
sistematica
Ramo delle scienze biologiche che si occupa dello studio e dell’identificazione delle relazioni tra esseri viventi e fossili e rappresenta tali relazioni in sistemi gerarchici che, a loro volta, ne costituiscono la classificazione.
S. filogenetica e s. evoluzionistica
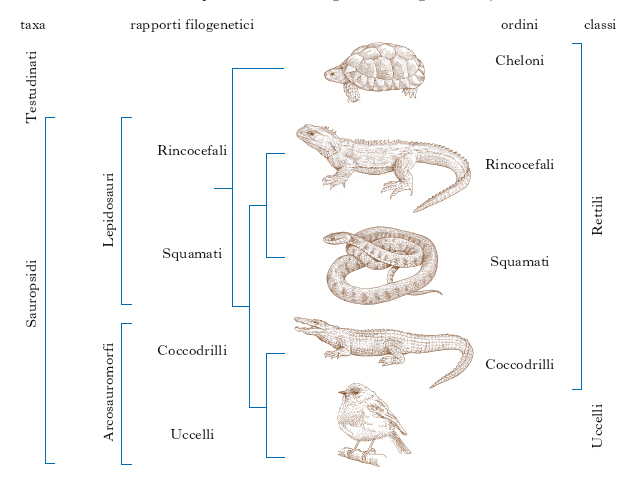
In base ai criteri tassonomici adottati, si distinguono due scuole: quella della s. filogenetica (detta anche cladistica) e quella della s. evoluzionistica, a seconda che la classificazione risultante tenga conto esclusivamente delle relazioni filogenetiche tra le specie o che, per la sua definizione, possano essere considerati altri elementi, come la comparsa di importanti novità evolutive. Sono un esempio gli Uccelli, collocati secondo la prima nella classe dei Rettili (essendo strettamente imparentati ai coccodrilli), mentre, per la seconda, essi costituiscono classe a sé per le novità evolutive presentate, quali la comparsa delle penne (fig. 1).
L’individuazione dei rapporti filogenetici, scopo principale della s., si avvale di tecniche mediate dalla biologia molecolare e dalla genetica (struttura delle proteine, studiata con metodi immunologici o elettroforetici; struttura degli acidi nucleici; tecniche di ibridizzazione del DNA). La tecnica di ibridizzazione degli acidi nucleici, applicata alla filogenesi dei vari taxa di Uccelli, ha dimostrato che molte forme ritenute filogeneticamente affini erano in realtà l’espressione di un’evoluzione convergente; un altro esempio, riferito invece ai Mammiferi, riguarda i panda, maggiore (Ailuropoda melanoleuca) e minore (Ailurus fulgens): i dati ottenuti dalle tecniche di ibridizzazione del DNA indicherebbero che il panda maggiore appartiene in effetti agli Ailuropodidi, famiglia affine agli Ursidi, mentre il minore apparterrebbe agli Ailuridi, famiglia maggiormente affine ai Procionidi. Nei suoi sviluppi più recenti, alla luce del rapido declino per fattori antropici della biodiversità, la s. ha assunto inoltre il ruolo vitale di descrivere il grande numero delle specie minacciate dall’estinzione. Nuovo impulso alle ricerche di s. deriva infine dalle sue numerose applicazioni in vari campi (agricoltura, industria, farmacologia).
Approccio biologico-molecolare
La s. dei viventi è soggetta a continue discussioni e aggiustamenti, a causa di sempre nuove acquisizioni di carattere morfologico, paleontologico, embriologico e genetico, che contribuiscono a ricostruire la storia evolutiva di un gruppo (e perciò la s. al suo interno) e le sue relazioni con altri gruppi. Soprattutto a partire dagli anni 1980 la biologia molecolare ha dato un notevole impulso a tale tipo di indagini, contribuendo in maniera sostanziale alla descrizione, a diversi livelli evolutivi, dell’albero della vita che lega, a partire da un progenitore comune vissuto circa 4 miliardi di anni fa, tutti gli organismi viventi. Molti degli studi che utilizzano i marcatori molecolari possono essere visti come tentativi di stimare la filogenesi, nel senso più vasto del termine, a uno dei vari livelli gerarchici di divergenza evolutiva; le relazioni filogenetiche fra i gruppi, così, possono essere studiate a livelli che vanno dalla micro- alla macroevoluzione. Le relazioni filogenetiche emerse a livello molecolare hanno dunque fornito opportunità senza precedenti alla comprensione dell’origine e della storia evolutiva degli organismi. Dal momento che la filogenesi descrive il ‘flusso dell’eredità’, solo i caratteri geneticamente trasmessi danno informazioni sulla stima della filogenesi stessa. Raramente i geni o gli alleli che controllano un determinato carattere morfologico, fisiologico o comportamentale possono essere specificati, e infatti molti caratteri tradizionalmente usati in tassonomia sono fortemente influenzati da fattori non genetici. È evidente che tali caratteri, se considerati come buoni descrittori in studi di s., possono risultare fuorvianti e condurre a conclusioni errate e prive di una reale base genetica.
