sviluppata; a complicare il quadro nello stesso sito sono stati rinvenuti resti cranici nei quali è possibile evidenziare una faccia simile a quella del 1470, denti simili al 1813 e capacità cranica assai ridotta pari a 580 ml. La grande variabilità di tutte queste forme ha fatto ritenere che esse debbano essere attribuite a due specie, Homo rudolfensis, la più antica, rappresentata dal reperto KNM ER 1470, e Homo habilis rappresentata dai reperti di Olduvai e dal reperto KNM ER 1813. Secondo alcuni tuttavia si tratterebbe di una variabilità intraspecifica legata a dimorfismo sessuale. Al momento è certo che a Homo habilis è associato il primo manufatto litico (Modo I o industria olduvaiana) costituito da semplici choppers, cioè pietre scheggiate su un solo lato (monofacciali), così da ottenere un margine tagliente da utilizzare in vario modo, ma non certo come arma di offesa. Homo habilis infatti non era un cacciatore ma si comportava da sciacallo quando aveva l’occasione di sfruttare la carcassa di un animale ucciso da un carnivoro, mentre le proteine animali che arricchivano la sua dieta, essenzialmente vegetariana, le doveva trovare in lucertole, vermi, uova, piccoli mammiferi.
Homo erectus. - È vissuto tra 1,8 e 0,4 milioni di anni. I numerosi reperti riferibili a tale specie provengono dall’Asia (i più tardivi) e dall’Africa (i più antichi). Le prime indagini (1891) condotte nell’isola di Giava (Indonesia) avevano messo in luce la presenza di quello che fu allora considerato ‘l’anello mancante’ nell’evoluzione degli O. e perciò classificato Pitecanthropus erectus. Ai reperti costituiti principalmente da una calotta cranica e un femore completo del tutto simile a quello moderno, si aggiunsero quelli rinvenuti a Zhoukoudian (Cina) che furono assegnati a un genere distinto: Sinanthropus pekinensis. In base ad attenta revisione tutti i reperti asiatici sono oggi attribuiti a Homo erectus e a tale specie sono da riferire anche molti reperti più antichi rinvenuti in Africa (Sudafrica, Algeria, Tanzania, Etiopia). A Koobi Fora sulle sponde orientali del Lago Turkana e a Nariokotome (sulle sponde occidentali) numerosi reperti, datati tra 1,8 e 1,9 milioni di anni, costituiscono il cosiddetto gruppo africano ‘precoce’ di Homo erectus, classificato oggi dalla maggior parte dei paleoantropologi Homo ergaster, distinto da Homo erectus asiatico (Homo erectus stricto sensu) che presenta caratteri peculiari: il cranio è notevolmente platicefalo e con una visiera soparorbitaria continua accompagnata da un marcato restringimento retroorbitario; l’occipitale presenta un’angolatura accentuata tra squama e piano nucale; la capacità cranica varia tra 950 ml del reperto di Giava e 1200 ml in quello cinese. In Asia Homo erectus stricto sensu non è accompagnato da industria litica di bifacciali; infatti l’arcaicità del cranio si accompagna a manufatti di Modo I; per tali ragioni alcuni paleoantropologi ritengono che vi sia stata una migrazione dall’Africa di una forma precedente Homo ergaster (Homo habilis?).
