
riparazione
Biologia
R. del DNA In biologia molecolare, la reintegrazione della funzionalità delle molecole di acido desossiribonucleico (DNA) che hanno subito un danno. Alcuni danni sono dovuti all’azione di sostanze mutagene o cancerogene ambientali, altri sono il risultato di errori che si verificano nel corso dei normali processi del metabolismo cellulare. La mancata r. del danno determina un accumulo di mutazioni, cioè di cambiamenti permanenti del DNA. Nelle cellule somatiche, l’accumulo di mutazioni aumenta la probabilità di insorgenza di tumori; nelle cellule germinali porta a un aumento delle anomalie ereditarie, spesso letali. I processi enzimatici di r. del DNA delle cellule sono pertanto fondamentali per impedire l’accumulo del danno, che si traduce in un aumento di mutazioni nelle generazioni cellulari successive.
La frequenza di mutazioni durante la duplicazione del DNA risulta molto più bassa rispetto a quella prevista in base alla frequenza con cui si formano appaiamenti errati (cioè 1 su 106 paia di basi). Le cellule, infatti, presentano due tipi di difesa in relazione agli errori di appaiamento che si verificano quando la DNA-polimerasi aggiunge un nucleotide sbagliato all’estremità crescente 3′ del nuovo filamento di DNA. Una prima forma di r. è fornita dalla stessa DNA-polimerasi che ha anche la funzione di nucleasi. Quando infatti non si verifica l’appaiamento corretto purina-pirimidina la conformazione diversa del DNA viene subito riconosciuta dalla polimerasi, che assume un’attività esonucleasica e quindi stacca la base errata dal filamento in crescita. Sia le polimerasi batteriche sia le polimerasi umane ε e δ presentano tale attività. Studi effettuati sui batteri e sui batteriofagi mutanti hanno dimostrato che questo processo riduce la quantità di mutazioni di un fattore di circa 1000 e, pertanto, la frequenza di mutazione diventa di 1 su 109 paia di basi replicate.
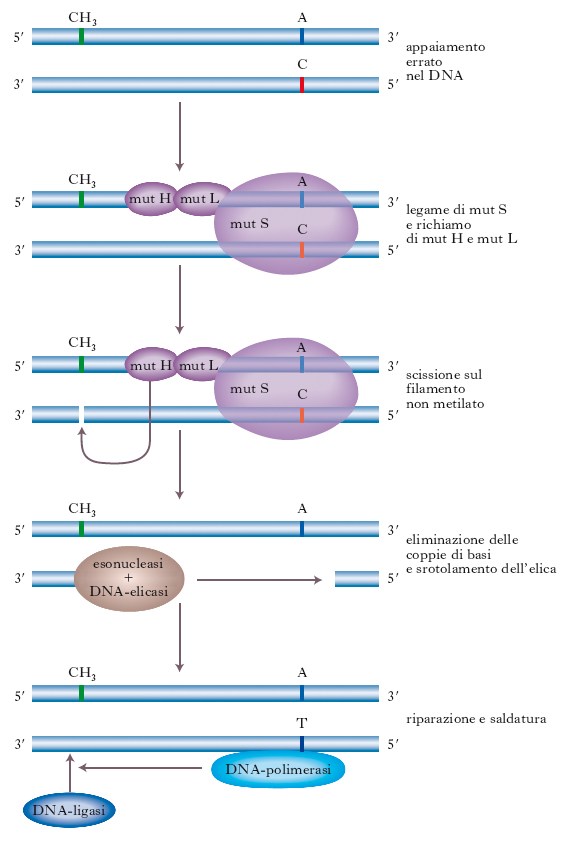
Non tutti gli appaiamenti sbagliati, tuttavia, vengono riconosciuti in tempo e si possono formare molecole che presentano un appaiamento non corretto di una base che determinerà, nel successivo ciclo di duplicazione, una mutazione. Per impedire questo interviene un sistema di r., detto r. dell’appaiamento errato guidato dal metile (v. fig.), ben studiato nei batteri ma che presenta, come quasi tutti gli altri sistemi di r., caratteristiche simili anche nei Mammiferi. Lo studio di questo sistema ha assunto un grande interesse da quando negli anni 1990 si è scoperto che un tipo di tumore ereditario umano, il tumore colon-rettale ereditario non polipoide (➔ HNPCC), è causato da mutazioni nei geni che riguardano il sistema di r. dell’appaiamento errato. Il processo è stato studiato inizialmente nei batteri con fenotipo detto mutatore (mut), che presentavano un’aumentata frequenza di mutazione spontanea in tutti i loci del genoma.