Omologie e analogie
Il problema centrale degli studi della filogenesi e della tassonomia dei viventi è sempre stato quello di poter distinguere le similarità dovute alla discendenza comune tra due specie (omologie) da quelle dovute a fenomeni di convergenza evolutiva (analogie). Ovviamente, la s. dei viventi dovrebbe riflettere solamente le prime, ma caratteri e comportamenti condivisi da più specie, che peraltro sono stati in genere quelli maggiormente tenuti in considerazione nelle varie s., sono risultati spesso il frutto di evoluzioni convergenti, vale a dire di risposte simili a stimoli ambientali simili da parte di specie non vicine filogeneticamente: l’ibridazione DNA-DNA ha, per es., evidenziato affinità tra i Ciconiformi e gli avvoltoi americani (➔ Catartiformi). È invece estremamente improbabile che affinità diffuse e complesse, riguardanti centinaia o migliaia di caratteristiche molecolari, derivino da fenomeni di convergenza, e, conseguentemente, tali affinità devono riflettere una reale discendenza comune tra due organismi.
Misure di divergenza genetica

L’approccio molecolare consente una comparazione diretta dei livelli relativi delle differenze genetiche praticamente tra tutti i gruppi di organismi. Si supponga, per es., di voler confrontare dal punto di vista evolutivo il grado di divergenza all’interno di un taxon di Pesci (genere o famiglia) con quella di un taxon corrispettivo di Uccelli. I caratteri morfologici tradizionalmente utilizzati nella s. dei Pesci (numero di squame lungo la linea laterale, raggi branchiostegali, posizione delle pinne ecc.) sono chiaramente di nessuna utilità in una ipotetica comparazione con i caratteri di rilevanza sistematica degli Uccelli (disposizione delle penne, struttura del siringe, numero e posizione delle dita del piede ecc.). Ciò ha obbligato i sistematici a sviluppare standard differenti per ognuno dei gruppi animali oggetto di studio. In altre parole, la s. di un gruppo animale non ha necessariamente lo stesso significato di quella di un gruppo diverso, perché diversi sono i criteri sui quali essa si basa. La fig. 2 mostra un grafico di comparazione genetica tra diversi taxa di Vertebrati: Uccelli appartenenti allo stesso genere mostrano una variabilità genetica molto più bassa di quanto non si osservi negli Anfibi e nei Rettili, a dispetto di una grande variabilità morfologica. Ciò può indicare che le specie congeneri degli Uccelli sono evolutivamente più giovani, e quindi hanno avuto meno tempo per divergere, oppure che le proteine degli Uccelli evolvono più lentamente; un risultato che conferma comunque come evoluzione morfologica e genetica possano, in molti casi, procedere a ritmi indipendenti. Tale tipo di analisi apre prospettive e potenzialità che la semplice comparazione morfologica non permetteva, consentendo, per es., di evidenziare, già a partire dalla metà degli anni 1970, come proteine e acidi nucleici di uomo e scimpanzé divergano in modo analogo a quanto misurato, in altri taxa, tra due specie vicine o addirittura congeneri. Ciò suggerisce che la tradizionale s. dei Primati, che pone uomo e scimpanzé in famiglie differenti (rispettivamente Ominidi e Pongidi), potrebbe essere basata solo su differenze morfologiche dovute a pochi geni regolatori che agiscono durante l’ontogenesi, o anche che le distinzioni morfologiche tra uomo e scimpanzé siano state enfatizzate a causa di un bias (errore sistematico) antropocentrico, abbiano cioè pesato oltre il loro genuino significato evolutivo.