Homo ergaster. - I reperti più significativi di Homo ergaster, ritrovati a Nariokotome, sono rappresentati da un cranio datato 1,8 milioni di anni e uno scheletro postcraniale (KNM WT 15.000) molto ben conservato (datato 1,5 milioni di anni) di un giovane di circa 12 anni (per questo chiamato ‘Turkana boy’), con capacità cranica pari a 850 ml, un peso stimato di 55 kg e una statura di 162 cm, il che comporta una stima di 180 cm nell’adulto. Homo ergaster rappresenta la specie chiave nel processo di ominazione per la morfologia moderna dello scheletro postcraniale adattato all’ambiente di savana caldo e arido. In tale ambiente infatti l’alta statura, indice di grande superficie corporea, è vantaggiosa per una buona dispersione del calore e i segmenti distali degli arti pelvici molto robusti, con impronte muscolari ben evidenti, parlano in favore di un adattamento per una maggiore resistenza alla lunga marcia su terreni aperti e accidentati; nella faccia la verticalizzazione del naso legata alla proiezione delle ossa nasali sul piano sagittale avrebbe comportato l’estensione della mucosa nasale utile per l’umidificazione dell’aria inspirata. La capacità e la morfologia del cranio sono invece ancora di tipo arcaico: cranio basso, con neurocranio ancora ridotto con forte aggetto delle arcate sopraorbitarie e splancnocranio con prognatismo accentuato; inoltre il diametro del canale vertebrale è ridotto a suggerire una minor efficienza del midollo spinale nel trasmettere gli stimoli dalla periferia ai centri encefalici e viceversa. A Homo ergaster è associata un’industria litica (Modo II o industria acheuleana) rappresentata dai cosiddetti bifacciali, ottenuti asportando per percussione dal blocco litico (arenaria, quarzite) alcune schegge su due lati opposti (il che contempla il concetto di simmetria), fino a ottenere un manufatto elaborato a forma di mandorla (amigdala) con la base adatta a essere impugnata e due margini funzionali. Per la prima volta fuori dall’Africa (Out-of-Africa I), a Dmanisi (Repubblica della Georgia), resti cranici (datati 1,7 milioni di anni) sono stati attribuiti a un discendente emerso dal gruppo di Homo ergaster (Homo ex gr. ergaster o Homo georgicus), per la loro grande somiglianza morfologica con H. ergaster, anche se la sua capacità cranica è più ridotta (650-780 ml).
Homo heidelbergensis. - Successiva a Homo ergaster, la specie Homo heidelbergensis si diffonde in Africa tra 0,6 e 0,3 milioni di anni (principali siti: Bodo, Etiopia; Kabwe, Zambia; Ndutu, Tanzania) e da qui raggiunge l’Europa a partire da 0,5 milioni di anni, come testimonia il reperto di Mauer, costituito da una mandibola assai robusta rinvenuta a Heidelberg (Germania). Arago (Francia), Petralona (Grecia), Atapuerca (Spagna) sono tra i siti in cui la presenza di Homo heidelbergensis è ben documentata; esso avrebbe raggiunto anche l’Asia (reperti di Dali, Cina). Rispetto a Homo erectus, Homo heidelbergensis presenta un toro sopraorbitario infossato nella regione glabellare, ossia una forte prominenza delle arcate sopraciliari, che ne preannuncia la scomparsa, una squama dell’occipitale più arrotondata, un cranio più espanso nella regione parietale, una faccia meno prognata, una capacità cranica pari a circa 1200 ml (nel reperto di Petralona). A Homo heidelbergensis è associata l’industria Modo II, ricca di strumenti litici adatti per scortecciare e raschiare materiale vegetale e per svolgere attività collegate con la caccia (tagliare, scuoiare, disarticolare, spezzare). Homo heidelbergensis era infatti un buon cacciatore che probabilmente cacciava in gruppo anche la megafauna (bovidi, cervidi, suidi ed elefanti, come testimoniano i ritrovamenti nei siti spagnoli di Torralba e Ambrona), anche se la raccolta di semi, radici, tuberi, miele, uova e insetti doveva costituire ancora una fonte di cibo importante. Campi base, accampamenti semistabili, siti di macellazione, atelier di taglio della pietra sono ben rappresentati in Europa, testimoniando l’organizzazione territoriale e sociale con la possibile divisione dei ruoli tra sessi e tra differenti classi generazionali.
Homo antecessor. - In Europa Homo heidelbergensis è stato preceduto dalla specie Homo antecessor, per prima descritta in Spagna nello strato inferiore della grotta della Gran Dolina ad Atapuerca (0,77 milioni di anni); l’antichità del reperto parla in favore di un popolamento dell’Europa a partire dall’Africa più antico di quanto si pensasse. Il ritrovamento a Ceprano (Lazio) di un cranio per certi versi simile a Homo antecessor e altrettanto antico (0,8 milioni di anni) confermerebbe la presenza precoce dell’uomo in Europa, dove Homo heidelbergensis precede la comparsa di Homo neanderthalensis, mentre in Africa la sua estinzione è marcata dalla comparsa di Homo sapiens.
