
neurotrasmettitore
Sostanza che veicola le informazioni neuronali attraverso la trasmissione sinaptica.
Modalità di liberazione
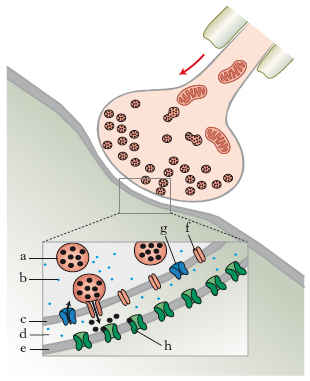
I n. sono sintetizzati nel citoplasma neuronale partendo da precursori molto comuni, disponibili nel neurone, attraverso numerose tappe enzimatiche. La trasmissione sinaptica è specifica, in quanto si basa sull’interazione tra il n. liberato dalla parte presinaptica e un recettore specifico presente sulla membrana postsinaptica: solo quando i due elementi, trasmettitore e recettore, sono compatibili, avviene la neurotrasmissione. I n. sono trasportati all’interno delle vescicole sinaptiche da trasportatori specifici e liberati in seguito alla stimolazione sinaptica. Nel momento in cui la membrana presinaptica è invasa dal potenziale d’azione, essa si depolarizza, entrano ioni Ca++ che favoriscono la fusione con la membrana plasmatica delle vescicole sinaptiche, ricche di n., le quali liberano il n. nello spazio sinaptico (fig. 1). Il n. liberato attiva i recettori postsinaptici e in questo modo il segnale si trasferisce da una cellula all’altra. La risposta che la cellula innervata produce quantitativamente è funzione del numero di molecole di n. uscito dalla parte presinaptica e del numero di recettori postsinaptici attivati.
Studi di biologia cellulare hanno dimostrato che vi è una grande differenza tra la sintesi dei due gruppi di n., i n. classici o convenzionali e i neuropeptidi; i primi sono sintetizzati a livello del citoplasma presinaptico e veicolati poi nelle vescicole sinaptiche tramite trasportatori, mentre i neuropeptidi sono sintetizzati nel reticolo endoplasmatico, nel corpo cellulare, e da lì trasferiti nei granuli di secrezione che vengono poi trasportati fino alla sinapsi; inoltre, le vescicole sinaptiche possono essere ricaricate del n. classico subito dopo averlo liberato, mentre i granuli che contengono i neuropeptidi non possono più essere ricaricati e la sinapsi deve essere rifornita di nuovi granuli dal corpo cellulare; infine, sembra che per liberare i neuropeptidi siano necessarie stimolazioni nervose più intense e prolungate. L’ipotesi attuale più accreditata è che i veri n. siano quelli di tipo classico, mentre i neuropeptidi agirebbero soprattutto da modulatori dell’attività neuronale.
Sintesi dei n. classici
Appartengono a questo gruppo l’acetilcolina (le sinapsi colinergiche nel sistema nervoso sono il 5-10 %), le monoammine (catecolammine, serotonina e istamina, secrete dall’1-2 % delle sinapsi cerebrali) e gli amminoacidi (GABA, glutammato, aspartato, taurina, secreti da circa il 60% delle sinapsi centrali). Ogni neurone sintetizza solo un n. di tipo convenzionale; si hanno così neuroni colinergici, dopaminergici, GABAergici ecc., che sono raggruppati in nuclei specifici del sistema nervoso centrale (SNC).

L’acetilcolina è sintetizzata a partire dalla colina, che entra nei neuroni attraverso uno specifico trasportatore per azione della colina-acetiltransferasi, ed è inattivata dalle acetilcolinesterasi, presenti nello spazio sinaptico. I neuroni colinergici sono raggruppati soprattutto nel nucleo magnocellulare di Maynert, nel nucleo del setto mediale e in alcuni nuclei pontini. Da lì essi si dirigono verso la corteccia in modo diffuso, al talamo e all’ippocampo. I recettori per l’acetilcolina si dividono in due tipi (fig. 2): i nicotinici, canali ionici per il sodio e il calcio, e i muscarinici, accoppiati alle proteine G (➔ proteina). In genere, l’acetilcolina è un mediatore eccitatorio.