Nel sistema di r. degli appaiamenti errati studiati nei batteri, gli enzimi della r. scorrono lungo il DNA dopo la duplicazione, individuano gli errori e ristabiliscono il corretto appaiamento. Il modo per riconoscere, fra i due filamenti di DNA, quello di nuova sintesi da correggere si basa sul fatto che quest’ultimo non è metilato. La metilazione (➔) è un fenomeno che si verifica sia nei procarioti sia negli eucarioti: i gruppi metilici vengono aggiunti al filamento di nuova sintesi solo dopo un certo periodo di tempo dal processo di duplicazione del DNA. Le proteine coinvolte in questo processo, individuate in Escherichia coli (chiamate proteine mutatrici o mut), sono mut S, mut L e mut H. La fig. mostra le varie fasi del processo: il filamento appena replicato non metilato contiene un appaiamento errato che determina una modificazione del DNA riconosciuta dalla proteina mut S; il legame di questa proteina al DNA richiama le proteine omologhe mut L e mut H. Quest’ultima è una endonucleasi che taglia il filamento di DNA non metilato opposto al sito 5′-GATC-3′; una esonucleasi e una elicasi scorrono sul DNA eliminando circa 100 paia di basi oltre all’appaiamento errato; la risultante interruzione nel DNA viene riparata mediante la DNA-polimerasi, che allinea nucleotidi secondo la sequenza dettata dal DNA di stampo; nell’ultima tappa interviene la DNA-ligasi, che salda il DNA. Nell’uomo le prove accumulate, soprattutto negli studi sulle mutazioni riguardanti individui affetti dal carcinoma colon-rettale non poliposico, indicano che vi sono almeno 5 loci genici implicati nel processo di riparazione. Le proteine codificate da questi geni rimuovono sia gli appaiamenti errati, sia le anse di 2-4 paia di basi fuori posto rispetto all’elica di DNA. Alcune proteine presentano un notevole grado di omologia con mut S e mut L e la via di r. sembra simile a quella presente nei batteri, anche se nell’uomo è più complessa e non se ne conoscono tutti i dettagli.
Botanica
Fenomeno di neoformazione per il quale l’organo di una pianta al quale sia stata asportata una parte, ripara la perdita ricostituendo la porzione asportata. Tali fenomeni sono frequenti nelle alghe, molto più rari nelle piante superiori (per es., l’estremo apice radicale).
Istruzione
Esame di r. Esame che l’alunno delle scuole statali, parificate o legalmente riconosciute, doveva sostenere nella sessione autunnale quando allo scrutinio finale o all’esame di idoneità della sessione estiva per i privatisti avesse dimostrato di possedere in qualche disciplina una preparazione insufficiente. Gli esami di r. sono stati aboliti nelle scuole elementare e media dalla l. 517/4 agosto 1977; dall’anno scolastico 1994-95 sono stati aboliti anche nella scuola secondaria superiore, dove sono stati sostituiti con il sistema dei debiti formativi (➔ debito).
Religione
Per la teologia cattolica, la cancellazione, a opera di Cristo, dell’offesa fatta dall’uomo a Dio con il peccato originale, ma anche le preghiere e le buone opere compiute come emendazione dei peccati.