Homo neanderthalensis. - Dopo il primo ritrovamento (1856) di una calotta cranica e molti resti dei segmenti appendicolari attribuiti all’Uomo di Neandertal, avvenuto in una grotta nella valle presso Düsseldorf, sono stati rinvenuti centinaia di resti di questa specie distribuiti in tutta l’Europa e nel Vicino e Medio Oriente (Israele, Iraq) in un arco di tempo compreso tra circa 0,2 e 0,03 milioni di anni. Il gruppo ‘precoce’ di forme precede l’ultima glaciazione di Würm (Pleistocene medio) che ha interessato gran parte dell’Europa. Tra i molti siti neandertaliani i più rappresentativi in ordine cronologico, dai più antichi, sono Swancombe (Inghilterra), Steinheim (Germania), La Chaise, Biache, Fontéchevade (Francia), Ehringsdorf (Germania), Montmaurin (Francia), Saccopastore (Italia). In tutte queste forme i caratteri neandertaliani sono meno marcati e nel complesso la morfologia è più moderna: il cranio, per es., è più arrotondato e la squama dell’occipitale è verticalizzata avvicinandosi alla forma tipicamente globosa del cranio umano. I Neandertaliani würmiani, vissuti tra 75.000 e 30.000 anni fa, per i caratteri peculiari costituiscono il gruppo dei cosiddetti Neandertaliani ‘classici’ la cui morfologia cranica è nettamente distinguibile da quella di Homo sapiens, così come da quella di Homo heidelbergensis. Infatti nei würmiani il cranio presenta una notevole platicefalia e un toro sopraorbitario accentuato; il frontale è molto sfuggente, l’occipitale presenta un toro occipitale trasverso (‘chignon’); la volta cranica è molto spessa, la faccia è prognata e oncognata (rigonfia a livello dei mascellari), con cavità paranasali sviluppate (pneumatizzazione dei mascellari); la mandibola manca di mento. Lo scheletro postcraniale è molto robusto: le proporzioni corporee ottenute rapportando il volume (peso corporeo stimato) alla superficie (statura stimata) danno un corpo tarchiato e con arti corti, specialmente nei segmenti distali, una costituzione brevilinea che è vantaggiosa nelle regioni fredde per ostacolare la perdita di calore. L’uomo di Neandertal era un valente cacciatore e un abile artigiano; l’industria musteriana (Modo III) è rappresentata da manufatti con punte, raschiatoi, lame che permettevano varie operazioni elaborate. Esso abitava in grotte o caverne nei pressi di corsi d’acqua, seppelliva i suoi morti (sono contestati da molti paleoantropologi i riti funebri), si prendeva cura dei disabili, non ricorreva a cannibalismo rituale, come si pensava un tempo. La socializzazione era favorita dal linguaggio verbale, come testimonierebbe la ricostruzione dell’apparato della fonazione. L’uomo di Neandertal crea tuttora problemi d’interpretazione: come rappresentante della ‘fase arcaica’ dell’umanità, potrebbe esserne considerato un antenato diretto, oppure un ramo che si è estinto durante l’evoluzione.
Homo sapiens
Due sono le teorie sull’origine di Homo sapiens. La prima ipotizza la sua comparsa in Africa intorno a 200.000 anni fa (teoria monocentrica-migrazionista, o Out of Africa II, proposta per la prima volta da C. Stringer), cominciando a diffondersi nell’arco degli ultimi 70.000 anni anche verso l’Australasia (dove arriva intorno a 60.000) e in Asia orientale, che raggiunge intorno ai 40-50.000 anni fa soppiantando Homo erectus stricto sensu; in Europa esso arriva intorno a 35.000 anni fa soppiantando Homo neanderthalensis. La seconda ipotizza l’origine di Homo sapiens come il prodotto dell’evoluzione regionale di tipo gradualistico di Homo erectus e di Homo neanderthalensis (teoria policentrica o multiregionale di M. Wolpoff); in questo caso l’uomo di Neandertal rappresenterebbe la ‘varietà’ umana arcaica classificata come Homo sapiens neanderthalensis per distinguerla dalla varietà ‘moderna’ europea di Homo sapiens sapiens; a loro volta le moderne popolazioni africane e asiatiche (orientali e australoasiatiche) rappresenterebbero il prodotto finale dell’evoluzione regionale di Homo erectus, per cui le attuali differenze fisiche delle diverse popolazioni continentali rappresenterebbero il frutto di centinaia di migliaia di anni di evoluzione graduale. Il modello monocentrico è quello su cui c’è un generale accordo.