Alla famiglia delle monoammine appartengono la noradrenalina, l’adrenalina, la dopammina, la serotonina e l’istamina. La noradrenalina è sintetizzata da neuroni confinati nel midollo allungato, nel ponte e nel nucleo del tratto solitario. Questi neuroni si diffondono a quasi tutte le regioni del sistema nervoso centrale, soprattutto alla corteccia, all’ipotalamo e al midollo spinale. I recettori adrenergici sono diversi, raggruppati in due grandi famiglie, α e β, accoppiati a proteine G. L’effetto dell’adrenalina o della noradrenalina dipende dal tipo di recettore eccitato e, in genere, è di tipo inibitorio. La dopammina è sintetizzata soprattutto in neuroni concentrati nella substantia nigra, nell’ipotalamo e nel bulbo olfattivo: essa è importante nel controllo della regolazione del movimento e nel controllo dell’affettività. La degenerazione delle cellule dopamminergiche porta a una patologia imponente, come quella del morbo di Parkinson, che può essere parzialmente alleviata somministrando un precursore della dopammina, la L-DOPA. Si conoscono almeno quattro recettori per la dopammina, i quali agiscono attraverso proteine G. La serotonina è sintetizzata a partire dal triptofano ed è inattivata per ossidazione dalle monoamminossidasi. I neuroni che la producono sono localizzati soprattutto nel rafe e nel midollo allungato e si irradiano alla corteccia, al midollo spinale e al cervelletto. La serotonina è certamente connessa con importanti processi affettivi e conoscitivi, nonché con il controllo di alcune funzioni importanti, come la regolazione della temperatura e l’appetito. I recettori serotoninergici sono di almeno cinque tipi: i recettori 5-HT1,2,4,5 sono legati a proteine G; il 5-HT3 è invece un canale ionico.
Gli amminoacidi costituiscono una famiglia di n. che è tra le più presenti nel sistema nervoso centrale; essi si suddividono in amminoacidi inibitori, come il GABA e la glicina, e in amminoacidi eccitatori, come il glutammato e l’aspartato. Il GABA è il più importante n. inibitore del sistema nervoso centrale, essendo contenuto in circa il 40% delle sinapsi. È sintetizzato soprattutto da piccoli interneuroni distribuiti in quasi tutto il sistema; la sua sintesi avviene anche in alcuni neuroni che si dirigono dal corpo striato alla substantia nigra. I recettori per il GABA sono di due tipi, i GABA A e i GABA B: i recettori GABA A sono canali ionici per il cloro e mediano i fenomeni inibitori, mentre i recettori GABA B sono accoppiati a proteine G. La glicina è il n. inibitorio presente nel midollo spinale e, anch’essa, attiva recettori-canale per il cloro. L’acido glutammico, che è il mediatore eccitatorio più diffuso nel sistema nervoso centrale, è sintetizzato da neuroni sparsi in molti nuclei del cervello; particolarmente studiati sono i nuclei dell’ippocampo, che forniscono un modello prezioso per analizzare i meccanismi con i quali si instaura e si mantiene la memoria.
Sintesi dei neuropeptidi
Tra i peptidi più studiati vi sono l’ossitocina e la vasopressina, gli ormoni ipotalamici, quali la somatostatina e il fattore per la liberazione delle gonadotropine (GnRH), la colecistochinina, il peptide vasoattivo intestinale (VIP) ecc. Questi peptidi possono essere sintetizzati dagli stessi neuroni che sintetizzano i n. classici; si realizza così, per alcuni neuroni, un duplice codice d’informazione attraverso mediatori differenti. Sembra, infatti, che i peptidi possano avere una funzione più modulatrice che non proprio neurotrasmettitrice: tra le associazioni ricordiamo, quelle tra noradrenalina e neuropeptide Y (NPY), acetilcolina e calcitonin gene-related peptide. La scoperta più rilevante in questo settore ha riguardato i peptidi oppioidi, come le endorfine e le encefaline, che stimolano i recettori per la morfina: la loro individuazione ha permesso di capire il meccanismo d’azione della morfina, cioè in che modo essa produca gli effetti analgesici, stimolanti o depressivi nel SNC.