Le testimonianze a favore della prima ipotesi sono paleontologiche. In Africa, a Omo Kibish (Etiopia) e a Border Cave e Klasies River (Sudafrica) sono stati rinvenuti i resti cranici più antichi, datati circa 190.000 anni, che presentano una morfologia del tutto moderna: frontale verticalizzato, presenza di mento, cranio arrotondato nella regione parietale, presenza della fossa canina a livello del mascellare, capacità cranica di circa 1350 ml. I ritrovamenti in Etiopia (regione di Afar, formazione di Bouri, Herto Member, datata 200.000 anni), costituiti da un cranio adulto molto ben conservato, colmerebbero il vuoto tra Homo heidelbergensis e Homo sapiens e sono classificati da alcuni paleoantropologi Homo sapiens idaltu («il più arcaico»), una varietà che per modificazione graduale avrebbe rappresentato il diretto antenato dell’uomo moderno. Nel campo di variabilità della morfologia moderna dell’uomo rientrerebbe anche il reperto rinvenuto a Liang Bua nell’isola di Flores (Indonesia) costituito da denti, un cranio e parte dello scheletro postcraniale datati 18.000 anni e appartenenti a un individuo adulto con una statura non superiore a 106 cm e una capacità cranica pari a 380 ml. Attribuito a una nuova specie, Homo floresiensis emersa da una popolazione di Homo erectus asiatico e in seguito diversificatasi per isolamento, è da alcuni attribuito a una varietà insulare di Homo sapiens per la sua somiglianza morfologica con la popolazione che abita l’isola (i pigmei Rampasasa); la capacità cranica estremamente ridotta dell’individuo sarebbe stata l’esito di una patologia subita durante l’accrescimento.
A favore della seconda ipotesi, le prove neontologiche basate sullo studio delle sequenze del DNA hanno confermato i dati paleontologici. In primo luogo l’uomo di Neandertal non è l’antenato di Homo sapiens ma è da attribuire a una specie divergente, Homo neanderthalensis. Il confronto fra il DNA della sequenza di 378 basi nucleotidiche del DNA mitocondriale (mtDNA) ottenuta per purificazione dalle polvere d’osso dell’omero dell’uomo di Neandertal confrontata con quello di individui appartenenti a popolazioni attuali di diversa origine geografica (Africani, Asiatici, Australoasiatici ed Europei) ha infatti evidenziato che in media vi sono 27 differenti mutazioni tra i neandertaliani e tutte le altre popolazioni attuali, un numero che è tre volte più grande di quello (circa 8) che differenzia queste ultime tra loro. La divergenza evolutiva tra i due rami della famiglia degli O. si sarebbe verificata intorno a 0,45 milioni di anni. Indagini successive condotte su 65.000 basi nucleotidiche ottenute dal DNA autosomico di reperti neandertaliani datati 38.000 anni provenienti da una grotta in Croazia, hanno confermato l’identità specifica dei neandertaliani. Tuttavia non può essere escluso che si sia verificato un certo grado di mescolamento tra l’uomo di Neandertal e l’uomo moderno, dal momento che in alcuni territori (per es., a Châtelperron, Francia meridionale) essi hanno convissuto per almeno 8.000 anni prima che i Neandertaliani si estinguessero, intorno o poco dopo 30.000 anni fa. A Gibilterra i reperti scheletrici neandertaliani risalgono a circa 28.000 anni e in Portogallo (Lagar Velho) resti scheletrici di età infantile sono datati 24.000 anni.
A Homo sapiens è associata l’industria aurignaziana (➔ Aurignaziano) costituita da un repertorio di manufatti ritoccati soprattutto in pietra (ma in anche legno, osso, avorio e corno), opera dell’uomo di Cro-Magnon riferibile a un moderno europeo. Strumenti aurignaziani si ritrovano associati anche agli ultimi Neandertaliani di Châtelperron, ma resta dubbio se si tratti di un’invenzione indipendente da parte dei Neandertaliani o se si tratti di acculturazione. In secondo luogo le testimonianze neontologiche date dal confronto del DNA di popolazioni africane con quello di popolazioni extrafricane, confermerebbero da un lato che la specie Homo sapiens è recente (con il mantenimento di una sostanziale omogeneità genetica), dall’altro che filogeneticamente le popolazioni africane rappresentano il ramo di partenza da cui, per mutazioni, si sarebbero separate le popolazioni extrafricane. Il modello evolutivo oggi più accreditato basato sul confronto delle sequenze genomiche ammette che il contributo più alto al pool genico della specie è dato dal continente africano. Gli studi più accreditati ipotizzano che dal nucleo originario di Homo sapiens, stimato in 10.000 unità riproduttive, si separarono in loco più sottogruppi che in seguito a isolamento persero parte della variabilità originaria (fenomeno di deriva genetica) diversificandosi, e, anche in seguito a migrazione dall’Africa, costituirono sottoinsiemi del pool genico originario. Partendo dall’analisi delle sequenze dell’mtDNA nelle attuali popolazioni dei diversi continenti, si è osservato che gli Africani subsahariani presentano una variabilità genetica più alta con l’mtDNA più simile al coalescente ovvero alla sequenza mitocondriale ancestrale da cui discenderebbero tutte le linee attuali presenti nella specie. Una volta migrate dall’Africa, le ‘nuove’ popolazioni si trovarono a fronteggiare situazioni ambientali diverse che portarono prima di tutto a una selezione dei caratteri somatici: le differenze fisiche che si riscontrano oggi tra le popolazioni dei vari continenti sono da considerare quindi quali varianti geografiche recenti, piuttosto che varianti ‘regionali’ seguendo l’ipotesi di Wolpoff. Per es., nel caso del colore della pelle dovuto alla melanina, il pigmento prodotto dai melanociti, le varietà più scure si accentrano nelle regioni tropico-equatoriali (Africa subsahariana, deserto occidentale dell’Australia, penisola indiana, Nuova Guinea), poiché risultano vantaggiose per proteggere la pelle dalle radiazioni ultraviolette eritematogene; un corpo longilineo (statura alta e peso relativamente basso) favorendo la dispersione del calore risulta vantaggioso tra le popolazioni subsahariane, all’opposto negli Inuit dell’estremo Nord è un corpo brevilineo a essere favorito. Sulla stessa base adattativa può essere interpretata la particolare morfologia della palpebra superiore degli asiatici (Mongoli, Cinesi), quella del naso e dei capelli, anche se per questi casi non si ha conferma sperimentale.
Alla diversificazione dell’uomo hanno contribuito in larga misura anche le vicende che hanno interessato il popolamento dei diversi territori, in primo luogo le grandi migrazioni. Esse hanno influito in maniera determinante sugli scambi genetici e/o culturali tra popolazioni di territori adiacenti, dove l’uso della stessa lingua ha favorito il mescolamento tra ‘diversi’ imponendosi sugli effetti selettivi dell’ambiente geografico. La correlazione tra la variabilità genetica e la variabilità linguistica è risultata positiva e altamente significativa a indicare quanto questo elemento culturale sia stato determinante durante la fase più recente del processo di diversificazione dell’uomo. Lo studio della variabilità dell’mtDNA si è dimostrato particolarmente utile per ricostruire le vicende del popolamento dei 5 continenti. Per es., nel caso dell’Europa è stato messo in evidenza che la variabilità genetica riscontratavi oggi è in gran parte quella originaria delle popolazioni arrivate dall’Africa durante il Paleolitico superiore, mentre l’apporto genetico delle popolazioni neolitiche migrate dal Medio Oriente – e diffusesi nel continente europeo in seguito all’affermazione dell’agricoltura – è assai limitato. Allo stesso risultato si è arrivati analizzando le sequenze del DNA del cromosoma Y.